
Control of Surface Properties of Hyaluronan/Chitosan ... - MDPI
Upload
independentCategory
view
1download
0
PATTERNS & PHENOTYPES
Three Vertebrate Hyaluronan Synthases AreExpressed During Mouse Development inDistinct Spatial and Temporal PatternsJanet Y.L. Tien1* and Andrew P. Spicer2
We have used in situ hybridization to study the expression of the vertebrate hyaluronan synthase (Has)gene family members, designated Has1, Has2, and Has3, during mouse development. At embryonic day (E)7.5, Has1 and Has2 are expressed throughout the gastrulating embryo. After E8.5, Has1 expressiondisappears, but Has2 continues to be strongly, albeit transiently, expressed in numerous tissues, includingthe branchial arches and craniofacial structures such as the palatal shelves and lens pit. Has2 is alsoexpressed during cardiac, skeletal, and tail development. Has3 transcripts are first detected at E10.5 in themaxillary and mandibular components of the first branchial arch. Notably, Has3 expression in thedeveloping teeth, vibrissae hair follicles, nasal cavity, and inner ear complements the expression pattern ofHas2. Our results indicate that, whereas Has2 is exclusively expressed in some tissues, its expressionpattern overlaps and/or complements that of Has1 and Has3 in others. Developmental Dynamics 233:130–141, 2005. © 2005 Wiley-Liss, Inc.
Key words: Hyaluronan synthase; HAS; expression; development; craniofacial; branchial arches; ectodermal placode;vibrissae follicle; tooth; inner ear
Received 20 April 2004; Revised 28 July 2004; Accepted 17 August 2004
INTRODUCTION
Hyaluronan (HA) is an extracellularmatrix glycosaminoglycan that isimportant during development andmorphogenesis (Toole, 2001; Spicerand Tien, 2004). Initially discoveredin the vitreous humor of the eye, thislinear polymer of repeating units ofD-glucuronic acid and N-acetylglu-cosamine can be found in virtuallyevery tissue and body fluid of verte-brates (Laurent, 1998). During em-bryogenesis, the extracellular andpericellular matrix surrounding mi-grating and proliferating cells is rich
in HA. This localized concentrationof HA decreases in differentiatingcells. HA is localized around migrat-ing neural crest cells and cushioncells of the heart, as well as in theelevating secondary palatal shelf,corneal stroma, semicircular canals,and limb buds. The critical role ofHA during development is furtherevidenced by the morphogenetic ab-normalities that resulted from re-moval of HA or inhibition of HAbinding to its associated proteins invivo or in organ cultures (Hay, 1980;Delpech et al., 1989; Haddon and
Lewis, 1991; Knudson et al., 1995;Toole, 1997; Camenisch et al., 2000).
Although simple in structure, HAis versatile in form and function. De-pending on its size, which can typi-cally vary between 200 and 4,000kDa, and concentration, HA hasdistinct effects on cell behavior. Ingeneral, lower concentrations of HAare able to promote aggregation,whereas higher concentrations havethe reverse effect, separating cellsfrom one another or from its substra-tum (Underhill and Toole, 1981;Green et al., 1988; Toole, 1991). As a
1Department of Cell Biology and Human Anatomy, University of California, School of Medicine, Davis, California2Center for Extracellular Matrix Biology, Institute of Biosciences and Technology, Texas A&M University System Health Science Center,Houston, TexasGrant sponsor: National Institutes of Health; Grant number: AR47433.*Correspondence to: Janet Y.L. Tien, Ph.D., Department of Cell Biology and Human Anatomy, University of California, Schoolof Medicine, 1 Shields Avenue, Davis, CA 95616. E-mail: [email protected]
DOI 10.1002/dvdy.20328Published online 11 March 2005 in Wiley InterScience (www.interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 233:130–141, 2005
© 2005 Wiley-Liss, Inc.

molecule that closely associates withcells and has the ability to bind tomultiple extracellular proteins, HAis an effective support structure inthe extracellular matrix, particu-larly in supporting tissues of verte-brates, including cartilage, skin,smooth muscle, and other connectivetissues (Spicer and Tien, 2004). As asignaling molecule, HA is able to ac-tivate several pathways throughbinding of HA receptors such asCD44 and receptor for hyaluronan-mediated motility (RHAMM; Bour-guignon et al., 2000; Lee and Spicer,2000; Lokeshwar and Selzer, 2000;Oliferenko et al., 2000; Slevin et al.,2002). There is recent evidence for arole of intracellular HA, possibly asa regulator of cell division (Evankoand Wight, 1999). With the range ofeffects HA can have on cell behavior,it is not surprising that abnormallevels of HA are often associatedwith pathological states such as os-teoarthritis, rheumatoid arthritis(Engstrom-Laurent, 1997), pulmo-nary fibrosis, as well as a variety ofmalignant cancers, including lung,mammary, and hepatic carcinoma(Knudson et al., 1989; Delpech et al.,1997). HA accumulation contributesto thyroid-associated ophthalmopa-thy, which is associated with Graves’disease (a thyroid glandular disease)and involves remodeling of the or-bital tissue (Cao et al., 1998). Tohelp understand the roles of HA inmorphogenesis in normal develop-ment, growth, and disease, we haveturned our attention to the enzymesresponsible for its biosynthesis: thehyaluronan synthases.
Hyaluronan synthases (Has) are afamily of plasma membrane glyco-syltransferases that synthesizehyaluronan (DeAngelis, 1999).Three highly conserved Has geneshave been identified in human,mouse, Xenopus, and zebrafish.Studies on HA synthases havehelped to illuminate the multiplefunctions of HA during morphogene-sis. Biochemical analyses revealedthat the three encoded polypeptideshave differential stability (half-life)and enzymatic activities. Overall,Has3 is most stable, Has2 is less,and Has1 is the least stable (Itano etal., 1999). In terms of in vitro enzy-matic activity, Has3 is able to poly-
merize low molecular mass HA (�2 � 105 Da) rather than mostly highmolecular mass HA (� 1 � 106 Da)as observed for Has1 and Has2(Spicer and McDonald, 1998). HAsynthesis can be regulated by manygrowth factors and cytokines, includ-ing transforming growth factor-beta/bone morphogenetic protein super-family members, interleukin-1 beta,tumor necrosis factor-alpha, epider-mal growth factor, and keratinocytegrowth factor, depending on the celltypes (Spicer and Tien, 2004). Regu-lation of HA synthesis by means ofthe HA synthases differs betweencell types in part because given celltypes express distinct profiles of oneor more Has genes.
