
IAP011 Primary solitary and multiple hydatid cysts of the brain

J. Eirkanmot. Microhinl., 19(2). 2002 pp. 175-1 82 0 2002 by the Society of Protomologists
Changes in Host and Parasite-Derived Cellular and Extracellular Matrix Components in Developing Cysts of Myxobolus pendula (Myxozoa)
AMANDA A. MARTYN, HENRY HONG, MAURICE J. RINGUETTE and SHERWIN S. DESSER Department of Zoology, University of Toronto, Toronto, Onturio M5S 3G.5, Canada
ABSTRACT. Cysts of Myxobolus yendulu from the gill arch of creek chub (Semotilus atromcrc~rlatus) were examined at various stages of development using light and electron microscopy. The subepithelial host connective tissue underwent dramatic changes, including degradation and remodelling of collagen and vascularisation. in response to the infection. Inflammatory cells lay in a fluid-filled space beneath the host’s connective tissue and surrounded a distinctive parasite-derived matrix, composed of collagen fibril bundles embedded in cellular processes of the underlying secretory cells. This collagen matrix was resistant to degradation and invasion by leukocytes. Secretion of a matrix by M. pendula as a structural support, and a protective barrier against the host inflammatory cells is a novel observation for cyst-forming Myxosporea.
Key Words. Collagen, development, electron microscopy, inflammation, light microscopy, matrix remodelling, Myxosporea, ultra- structure, vascularisation.
HE phylum Myxozoa is comprised of parasitic organisms T with a unique life cycle that involves alternation between fish and worm hosts (Wolf and Markiw 1984). Although more than 1,300 species of Myxozoa have been described, less than 30 complete life cycles have been demonstrated (Kent et al. 2001). Depending on the species, development of the myxospo- rean stage in the fish host can occur in the blood, on the lumenal surface of organs, or in tissues where cyst-like xenomas are often formed (Molnkr 1994). While several authors have de- scribed a host-derived connective tissue layer surrounding ma- ture myxozoan cysts (see MolnSu 1994; Shul’man 1988), most of the studies of histozoic species have focussed mainly on sporogenesis, so little is known about the structural changes of cysts during their development.
Guilford (1 967) studied sporogenesis of Myxobolus pendida in globular cysts on the gill arch of creek chub, Sernoti1Li.s atru- maculatus Mitchill, 1818. Based on observations of whole cysts and histological sections, he described a distinct “ectoplasm’ ’ associated with the parasite and a parasitic “endoplasm” where sporogenesis occurs. He also noted a fluid-filled space contain- ing granulocytes that separated the parasite from the host mu- cous membrane.
In order to elucidate the role of the host and parasite during the formation of the complex cysts of M. pendula, we examined cysts at different stages of development by light and electron microscopy.
MATERIALS AND METHODS Semotilus atromaculatus (N = 21 1) were caught in Lake Sa-
sejewun (45” 35‘ 3 0 N, 78” 31‘ 30” w) in Algonquin Park, Ontario, from 30 May to 1 August 2000 and from 1 to 5 May 2001, using Gee@ minnow traps (Dynamic Aqua-Supply Ltd., Surrey, BC) baited with Purina puppy chow@ (Ralston Purina Canada Inc., Mississauga, ON). These fish were pithed and their gills were examined for cysts using a dissecting microscope. A total of 31 cysts were isolated and excised. They were assigned to four groups on the basis of their development: 1) “young” cysts (early May), which were the first stages visible on the gills; 2) “early intermediate” (late May to early June); 3) “late intermediate” (late June to early July) cysts; and 4) “mature” cysts (mid- to late July).
Unfixed cysts were photographed using a Leica MZ8 dis- secting microscope with a Photometrics Coolsnap colour digital camera. For transmission electron microscopy (TEM), cysts from each stage of development, and uninfected gill tissue as a control, were fixed at room temperature in 2.5% glutaralde-
Corresponding Author: S. Desser-Telephone number: 416-978- 6956; FAX number: 416-978-8532; E-mail: [email protected].
hyde in 0.1 M Sorenson’s phosphate buffer (pH 7.0), postfixed at room temperature with 1.0% OsO, in 0.15 M Sorenson’s phosphate buffer (pH 7.0), dehydrated through an ascending ethanol series, infiltrated and embedded in Spurr’s resin, and polymerised overnight at 65 “C. Semithin sections, stained with 0.2% toluidine blue and 0.5% methylene blue in 0.5% sodium borate, were photographed using an Olympus BX60 microscope with a Hitachi KP-D5O colour digital camera. Ultrathin sections stained with 3% uranyl acetate in 50% methanol and Reynolds’ lead citrate stain (Reynolds 1963) were examined with a Hitachi H7000 transmission electron microscope. All sections examined were through the middle of each cyst.
For scanning electron microscopy (SEM), cysts were fixed at room temperature in 2.5% glutaraldehyde in 0.1 M Soren- son’s phosphate buffer (pH 7.0), snap-frozen in liquid nitrogen, cryofractwed, postfixed at room temperature with 1.0% OsO,
Fig. 1. Mature cyst of Myxobolus penduln attached to a gill arch of the creek chub, Semotilus atromaculutus. An anastomosing network of blood vessels extends from the efferent branchial artery into the cyst wall. Bar = 250 km.
