
Reduced Susceptibility of Magnocellular Neuroendocrine Nuclei of the Rat Hypothalamus to Transient...
12
Reduced Susceptibility of Magnocellular Neuroendocrine Nuclei of the Rat Hypothalamus to Transient Focal Ischemia Produced by Middle Cerebral Artery Occlusion Margarita C. Curra ´ s-Collazo,* Urvish B. Patel,* and Mohamed O. Hussein† *Department of Cell Biology and Neuroscience, University of California at Riverside, Riverside, California 92521 and †Department of Basic Sciences, Cleveland Chiropractic College, Los Angeles, California 90004 Received December 21, 2001; accepted July 31, 2002 Intraparenchymal injections of glutamate ana- logues into the diencephalon near the supraoptic (SON) and paraventricular nucleus (PVN) of the hypo- thalamus selectively spare magnocellular neuroendo- crine cells. In this study we investigated for the first time the susceptibility of this neuronal population to ischemia. Temporary focal ischemia was produced us- ing a three-vessel occlusion method involving unilat- eral middle cerebral artery and bilateral common ca- rotid artery occlusion (MCAO/CCAO). Most of the 3-h ischemic period was maintained without anesthesia and reversed by microclip removal of the contralat- eral common carotid artery occlusion. In one subset of rats transcardial perfusion with India ink was used to estimate the degree of ischemia produced during MCAO/CCAO in the SON, lateral magnocellular nu- cleus of the PVN (PVL), caudoputamen (CP), and fron- toparietal cortex (COR). Computer-assisted densitom- etry measurements of ink density indicated significant reductions in ink penetration in the territory of the occluded MCA within the SON (46%), PVL (45%), CP (53%), and COR (76%). In contrast, neither sham-oper- ated rats nor rats subjected to occlusion of the MCA alone showed differences in ink optical densities be- tween the sides ipsilateral and contralateral to MCAO. The other subset of rats were perfused 48 –72 h after recovery and brain sections were examined for neuro- degenerative changes. While the incidences of cere- bral and caudoputamen infarction after MCAO/CCAO were 98.4 and 52%, respectively, the histological fea- tures of the SON or PVL in ischemic rats were similar to those of control rats. Reduced susceptibility of mag- nocellular neuroendocrine cells to ischemia may be due to a number of mechanisms including neuronal resilience, neuroprotection by glia and vascular/ perivascular cells, and access to perivascular cerebro- spinal fluid. © 2002 Elsevier Science (USA) Key Words: supraoptic nucleus; paraventricular nu- cleus; survival; osmoregulation; vasopressin; excito- toxicity; stroke; neuroprotection. INTRODUCTION Severe ischemia produced by occluding an intracra- nial artery results in irreversible neurologic injury. Permanent occlusion of the middle cerebral artery last- ing 24 h or longer, for example, leads to widespread neuronal necrosis in the cerebral cortex (1). Previous studies have used varied methods to study the effects of ischemia on neurologic deficits and brain histopa- thology in the cortex and striatum (1– 6). Subcortical regions such as the thalamus and hypothalamus have received relatively less attention in this regard (7–9). In particular, the magnocellular neuroendocrine nuclei have been relatively understudied, in spite of the fact that they are supplied by the middle cerebral artery (10, 11). The neurosecretory cells of the paraventricu- lar nucleus (PVN) and the supraoptic nucleus (SON) of the hypothalamus, which produce the hormones oxyto- cin and vasopressin, are involved in body fluid and salt balance as well as cardiovascular function (12–18). Un- like neurons in the cortex or other hypothalamic re- gions, these magnocellular neurosecretory cells are re- sistant to intracranial or systemic injections of gluta- mate analogues (19 –25), indicating that they may be selectively spared during stroke as well. The objectives of this initial study were (1) to deter- mine if occlusion of the middle cerebral artery (MCA) could result in reduced perfusion in the PVN and SON and (2) to investigate whether experimental ischemia had adverse effects on these nuclei after a 48- to 72-h survival period, a period sufficient to produce neuronal necrosis (8, 26). The former goal was achieved by quan- tifying ink penetration in local blood vessels during arterial occlusion. To investigate the delayed effects of ischemia, a second group of rats were allowed 48 –72 h of recovery after MCA and bilateral common carotid artery occlusion (MCAO/CCAO) and sections from dif- ferent brain areas were examined for histopathologic changes using cresyl violet and hematoxylin/eosin staining. Comparisons were made with the cerebral Experimental Neurology 178, 268 –279 (2002) doi:10.1006/exnr.2002.8032 268 0014-4886/02 $35.00 © 2002 Elsevier Science (USA) All rights reserved.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Reduced Susceptibility of Magnocellular Neuroendocrine Nuclei of the Rat Hypothalamus to Transient...

Experimental Neurology 178, 268–279 (2002)doi:10.1006/exnr.2002.8032
Reduced Susceptibility of Magnocellular Neuroendocrine Nucleiof the Rat Hypothalamus to Transient Focal Ischemia Produced
by Middle Cerebral Artery OcclusionMargarita C. Curras-Collazo,* Urvish B. Patel,* and Mohamed O. Hussein†
*Department of Cell Biology and Neuroscience, University of California at Riverside, Riverside, California 92521 and †Department ofBasic Sciences, Cleveland Chiropractic College, Los Angeles, California 90004
Intraparenchymal injections of glutamate ana-logues into the diencephalon near the supraoptic(SON) and paraventricular nucleus (PVN) of the hypo-thalamus selectively spare magnocellular neuroendo-crine cells. In this study we investigated for the firsttime the susceptibility of this neuronal population toischemia. Temporary focal ischemia was produced us-ing a three-vessel occlusion method involving unilat-eral middle cerebral artery and bilateral common ca-rotid artery occlusion (MCAO/CCAO). Most of the 3-hischemic period was maintained without anesthesiaand reversed by microclip removal of the contralat-eral common carotid artery occlusion. In one subset ofrats transcardial perfusion with India ink was used toestimate the degree of ischemia produced duringMCAO/CCAO in the SON, lateral magnocellular nu-cleus of the PVN (PVL), caudoputamen (CP), and fron-toparietal cortex (COR). Computer-assisted densitom-etry measurements of ink density indicated significantreductions in ink penetration in the territory of theoccluded MCA within the SON (46%), PVL (45%), CP(53%), and COR (76%). In contrast, neither sham-oper-ated rats nor rats subjected to occlusion of the MCAalone showed differences in ink optical densities be-tween the sides ipsilateral and contralateral to MCAO.The other subset of rats were perfused 48–72 h afterrecovery and brain sections were examined for neuro-degenerative changes. While the incidences of cere-bral and caudoputamen infarction after MCAO/CCAOwere 98.4 and 52%, respectively, the histological fea-tures of the SON or PVL in ischemic rats were similarto those of control rats. Reduced susceptibility of mag-nocellular neuroendocrine cells to ischemia may bedue to a number of mechanisms including neuronalresilience, neuroprotection by glia and vascular/perivascular cells, and access to perivascular cerebro-spinal fluid. © 2002 Elsevier Science (USA)
Key Words: supraoptic nucleus; paraventricular nu-cleus; survival; osmoregulation; vasopressin; excito-toxicity; stroke; neuroprotection.
© 2002 Elsevier Science (USA)All rights reserved.