Northern blot analyses haveshown that in the adult human andmouse, Has1 and Has3 are ex-pressed at higher levels and in moretissues than Has2. Conversely, inthe embryo, Has2 is widely ex-pressed at high levels throughoutgestation, whereas Has1 and Has3have restricted expression at earlyand later stages, respectively (Spicerand McDonald, 1998). Loss-of-func-tion studies in the mouse establishedthat Has2-dependent HA synthesisis critical for normal embryonic de-velopment (Camenisch et al., 2000).Of interest, Has1 and Has3 homozy-gous null animals are viable andfertile (unpublished data; Bai et al.,manuscript submitted for publica-tion). Thus, Has2 is the major HAsynthase during embryogenesis.From the targeted inactivation ofHas2, we now know that HA is re-quired for the establishment and/ormaintenance of extracellular matrix-defined spaces and that HA has acritical role in the epithelial–mesen-chymal transformation of endocar-dial cushion cells.
To understand further the func-tions of HA and the differences be-tween the three HA synthases, wechose to elucidate the spatial andtemporal distribution of Has genetranscripts during mouse develop-ment using in situ hybridization.The results of these studies revealimportant relationships between HAsynthases and suggest novel rolesfor HA during the development ofcertain structures.
RESULTS AND DISCUSSION
Has1 and Has2 AreExpressed DuringGastrulation
Shortly after implantation, the earlyembryo will rapidly undergo gastrula-tion to establish three primary germlayers. In this process, epiblast cellsfrom the inner cell mass will migratein streams to form the embryonic ec-toderm, mesoderm and endoderm.This migration may be hyaluronan-dependent (Solursh and Morriss,1977; Gilbert, 1997; Mullegger andLepperdinger, 2002; Bakkers et al.,2004). We asked which HA synthasewas responsible for the production ofhyaluronan during gastrulation in themouse. Using in situ hybridizationwith digoxigenin-labeled probes forHas1, Has2, and Has3 on whole-mounts, we observed that Has1 andHas2, but not Has3, were expressed ator around embryonic day (E) 7.5 (Fig.1A–C). At this stage, Has1 and Has2are expressed in the embryo and ex-traembryonic membranes, althoughHas1 appears to be more evenly dis-tributed than Has2. To better visual-ize the expression patterns, we hy-bridized radioactive RNA probes on12-micron frozen sections. Has1 andHas2 are both expressed in the allan-tois, amnion, and yolk sac, as well asthe neural ectoderm of the embryonichead folds and throughout the ce-phalic mesenchyme (Fig. 1D,E). Has2transcripts appear to be concentratedat the location of the prospective peri-cardial cavity (Fig. 1E, arrows). Inmore posterior sections than thoseshown in Figure 1D,E, Has1 and Has2continue to be expressed in the em-bryo proper, including the embryonicmesoderm and possibly the primitivestreak (Fig. 1G,H). In situ hybridiza-tion using a Has3 or sense probe didnot produce specific signal (Fig.1C,F,I).
Has2 Is Expressed inMultiple Tissues, IncludingCraniofacial and CardiacStructures
In E8.5 and older embryos, Has1 ex-pression is diminished (Fig. 1J). How-ever, Has2 expression was evident atall stages examined in a variety of
HYALURONAN SYNTHASE EXPRESSION 131

Fig. 1. Expression of hyaluronan synthase (Has) 1 and Has2 during mouse development. In situ hybridization on whole-mount specimens (A–C, J–Q)and cryosections (D–I), counterstained with Hoechst (E�, H�). A–C: E7.25–E7.5 mice hybridized with Has1, Has2, and Has3 digoxigenin (DIG) -labeledriboprobes. D,E,G,H: Radiolabeled Has1 (D,G) and Has2 (E,H) probes used on embryonic day (E) 7.75 transverse cryosections. F,I: A sense probe wasused on adjacent sections. J,K: E8.5 mice hybridized with Has1 (J) and Has2 (K) DIG-probes. L,K: L is a dorsal view of K. M: Branchial arches arenumbered 1-4/6. Inset in M is an anterior view of the E9.5 tail. N: E10.5 heart from the embryo’s right side (Ntop inset); anterior view of the E10.5 limb(bottom inset). Asterisks mark distal Has2 expression. O–Q: Has2 expression in craniofacial regions at E11.5 and E12.5. Q is a frontal view of P.Specific regions are labeled as follows: amnion (am), endocardial cushions (ec), limb bud (lb), lens pit (le), cephalic mesenchyme (me), mandible (mn),maxilla (mx), nasal process (np), neural groove (ng), neuroepithelium (ne), outflow tract (ot), future pericardial cavity (pc), posterior neuropore (pn),somites (so), ventricle (v), yolk sac (yo). Scale bars � 100 �m in C (applies to A–C), H� (applies to D–I), L (applies to J–L); 500 �m in M.
132 TIEN AND SPICER

tissues. In the E8.5 mouse, Has2 isexpressed in the areas associated withmigration of neural crest cells duringbranchial arch formation (Fig. 1K).Dorsally, the expression pattern canbe discerned as an upside-down “V”,beginning above the heart in the neu-ral groove region and ending on eitherside of the heart (Fig. 1L). At E9.5,Has2 is expressed in all of thebranchial arches and in the area offuture cardiac structure development(Fig. 1M), areas where active cranialneural crest cell migration is known tooccur (Gilbert, 1997; Chai et al., 2000;Zhang et al., 2003). Has2 expressionin the E9.5 heart is also consistentwith our previous results showing ex-pression in the atrial and ventricularendothelium (Camenisch et al., 2000).Additionally, at E9.5, Has2 is ex-pressed in the posterior somites (Fig.1M, so, arrowheads), on either side ofthe posterior neuropore (pn, arrows),and at the tip of the tail (Fig. 1M,white arrow and inset). Has2 expres-sion in the tail persists until at leastE13.5 (data not shown).