175
176 J. EUKARYOT. MICROBIOL., VOL. 49, NO. 2, MARCH-APRIL 2002
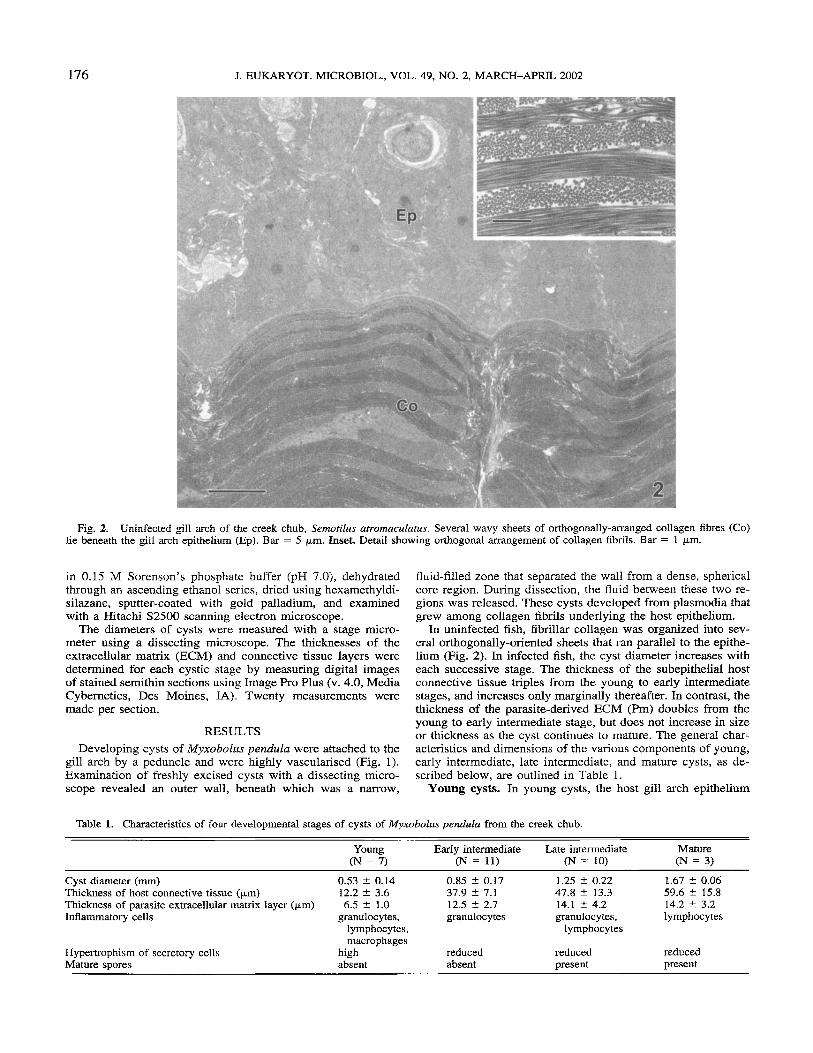
Fig. 2. Uninfected gill arch of the creek chub, Semofilits urroniaculatus. Several wavy sheets of orthogonally-asranged collagen fibres (Co) lie beneath the gill arch epithelium (Ep). Bar = 5 pm. Inset. Detail showing orthogonal arrangement of collagen fibrils. Bar = 1 pm.
in 0.15 M Sorenson’s phosphate buffer (pH 7.0), dehydrated through an ascending ethanol series, dried using hexamethyldi- silazane, sputter-coated with gold palladium, and examined with a Hitachi S2500 scanning electron microscope.
The diameters of cysts were measured with a stage micro- meter using a dissecting microscope. The thicknesses of the extracellular matrix (ECM) and connective tissue layers were determined for each cystic stage by measuring digital images of stained semithin sections using Image Pro Plus (v. 4.0, Media Cybernetics, Des Moines, IA). Twenty measurements were made per section.
RESULTS Developing cysts of Myxobolzis pendula were attached to the
gill arch by a peduncle and were highly vascularised (Fig. 1). Examination of freshly excised cysts with a dissecting micro- scope revealed an outer wall, beneath which was a narrow,
fluid-filled zone that separated the wall from a dense, spherical core region. During dissection, the fluid between these two re- gions was released. These cysts developed from plasmodia that grew among collagen fibrils underlying the host epithelium.
In uninfected fish, fibrillar collagen was organized into sev- eral orthogonally-oriented sheets that ran parallel to the epithe- lium (Fig. 2). In infected fish, the cyst diameter increases with each successive stage. The thickness of the subepithelial host connective tissue triples from the young to early intermediate stages, and increases only marginally thereafter. In contrast, the thickness of the parasite-derived ECM (Pm) doubles from the young to early intermediate stage, but does not increase in size or thickness as the cyst continues to mature. The general char- acteristics and dimensions of the various components of young, early intermediate, late intermediate, and mature cysts, as de- scribed below, are outlined in Table 1.
Young cysts. In young cysts, the host gill arch epithelium
Table 1. Characteristics of four developmental stages of cysts of Myxobolus pendula from the creek chub.