INTRODUCTION
Severe ischemia produced by occluding an intracra-nial artery results in irreversible neurologic injury.Permanent occlusion of the middle cerebral artery last-ing 24 h or longer, for example, leads to widespreadneuronal necrosis in the cerebral cortex (1). Previousstudies have used varied methods to study the effectsof ischemia on neurologic deficits and brain histopa-thology in the cortex and striatum (1–6). Subcorticalregions such as the thalamus and hypothalamus havereceived relatively less attention in this regard (7–9).In particular, the magnocellular neuroendocrine nucleihave been relatively understudied, in spite of the factthat they are supplied by the middle cerebral artery(10, 11). The neurosecretory cells of the paraventricu-lar nucleus (PVN) and the supraoptic nucleus (SON) ofthe hypothalamus, which produce the hormones oxyto-cin and vasopressin, are involved in body fluid and saltbalance as well as cardiovascular function (12–18). Un-like neurons in the cortex or other hypothalamic re-gions, these magnocellular neurosecretory cells are re-sistant to intracranial or systemic injections of gluta-mate analogues (19–25), indicating that they may beselectively spared during stroke as well.
The objectives of this initial study were (1) to deter-mine if occlusion of the middle cerebral artery (MCA)could result in reduced perfusion in the PVN and SONand (2) to investigate whether experimental ischemiahad adverse effects on these nuclei after a 48- to 72-hsurvival period, a period sufficient to produce neuronalnecrosis (8, 26). The former goal was achieved by quan-tifying ink penetration in local blood vessels duringarterial occlusion. To investigate the delayed effects ofischemia, a second group of rats were allowed 48–72 hof recovery after MCA and bilateral common carotidartery occlusion (MCAO/CCAO) and sections from dif-ferent brain areas were examined for histopathologicchanges using cresyl violet and hematoxylin/eosin
Received December 21, 2
; accepted July 31, 2002staining. Comparisons were made with the cerebral
001
2680014-4886/02 $35.00

cortex, where significant progress has been made in thecharacterization of ischemia-induced infarction. Toproduce ischemia we used MCAO in tandem withCCAO (6). This model prevented the need for retrac-tion of the brain or mechanical ventilation of the rats.Further, the ischemic period of 3 h was carried out, forthe most part, while animals were unanesthetized. Ourfindings suggest that ischemia can be produced in theSON and PVN using this model. After a 3-day period ofsurvival, magnocellular neuroendocrine cells (MNCs)in these nuclei appeared normal, while other territo-ries of the MCA manifested signs of neurodegenera-tion. These findings have been published previously inpreliminary form (27).
MATERIALS AND METHODS
Animals
All animal use procedures were conducted in strictaccordance to the guidelines endorsed by the NationalInstitutes of Health Guide for the Care and Use ofLaboratory Animals (1996) and the University of Cal-ifornia at Riverside Chancellor’s Committee on Labo-ratory Animal Care. Male Fisher rats (210–310 g) werehoused individually in a vivarium with a 12:12 h light–dark cycle and were allowed ad libitum access to Pu-rina rat chow and tap water except on the day beforesurgery, when they were given only water to drink.Rats were divided into four groups; a unilateral MCAOonly group, MCAO combined with bilateral CCAO,sham-operated, and intact controls. Before surgeryeach animal was anesthetized with one ip injection ofNembutal (50 mg/kg).
Ischemic Injury
A three-vessel model including MCA occlusion wasadopted as described (6) in order to investigate thepossibility of inducing hypothalamic ischemia. Briefly,anesthetized rats were placed in a steriotaxic framemodified with a head holder which could be rotated. Askin incision was made on the right side of the headand the underlying temporalis muscle was cut andreflected. Using a stereomicroscope, the right MCA wasvisualized through a small craniotomy using an elec-tric microdrill under continuous saline irrigation. TheMCA was visualized with reference to the inferior ce-rebral vein, at a site proximal to the MCA bifurcation,but distal to the origin of the lenticulostriate arteries(28). The MCA was exposed and permanently ligatedwith 10-0 surgical suture. Compromised blood flow inthe occluded MCA was verified visually. Subsequently,a midline incision was made in the neck to isolate thecommon carotid arteries (CCA). The ipsilateral CCAwas permanently occluded with 3-0 suture while thecontralateral CCA was temporarily occluded with a
surgical microclip. Wounds were closed with surgicalwound clips and the rats returned to their individualcages. This model offered less invasive surgery and lessanimal discomfort than proximal occlusion models. Byoccluding the carotids in tandem with MCAO, Buchanand Xue (1992) have shown marked cerebral blood flowreduction and larger predictable infarcts compared toother distal occlusion models (5).
A subset of rats (n � 30) were assigned to an indiaink group (see below) and perfused during the ischemicperiod to estimate the extent of ischemia in the SON,PVN, caudoputamen (CP), and frontoparietal cortex(COR). In another subset of rats assigned to the sur-vival group (n � 63), surgery and occlusions wereperformed under Nembutal anesthesia (50 mg/kg).Rats began to emerge from the anesthesia about a halfhour after the last of the three blood vessels was oc-cluded. After an additional 3 h of ischemia the con-tralateral CCA clamp was removed without anesthesiaand rats were allowed to survive for an additional48–72 h. Core temperature was monitored and main-tained near 37°C during the ischemic period and arecovery period of 3 h more using a heating pad and/orlamp. Sham-operated control rats (n � 24) weretreated in an identical manner to the experimental ratsexcept that the actual occlusions were not performed.Intact controls (n � 10) did not receive anesthesia orsurgery. Neurological tests were performed at the endof the survival period. Subsequently, rats were deeplyanesthetized with Nembutal and transcardially per-fused with 100 ml of cold heparinized (2.5 Units/ml)phosphate-buffered saline (PBS) followed by 4, 6, or 8%cold PFA. Brains were removed and postfixed for 1–3days.
Of one hundred and sixty-two rats, 14 rats diedduring the survival period, resulting in a mortality rateof 8.6%. Those displaying uncontrolled bleeding duringsurgery, respiratory distress, or epileptic seizures wereeuthanized (n � 18) and their measurements were notincluded in the data reported in this study. In addition,five brains were damaged during processing.
India Ink Perfusions
During the early phase of the ischemic period, whilestill under deep anesthesia, rats were transcardiallyperfused with 100 ml of cold PBS followed by 200 ml offiltered cold PBS containing 20% black India ink (De-sign Higgins, Sanford). A flow rate of 10.8 ml/min wasdetermined to provide the best ink retention in smallblood vessels such as capillaries without imposingdamage. Gross inspection of brains indicated that con-trol rats had well perfused brains, displaying dark grayto black staining in major and minor blood vessels aswell as the brain parenchyma. In contrast, MCAO/CCAO rats showed obvious discontinuity in ink distri-
269REDUCED SUSCEPTIBILITY TO ISCHEMIA IN NEUROENDOCRINE CELLS

bution in the territory of the occluded MCA, in bothcortical and subcortical regions.