At E10.5, Has2 continues to be ex-pressed in the developing heart, nowclearly visible in the endocardial cush-ion cells of the future atrioventricularcanal and in the outflow tract (Fig.1N, top inset). The expression of Has2in the heart is consistent with our pre-vious observations of Has2 transcriptsand HA localization in tissue sections(Camenisch et al., 2000). Cardiac neu-ral crest cells contribute to the forma-tion of the outflow tract and endocar-dial cushion cells (Gilbert, 1997;Hutson and Kirby, 2003) and a highconcentration of HA was thought toprovide only the physical driving forcebehind expansion of the cushions. Wenow know that HA, synthesized byHas2, also plays a role in the Ras-dependent signaling pathway in-volved in instructing the transforma-tion of cardiac endothelial cells intomesenchyme (Camenisch et al., 2000).
Has2 is strongly expressed in thenasal processes at E10.5 (Fig. 1N),where HA has been implicated in themolding of the nasal folds (Burk,1985). Has2 is also expressed in mul-tiple locations in and around the man-dibular component of the firstbranchial arch and in the invaginat-ing lens pit. It is at this stage that wefirst clearly observe the expression of
Has2 during limb development. In theforelimb bud, Has2 transcripts arepresent medial to the signaling centerknown as the apical ectodermal ridgeand on the dorsal half of the proximallimb bud (Fig. 1N, bottom inset).
We have shown recently that Has2continues to be expressed in the limbin a distinct pattern during chondro-genesis and joint development; prolif-erating, and precondensing mesen-chymal cells of the cartilage anlagenexpress a high level of Has2, but thisexpression is down-regulated immedi-ately before aggregation of the mesen-chymal cells (Lee et al., 2002; manu-script in preparation). Mountingevidence points to HA as a pivotalplayer during specific stages of skele-tal development. Initially, HA is asso-ciated with the migration of sclero-tomal, mesodermal, and neural crestcells that give rise to the axial skele-ton, the limb skeleton, and thebranchial arches, craniofacial bones,and cartilage (Solursh et al., 1979;Knudson and Toole, 1985; Perris etal., 1991). During limb bud outgrowth,a large amount of extracellular HAmay facilitate cell division (Knudson,2003). HA levels rapidly decrease atthe time of condensation when cellsaggregate before the formation of in-dividual cartilage or bone elements.The interaction between HA and a cellsurface receptor, possibly CD44, arerequired for this early adhesive cell–cell interactions of limb bud mesen-chyme (Maleski and Knudson, 1996).During synovial joint development ahigh level of localized HA production,influenced by mechanical stimuli, maymediate joint cavitation to form a cell-free space (Craig et al., 1990; Archeret al., 2003; Pitsillides, 2003). We areworking currently to elucidate furtherthe potential roles and regulation ofHA synthesis by Has2 during carti-lage and joint development.
At E11.5, Has2 expression in thenasal processes and the maxillaryand mandibular components of thefirst branchial arch continues to bevisible, but in an increasingly nar-row pattern (Fig. 1O). Noticeably,the otic vesicle begins to expressHas2 (Fig. 1O). In particular, Has2transcripts appear to be localizedto the lateral wall region of the oticvesicle (data not shown). Accordingto fate mapping experiments in
mice, the lateral wall will eventuallycontribute to the development ofsemicircular canals. At E12.5, Has2expression during craniofacial devel-opment is apparent in and aroundthe vibrissae (whisker) hair follicles,including those above the orbit of theeye (Fig. 1P). Has2 transcripts arealso abundant around the entranceto the nasal cavity (Fig. 1Q).
Has2 Is Expressed in theDeveloping Eye
We show that Has2 is the major HAsynthase that is expressed during eyedevelopment. Within the invaginatinglens placode, or lens pit, at E10.5,Has2 is expressed at high levels (Fig.2A). Has2 transcripts are also local-ized to the cranial mesenchyme me-dial to the outer layer of the optic cup(future pigmented layer of the retina)and surrounding the forming opticstalk. When the lens vesicle hasformed by E11.5, Has2 expression dis-appears from the lens (Fig. 2B). Theexpression of Has2 medial to the outerlayer of the optic cup persists, at firstin the area of mesenchymal condensa-tion, then at E13.5 in a layer juxta-posed to the pigmented retinal layer(Fig. 2C). The layer of Has2-express-ing cells extends toward and adjacentto the future cornea at the lip of theretinal layers. Has2 is also expressedin the ectodermal protrusions of thefuture eyelids.
Corneal development has longbeen a model example for the space-creating role of hyaluronan (Hay,1980). At the junction between thelens and retina, capillary endothelialcells migrate under the corneal epi-thelium and secrete hyaluronan,causing the stroma to swell. Thisprocess creates a cell-free substra-tum for neural-crest– derived mesen-chymal cells to migrate. Once the fu-ture corneal cells have entered, theywill secrete hyaluronidase, causingthe stroma to shrink. Here, we showthat Has2 is responsible for the ini-tial synthesis of HA during eye de-velopment (Fig. 2). We followed theexpression of Has2 in the otic pit andthen later as it became more region-alized in and around the developingeye.
HYALURONAN SYNTHASE EXPRESSION 133

Has3 Is Expressed DuringTooth and VibrissaDevelopment
In the whole-mount analyses, Has3transcripts did not make an appear-
ance until E10.5. Has3 expressionfirst appears in an extremely localizedmanner; two discrete spots on themaxillary portion of the first branchialarch and, similarly, in two spots onthe mandibular portion (Fig. 3A and
3B, respectively). From E11.5 toE15.5, the expression of Has3 is local-ized to the developing incisors (notshown). Thus, the spots of expressionin the E10.5 mouse most likely corre-spond to areas of ectodermal conden-
Fig. 2. A–C�: Expression of hyaluronan synthase (Has) 2 in the developing eye using in situ hybridization on cryosections (A–C), counterstained withHoechst nuclear stain (A�–C�). Frontal sections through the eye region of E10.5 (A), E11.5 (B), and E13.5 (C) were hybridized with a radiolabeled Has2probe. Prominent expression of Has2 is labeled with arrows and arrowheads in corresponding structures. The asterisk in C is nonspecific signal. Scalebars � 100 �m in A (applies to A,A�), in B (applies to B,B�), in C (applies to C,C�).
