
Carbonic Anhydrases in Anthozoan Corals–A Review
14
Carbonic anhydrases in anthozoan corals—A review Anthony Bertucci a,b, ,à , Aurélie Moya c,d,e,à , Sylvie Tambutté a , Denis Allemand a , Claudiu T. Supuran f , Didier Zoccola a,⇑ a Centre Scientifique de Monaco, Avenue Saint Martin, MC 98000, Monaco b Université de Nice Sophia Antipolis, Faculté des Sciences, Parc Valrose, 28 Avenue Valrose, 06108 Nice Cedex 2, France c INSU–CNRS, Laboratoire d’Océanographie de Villefranche, B.P. 28, 06234 Villefranche-sur-mer Cedex, France d UPMC University of Paris 06, Observatoire Océanologique de Villefranche, 06230 Villefranche-sur-mer, France e ARC Centre of Excellence for Coral Reef Studies, James Cook University, Townsville QLD 4811, Australia f Università degli Studi di Firenze, Laboratorio di Chimica Bioorganica, Rm. 188, Via della Lastruccia 3, I-50019 Sesto Fiorentino (Firenze), Italy article info Article history: Available online 30 October 2012 Keywords: Anthozoa Symbiosis Calcification Dissolved inorganic carbon Carbonic anhydrases abstract Coral reefs are among the most biologically diverse and economically important ecosystems on the pla- net. The deposition of massive calcium carbonate skeletons (biomineralization or calcification) by scle- ractinian corals forms the coral reef framework/architecture that serves as habitat for a large diversity of organisms. This process would not be possible without the intimate symbiosis between corals and photosynthetic dinoflagellates, commonly called zooxanthellae. Carbonic anhydrases play major roles in those two essential processes of coral’s physiology: they are involved in the carbon supply for calcium carbonate precipitation as well as in carbon-concentrating mechanisms for symbiont photosynthesis. Here, we review the current understanding of diversity and function of carbonic anhydrases in corals and discuss the perspective of theses enzymes as a key to understanding impacts of environmental changes on coral reefs. Ó 2012 Elsevier Ltd. All rights reserved. 1. Introduction The phylum Cnidaria is highly diverse with between 9000 and 13,000 living species that have been described by systematists. 1,2 The Class Anthozoa contains sessile organisms such as sea anemo- nes, sea whips and reef-building corals (order Scleractinia) (Fig. 1). Coral reefs are among the most biologically diverse, productive and economically important ecosystems on the planet. They determine the physical and ecological characteristics of entire coastlines. Reef structures represent essential habitat for a wide variety of marine animals. 3 Five hundred million people depend on healthy coral reefs for food, coastal protection, renewable resources, and income (largely due to tourism), with an estimated 30 million of the world’s poorest people depending entirely on coral reefs for food. 4 The formation of coral reefs is the result of two major aspects of scleractinian physiology: (i) Symbiosis, in the form of a mutualistic partnership between the cnidarian host and endosymbiotic Dinoflagellates of the genus Symbiodinium (referred to as zooxanthellae). Almost half of the cnidarians harbor unicellular Dinoflagellate sym- bionts in their tissue. 5 Anthozoa/Dinoflagellate symbioses are widespread in the marine environment and their impor- tant ecological role is widely documented. 6 Zooxanthellae gain protection from herbivores, exposure to light, and nutrients from the animal host, which allow them to fix large quantities of carbon dioxide (CO 2 ). In exchange, up to 95% of photosynthetic products are used by the host as source of energy. 7 (ii) Calcification, or the formation of an external skeleton through the transformation of dissolved inorganic carbon, into calcium carbonate (CaCO 3 ), in the form of aragonite. This process accounts for about 15% of the global calcium carbonate production. 8 Photosynthesis and calcification do not occur independently. Photosynthetic fixation of carbon dioxide and the precipitation of calcium carbonate are intimately linked both spatially and tempo- rally as it has been shown that calcification is enhanced by photo- synthesis (see Allemand et al. 9 for a review). Therefore coral physiology is tightly linked to the availability of dissolved inor- ganic carbon (DIC) in seawater, and DIC plays a major role at the interface between symbiosis and calcification. In the open ocean, where the average pH is about 8.2, the majority of DIC exists in the form of bicarbonate (HCO 3 ). This 0968-0896/$ - see front matter Ó 2012 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.bmc.2012.10.024 ⇑ Corresponding author. Tel.: +377 97 77 08 73; fax: +377 92 16 79 81. E-mail address: zoccola@centrescientifique.mc (D. Zoccola). Present address: ARC Centre of Excellence for Coral Reef Studies, James Cook University, Townsville QLD 4811, Australia. à These authors contributed equally to this work. Bioorganic & Medicinal Chemistry 21 (2013) 1437–1450 Contents lists available at SciVerse ScienceDirect Bioorganic & Medicinal Chemistry journal homepage: www.elsevier.com/locate/bmc
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Carbonic Anhydrases in Anthozoan Corals–A Review

Bioorganic & Medicinal Chemistry 21 (2013) 1437–1450
Contents lists available at SciVerse ScienceDirect
Bioorganic & Medicinal Chemistry
journal homepage: www.elsevier .com/locate /bmc
Carbonic anhydrases in anthozoan corals—A review
Anthony Bertucci a,b,�,�, Aurélie Moya c,d,e,�, Sylvie Tambutté a, Denis Allemand a, Claudiu T. Supuran f,Didier Zoccola a,⇑a Centre Scientifique de Monaco, Avenue Saint Martin, MC 98000, Monacob Université de Nice Sophia Antipolis, Faculté des Sciences, Parc Valrose, 28 Avenue Valrose, 06108 Nice Cedex 2, Francec INSU–CNRS, Laboratoire d’Océanographie de Villefranche, B.P. 28, 06234 Villefranche-sur-mer Cedex, Franced UPMC University of Paris 06, Observatoire Océanologique de Villefranche, 06230 Villefranche-sur-mer, Francee ARC Centre of Excellence for Coral Reef Studies, James Cook University, Townsville QLD 4811, Australiaf Università degli Studi di Firenze, Laboratorio di Chimica Bioorganica, Rm. 188, Via della Lastruccia 3, I-50019 Sesto Fiorentino (Firenze), Italy
a r t i c l e i n f o
Article history:Available online 30 October 2012
Keywords:AnthozoaSymbiosisCalcificationDissolved inorganic carbonCarbonic anhydrases
0968-0896/$ - see front matter � 2012 Elsevier Ltd. Ahttp://dx.doi.org/10.1016/j.bmc.2012.10.024
⇑ Corresponding author. Tel.: +377 97 77 08 73; faxE-mail address: [email protected] (D.
� Present address: ARC Centre of Excellence for CoUniversity, Townsville QLD 4811, Australia.
� These authors contributed equally to this work.
a b s t r a c t
Coral reefs are among the most biologically diverse and economically important ecosystems on the pla-net. The deposition of massive calcium carbonate skeletons (biomineralization or calcification) by scle-ractinian corals forms the coral reef framework/architecture that serves as habitat for a large diversityof organisms. This process would not be possible without the intimate symbiosis between corals andphotosynthetic dinoflagellates, commonly called zooxanthellae. Carbonic anhydrases play major rolesin those two essential processes of coral’s physiology: they are involved in the carbon supply for calciumcarbonate precipitation as well as in carbon-concentrating mechanisms for symbiont photosynthesis.Here, we review the current understanding of diversity and function of carbonic anhydrases in coralsand discuss the perspective of theses enzymes as a key to understanding impacts of environmentalchanges on coral reefs.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
The phylum Cnidaria is highly diverse with between 9000 and13,000 living species that have been described by systematists.1,2
The Class Anthozoa contains sessile organisms such as sea anemo-nes, sea whips and reef-building corals (order Scleractinia) (Fig. 1).Coral reefs are among the most biologically diverse, productive andeconomically important ecosystems on the planet. They determinethe physical and ecological characteristics of entire coastlines. Reefstructures represent essential habitat for a wide variety of marineanimals.3 Five hundred million people depend on healthy coralreefs for food, coastal protection, renewable resources, and income(largely due to tourism), with an estimated 30 million of theworld’s poorest people depending entirely on coral reefs for food.4
The formation of coral reefs is the result of two major aspects ofscleractinian physiology:
(i) Symbiosis, in the form of a mutualistic partnership betweenthe cnidarian host and endosymbiotic Dinoflagellates of thegenus Symbiodinium (referred to as zooxanthellae). Almost
ll rights reserved.
