
Chain-length determination mechanism of isoprenyl diphosphate synthases and implications for...
7
TIBS 24 – NOVEMBER 1999 445 REVIEWS Waterfield, M. D. (1999) Curr. Opin. Cell Biol. 11, 219–225 18 Rameh, L. E. et al. (1997) J. Biol. Chem. 272, 22059–22066 19 Ferguson, K. M., Lemmon, M. A., Schlessinger, J. and Sigler, P. B. (1995) Cell 83, 1037–1046 20 Fukuda, M., Kojima, T., Kabayama, H. and Mikoshiba, K. (1996) J. Biol. Chem. 271, 30303–30306 21 Bromann, P. A., Boetticher, E. E. and Lomasney, J. W. (1997) J. Biol. Chem. 272, 16240–16246 22 Isakoff, S. J. et al. (1999) EMBO J. 17, 5374–5387 23 Salim, K. et al. (1996) EMBO J. 15, 6241–6250 24 Hall, A. (1998) Science 279, 509–514 25 Soisson, S. M. et al. (1998) Cell 95, 259–268 26 Liu, X. et al. (1998) Cell 95, 269–277 27 Aghazadeh, B. et al. (1998) Nat. Struct. Biol. 5, 1019–1107 28 Zheng, J. et al. (1997) J. Biol. Chem. 272, 30340–30344 29 Nimnual, A. S., Yatsula, B. A. and Bar-Sagi, D. (1998) Science 279, 560–563 30 Qian, X. et al. (1998) Mol. Cell. Biol. 18, 771–778 31 Han, J. et al. (1998) Science 279, 558–563 32 Jiang, Y. et al. (1998) Nature 395, 808–813 33 Klarlund, J. K. et al. (1998) J. Biol. Chem. 273, 1859–1862 34 Nagel, W. et al. (1998) J. Biol. Chem. 273, 14853–14861 35 Venkateswarlu, K., Oatey, P. B., Tavare, J. M. and Cullen, P. J. (1998) Curr. Biol. 8, 463–466 36 Olson, M. F. et al. (1997) Oncogene 15, 2827–2831 37 Nicholls, A., Sharp, K. and Honig, B. (1991) Proteins 11, 281 38 Harlan, J. E., Yoon, H. S., Hajduk, P. J. and Fesik, S. W. (1995) Biochemistry 34, 9859–9864 39 Hyvönen, M. et al. (1995) EMBO J. 14, 4676–4685 40 Lemmon, M. A. et al. (1995) Proc. Natl. Acad. Sci. U. S. A. 92, 10472–10476 41 Frech, M. et al. (1997) J. Biol. Chem. 272, 8474–8481 42 Klein, D. E. et al. (1998) J. Biol. Chem. 273, 27725–27733 43 Fushman, D. et al. (1998) J. Biol. Chem. 273, 2835–2843 44 Kubiseski, T. J. et al. (1997) J. Biol. Chem. 272, 1799–1804 45 Koshiba, S. et al. (1997) J. Mol. Biol. 269, 579–591 ISOPRENOIDS ARE THE most chemically diverse family of naturally occurring com- pounds that are widely distributed among Archaea, Bacteria and Eukarya. Many types of isoprenoids (e.g. steroids, choles- terol, retinoids, carotenoids, ubiquinones, prenyl proteins) are essential compo- nents of the cellular machinery of all organisms due to their roles in a variety of biological processes (Fig. 1). For example, retinoids are involved in morphogenesis and phototransduction, whereas prenyl proteins, including Ras and Rho GTPases, work as signal transducers. Moreover, steroids have distinct biological functions ranging from the regulation of sodium re- absorption by aldosterone to the control of female sexual development by estro- gens. Isoprenoids vary greatly in size but are all built from repetitions of the same C 5 isoprene unit: for example, steroids and cholesterol are derived from the C 15 farne- syl diphosphate (FPP), whereas retinoids, carotenoids and chlorophyll originate from geranylgeranyl diphosphate (GGPP), a C 20 isoprenoid. Natural rubbers are composed of over 5000 isoprene units. Isoprenyl diphosphate synthases, also referred to as prenyltransferases, cata- lyze the consecutive condensation of iso- pentenyl diphosphate (IPP) with allylic diphosphates to produce linear precur- sors of all isoprenoids (Fig. 1). Each member of this enzyme family is classi- fied according to the length of its final product and the geometry of the newly formed double bonds (E or Z). FPP syn- thase catalyzes the formation of a C 15 product with E-double bonds, whereas undecaprenyl diphosphate synthase gives a C 55 product with Z-double bonds. Interestingly, although the chemical mechanisms of these condensation reac- tions are identical, the chain lengths of the final products never exceed the limit determined by the specificities of each individual enzyme. Comparison of the primary amino acid sequences of prenyl- transferases suggests that all the E-type enzymes evolved from a common ances- tor 1 but that the Z-type enzymes seem to have a different origin 2 . However, despite the increasing availability of DNA se- quences and recent reports of X-ray struc- tures, several important questions remain. Why does the condensation terminate precisely at a particular step? What is the exact molecular mechanism responsible for chain-length determination? How has Chain-length determination mechanism of isoprenyl diphosphate synthases and implications for molecular evolution Kevin Wang and Shin-ichi Ohnuma In the synthesis of isoprenoids, isoprenyl diphosphate synthases catalyze the consecutive condensation of isopentenyl diphosphate with allylic diphosphates to produce a variety of prenyl diphosphates with well-defined chain lengths. Site-directed mutagenesis in conjunction with X-ray crystal- lographic studies have identified specific amino acid residues responsible for chain-length determination. Simple combinations of these residues within a characteristic motif are not only sufficient to confer product spe- cificities to all isoprenyl diphosphate synthases but represent structural features that reflect the enzyme family’s evolutionary course. K. Wang is at the University of California San Francisco, School of Medicine, S 245, Box 0454, San Francisco, CA 94143, USA; and S-i. Ohnuma is at the Dept of Anatomy, University of Cambridge, Downing St, Cambridge, UK CB2 3DY. Email: [email protected] 0968 – 0004/99/$ – See front matter © 1999, Elsevier Science Ltd. All rights reserved. PII: S0968-0004(99)01464-4 Box 1. A note on nomenclature In this article, the name isoprenyl diphos- phate synthase is used as the generic name for the subclass of prenyl transferases that catalyze chain enlongation. Another alterna- tive name is prenyl diphosphate synthase. The term prenyl transferase was avoided as this also includes protein prenyltransferases.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Chain-length determination mechanism of isoprenyl diphosphate synthases and implications for...