134 TIEN AND SPICER

sations that would lead ultimately totooth formation. Below, we describe ingreater detail the expression of Has3during tooth development.
Has3 expression was also detectedin another tissue in the whole-mounts: the developing follicles ofvibrissae and hair. At E11.5, Has3 isexpressed in the earliest developingvibrissae follicles (not shown). Upperlip vibrissa develop along a proximo-distal progression, so that, at a giventime point, hair follicles closer to theeye are more mature than folliclescloser to the nose and mouth. Afterthis temporal and spatial progression,Has3 is expressed during the onset offollicle development, first appearingas superficial round spots closer to theeye, then transiently opening into awreath and quickly disappearing fromthe surface after 24–48 hr as thesefollicles become more mature (Fig.3C,D). At E12.5, Has3 is expressed atthe location corresponding to the fu-
ture hair follicle above the eye (su-praorbital) and the two below the eye(postorbital and postoral; Fig. 3C, ar-rows). Hair follicles of the nose, edgesof the lips and chin, as well as the restof the head, body, and limbs also ex-press Has3 as of E14.5 (Fig. 3E,F, anddata not shown).
Has2 and Has3 AreExpressed During Tooth andVibrissa Hair FollicleDevelopment in aComplementary Pattern
To clarify which tissues were express-ing each HA synthase, we performedin situ hybridization on frozen sec-tions using radiolabeled Has2, Has3,or sense probes. Of interest, we ob-served that Has2 and Has3 were ex-pressed during tooth and vibrissae de-velopment in a complementarypattern. Teeth and hair are ectoder-
mal in origin. Ectodermal organs de-velop similarly, beginning with a con-densation or aggregation of epidermal(ectodermal) cells that thicken to forma placode. This placode increases indepth, while sending molecular in-structions to the underlying mesen-chyme to differentiate. Ultimately,the interacting epidermal and mesen-chymal components will transforminto a complete organ (Pispa andThesleff, 2003).
Mammalian teeth develop from theepithelia of the first branchial archand neural crest-derived mesen-chyme. During early tooth develop-ment, Has3 is first expressed at E10.5(Fig. 3A,B). In sections through theE11.5 day mouse, we show that Has3is expressed in the dental epithelialplacode (Fig. 4B). In contrast, Has2mRNA is undetectable or is present atlow levels in the dental epithelium(Fig. 4A). In E13.5 (not shown), E14.5and E15.5 developing molars, Has2 is
Fig. 3. Expression of hyaluronan synthase (Has) 3 during tooth and hair follicle formation using in situ hybridization on whole-mount specimens atembryonic day (E) 10.5–E15.5. A,B: Inset in B is a cartoon of an E10.5 embryo; the dashed line indicates the plane of dissection used to visualize thebranchial arch components shown in A (maxillary) and B (mandibular). C–F: Has3 expression is detected in the developing vibrissa hair folliclesE12.5–E14.5 (C–E) and hair follicles on the head at E15.5 (F). Arrows indicate specific Has3 expression. Scale bar � 500 �m in B (applies to A,B).
HYALURONAN SYNTHASE EXPRESSION 135
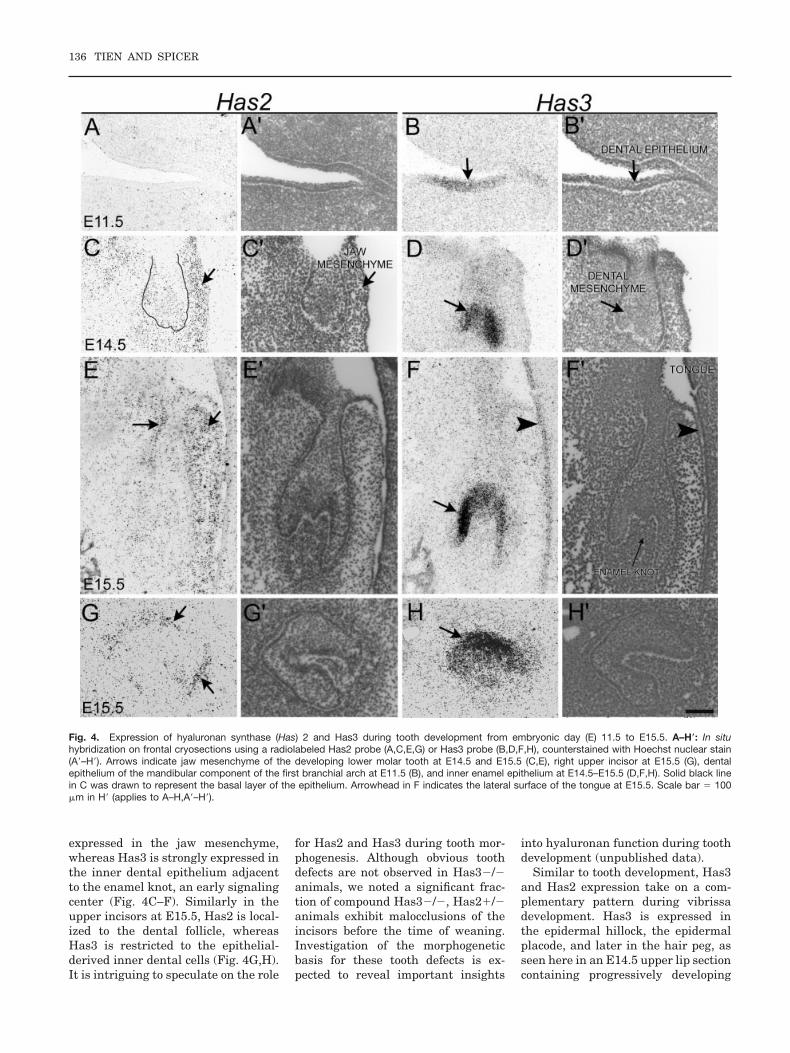
expressed in the jaw mesenchyme,whereas Has3 is strongly expressed inthe inner dental epithelium adjacentto the enamel knot, an early signalingcenter (Fig. 4C–F). Similarly in theupper incisors at E15.5, Has2 is local-ized to the dental follicle, whereasHas3 is restricted to the epithelial-derived inner dental cells (Fig. 4G,H).It is intriguing to speculate on the role
for Has2 and Has3 during tooth mor-phogenesis. Although obvious toothdefects are not observed in Has3�/�animals, we noted a significant frac-tion of compound Has3�/�, Has2�/�animals exhibit malocclusions of theincisors before the time of weaning.Investigation of the morphogeneticbasis for these tooth defects is ex-pected to reveal important insights
into hyaluronan function during toothdevelopment (unpublished data).