Brains were removed and postfixed in 4–8% para-formaldehyde (PFA) for an additional 1–3 days andthen blocked. Coronal sections (70 �m) were obtainedusing a vibratome and then mounted, air dried, andcounterstained with cresyl violet to determine the ex-act locations of neurons in question. Sections wereexamined at 200� using a conventional light micro-scope (Nikon Axiophot) and analyzed for ink penetra-tion. Sections from control brains showed widespreadink penetration, whereas sections from ischemic brainshad relatively less to no ink penetration in the territorysupplied by the occluded MCA (Fig. 1). Microscopeimages of the middle aspect of the SON, the PVL, andthe parietal or frontal cortex were acquired with aSony 3CCD color video camera and digitized. Usingcomputer-assisted densitometry (Image-Pro Plus;Media Cybernetics), images were segmented based onan RGB color model in order to isolate black objects.Integrated optical densities (IODs) for ink-containingobjects such as small blood vessels and capillaries werecalculated and summed for each image. Summed IODsfrom each of three images obtained from each side (twosides) of each brain region (four regions) were analyzedfor each rat. IODs for all rats in each group were pooledto obtain mean IODs � SEM.
Histopathological Analysis
After 48–72 h postischemia, rats were transcardiallyperfused and brains were blocked rostrally through thefrontal lobe of the cerebrum, 2 mm in front of the opticchiasm, and posteriorly through the temporal lobe, 2mm behind the superficial boundary of the optic tracts.Sections were cut at 70 �m, stained with 0.001% cresylviolet acetate, coverslipped, and examined under lightmicroscopy at 200 and 400�. Most sections were coun-terstained with cresyl violet to identify infarcted areasor to examine the distribution of India ink perfusionwithin selected brain regions. Viable cells had violet-blue somata with normal proportions (such as those incontrol areas). Histological features used to identifythe lesion included neuronal shrinkage, necroticperikarya, absence of neurons, or alterations in thetissue stainability.
In order to resolve more discrete changes in cellularmorphology tissue sections were processed for hema-toxylin and eosin staining. Dehydration and infiltra-tion with polyethylene glycol was followed by section-ing at 10–14 �m using a microtome and mounting asdescribed by Smithson and colleagues (29). Sectionswere air-dried and stained with Gill’s No. 1 hematoxi-lin and eosin Y solution. Histological features used toidentify the lesion at high magnification included neu-ronal shrinkage with scalloped appearance, reducedstainability of neurons, and vacuolation (sponginess) of
the neuropil or diffuse pallor of the eosinophilic back-ground due to a reduced presence of viable cells asdescribed by Garcia and others (8). At 400� magnifi-cation infarction was counted if the necrotic area en-compassed at least 10% of the field (0.31 mm2). Anincidence of infarction for selected cortical and subcor-tical regions was calculated as the proportion of thetotal number of rats that contained an infarcted sam-ple. Control samples were derived from the contralat-eral hemisphere, the ipsilateral areas not supplied bythe MCA, and sham-operated (n � 24) and intactanimals (n � 10).
Statistical Analysis
India ink penetration. Statistical evaluation of thedifferences in ink penetration between contralateraland ipsilateral sides (MCAO side) and among selectedbrain regions was performed using a two-way analysisof variance (ANOVA) with interaction. Posthoc analy-sis was performed using Tukey’s test for multiple com-parisons with a 5% experiment-wide error rate. Weconsidered statistical test significance if the corre-sponding P value was �0.05.
RESULTS
Ink Perfusion
In order to begin to address the effect of MCAO/CCAO on the magnocellular neuroendocrine nuclei ofthe hypothalamus we measured the ability of transcar-dially perfused India ink to penetrate these regionsimmediately after artery occlusion. For the MCAO/CCAO model (n � 30) ANOVA indicated a significanteffect of sides (df � 29; F � 317.49, P � 0.0001) andregions (df � 29; F � 142.51, P � 0.0001) and in theinteraction between sides and regions (df � 29; F �11.83, P � 0.001). This prompted the subsequentanalysis using Tukey’s test for multiple comparisons.In order to quantitate any potential differences be-tween regions we statistically analyzed the effect ofMCAO/CCAO on regional mean ink IODs for SON,PVL, CP, and COR. For each brain region examined,Table 1 lists the mean ink IODs (�106) � SEM forcontralateral (unoccluded) and ipsilateral (occluded)sides combined. Tukey’s test for multiple comparisonsyielded the following regional rank order of means (�SEM) from high to low staining density: PVL (1.898 �0.928) � SON (1.300 � 0.669) � COR (0.903 � 0.695) �CP (0.577 � 0.344), suggesting a high degree of vascu-larization in the magnocellular neuroendocrine nuclei.When contralateral IODs were analyzed alone an iden-tical rank order was determined for these four brainregions.
More importantly, our analysis confirmed the effec-tiveness of the MCAO/CCAO model in producing uni-
270 CURRAS-COLLAZO, PATEL, AND HUSSEIN

FIG. 1. Extent of India ink distribution after MCAO/CCAO. Rats were perfused with 20% India ink immediately after three-vesselocclusion. Sections taken through the territory of the occluded MCA showed minimal ink penetration in the frontoparietal cortex (B) andreduced perfusion in the caudoputamen (D), SON (F), and lateral magnocellular nucleus of the PVL (H). The side contralateral to occlusiondisplayed significantly greater ink distribution (A, C, E, and G), as confirmed by computer-assisted densitometry. Calibration bar, 50 �m.
271REDUCED SUSCEPTIBILITY TO ISCHEMIA IN NEUROENDOCRINE CELLS

lateral ischemia. The major outcome of Tukey’s analy-sis was that contralateral mean ink IODs were greaterthan ipsilateral values in each brain region studied.Table 2 lists the average ink penetration for ipsilateraland contralateral sides in SON, PVL, COR, and CPmeasured using the mean ink IODs acquired from 30rats. For SON, the mean IODs � SEM for contralateraland ipsilateral sides were 1.687 � 0.704 and 0.913 �0.452, respectively. For PVL, these values were2.452 � 0.796 and 1.344 � 0.700, respectively. Corre-sponding values for COR were 1.452 � 0.612 for con-tralateral and 0.355 � 0.266 for ipsilateral sides. MeanIOD values for CP were lower overall but still differentbetween contralateral and ipsilateral sides, respec-
tively (0.784 � 0.355 and 0.369 � 0.217). An estimateof the extent of ischemia in each brain region wascalculated as the difference between the mean IODs forcontralateral and ipsilateral sides and expressed asfractional change in ink perfusion (Table 2). Thesevalues were 0.756 (COR), 0.529 (CP), 0.459 (SON), and0.452 (PVL), where 1.0 represented complete ischemia.These data suggest a significant effect of MCAO/CCAOin decreasing blood perfusion to these brain regions.Fractional changes in ink perfusion ranged from 0.02to 0.98 for SON, from 0.02 to 0.99 for PVL, from 0.03 to0.99 for CP, and from 0.06 to 0.99 for COR, indicatingthat in a few cases almost complete ischemia was at-tained in the SON and PVN.
Hypothalamic ischemia is evident in Fig. 1, whichshows representative images from sections through theSON, PVL, as well as CP and COR of MCAO/CCAOrats perfused with black India ink during the three-artery occlusion period. Extensive vascular branchingcontaining the ink can be seen on the side contralateralto the MCA occlusion (left panels). In the SON andPVL these branches course intimately between thelarge magnocellular neuroendocrine cells (�20 �m indehydrated tissue). On the ipsilateral side, however,MCAO/CCAO reduced ink penetration most notably inCOR (B) but also in CP (D), SON (F), and PVL (H).
