
The incidence of focal and non-focal EEG abnormalities in clinical epilepsy
Upload
independentCategory
view
0download
0
Molecular Brain Research 89 (2001) 133–146www.elsevier.com/ locate /bres
Research report
Caspase mRNA expression in a rat model of focal cerebral ischemiaa , a b a*David C. Harrison , Robert P. Davis , Brian C. Bond , Colin A. Campbell ,
a a aMichael F. James , Andrew A. Parsons , Karen L. PhilpottaDepartment of Neurology, GlaxoSmithKline, New Frontiers Science Park, Third Avenue, Harlow, Essex CM19 5AW, UK
bDepartment of Statistical Sciences, GlaxoSmithKline, New Frontiers Science Park, Third Avenue, Harlow, Essex CM19 5AW, UK
Accepted 6 February 2001
Abstract
Proteins of the caspase family are involved in the signalling pathway that ultimately leads to programmed cell death (apoptosis), whichhas been reported to occur in some experimental models of stroke. In a previous paper we used quantitative reverse transcription andpolymerase chain reaction (RT-PCR) to characterise changes in the mRNA expression of one member of this family, caspase-3, in a ratmodel of permanent focal ischemia. Here we have used this technique to study the expression of a further three caspases which areinvolved in different aspects of caspase signalling. Caspase-8, involved in Fas-mediated apoptosis, was upregulated in the cortex ofischemic rats. Caspase-11, which leads to the synthesis of the functional form of the cytokine interleukin-1b, also showed increasedexpression, but with a different temporal profile from caspase-8. In contrast, caspase-9, which forms part of the pathway signallingthrough the mitochondria, showed a decrease in expression. The expression of a further four caspases (1, 2, 6 and 7) has also beencharacterised in a simpler experiment. These caspases all showed distinctive patterns of expression following the induction of ischemia.These data lead us to conclude that caspase expression as a whole is under very strict transcriptional control in this model. Certainelements of caspase signalling, such as the Fas-induced pathway and the events upstream of IL-1b processing, are upregulated, whileothers are not. This may be due to some form of genetic program activated in response to ischemia in the brain and may highlight whichbiological pathways are modulated. 2001 Elsevier Science B.V. All rights reserved.
Theme: Disorders of the nervous system
Topic: Ischemia
Keywords: Caspase; Ischemia; Gene expression; Apoptosis
1. Introduction from internucleosomal cleavage of DNA in apoptosis.This, and other features of apoptosis, are observed in brain
The loss of cells following an ischemic insult to the tissue from animal models of focal ischemia, and whilebrain results from a wide range of noxious stimuli in and there is widespread necrosis occurring within the area ofaround the affected area of tissue. These stimuli include injury, apoptosis is also thought to play an important roledeprivation of oxygen and glucose necessary for normal in ischemic cell death in the brain [38,40,44].cellular survival, increased concentrations of excitotoxic In C. elegans the CED-3 protein is essential for apop-amino acids and release of cytotoxic mediators such as tosis occurring during normal development. The ced-3cytokines [36]. Despite the variety of death-promoting gene was found to have homology to mammalianstimuli, cell death is considered to occur by only two basic interleukin-1b converting enzyme (ICE), an enzyme re-mechanisms: necrosis and apoptosis, which can be dis- sponsible for the generation of the pro-inflammatorytinguished by their distinctive morphological and bio- cytokine interleukin-1b (IL-1b) from its precursor [66].chemical features, most notably the laddering which results Subsequently, ICE has been found to belong to a family
of at least 12 related proteases, known as caspases, whichshare an active site, QACXG, and the ability to cleave*Corresponding author. Tel.: 144-1279-622-728; fax: 144-1279-622-substrates at sites with aspartate in the P1 position. A main371.
E-mail address: david c [email protected] (D.C. Harrison). functional role of caspases is to form a signalling cascade] ]
0169-328X/01/$ – see front matter 2001 Elsevier Science B.V. All rights reserved.PI I : S0169-328X( 01 )00058-4

134 D.C. Harrison et al. / Molecular Brain Research 89 (2001) 133 –146
between death-inducing stimuli such as activation of the drug targets themselves. Alternatively, by understandingdeath receptor Fas, and cleavage of protein substrates possible transcriptional programs activated in the ischemictargeted during apoptosis [7,12,51]. brain, it may be possible to target signalling mechanisms
In a variety of animal models of cerebral ischemia, upstream of transcription and inhibit the execution of suchincluding the model employed in this study, increased a program as a whole.caspase activity has been observed [6,17] and caspaseinhibitors can reduce the infarct volume [14,23,43]. Lesionvolumes are also reduced in animals with mutations ordeletions of caspase genes [22,55]. 2. Materials and methods
Apoptosis requires de novo synthesis of mRNA andprotein to take place [46], although there is an overall Animal procedures were performed in accordance withdecrease in RNA transcription [10]. Taken together, these the Home Office regulations as outlined in the Animalsobservations suggest that there may be a distinct genetic (Scientific Procedures) Act 1986. All procedures wereprogram involving selective transcription of certain critical critically reviewed by colleagues experienced in animalgenes required for apoptosis to proceed [15]. research, in accordance with company regulations. All
There is increasing evidence of an upregulation of animals were housed under standardised environmentalcaspases at a transcriptional level during apoptosis in vitro conditions (12 h light /dark cycle, 21618C and 5565%[5,13,54,65]. Increased levels of caspase expression have humidity) and allowed free access to food and water for atalso been demonstrated in several animal models of least 5 days acclimatisation prior to use.cerebral ischemia [1,4,27,29,32,49]. However, while most Permanent middle cerebral artery occlusion (MCAO)members of the caspase family have been studied to some was carried out according to the method of Zea-Longa etextent in in vitro systems, until now work in in vivo al. [68]. Male Sprague Dawley rats weighing 300–350 gsystems has principally focussed on the more extensively were anesthetised with halothane. The left middle cerebralstudied members of the family, namely caspases-1, -2 and artery was occluded by an intraluminal filament at its-3. origin from the circle of Willis. The rats were euthanased
Recent advances in the use of fluorogenic probes in 3, 6, 12 or 24 h after surgery by halothane overdose.conjunction with PCR have enabled the measurement of an Parallel groups of rats either received sham surgery, inaccumulating PCR product in real time [19,26,39]. This which an identical procedure was followed but withoutallows the rapid generation of quantitative data showing inserting the filament, or no treatment at all prior to