Young Early intermediate Late intermediate Mature (N = 7 ) (N = 11) (N = 10) (N = 3)
Cyst diameter (mm) 0.53 2 0.14 0.85 ? 0.17 1.25 2 0.22 1.67 ? 0.06 Thickness of host connective tissue (pm) 12.2 i 3.6 37.9 t 7.1 47.8 2 13.3 59.6 rt 15.8 Thickness of parasite extracellular matrix layer (pm) 6.5 rt 1 .O 12.5 -+ 2.7 14.1 % 4.2 14.2 L 3.2 Inflammatory cells granulocytes, granulocytes granulocytes, lymphocytes
lymphocytes, lymphocytes macrophages
Hypertrophism of secretory cells high reduced reduced reduced Mature spores absent absent present present

M m T Y N ET AL.- DEVELOPMENT OF MYXOBOLUS PENDULA CYSTS 177
Fig. 3. Section through a young cyst showing host epithelium (Ep) and subepithelial collagen (arrow) of the creek chub, Semotilus ufru- maculatus, and hypertrophic secretory cells (Sc) and sporogenic region (Sp) of Myxobolus pendula. Macrophages (Ma) and lymphocytes (Lc) are seen between the collagen sheet and a thin parasite ECM (Pm). Bar = 10 urn.
overlaid a loose connective tissue layer that contained a colla- gen matrix with a dramatically different supramolecular orga- nization (Fig. 3) than that of the subepithelial collagen network observed in uninfected gills (Fig. 2). A wavy sheet of fibrillar collagen surrounded by unorganized collagen fibrils was visi- ble. Beneath this collagenous sheet was a lucent zone inter- spersed with numerous inflammatory cells, including macro-
phages, lymphocytes, and granulocytes. Underlying this loose connective tissue and associated with the parasite domain was an extracellular matrix, composed of collagen fibril bundles em- bedded between cellular processes. A layer of elongate hyper- trophic cells rich in mitochondi-ia and endoplasinic reticulum lay in intimate contact between the ECM and the centrally- located generative cells.
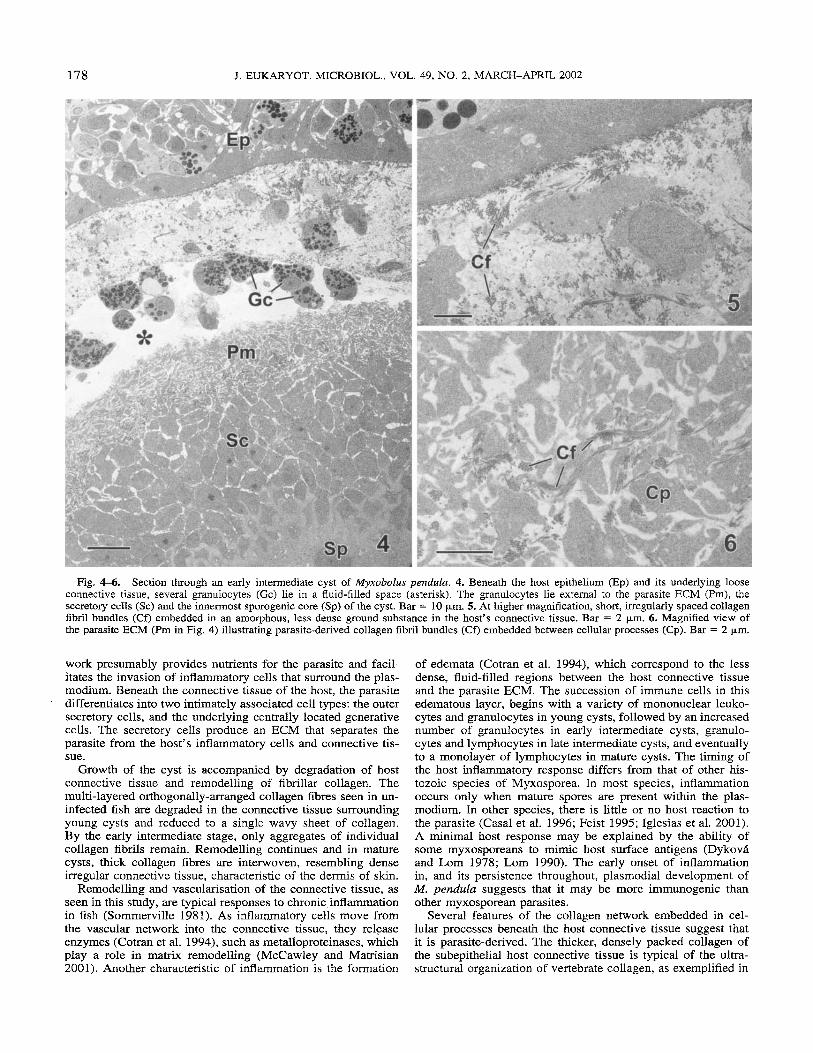
Early intermediate cysts. (Fig. 4-6) By this stage of de- velopment, the connective tissue beneath the host epithelium had undergone further remodelling (Fig. 4). The single sheet of fibrillar collagen was replaced by thin collagen fibres with no specific orientation, embedded in a less dense ground substance (Fig. 5) . Granulocytes, the predominant inflammatory cells at this stage, were observed in a fluid-filled space, devoid of ma- trix. The underlying parasite-associated ECM had doubled in size (Fig. 6). The cells between the Pm and the central sporo- genic region were more numerous than in young cysts, although they appeared smaller, more rounded, and less hypertrophic.