We also examined whether three-blood-vessel occlu-sion could decrease ink penetration in the parvocellu-lar PVN using a subset of the 30 total rats for whichboth parvocellular and magnocellular PVN regionswere present. Statistical analysis of mean IODs ob-tained from ipsilateral and contralateral sides in theparvocellular and magnocellular regions of 21 ratsyielded a statistically significant interaction betweenregions and sides (F � 87.01; P � 0.0001). However,ad hoc analysis using Tukey’s test yielded a significantdifference only in the magnocellular region (T �11.98; P � 0.0001). Mean ink IODs were no different
TABLE 1
Total India Ink Penetration by Regionin MCAO/CCAO Rats
COR CP SON PVL
Mean 0.903 0.577 1.300* 1.898*SEM 0.695 0.344 0.669 0.928Rats 30 30 30 30Sections 180 180 180 180
Note. Digital images of ink penetration during three-blood-vesselocclusion in selected brain regions were analyzed using computer-assisted densitometry. IOD values (�106) are expressed as means ofthe total number of samples analyzed. Mean ink IODs for SON andPVL were greater than those for COR and CP, confirming a greaterdegree of vascularization in the magnocellular neuroendocrine nu-clei. Mean values for the different brain regions represent the pooleddata from both contralateral and ipsilateral sides. The number ofrats per group (N � 30) and the number of sections sampled pergroup are indicated (3 sections � 2 sides � 30 rats). Standard errorsreported are based on the observations used to calculate the means.The standard errors on common variance would be 0.058 and 0.048(�106) for region and side means, respectively.
* P � 0.05, difference in mean IODs for cortex (COR), caudopu-tamen (CP), supraoptic nucleus (SON), and lateral magnocellularnucleus of PVN (PVL) pooled from both sides.
TABLE 2
Degree of Ischemia in Selected Brain Regions Produced by MCAO/CCAO
COR CP SON PVL
Contralateral 1.452 � 0.612 0.784 � 0.335 1.687 � 0.704 2.452 � 0.796Ipsilateral 0.355 � 0.266* 0.369 � 0.217 0.913 � 0.452* 1.344 � 0.700*Difference in sides 1.097 0.415 0.774 1.108Degree of ischemia 0.756 0.529 0.459 0.452Rats 30 30 30 30Sections (each side) 90 90 90 90
Note. Digital images of ink penetration on ipsilateral (occluded) and contralateral (unoccluded) sides were analyzed using computer-assisted densitometry. IOD values (�106) are expressed as means � SEM. The number of rats per group (N � 30) and the number ofsections/side for each region sampled are indicated (3 sections � 30 rats). The degree of ischemia was calculated as the fractional change inink penetration (IODcontra � IODipsi/IODcontra); a value of 0 represents normal brain perfusion and 1.0 represents complete ischemia. For eachregion analyzed the mean ink IOD for contralateral was greater than that on the ipsilateral side. Standard errors reported are based on theobservations used to calculate the means. The standard error on common variance would be 0.0824 (�106) for interaction means.
* P � 0.05, difference in IODs between contralateral and ipsilateral sides for individual regions.
272 CURRAS-COLLAZO, PATEL, AND HUSSEIN
between ipsilateral and contralateral sides in the par-vocellular PVN (T � 1.081; P � 0.7013). Therefore,the three-blood-vessel occlusion model of ischemiaused in our study could not provide a sufficient block ofink penetration in the parvocellular PVN. Data on theparvocellular PVN, then, are inconclusive regardingthe susceptibility of these cells to ischemia.
For the MCAO only group (n � 5) F and P values forthe interaction mean for ink IOD were within normallimits (df � 4; F � 0.36, P � 0.698) even though theeffect of region showed a statistically significant differ-ence, as expected since cortex is almost exclusivelysupplied by the MCA (df � 4; F � 9.73; P � 0.0001).Tukey’s posthoc analysis yielded no difference betweenipsilateral and contralateral sides in either SON (T ��2.62; P � 0.1036) or PVL (T � �2.283; P �0.2131) in the MCAO only group. A similar analysis ofthe control group (n � 19) showed no statistical sig-nificance in the effect of sides (df � 18; F � 1.81, P �0.180), regions (df � 18; F � 1.37, P � 0.255), or theinteraction between sides and regions (df � 18; F �0.56, P � 0.571). The control group was composed ofsham-operated (n � 12) and intact control rats (n � 7)since no differences in each of these sets of rats werefound between sides or regions nor in the interactionbetween sides and regions.
Based on these results, the remaining rats were sub-jected to MCAO/CCAO in order to achieve the mostintense ischemia possible in the SON and PVL. Usingthe MCAO/CCAO model our mortality rate was 8.6%.This percentage is higher than that reported by Garciaand colleagues (8) but much lower that that reportedby Nagasawa and Kogure (4) using single-artery occlu-sion.
Histological Evaluation
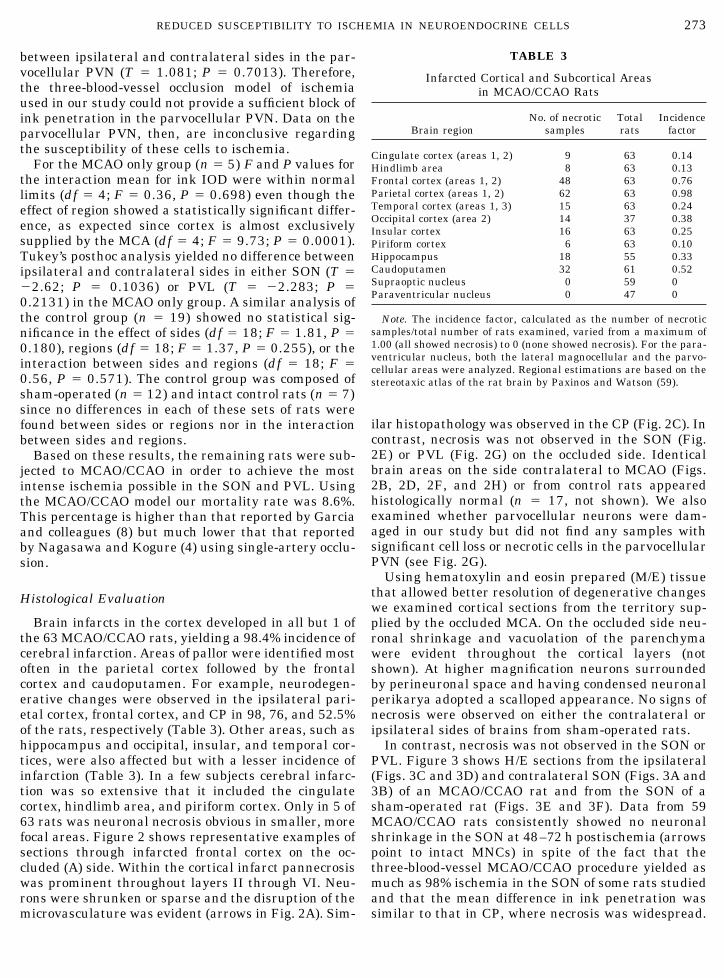
Brain infarcts in the cortex developed in all but 1 ofthe 63 MCAO/CCAO rats, yielding a 98.4% incidence ofcerebral infarction. Areas of pallor were identified mostoften in the parietal cortex followed by the frontalcortex and caudoputamen. For example, neurodegen-erative changes were observed in the ipsilateral pari-etal cortex, frontal cortex, and CP in 98, 76, and 52.5%of the rats, respectively (Table 3). Other areas, such ashippocampus and occipital, insular, and temporal cor-tices, were also affected but with a lesser incidence ofinfarction (Table 3). In a few subjects cerebral infarc-tion was so extensive that it included the cingulatecortex, hindlimb area, and piriform cortex. Only in 5 of63 rats was neuronal necrosis obvious in smaller, morefocal areas. Figure 2 shows representative examples ofsections through infarcted frontal cortex on the oc-cluded (A) side. Within the cortical infarct pannecrosiswas prominent throughout layers II through VI. Neu-rons were shrunken or sparse and the disruption of themicrovasculature was evident (arrows in Fig. 2A). Sim-
ilar histopathology was observed in the CP (Fig. 2C). Incontrast, necrosis was not observed in the SON (Fig.2E) or PVL (Fig. 2G) on the occluded side. Identicalbrain areas on the side contralateral to MCAO (Figs.2B, 2D, 2F, and 2H) or from control rats appearedhistologically normal (n � 17, not shown). We alsoexamined whether parvocellular neurons were dam-aged in our study but did not find any samples withsignificant cell loss or necrotic cells in the parvocellularPVN (see Fig. 2G).