TIBS 24 – NOVEMBER 1999
445
REVIEWSWaterfield, M. D. (1999) Curr. Opin. Cell Biol.11, 219–225
18 Rameh, L. E. et al. (1997) J. Biol. Chem. 272,22059–22066
19 Ferguson, K. M., Lemmon, M. A., Schlessinger,J. and Sigler, P. B. (1995) Cell 83, 1037–1046
20 Fukuda, M., Kojima, T., Kabayama, H. andMikoshiba, K. (1996) J. Biol. Chem. 271,30303–30306
21 Bromann, P. A., Boetticher, E. E. and Lomasney,J. W. (1997) J. Biol. Chem. 272, 16240–16246
22 Isakoff, S. J. et al. (1999) EMBO J. 17,5374–5387
23 Salim, K. et al. (1996) EMBO J. 15, 6241–6250
24 Hall, A. (1998) Science 279, 509–51425 Soisson, S. M. et al. (1998) Cell 95, 259–26826 Liu, X. et al. (1998) Cell 95, 269–277
27 Aghazadeh, B. et al. (1998) Nat. Struct. Biol. 5,1019–1107
28 Zheng, J. et al. (1997) J. Biol. Chem. 272,30340–30344
29 Nimnual, A. S., Yatsula, B. A. and Bar-Sagi, D.(1998) Science 279, 560–563
30 Qian, X. et al. (1998) Mol. Cell. Biol. 18,771–778
31 Han, J. et al. (1998) Science 279, 558–56332 Jiang, Y. et al. (1998) Nature 395, 808–81333 Klarlund, J. K. et al. (1998) J. Biol. Chem. 273,
1859–186234 Nagel, W. et al. (1998) J. Biol. Chem. 273,
14853–1486135 Venkateswarlu, K., Oatey, P. B., Tavare, J. M.
and Cullen, P. J. (1998) Curr. Biol. 8, 463–46636 Olson, M. F. et al. (1997) Oncogene 15,
2827–2831
37 Nicholls, A., Sharp, K. and Honig, B. (1991)Proteins 11, 281
38 Harlan, J. E., Yoon, H. S., Hajduk, P. J. and Fesik,S. W. (1995) Biochemistry 34, 9859–9864
39 Hyvönen, M. et al. (1995) EMBO J. 14,4676–4685
40 Lemmon, M. A. et al. (1995) Proc. Natl. Acad.Sci. U. S. A. 92, 10472–10476
41 Frech, M. et al. (1997) J. Biol. Chem. 272,8474–8481
42 Klein, D. E. et al. (1998) J. Biol. Chem. 273,27725–27733
43 Fushman, D. et al. (1998) J. Biol. Chem. 273,2835–2843
44 Kubiseski, T. J. et al. (1997) J. Biol. Chem. 272,1799–1804
45 Koshiba, S. et al. (1997) J. Mol. Biol. 269,579–591
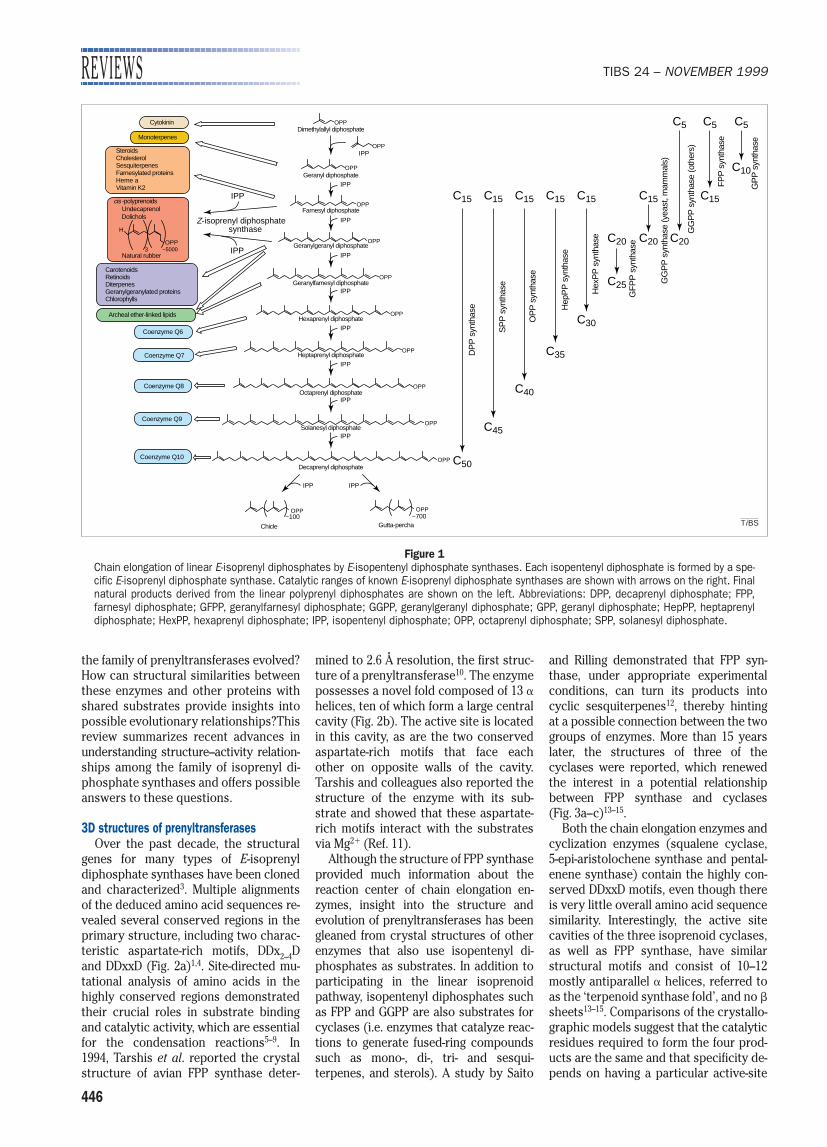
ISOPRENOIDS ARE THE most chemicallydiverse family of naturally occurring com-pounds that are widely distributed amongArchaea, Bacteria and Eukarya. Manytypes of isoprenoids (e.g. steroids, choles-terol, retinoids, carotenoids, ubiquinones,prenyl proteins) are essential compo-nents of the cellular machinery of all
organisms due to their roles in a variety ofbiological processes (Fig. 1). For example,retinoids are involved in morphogenesisand phototransduction, whereas prenylproteins, including Ras and Rho GTPases,work as signal transducers. Moreover,steroids have distinct biological functionsranging from the regulation of sodium re-absorption by aldosterone to the controlof female sexual development by estro-gens. Isoprenoids vary greatly in size butare all built from repetitions of the sameC5 isoprene unit: for example, steroids andcholesterol are derived from the C15 farne-syl diphosphate (FPP), whereas retinoids,carotenoids and chlorophyll originate
from geranylgeranyl diphosphate (GGPP),a C20 isoprenoid. Natural rubbers arecomposed of over 5000 isoprene units.