¨changes in transcript numbers in tissue samples. An euthanasia (n 5 2 naıve rats, four sham and four MCAOincrease in throughput not only allows a greater number of for 3, 6 and 12 h time points, three sham and five MCAOtranscripts to be studied, but also allows a greater number for the 24 h time point).of animals and experimental conditions to be included in a A neurological assessment of motor and behaviouralstudy, enabling a more sensitive and reliable statistical changes was carried out on all sham-operated and MCAOanalysis of the data. animals 1 h after surgery (to determine success of occlu-
We have previously used this technique to study the sion), and immediately prior to sacrifice. Rats were scoredexpression of caspase-3 following permanent occlusion of on the following six point scale: 0, no deficit; 1, failure tothe middle cerebral artery in rats, a severe model of extend right forepaw fully; 2, decreased resistance tocerebral ischemia [24]. In the present study we have lateral push, or decreased grip of right forelimb while tailcarried out further analysis of caspase-3 expression and pulled; 3, spontaneous circling or walking to contralateralassessed the pattern and time course of expression of a side; 4, walks only when stimulated; 5, unresponsive tofurther seven caspases in this model. stimulation with a depressed level of consciousness. The
We found marked differences in the expression profiles inclusion of an animal in the study was dependent on aof the various caspases following induction of ischemia, neurological score of $3 for MCAO rats or 0 for sham-with caspases-1, -3, -6, -7, -8 and -11 showing increased operated rats 1 h after surgery.expression, although with differing time courses. In con- The left and right cerebral cortices were dissected fromtrast, caspase-2 expression did not change and caspase-9 each rat, snap frozen in liquid nitrogen and stored atmRNA was reduced in the ischemic cortex. Taken together 2808C. The frozen tissue samples were homogenised inthese data suggest that the caspases have functionally TRIzol reagent (Life Technologies, Gaithersburg, MD,distinct roles in the development of the lesion, and give an USA) using 1 ml of TRIzol per 50 mg of tissue. Totalindication of which caspases may be regulated in this RNA was extracted from the tissue according to theprocess. This in turn may shed light on possible genetic manufacturer’s suggested protocol with the addition of anprograms activated in the ischemic brain. extra chloroform extraction step and phase separation, and
Increased understanding of both of these areas is of an extra wash of the isolated RNA in 70% ethanol. Thepotential value in the search for therapeutic agents for RNA was resuspended in PCR grade water and thestroke. Individual caspases may be viewed as potential concentration calculated by A measurement. RNA260

D.C. Harrison et al. / Molecular Brain Research 89 (2001) 133 –146 135
quality was assessed by electrophoresis on a 1% agarose probe sequences together with Genbank accession numbersgel. An aliquot of each RNA sample was taken and equal where applicable are shown in Table 1.quantities of RNA were pooled from all rats according to The way in which particular transcripts change betweentreatment group, time point and tissue. Data was generated treatment groups and their behavior over time was assessedfor all of the caspases in the study using these pooled RNA by analysis of variance (ANOVA) on log transformed datasamples. In addition, RNA samples from individual rats accommodating for expression of mRNA for the house-were used for caspases-3, -8, -9 and -11. For this latter keeping gene GAPDH by incorporating this as a covariate.group of caspases, only data from individual rats is ANOVAs of data from individual rats were split accordingreported here. to the hierarchical random error structure (i.e. between
First strand cDNA was synthesised from 1 mg of each animals, between samples within animals and between RTRNA sample; 0.01 M DTT, 0.5 mM each dNTP, 0.5 mg replicates within sample) and treatment factors testedoligo(dT) primer, 40 U RNAseOUT ribonuclease inhibitor against the appropriate error term. Due to pooling of(Life Technologies), 200 U SuperscriptII reverse tran- samples, ANOVAs of the remaining data had a simplerscriptase (Life Technologies). Triplicate reverse transcrip- structure with only one error term (i.e. between RTtion reactions were performed along with an additional replicates within sample). Resulting treatment means fromreaction in which the reverse transcriptase enzyme was analyses are adjusted for the housekeeping gene. Pairwise
¨omitted to allow for assessment of genomic DNA contami- comparisons of each treatment group relative to the naıvenation in each sample. The resulting cDNA products were group of animals were made at each time point and thedivided into 20 aliquots using a Hydra 96 robot (Robbins 95% (for individual data) or 99% (for pooled data)Scientific, Sunnyvale, CA, USA) for parallel Taqman PCR confidence interval for the fold difference was calculatedreactions using different primer and probe sets for quantifi- using the appropriate pooled estimates of variability fromcation of multiple cDNA sequences. Where pooled RNA the ANOVA. Error between RT replicates is naturallywas used, all cDNAs were accommodated on one 96 well smaller than that between animals and between samples,plate, however individual rat samples were assayed on four and hence the necessity to use this in the calculation ofplates, with one time point on each plate, together with confidence intervals for pooled data was compensated by
¨replicate samples from the naıve animals to control for the calculation of 99% confidence intervals. In addition,plate effects. for caspases where data from individual rats was used,
Taqman PCR was carried out using an ABI prism 7700 comparisons were made between ipsilateral and contrala-sequence detector (Perkin-Elmer Applied Biosystems, teral cortex of MCAO rats, and also ipsilateral cortex ofFoster City, CA, USA) on the cDNA aliquots; 2.5 mM MCAO rats and the corresponding (left) cortex of sham-MgCl , 0.2 mM dATP, dCTP, dGTP and dUTP, 0.1 mM operated rats.2
each primer, 0.05 mM Taqman probe, 0.01 U AmpErase Three variance components analyses were used touracil-N-glycosylase, 0.0125 U Amplitaq Gold DNA poly- investigate the magnitude of the different sources ofmerase (all reagents from Applied Biosystems) 508C for 2 random variability in the two experiment for each gene.min, 958C for 10 min followed by 40 cycles of 958C for 15 The first two analyses were performed on the individuals, 608C for 1 min. Additional reactions were performed on data where available. One on the sham and MCAO data
¨each 96 well plate using known dilutions of rat genomic together and the other on the data from naıve rats alone.¨DNA (Clontech Laboratories, Palo Alto, CA, USA) or in The analysis of the naıve data enabled the investigation of
the case of caspases-2 and -9, where the amplicon lies plate to plate variability, which was aliased with the timeacross an intron /exon boundary, plasmid DNA. This factor in the sham and MCAO data. The third varianceprovided a PCR template to allow construction of a component analysis was performed on the pooled data.standard curve relating threshold cycle to template copy These are presented in the original units and additionallynumber. as percentages of the total amount of random variability in