: +377 92 16 79 81.Zoccola).ral Reef Studies, James Cook
half of the cnidarians harbor unicellular Dinoflagellate sym-bionts in their tissue.5 Anthozoa/Dinoflagellate symbiosesare widespread in the marine environment and their impor-tant ecological role is widely documented.6 Zooxanthellaegain protection from herbivores, exposure to light, andnutrients from the animal host, which allow them to fixlarge quantities of carbon dioxide (CO2). In exchange, up to95% of photosynthetic products are used by the host assource of energy.7
(ii) Calcification, or the formation of an external skeletonthrough the transformation of dissolved inorganic carbon,into calcium carbonate (CaCO3), in the form of aragonite.This process accounts for about 15% of the global calciumcarbonate production.8
Photosynthesis and calcification do not occur independently.Photosynthetic fixation of carbon dioxide and the precipitation ofcalcium carbonate are intimately linked both spatially and tempo-rally as it has been shown that calcification is enhanced by photo-synthesis (see Allemand et al.9 for a review). Therefore coralphysiology is tightly linked to the availability of dissolved inor-ganic carbon (DIC) in seawater, and DIC plays a major role at theinterface between symbiosis and calcification.
In the open ocean, where the average pH is about 8.2, themajority of DIC exists in the form of bicarbonate (HCO�3 ). This

Figure 1. Simplified phylogeny of Metazoa. Porifera (sponges) is the most ancestral Phylum. Cnidaria appears as a sister group of Bilateria. Inside Cnidaria, the Anthozoanclass is basal. The red circle indicates the last cnidarian common ancestor. Some cnidarian species cited in this review are listed in their group-corresponding boxes(Scleractinia or Actiniaria).77
1438 A. Bertucci et al. / Bioorg. Med. Chem. 21 (2013) 1437–1450
external HCO�3 must be converted into CO2 to supply the photosyn-thetic apparatus. The source of DIC for calcification may originateeither from metabolic CO2 or from external HCO�3 .10,11 The inter-conversion between these different forms of DIC is crucial for bothphotosynthesis and calcification. In the animal kingdom, carbonicanhydrases (CAs) facilitate/catalyze the interconversion of CO2 intoHCO�3 and, thus, are likely to play a crucial role in coral physiology.Since 1959 with pioneering works of Goreau,12 numerous studieshave attempted to characterize CAs and their roles of CAs in antho-zoan physiology (see Table 1), either in symbiosis or calcificationprocesses. In this review, we discuss the current knowledge aboutanthozoan carbonic anhydrases diversity and function. In addition,we highlight recent molecular studies that investigate the responseof CAs to environmental parameters and provide insight into theresponse of corals to global change and the comprehension of coralphysiology.
2. Generalities
2.1. Carbonic anhydrases
The hydration of carbon dioxide (CO2) to bicarbonate (HCO�3 )and protons (H+) is a simple but essential reaction. These threechemical species are essential for multiple physiological processesin all organisms throughout the tree of life, but the reaction is tooslow to meet the physiological requirements of most organisms inwhich CO2 is produced in high amounts (e.g. aerobic respiration) orwhere CO2 is needed in high concentrations (e.g. in cyanobacteria,algae, plants, and other photosynthesizing organisms).13–15 Tomeet these requirements, the reaction must be catalysed by asuperfamily of enzymes known as carbonic anhydrases (CAs, EC4.2.1.1). These proteins evolved independently at least five timesin a variety of organisms found in all three life domains (Bacteria,Archaea, and Eukarya), and are present in virtually all living organ-isms and in relatively high amounts in various tissues/cell com-partments.16–18
The first, and best studied, class of these enzymes is the a-CAs.Members of this class are found in vertebrates, protozoa, algae, inthe cytoplasm of green plants, and in some Bacteria. A second class,the b-CAs are predominantly found in Bacteria, algae, and the chlo-roplasts of both mono- and dicotyledons, but also occur in manyfungi and some Archaea. The c-CAs are present in Archaea andsome Bacteria, and the final two classes, the d- and f-CAs, appearto occur exclusively in marine diatoms. The catalytic mechanismof a-CAs is well known.17,18 The cofactor associated with a-carbonic anhydrase is generally Zn2+ ion. In vertebrates, aCAs areinvolved in crucial physiological processes such as respirationand transport of CO2/HCO�3 , pH and inorganic carbon homeostasis,electrolyte secretion in a variety of tissues/organs, biosyntheticreactions (such as gluconeogenesis, lipogenesis, and ureagenesis),
bone resorption, calcification, tumorigenicity, and many otherphysiological or pathological processes.16,19,20 In algae, plants,and some bacteria, a-CAs play an important role in photosynthesisand other biosynthetic reactions in which CO2/HCO�3 aresubstrates.21
Alpha-CA isoforms differ in their catalytic activity, their sensi-tivity to inhibitors and activators, and their subcellular and tissuedistribution.16,22 Sixteen different a-CA isozymes have been identi-fied in mammals: four cytosolic isozymes (CA I–III, CA VII and XIII),five membrane-bound isozymes (CA IV, CA IX, CA XII, CA XIV andCA XV), two mitochondrial isozymes (CA VA and VB), and one se-creted CA isozyme, CA VI. The CA-related proteins (CARPs) havelost classical CA activity and their physiological function remainsunknown (CARP VIII, CARP X and CARP XI). However, their highlyconserved nature suggests a very important role in vertebrates.23
It should be noted that primates lack CA XV.19
2.2. Histology of Anthozoa
The anatomy of Cnidarians appears relatively simple as the tis-sues are made of two cellular layers (diploblastic), the ectoderm(facing the environment or the substrate) and the endoderm (fac-ing the coelenteron), separated by a cell-free connective layer,the mesoglea. The body of Anthozoa is a column-shaped structurewith an apical mouth surrounded by tentacles. The mouth servesas the interface between seawater and the internal cavity, the coe-lenteron. The oral tissue of the polyp is in contact with the externalmedium, while the aboral tissue is in contact with the substrate. Inzooxanthellate cnidarians, the symbiotic Dinoflagellates are lo-cated in the cytoplasm of oral endodermal cells. Zooxanthellaeare surrounded by a host-derived membrane vesicle called theperisymbiotic or perialgal membrane.24 In scleractinian corals,aboral tissue is referred to as calicoblastic epithelium and isresponsible for the secretion of the skeleton. The coral animal hostand its symbiotic Dinoflagellates (as well as other associated pro-karyotic organisms) brought together are referred to as holobiont.The structural anatomy and histology of a symbiotic scleractinianare presented in Figure 2 (modified from Bertucci et al.25 and Moyaet al.26).
2.3. Symbiosis and constraints on the availability of DIC forphotosynthesis
Both partners, the animal host and the intracellular Dinoflagel-lates, have adapted to an unusual situation by evolving a set of newcharacteristics (for a review, see Furla et al.27). The first conse-quence of this symbiotic way of life is the constraint for the hostto live in shallow waters highly exposed to light. The intracellularlocation of the algal partner and its isolation from the externalmedium are thought to allow the host to control symbiont density
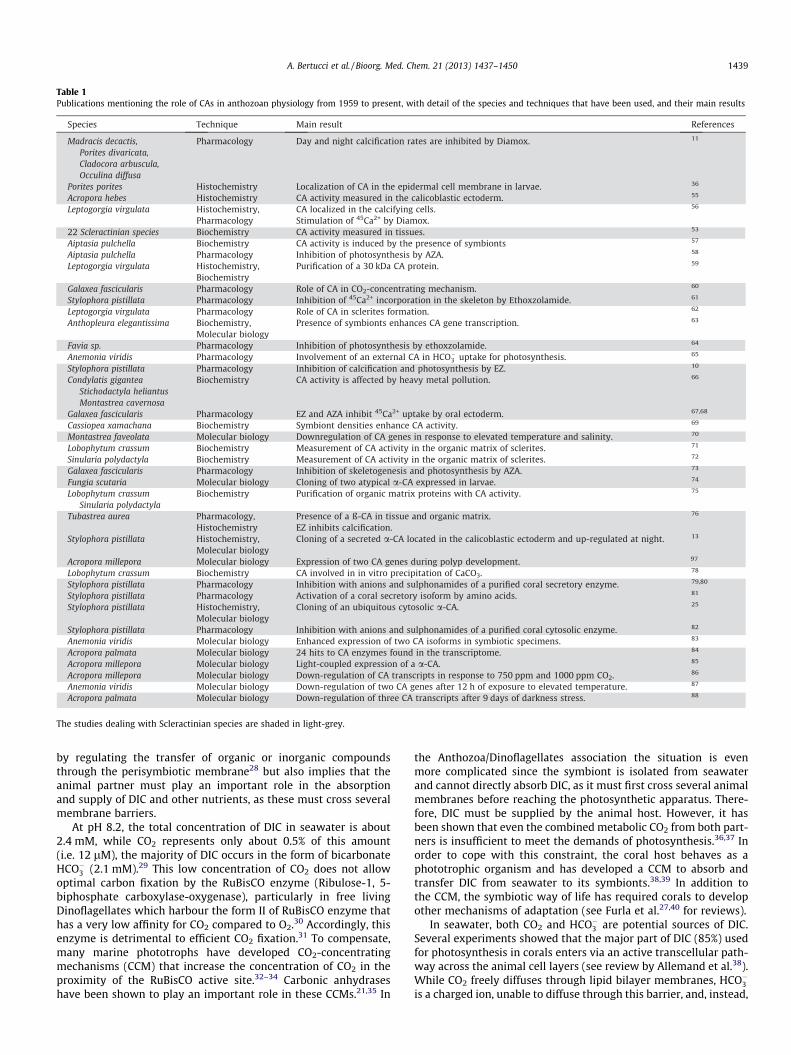
Table 1Publications mentioning the role of CAs in anthozoan physiology from 1959 to present, with detail of the species and techniques that have been used, and their main results
The studies dealing with Scleractinian species are shaded in light-grey.