Isoprenyl diphosphate synthases, alsoreferred to as prenyltransferases, cata-lyze the consecutive condensation of iso-pentenyl diphosphate (IPP) with allylicdiphosphates to produce linear precur-sors of all isoprenoids (Fig. 1). Eachmember of this enzyme family is classi-fied according to the length of its finalproduct and the geometry of the newlyformed double bonds (E or Z). FPP syn-thase catalyzes the formation of a C15product with E-double bonds, whereasundecaprenyl diphosphate synthasegives a C55 product with Z-double bonds.Interestingly, although the chemicalmechanisms of these condensation reac-tions are identical, the chain lengths ofthe final products never exceed the limitdetermined by the specificities of eachindividual enzyme. Comparison of theprimary amino acid sequences of prenyl-transferases suggests that all the E-typeenzymes evolved from a common ances-tor1 but that the Z-type enzymes seem tohave a different origin2. However, despitethe increasing availability of DNA se-quences and recent reports of X-ray struc-tures, several important questions remain.Why does the condensation terminateprecisely at a particular step? What is theexact molecular mechanism responsiblefor chain-length determination? How has
Chain-length determinationmechanism of isoprenyl
diphosphate synthases andimplications for molecular
evolution
Kevin Wang and Shin-ichi OhnumaIn the synthesis of isoprenoids, isoprenyl diphosphate synthases catalyzethe consecutive condensation of isopentenyl diphosphate with allylicdiphosphates to produce a variety of prenyl diphosphates with well-definedchain lengths. Site-directed mutagenesis in conjunction with X-ray crystal-lographic studies have identified specific amino acid residues responsiblefor chain-length determination. Simple combinations of these residueswithin a characteristic motif are not only sufficient to confer product spe-cificities to all isoprenyl diphosphate synthases but represent structuralfeatures that reflect the enzyme family’s evolutionary course.
K. Wang is at the University of California SanFrancisco, School of Medicine, S 245, Box 0454, San Francisco, CA 94143, USA; andS-i. Ohnuma is at the Dept of Anatomy,University of Cambridge, Downing St, Cambridge, UK CB2 3DY. Email: [email protected]
0968 – 0004/99/$ – See front matter © 1999, Elsevier Science Ltd. All rights reserved. PII: S0968-0004(99)01464-4
Box 1. A note on nomenclature
In this article, the name isoprenyl diphos-phate synthase is used as the generic namefor the subclass of prenyl transferases thatcatalyze chain enlongation. Another alterna-tive name is prenyl diphosphate synthase.The term prenyl transferase was avoided asthis also includes protein prenyltransferases.
REVIEWS TIBS 24 – NOVEMBER 1999
446
and Rilling demonstrated that FPP syn-thase, under appropriate experimentalconditions, can turn its products intocyclic sesquiterpenes12, thereby hintingat a possible connection between the twogroups of enzymes. More than 15 yearslater, the structures of three of the cyclases were reported, which renewedthe interest in a potential relationship between FPP synthase and cyclases (Fig. 3a–c)13–15.
Both the chain elongation enzymes andcyclization enzymes (squalene cyclase,5-epi-aristolochene synthase and pental-enene synthase) contain the highly con-served DDxxD motifs, even though thereis very little overall amino acid sequencesimilarity. Interestingly, the active sitecavities of the three isoprenoid cyclases,as well as FPP synthase, have similarstructural motifs and consist of 10–12mostly antiparallel a helices, referred toas the ‘terpenoid synthase fold’, and no bsheets13–15. Comparisons of the crystallo-graphic models suggest that the catalyticresidues required to form the four prod-ucts are the same and that specificity de-pends on having a particular active-site
the family of prenyltransferases evolved?How can structural similarities betweenthese enzymes and other proteins withshared substrates provide insights intopossible evolutionary relationships?Thisreview summarizes recent advances inunderstanding structure–activity relation-ships among the family of isoprenyl di-phosphate synthases and offers possibleanswers to these questions.
3D structures of prenyltransferasesOver the past decade, the structural
genes for many types of E-isoprenyldiphosphate synthases have been clonedand characterized3. Multiple alignmentsof the deduced amino acid sequences re-vealed several conserved regions in theprimary structure, including two charac-teristic aspartate-rich motifs, DDx2–4Dand DDxxD (Fig. 2a)1,4. Site-directed mu-tational analysis of amino acids in thehighly conserved regions demonstratedtheir crucial roles in substrate bindingand catalytic activity, which are essentialfor the condensation reactions5–9. In1994, Tarshis et al. reported the crystalstructure of avian FPP synthase deter-
mined to 2.6 Å resolution, the first struc-ture of a prenyltransferase10. The enzymepossesses a novel fold composed of 13 ahelices, ten of which form a large centralcavity (Fig. 2b). The active site is locatedin this cavity, as are the two conservedaspartate-rich motifs that face eachother on opposite walls of the cavity.Tarshis and colleagues also reported thestructure of the enzyme with its sub-strate and showed that these aspartate-rich motifs interact with the substratesvia Mg21 (Ref. 11).