Primer and probe sets for quantitative RT-PCR were that particular gene (Table 2).designed using Primer Express v.1.0 software (AppliedBiosystems). For caspases-1, -2, -3 and -11 and thehousekeeping gene glyceraldehyde-3-phosphate dehydro- 3. Resultsgenase (GAPDH), the rat mRNA sequence was obtainedfrom the GenEMBL database. For caspases-6, -7, -8 and -9 We have carried out a quantitative analysis of caspases-no published rat sequence was available. In these cases, 8, -9 and -11 in the permanent MCA occlusion modelsmall fragments of rat sequence were generated by PCR using measurements taken from individual rats. For com-using rat brain cDNA together with primers designed from parison, further analysis has also been performed on thepublished human or mouse sequences. The PCR products data for caspase-3 reported previously [24]. Covariatewere sequenced, and the partial rat caspase sequences thus analysis has been used to normalise data using valuesgenerated were sufficient for the design of rat-specific obtained for mRNA expression of the housekeeping geneprimers and probes for quantitative RT-PCR. Primer and GAPDH. This allows one to correct for fluctuations in

136 D.C. Harrison et al. / Molecular Brain Research 89 (2001) 133 –146
Table 1Primer and probe sequences
Caspase-1 U14647 Forward primer 59-GACAGGTCCTGAGGGCAAAGRat Reverse primer 59-AAAAGTTCATCCAGCAATCCATTT
Taqman probe 59-TTGTCCCTACACTCACTGAGTTGATAAATTGCTTCaspase-2 U34684 Forward primer 59-CTCTTCAAGCTTTTGGGCTACAA
Rat Reverse primer 59-TTCTGAAGTTTCTCTTGCATTTCCTTaqman probe 59-TCCATGTGCTGTATGACCAGACTGCA
Caspase-3 U49930 Forward primer 59-AATTCAAGGGACGGGTCATGRat Reverse primer 59-GCTTGTGCGCGTACAGTTTC
Taqman probe 59-TTCATCCAGTCACTTTGCGCCATGCaspase-6 AF025670 Forward primer 59-AGCATGACGTGCCATTGGT
Rat Reverse primer 59-ACGTTGTCGTCCAGCTTGTCTTaqman probe 59-TCTGATGATCCACCACGTCCAGAGG
aCaspase-7 U67321 Forward primer 59-GTCCTTGCCATGCTCATTCAGMouse Reverse primer 59-CCCAGGGAAAGGGCTCCT
Taqman probe 59-TTGTGCAGGCCCTCTGCTCCATaCaspase-8 AF067834 Forward primer 59-TAAGACCTTTAAGGAGCTTCATTTTGA
Mouse Reverse primer 59-AGGATACTAGAACCTCATGGATTTGACTaqman probe 59-ATCGTGTCCTTCAGTGATTGCACAGCA
aCaspase-9 U60521 Forward primer 59-GAGGGAAGCCCAAGCTGTTCHuman Reverse primer 59-GCCACCTCAAAGCCATGGT
Taqman probe 59-TTTCTGCTCACCACCACAGGCCTGaCaspase-11 Y13089 Forward primer 59-CAGACAGTCACATTCCTGGTGTTAA
Mouse Reverse primer 59-GCTTCACTGTGCATTGTTCCATaqman probe 59-CTCATGGCACATTGTCAGGGCCTT
GAPDH U75401 Forward primer 59-GAACATCATCCCTGCATCCARat Reverse primer 59-CCAGTGAGCTTCCCGTTCA
Taqman probe 59-CTTGCCCACAGCCTTGGCAGCa In the case of caspases-7, -8, -9 and -11, these orthologs were used to design primers for the generation of a fragment of rat mRNA sequence by RT-PCRfrom rat brain mRNA.
RNA quality and efficiency of the reverse transcription crease over the time course following MCAO which wasreaction. We have previously demonstrated that GAPDH specific to the ipsilateral cortex of the ischemic animals
¨mRNA is unaltered in this model [24]. (Fig. 2a). Compared to naıve rats the mRNA was elevatedby 65, 65 and 223% in ipsilateral MCAO cortex at 6, 12
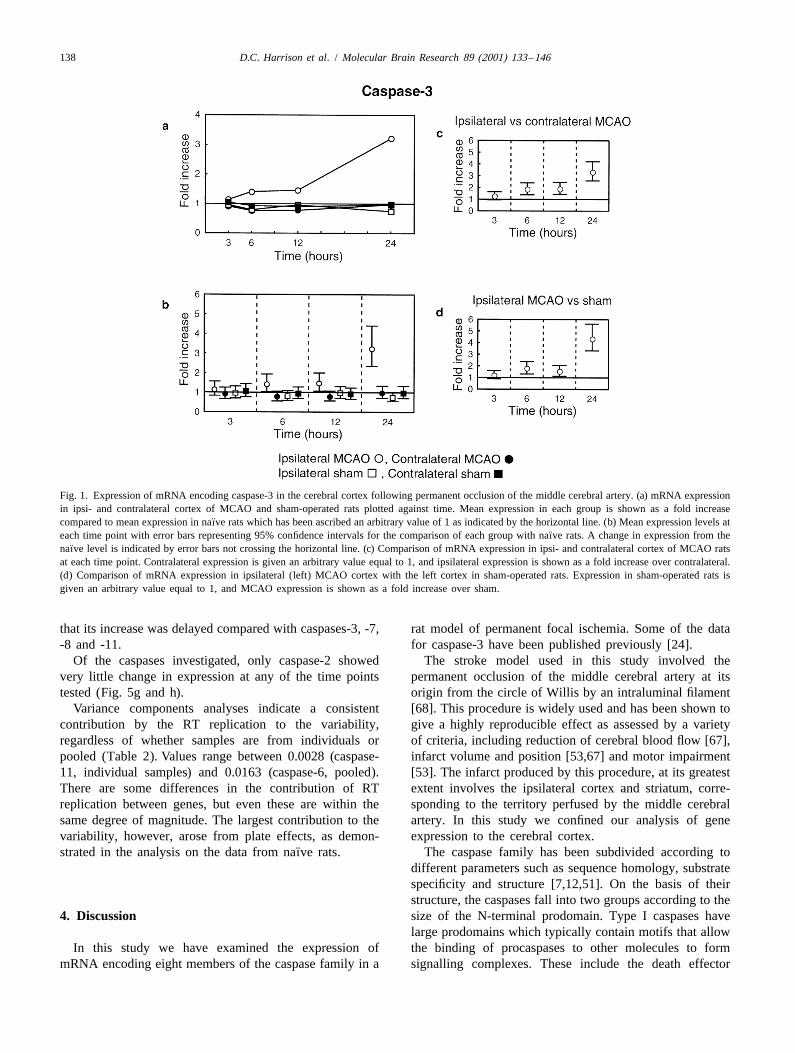
3.1. Caspase-3 and 24 h, respectively. No such changes were observed inthe contralateral cortex from MCAO rats or in sham-
Caspase-3 mRNA expression, as reported previously operated rats (Fig. 2b). That these changes are infarct[24], underwent a steady increase over the time course dependent is indicated by comparison with the otherfollowing MCA occlusion (Fig. 1a). This increased expres- control groups; a significant increase in caspase-8 mRNAsion was significant (P , 0.05) at 6, 12 and 24 h post- was observed at 12 and 24 h, but not at 6 h, in contralateral
¨MCAO when compared to expression levels in naıve rats cortex from MCAO rats (Fig. 2c), and at 6, 12 and 24 h(Fig. 1b), representing increases of 41, 46 and 220%, compared to ipsilateral cortex in sham-operated rats (Fig.respectively. By contrast, there was no significant altera- 2d).tion in expression levels in the contralateral cortex in At the 3 h time point the mRNA for caspase-8 wasMCAO rats or in either cortex in sham rats (Fig. 1b). reduced significantly in both sides of the cortex in MCAO
In this study, further analysis was carried out to compare rats, but not in sham-operated rats when compared to the¨mRNA in the ipsilateral cortex of MCAO rats to the other naıve group (Fig. 2b). There was no difference in caspase-
control groups, namely the ipsilateral (left) cortex of sham- 8 mRNA between ipsi- and contralateral MCAO cortex atoperated rats, and the contralateral cortex of MCAO rats. this time point.Caspase-3 mRNA was found to be significantly elevated at6, 12 and 24 h, but not 3 h, in ipsilateral MCAO cortex 3.3. Caspase-11when compared with contralateral MCAO (Fig. 1c) andipsilateral sham (Fig. 1d) cortices, showing that increases Caspase-11 mRNA was also increased in the ipsilateralwere infarct dependent. cortex of ischemic rats, but the changes have a somewhat
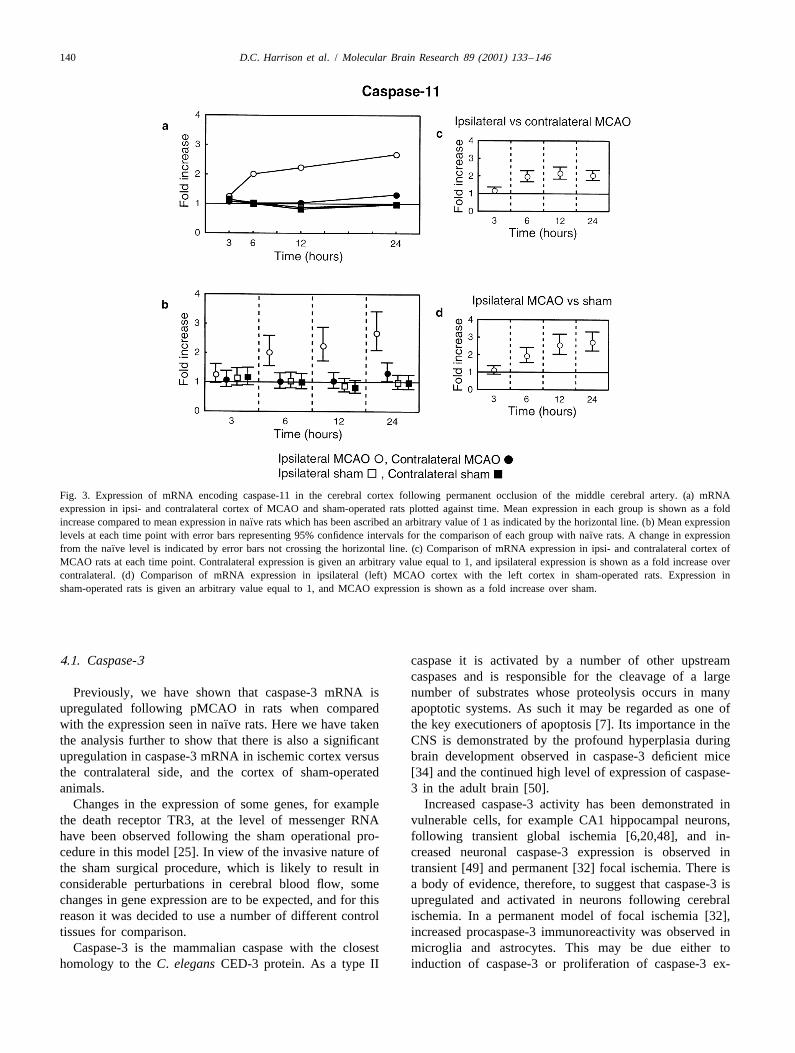
different profile from those observed for caspases-3 and -8.3.2. Caspase-8 The largest increase in expression occurred between 3 h,
when the RNA level is not significantly different from that¨Caspase-8 mRNA expression underwent a steady in- observed in naıve rats, and 6 h where an increase of 101%
D.C. Harrison et al. / Molecular Brain Research 89 (2001) 133 –146 137
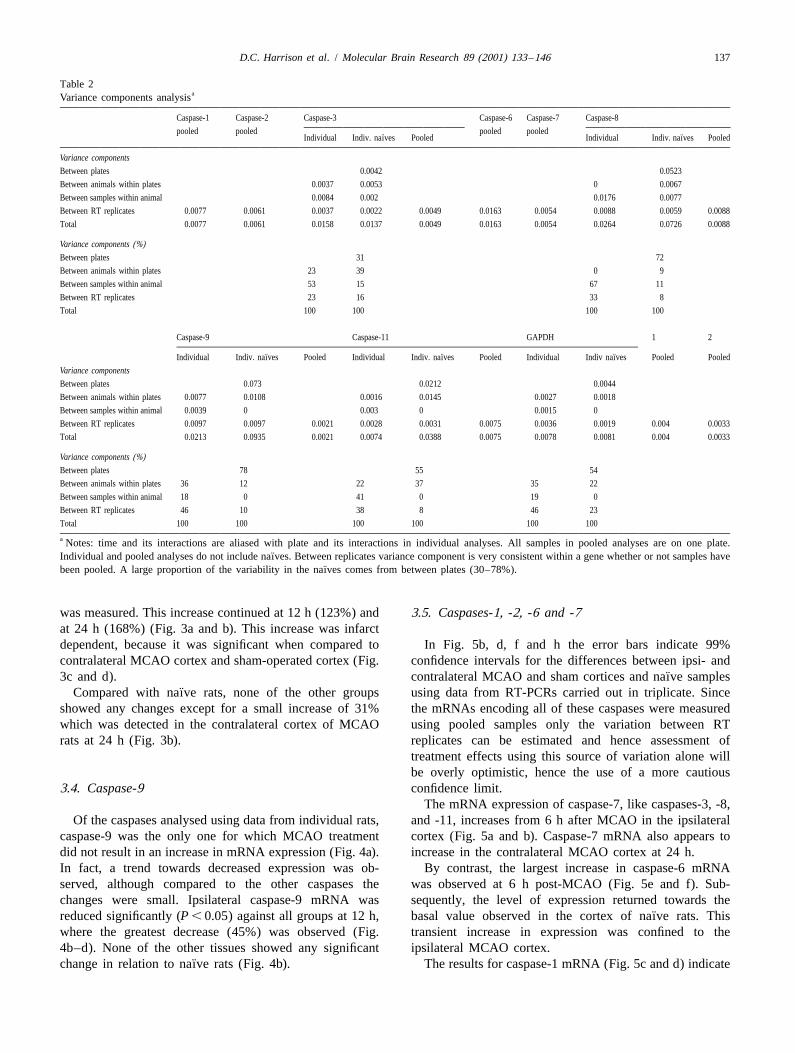
Table 2aVariance components analysis
Caspase-1 Caspase-2 Caspase-3 Caspase-6 Caspase-7 Caspase-8
pooled pooled pooled pooled¨ ¨Individual Indiv. naıves Pooled Individual Indiv. naıves Pooled
Variance components
Between plates 0.0042 0.0523
Between animals within plates 0.0037 0.0053 0 0.0067
Between samples within animal 0.0084 0.002 0.0176 0.0077
Between RT replicates 0.0077 0.0061 0.0037 0.0022 0.0049 0.0163 0.0054 0.0088 0.0059 0.0088
Total 0.0077 0.0061 0.0158 0.0137 0.0049 0.0163 0.0054 0.0264 0.0726 0.0088
Variance components (%)Between plates 31 72
Between animals within plates 23 39 0 9
Between samples within animal 53 15 67 11
Between RT replicates 23 16 33 8
Total 100 100 100 100
Caspase-9 Caspase-11 GAPDH 1 2
¨ ¨ ¨Individual Indiv. naıves Pooled Individual Indiv. naıves Pooled Individual Indiv naıves Pooled Pooled
Variance components
Between plates 0.073 0.0212 0.0044
Between animals within plates 0.0077 0.0108 0.0016 0.0145 0.0027 0.0018
Between samples within animal 0.0039 0 0.003 0 0.0015 0
Between RT replicates 0.0097 0.0097 0.0021 0.0028 0.0031 0.0075 0.0036 0.0019 0.004 0.0033
Total 0.0213 0.0935 0.0021 0.0074 0.0388 0.0075 0.0078 0.0081 0.004 0.0033
Variance components (%)Between plates 78 55 54
Between animals within plates 36 12 22 37 35 22
Between samples within animal 18 0 41 0 19 0
Between RT replicates 46 10 38 8 46 23
Total 100 100 100 100 100 100
a Notes: time and its interactions are aliased with plate and its interactions in individual analyses. All samples in pooled analyses are on one plate.¨Individual and pooled analyses do not include naıves. Between replicates variance component is very consistent within a gene whether or not samples have
¨been pooled. A large proportion of the variability in the naıves comes from between plates (30–78%).