Similar to tooth development, Has3and Has2 expression take on a com-plementary pattern during vibrissadevelopment. Has3 is expressed inthe epidermal hillock, the epidermalplacode, and later in the hair peg, asseen here in an E14.5 upper lip sectioncontaining progressively developing
Fig. 4. Expression of hyaluronan synthase (Has) 2 and Has3 during tooth development from embryonic day (E) 11.5 to E15.5. A–H�: In situhybridization on frontal cryosections using a radiolabeled Has2 probe (A,C,E,G) or Has3 probe (B,D,F,H), counterstained with Hoechst nuclear stain(A�–H�). Arrows indicate jaw mesenchyme of the developing lower molar tooth at E14.5 and E15.5 (C,E), right upper incisor at E15.5 (G), dentalepithelium of the mandibular component of the first branchial arch at E11.5 (B), and inner enamel epithelium at E14.5–E15.5 (D,F,H). Solid black linein C was drawn to represent the basal layer of the epithelium. Arrowhead in F indicates the lateral surface of the tongue at E15.5. Scale bar � 100�m in H� (applies to A–H,A�–H�).
136 TIEN AND SPICER

follicles (Fig. 5B). In contrast, Has2 isconspicuously restricted to the under-lying dermal condensation and inter-follicular dermal mesoderm (Fig. 5A).In more mature follicles, Has3 expres-sion becomes restricted to the hairmatrix cells overlying the dermal pa-pilla (Fig. 5D), whereas Has2 tran-scripts are distributed along the de-veloping hair shaft in the dermalsheath area (Fig. 5C).
Has2 and Has3 AreExpressed DuringCraniofacial Development inOverlapping andComplementary Patterns
Has2 and Has3 overlap in expressionin the cartilage primordium of the na-sal capsule and the orbitosphenoid
bone, (Fig. 6A,B) as well as Meckel’scartilage, hyoid, and manubriumsterni (data not shown). On the otherhand, Has2 and Has3 are expressed ina complementary manner in the carti-lage primordium of the nasal septum;Has2 is present in a layer encompass-ing the future septum whereas Has3is in the center of the anlagen (Fig.6A–D). In terms of epithelial expres-sion, Has2 transcripts are present inthe olfactory epithelium and the epi-thelium of the developing vomerona-sal organ (Fig. 6A,C). Has3 tran-scripts appear to be localized to therespiratory epithelium (Fig. 6D). Inthe developing turbinates, Has2 is theonly HA synthase expressed (Fig. 6C).
Has2 only is expressed in the as-cending palatal shelves (Fig. 6A)where, previously, HA has been local-
ized at high concentrations and isthought to be necessary for shelf ele-vation (Pratt et al., 1973). Has2 ex-pression in the palatal shelves laststhrough shelf elevation at E15.5 (notshown). Has3 is strongly expressed onthe epithelium of the tongue and oralcavity (Figs. 6B,F, 4F). In contrast,Has2, but not Has3, is expressed inthe developing foliate papillae at thebase of the tongue (Fig. 6E).
Has2 and Has3 AreExpressed During Inner EarDevelopment in Overlappingand Complementary Patterns
During inner ear development, Has3,and Has2 to a lesser extent, are ex-pressed in the otic capsule, the precur-sor to the endochondral bony laby-
Fig. 5. Expression of hyaluronan synthase (Has) 2 and Has3 during vibrissa follicle development in the embryonic day (E) 14.5 mouse. A–D�: In situhybridization on transverse cryosections of vibrissa follicles at various stages using a radiolabeled Has2 probe (A,C) or Has3 probe (B,D),counterstained with Hoechst (A�–D�). A–D: The drawn solid line represents the basal layer of the epidermis. Arrows and arrowheads indicate positivesignal for Has2 or Has3 transcripts in the corresponding labeled structures. Scale bars � 100 �m in B� (applies to A,A�,B,B�), D� (applies to C,C�,D,D�).
HYALURONAN SYNTHASE EXPRESSION 137

rinth (Fig. 6G,H; Liu et al., 2002).Only Has2 is expressed at a high levelat the inner rim of the semicircularcanals, and at lower levels in the mes-enchyme surrounding the vestibularand cochlear structures (Fig. 6G). Ofinterest, only Has3 is expressed in the
vestibulocochlear ganglion and con-tinues to be expressed in later stagesin the vestibular and spiral (cochlear)ganglia (Fig. 6H, and data not shown).
The inner ear develops like otherectodermally derived sensory organs,with the formation of a thickened ec-
toderm, the otic placode, which invagi-nates, then pinches off into a vesicle(Graham and Begbie, 2000; Francis-West et al., 2002). The otic vesicletransforms under the instruction ofmultiple molecular cues to become amembranous labyrinth for balance
Fig. 6. A–H: Expression of Has2 and Has3 during craniofacial and inner ear development. In situ hybridization on frontal (A,B) and transverse(C,D,G,H) sections of an E13.5 mouse and transverse sections of an E15.5 mouse (E,F), counterstained with Hoechst (A�,C�,E�,G�). Radiolabeled Has2(A,C,E,G) or Has3 probes (B,D,F,H) were used. Arrowheads indicate overlapping Has2 and Has3 expression. Arrows indicate generally exclusiveexpression. Specific regions are labeled as follows: foliate papillae (fp), lateral semicircular canal (lsc), nasal capsule (ns), nasal septum (ns), olfactoryepithelium (oe), orbitosphenoid bone (or), otic capsule (oc), posterior semicircular canal (psc), secondary palatal shelves (ps), respiratory epithelium (re),tongue epithelium (te), vestibulocochlear ganglion (vcg), and vomeronasal organ (vn). Scale bars � 500 �m in C� (applies to A–D,A�,C�), 500 �m in E�(applies to E,E�,F); 500 �m in G� (applies to G,G�,H).