Although the structure of FPP synthaseprovided much information about the reaction center of chain elongation en-zymes, insight into the structure and evolution of prenyltransferases has beengleaned from crystal structures of otherenzymes that also use isopentenyl di-phosphates as substrates. In addition toparticipating in the linear isoprenoidpathway, isopentenyl diphosphates suchas FPP and GGPP are also substrates for cyclases (i.e. enzymes that catalyze reac-tions to generate fused-ring compoundssuch as mono-, di-, tri- and sesqui-terpenes, and sterols). A study by Saito
Coenzyme Q10
Coenzyme Q9
Coenzyme Q8
Coenzyme Q7
Coenzyme Q6
Archeal ether-linked lipids
CarotenoidsRetinoidsDiterpenesGeranylgeranylated proteinsChlorophylls
SteroidsCholesterolSesquiterpenesFarnesylated proteinsHeme aVitamin K2
Monoterpenes
OPP
OPP
OPP
OPP
OPP
OPP
OPP
OPP
OPP
OPPOPP
OPP
OPP
IPP
IPP
IPP
IPP
IPP
IPP
IPP
IPP
Dimethylallyl diphosphate
Geranyl diphosphate
Farnesyl diphosphate
Geranylgeranyl diphosphate
Geranylfarnesyl diphosphate
Hexaprenyl diphosphate
Heptaprenyl diphosphate
Octaprenyl diphosphate
Solanesyl diphosphate
Decaprenyl diphosphate
IPP
Chicle Gutta-percha~100 ~700
IPP IPP
H
OPP
cis -polyprenoids Undecaprenol Dolichols
Natural rubber~50003
IPP
IPP
Z-isoprenyl diphosphatesynthase
DP
P s
ynth
ase
SP
P s
ynth
ase
OP
P s
ynth
ase
Hep
PP
syn
thas
e
Hex
PP
syn
thas
e
GF
PP
syn
thas
e
GG
PP
syn
thas
e (y
east
, mam
mal
s)
GP
P s
ynth
ase
GG
PP
syn
thas
e (o
ther
s)
FP
P s
ynth
ase
C15
C50
C15
C30
C15 C15
C5
C20C20C20
C25
C5 C5
C10
C15
C35
C15
C40
C15
C45
Cytokinin
Ti BS
Figure 1Chain elongation of linear E-isoprenyl diphosphates by E-isopentenyl diphosphate synthases. Each isopentenyl diphosphate is formed by a spe-cific E-isoprenyl diphosphate synthase. Catalytic ranges of known E-isoprenyl diphosphate synthases are shown with arrows on the right. Final natural products derived from the linear polyprenyl diphosphates are shown on the left. Abbreviations: DPP, decaprenyl diphosphate; FPP, farnesyl diphosphate; GFPP, geranylfarnesyl diphosphate; GGPP, geranylgeranyl diphosphate; GPP, geranyl diphosphate; HepPP, heptaprenyldiphosphate; HexPP, hexaprenyl diphosphate; IPP, isopentenyl diphosphate; OPP, octaprenyl diphosphate; SPP, solanesyl diphosphate.

TIBS 24 – NOVEMBER 1999
conformation, determined by the sur-rounding layers. In addition, protein far-nesyltransferase, which activates pro-teins such as Ras via addition of afarnesyl residue to the cysteine residueof C-terminal CAAx motifs, also has a similar structural fold formed by ahelices (Fig. 3d)16. Taken together, the 3Ddata lend support to possible evolution-ary relationships between some chainelongation and cyclase enzymes.
Conservation between E- and Z-isoprenyldiphosphate synthases?
Recently, Shimizu et al. cloned thegene for undecaprenyl diphosphate synthase (UPP), thereby providing thefirst identification and purification of aZ-isoprenyl diphosphate synthase2. Theprimary structure of UPP synthase iscompletely different from those of E-isoprenyl diphosphate synthases and,even though a DDxxD motif-like struc-ture is present in the enzyme, it seemspremature to designate it as an equallycrucial region corresponding to that ofthe E-isoprenyl diphosphate synthases.It would be interesting to examine similarities and differences in the 3Dstructures of these two types of prenyl-transferases and to explore whether the Z-isoprenyl diphosphate synthasefamily evolved via comparable molecu-lar mechanisms. The remainder of thisreview will focus on the more exten-sively studied E-isoprenyl diphosphatesynthases.
REVIEWS
Mechanism of chain-length determination byE-isoprenyl diphosphate synthases
Some of the most interesting researchinto the catalytic mechanism of prenyl-transferases is aimed at understanding
the mechanism by which individual enzymes recognize chain lengths of sub-strates or products, or both. As men-tioned previously, despite identical con-densation mechanisms, these enzymes
Figure 2Structure of E-isoprenyl diphosphate synthase. (a) Conserved primary structure of E-isoprenyl diphosphate synthases and amino acid com-parisons of the chain-length determination (CLD) region. E-isoprenyl diphosphate synthases have seven conserved domains including two aspartate-rich motifs, the first aspartate-rich motif (FARM) and the second aspartate-rich motif (SARM). In the sequence alignment, con-served aspartic acids, the fourth and the fifth amino acids before the FARM motif, two amino-acid insert in the FARM motif are indicated inred, blue and orange, respectively. Glutamic acid in the FARM motif of yeast or human GGPP synthases is indicated in green. (b) 3D structureof avian FPP synthase mutant (F112A/F113S) bound to its substrate FPP (Ref. 11). Two side chains of amino acids at the fourth (S113) andthe fifth (A112) positions before the FARM motif interact directly with the v-terminal of FPP. The second SARM motif seems to bind IPP.Abbreviations: FPP, farnesyl diphosphate; GGPP, geranylgeranyl diphosphate; HexPP, hexaprenyl diphosphate; OPP, octaprenyl diphosphate.
Figure 33D structures of proteins that utilize E-isoprenyl diphosphates. (a) Squalene-hopane cy-clase13, (b) pentalenene synthase15, (c) 5-epi-aristolochene synthase14 and (d) protein farne-syltransferase16, which all use farnesyl diphosphate (FPP) or its derivative as substrate, have3D structures similar to avian FPP synthase. The terpenoid-synthase fold is formed solely by ahelices.
447

REVIEWS TIBS 24 – NOVEMBER 1999
448
rarely catalyze the formation of productsgreater than a pre-determined lengthspecific to each prenyltransferase. Forexample, FPP synthase produces a C15compound and hardly any C20. A differentenzyme, GGPP synthase, manages theproduction of C20. However, even thoughthe original crystal structure of avianFPP synthase and mutagenesis studiesof conserved amino acids contributedvaluable insights into the condensationreaction, these experiments did not pro-vide answers to the question of chain-length specificity5–9. Further randomchemical mutagenesis and X-ray crystal-lography of enzymes with their sub-strates were breakthrough experimentsthat identified specific amino acid resi-dues that are essential for chain-lengthdetermination of the ultimate product.