A. Bertucci et al. / Bioorg. Med. Chem. 21 (2013) 1437–1450 1439
by regulating the transfer of organic or inorganic compoundsthrough the perisymbiotic membrane28 but also implies that theanimal partner must play an important role in the absorptionand supply of DIC and other nutrients, as these must cross severalmembrane barriers.
At pH 8.2, the total concentration of DIC in seawater is about2.4 mM, while CO2 represents only about 0.5% of this amount(i.e. 12 lM), the majority of DIC occurs in the form of bicarbonateHCO�3 (2.1 mM).29 This low concentration of CO2 does not allowoptimal carbon fixation by the RuBisCO enzyme (Ribulose-1, 5-biphosphate carboxylase-oxygenase), particularly in free livingDinoflagellates which harbour the form II of RuBisCO enzyme thathas a very low affinity for CO2 compared to O2.30 Accordingly, thisenzyme is detrimental to efficient CO2 fixation.31 To compensate,many marine phototrophs have developed CO2-concentratingmechanisms (CCM) that increase the concentration of CO2 in theproximity of the RuBisCO active site.32–34 Carbonic anhydraseshave been shown to play an important role in these CCMs.21,35 In
the Anthozoa/Dinoflagellates association the situation is evenmore complicated since the symbiont is isolated from seawaterand cannot directly absorb DIC, as it must first cross several animalmembranes before reaching the photosynthetic apparatus. There-fore, DIC must be supplied by the animal host. However, it hasbeen shown that even the combined metabolic CO2 from both part-ners is insufficient to meet the demands of photosynthesis.36,37 Inorder to cope with this constraint, the coral host behaves as aphototrophic organism and has developed a CCM to absorb andtransfer DIC from seawater to its symbionts.38,39 In addition tothe CCM, the symbiotic way of life has required corals to developother mechanisms of adaptation (see Furla et al.27,40 for reviews).
In seawater, both CO2 and HCO�3 are potential sources of DIC.Several experiments showed that the major part of DIC (85%) usedfor photosynthesis in corals enters via an active transcellular path-way across the animal cell layers (see review by Allemand et al.38).While CO2 freely diffuses through lipid bilayer membranes, HCO�3is a charged ion, unable to diffuse through this barrier, and, instead,

Figure 2. (A) Close-up of an individual coral polyp of the coral Pocillopora damicornis (credit Gergely Torda). (B) Schematic representation of a longitudinal section of a polyp(modified from Moya et al.26). (C) Histological organization of a symbiotic and calcifying Anthozoan. Cal., calicoblastic epithelium; Cn., cnidocyte; Ecto., ectoderm; Endo.,endoderm; M., mesoglea; Zoox., zooxanthellae.
1440 A. Bertucci et al. / Bioorg. Med. Chem. 21 (2013) 1437–1450
requires a carrier protein. Despite this additional cost, it has beenshown that the host uses the seawater HCO�3 as primary sourceof DIC for photosynthesis (see review by Allemand et al.38). CAshave undoubtedly played a crucial role in this adaptation wherethe animal partner must fulfill the DIC requirements of itssymbionts.
2.4. Biomineralization process and DIC equilibrium
Biomineralization is seen in representatives of the majority ofanimal phyla, with manifestation of a spectacular diversity ofendo- and exoskeletons as well as sensory and protective struc-tures composed from a variety of minerals.41 Common threadsconnecting basic structures in different taxa are of considerableinterest. It has recently been suggested that carbonic anhydraseswere components of a core molecular biomineralization toolkitpresent in the first metazoans and subsequently inherited by var-ious metazoan lineages as a toolkit for further development of keyelements of calcification.42 While the involvement of CA is well ac-cepted, the exact role that this enzyme plays in biomineralizationis still not known.43–46
Scleractinian corals precipitate an aragonitic calcium carbonate(CaCO3) skeleton resulting from the precipitation of calcium andinorganic carbon at the site of calcification. The source of DIC forcalcification is uncertain (see discussion in Allemand et al.9). Somestudies suggested that external HCO�3 is the preferential substratefor skeletogenesis,47 while others show that a large part (60–80%) of the carbon used for CaCO3 formation may originate fromrespired (metabolic) CO2.10,11
Depending on the source of carbon delivered to the site of cal-cification, CA could facilitate biomineralization in one of two ways:
(i) If metabolic CO2 is the major source of carbon for calcifica-tion, CA may accelerate the conversion of this CO2, whichdiffuses from the tissue to the skeletogenic fluid due to the
higher pH of the calcifying region,48,49 into HCO�3 , asdescribed by the following equations (⁄indicates the reactioncatalysed by CA activity):
�CO2 þH2O¡Hþ þHCO�3
HCO�3 ¡CO2�3 þHþ
CO2 þH2O¡2Hþ þ CO2�3
The dissociation of the second reaction has a pK near 10. At pHbelow 8, only a minor quantity of CO2�
3 can be present ([CO2�3 ]/
[ HCO�3 ] <0.01). The two H+ produced by this set of reactions maythen be removed from the site of calcification by a Ca2+-ATPasepresent within the calicoblastic epithelium50 which catalyzes theexchange 2H+/Ca2+.
(i) If seawater HCO�3 is the source of DIC for calcification, CAmay then accelerate the conversion to CO2 to buffer the acid-ity produced by the conversion of HCO�3 into CO2�
3 . In orderto proceed, this reaction needs a low [CO2] and a pH above8.51 Such a mechanism was initially suggested by Sikeset al.52 for coccolithophorids. This reaction is described bythe following equations (⁄indicates the reaction catalyzedby CA activity):
HCO�3 ¡CO2�3 þHþ
�Hþ þHCO�3 ¡CO2 þH2O
2HCO�3 ¡CO2�3 þ CO2 þH2O
While this chapter demonstrates the importance of CAs to bothsymbiosis and calcification, the following chapter describes howthe study of CA activity has led to an improved understanding ofDIC utilization in these processes.

Figure 3. Number of publications dealing with carbonic anhydrases in Anthozoans by decade. The number of publications in each field (Pharmacology, Histochemistry,Biochemistry and Molecular biology) is indicated in the corresponding portions of each bar. Some studies cover two or more fields and might be represented several times in asame decade bar; the total number of records is thus different from Table 1.
A. Bertucci et al. / Bioorg. Med. Chem. 21 (2013) 1437–1450 1441
3. The study of CAs in Anthozoans
Although CAs have multiple biological functions, including cel-lular transport and metabolism,53 research in Anthozoan mainlyfocussed on their involvement in photosynthesis and biominerali-zation (see Fig. 3 and Table 1). Until recently, the role of these en-zymes in corals was only inferred by looking at the effect of twochemical inhibitors, acetazolamide (Diamox, AZA) and ethoxzola-mide (EZ), on photosynthetic efficiency and/or calcification rate.The use of this method and potential improvements will be dis-cussed later in this review. The study of CAs in Cnidaria startedin 195912 but the number of publication records remains low till1999. It then increased sharply since the year 2000. This increaseis likely to be due to the development and the use of molecularbiology in Cnidarian research as shown by the greater proportionof this approach in Figure 3. These studies have detected CA activ-ity in all species investigated. For example Weis et al.54 detectedCA activity in 22 species of tropical cnidarians. It has also beenshown that CA activity is higher in host tissues harbouring symbi-otic algae.55
Table 1 also highlights the predominant use of Scleractinianspecies compared to other Anthozoans (anemones and soft corals),as Scleractinians represent 25 of the forty publications listed in Ta-ble 1. Of these 25 publications, only nine and five describe a clearlink between CA and biomineralization and photosynthesis,respectively. This low number of evidences most likely comes fromthe complex link between calcium carbonate deposition and pho-tosynthesis (e.g. phenomenon of light-enhanced calcifica-tion8,56,57), and the difficulty to study one of these processesindependently, a problem that does not exist when studyingnon-calcifying organisms such as anemones (Anemonia viridis,Aiptasia pulchella, Anthopleura elegantissima). It is interesting tonote that the two soft corals species (Sinularia polydactyla andLobophytum crassum), even if zooxanthellate species, have onlybeen used to study the role of CAs in biomineralization.