Using hematoxylin and eosin prepared (M/E) tissuethat allowed better resolution of degenerative changeswe examined cortical sections from the territory sup-plied by the occluded MCA. On the occluded side neu-ronal shrinkage and vacuolation of the parenchymawere evident throughout the cortical layers (notshown). At higher magnification neurons surroundedby perineuronal space and having condensed neuronalperikarya adopted a scalloped appearance. No signs ofnecrosis were observed on either the contralateral oripsilateral sides of brains from sham-operated rats.
In contrast, necrosis was not observed in the SON orPVL. Figure 3 shows H/E sections from the ipsilateral(Figs. 3C and 3D) and contralateral SON (Figs. 3A and3B) of an MCAO/CCAO rat and from the SON of asham-operated rat (Figs. 3E and 3F). Data from 59MCAO/CCAO rats consistently showed no neuronalshrinkage in the SON at 48–72 h postischemia (arrowspoint to intact MNCs) in spite of the fact that thethree-blood-vessel MCAO/CCAO procedure yielded asmuch as 98% ischemia in the SON of some rats studiedand that the mean difference in ink penetration wassimilar to that in CP, where necrosis was widespread.
TABLE 3
Infarcted Cortical and Subcortical Areasin MCAO/CCAO Rats
Brain regionNo. of necrotic
samplesTotalrats
Incidencefactor
Cingulate cortex (areas 1, 2) 9 63 0.14Hindlimb area 8 63 0.13Frontal cortex (areas 1, 2) 48 63 0.76Parietal cortex (areas 1, 2) 62 63 0.98Temporal cortex (areas 1, 3) 15 63 0.24Occipital cortex (area 2) 14 37 0.38Insular cortex 16 63 0.25Piriform cortex 6 63 0.10Hippocampus 18 55 0.33Caudoputamen 32 61 0.52Supraoptic nucleus 0 59 0Paraventricular nucleus 0 47 0
Note. The incidence factor, calculated as the number of necroticsamples/total number of rats examined, varied from a maximum of1.00 (all showed necrosis) to 0 (none showed necrosis). For the para-ventricular nucleus, both the lateral magnocellular and the parvo-cellular areas were analyzed. Regional estimations are based on thestereotaxic atlas of the rat brain by Paxinos and Watson (59).
273REDUCED SUSCEPTIBILITY TO ISCHEMIA IN NEUROENDOCRINE CELLS

FIG. 2. Necrotic changes after MCAO/CCAO were apparent in the frontoparietal cortex and caudoputamen but not in the SON or PVL.A subset of rats were allowed to survive for 48–72 h after MCAO/CCAO for 3 h followed by reperfusion. Photomicrographs of sections throughthe frontoparietal cortex (A and B) showed a diffuse pallor, neuronal shrinkage, and a disruption of the microvasculature only in thehemisphere ipsilateral to the occluded MCA (A). In sections of caudoputamen (C and D) a relative loss of neurons is evident on the occludedside (C; arrows). In comparison, caudoputamen neurons on the contralateral side appear normal (arrowheads). Magnocellular neuroendo-crine cells on both sides of the SON (E and F) and PVL (G and H) appear normal (arrowheads). Calibration bar, 50 �m.
274 CURRAS-COLLAZO, PATEL, AND HUSSEIN

No vacuolation was observed in the SON and similarresults were obtained in the PVL (n � 47; Fig. 4).Figure 4 shows that the majority of neurons in the PVLwere intact on the MCA-occluded side (Figs. 4C and4D; arrows). Parvocellular neurons do not show anysigns of necrosis.
DISCUSSION
In this study we have examined the effectiveness ofa three-vessel occlusion model in producing unilateral
ischemia and neurodegeneration in the magnocellularneuroendocrine nuclei of the hypothalamus. Statisticalanalysis confirmed that the differences in ink penetra-tion between sides as measured by India ink perfusionwere significant in the SON and PVL, respectively,yielding estimated degrees of ischemia of 45.9 and45.2%. A significant reduction of ink penetration wasmeasured in the CP (52.9%), while that in the COR wasgreater (75.6%). India ink has been used as an intra-vascular tracer in previous studies to study the micro-vascular anatomy of striate vessels in dogs (30). Other
FIG. 3. Absence of neurodegenerative changes in the SON after MCAO/CCAO. Rats subjected to MCAO/CCAO and reperfusion 3 h laterwere sacrificed after 48–72 h of survival. Ten- to 14-�m sections were cut from MCAO/CCAO rat brains and counterstained with hematoxylinand eosin. Neurons in the SON ipsilateral to the MCAO (C) had a normal morphology, as did those on the contralateral SON (A) and thosein the SON from a sham-operated rat (E). At higher magnification (400�) no signs of necrosis were observed on the ipsilateral SON (D).Arrows point to healthy magnocellular neuroendocrine cells. Blood vessels can be seen coursing between the neurosecretory cells (asterisksin D and F). Calibration bar, 50 �m.
275REDUCED SUSCEPTIBILITY TO ISCHEMIA IN NEUROENDOCRINE CELLS

investigators have perfused four-vessel occluded ratswith Evan’s blue dye but quantitation of ischemia wasnot carried out (31).
The lower degree of ischemia in the SON, PVL, andCP as compared to the COR of MCAO/CCAO rats maybe due to the fact that these nuclei receive blood supplyfrom the anterior cerebral artery as well as the MCA(10, 11, 32). In addition, the MCA territory in rats ispartially supplied by interarterial anastomoses be-tween the MCA and the ACA (33) as well as collaterals
(34). Assuming that blood flow in the CCAs is alreadycompromised during MCAO/CCAO, it is unlikely thatthe collateral blood supply in the rat contributes to thelower degree of ischemia in the SON and PVL.