was measured. This increase continued at 12 h (123%) and 3.5. Caspases-1, -2, -6 and -7at 24 h (168%) (Fig. 3a and b). This increase was infarctdependent, because it was significant when compared to In Fig. 5b, d, f and h the error bars indicate 99%contralateral MCAO cortex and sham-operated cortex (Fig. confidence intervals for the differences between ipsi- and
¨3c and d). contralateral MCAO and sham cortices and naıve samples¨Compared with naıve rats, none of the other groups using data from RT-PCRs carried out in triplicate. Since
showed any changes except for a small increase of 31% the mRNAs encoding all of these caspases were measuredwhich was detected in the contralateral cortex of MCAO using pooled samples only the variation between RTrats at 24 h (Fig. 3b). replicates can be estimated and hence assessment of
treatment effects using this source of variation alone willbe overly optimistic, hence the use of a more cautious
3.4. Caspase-9 confidence limit.The mRNA expression of caspase-7, like caspases-3, -8,
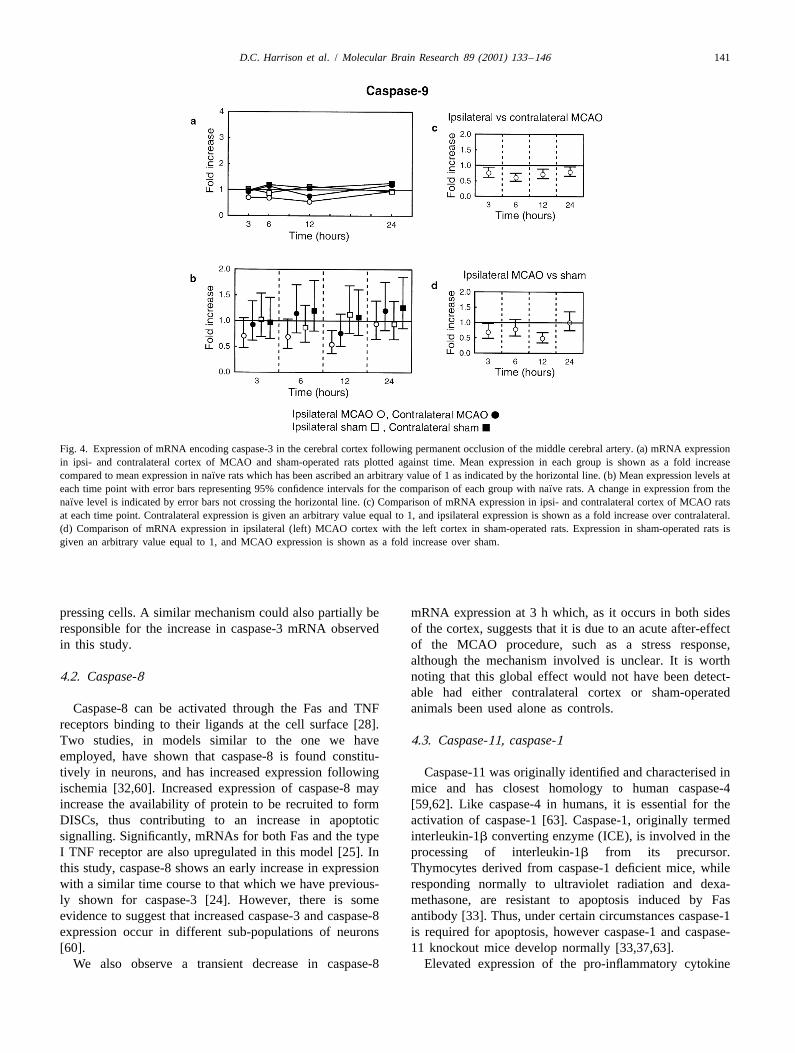
Of the caspases analysed using data from individual rats, and -11, increases from 6 h after MCAO in the ipsilateralcaspase-9 was the only one for which MCAO treatment cortex (Fig. 5a and b). Caspase-7 mRNA also appears todid not result in an increase in mRNA expression (Fig. 4a). increase in the contralateral MCAO cortex at 24 h.In fact, a trend towards decreased expression was ob- By contrast, the largest increase in caspase-6 mRNAserved, although compared to the other caspases the was observed at 6 h post-MCAO (Fig. 5e and f). Sub-changes were small. Ipsilateral caspase-9 mRNA was sequently, the level of expression returned towards the
¨reduced significantly (P , 0.05) against all groups at 12 h, basal value observed in the cortex of naıve rats. Thiswhere the greatest decrease (45%) was observed (Fig. transient increase in expression was confined to the4b–d). None of the other tissues showed any significant ipsilateral MCAO cortex.
¨change in relation to naıve rats (Fig. 4b). The results for caspase-1 mRNA (Fig. 5c and d) indicate
138 D.C. Harrison et al. / Molecular Brain Research 89 (2001) 133 –146
Fig. 1. Expression of mRNA encoding caspase-3 in the cerebral cortex following permanent occlusion of the middle cerebral artery. (a) mRNA expressionin ipsi- and contralateral cortex of MCAO and sham-operated rats plotted against time. Mean expression in each group is shown as a fold increase
¨compared to mean expression in naıve rats which has been ascribed an arbitrary value of 1 as indicated by the horizontal line. (b) Mean expression levels at¨each time point with error bars representing 95% confidence intervals for the comparison of each group with naıve rats. A change in expression from the
¨naıve level is indicated by error bars not crossing the horizontal line. (c) Comparison of mRNA expression in ipsi- and contralateral cortex of MCAO ratsat each time point. Contralateral expression is given an arbitrary value equal to 1, and ipsilateral expression is shown as a fold increase over contralateral.(d) Comparison of mRNA expression in ipsilateral (left) MCAO cortex with the left cortex in sham-operated rats. Expression in sham-operated rats isgiven an arbitrary value equal to 1, and MCAO expression is shown as a fold increase over sham.
that its increase was delayed compared with caspases-3, -7, rat model of permanent focal ischemia. Some of the data-8 and -11. for caspase-3 have been published previously [24].
Of the caspases investigated, only caspase-2 showed The stroke model used in this study involved thevery little change in expression at any of the time points permanent occlusion of the middle cerebral artery at itstested (Fig. 5g and h). origin from the circle of Willis by an intraluminal filament
Variance components analyses indicate a consistent [68]. This procedure is widely used and has been shown tocontribution by the RT replication to the variability, give a highly reproducible effect as assessed by a varietyregardless of whether samples are from individuals or of criteria, including reduction of cerebral blood flow [67],pooled (Table 2). Values range between 0.0028 (caspase- infarct volume and position [53,67] and motor impairment11, individual samples) and 0.0163 (caspase-6, pooled). [53]. The infarct produced by this procedure, at its greatestThere are some differences in the contribution of RT extent involves the ipsilateral cortex and striatum, corre-replication between genes, but even these are within the sponding to the territory perfused by the middle cerebralsame degree of magnitude. The largest contribution to the artery. In this study we confined our analysis of genevariability, however, arose from plate effects, as demon- expression to the cerebral cortex.
¨strated in the analysis on the data from naıve rats. The caspase family has been subdivided according todifferent parameters such as sequence homology, substratespecificity and structure [7,12,51]. On the basis of theirstructure, the caspases fall into two groups according to the
4. Discussion size of the N-terminal prodomain. Type I caspases havelarge prodomains which typically contain motifs that allow
In this study we have examined the expression of the binding of procaspases to other molecules to formmRNA encoding eight members of the caspase family in a signalling complexes. These include the death effector

D.C. Harrison et al. / Molecular Brain Research 89 (2001) 133 –146 139
Fig. 2. Expression of mRNA encoding caspase-8 in the cerebral cortex following permanent occlusion of the middle cerebral artery. (a) mRNA expressionin ipsi- and contralateral cortex of MCAO and sham-operated rats plotted against time. Mean expression in each group is shown as a fold increase
¨compared to mean expression in naıve rats which has been ascribed an arbitrary value of 1 as indicated by the horizontal line. (b) Mean expression levels at¨each time point with error bars representing 95% confidence intervals for the comparison of each group with naıve rats. A change in expression from the
¨naıve level is indicated by error bars not crossing the horizontal line. (c) Comparison of mRNA expression in ipsi- and contralateral cortex of MCAO ratsat each time point. Contralateral expression is given an arbitrary value equal to 1, and ipsilateral expression is shown as a fold increase over contralateral.(d) Comparison of mRNA expression in ipsilateral (left) MCAO cortex with the left cortex in sham-operated rats. Expression in sham-operated rats isgiven an arbitrary value equal to 1, and MCAO expression is shown as a fold increase over sham.
domain (DED) of caspase-8 (and caspase-10) which is signalling pathways, and one, caspase-3, because it isinvolved in the formation of a death-inducing signalling thought to be a key effector caspase acting downstream.complex (DISC), together with the adapter molecule Data from pooled RNA samples for these caspases is notFADD and the death receptor Fas [28]. Similarly, the reported. Table 2 shows the sources of random variabilitycaspase recruitment domain (CARD) of caspase-9 allows arising between animals, between samples within animalsthe formation of the apoptosome, together with the mole- and between RT-PCR replicates from the same sample.cules Apaf-1 and cytochrome c [69]. Activation of type I These data are compared with variability between repli-caspases occurs upon formation of such complexes, which cates from pooled RNA samples.autocatalyse cleavage of the caspase to initiate further It is critical to assess changes over time in sham and
¨caspase signalling. In contrast, type II caspases possess a MCAO groups relative to the naıve group as strong platemuch shorter prodomain. Activation of these caspases effects were observed for most genes (Table 2). Thisoccurs due to cleavage, principally by other caspases. enables the time effect to be studied independently of anyWhile there may be commonality in the signalling pathway plate effects. When designing studies which require theat the level of type II caspases, the type I caspases utilise division of samples between multiple plates, it is essentialdistinct pathways involved in the initiation of apoptosis. that this is taken into consideration.mRNA expression of all eight caspases was examined Naturally, additional variation is introduced betweenusing pooled RNA samples, in the interests of establishing animals and between samples within animals. These area rapid method for screening mRNA expression in disease not insubstantial (Table 2) and therefore the appropriatemodels. In addition, we studied the mRNA expression of error term incorporating these must be utilised. For exam-four caspases in detail, using RNA samples derived from ple, the different experimental procedures applied toindividual rats. Three of these (caspases-8, -9 and -11) individual animals should be assessed relative to errorwere chosen as representatives of the principal caspase composed of all three types of variation.