3.1. Role of CA activity in symbiosis
3.1.1. CA and photosynthesisThe role of CA in the epithelial transport of HCO�3 for photosyn-
thesis is well documented in marine algae and cyanobacte-ria.21,35,58 In symbiotic Cnidarians, CA activity was first describedeither in whole tissue extract or the entire organism.54,59–61 Ithas been shown that CA activity is important for photosyntheticactivity and appears to be linked to the symbiotic state. Indeedthe CA activity measured in Dinoflagellates-associated species is,on average, 29-times higher than that measured in non-symbioticcnidarian species54 and the presence of symbionts induces CAactivity in the tissues of the sea anemone A. pulchella. This activityis reduced 2.5-fold in aposymbiotic specimen, that is, hosts thathave lost their symbionts.62 These authors then suggested a roleof CAs as a ‘CO2 supply’.54 This role has also been inferred from sev-eral pharmacological experiments using AZA and/or EZ. The use of0.1–1 mM AZA inhibits 50–80% of the photosynthesis in Milleporaalcicornis and Aiptasia pallida.54,59 AZA is supposed to act preferen-tially on extracellular CAs whereas EZ is a more permeable mole-cule able to affect intracellular enzymes.63 AZA and EZ havesimilar effects on freshly isolated zooxanthellae (i.e. symbiontsout of animal tissues but still surrounded by their perisymbioticmembrane) from the coral Galaxea fascicularis.60 EZ has beenshown to have (i) a higher inhibitory effect than AZA in the holobi-ont (80 vs 60%) and (ii) a greater effect on the holobiont than oncultured zooxanthellae (80 vs 60%).60 Taken together, and withthe caveats that AZA and EZ have very diverse affinity for eachCA isoform16 and that Dinoflagellates may possess different classesof enzymes, these results led the authors to stipulate that only 20%of the CA activity involved in DIC uptake for photosynthesis is fromanimal origin. This percentage is probably underestimated due to alack of specificity of AZA and EZ. It is also interesting to note thatthese chemicals may have different effects depending on thetissue layer, for instance ectoderm versus endoderm.61,64 This

1442 A. Bertucci et al. / Bioorg. Med. Chem. 21 (2013) 1437–1450
observation suggested for the first time a differential distributionof the total amount of CAs and/or the existence of differentisoforms of CAs in coral compartments. Using antibodies raisedagainst human isoforms, CA has first been localized around thezooxanthellae60 and near or to the membrane surrounding them,59
confirming a role in DIC supply for photosynthesis.
3.1.2. CA and inorganic carbon uptakeIn addition, CA activity has been shown to play a role in DIC
absorption by the oral tissue of the symbiotic Mediterranean seaanemone A. viridis.61 Furla et al.65 measured this activity in purifiedectodermal and endodermal membranes, and in the cytosol of oralcell layers. The presence of CA activity associated with the oralectodermal cells, together with an externally directed H+-ATPaseactivity likely responsible for the acidification of the extracellularmedium, argues in favour of a DIC uptake based on primary extra-cellular dehydration of HCO�3 into CO2 by a membrane-bound CA.This mechanism is similar to that found in renal proximal tubulesof vertebrates.66 CAs in corals are therefore involved in the initialstep of inorganic carbon uptake. Once converted to CO2, DIC canreadily diffuse into the ectodermal cell, following the concentra-tion gradient created by the extrusion of H+ to the external med-ium. In the animal cytoplasm, where the pH is 7.4,67 CO2 isequilibrated with HCO�3 and a CA activity, identified by the previ-ously cited authors,11 prevents back-diffusion of CO2 and leakageof DIC.
3.1.3. CAs in DinoflagellatesThe mechanisms of HCO�3 transfer from ectodermal to endoder-
mal cells and across the symbiosome membrane to the zooxanthel-lae are still poorly understood but might involve HCO�3transporters, since the process is highly sensitive to anion trans-port inhibitors such as DIDS (4,40-Diisothiocyano-2,20-stilbenedi-sulfonic acid).60 Experiments on the kinetics photosynthesis,conducted under conditions of controlled inorganic carbon concen-trations, showed that marine eukaryotic and prokaryotic phyto-plankton has the ability to concentrate inorganic carbon forphotosynthesis (for a review, see Reinfelder et al.31). A role ofCAs in these CCMs was hypothesized and CA activity has sincebeen measured in Dinoflagellates (e.g. Lapointe et al.68). The exactrole of CAs in these organisms remains unclear but is certainly cru-cial since, as stated previously, animal CAs appear to account onlyfor 20% of the total CA activity involved in photosynthesis. So far,two models, based either on an external CA activity to dehydrateHCO�3 and/or an intracellular CA activity to convert CO2 are ac-cepted. Since the ionic environment is very different between sea-water (free-living lifestyle) and in the symbiosome, Dinoflagellatesneed to adapt to the symbiotic way of life. The evidence for a CCMinvolving a light-enhanced CA activity among in hospite Symbiodiniumis provided by Baillie and Yellowlees69 and Leggat et al.39,70 in thegiant clam Tridacna gigas. Al-Moghrabi et al.60 and Goiran et al.71
have extensively studied the DIC uptake in Symbiodinium sp.associated with the coral G. fascicularis or in culture. Their worksuggests that, in the symbiotic state, the transported HCO�3 isfinally protonated into CO2 by an H+-ATPase belonging to the zoo-xanthellae,72 with the aid of a CA activity located close to, or near,the symbiont membrane as suggested by immunolocalization;59,60
while in the free-living stage, this CA appears to be repressed andDIC is mainly absorbed by a Na+/HCO�3 co-transporter.
The contradictory evidences regarding CO2/HCO�3 uptake andexternal/intracellular location of CAs in Dinoflagellates may comefrom their complex phylogenetic history that could explain the factthat different genera or species express various modes or mecha-nisms of DIC acquisition.31 So far, the only fully characterized CAfrom a free-living Dinoflagellate is an external membrane-associatedo-CA from Lingulodinium polyedrum68 although two partially
characterized CAs are available in zooxanthellae where a 32 anda 70 kDa protein have been isolated.39,69
In conclusion, in the currently accepted model, CAs are involvedin all stages of DIC uptake and transport from seawater to symbi-onts (see upper part of Figs. 2C and 4), including the uptake ofDIC from the external medium, in the equilibration of HCO�3 /CO2
in the cytosol, and in the delivery of DIC to the photosyntheticmachinery of the algal partner.
3.2. CA activity in biomineralization
The first author seeking to demonstrate the role of carbonicanhydrases in the coral calcification was Goreau.12 This authorshowed that in four species of Scleractinian corals (Madracis decactis,Porites divaricata, Cladocora arbuscula, Oculina diffusa), AZA decreasedcoral calcification rates by approximately 50%, and suggested thatthe enzyme aids calcification by removing carbonic acid producedby calcification itself from the skeletogenic site. In agreement withthis findings, Isa and Yamazato73 demonstrated that CA activitywas correlated with the rate of calcification and similar results wereobtained with AZA or EZ on two Scleractinian species, Stylophorapistillata11,74 and G. fascicularis.75,76 In these studies, calcificationdecreased by as much as 73% but was not completely abolishedcalcification, suggesting that CA, although important, is notabsolutely necessary for the calcification process.
Inhibition also varies according to light conditions. Goreau12
found that when incubated in the light with 1 mM AZA, calcifica-tion of P. divaricata or O. diffusa decreased to a level similar tothe dark rate (51% inhibition). When incubated in the dark, the rateof calcification upon addition of CA inhibitor further decrease (34%inhibition) suggesting that the action of animal CA and that of pho-tosynthesizing zooxanthellae are ‘similar and probably synergistic’.However, the subsequent inhibition observed at night suggests anadditional role of CA in the calcification process.