138 TIEN AND SPICER

and hearing with its three semicircu-lar canals, vestibule (saccule and utri-cle), and cochlea (Morsli et al., 1998).Among those molecules proposed to becritical for development of the innerear is HA. To form the semicircularcanals, the otic vesicle develops a setof three flattened outpockets. The cen-ters of each of the outpockets will in-dent, and the epithelial sheets of theindentation will eventually make con-tact and fuse, leaving a tube, the semi-circular canal. HA was necessary toform these indentations in a Xenopuslaevis model system (Haddon andLewis, 1991). We show here that Has2is the major HA synthase expressed inoutpocket regions of the developingsemicircular canals (Fig. 6G).
SUMMARY
Overall, we have learned that Has1overlaps in expression with Has2early in development. Has2 expres-sion overlaps with Has3 in a limitednumber of tissues. Where Has3 is ex-pressed, Has2 is generally expressedin adjacent mesodermal tissues. Be-cause the different Has genes areresponsible for possibly different en-zymatic activities and molecularweights of the HA polymer synthe-sized, the complementary expressionpattern of Has2 and Has3 in certaintissues may further support distinctfunctions of the HA synthases. Has2remains the major HA synthase dur-ing development as it is the mostwidely expressed, which is consistentwith the phenotype observed in Has2-deficient animals. Here, we revealthat Has2 is the only HA synthaseexpressed in a narrow window of de-velopment, between E8.5 and E9.5.The exclusive expression of Has2would help explain the lethality ofHas2-deficient animals at this stage,suggesting that hyaluronan may benecessary before E8.5, but Has1 mayfulfill this requirement in the absenceof Has2.
We have shown that HA synthasetranscripts are generally distributedin a spatial and temporal manner thatis consistent with the previously re-ported localization of HA, especially inmorphogenetically active systems(Fenderson et al., 1993). Surprisingly,Has3 transcripts were detected in lo-cales where HA has not been reported
to exist and where Has2 transcriptswere not detected, for instance, in thedental lamina and dental mesen-chyme (Yu and Toole, 1997). This find-ing will, no doubt, invite further spec-ulation on the differences between thethree HA synthases. Based upon theexpression patterns we have observed,it would be extremely informative tofurther investigate phenotypes of theviable Has1- and Has3-deficient micein an effort to detect subtle yet impor-tant developmental defects that havegone undetected to date.
Based upon our data showing ex-pression of Has2 and Has3 during thedevelopment of ectodermally derivedorgans that originate as a placode (ol-factory, otic, lens, hair, teeth), itwould be interesting to investigatewhether Has2 and Has3 are expressedduring the development of other ecto-dermally derived organs. These in-clude the adenohypophysis placodethat gives rise to the anterior pitu-itary, the trigeminal placode thatgives rise to the somatosensory neu-rons of the trigeminal ganglion, thelateral line placode of fish and am-phibians that gives rise to receptorcells and sensory neurons, and theepibranchial placode that is located inthe branchial arches and gives rise tothe neurons of the distal facial, glos-sopharyngeal, and vagal ganglia (Gra-ham and Begbie, 2000; Baker andBronner-Fraser, 2001; Francis-Westet al., 2002; Pispa and Thesleff, 2003).It is possible that the complementaryexpression pattern that we observedbetween Has2 and Has3 during devel-opment of some ectodermally derivedtissues is repeated in other similarlyderived structures, thus suggesting acommon and conservative mechanismfor this morphogenetic event.
A model of the conserved nature ofthe Has genes is clearly emerging. To-gether with our results in the mouse,along with the results of other inves-tigators in zebrafish and Xenopus, thepattern of Has expression during de-velopment is this: Has1 is promi-nently expressed in the early stages ofmidblastula, gastrulation, and neuru-lation (Bakkers et al., 2004; Nardini etal., 2004); Has3 is expressed later indevelopment when sensory organs be-gin to form (Vigetti et al., 2003; Nar-dini et al., 2004); and Has2 is ex-pressed throughout all stages of the
embryo (Bakkers et al., 2004; Nardiniet al., 2004). Further analyses in spe-cific tissue types between species willbe necessary to elucidate the extent ofthe conserved expression of the threeHas genes.
EXPERIMENTALPROCEDURES
Tissue Preparation
Embryos were collected at 7.5 through15.5 days postcoitus from C57BL/6J orCD-1 mice, dissected in ice-cold phos-phate-buffered saline (PBS), and fixedwith 4% paraformaldehyde for 2–20hr at 4°C with slow shaking. Forwhole-mount in situ hybridization,fixed embryos were washed with 0.1%Tween-20 in PBS (PBT), 3 times for 5min at 4°C, dehydrated in increasingconcentrations of methanol in PBT(25%, 50%, 75%, 100%) for 10 mineach at 4°C, and stored in fresh 100%methanol at �20°C until use. For ra-dioactive in situ hybridization on fro-zen sections, fixed embryos werewashed twice with PBS for 30 min at4°C and subsequently incubated over-night in 20% sucrose at 4°C with slowshaking. Intact embryos or dissectedtissues were then embedded in Tis-sue-Tek O.C.T. compound, frozen in amethylbutanol/dry ice bath, andstored at �80°C.