We speculate that the extent of ischemia observed inthe SON and PVL is sufficient to produce irreversibleinjury. A comparison of our ink penetration with cere-bral blood flow (CBF) values obtained in previous stud-ies can be attempted. Using the same MCAO/CCAOmodel Buchan and colleagues (6) reported CBF reduc-
FIG. 4. Lack of histopathological changes in the PVL of MCAO/CCAO rats. Rats subjected to MCAO/CCAO and reperfusion 3 h later weresacrificed after 48–72 h of survival. Ten- to 14-�m sections were cut from rat brains and counterstained with hematoxylin and eosin.Photomicrographs of PVL sections through the occluded (C) and contralateral (A) sides as well as from a control rat (E) showed a similarpattern of staining. At higher magnification neither shrunken magnocellular neurons nor vacuolation were obvious. Instead, MNCs on theside ipsilateral to MCAO had normal proportions (arrows in D). The faded appearance of a few neurons on the ipsilateral side was alsodetected in some sections of the contralateral PVL and control PVL (arrowheads). Blood vessels could be seen coursing between theneurosecretory cells (asterisks in B, D, and F). Calibration bar, 50 �m except in E (43 �m).
276 CURRAS-COLLAZO, PATEL, AND HUSSEIN

tions in the occluded cortex to about 8% of baselinelevels. Based on control data from MCAO alone orMCAO in tandem with unilateral CCAO, absolute CBFvalues may be approximately 2.2–10 ml/min/100 g dur-ing MCAO/CCAO (35, 36). Assuming similar controlCBF values and equal penetration of ink particles inthe SON, PVL, CP, and COR, the reduced CBF on theoccluded side should extrapolate to 4.5–20.8 ml/min/100 g in our model. These estimates are below the 25ml/min/100 g threshold required for induction of isch-emia (37, 38). CBF at this level can result in infarctionif maintained for 3 to 4 h (6, 37). Further studies areneeded to measure CBF directly in these deep regionsof the brain.
Our histological data suggest that neither the SONnor the PVL showed signs of necrosis after 48–72 h ofsurvival. No histopathological changes were observedin the ipsilateral SON and PVL of any rat. The absenceof neuronal shrinkage and the presence of normalstainability in MNCs suggests a reduced susceptibilityof magnocellular neuroendocrine cells of the hypothal-amus to ischemia. In contrast, a significant incidence ofinfarction (52%) was observed in the caudoputamen.The frontoparietal cortex was the most affected, withall but 1 of 63 rats displaying cerebral infarction. Mostbrains showed necrotic changes that extended into sev-eral cerebral lobes even though these varied from rat torat.
Our current findings showing reduced susceptibilityof the SON and PVL to ischemia-induced neurodegen-eration support the results of previous studies showingresistance of MNCs to glutamate analogues. This isrelevant since in vivo microdialysis studies of cortexhave demonstrated that extracellular levels of gluta-mate and glycine are elevated in rat and gerbil focaland global models of ischemia (39, 40). In addition,chronic exposure of the brain to glutamate results inneurodegenerative changes similar to those producedby ischemia in vivo (7, 25) or hypoxic-induced excito-toxicity in vitro (41, 42). The critical event mediatingglutamate-induced cell death is dependent on Ca2� ions(42), which accumulate intracellularly as a result of theactivation of N-methyl-D-aspartate-type glutamate re-ceptors (NMDA). Indeed, NMDA receptor antagonistsgiven prior to or a few hours after an ischemic insultcan protect against ensuing necrotic changes (43–45).Intracranial injections of glutamate receptor agonistssuch as ibotenic acid (20, 23, 24), kainic acid (21, 22),quisqualic acid (24), and NMDLA (21, 24) do not pro-duce cell death in MNCs of the SON and PVN. Arelative insensitivity to EAAs has been verified in ourlaboratory using an in vitro model of excitotoxicity.Specifically, SON cultures are much less susceptible tochronic NMDA application (46) and to saturating dosesof exogenous glutamate compared to cortical cultures;the latter may be due, in part, to enhanced glutamateuptake, calcium tolerance in the SON, but not to a lack
of NMDA receptors (47–50). In combination, thesefindings suggest the possibility that MNCs are resis-tant to ischemia for the same reasons they are resis-tant to glutamate receptor agonists.
We hypothesize that more prolonged ischemia wouldnot be expected to induce neurodegeneration in theSON and PVL. Using the same three-vessel occlusionmethod Buchan and colleagues (1992) demonstratedthat damage incurred by 3 h of ischemia followed by 20or 21 h of reperfusion was indistinguishable from thatseen after 24 h of permanent ischemia. Furthermore,the lack of necrosis in PVN and SON MNCs is notlikely to be due to insufficient duration of the postisch-emic period. The time lapse, 48–72 h after MCAocclusion, is sufficient to detect irreversible neuronalnecrosis in the cerebral cortex, with no significantincreases in necrotic neurons seen after 12 h postisch-emia (8). It is unlikely that SON and PVN neurons diemore slowly after an ischemic insult than cortical neu-rons, although we cannot rule out this possibility.
The focal ischemia model used in the present studywas chosen in order to augment the likelihood of in-farction and to reduce the time under anesthesia re-ported using single-artery occlusion (5, 51, 52). Sincedistal occlusion of the MCA alone results in less fre-quent and extensive necrosis compared with that inanimals with proximal occlusions (51, 53), the combi-nation of common carotid artery occlusion with MCAOhas been recommended by previous studies to improvereproducibility in cerebral infarction (6, 54, 55). Ac-cordingly, our results indicate a 98.4% incidence ofcerebral infarction with MCAO/CCAO. Also, using athree-way artery occlusion, Rubino and Young (56)have shown cortical infarcts in 30–40% of the ischemicrats. In their study (as in ours) branches of the MCAdistal to the crossing of the inferior cerebral vein wereoccluded. Our much higher incidence of infarction maybe attributed to the longer period of ischemia (3 hcompared to 30 min).
Using MCAO/CCAO Buchan and colleagues (6) havereported a similar moderate incidence of striatal in-farction. Rats subjected to transient unilateral occlu-sion of the proximal MCA using the intraluminalmethod show subcortical damage in CP. Memezawaand others (57) reported a high incidence of infarctionin both CP (87.5%) and neocortex (100%) with 1 h ofischemia. In contrast, unilateral occlusion of the MCAbeginning 1 mm distal to the inferior cerebral veinyields no infarction under these conditions (5, 53).Therefore, the intermediate incidence of striatal infarc-tion measured here (52%) is probably not due to block-ade of blood supply via lenticulostriate and TOLbranches of the MCA (51) but may be due to reductionof blood flow in branches supplying the striatum re-sulting from MCA and CCA occlusion.
It is not surprising that the degree of ischemia wasdifferent in the parvocellular and magnocellular re-
277REDUCED SUSCEPTIBILITY TO ISCHEMIA IN NEUROENDOCRINE CELLS

gions of the PVN since they are not supplied by iden-tical arterial components. For example, the lower (par-vocellular) region is supplied by the lateral chiasmaticartery, a branch of the retrochiasmatic artery, whilethe upper (magnocellular) region gets supplied by boththe lateral hypothalamic artery and the medial sub-commissural artery. While both lateral chiasmatic andlateral hypothalamic arteries usually arrive from theinternal carotid via branching from the retrochias-matic artery, the medial commissural artery rises fromthe anterior cerebral artery. Complexity is added bythe fact that the lateral hypothalamic artery some-times rises directly from the internal carotid artery(58). Moreover, the density of capillaries is much lowerin the parvocellular than in the magnocellular regionsof the PVN (11), possibly affecting resistance and thusblood flow in both areas.