The involvement of CA in the calcification is also indicated bylocalization of this enzyme in the calicoblastic epithelium in con-tact with the skeleton. By histochemistry, Hayes and Goreau36
localized CA in the membrane of the epidermal cells of planulaeof the coral Porites porites. By a similar method, Isa and Yamazato73
localized CA within the calicodermis of the coral Acropora hebes.The role of CA in the biomineralization process is not restricted
to Scleractinian zooxanthellate corals. Indeed, in the non-symbioticoctocoral Leptogorgia virgulata, Lucas and Knapp79 showed that CAplays a pivotal role in the formation of calcium carbonate spicules.In this cnidarian, CA involved in the calcification process have beenlocalized to the spicule vacuole membrane as well as to the insideof electron-dense Golgi-derived vesicles.78,79 Curiously, in the octo-coral L. virgulata, treatment with sulfonamide causes a 30% in-crease in the rate of 45Ca2+ incorporation in the spicules.78 One ofthe explanations proposed by the authors is that inorganic carbonwas not limiting for calcification in this species. Another proposedhypothesis is that CA may have an inhibitory role in the calcifica-tion process for some species, as observed in other organisms suchas Molluscs.45
Several organic matrix proteins with CA domains and activitieshave been characterized in other gorgonians.80–83 Rahman et al.83
suggest that organic matrix proteins in alcyonarian are not onlystructural proteins, but also catalysts. Similar to the nacrein iden-tified in molluscs, these authors reported a protein that has bothcalcium-binding and CA activity.83 They showed that this proteininhibited the precipitation of CaCO3 from a saturated solution con-taining CaCl2 and NaHCO3, indicating that it can act as a negativeregulator for calcification in sclerites of alcyonarians. They sug-gested that the organic matrix proteins in alcyonarian corals possi-bly binds calcium82 and works as a CA that catalyses the HCO�3
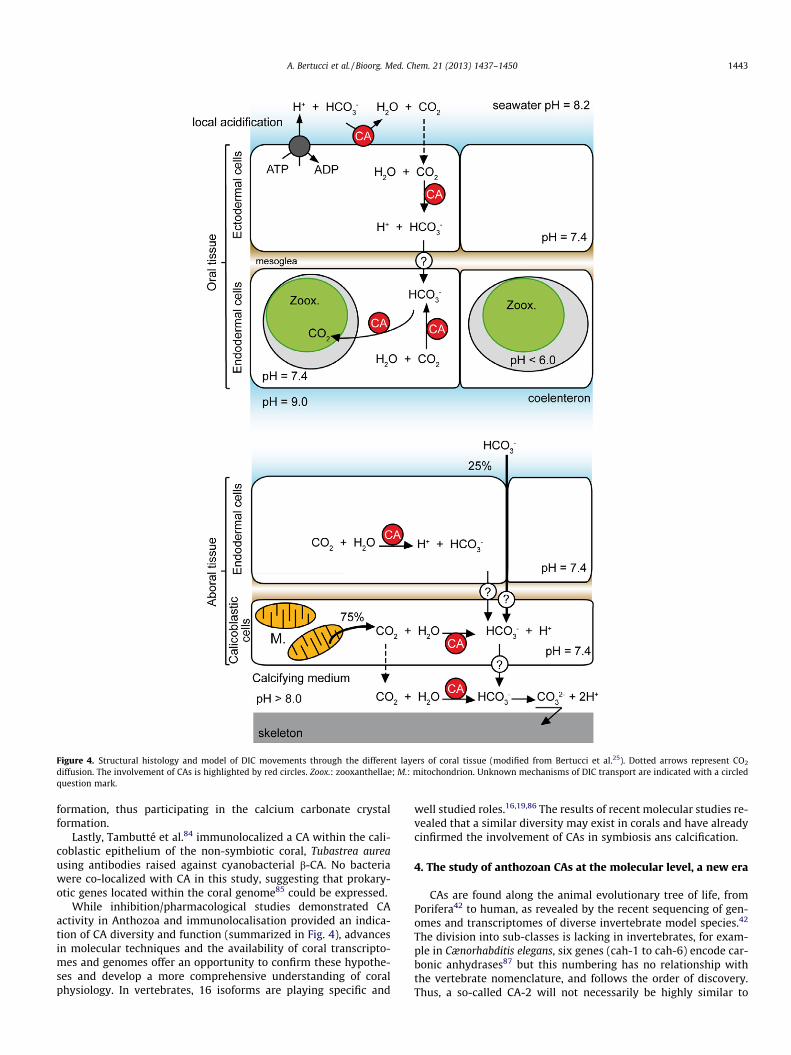
Figure 4. Structural histology and model of DIC movements through the different layers of coral tissue (modified from Bertucci et al.25). Dotted arrows represent CO2
diffusion. The involvement of CAs is highlighted by red circles. Zoox.: zooxanthellae; M.: mitochondrion. Unknown mechanisms of DIC transport are indicated with a circledquestion mark.
A. Bertucci et al. / Bioorg. Med. Chem. 21 (2013) 1437–1450 1443
formation, thus participating in the calcium carbonate crystalformation.
Lastly, Tambutté et al.84 immunolocalized a CA within the cali-coblastic epithelium of the non-symbiotic coral, Tubastrea aureausing antibodies raised against cyanobacterial b-CA. No bacteriawere co-localized with CA in this study, suggesting that prokary-otic genes located within the coral genome85 could be expressed.
While inhibition/pharmacological studies demonstrated CAactivity in Anthozoa and immunolocalisation provided an indica-tion of CA diversity and function (summarized in Fig. 4), advancesin molecular techniques and the availability of coral transcripto-mes and genomes offer an opportunity to confirm these hypothe-ses and develop a more comprehensive understanding of coralphysiology. In vertebrates, 16 isoforms are playing specific and
well studied roles.16,19,86 The results of recent molecular studies re-vealed that a similar diversity may exist in corals and have alreadycinfirmed the involvement of CAs in symbiosis ans calcification.
4. The study of anthozoan CAs at the molecular level, a new era
CAs are found along the animal evolutionary tree of life, fromPorifera42 to human, as revealed by the recent sequencing of gen-omes and transcriptomes of diverse invertebrate model species.42
The division into sub-classes is lacking in invertebrates, for exam-ple in Cænorhabditis elegans, six genes (cah-1 to cah-6) encode car-bonic anhydrases87 but this numbering has no relationship withthe vertebrate nomenclature, and follows the order of discovery.Thus, a so-called CA-2 will not necessarily be highly similar to

1444 A. Bertucci et al. / Bioorg. Med. Chem. 21 (2013) 1437–1450
the human CA II, either in term of sequence, activity or location.However, among invertebrates, phylogenetic comparisons withvertebrates and a detailed analysis of CA sequences can offer in-sight into the function and the location of the protein.
In Cnidarians, the first CA to be cloned was one in the sea anem-one A. elegantissima.88 Since then, different CAs have been charac-terized, either by EST sequencing or by conventional techniques.Based on the available amino acid sequences in Genbank data li-brary (cloned RNA sequences and Acropora digitifera, Nematostellavectensis and Hydra magnipapillata genome sequences) and ESTdata mining from A. viridis,89 Acropora millepora,90,91 Pocilloporadamicornis92 and non-published results (see for instance data fromMatz lab—http://www.bio.utexas.edu/research/matz_lab/matzlab/Data.html), we performed a phylogenetic analysis comparing hu-man and cnidarian CAs (Fig. 5). As found in other studies,93,94 thephylogenetic tree reveals three main clusters: (1) the membrane-bound or secreted proteins, (2) the cytosolic and mitochondrialproteins, (3) and the CARPs, which suggests that these 3 familiesevolved from a common ancestor prior to the separation betweenCnidaria and Bilateria. However, because human CAs are notgrouped with the anthozoan ones, we suggest that the subclassesappeared independently in each phylum. In each cluster, we couldfind redundancy in some species (Fig. 5). Gene duplication is pos-sible but evidence for these events must be found in genome asduplication can arise as an artefact due to sequencing techniques.Until now, only two anthozoan genomes, N. vectensis and A. digitifera,are available but additional genome sequencing will offer insightsinto the question of these redundancies. Finally regarding evolu-tion, as expected95 the CAs of Actiniaria (sea anemones) appearto have evolved before the coral ones in each cluster.
4.1. Secreted or membrane-bound a-CA
The first two membrane-bound CAs cloned in cnidarians wereboth from the coral Fungia scutaria.96 These two sequences, namedFCA a and b, contain 201 and 225 amino acids respectively. Bothsequences are shorter than usual CAs. The FCA a and b sequencespossess a truncated catalytic site containing the three histidine res-idues but lacking respectively 14 and 6 amino acids from the C-terminus. No catalytic studies were performed on these CAs. Recently,other secreted or membrane-bound CAs have been cloned from theScleractinian corals S. pistillata,14 A. millepora97 and A. digitifera.98
They form a distinct group (group III in Figure 5) among the se-creted/membrane bound CAs. The a-CA from S. pistillata, referredto as STPCA (ACA53457), consists of 324 amino acids and containsa complete CA catalytic site, as predicted for functional CAs.93 Sim-ilarly to FCA a and b, the presence of a signal peptide indicates thatSTPCA protein is not cytosolic but secreted or membrane-bound.However, the protein does not contain a predictive GPI anchor siteor transmembrane domain, suggesting that it is secreted ratherthan membrane-bound. Activity and in vitro inhibition studieshave been performed on this enzyme (see below). STPCA was alsolocalized by immunochemistry within the calicoblastic ectodermsuggesting that this isoform is directly linked to calcification. Fur-thermore, Moya et al.14 demonstrated, using real-time PCR, thatexpression of this secreted CA isoform increased two-fold in thedark relative to the light. This result lead the authors to hypothe-size that up-regulation of this CA gene allows the coral to copewith night acidosis and reinforces the pharmacological experi-ments of Goreau12 that suggested an additional role of CA at night.
Another group of secreted/membrane isoforms (group II inFig. 5) groups with the A. millepora CA C007-E7 (ACJ64662). ThisCA groups with vertebrate IV/XV-type carbonic anhydrases, andsimilar to other members of this group, is predicted to be GPI an-chored. This CA is expressed most strongly in a restricted area ofthe aboral end of the metamorphosing larva and primary polyp,97
an observation that is consistent with involvement in the onsetof calcification. In slightly older polyps, the expression in the aboraldisc declines and shifts to the tentacles. In a microarray experi-ment on A. millepora, this CA was found to be up-regulated duringdevelopment from a non-calcifying planula to a calcifyingpolyp.97 Homologs of this CA are found in A. tenuis, P. astreoides,and S. pistillata. In S. pistillata, a new CA (STPCA-4) has recentlybeen cloned and is part of another group (Fig. 5, group I). The studyof its activity is in progress (Bertucci et al., personal data).Combined Blast searches of the A. millepora transcriptome andphylogenetic analysis led to the identification of a total of 10membrane-bound or secreted isoforms,26 but their localizationand role, again, remain to be determined. Interestingly, only onesecreted or membrane-bound CA is found in the A. viridis EST data-base89 and two in N. vectensis genome suggesting that Actiniariansmay possess an ancestral and less diversified set of secreted CAs.