140 D.C. Harrison et al. / Molecular Brain Research 89 (2001) 133 –146
Fig. 3. Expression of mRNA encoding caspase-11 in the cerebral cortex following permanent occlusion of the middle cerebral artery. (a) mRNAexpression in ipsi- and contralateral cortex of MCAO and sham-operated rats plotted against time. Mean expression in each group is shown as a fold
¨increase compared to mean expression in naıve rats which has been ascribed an arbitrary value of 1 as indicated by the horizontal line. (b) Mean expression¨levels at each time point with error bars representing 95% confidence intervals for the comparison of each group with naıve rats. A change in expression
¨from the naıve level is indicated by error bars not crossing the horizontal line. (c) Comparison of mRNA expression in ipsi- and contralateral cortex ofMCAO rats at each time point. Contralateral expression is given an arbitrary value equal to 1, and ipsilateral expression is shown as a fold increase overcontralateral. (d) Comparison of mRNA expression in ipsilateral (left) MCAO cortex with the left cortex in sham-operated rats. Expression insham-operated rats is given an arbitrary value equal to 1, and MCAO expression is shown as a fold increase over sham.
4.1. Caspase-3 caspase it is activated by a number of other upstreamcaspases and is responsible for the cleavage of a large
Previously, we have shown that caspase-3 mRNA is number of substrates whose proteolysis occurs in manyupregulated following pMCAO in rats when compared apoptotic systems. As such it may be regarded as one of
¨with the expression seen in naıve rats. Here we have taken the key executioners of apoptosis [7]. Its importance in thethe analysis further to show that there is also a significant CNS is demonstrated by the profound hyperplasia duringupregulation in caspase-3 mRNA in ischemic cortex versus brain development observed in caspase-3 deficient micethe contralateral side, and the cortex of sham-operated [34] and the continued high level of expression of caspase-animals. 3 in the adult brain [50].
Changes in the expression of some genes, for example Increased caspase-3 activity has been demonstrated inthe death receptor TR3, at the level of messenger RNA vulnerable cells, for example CA1 hippocampal neurons,have been observed following the sham operational pro- following transient global ischemia [6,20,48], and in-cedure in this model [25]. In view of the invasive nature of creased neuronal caspase-3 expression is observed inthe sham surgical procedure, which is likely to result in transient [49] and permanent [32] focal ischemia. There isconsiderable perturbations in cerebral blood flow, some a body of evidence, therefore, to suggest that caspase-3 ischanges in gene expression are to be expected, and for this upregulated and activated in neurons following cerebralreason it was decided to use a number of different control ischemia. In a permanent model of focal ischemia [32],tissues for comparison. increased procaspase-3 immunoreactivity was observed in
Caspase-3 is the mammalian caspase with the closest microglia and astrocytes. This may be due either tohomology to the C. elegans CED-3 protein. As a type II induction of caspase-3 or proliferation of caspase-3 ex-
D.C. Harrison et al. / Molecular Brain Research 89 (2001) 133 –146 141
Fig. 4. Expression of mRNA encoding caspase-3 in the cerebral cortex following permanent occlusion of the middle cerebral artery. (a) mRNA expressionin ipsi- and contralateral cortex of MCAO and sham-operated rats plotted against time. Mean expression in each group is shown as a fold increase
¨compared to mean expression in naıve rats which has been ascribed an arbitrary value of 1 as indicated by the horizontal line. (b) Mean expression levels at¨each time point with error bars representing 95% confidence intervals for the comparison of each group with naıve rats. A change in expression from the
¨naıve level is indicated by error bars not crossing the horizontal line. (c) Comparison of mRNA expression in ipsi- and contralateral cortex of MCAO ratsat each time point. Contralateral expression is given an arbitrary value equal to 1, and ipsilateral expression is shown as a fold increase over contralateral.(d) Comparison of mRNA expression in ipsilateral (left) MCAO cortex with the left cortex in sham-operated rats. Expression in sham-operated rats isgiven an arbitrary value equal to 1, and MCAO expression is shown as a fold increase over sham.
pressing cells. A similar mechanism could also partially be mRNA expression at 3 h which, as it occurs in both sidesresponsible for the increase in caspase-3 mRNA observed of the cortex, suggests that it is due to an acute after-effectin this study. of the MCAO procedure, such as a stress response,
although the mechanism involved is unclear. It is worth4.2. Caspase-8 noting that this global effect would not have been detect-
able had either contralateral cortex or sham-operatedCaspase-8 can be activated through the Fas and TNF animals been used alone as controls.
receptors binding to their ligands at the cell surface [28].Two studies, in models similar to the one we have 4.3. Caspase-11, caspase-1employed, have shown that caspase-8 is found constitu-tively in neurons, and has increased expression following Caspase-11 was originally identified and characterised inischemia [32,60]. Increased expression of caspase-8 may mice and has closest homology to human caspase-4increase the availability of protein to be recruited to form [59,62]. Like caspase-4 in humans, it is essential for theDISCs, thus contributing to an increase in apoptotic activation of caspase-1 [63]. Caspase-1, originally termedsignalling. Significantly, mRNAs for both Fas and the type interleukin-1b converting enzyme (ICE), is involved in theI TNF receptor are also upregulated in this model [25]. In processing of interleukin-1b from its precursor.this study, caspase-8 shows an early increase in expression Thymocytes derived from caspase-1 deficient mice, whilewith a similar time course to that which we have previous- responding normally to ultraviolet radiation and dexa-ly shown for caspase-3 [24]. However, there is some methasone, are resistant to apoptosis induced by Fasevidence to suggest that increased caspase-3 and caspase-8 antibody [33]. Thus, under certain circumstances caspase-1expression occur in different sub-populations of neurons is required for apoptosis, however caspase-1 and caspase-[60]. 11 knockout mice develop normally [33,37,63].
We also observe a transient decrease in caspase-8 Elevated expression of the pro-inflammatory cytokine
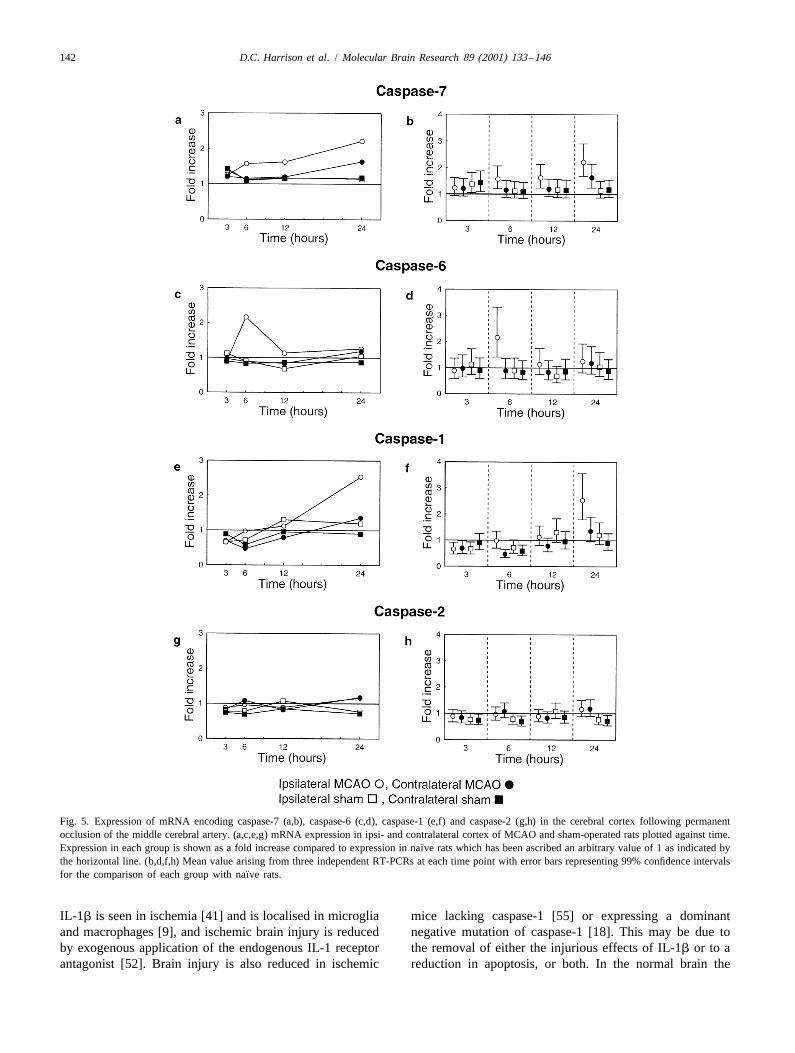
142 D.C. Harrison et al. / Molecular Brain Research 89 (2001) 133 –146
Fig. 5. Expression of mRNA encoding caspase-7 (a,b), caspase-6 (c,d), caspase-1 (e,f) and caspase-2 (g,h) in the cerebral cortex following permanentocclusion of the middle cerebral artery. (a,c,e,g) mRNA expression in ipsi- and contralateral cortex of MCAO and sham-operated rats plotted against time.