Whole-Mount andRadioactive In SituHybridization
RNA in situ hybridizations on wholeembryos were carried out essentiallyas described, with slight modifications(Conlon and Rossant, 1992). Digoxige-nin (DIG)-labeled riboprobes weresynthesized and purified using a DIG-RNA labeling kit (Roche, Basel, Swit-zerland). Radioactive in situ hybrid-ization was carried out on 12-�mfrozen tissue sections essentially asdescribed (Wilkinson, 1992). 35S-uri-dine triphosphate–labeled riboprobesat 105 dpm per �l hybridization solu-tion were layered onto sections andincubated at 50°C for �16 hr in achamber humidified with 50% form-amide and 4� standard saline citrate.The sense and Has2 antisense probeswere generated from a plasmid con-taining the mouse Has2 open-reading
HYALURONAN SYNTHASE EXPRESSION 139

frame. The Has1 and Has3 probeswere similarly generated from plas-mids containing the complete open-reading frames in each instance. Thespecificity of each probe for the specificHA synthase transcripts was verifiedby Northern analyses before use inthese experiments. No cross-hybrid-ization was observed between thethree Has riboprobes. No specific sig-nal was detected using sense probes.
ACKNOWLEDGMENTSWe thank Paul FitzGerald, RichardTucker, and Tom Blankenship for in-valuable advice and support; Rob Kosfor technical advice; and Doris Wu forhelpful discussions.
REFERENCES
Archer CW, Dowthwaite GP, Francis-WestP. 2003. Development of synovial joints.Birth Defects Res C Embryo Today 69:144–155.
Baker CV, Bronner-Fraser M. 2001. Verte-brate cranial placodes I. Embryonic in-duction. Dev Biol 232:1–61.
Bakkers J, Kramer C, Pothof J, QuaedvliegNE, Spaink HP, Hammerschmidt M.2004. Has2 is required upstream of Rac1to govern dorsal migration of lateral cellsduring zebrafish gastrulation. Develop-ment 131:525–537.
Bourguignon LY, Zhu H, Shao L, ChenYW. 2000. CD44 interaction with tiam1promotes Rac1 signaling and hyaluronicacid-mediated breast tumor cell migra-tion. J Biol Chem 275:1829–1838.
Burk DT. 1985. Morphological effects ofstreptomyces hyaluronidase treatmenton the outgrowth of the nasal processesin mouse embryos. J Craniofac GenetDev Biol 5:385–398.
Camenisch TD, Spicer AP, Brehm-GibsonT, Biesterfeldt J, Augustine ML, CalabroA, Jr., Kubalak S, Klewer SE, McDonaldJA. 2000. Disruption of hyaluronan syn-thase-2 abrogates normal cardiac mor-phogenesis and hyaluronan-mediatedtransformation of epithelium to mesen-chyme. J Clin Invest 106:349–360.
Cao HJ, Wang HS, Zhang Y, Lin HY,Phipps RP, Smith TJ. 1998. Activation ofhuman orbital fibroblasts through CD40engagement results in a dramatic induc-tion of hyaluronan synthesis and prosta-glandin endoperoxide H synthase-2 ex-pression. Insights into potentialpathogenic mechanisms of thyroid-asso-ciated ophthalmopathy. J Biol Chem 273:29615–29625.
Chai Y, Jiang X, Ito Y, Bringas P, Jr., HanJ, Rowitch DH, Soriano P, McMahon AP,Sucov HM. 2000. Fate of the mammaliancranial neural crest during tooth andmandibular morphogenesis. Develop-ment 127:1671–1679.
Conlon RA, Rossant J. 1992. Exogenousretinoic acid rapidly induces anterior ec-topic expression of murine Hox-2 genesin vivo. Development 116:357–368.
Craig FM, Bayliss MT, Bentley G, ArcherCW. 1990. A role for hyaluronan in jointdevelopment. J Anat 171:17–23.
DeAngelis PL. 1999. Hyaluronan syn-thases: fascinating glycosyltransferasesfrom vertebrates, bacterial pathogens,and algal viruses. Cell Mol Life Sci 56:670–682.
Delpech B, Delpech A, Bruckner G, GirardN, Maingonnat C. 1989. Hyaluronan andhyaluronectin in the nervous system. In:The biology of hyaluronan. Chichester:Wiley. p 208–232.
Delpech B, Girard N, Bertrand P, CourelMN, Chauzy C, Delpech A. 1997. Hyalu-ronan: fundamental principles and ap-plications in cancer. J Intern Med 242:41–48.
Engstrom-Laurent A. 1997. Hyaluronan injoint disease. J Intern Med 242:57–60.
Evanko SP, Wight TN. 1999. Intracellularlocalization of hyaluronan in proliferat-ing cells. J Histochem Cytochem 47:1331–1342.
Fenderson BA, Stamenkovic I, Aruffo A.1993. Localization of hyaluronan inmouse embryos during implantation,gastrulation and organogenesis. Differ-entiation 54:85–98.
Francis-West PH, Ladher RK, SchoenwolfGC. 2002. Development of the sensoryorgans. Sci Prog 85:151–173.
Gilbert SF. 1997. Developmental biology.Sunderland, MA: Sinauer Associates,Inc. 245 p.
Graham A, Begbie J. 2000. Neurogenic pla-codes: a common front. Trends Neurosci23:313–316.
Green SJ, Tarone G, Underhill CB. 1988.Aggregation of macrophages and fibro-blasts is inhibited by a monoclonal anti-body to the hyaluronate receptor. ExpCell Res 178:224–232.
Haddon CM, Lewis JH. 1991. Hyaluronanas a propellant for epithelial movement:the development of semicircular canalsin the inner ear of Xenopus. Develop-ment 112:541–550.
Hay ED. 1980. Development of the verte-brate cornea. Int Rev Cytol 63:263–322.
Hutson MR, Kirby ML. 2003. Neural crestand cardiovascular development: a 20-year perspective. Birth Defects Res CEmbryo Today 69:2–13.
Itano N, Sawai T, Yoshida M, Lenas P,Yamada Y, Imagawa M, Shinomura T,Hamaguchi M, Yoshida Y, Ohnuki Y,Miyauchi S, Spicer AP, McDonald JA,Kimata K. 1999. Three isoforms of mam-malian hyaluronan synthases have dis-tinct enzymatic properties. J Biol Chem274:25085–25092.