4.2. Cytosolic and mitochondrial a-CAs
To date, 18 intracellular CAs have been identified in Anthozoans(Fig. 5). In addition to the first cloned A. elangantissima sequence(AAD32675), other cytosolic CAs have been found in symbiotic(A. viridis) and non-symbiotic (N. vectensis) Actiniarians. Becausethis isoform is strongly up-regulated when the facultative symbi-otic anemone takes up endosymbionts,88 the A. elangantissima genewas described as a ‘‘symbiosis gene’’. Similar results have been ob-tained in other species. For instance, Ganot et al.99 identified a CAisoform that is related to the mammalian cytosolic isoform CAI/II,and shows an up-regulated transcription in symbiotic specimensof A. viridis. However, clear counterparts of this soluble cytosolicCA are present in both N. vectensis and H. magnipapillata (Hydro-zoa), neither of which harbour symbionts (Fig. 5, group V).
In corals, the phylogenetic tree shows that intracellular CAs fallinto two clusters that are equally similar to both hCAIII or hCA IIisozymes, as determined by a BLAST comparison.
Concerning the CAs related to the hCAII (group IV in Fig. 5),Bertucci et al.25 characterized a widespread isoform in S. pistillata,named STPCA-2 (ACE95141). The presence of this CA in the cyto-plasm of cells containing symbionts, as well as in aboral cells,has been shown by immunolocalization using specifically raisedantibodies. STPCA-2 might be the same CA that was previouslyidentified11,59,60 and hypothesized to play a role as a HCO�3 /CO2
equilibration tool in the cytosol and/or might serve to deliverCO2 to the symbionts.
Concerning the CAs related to the hCAIII (group V), the first CAwas found in A. millepora (ACJ64633).97 This CA is expressed in theoral half of the metamorphosing larva and the entire ectoderm ofthe primary polyp, except the aboral disc and the oral pore. In olderpolyps this carbonic anhydrase is expressed in the septa, wherecalcification occurs to form adult structures. Other cytosolic CAswere found in A. digitifera, Porites astreoides, S. pistillata (STPCA-3,Bertucci et al., unpublished data) and P. damicornis.
Combined Blast search in the A. millepora transcriptome andphylogenetic analysis led to the identification of a total of 5 cyto-solic isoforms26 but their localization and specific role are un-known. Since the diversity of CA isoforms in corals is likely to beas great as that in vertebrates (15 isoforms in human), further anal-ysis must be performed in order to completely understand the roleof CAs in anthozoan physiology.
4.3. CA-related proteins
As previously mentioned, CARPs have lost their classical CAactivity. Four cnidarian proteins group with the cytosolic formsof CARPs (i.e. hCARP X and XI). hCARP VIII homologous could notbe found in any of the available cnidarian libraries or genomes. It

Figure 5. Consensus tree of human and anthozoan carbonic anhydrase domain protein sequences. Alignment of sequences was done with ClustalW. Tree was thenconstructed using PhyML 3.0137 starting from a random tree, using the best-fitting model of protein sequence evolution by using ProtTest 1.3.138 cpREV + I + G + F substitutionmodel with gamma-distributed rate of 0.265 was then used. ML bootstrap proportions were obtained after 1000 bootstrap replicates. Only values above 50% are indicated.The branch lengths are proportional to the number of substitutions per site (see scale bar).
A. Bertucci et al. / Bioorg. Med. Chem. 21 (2013) 1437–1450 1445

1446 A. Bertucci et al. / Bioorg. Med. Chem. 21 (2013) 1437–1450
is not a surprise since no CARP VIII orthologs also do not occur inbilaterian protostomes, and it seems likely that the origin of CARPVIII occurred after the separation of the Protostomia and Deutero-stomia lineages. Comparison of human and cnidarian CARPs showsthat the His94 is replaced by an arginine (Aspatwar et al.100 for re-view) but also by a lysine (in N. vectensis). Furthermore, as inhCARP XI, the His96 is replaced by an arginine in S. pistillata. At last,in most of the cnidarian CARPs, the His119 is replaced by a gluta-mine (data not shown). So as for the human CARPs, it is not possi-ble for Zn2+ to interact with the histidine residues crucial for activeCAs. In mammals, CARP X and XI are mostly expressed in the brain,suggesting that they may contribute to the development of thenervous system.101,102 It remains unknown whether the cnidarianCARPs are also expressed in nerve cells. Alternatively, CARPs inCnidaria may play a role in calcium signalling, as some CARPs areassociated with the calcium signalling-mediated inositol 1,4,5 tri-phosphate (IP3) receptor,103 Miyamoto104 suggested that, in themantle of the pearl oyster, CARP might be involved in shell forma-tion regulated by IP3 receptor signalling. This role might exist alsoin Cnidaria.
We are just at the beginning of the molecular era of Anthozoanresearch, as the first new generation genomes became availableonly recently.98,105 Structure/function data will be useful to betterunderstand not only how anthozoan CAs work but also bring newinsight on the evolution of vertebrate CAs.
5. Impact of environmental stresses on CA transcription andactivity
Symbiotic cnidarians are very sensitive to environmental stres-ses such as increased temperature and carbon dioxide partial pres-sure, but also pathogens, pollutions and changes in salinity.106–108
Considering the central role of carbonic anhydrases in coral phys-iology, it is reasonable to hypothesize that environmental stressesthat influence coral physiology might influence CA gene expressionor enzyme activity. Indeed, recent experiments demonstrate thatthis is the case and offer confirmation of, and insight into, coralphysiology and response to stressors.
5.1. Elevated temperature
Temperature is certainly the most studied stressor in the coralliterature,108 as it is known to induce symbiosis breakdown, lead-ing to zooxanthellae and/or pigments loss, referred to as bleaching.Three studies, to date, measured the impact of increased tempera-ture on genes coding for carbonic anhydrases, and all of themshowed a significant decline in response to elevated tempera-ture.109–111 More specifically, Moya et al.111 showed that twoA. viridis carbonic anhydrases (CA2-c and CA2-m) were profoundlydown-regulated in response to elevated temperature, as early as12 h into the experiment. Interestingly, the same CAs were previ-ously shown to be highly up-regulated in symbiotic specimenscompared to aposymbiotic specimens99 as mentioned in the para-graph 4.2. Authors suggest that immediate down-regulation ofthose CAs is consistent with a rapid limiting CO2 supply after ther-mal stress, although it is still to be determined whether drop of CAsis part of a CCM limitation strategy imposed by the host or whetherit is to create resilience to limited CO2 demand, subsequent to thechloroplast dysfunction.
5.2. Ultraviolet radiation
Still in the study of Moya et al.,111 UVR exposure (1.25 mW/cm2) had no effect on the transcription of the two CA genes(CA2-c and CA2-m) in A. viridis. On the contrary, in the coral
Montastrea faveolata,109 the only observed CA gene showed signif-icant up-regulation under UVR exposure.
5.3. Ocean acidification (OA)
As the world’s oceans are sinks for CO2 released into the atmo-sphere, elevated emissions are leading to a decrease in seawaterpH. A process known as Ocean Acidification (OA).112 Although sev-eral studies have demonstrated the negative effects of OA on calci-fication rates in corals,106,113–115 other studies report oppositeresults on gross calcification.116–118 Considering the role of CAs incalcification, it is interesting to determine the impact of increasedpCO2 on CA expression/activity. To our knowledge, only one studyhas investigated the effect of pCO2 increase on CA gene expressionin cnidarians. Moya et al.26 showed that in nonsymbiotic coraljuveniles, the expression of a large number of the genes encodingCAs (4 and 8 of the total of 15 CAs identified in the A. milleporatranscriptome), decreased significantly under 750 and 1000 ppmCO2, reflecting the medium and high CO2 scenarios for the 21stcentury predicted by the IPCC.119 The reasons for this are not clear;presumably, there is a reduced requirement for carbonic anhydraseactivity in the extracellular calcifying medium and in the calicob-lastic ectoderm under acidification, possibly to maintain appropri-ate pH at the site of calcification, and/or in other coral tissues. Incontrast, Vidal-Dupiol et al.120 have investigated global transcrip-tomic changes in adult scleractinian coral P. damicornis exposedto acidification (pH 7.4) during a 3-week period and found that car-bonic anhydrases were up-regulated at low pH values (7.8 and 7.4)but down-regulated at pH 7.2. This discrepancy between resultsmay be due to a species differences and / or stage-specificresponses.
5.4. Salinity
Edge et al.109 measured a significant increase in the expressionof a CA gene under two elevated salinities. Elevated salinity isknown to affect the ionic regulation,121 acid–base balance122 andosmoregulation in marine organisms,122,123 giving a possibleexplanation for the up-regulation of a carbonic anhydrase in thisstudy.