¨Expression in each group is shown as a fold increase compared to expression in naıve rats which has been ascribed an arbitrary value of 1 as indicated bythe horizontal line. (b,d,f,h) Mean value arising from three independent RT-PCRs at each time point with error bars representing 99% confidence intervals
¨for the comparison of each group with naıve rats.
IL-1b is seen in ischemia [41] and is localised in microglia mice lacking caspase-1 [55] or expressing a dominantand macrophages [9], and ischemic brain injury is reduced negative mutation of caspase-1 [18]. This may be due toby exogenous application of the endogenous IL-1 receptor the removal of either the injurious effects of IL-1b or to aantagonist [52]. Brain injury is also reduced in ischemic reduction in apoptosis, or both. In the normal brain the

D.C. Harrison et al. / Molecular Brain Research 89 (2001) 133 –146 143
localisation of caspase-1 and IL-1b has been shown to Caspase-6 has been shown to activate caspase-3 [42],reside in cells of the vascular system [64] and in neurons. and in Jurkat cells, caspase-9 dependent activation ofFollowing transient global ischemia in gerbils, a model caspases-8 and -10 is blocked by inhibition of caspase-6where neuronal death is observed, microglial expression of [57]. Thus the role of caspase-6 may not simply be as acaspase-1 is induced, but neuronal expression does not late stage ‘executioner’ caspase, but may be more com-change [4]. Microglial caspase-1 immunoreactivity has plex. An increased requirement for caspase-6 at an earlyalso been described in the corpus callosum, but not in the time point may indicate its involvement in such a mecha-infarct or penumbra in a permanent model of focal nism. Interestingly, a transient increase in caspase-6 im-ischemia [32]. Again, this points to the major effect of the munoreactivity in astrocytes has been reported in perma-caspase-11/caspase-1 pathway being pro-inflammatory in nent focal ischemia [32]. This effect did not occur until 12the ischemic brain. to 24 h after surgery, however the model employed was
The increase in contralateral expression of caspase-11 at less severe than the one in our study and it is therefore not24 h is in keeping with this hypothesis. Elevated levels of possible to compare the two time courses directly.IL-1b protein start to appear in the contralateral hemi- Caspase-7 has not been reported to participate in anysphere at a late stage in this model. Increased caspase-1 feedback mechanism in the same manner as caspase-6. Itsand caspase-11 would be expected for the processing of late increase in expression would suggest that its role inthis molecule. Again, detection of this contralateral change ischemia induced apoptosis is limited to cleavage ofis only made possible by inclusion of all of the appropriate proteins downstream of the caspase pathway, or that it iscontrols in the study. found in a different population of cells whose requirement
for caspase transcription occurs at a different stage in the4.4. Caspase-9 overall progression of the developing lesion.
Caspase-9 can be activated by apoptotic stimuli which 4.6. Caspase-2do not activate caspase-8; cells derived from mice lackingcaspase-9 undergo cell death triggered by Fas, but are Our data in this study show no overall changes in theresistant to apoptosis induced by dexamethasone or gamma expression of mRNA for caspase-2. This protein exists inradiation [21]. Caspase-9 knockout mice display CNS two isoforms, long and short, encoded by alternatelymalformations and caspase-9 is constitutively expressed in spliced variants of the mRNA. The long and short forms ofneurons both during development and in the adult, indicat- caspase-2 act as positive and negative regulators of apop-ing an important role for caspase-9 in neuronal apoptosis tosis, respectively [61]. The primers and probe we used are[21,35]. specific for the long, pro-apoptotic form of caspase-2. In
We have detected a decrease in caspase-9 expression models of both focal [1] and global ischemia [29], afollowing cerebral ischemia. If caspase-9 is activated under transient increase in mRNA for both isoforms of caspase-2ischemic conditions, the levels of this protein normally mRNA has been reported. The different results seen herefound in cells undergoing apoptosis must be sufficient for may be due to the precise model employed or, in the caseapoptosome formation and for signalling to occur normal- of the focal ischemia study, to the tissues sampled; ourly. The decrease observed here suggests either a loss of study was restricted to the cerebral cortex, whereas Asahicaspase-9 expressing cells in the region affected, or a et al. sampled the whole cerebral hemisphere. Sincerelative decrease in its expression. The appearance of different regions of the brain are prone to apoptosis todecreased caspase-9 as early as 3 h post-MCAO, when differing degrees it is likely that changes in caspasethere is little evidence of any major changes in the cellular expression will also differ between one region and another.composition of the infarct, would point to the latter Kinoshita et al. [29], using a gerbil model of transientscenario. If this caspase has a role in apoptosis in cerebral global ischemia, report a greater increase in the CA1–4ischemia it appears that this employs the caspase-9 already fields of the hippocampus which readily undergo apop-present, and that little or no de novo synthesis of caspase-9 tosis, than in cortex which is more resistant to cell deathis required. [30]. In caspase-2 deficient mice there is no reduction in
neurological damage in models of either permanent focal4.5. Caspases-6 and -7 or transient ischemia when compared with wild type
littermates [3].Caspases-6 and -7 are both type two caspases with close When evaluating changes in gene expression in an in
homology to caspase-3. Both have been reported to be vivo situation such as the one described in this study it isexpressed at very low levels, if not absent, in normal brain important to consider any changes taking place in cellular[59], therefore any involvement of these caspases is likely composition of the tissue. Following focal cerebral is-to require de novo synthesis of mRNA and protein. Our chemia a number of substantial changes take place in thedata show increased transcription of both caspases, but affected area. Neurons and oligodendrocytes die within thewith different time courses. ischemic infarct [2,45], particularly after 12 h of ischemia.

144 D.C. Harrison et al. / Molecular Brain Research 89 (2001) 133 –146
Astrocytes and microglia are decreased in number in the although their importance as mediators of inflammation viacore region of the lesion and proliferation of both of these IL-1b would suggest a different or additional role.cell types occurs in the marginal areas [8]. Macrophages Caspases-2 and -9, which lie on separate pathways,and polymorphonuclear leukocytes invade the margins of would appear not to be under control of such a program,the lesion at a late stage, after around 24 h of ischemia although the lack of increased transcription does not rule[8,31]. These changes become most pronounced only after out an important role for these enzymes in this model of12 to 24 h of ischemia. At earlier time points, therefore, ischemia.changes in observed mRNA expression are likely to be In conclusion, we have presented a detailed analysis ofrepresentative of regulation of gene expression within the temporal changes in caspase mRNA expression follow-normal tissue, whereas later differences in mRNA levels ing permanent occlusion of the middle cerebral artery. Thisare more likely to be due to overall changes in the cellular has revealed distinct patterns of transcription pointing tocomposition of the tissue sample. the roles of different pathways and individual caspases
Studies which examine expression at the mRNA level during the development of the lesion. Such informationsuffer from the inherent disadvantage that changes seen in provides a platform for the development of novel drugmRNA do not necessarily correspond to changes seen in targets and gives insight into time windows when com-protein expression. Furthermore, it has been suggested that pounds designed against such targets could be effective.the coupling of mRNA transcription to protein synthesis isin some way disrupted under ischemic conditions. Whilethere can be no doubt that normal physiological function is Referencesdisturbed following ischemia, protein synthesis may stillbe important because its inhibition blocks the development [1] M. Asahi, M. Hoshimaru, Y. Uemura, T. Tokime, M. Kojima, T.
Ohtsuka, N. Matsuura, T. Aoki, K. Shibahara, H. Kikuchi, Expres-of the ischemic infarct [40].sion of interleukin-1 converting enzyme gene family and bcl-2 geneOur results suggest that the transcriptional activationfamily in rat brain following permanent occlusion of the middle
patterns of the eight caspases studied fall into three (or cerebral artery, J. Cereb. Blood Flow Metab. 17 (1997) 11–18.possibly four) groups. In the first group, a modest increase [2] R.T. Bartus, R.L. Dean, K. Cavanaugh, D. Eveleth, D.L. Carriero,in expression is observed at 6 h post-MCAO and continues G. Lynch, Time related neuronal changes following middle cerebral
artery occlusion: implications for therapeutic intervention and theto rise. This pattern includes caspases-3, -7 and -8 and -11.role of calpain, J. Cereb. Blood Flow Metab. 15 (1995) 969–979.Whereas caspases-3 and -7 are thought to represent type II
[3] L. Bergeron, G.I. Perez, G. MacDonald, L. Shi, Y. Sun, A.caspases, caspase 8 (and possibly caspase-11) are type I Jurisicova, S. Varmuza, K.E. Latham, J.A. Flaws, J.C.M. Salter, H.caspases. In fact, the activation mechanism of caspase-11 Hara, M.A. Moskowitz, E. Li, A. Greenberg, J.L. Tilly, J. Yuan,is unknown, although its large prodomain suggests a type-I Defects in regulation of apoptosis in caspase-2-deficient mice, Genes
Dev. 12 (1998) 1304–1314.mechanism. Caspase-1 may also fit into this group, al-[4] R.V. Bhat, R. DiRocco, V.R. Marcy, D.G. Flood, Y. Zhu, P.though its increase in expression is delayed compared with
Dobrzanski, R. Siman, R. Scott, P.C. Contreras, M. Miller, Increasedcaspases-3, -7, -8 and -11. Caspase-6 appears to be expression of IL-1b converting enzyme in hippocampus afterseparately grouped because its mRNA increased only ischemia: selective localisation in microglia, J. Neurosci. 16 (1996)transiently at 6 h. Caspases-2 and -9 form a third grouping 4146–4154.