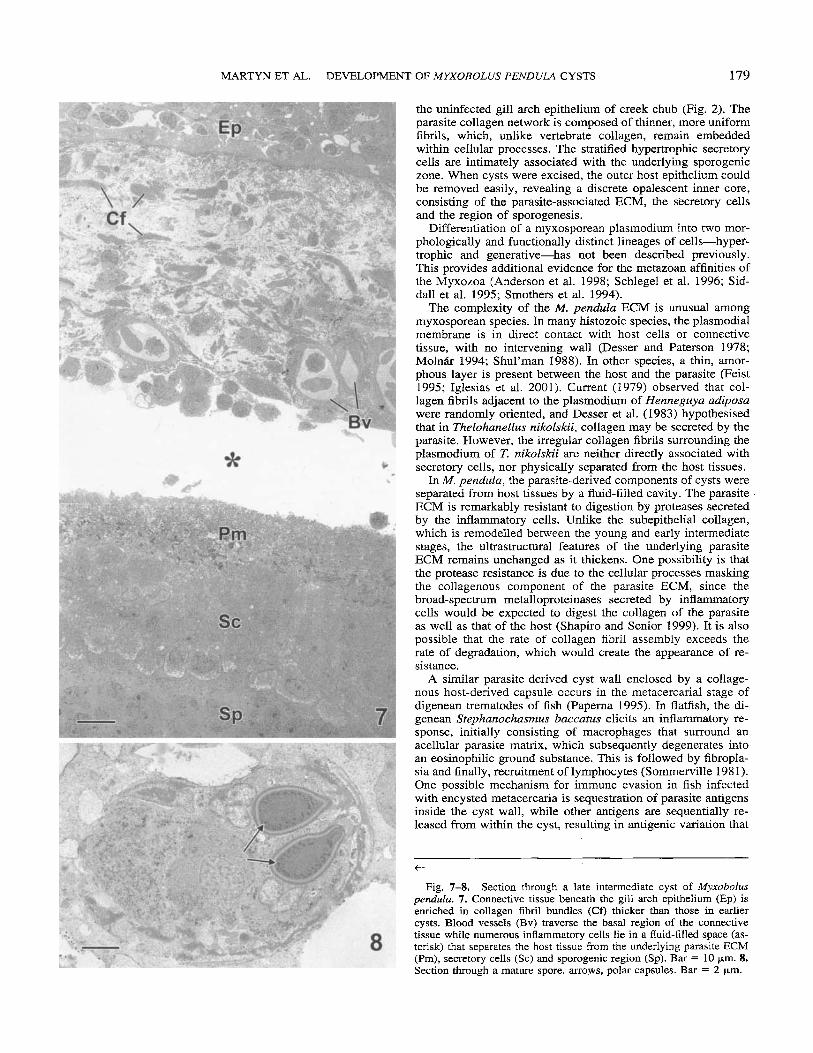
Late intermediate cysts. (Fig. 7, 8) Several morphological changes had occurred within the cyst wall between early and late intermediate stages. The layer of connective tissue under- lying the host epithelium was somewhat thicker and many of the collagen fibres had increased in diameter (Fig. 7). At this stage, blood vessels were prominent, and granulocytes and lym- phocytes represented the majority of the immune cells. The un- derlying parasite-associated ECM had thicker collagen fibril bundles than those observed in early intermediate cysts. How- ever, there was no significant change in the appearance of se- cretory cells between early and late intermediate cysts. Mature spores were visible for the first time within the sporogenic core (Fig. 8).
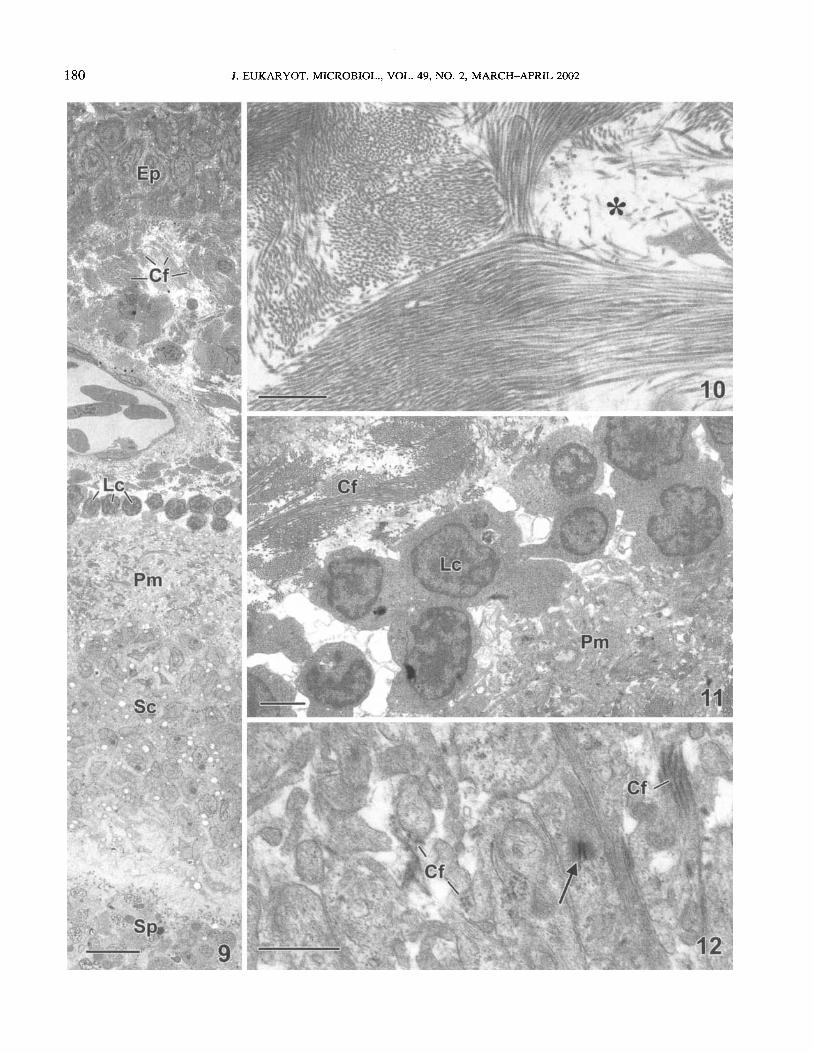
Mature cysts. In mature cysts (Fig. 9-12), the connective tissue layer beneath the epithelium had increased in density and marginally in size. Both the ratio of fibrillar collagen to the less dense matrix, and the size of the collagen fibres had increased (Fig. 9). Interspersed within collagen bundles were less dense areas containing small aggregates of fibrils (Fig. 10). Numerous blood vessels were visible, consistent with the extensive vas- cularisation observed in whole, unfixed cysts (Fig. 1). A thin layer of lymphocytes (Fig. 9, 11) surrounded the parasite-as- sociated ECM. At higher magnification, the collagen fibres as- sociated with the host connective tissue appeared densely packed (Fig. 10). In contrast, the collagen fibres associated with the parasite-derived cellular processes were smaller and more dispersed. representing a lower proportion of the total matrix volume. Desmosome-like junctional complexes adjoined the parasite-cellular processes (Fig. 12). The layer of secretoiy cells underlying the Pm resembled those in early and late interme- diate cysts (Fig. 9).