5.5. Pollutants
Gilbert and Guzman124 demonstrated that carbonic anhydraseactivity decreased in two anemones, Condylactis gigantea andStichodactyla helianthus, that were exposed to metal contamination(copper, nickel, lead, and vanadium). Similarly, Montastrea cavern-osa collected from metal-contaminated sites compared to levelsfound in specimens from pristine sites. Also, Bielmyer et al.125
observed that total CA activity decreased in the corals Acroporacervicornis and M. faveolata when exposed to copper, however,while the effects were not significant in the coral P. damicornis.In the latter study, a strong correlation was observed between CAactivity and growth. In both studies, CA activity was affected beforeany observation of algae expulsion, leading Bielmyer et al.125 tosuggest that this decrease in CA activity might have affected thesymbionts by reducing the CO2 available for photosynthesis. Suppres-sion of CA activity could also have decreased the substrate availablefor use in calcification and skeletal growth of the animal host.
To conclude, cnidarian carbonic anhydrases are negatively af-fected by temperature, ocean acidification, and pollutants, but ap-pear to be unaffected or positively affected by UVR exposure andincrease in salinity. Together, those studies show that CA geneexpression/protein activity may be an important biomonitoringtool for assessing coral health prior to irreversible damage.

Table 3Inhibition constants of anionic inhibitors against human isozymes hCA I, II and VI, andthe CAs from the coral Stylophora pistillata, STPCA and STPCA-2 for the CO2 hydrationreaction at 20 �C126
Inhibitor Ki (nM)
A. Bertucci et al. / Bioorg. Med. Chem. 21 (2013) 1437–1450 1447
However, the method chosen is important, as the carbonic anhydraserepertoire in cnidarians is relatively complex26 (paragraph 4 in thisreview). Hence, measuring the activity of all CA proteins in anorganism might not be as precise as measuring gene expressionof a single and specific isoform of CA (see paragraph 6). Thisstatement is reinforced by the fact that CA play various roles incnidarian physiology, from symbiosis to calcification, acid–basebalance, or again ion regulation, and it is likely that environmentalstresses affect those isoforms differently.
6. Pharmacological study of Cnidarian CAs
Historically, the study of carbonic anhydrases in cnidarians hasprimarily relied on the application of AZA (acetazolamide) and EZ(ethoxzolamide) to whole, living organisms. Although these exper-iments highlighted the importance of CA and the comprehension ofbasal coral physiology (see paragraphs 3.1 and 3.2 in this review),they suffer from one major limitation: AZA and EZ do not show anystrong specificity for individual isoforms. Their inhibition con-stants (ki) for human isoforms range from 2.5 to 250 nM and from0.8 to 93 nM respectively (hCA III being insensitive to sulphona-mides/sulphamates/sulphamides18). In the coral literature relatedto calcification, the final concentration of inhibitors used rangesfrom 300 lM74 to 1 mM48 of EZ and from 1 lM78 to 1 mM12,76 ofAZA. This is roughly similar to what has been used for the studyof photosynthesis in corals, with 100 lM59,60 to 1 mM54 of AZAand 10060 to 600 lM11,64 of EZ. Which, regarding the diversity ofisoforms and functions in corals, makes any interpretation difficult.
However, the recent cloning and purification of recombinantisoforms of CAs from S. pistillata14,25 paves the way to a more accu-rate study of this class of enzymes in corals. The productions ofSTPCA and STPCA-2 by heterologous expression in mammaliancells and study of their catalytic study revealed that the kineticparameters of these cnidarian CAs are similar to human enzymes(see Table 2). The catalytic constant (kcat) of STPCA(3.1 � 105 s�1) is very similar to the human secreted CA VI(3.4 � 105 s�1) whereas STPCA-2 has a higher activity level(5.6 � 105 s�1). With a kcat/Km ratio of 8.3 � 107 M�1 s�1, STPCA-2appears as a very efficient enzyme. Moreover it may also be notedthat CAs from S. pistillata, as with human isoforms, have inhibitionconstants of the same order of magnitude concerning acetazola-mide (16 vs 74 nM) and ethoxzolamide (39.4 vs 105 nM).126 Theseresults clearly show that, so far, pharmacological approach has notbeen specific enough for the study of individual CAs in corals. In-deed, if we hypothesize that any other hypothetical cnidarian iso-forms would have kinetic parameters in the range of the humanones, the concentrations cited above are able to inhibit the totalCA activity of a living coral.
Table 2Relative activity level and kinetic parameters for the CO2 hydration reaction catalysedat 20 �C and pH 7.5 in 10 mM Hepes buffer by the human carbonic anhydrasesisoforms I, II and VI, as well as the Stylophora pistillata enzymes STPCA and STPCA-2,and their inhibition data with AZA and EZ (from139 and25)
Isozyme Relative activity kcat (s�1) kcat/Km (M�1 s�1) ki (10�9 M)
AZA EZ
hCA I Moderate 2.0 � 105 5.0 � 107 250 25hCA II Very high 1.4 � 106 1.5 � 108 12 8hCA VI Moderate 3.4 � 105 4.9 � 107 11 43hCA IX High 3.8 � 105 5.5 � 107 25 34hCA XII Moderate 4.2 � 105 3.5 � 107 5.7 22STPCA Moderate 3.1 � 105 4.6 � 107 16 39.4STPCA-2 High 5.6 � 105 8.3 � 107 74 105
Values are means of three measurements.
As they are involved in important physiological processes,mammalian CAs have been identified as therapeutic targets withthe potential to be inhibited or activated to treat a wide range ofdisorders, including glaucoma, epilepsy, obesity, and can-cer.18,86,127 Similarly, isoforms from other organisms have beencharacterized in the last few years and constitute new drug targetsto treat malaria or fungal and bacterial infection (e.g. Innocentiet al.128). The use of such drugs in non-model organisms, likecorals, might be helpful to decipher the role of CAs. The activityof purified recombinant STPCA and STPCA-2 have been testedwith two classes of classical CA inhibitors, anions126,129 andsulphonamides.126,130 The inhibition data are presented in Tables3 and 4 respectively, where data of human isozymes are includedfor comparison purposes.
Inhibition data reveal that STPCA and STPCA-2 show differentprofiles. STPCA appears to be slightly more sensitive to anions thanSTPCA-2, even if the inhibitory constants are in the same nanomo-lar range for both coral enzymes. Iodide (I�) and sulfamide(H2NSO2NH2) are the best inhibitors of STPCA and STPCA-2 respec-tively. Iodide is more than 3000 times more efficient on the se-creted isoform whereas the best selectivity for STPCA-2 isobtained with sulfamic acid (H2NSO3H), with a lower ratio of about95 times. It is interesting to note that some authors have utilizediodide instead of stilbene derivatives (e.g. DIDS) as an inhibitor ofHCO�3 transporter in order to study the transport of inorganic car-bon in coral tissues for photosynthesis and calcification (e.g. 2 mMused by Furla et al.11). Even if no effect has been measured on cal-cification, the use of iodide might lead to misinterpretation of re-sults and should be used with care for in vivo studies in corals.Among the best anionic inhibitors, it can be surprising to find car-bonate ion as a relatively good inhibitor of STPCA. Indeed, STPCA issecreted by calicoblastic cells in the external calcifying medium(ECM) where CO2�
3 is the ultimate form of inorganic carbon usedfor skeleton deposition (see paragraph 2.4). This result could reveala negative feedback on STPCA in order to regulate the biomineral-ization process.