[5] C. Bowen, H.J. Voeller, K. Kikly, E.P. Gelmann, Synthesis ofin that their mRNA was unchanged or even reduced duringprocaspases-3 and -7 during apoptosis in prostate cancer cells, Cellthe time course of the experiment. These results couldDeath Differ. 6 (1999) 394–401.
suggest that the transcription of these enzymes is under the [6] J. Chen, T. Hagayama, K. Jin, R.A. Stetler, R.L. Zhu, S.H. Graham,control of distinct mechanisms. In view of the finding that R.P. Simon, Induction of caspase-3-like protease may mediatemRNA transcription and translation are required for apop- delayed neuronal death in the hippocampus after transient cerebral
ischemia, J. Neurosci. 18 (1998) 4914–4928.tosis to take place [46], these findings raise interesting[7] G.M. Cohen, Caspases: the executioners of apoptosis, Biochem. J.possibilities regarding possible genetic programs activated
326 (1997) 1–16.in this model. [8] C.A. Davies, S.A. Loddick, R.P. Stroemer, J. Hunt, N.J. Rothwell,
Caspases-8 and -10 are the key initiator caspases in TNF An integrated analysis of the progression of cell responses inducedand Fas-induced apoptosis [28]. Caspases-3 and -7 can by permanent middle cerebral artery occlusion in the rat, Exp.
Neurol. 154 (1998) 199–212.both be activated by caspase-8 [58] and caspase-10 [16].[9] C.A. Davies, S.A. Loddick, S. Toulmond, R.P. Stroemer, J. Hunt,Furthermore, both of these enzymes are activated them-
N.J. Rothwell, The progression and topographic distribution ofselves during TNF and Fas-induced apoptosis [11,56]. interleukin-1b expression after permanent middle cerebral arterymRNA for Fas and TNF is substantially increased in focal occlusion in the rat, J. Cereb. Blood Flow Metab. 19 (1999) 87–98.ischemia [25] and Fas has been shown to play an important [10] T.L. Deckwerth, E.M. Johnson Jr., Temporal analysis of events
associated with programmed cell death (apoptosis) of sympatheticrole in neuronal apoptosis in vivo and in focal ischemianeurons deprived of nerve growth factor (NGF), J. Cell Biol. 123[47]. The concurrent increases in the mRNA for caspases-(1993) 1207–1222.
8, -3 and -7 may represent an apoptotic program for a [11] H. Duan, A.M. Chinnaiyan, P.L. Hudson, J.P. Wing, W.-W. He, V.M.pathway that clearly has an important role in this model. Dixit, ICE-LAP3, a novel mammalian homologue of the Caenor-
Caspases-1 and -11 may also be part of this program, habditis elegans cell death protein Ced-3 is activated during Fas-

D.C. Harrison et al. / Molecular Brain Research 89 (2001) 133 –146 145
and tumor necrosis factor-induced apoptosis, J. Biol. Chem. 271 caspase-8 /FLICE in the initiation of the Fas-induced apoptotic(1996) 1621–1625. cascade, Curr. Biol. 8 (1998) 1001–1008.
[12] W.C. Earnshaw, L.M. Martins, S.H. Kaufmann, Mammalian cas- [29] M. Kinoshita, H. Tomimoto, A. Kinoshita, S. Kumar, M. Noda,pases: structure, activation, substrates and functions during apop- Up-regulation of the nedd2 gene encoding an ICE/Ced-3-liketosis, Annu. Rev. Biochem. 68 (1999) 383–424. cysteine protease in the gerbil brain after transient global ischemia,
J. Cereb. Blood Flow Metab. 17 (1997) 507–514.[13] B.A. Eldadah, A.G. Yakovlev, A.I. Faden, The role of CED-3 relatedcysteine proteases in apoptosis of cerebellar granule cells, J. [30] T. Kirino, Delayed neuronal death in the gerbil hippocampusNeurosci. 17 (1997) 6105–6113. following ischemia, Brain Res. 239 (1982) 57–69.
[14] M. Endres, S. Namura, M. Shimuzu-Sasamata, C. Waeber, L. Zhang, [31] P.M. Kochanek, J.M. Hallenbeck, Polymorphonuclear leukocytesT. Gomez-Isla, B.T. Hyman, M.A. Moskowitz, Attenuation of and monocytes /macrophages in the pathogenesis of cerebral is-delayed neuronal death after mild focal ischemia in mice by chemia and stroke, Stroke 23 (1992) 1367–1379.inhibition of the caspase family, J. Cereb. Blood Flow Metab. 18 [32] J. Krupinski, E. Lopez, E. Marti, I. Ferrer, Expression of caspases(1998) 238–247. and their substrates in the rat model of focal cerebral ischemia,
[15] S. Estus, W.J. Zaks, R.S. Freeman, M. Gruda, R. Bravo, E.M. Neurobiol. Dis. 7 (2000) 332–342.Johnson Jr., Altered gene expression in neurons during programmed [33] K. Kuida, J.A. Lippke, G. Ku, M.W. Harding, D.J. Livingston,cell death: identification of c-jun as necessary for neuronal apop- M.S.-S. Su, R.A. Flavell, Altered cytokine export and apoptosis intosis, J. Cell Biol. 127 (1994) 1717–1727. mice deficient in interleukin-1b converting enzyme, Science 267
[16] T. Fernandes-Alnemri, R.C. Armstrong, J. Krebs, S.M. Srinivasula, (1995) 2000–2003.L. Wang, F. Bullrich, L.C. Fritz, J.A. Trapani, K.J. Tomaselli, G. [34] K. Kuida, T.S. Zheng, S. Na, C. Kuan, D. Yang, H. Karasuyama, P.Litwack, E.S. Alnemri, In vitro activation of CPP32 and Mch3 by Rakic, R.A. Flavell, Decreased apoptosis in the brain and prematureMch4, a novel human apoptotic cysteine protease containing two lethality in CPP32-deficient mice, Nature 384 (1996) 368–372.FADD-like domains, Proc. Natl. Acad. Sci. USA 93 (1996) 7464– [35] K. Kuida, T.F. Haydar, C.-Y. Kuan, Y. Gu, C. Taya, H. Karasuyama,7469. M.S.-S. Su, P. Rakic, R.A. Flavell, Reduced apoptosis and cyto-
[17] K. Fink, J. Zhu, S. Namura, M. Shimuzu-Sasamata, M. Endres, J. chrome c-mediated caspase activation in mice lacking caspase-9,Ma, T. Dalkara, J. Yuan, M.A. Moskowitz, Prolonged therapeutic Cell 94 (1998) 325–337.window for ischemic brain damage caused by delayed caspase [36] J.-M. Lee, G.J. Zipfel, D.W. Choi, The changing landscape ofactivation, J. Cereb. Blood Flow Metab. 18 (1998) 1071–1076. ischaemic brain injury mechanisms, Nature 399 (Suppl.) (1999)
[18] R.M. Friedlander, V. Gagliardini, H. Hara, K.B. Fink, W. Li, G. A7–A14.MacDonald, M.C. Fishman, A.H. Greenberg, M.A. Moskowitz, J. [37] P. Li, H. Allen, S. Banerjee, S. Franklin, L. Herzog, C. Johnston, J.Yuan, Expression of a dominant negative mutant of interleukin-1b McDowell, M. Paskind, L. Rodman, J. Salfeld, E. Towne, D.converting enzyme in transgenic mice prevents neuronal cell death Tracey, S. Wardwell, F.-Y. Wei, W. Wong, R. Kamen, T. Seshadri,induced by trophic factor withdrawal and ischemic brain injury, J. Mice deficient in IL-1b converting enzyme are defective in pro-Exp. Med. 185 (1997) 933–940. duction of mature IL-1b and resistant to endotoxic shock, Cell 80
[19] U.E.M. Gibson, C.A. Heid, P.M. Williams, A novel method for real (1995) 401–411.time quantitative RT-PCR, Genome Methods 6 (1996) 995–1001. [38] Y. Li, M. Chopp, N. Jiang, F. Yao, C. Zaloga, Temporal profile of in
¨[20] F. Gillardon, B. Bottiger, B. Schmitz, M. Zimmermann, K.-A. situ DNA fragmentation after transient middle cerebral arteryHossmann, Activation of CPP-32 protease in hippocampal neurons occlusion in the rat, J. Cereb. Blood Flow Metab. 15 (1995)following ischemia and epilepsy, Brain Res. Mol. Brain Res. 50 389–397.(1997) 16–22. [39] Y.S. Lie, C.J. Petropoulos, Advances in quantitative PCR technolo-
[21] R. Hakem, A. Hakem, G.S. Duncan, J.T. Henderson, M. Woo, M.S. gy: 59 nuclease assays, Curr. Opin. Biotechnol. 9 (1998) 43–48.Soenges, A. Elia, J.L. De la pompa, D. Kaji, W. Khoo, J. Potter, R. [40] M.D. Linnik, R.H. Zobrist, M.D. Hatfield, Evidence supporting aYoshida, S.A. Kaufman, S.W. Lowe, J.M. Penninger, T.W. Mak, role for programmed cell death in focal ischemia in rats, Stroke 24Differential requirement for Caspase-9 in apoptotic pathways in (1993) 2002–2009.vivo, Cell 94 (1998) 339–352. [41] T. Liu, P.C. McDonnell, P.R. Young, R.F. White, A.L. Siren, J.M.