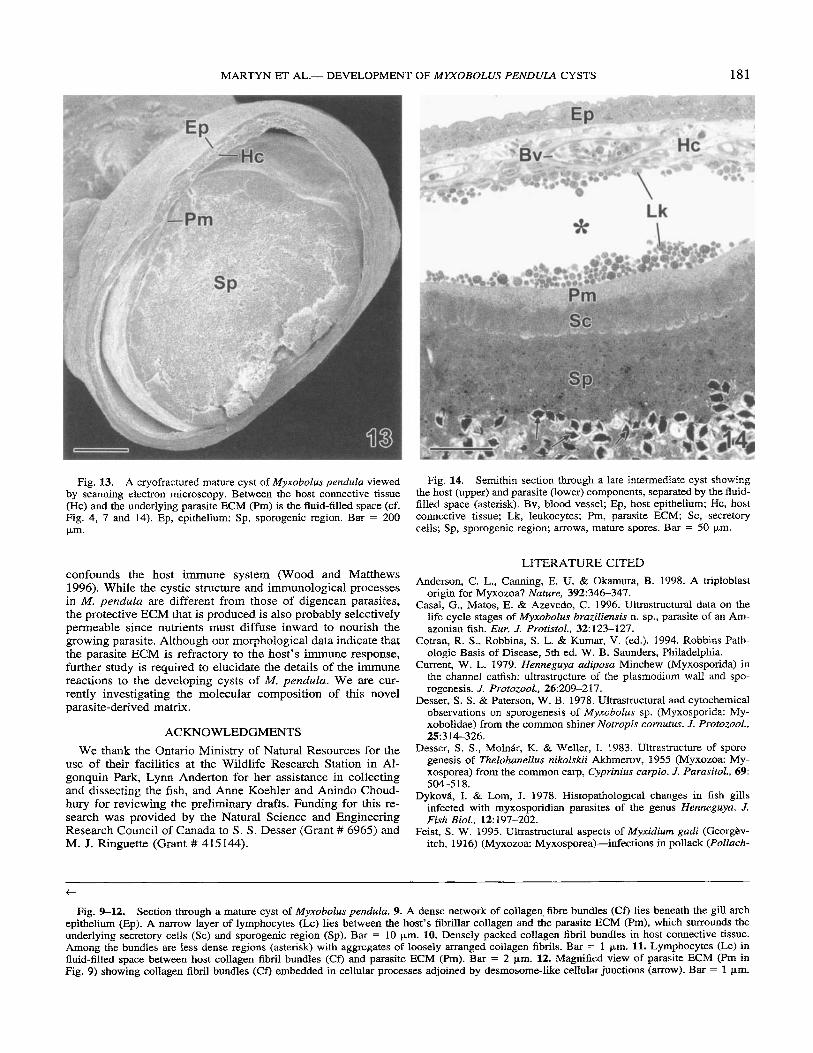
Scanning electron microscopy of a cryofractured mature cyst provided an overview of the relative thicknesses and boundaries of the layers (Fig 13). The epithelium and the collagen-rich connective tissue were in direct contact. In many regions, these layers were noticeably separated from the Pm, which surround- ed the secretory cells and sporogenic core. The distinct bound- aries between layers were also apparent in semithin sections (Fig. 14).