In 1996, Tarshis et al. performed X-rayanalyses of the structure of mutatedavian FPP synthase in the presence andabsence of bound allylic substrates11.Combined analyses of normal and mu-tated avian FPP synthases led to the sug-gestion that the ultimate length of thepolyisoprenoid chain is governed by thesize of a hydrophobic pocket in the inte-rior of the enzyme that binds the growinghydrocarbon chain. Two benzyl groupsin Phe112 and Phe113, located directlyN-terminal to the first DDxxD sequence,form the floor of the pocket and replace-ment of these aromatic rings withsmaller side chains led to the formationof longer chain products (Fig. 2b).
Tackling the same problem from a different angle, Ohnuma et al. carried outrandom mutagenesis experiments aimedat altering the chain-length selectivity ofprenyltransferases, using the genes encoding FPP and GGPP synthases from Bacillus stearothermophilus andSulfolobus acidocaldarius, respectively, asa representative model17,18. In combi-nation with an in vivo selection methodthat utilizes carotenoid biosyntheticgenes as reporter genes, FPP synthaseswere successfully converted to GGPPsynthase17. Furthermore, the chainlength of the final product is dramaticallychanged by replacement of only oneamino acid, Tyr81 of FPP synthase, situ-ated at the fifth amino acid N-terminal tothe first aspartate-rich motif (FARM).Assuming that the essential structure ofthe B. stearothermophilus FPP synthasebinding cavity is similar to that of theavian FPP synthase, Tyr81 of B. stearo-thermophilus FPP synthase is probablysituated at ~11–12 Å from the FARMmotif, and is thought to bind the diphos-phate moiety of an allylic substrate via
magnesium ions5,6,19, in the large centralcavity of the FPP synthase. Comparisonof 19 different mutated FPP synthaseswith the wild-type enzyme, each with adifferent amino acid substitution at pos-ition 81, suggests that the average chainlength of a final product is inversely pro-portional to the accessible surface areaof the substituted amino acid20.
In another study of chain-length se-lectivity, the authors used a mutantyeast strain with a defect in hexaprenyldiphosphate synthase to isolate mu-tated GGPP synthases capable of syn-thesizing geranylfarnesyl diphosphate(C25) and hexaprenyl diphosphate (C30)from a library of randomly mutatedGGPP synthases18. Sequence analysis ofisolated mutants revealed that the mu-tations were located at the fifth aminoacid N-terminal to the FARM motif,which corresponds to the same substi-tuted position in the FPP synthases thatdisplayed GGPP synthase activity, described above. Presumably, this was aresult of deepening of the bindingpocket, as in the case of the avian mu-tants reported by Tarshis et al.11 Takentogether, these experiments indicatethat FPP synthases from B. stearo-thermophilus and GGPP synthase from S. acidocaldarius all use an amino acid ata fixed position five residues N-terminalto the FARM motif to limit productlength. The next logical question is then,what difference(s) gives rise to the dis-tinct product specificities between FPPsynthase and GGPP synthase?
Ohnuma et al. discovered that allchimeric GGPP synthases with approxi-mately ten amino acids around the FARMmotif, when replaced with those of FPPsynthases from several different organ-isms, showed FPP synthase activity,which indicates that the region aroundthe FARM determines the product speci-ficity of FPP synthases and GGPP syn-thases21. More precise mutational analy-ses suggested that, depending on theorganism, two different mechanisms de-termine the product specificity of FPPsynthases. For eukaryotes, both of thetwo aromatic amino acids at the fifth andthe fourth positions N-terminal to theFARM motif in FPP synthase directlyblock further condensation of com-pounds greater than C15 and provide theproduct specificity, whereas insertions oftwo amino acids within the FARM motifconfer product specificity to bacterialFPP synthases21. The two amino acid in-sertions might act by changing the struc-ture of the FARM motif. Independently,Tarshis et al. also suggested the possible
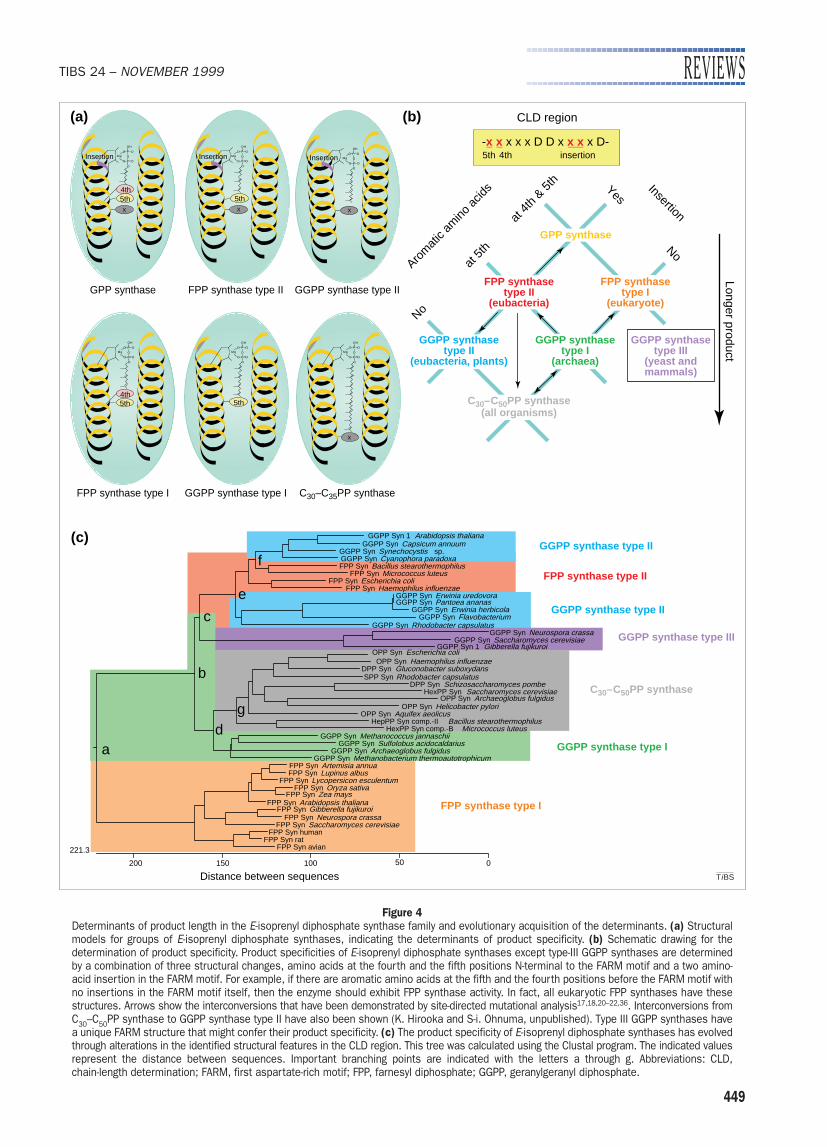
significance of the fourth amino acid N-terminal to the FARM motif and its in-fluence on the size of the active-sitepocket11. Moreover, a mutated isoprenyldiphosphate synthase with both charac-teristics (i.e. aromatic amino acids at thefourth and the fifth positions and thetwo-amino-acid insertion) exclusivelyproduces GPP, a C10 compound, whichsuggests that the two structures act inde-pendently to shorten product length22.On the basis of these results, it is pro-posed that the region from the fifth pos-ition N-terminal to the FARM motif to theend of the FARM sequence determinesthe final products of all E-isoprenyldiphosphate synthases and can thus bedesignated as the chain-length determi-nation (CLD) region (Fig. 2a). A series ofmutational analyses has revealed thatsimple combinations of characteristicamino acids in the CLD region conferproduct specificities to E-isoprenyldiphosphate synthases (Fig. 4a,b).Moreover, as shown below, this intercon-version of product specificities mighthave been employed during evolution.