CONCLUSIONS
Our study has shown for the first time that MNCs ofthe SON and PVN are remarkably resistant to isch-emia-induced neurodegeneration relative to frontopa-rietal cortex and caudoputamen neurons. In futurework it will become important to assess the functionalintegrity of these neurosecretory cells after an ischemicinsult. The full set of mechanisms contributing to theselective sparing of MNCs remains to be elucidated.
ACKNOWLEDGMENTS
We express our sincere thanks to Dr. Dong Xue for his help indemonstrating the surgical procedure. In addition, we are grateful toC. M. Collazo for help with graphics and figures, to S. Desai, K.Flotildes, H. Mussenden, P. Peiris, M. Pugh, E. Snissarenko, G.Soliman, and E. Tzeng for their technical assistance, and to B.Cowden (Media Cybernetics) and R. Logeman (A.G. Heinz) for soft-ware support. A special thanks is extended to Dr. R. Beaver (UCR)for advice on the statistical analysis. This work was funded by anAmerican Heart Association Grant-in-Aid to M.C.C.-C.
REFERENCES
1. Garcia, J. H., K. F. Liu, and K. L. Ho. 1995. Neuronal necrosisafter middle cerebral artery occlusion in Wistar rats progressesat different time intervals in the CP and the cortex. Stroke26(4): 636–642.
2. Tamura, A., D. I. Graham, J. McCulloch, and G. M. Teasdale.1981. Focal cerebral ischemia in the rat. I: Description of tech-nique and early neuropathological consequences following mid-dle cerebral artery occlusion. J. Cereb. Blood Flow Metab. 1:53–60.
3. Zea Longa, E., P. R. Weinstein, S. Carlson, and R. Cummins.1989. Reversible middle cerebral artery occlusion without cra-niotomy in rats. Stroke 20: 84–91.
4. Nagasawa, H., and K. Kogure. 1989. Correlation between cere-bral blood flow and histologic changes in a new rat model ofmiddle cerebral artery occlusion. Stroke 20: 1037–1043.
5. Bederson, J. B., L. H. Pitts, M. Tsuji, M. C. Nishimura, R. L.Davis, and H. Bartkowski. 1986. Middle cerebral artery occlu-
sion: evaluation of the model and development of a neurologicexamination. Stroke 17(3): 472–476.
6. Buchan, A. M., D. Xue, and A. Slivka. 1992b. A new model oftemporary focal neocortical ischemia in the rat. Stroke 23: 273–279.
7. Ikonomidou, C., M. T. Price, J. L. Mosinger, G. Frierdich, J. La-bruyere, K. Shahid Salles, and J. W. Olney. 1989. Hypobaric–ischemic conditions produce glutamate-like cytopathology ininfant rat brain. J. Neurosci. 9(5): 1693–1700.
8. Garcia, J. H., Y. Yoshida, H. Chen, Y. Li, Z. G. Zhang, J. Lian,S. Chen, and M. Chopp. 1993. Progression from ischemic injuryto infarct following middle cerebral artery occlusion in the rat.Am. J. Pathol. 142: 623–635.
9. Zhang, R.-L., M. Chopp, H. Chen, and J. H. Garcia. 1994.Temporal profile of ischemic tissue damage, neutrophil re-sponse, and vascular plugging following permanent and tran-sient (2H) middle cerebral artery occlusion in the rat. J. Neurol.Sci. 125: 3–10.
10. Ambach, G., and M. Palkovits. 1974. Blood supply of the rathypothalamus: I. nucleus supraopticus. Acta Morphol. Acad.Sci. Hung. 22(3–4): 291–310.
11. Ambach, G., and M. Palkovits. 1974. Blood supply of the rathypothalamus: II. Nucleus paraventricularis. Acta Morphol.Acad. Sci. Hung. 22(3–4): 311–320.
12. Robinson, I. C. A. F. 1983. Neurohypophysial peptides in cere-brospinal fluid. In The Neurohypophysis: Structure, Functionand Control (B. A. Cross and G. Leng, Eds.), Vol. 60, pp. 129–145. Elsevier, Amsterdam.
13. Roberts, J. S. 1977. Oxytocin. Eden Press, Montreal.14. Shrier, R. W. 1985. Vasopressin. Raven Press, New York.15. Dyball, R. E. J., and P. S. Poutney. 1973. Discharge patterns of
supraoptic and paraventricular neurones in rats given 2% NaClsolution instead of drinking water. J. Endocrinol. 56: 91–98.
16. Dutton, A., and R. E. J. Dyball. 1979. Phasic firing enhancesvasopressin release from the rat neurohypophysis. J. Physiol. 290:433–440.
17. Sjoquist, M., W. Huang, E. Jacobsson, O. Skott, E. M. Stricker,and A. F. Sved. 1999. Sodium excretion and renin secretionafter continuous versus pulsatile infusion of oxytocin in rats.Endocrinology 140(6): 2814–2818.
18. Brimble, M. J., R. E. J. Dyball, and M. L. Forsling. 1978. Oxytocinrelease following osmotic activation of oxytocin neurones in theparaventricular and supraoptic nuclei. J. Physiol. 278: 69–78.
19. Nadler, J. V., B. W. Perry., and C. W. Cotman. 1978. Intraven-tricular kainic acid preferentially destroys hippocampal cells.Nature 271: 676–677.
20. Hu, B., T. Cunningham, R. Nissen, L. P. Renaud, and C. W.Bourque. 1992. Rat supraoptic neurons are resistant to gluta-mate neurotoxicity. NeuroReport 3: 87–90.
21. Olney, J. W., and T. de Gubareff. 1978. Extreme sensitivity ofolfactory cortical neurons to kainic acid toxicity. Kainic Acid asa Tool in Neurobiology (E. G. McGeer et al., Eds.), pp. 201–217.Raven Press, New York.
22. Peterson, G. M., and R. Y. Moore. 1980. Selective effects ofkainic acid on diencephalic neurons. Brain Res. 202: 165–182.
23. Herman, J. P., and S. J. Wiegand. 1986. Ibotenate-induced celldeath in the hypothalamic paraventricular nucleus: Differen-tial susceptibility of magnocellular and parvicellular neurons.Brain Res. 383: 367–372.
24. Hastings, M. H., P. Winn, and S. B. Dunnett. 1985. Neurotoxicamino acid lesions of the lateral hypothalamus: A parametriccomparison of the effects of ibotenate, N-methyl-D-aspartateand quisqualate in the rat. Brain Res. 360: 248–256.
278 CURRAS-COLLAZO, PATEL, AND HUSSEIN

25. Olney, J. W., and L. G. Sharpe. 1969. Brain lesion in an infantrhesus monkey treated with monosodium glutamate. Science166: 386–388.
26. Garcia, J. H., S. Wagner, K.-F. Liu, and X.-J. Hu. 1995.Neurological deficit and extent of neuronal necrosis attribut-able to middle cerebral artery occlusion in rats. Stroke 26:627–635.
27. Hussein, M. O., and M. C. Curras-Collazo. 1998. Middle cerebralartery occlusion is effective in producing ischemia of the supraop-tic and paraventricular nuclei of the hypothalamus. J. Vasc. Res.35(Suppl. 3): 8–9.