DISCUSSION Our study of developing cysts of Myxobohs pendula re-
vealed that both host and parasite components undergo dramatic structural changes. As the plasmodia1 cells differentiate and di- vide, the host epithelium and dermis overlying the expanding plasmodium are stretched outward, forming the cyst wall. Growth of the cyst is associated with angiogenesis in the host connective tissue surrounding the parasite. This vascular net-
178 J . EUKARYOT. MICROBIOL., VOL. 49, NO. 2, MARCH-APRIL 2002
Fig. 4-6. Section through an early intermediate cyst of Myxobolus peizdulu. 4. Beneath the host epithelium (Ep) and its underlying loose connective tissue, several granulocytes (Gc) lie in a fluid-filled space (asterisk). The granulocytes lie external to the parasite ECM (Pm), the secretory cells (Sc) and the innermost sporogenic core (Sp) of the cyst. Bar = 10 pm. 5. At higher magnification, short, irregularly spaced collagen fibril bundles (Cf) embedded in an amorphous, less dense ground substance in the host’s connective tissue. Bar = 2 pm. 6. Magnified view of the parasite ECM (Pm in Fig. 4) illustrating parasite-derived collagen fibril bundles (Cf) embedded between cellular processes (Cp). Bar = 2 pm.
work presumably provides nutrients for the parasite and facil- itates the invasion of inflammatory cells that surround the plas- modium. Beneath the connective tissue of the host, the parasite differentiates into two intimately associated cell types: the outer secretory cells, and the underlying centrally located generative cells. The secretory cells produce an ECM that separates the parasite from the host’s inflammatory cells and connective tis- sue.
Growth of the cyst is accompanied by degradation of host connective tissue and remodelling of fibrillar collagen. The multi-layered orthogonally-arranged collagen fibres seen in un- infected fish are degraded in the connective tissue surrounding young cysts and reduced to a single wavy sheet of collagen. By the early intermediate stage, only aggregates of individual collagen fibrils remain. Remodelling continues and in mature cysts, thick collagen fibres are interwoven, resembling dense irregular connective tissue, characteristic of the dermis of skin.
Remodelling and vascularisation of the connective tissue, as seen in this study, are typical responses to chronic inflammation in fish (Sommerville 1981). As inflammatory cells move from the vascular network into the connective tissue, they release enzymes (Cotran et al. 1994), such as metalloproteinases, which play a role in matrix remodelling (McCawley and Matrisian 2001). Another characteristic of inflammation is the formation
of edemata (Cotran et al. 1994), which correspond to the less dense, fluid-filled regions between the host connective tissue and the parasite ECM. The succession of immune cells in this edematous layer, begins with a variety of mononuclear leuko- cytes and granulocytes in young cysts, followed by an increased number of granulocytes in early intermediate cysts, granulo- cytes and lymphocytes in late intermediate cysts, and eventually to a monolayer of lymphocytes in mature cysts. The timing of the host inflammatory response differs from that of other his- tozoic species of Myxosporea. In most species, inflammation occurs only when mature spores are present within the plas- modium. In other species, there is little or no host reaction to the parasite (Casal et al. 1996; Feist 1995; Iglesias et al. 2001). A minimal host response may be explained by the ability of some myxosporeans to mimic host surface antigens (Dykov6 and Lom 1978; Lom 1990). The early onset of inflammation in, and its persistence throughout, plasmodia1 development of M. pendulu suggests that it may be more immunogenic than other myxosporean parasites.
Several features of the collagen network embedded in cel- lular processes beneath the host connective tissue suggest that it is parasite-derived. The thicker, densely packed collagen of the subepithelial host connective tissue is typical of the ultra- structural organization of vertebrate collagen, as exemplified in
MARTYN ET AL.- DEVELOPMENT OF MYXOBOLUS PENDULA CYSTS 179
the uninfected gill arch epithelium of creek chub (Fig. 2). The parasite collagen network is composed of thinner, more uniform fibrils, which, unlike vertebrate collagen, remain embedded within cellular processes. The stratified hypertrophic secretory cells are intimately associated with the underlying sporogenic zone. When cysts were excised, the outer host epithelium could be removed easily, revealing a discrete opalescent inner core, consisting of the parasite-associated ECM, the secretory cells and the region of sporogenesis.
Differentiation of a myxosporean plasmodium into two mor- phologically and functionally distinct lineages of cells-hyper- trophic and generative-has not been described previously. This provides additional evidence for the metazoan affinities of the Myxozoa (Anderson et al. 1998; Schlegel et al. 1996; Sid- dall et al. 1995; Smothers et al. 1994).
The complexity of the M. pendula ECM is unusual among myxosporean species. In many histozoic species. the plasmodia1 membrane is in direct contact with host cells or connective tissue, with no intervening wall (Desser and Paterson 1978; Moln6r 1994; Shul’man 1988). In other species, a thin, amor- phous layer is present between the host and the parasite (Feist 1995; Iglesias et al. 2001). Current (1979) observed that col- lagen fibrils adjacent to the plasmodium of Henneguya udiposa were randomly oriented, and Desser et al. (1983) hypothesised that in Thelohunellus nikolskii, collagen may be secreted by the parasite. However, the irregular collagen fibrils surrounding the plasmodium of T. nikolskii are neither directly associated with secretory cells, nor physically separated from the host tissues.
In M. pendula, the parasite-derived components of cysts were separated from host tissues by a fluid-filled cavity. The parasite ECM is remarkably resistant to digestion by proteases secreted by the inflammatory cells. Unlike the subepithelial collagen, which is remodelled between the young and early intermediate stages, the ultrastructural features of the underlying parasite ECM remains unchanged as it thickens. One possibility is that the protease resistance is due to the cellular processes masking the collagenous component of the parasite ECM, since the broad-spectrum metalloproteinases secreted by inflammatory cells would be expected to digest the collagen of the parasite as well as that of the host (Shapiro and Senior 1999). It is also possible that the rate of collagen fibril assembly exceeds the rate of degradation, which would create the appearance of re- sistance.