Knudson CB. 2003. Hyaluronan and CD44:strategic players for cell-matrix interac-tions during chondrogenesis and matrixassembly. Birth Defects Res C EmbryoToday 69:174–196.
Knudson CB, Toole BP. 1985. Changes inthe pericellular matrix during differenti-
ation of limb bud mesoderm. Dev Biol112:308–318.
Knudson W, Biswas C, Li X-Q, Nemec R,Toole B. 1989. The role and regulation oftumour-associated hyaluronan. In: Thebiology of hyaluronan. Chichester:Wiley. p 150–169.
Knudson CB, Munaim SI, Toole BP. 1995.Ectodermal stimulation of the produc-tion of hyaluronan-dependent pericellu-lar matrix by embryonic limb mesoder-mal cells. Dev Dyn 204:186–191.
Laurent TC. 1998. The chemistry, biologyand medical applications of hyaluronanand its derivatives. London: PortlandPress.
Lee JY, Spicer AP. 2000. Hyaluronan: amultifunctional, megaDalton, stealthmolecule. Curr Opin Cell Biol 12:581–586.
Lee JY, Rountree RB, Kingsley DM, SpicerAP. 2002. In vivo investigation of hyalu-ronan and hyaluronan synthase-2 func-tion during cartilage and joint develop-ment. In: Kuettner KE, editor. The manyfaces of osteoarthritis. Basel, Switzer-land: Birkhauser. p 213–218.
Liu W, Li G, Chien JS, Raft S, Zhang H,Chiang C, Frenz DA. 2002. Sonic hedge-hog regulates otic capsule chondrogene-sis and inner ear development in themouse embryo. Dev Biol 248:240–250.
Lokeshwar VB, Selzer MG. 2000. Differ-ences in hyaluronic acid-mediated func-tions and signaling in arterial, microves-sel, and vein-derived human endothelialcells. J Biol Chem 275:27641–27649.
Maleski MP, Knudson CB. 1996. Hyaluro-nan-mediated aggregation of limb budmesenchyme and mesenchymal conden-sation during chondrogenesis. Exp CellRes 225:55–66.
Morsli H, Choo D, Ryan A, Johnson R, WuDK. 1998. Development of the mouse in-ner ear and origin of its sensory organs.J Neurosci 18:3327–3335.
Mullegger J, Lepperdinger G. 2002. Hyalu-ronan is an abundant constituent of theextracellular matrix of Xenopus em-bryos. Mol Reprod Dev 61:312–316.
Nardini M, Ori M, Vigetti D, Gornati R,Nardi I, Perris R. 2004. Regulated geneexpression of hyaluronan synthases dur-ing Xenopus laevis development. GeneExpr Patterns 4:303–308.
Oliferenko S, Kaverina I, Small JV, HuberLA. 2000. Hyaluronic acid (HA) bindingto CD44 activates Rac1 and induces la-mellipodia outgrowth. J Cell Biol 148:1159–1164.
Perris R, Krotoski D, Lallier T, Domingo C,Sorrell JM, Bronner-Fraser M. 1991.Spatial and temporal changes in the dis-tribution of proteoglycans during avianneural crest development. Development111:583–599.
Pispa J, Thesleff I. 2003. Mechanisms ofectodermal organogenesis. Dev Biol 262:195–205.
Pitsillides AA. 2003. Identifying and char-acterizing the joint cavity-forming cell.Cell Biochem Funct 21:235–240.
Pratt RM, Goggins JF, Wilk AL, King CT.1973. Acid mucopolysaccharide synthe-
140 TIEN AND SPICER

sis in the secondary palate of the devel-oping rat at the time of rotation and fu-sion. Dev Biol 32:230–237.
Slevin M, Kumar S, Gaffney J. 2002. An-giogenic oligosaccharides of hyaluro-nan induce multiple signaling path-ways affecting vascular endothelial cellmitogenic and wound healing re-sponses. J Biol Chem 277:41046 –41059.
Solursh M, Morriss GM. 1977. Glycosami-noglycan synthesis in rat embryos dur-ing the formation of the primary mesen-chyme and neural folds. Dev Biol 57:75–86.
Solursh M, Fisher M, Meier S, Singley CT.1979. The role of extracellular matrix inthe formation of the sclerotome. J Em-bryol Exp Morphol 54:75–98.
Spicer AP, McDonald JA. 1998. Character-ization and molecular evolution of a ver-tebrate hyaluronan synthase gene fam-ily. J Biol Chem 273:1923–1932.
Spicer AP, Tien JYL. 2004. HA and mor-phogenesis. Birth Defects Res EmbryoToday 72:89–108.
Toole BP. 1991. Proteoglycans and hyalu-ronan in morphogenesis and differentia-tion. New York: Plenum Press. p 305–341.
Toole BP. 1997. Hyaluronan in morphogen-esis. J Intern Med 242:35–40.
Toole BP. 2001. Hyaluronan in morphogen-esis. Semin Cell Dev Biol 12:79–87.
Underhill CB, Toole BP. 1981. Receptorsfor hyaluronate on the surface of parentand virus-transformed cell lines: bindingand aggregation studies. Exp Cell Res131:419–423.
Vigetti D, Viola M, Gornati R, Ori M, NardiI, Passi A, De Luca G, Bernardini G.2003. Molecular cloning, genomic organi-zation and developmental expression ofthe Xenopus laevis hyaluronan synthase3. Matrix Biol 22:511–517.
Wilkinson DG. 1992. In situ hybridization:a practical approach. Oxford; New York:IRL Press at Oxford University Press. p15–32.
Yu Q, Toole BP. 1997. Common pattern ofCD44 isoforms is expressed in morpho-genetically active epithelia. Dev Dyn 208:1–10.
Zhang Y, Wang S, Song Y, Han J, Chai Y,Chen Y. 2003. Timing of odontogenicneural crest cell migration and tooth-forming capability in mice. Dev Dyn 226:713–718.
HYALURONAN SYNTHASE EXPRESSION 141
Copyright © 2022 FDOKUMEN




