28. Coyle, P. 1975. Arterial patterns of the rat rhinencephalon andrelated structures. Exp. Neurol. 49(3): 671–690.
29. Smithson, K. G., B. A. MacVicar, and G. I. Hatton. 1983. Poly-ethylene glycol embedding: A technique compatible with immu-nocytochemistry, enzyme histochemistry, histofluorescence andintracellular staining. J. Neurosci. Methods 7(1): 27–41.
30. St. Jacques, R., W. Gorczyca, and G. Mohr. 1996. Microvascularanatomy of striate vessels in dogs: Contribution to an experi-mental model of forebrain ischemia. Neurol. Res. 18(2): 157–162.
31. Pulsinelli, W. A., and J. B. Brierly. 1979. A new model of bilateralhemispheric ischemia in the unanesthetized rat. Stroke 10(3):267–272.
32. Rieke, G. K., D. E. Bowers, and P. Penn. 1981. Vascular supplypattern to rat caudatoputamen and globus pallidus: Scanningelectronmicroscopic study of vascular endocasts of stroke-pronevessels. Stroke 12(6): 840–847.
33. Coyle, P., and P. T. Jokelainen. 1982. Dorsal cerebral arterialcollaterals of the rat. Anat. Rec. 203(3): 397–404.
34. Lee, R. M. 1995. Morphology of cerebral arteries. Pharmacol.Ther. 66(1): 149–73.
35. Drummond, J. C., Y.-S. Oh, D. J. Cole, and H. M. Shapiro. 1989.Phenylephrine-induced hypertension reduces ischemia follow-ing middle cerebral artery occlusion in rats. Stroke 20: 1538–1544.
36. Salgado, A. V., S. C. Jones, A. J. Furlan, E. Korfali, S. A. Marshall,and J. R. Little. 1989. Bimodal treatment with nimodipine andlow-molecular-weight dextran for focal cerebral ischemia in therat. Ann. Neurol. 26(5): 621–7.
37. Kaplan, B., S. Brint, J. Tanabe, M. Jacewicz, X.-J. Wang, andW. Pulsinelli. 1991. Temporal thresholds for neocortical infarc-tion in rats subjected to reversible cerebral ischemia. Stroke 22:1032–1039.
38. Tyson, G. W., G. M. Teasdale, D. I. Graham, and J. McCullogh.1984. Focal cerebral ischemia in the rat: Topography of hemo-dynamic and histopathological changes. Ann. Neurol. 15: 559–567.
39. Benveniste, H., J. Drejer, A. Schousboe, and N. H. Diemer.1984. Elevation of the extracellular concentrations of glutamateand aspartate in rat hippocampus during transient cerebralischemia monitored by intracerebral microdialysis. J. Neuro-chem. 43(5): 1369–1374.
40. Globus, M. Y., R. Busto, W. D. Dietrich, E. Martinez, I. Valdes,and M. D. Ginsberg. 1988. Effect of ischemia on the in vivorelease of striatal dopamine, glutamate, and gamma-aminobu-tyric acid studied by intracerebral microdialysis. J. Neurochem.51(5): 1455–1464.
41. Rothman, S. M. 1983. Synaptic activity mediates death of hy-poxic neurons. Science 220: 536–537.
42. Choi, D. W. 1987. Ionic dependence of glutamate neurotoxicityin cortical cell culture. J. Neurosci. 7: 369–379.
43. Palmer, G. C., E. F. Cregan, A. R. Borrelli, and F. Willett. 1995.Neuroprotective properties of the uncompetitive NMDA recep-tor antagonist remacemide hydrochloride. Ann. N. Y. Acad. Sci.765: 236–247.
44. Ikonomidou, C., and L. Turski. 1995. Excitotoxicity and neuro-degenerative diseases. Curr. Opin. Neurol. 8: 487–497.
45. Meldrum, B. S. 1990. Protection against ischemia neuronaldamage by drugs acting on excitatory neurotransmission. Ce-rebrovasc. Brain Metab. Rev. 2: 27–57.
46. Pak, C. W., and M. C. Curras. 1998. Resistance to NMDAexcitotoxicity is shown by supraoptic neurons of the hypothal-amus in vitro. Soc. Neurosci. Abstr. 24(1): 464.
47. Pak, C. W., and M. C. Curras-Collazo. 2000. Resistance ofsupraoptic neurons to glutamate excitotoxicity may be partiallydue to enhanced glutamate clearance. FASEB J. 14(4): A633.
48. Pak, C. W., M. C. Curras-Collazo, and R. B. Meeker. 2001.Mechanism of excitotoxic resistance in supraoptic neurons mayinvolve higher tolerance to calcium load. Soc. Neurosci. Abstr.27: Program 178.4.
49. Curras, M. C., P. Rack, and R. B. Meeker. 1998. Channelproperties of NMDA glutamate receptors on magnocellular neu-roendocrine cells cultured from the rat supraoptic nucleus.Brain Res. 789: 181–193.
50. Hu, B., and C. W. Bourque. 1991. Functional N-methyl-D-as-partate and non-N-methyl-D-aspartate receptors are expressedby rat supraoptic neurosecretory cells in vitro. J.Neuroendocri-nol. 3(5): 509–514.
51. Niiro, M., R. P. Simon, K. Kadota, and T. Asakura. 1996. Prox-imal branching patterns of middle cerebral artery (MCA) inrats and their influence on the infarct size produced by MCAocclusion. J. Neurosci. Methods 64: 19–23.
52. Fox, G., D. Gallacher, S. Shevde, J. Loftus, and G. Swayne.1993. Anatomic variation of the middle cerebral artery in theSprague-Dawley rat. Stroke 24: 2087–2093.
53. Shigeno, T., J. McCulloch, D. I. Graham, A. D. Mendelow, andG. M. Teasdale. 1985. Pure cortical ischemia versus striatalischemia. Circulatory, metabolic, and neuropathologic conse-quences. Surg. Neurol. 24(1): 47–51.
54. Chen, S. T., C. Y. Hsu, E. L. Hogan, H. Maricq, and J. D.Balentine. 1986. A model of focal ischemic stroke in the rat:reproducible extensive cortical infarction. Stroke 17: 738–743.
55. Brint, S., M. Jacewicz, M. Kiessling, J. Tanabe, and W.Pulsinelli. 1988. Focal brain ischemia in the rat: Methods forreproducible neocortical infarction using tandem occlusion ofthe distal middle cerebral and ipsilateral common carotid ar-teries. J. Cereb. Blood Flow Metab. 8: 474–485.
56. Rubino, G. J., and W. Young. 1988. Ischemic cortical lesionsafter permanent occlusion of individual middle cerebral arterybranches in rats. Stroke 19(7): 870–877.
57. Memenzawa, H., M. L. Smith, and B. K. Siesjo. 1992. Penum-bral tissues salvaged by reperfusion following middle cerebralartrery occlusion in rats. Stroke 23: 552–559.
58. Ambach, G., and M. Palkovits. 1979. The blood supply of thehypothalamus in the rats. In Handbook of the Hypothalamus—Anatomy of the Hypothalamus, (P.J. Morgane and J. Panksepp,Eds.), Marcel Dekker, Inc., New York. pp. 267–377.
59. Paxinos, G., and C. Watson. 1986. The Rat Brain in StereotaxicCoordinates. Academic Press, San Diego.
279REDUCED SUSCEPTIBILITY TO ISCHEMIA IN NEUROENDOCRINE CELLS




