Creating evolutionary diversity fromchanges in CLD composition
The idea that E-isoprenyl diphosphatesynthases were produced by divergentevolution was first proposed in 1994 byChen et al., who compared amino acidsequences of 13 distinct enzymes in thefamily that were available at the time1.One of the main conclusions from thestudy was that Methanobacterium ther-moautotrophicum GGPP synthase, the bi-functional archaeal enzyme that pro-duces FPP as an intermediate23, is anancient ‘pre-enzyme’ and that the earli-est branch in isoprenyl diphosphate syn-thase evolution was a functional segre-gation that separated the long-chainfrom the short-chain synthases, as illus-trated by the divergence between GGPPsynthase and yeast hexaprenyl diphos-phate synthase. Fine-tuning of chain-length control then evolved indepen-dently after the bifunctional GGPPsynthase segregated into Archaea,Bacteria and Eukarya. This idea is alsoconsistent with the hypothesis ofReardon and Farber that enzymes partici-pating in sequential steps of a metabolicpathway evolve one from another24.
An updated phylogenetic tree con-structed from more than 50 of the E-iso-prenyl diphosphate synthases currentlyavailable in the GenBank database isshown in Fig. 4c. Almost all organismshave three kinds of polyprenyl diphos-phate synthases: FPP synthase, GGPP
TIBS 24 – NOVEMBER 1999
449
REVIEWS
0
221.3
50100150200
GGPP Syn 1
GGPP synthase type II
GGPP synthase type II
GGPP synthase type III
GGPP synthase type I
C30–C50PP synthase
FPP synthase type II
FPP synthase type I
Arabidopsis thalianaGGPP Syn Capsicum annuum
GGPP Syn Synechocystis sp.GGPP Syn Cyanophora paradoxaFPP Syn Bacillus stearothermophilus
FPP Syn Micrococcus luteusFPP Syn Escherichia coli
FPP Syn Haemophilus influenzaeGGPP Syn Erwinia uredovoraGGPP Syn Pantoea ananas
GGPP Syn Erwinia herbicolaGGPP Syn Flavobacterium
GGPP Syn Rhodobacter capsulatusGGPP Syn Neurospora crassa
GGPP Syn Saccharomyces cerevisiaeGGPP Syn 1 Gibberella fujikuroi
OPP Syn Escherichia coliOPP Syn Haemophilus influenzae
DPP Syn Gluconobacter suboxydansSPP Syn Rhodobacter capsulatus
DPP Syn Schizosaccharomyces pombeHexPP Syn Saccharomyces cerevisiae
OPP Syn Archaeoglobus fulgidusOPP Syn Helicobacter pylori
OPP Syn Aquifex aeolicusHepPP Syn comp.-II Bacillus stearothermophilus
HexPP Syn comp.-B Micrococcus luteusGGPP Syn Methanococcus jannaschii
GGPP Syn Sulfolobus acidocaldariusGGPP Syn Archaeoglobus fulgidus
GGPP Syn Methanobacterium thermoautotrophicumFPP Syn Artemisia annuaFPP Syn Lupinus albus
FPP Syn Lycopersicon esculentumFPP Syn Oryza sativa
FPP Syn Zea maysFPP Syn Arabidopsis thaliana
FPP Syn Gibberella fujikuroiFPP Syn Neurospora crassa
FPP Syn Saccharomyces cerevisiaeFPP Syn human
FPP Syn ratFPP Syn avian
a
b
c
d
e
f
g
Distance between sequences
(c)
Mg
O
O
P
P
O
OO
OH
O
4th5thX
Mg
O
O
P
P
O
OO
OH
O
5th
Mg
O
O
P
P
O
OO
OH
O
X
Mg
O
O
P
P
O
OO
OH
O
5th4th
Mg
O
O
P
P
O
OO
OH
O
5thX X
Mg
O
O
P
P
O
OO
OH
O
GPP synthase FPP synthase type II GGPP synthase type II
FPP synthase type I GGPP synthase type I C30–C35PP synthase
Insertion
(a)
Insertion Insertion
-x x x x x D D x x x x D-
Longer product
Arom
atic
amino
acid
s Insertion
at 5
th
at 4
th &
5th
No
Yes
No
CLD region
GPP synthase
FPP synthasetype I
(eukaryote)
FPP synthasetype II
(eubacteria)
C30–C50PP synthase(all organisms)
GGPP synthasetype II
(eubacteria, plants)
GGPP synthasetype I
(archaea)
GGPP synthasetype III
(yeast andmammals)
4th5th insertion
(b)
Ti BS
Figure 4Determinants of product length in the E-isoprenyl diphosphate synthase family and evolutionary acquisition of the determinants. (a) Structuralmodels for groups of E-isoprenyl diphosphate synthases, indicating the determinants of product specificity. (b) Schematic drawing for the determination of product specificity. Product specificities of E-isoprenyl diphosphate synthases except type-III GGPP synthases are determinedby a combination of three structural changes, amino acids at the fourth and the fifth positions N-terminal to the FARM motif and a two amino-acid insertion in the FARM motif. For example, if there are aromatic amino acids at the fifth and the fourth positions before the FARM motif withno insertions in the FARM motif itself, then the enzyme should exhibit FPP synthase activity. In fact, all eukaryotic FPP synthases have thesestructures. Arrows show the interconversions that have been demonstrated by site-directed mutational analysis17,18,20–22,36. Interconversions fromC30–C50PP synthase to GGPP synthase type II have also been shown (K. Hirooka and S-i. Ohnuma, unpublished). Type III GGPP synthases havea unique FARM structure that might confer their product specificity. (c) The product specificity of E-isoprenyl diphosphate synthases has evolvedthrough alterations in the identified structural features in the CLD region. This tree was calculated using the Clustal program. The indicated values represent the distance between sequences. Important branching points are indicated with the letters a through g. Abbreviations: CLD, chain-length determination; FARM, first aspartate-rich motif; FPP, farnesyl diphosphate; GGPP, geranylgeranyl diphosphate.