A similar parasite-derived cyst wall enclosed by a collage- nous host-derived capsule occurs in the metacercarial stage of digenean trematodes of fish (Paperna 1995). In flatfish, the di- genean Stephunochasinus bnccatiis elicits an inflammatory re- sponse, initially consisting of macrophages that surround an acellular parasite matrix, which subsequently degenerates into an eosinophilic ground substance. This is followed by fibropla- sia and finally, recruitment of lymphocytes (Sommerville 1981). One possible mechanism for immune evasion in fish infected with encysted metacercaria is sequestration of parasite antigens inside the cyst wall, while other antigens are sequentially re- leased from within the cyst, resulting in antigenic variation that
t
Fig. 7-8. Section through a late intermediate cyst of Myxobolus peiidula. 7. Connective tissue beneath the gill arch epithelium (Ep) is enriched in collagen fibril bundles (Cf) thicker than those in earlier cysts. Blood vessels (Bv) traverse the basal region of the connective tissue while numerous inflammatory cells lie in a fluid-filled space (as- terisk) that separates the host tissue from the underlying parasite ECM (Pm), secretory cells (Sc) and sporogenic region (Sp). Bar = 10 pm. 8. Section through a mature spore. arrows, polar capsules. Bar = 2 pm.
180 J. EUKARYOT. MICROBIOL., VOL. 49, NO. 2, MARCH-APRIL 2002
MARTYN ET AL.- DEVELOPMENT OF MXXOBOLUS PENDULA CYSTS 181
Fig. 13. A cryofractured mature cyst of M y x o b o h pendda viewed by scanning electron microscopy. Between the host connective tissue (Hc) and the underlying parasite ECM (Pm) is the fluid-filled space (cf. Fig. 4, 7 and 14). Ep, epithelium; Sp, sporogenic region. Bar = 200 pm.
Fig. 14. Semithin section through a late intermediate cyst showing the host (upper) and parasite (lower) components, separated by the fluid- filled space (asterisk). Bv, blood vessel; Ep, host epithelium; Hc, host connective tissue; Lk, leukocytes; Pm, parasite ECM; Sc, secretory cells; Sp, sporogenic region; arrows, mature spores. Bar = 50 pi”.
confounds the host immune system (Wood and Matthews 1996). While the cystic structure and immunological processes in M. pendula are different from those of digenean parasites, the protective ECM that is produced is also probably selectively permeable since nutrients must diffuse inward to nourish the growing parasite. Although our morphological data indicate that the parasite ECM is refractory to the host’s immune response, further study is required to elucidate the details of the immune reactions to the developing cysts of M. pendula. We are cur- rently investigating the molecular composition of this novel parasite-derived matrix.
ACKNOWLEDGMENTS We thank the Ontario Ministry of Natural Resources for the
use of their facilities at the Wildlife Research Station in Al- gonquin Park, Lynn Anderton for her assistance in collecting and dissecting the fish, and Anne Koehler and Anindo Choud- hury for reviewing the preliminary drafts. Funding for this re- search was provided by the Natural Science and Engineering Research Council of Canada to S. S. Desser (Grant # 6965) and M. J. Ringuette (Grant # 415144).
LITERATURE CITED Anderson, C. L., Canning, E. U. & Okamura, B. 1998. A triploblast
origin for Myxozoa? Nature, 392:346-347. Casal, G., Matos, E. & Azevedo, C. 1996. Ultrastructurid data on the
life cycle stages of Myxobolus braziliensis n. sp., parasite of an Am- azonian fish. Eur. J. Protistol., 32: 123-127.
Cotran, R. S., Robbins, S. L. & Kumru;, V. (ed.). 1994. Robbins Path- ologic Basis of Disease, 5th ed. W. B. Saunders, Philadelphia.
Current, W. L. 1979. Hennegu,yu adiposu Minchew (Myxosporida) in the channel catfish: ultrastmcture of the plasmodium wall and spo- rogenesis. J. Protozoal., 26:209-217.
Desser, S. S. & Paterson, W. B. 1978. Ultrastructural and cytochemical observations on sporogenesis of Myxobolrrs sp. (Myxosporida: My- xobolidae) from the common shiner Notropis cornuhis. J. Protozoal., 25314326.
Desser, S . S., Molnhr, K. & Weller, I. 1983. Ultrastructure of sporo- genesis of Thelohanellus nikolskii Akhmerov, 1955 (Myxozoa: My- xosporea) from the common carp, Cyprinius curpio. J. PurasiroL, 69:
Dykovft, I. & Lom, J. 1978. Histopathologlcal changes in fish gills infected with myxosporidian parasites of the genus Henneguyu. J. Fish Biol., 12: 197-202.
Feist, S. W. 1995. Ultrastructural aspects of Myxidium gudi (Georgbv- itch, 1916) (Myxozoa: Myxosporea)-infections in pollack (Polluch-
504-5 18.
t
Fig. 9-12. Section through a mature cyst of Myxobolus pendulu. 9. A dense network of collagen fibre bundles (Cf) lies beneath the gill arch epithelium (Ep). A narrow layer of lymphocytes (Lc) lies between the host’s fibrillar collagen and the parasite ECM (Pm), which surrounds the underlying secretory cells (Sc) and sporogenic region (Sp). Bar = 10 pm. 10. Densely packed collagen fibril bundles in host connective tissue. Among the bundles are less dense regions (asterisk) with aggregates of loosely arranged collagen fibrils. Bar = 1 pm. 11. Lymphocytes (Lc) in fluid-filled space between host collagen fibril bundles (Cf) and parasite ECM (Pm). Bar = 2 pm. 12. Magnified view of parasite ECM (Pm in Fig. 9) showing collagen fibril bundles (Cf) embedded in cellular processes adjoined by desmosome-like cellular junctions (arrow). Bar = 1 km.