[22] H. Hara, K. Fink, M. Endres, R.M. Friedlander, V. Gagliardini, J. Hallenbeck, F.C. Barone, G.Z. Feuerstein, Interleukin-1 beta mRNAYuan, M.A. Moskowitz, Attenuation of transient focal ischemic expression in ischemic rat cortex, Stroke 24 (1993) 1746–1750.injury in transgenic mice expressing a mutant ICE inhibitory protein, [42] X. Liu, C.N. Kim, J. Pohl, X. Wang, Purification and characterisa-J. Cereb. Blood Flow Metab. 17 (1997) 370–375. tion of an interleukin-1beta-converting enzyme family protease that
[23] H. Hara, R.M. Friedlander, V. Gagliardini, C. Ayata, K. Fink, Z. activates cysteine protease P32, J. Biol. Chem. 271 (1996) 13371–Huang, M. Shimuzu-Sasamata, J. Yuan, M.A. Moskowitz, Inhibition 13376.of interleukin-1b converting enzyme family proteases reduces [43] S.A. Loddick, A. MacKenzie, N.J. Rothwell, An ICE inhibitorischemic and excitotoxic neuronal damage, Proc. Natl. Acad. Sci. z-VAD-DCB attenuates ischaemic brain damage in the rat, NeuroRe-USA 94 (1997) 2007–2012. port 7 (1996) 1465–1468.
[24] D.C. Harrison, A.D. Medhurst, B.C. Bond, C.A. Campbell, R.P. [44] J.P. MacManus, I.E. Hill, Z.-G. Huang, I. Rasquinha, D. Xue, A.M.Davis, K.L. Philpott, The use of quantitative RT-PCR to measure Buchan, DNA damage consistent with apoptosis in transient focalmRNA expression in a rat model of focal ischemia — caspase-3 as a ischemic neocortex, NeuroReport 5 (1994) 493–496.case study, Brain Res. Mol. Brain Res. 75 (2000) 143–149. [45] K. Mandai, M. Matsumoto, K. Kitagawa, K. Matsushita, T. Ohtsuki,
[25] D.C. Harrison, J. Roberts, C.A. Campbell, B. Crook, R. Davis, K. T. Mabuchi, D.R. Colman, T. Kamada, T. Yanagihara, IschemicDeen, J. Meakin, D. Michalovich, J. Price, M. Stammers, P.R. damage and subsequent proliferation of oligodendrocytes in focalMaycox, TR3 death receptor expression in normal and ischaemic cerebral ischemia, Neuroscience 77 (1997) 849–861.brain, Neuroscience 96 (2000) 147–160. [46] D.P. Martin, R.E. Schmidt, P.S. DiStefano, O.H. Lowry, J.G. Carter,
[26] C.A. Heid, J. Stevens, K.J. Livak, P.M. Williams, Real time E.M. Johnson Jr., Inhibitors of RNA synthesis and protein synthesisquantitative PCR, Genome Methods 6 (1996) 986–994. prevent neuronal death caused by nerve growth factor deprivation, J.
[27] J. Honkaniemi, S.M. Massa, M. Breckinridge, F.R. Sharp, Global Cell Biol. 106 (1988) 829–844.ischemia induces apoptosis-associated genes in hippocampus, Brain [47] A. Martin-Villalba, I. Herr, I. Jeremias, M. Hahne, R. Brandt, J.Res. Mol. Brain Res. 42 (1996) 79–88. Vogel, J. Schenkel, T. Herdegen, K.-M. Debatin, CD95 ligand
[28] P. Juo, C.J. Kuo, J. Yuan, J. Blenis, Essential requirement for (Fas-L/APO-1L) and tumor necrosis factor-related apoptosis-induc-

146 D.C. Harrison et al. / Molecular Brain Research 89 (2001) 133 –146
ing ligand mediate ischemia-induced apoptosis in neurons, J. Fas /APO-1 protease Mch5 is a CrmA-inhibitable protease thatNeurosci. 19 (1999) 3809–3817. activates multiple Ced-3 / ICE-like cysteine proteases, Proc. Natl.
[48] S. Namura, J. Zhu, K. Fink, M. Endres, A. Srinivasan, K.J. Acad. Sci. USA 93 (1996) 14486–14491.Tomaselli, J. Yuan, M.A. Moskowitz, Activation and cleavage of [59] M. Van de Craen, P. Vandenabeele, W. Declercq, I. Van den Brande,caspase-3 in apoptosis induced by experimental cerebral ischemia, J. G. Van Loo, F. Molemans, P. Schotte, W. Van Criekinge, R. Beyaert,Neurosci. 18 (1998) 3659–3668. W. Fiers, Characterisation of seven murine caspase family members,
[49] B. Ni, X. Wu, Y. Su, D. Stephenson, E.B. Smalstig, J. Clemens, S.M. FEBS Lett. 403 (1997) 61–69.Paul, Transient global forebrain ischemia induces a prolonged [60] J.J. Velier, J.A. Ellison, K.K. Kikly, P.A. Spera, F.C. Barone, G.Z.expression of the caspase-3 mRNA in rat hippocampal CA1 Feuerstein, Caspase-8 and Caspase-3 are expressed by differentpyramidal neurons, J. Cereb. Blood Flow Metab. 18 (1998) 248– populations of cortical neurons undergoing delayed cell death after256. focal stroke in the rat, J. Neurosci. 19 (1999) 5932–5941.
[50] B. Ni, X. Wu, Y. Du, Y. Su, E. Hamilton-Byrd, P.K. Rockey, P. [61] L. Wang, M. Miura, L. Bergeron, H. Zhu, J. Yuan, Ich-1, anRosteck Jr., G.G. Poirier, S.M. Paul, Cloning and expression of a rat Ice /ced-3-related gene, encodes both positive and negative reg-brain interleukin-1b-converting enzyme (ICE)-related protease ulators of programmed cell death, Cell 78 (1994) 739–750.(IRP) and its possible role in apoptosis of cultured cerebellar [62] S. Wang, M. Miura, H. Zhu, V. Gagliardini, J. Yuan, Identificationgranule neurons, J. Neurosci. 17 (1997) 1561–1569. and characterisation of caspase-11, a member of the ICE/Ced-3
[51] D.W. Nicholson, N.A. Thornberry, Caspases: killer proteases, Trends family and an upstream regulator of ICE, J. Biol. Chem. 271 (1996)Biochem. Sci. 22 (1997) 299–306. 20580–20587.
[52] J.K. Relton, N.J. Rothwell, Interleukin-1 receptor antagonist inhibits [63] S. Wang, M. Miura, Y.-K. Jung, H. Zhu, E. Li, J. Yuan, Murineischemic and excitotoxic neuronal damage in the rat, Brain Res. caspase-11, an ICE-interacting protease, is essential for the activa-Bull. 29 (1992) 243–246. tion of ICE, Cell 92 (1998) 501–509.
[53] D.C. Rogers, C.A. Campbell, J.L. Stretton, K.B. Mackay, Correla- [64] M.-L. Wong, P.B. Bongiorno, P.W. Gold, J. Licinio, Localization oftion between motor impairment and infarct volume after permanent interleukin-1beta converting enzyme mRNA in rat brain vasculature:and transient middle cerebral artery occlusion in the rat, Stroke 28 evidence that the genes encoding the interleukin-1 system are(1997) 2060–2066. constitutively expressed in brain blood vessels, Neuroimmunomodu-
[54] P. Rong, A.M. Bennie, W.R. Epa, G.L. Barrett, Nerve growth factor lation 2 (1995) 141–148.determines survival and death of PC12 cells by regulation of the [65] J. Yao, R.W. Johnson, Induction of interleukin-1b-converting en-bcl-x, bax and caspase-3 genes, J. Neurochem. 72 (1999) 2294– zyme (ICE) in murine microglia by lipopolysaccharide, Brain Res.2300. Mol. Brain Res. 51 (1997) 170–178.
[55] G.P. Schielke, G.-Y. Yang, B.D. Shiver, A.L. Betz, Reduced is- [66] J. Yuan, S. Shaham, S. Ledoux, H.M. Ellis, H.R. Horvitz, The C.chemic brain injury in interleukin-1b converting enzyme-deficient elegans cell death gene ced-3 encodes a protein similar to mam-mice, J. Cereb. Blood Flow Metab. 18 (1998) 180–185. malian interleukin-1b-converting enzyme, Cell 75 (1993) 641–652.
[56] J. Schlegel, I. Peters, S. Orrenius, D.K. Miller, N.A. Thornberry, [67] G.J. Zarow, H. Karibe, B.A. States, S.H. Graham, P.R. Weinstein,T.-T. Yamin, D.W. Nicholson, CPP32/apopain is a key interleukin- Endovascular suture occlusion of the middle cerebral artery in rats:1b converting enzyme-like protease involved in Fas-mediated effect of suture insertion distance on cerebral blood flow, infarctapoptosis, J. Biol. Chem. 271 (1995) 1841–1844. distribution and infarct volume, Neurol. Res. 19 (1997) 409–416.
[57] E.A. Slee, M.T. Harte, R.M. Kluck, B.B. Wolf, C.A. Casiano, D.D. [68] E. Zea-Longa, P.R. Weinstein, S. Carlson, R. Cummins, ReversibleNewmeyer, H.-G. Wang, J.C. Reed, D.W. Nicholson, E.S. Alnemri, middle cerebral artery occlusion without craniectomy in rats, StrokeD.R. Green, S.J. Martin, Ordering the cytochrome c-initiated caspase 20 (1989) 84–91.cascade: hierarchical activation of caspases-2, -3, -6, -7, -8, and -10 [69] H. Zou, W.J. Henzel, X. Liu, A. Lutschg, X. Wang, Apaf-1, a humanin a caspase-9-dependent manner, J. Cell Biol. 144 (1999) 281–292. protein homologous to C. elegans CED-4, participates in cyto-
[58] S.M. Srinivasula, M. Ahmad, T. Fernades-Alnemri, G. Litwack, E.S. chrome c-dependent activation of caspase-3, Cell 90 (1997) 405–Alnemri, Molecular ordering of the Fas-apoptotic pathway: the 413.
Copyright © 2022 FDOKUMEN




