REVIEWS TIBS 24 – NOVEMBER 1999
450
synthase and long-chain polyprenyldiphosphate synthase. A comparison ofthe amino acid sequences of E-isoprenyldiphosphate synthases, specificallyaround the conserved FARM motif, showsa common profile. All FPP synthases andarchaeal GGPP synthases possess anaromatic amino acid (phenylalanine ortyrosine) five residues upstream of theFARM motif. In contrast, E-isoprenyldiphosphate synthases that catalyzecondensations beyond GGPP (C20) andGGPP synthases from organisms otherthan Archaea have a nonaromatic aminoacid at the corresponding position. Forexample, 11 E-isoprenyl diphosphate syn-thases that catalyze consecutive conden-sations beyond GGPP have been cloned,including hexaprenyl diphosphate syn-thase from Saccharomyces cerevisiae25,heptaprenyl diphosphate synthase fromB. stearothermophilus26, octaprenyldiphosphate synthase from E. coli27,solanesyl diphosphate synthase from R. capsulatus28 and decaprenyl diphos-phate synthase from S. pombe29. Theseenzymes all have an alanine residue atposition five N-terminal to the FARMmotif. Studies by Ohnuma et al.17,18 withB. stearothermophilus FPP synthase andS. acidocaldarius GGPP synthase havealso shown that substitution of the aro-matic amino acid at the position five N-terminal to FARM brings about the con-versions of FPP synthase to GGPP syn-thase and of GGPP synthase to C30–C50synthase. Moreover, in heptaprenyldiphosphate synthase from B. stearother-mophilus, replacement of alanine at thisposition with aromatic amino acids re-sulted in conversion in the reverse direc-tion to GGPP synthase. These data sup-port the idea that this amino acid plays avital role in determining whether or notthe enzymes can catalyze the formationof products longer than GGPP. Ex-trapolating the data a step further, onecould argue that the fifth amino acid N-terminal to the FARM motif is biologi-cally significant for the determination ofchain length in the ultimate product and,consequently, of the enzyme responsiblefor its synthesis. The change betweenalanine and aromatic amino acids at thisposition has occurred during evolutionat branch d as shown in Fig. 4c.
How has fine-tuning of chain-lengthcontrol taken place during evolution?For instance, how did FPP synthasesevolve and what changes gave these enzymes the ability to synthesize FPPbut not GGPP? Upon further exami-nation of the chain-elongation modelthat was proposed on the basis of mu-
tagenesis results, and taking into ac-count the suggested mechanism byPoulter’s group from X-ray crystallogra-phy, one can piece together a workingmodel (Fig. 4a,b) that explains the observed evolutionary relationships.
Replacement of small amino acids atpositions four and five N-terminal to theFARM motif with aromatic amino acids orinsertion of two amino acids (proline andserine) in the FARM motif of archaealGGPP synthase caused changes in cata-lytic activity, which led to its conversioninto FPP synthase. FPP synthases can beclassified into two types according totheir sequences in the CLD region and thestructural requirements for chain-lengthdetermination in eukaryotic FPP synthase(type I) are different from those inprokaryotic FPP synthase (type II)21.These findings suggest that FPP syn-thases have evolved from the same an-cestral origin in two ways: if archaealGGPP synthase is the ‘pre-enzyme’, type-IFPP synthase activity would have beenconferred by a substitution at the fourthamino acid N-terminal to the FARM motif(branch a in Fig. 4c). For example, boththe avian and human FPP synthases differfrom the isoprenyl diphosphate synthase(idsA) of M. thermoautotrophicum in thatthey have an aromatic amino acid(phenylalanine) at the fourth position N-terminal to the FARM motif and havepresumably evolved with other eukary-otic FPP synthases. For type-II FPP syn-thases, insertion of two amino acidresidues in the FARM motif (branch c)and other, yet unknown modificationsprobably occurred to fine-tune their prod-uct specificity. The presence of a two-amino-acid insertion within FARM and thelack of change (compared with the ‘pre-enzyme’) at the fourth position N-termi-nal to the FARM motif in E. coli and B. stearothermophilus FPP synthases sub-stantiate this hypothesis (Fig. 2a, 4a,b).
Extending the idea even further, if evo-lution starts from type-II FPP synthase,keeping intact the two amino acid inser-tions but changing the fifth amino acidbefore the FARM to a non-aromatic one,the enzyme would acquire new catalyticactivity resulting in the formation of type-II GGPP synthase (branch e and f). Bothplant and bacterial GGPP synthases illus-trate this point well: they have amino acidinsertions and non-aromatic (alanine)residues (Fig. 2a and 4a). Branch f alsoindicates that plant GGPP synthases werenot derived from plant FPP synthasesbut from bacterial FPP synthases (type-IIFPP synthase), probably through a sym-biotic relationship with bacteria30. Clearly,
the three important structural featuresfor chain-length determination have alsoplayed significant roles in evolution.
One exception to this structural deter-mination of product specificity model isthe GGPP synthases from yeast andhuman (type-III GGPP synthase)31,32 rep-resented by branch c in the phylogenetictree (Fig. 4c). All known members of thissubgroup have an acidic amino acid, glu-tamic acid, in the FARM motif (Fig. 2a),which could change the structure of theCLD region and serve as a fourth deter-minant of product specificity. It is note-worthy that glutamic acid might also af-fect substrate specificity as these type-IIIGGPP synthase enzymes cannot acceptdimethylallyl diphosphate (DMAPP) astheir substrate33.