182 J. EUKARYOT. MICROBIOL., VOL. 49, NO. 2, MARCH-APRIL 2002
ius polltrcltiirs L. ) and saithe (P. virens L.). Eur. J . Protbtol., 31:
Guilford, H. G. 1967. Myxosoma pendicla n. sp. (Protozoa, Myxospor- ida) from the creek chub Senzorihs atromaculatrs. J. Protozool., 14: 196-198.
Iglesias, R., Parami, A,, Alvarez, F.. Leiro, J. & Sanmartin, M. L. 2001. Myxobolid parasites infecting the gills of the Iberian nase, Chon- drostonm polylepis (Steindacher), in Galicia (NW Spain), with a de- scription of Myxobolus gallaicus sp. nov. J. Fish Dis., 24: 125-134.
Kent, M. L., Andree, K. B., Bartholomew, J. L., El-Matbouli, M., Des- ser, S. S., Devlin, R. H., Feist, S. W., Hedrick, R. P., Hoffmann, R. W., Khattra, J., Hallett, S. L., Lester, R. J. G., Longshaw, M., Pal- enzuela, O., Siddall, M. E. & Xiao, C . 2001. Recent advances in our knowledge of the Myxozoa. J. Eukuiyot. Microbiol., 48:395413.
Lom, J . 1990. Phylum Myxozoa. In: Margnlis, L., Corliss, J. O., Mel- konian, M. & Chapman, D. J (ed.), Handbook of Protoctista. Jones & Bartlett, Boston. 4:36-52.
McCawley, L. J. & Matrisian, L. M. 2001. Matrix metalloproteinases: they’re not just for matrix anymore! Curr. Opin. CeN Biol., 13534- 540.
Molnfir, K. 1994. Comments on the host, organ and tissue specificity of fish myxosporeans and on the types of their intrapiscine devel- opment. Hung. SOC. Parasitol., 275-20.
Paperna, I. 1995. Digenea (Phylum Platyhelminthes) In: Woo, P. T. K. (ed.), Fish Diseases and Disorders. Vol. 1. Protozoan and Metazoan Infections. C.A.B. Int‘l., Wallingford, U.K. 9:329-389.
Reynolds. E. S. 1963. The use of lead citrate at high pH as an electron- opaque stain in electron microscopy. J. Cell Biof.. 17:208-212.
309-317. Schlegel, M., Lom, J., Stechmann, A,, Bernhdrd, D., Leipe, D., DykovB,
I. & Sogin, M. L. 1996. Phylogenetic analysis of complete small subunit ribosomal RNA coding region of Myxidium lieberkuehni: ev- idence that Myxozoa are Metazoa and related to the Bilateria. Arch. Protistenkd., 147: 1-9.
Shapiro, S. D. & Senior, R. M. 1999. Matrix metalloproteinases. Matrix degradation and more. Am. J. Respir. Cell Mol. Biol.. 2O:llOO-1102.
Shul’man, S. S. 1988. Myxosporidia of the USSR. Amerind Publishing Co. Pvt. Ltd., New Delhi.
Siddall, M. E., Martin, D. S., Bridge, D., Desser, S. S. & Cone, D. K. 1995. The demise of a phylum of protists: phylogeny of Myxozoa and other parasitic cnidaria. J. Purusitol., 81:961-967.
Smothers, J. F., von Dohlen, C. D., Smith, L. H., Jr.& Spall, R. D. 1994. Molecular evidence that the myxozoan protists are metazoans. Science, 265:1719-1721.
Soinmerville, C. 1981. A comparative study of the tissue response to invasion and encystment by Stephanochasrnus baccutus (Nicoll, 1907) (Digenea: Acanthocolpidae) in four species of flatfish. J. Fish Dis., 453-68.
Wolf, K. & Markiw, M. E. 1984. Biology contravenes taxonomy in the Myxozoa: new discoveries show alternation of invertebrate and ver- tebrate hosts. Science, 225:1449-1452.
Wood, B. P. & Matthews, R. A. 1996. Reactions of sera from immu- nised and experimentally infected thick-lipped grey mullet, Cheylnn lubrosus (Risso, 1826), with isolated proteins of Crptocotyle lingua (Creplin, 1825). Fish Shelljish Immzcnol., 6:483-492.
Received 12/I4/2001, 02/04/02; accepted 02/19/02
54th Annual Meeting of the
Society of rotozoologists
Salt Lake City, Utah
May 117, 18, & 19, 2082
The Society of Protozoologists (SP) is pleased to announce the 54th annual meeting of the Society in Salt Lake City, Utah at the Grand America Hotel.
ScientiJc Program Chair: Dr. Rebecca J . Gast Woods Hole Oceanographic Institution MS #32 3-24 Redfield Bldg Woods Hole, MA 02543 Tel: 508/289-3209
E-mail: rgast @ whoi.edu FASX: 508/457-2169
Copyright © 2022 FDOKUMEN




