The evolution of E-isoprenyl diphos-phate synthases is described here fromthe viewpoint of a relationship betweenproduct specificity and the structure ofCLD regions. However, the evolutionarycourse for each member of the C30–C50PPsynthase group and its fine-tuning mecha-nisms for product specificity are notknown. It has been reported that thisgroup can be divided into two subgroupsaccording to the quaternary structure.For example, heptaprenyl diphosphatesynthase from B. stearothermophilus andhexaprenyl diphosphate synthase fromM. luteus B-P 26 are constructed with twodifferent subunits encoded in a singleoperon2,26,34. These heteromeric structuresare essential for the enzymes’ proper activities. In contrast, octaprenyl diphos-phate synthase from E. coli and solanesyldiphosphate synthase from M. luteushave homodimer structures much likethose of FPP and GGPP synthases35. Thisdifference in subunit composition musthave occurred around branch g in Fig. 4c.GGPP synthase from S. acidocaldarius andFPP synthase from B. stearothermophiluswith multiple mutations at positions five,eight and 11 N-terminal to the FARM motifcan produce longer products than mu-tants with a single mutation at positionfive36. Moreover, product lengths are de-pendent on the nature of the mutatedamino acids. For example, a mutated en-zyme with double mutations of L74G andF77G produces a C35 product, whereas anenzyme with triple mutations of I71G,L74G and F77G produces a C40 compoundas the main product36. This result sug-gests that product specificity in theC30–C50PP synthase group can be deter-mined around the mutated region.However, there is no known significant re-lationship between the sequences aroundthe region and the product specificities in

TIBS 24 – NOVEMBER 1999
451
this group. Also, members in this branchof C30–C50PP synthase in the evolutionarytree are not organized according to theirproduct specificities (Fig. 4c), therebypointing to the possibility that each mem-ber independently derived its productspecificity following the appearance of itshost organism.
Concluding remarksThis review presents evidence for a re-
lationship between the structural require-ments for product specificities and thephylogenetic tree of E-isoprenyl diphos-phate synthases and suggests a possibleevolutionary course with the assumptionthat archaeal GGPP synthase is closest intime to the common ancestor. However,this might not be a valid assumption tomake. In the future, more precise evolu-tionary analyses need to be integrated,by using a larger number of samples withphylogenetic data that include isoprenoidmetabolic enzymes other than isoprenyldiphosphate synthases, in order to fullyunderstand the evolutionary mechanismof isoprenoid biosynthesis. Researcherswill also need to examine why enzymesthat use linear isoprenyl diphosphatesshare a common basic 3D structure, the ‘terpenoid synthase fold’. Mutationalanalysis of isoprenoid biosynthetic en-zymes might convert the enzymatic activ-ities from polymerization to cyclization or vice versa, providing a more global picture of the evolutionary process.
AcknowledgementsS-i. Ohnuma thanks K. Ogura,
T. Koyama, T. Nishino and H. Sagami for support and encouragement. S-i.Ohnuma also thanks members of theNishino laboratory.
References1 Chen, A., Kroon, P. A. and Poulter, C. D. (1994)
Protein Sci. 3, 600–6072 Shimizu, N., Koyama, T. and Ogura, K. (1998)
J. Biol. Chem. 273, 19476–194813 Ogura, K. and Koyama, T. (1998) Chem. Rev.
98, 1263–12764 Koyama, T. et al. (1994) Acta Biochim. Pol. 41,
281–2925 Joly, A. and Edwards, P. A. (1993) J. Biol. Chem.
268, 26983–269896 Song, L. and Poulter, C. D. (1994) Proc. Natl.
Acad. Sci. U. S. A. 91, 3044–30487 Koyama, T. et al. (1994) Biochemistry 33,
12644–126488 Koyama, T. et al. (1995) Biochem. Biophys.
Res. Commun. 212, 681–6869 Koyama, T. et al. (1996) Biochemistry 35,
9533–953810 Tarshis, L. C. et al. (1994) Biochemistry 33,
10871–1087711 Tarshis, L. C. et al. (1996) Proc. Natl. Acad. Sci.
U. S. A. 93, 15018–1502312 Saito, A. and Rilling, H. C. (1981) Arch.
Biochem. Biophys. 208, 508–51113 Wendt, K. U., Poralla, K. and Schulz, G. E.
(1997) Science 277,1811–1815
14 Starks, C. M. et al.(1997) Science 277,1815–1820
15 Lesburg, C. A. et al.(1997) Science 277,1820–1824
16 Long, S. B., Casey, P. J.and Beese, L. S.(1998) Biochemistry37, 9612–9618
17 Ohnuma, S. et al.(1996) J. Biol. Chem.271, 10087–10095
18 Ohnuma, S. et al.(1996) J. Biol. Chem.271, 18831–18837
19 Marrero, P. F., Poulter, C. D. andEdwards, P. A. (1992)J. Biol. Chem. 267,21873–21878
20 Ohnuma, S. et al.(1996) J. Biol. Chem.271, 30748–30754
21 Ohnuma, S. et al.(1997) J. Biol. Chem.272, 5192–5198
22 Narita, K. et al. J. Biochem. (Tokyo)(in press)
23 Chen, A. and Poulter,C. D. (1993) J. Biol.Chem. 268,11002–11007
24 Reardon, D. andFarber, G. K. (1995)FASEB J. 9, 497–503
25 Ashby, M. N. andEdwards, P. A. (1990)J. Biol. Chem. 265,13157–13164
26 Koike-Takeshita, A. et al. (1995) J. Biol.Chem. 270,18396–18400
27 Asai, K. et al. (1994)Biochem. Biophys.Res. Commun. 202,340–345
28 Okada, K. et al.(1997) J. Bacteriol.179, 5992–5998
29 Suzuki, K. et al.(1997) J. Biochem.(Tokyo) 121,496–505
30 Shih, M. C., Lazar, G.and Goodman, H. M.(1986) Cell 47,73–80
31 Jiang, Y. et al. (1995)J. Biol. Chem. 270,21793–21799
32 Kuzuguchi, T. et al.(1999) J. Biol. Chem.274, 5888–5894
33 Sagami, H., Morita, Y.and Ogura, K. (1994)J. Biol. Chem. 269,20561–20566
34 Zhang, Y. W., Koyama,T. and Ogura, K.(1997) J. Bacteriol.179, 1417–1419
35 Ohnuma, S., Koyama,T. and Ogura, K.(1991) J. Biol. Chem.266, 23706–23713
36 Ohnuma, S. et al.(1998) J. Biol. Chem.273, 26705–26713
REVIEWS
Pete Jeffs is a freelancer working in Paris, France.




