The nature and composition of the ECM is still a matter ofdebate in coral biology as it is still unclear if it is isolated from oropen to the outside environment.8,131 Consequently its physico-chemical and ionic composition is debated.75,132–134 The fact that
hCA I hCA II hCA VI STPCA STPCA-2
F� >300 >300 0.60 0.62 0.92Cl� 6 200 0.72 0.50 0.53Br� 4 63 0.73 0.0097 0.96I� 0.3 26 0.81 0.0090 33.0CNO� 0.0007 0.03 0.69 0.59 0.69SCN� 0.2 1.6 0.89 0.68 0.51CN� 0.0005 0.02 0.07 0.58 0.86N�3 0.0012 1.5 0.07 0.52 4.68HCO�3 12 85 0.80 0.45 7.81
CO2�3
15 73 0.69 0.010 0.24
NO�3 7 35 0.76 0.56 0.99NO�2 8.4 63 0.82 0.77 3.15HS� 0.0006 0.04 0.71 0.58 3.94HCO�3 18 89 14.2 0.41 0.43
SO2�4
63 >200 9.9 0.91 0.33
H2NSO2NH2 0.31 1.13 0.07 0.010 0.0057H2NSO3H 0.021 0.39 0.09 0.81 0.0085Ph-B(OH)2 58.6 23.1 0.82 0.68 0.0081Ph-AsO3H2 31.7 49.2 13.9 0.78 0.0672
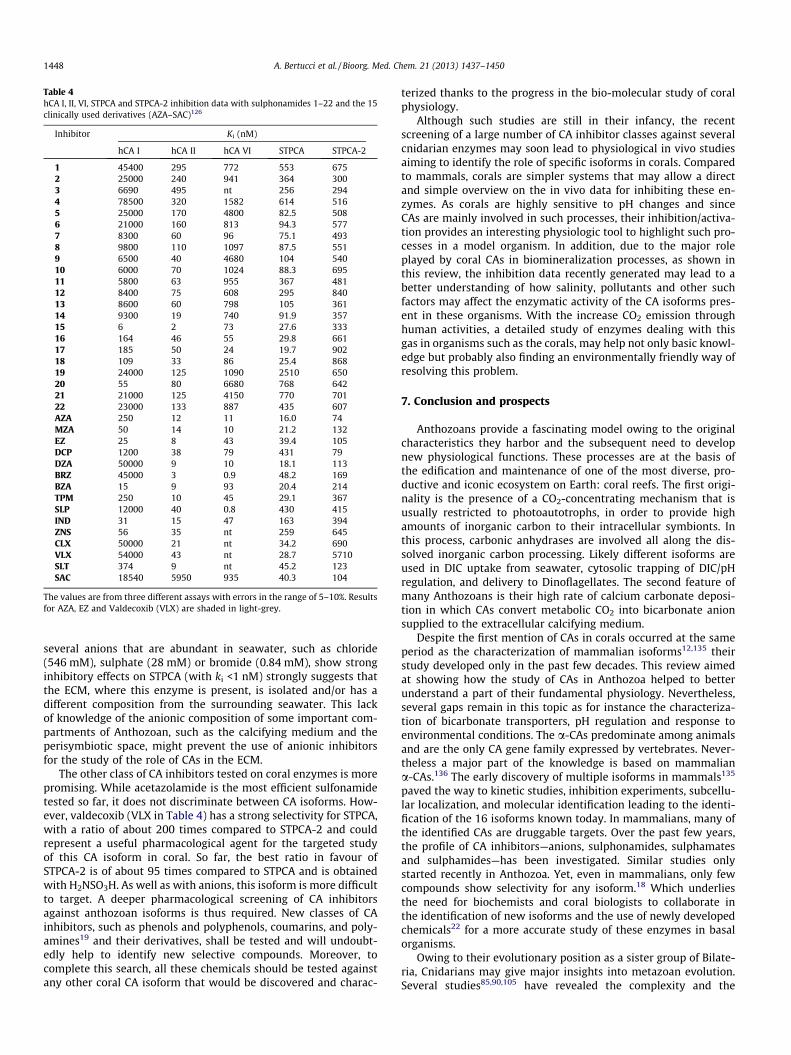
Table 4hCA I, II, VI, STPCA and STPCA-2 inhibition data with sulphonamides 1–22 and the 15clinically used derivatives (AZA–SAC)126
Inhibitor Ki (nM)
hCA I hCA II hCA VI STPCA STPCA-2
1 45400 295 772 553 6752 25000 240 941 364 3003 6690 495 nt 256 2944 78500 320 1582 614 5165 25000 170 4800 82.5 5086 21000 160 813 94.3 5777 8300 60 96 75.1 4938 9800 110 1097 87.5 5519 6500 40 4680 104 54010 6000 70 1024 88.3 69511 5800 63 955 367 48112 8400 75 608 295 84013 8600 60 798 105 36114 9300 19 740 91.9 35715 6 2 73 27.6 33316 164 46 55 29.8 66117 185 50 24 19.7 90218 109 33 86 25.4 86819 24000 125 1090 2510 65020 55 80 6680 768 64221 21000 125 4150 770 70122 23000 133 887 435 607AZA 250 12 11 16.0 74MZA 50 14 10 21.2 132EZ 25 8 43 39.4 105DCP 1200 38 79 431 79DZA 50000 9 10 18.1 113BRZ 45000 3 0.9 48.2 169BZA 15 9 93 20.4 214TPM 250 10 45 29.1 367SLP 12000 40 0.8 430 415IND 31 15 47 163 394ZNS 56 35 nt 259 645CLX 50000 21 nt 34.2 690VLX 54000 43 nt 28.7 5710SLT 374 9 nt 45.2 123SAC 18540 5950 935 40.3 104
The values are from three different assays with errors in the range of 5–10%. Resultsfor AZA, EZ and Valdecoxib (VLX) are shaded in light-grey.
1448 A. Bertucci et al. / Bioorg. Med. Chem. 21 (2013) 1437–1450
several anions that are abundant in seawater, such as chloride(546 mM), sulphate (28 mM) or bromide (0.84 mM), show stronginhibitory effects on STPCA (with ki <1 nM) strongly suggests thatthe ECM, where this enzyme is present, is isolated and/or has adifferent composition from the surrounding seawater. This lackof knowledge of the anionic composition of some important com-partments of Anthozoan, such as the calcifying medium and theperisymbiotic space, might prevent the use of anionic inhibitorsfor the study of the role of CAs in the ECM.
The other class of CA inhibitors tested on coral enzymes is morepromising. While acetazolamide is the most efficient sulfonamidetested so far, it does not discriminate between CA isoforms. How-ever, valdecoxib (VLX in Table 4) has a strong selectivity for STPCA,with a ratio of about 200 times compared to STPCA-2 and couldrepresent a useful pharmacological agent for the targeted studyof this CA isoform in coral. So far, the best ratio in favour ofSTPCA-2 is of about 95 times compared to STPCA and is obtainedwith H2NSO3H. As well as with anions, this isoform is more difficultto target. A deeper pharmacological screening of CA inhibitorsagainst anthozoan isoforms is thus required. New classes of CAinhibitors, such as phenols and polyphenols, coumarins, and poly-amines19 and their derivatives, shall be tested and will undoubt-edly help to identify new selective compounds. Moreover, tocomplete this search, all these chemicals should be tested againstany other coral CA isoform that would be discovered and charac-
terized thanks to the progress in the bio-molecular study of coralphysiology.
Although such studies are still in their infancy, the recentscreening of a large number of CA inhibitor classes against severalcnidarian enzymes may soon lead to physiological in vivo studiesaiming to identify the role of specific isoforms in corals. Comparedto mammals, corals are simpler systems that may allow a directand simple overview on the in vivo data for inhibiting these en-zymes. As corals are highly sensitive to pH changes and sinceCAs are mainly involved in such processes, their inhibition/activa-tion provides an interesting physiologic tool to highlight such pro-cesses in a model organism. In addition, due to the major roleplayed by coral CAs in biomineralization processes, as shown inthis review, the inhibition data recently generated may lead to abetter understanding of how salinity, pollutants and other suchfactors may affect the enzymatic activity of the CA isoforms pres-ent in these organisms. With the increase CO2 emission throughhuman activities, a detailed study of enzymes dealing with thisgas in organisms such as the corals, may help not only basic knowl-edge but probably also finding an environmentally friendly way ofresolving this problem.
7. Conclusion and prospects
Anthozoans provide a fascinating model owing to the originalcharacteristics they harbor and the subsequent need to developnew physiological functions. These processes are at the basis ofthe edification and maintenance of one of the most diverse, pro-ductive and iconic ecosystem on Earth: coral reefs. The first origi-nality is the presence of a CO2-concentrating mechanism that isusually restricted to photoautotrophs, in order to provide highamounts of inorganic carbon to their intracellular symbionts. Inthis process, carbonic anhydrases are involved all along the dis-solved inorganic carbon processing. Likely different isoforms areused in DIC uptake from seawater, cytosolic trapping of DIC/pHregulation, and delivery to Dinoflagellates. The second feature ofmany Anthozoans is their high rate of calcium carbonate deposi-tion in which CAs convert metabolic CO2 into bicarbonate anionsupplied to the extracellular calcifying medium.
Despite the first mention of CAs in corals occurred at the sameperiod as the characterization of mammalian isoforms12,135 theirstudy developed only in the past few decades. This review aimedat showing how the study of CAs in Anthozoa helped to betterunderstand a part of their fundamental physiology. Nevertheless,several gaps remain in this topic as for instance the characteriza-tion of bicarbonate transporters, pH regulation and response toenvironmental conditions. The a-CAs predominate among animalsand are the only CA gene family expressed by vertebrates. Never-theless a major part of the knowledge is based on mammaliana-CAs.136 The early discovery of multiple isoforms in mammals135
paved the way to kinetic studies, inhibition experiments, subcellu-lar localization, and molecular identification leading to the identi-fication of the 16 isoforms known today. In mammalians, many ofthe identified CAs are druggable targets. Over the past few years,the profile of CA inhibitors—anions, sulphonamides, sulphamatesand sulphamides—has been investigated. Similar studies onlystarted recently in Anthozoa. Yet, even in mammalians, only fewcompounds show selectivity for any isoform.18 Which underliesthe need for biochemists and coral biologists to collaborate inthe identification of new isoforms and the use of newly developedchemicals22 for a more accurate study of these enzymes in basalorganisms.
Owing to their evolutionary position as a sister group of Bilate-ria, Cnidarians may give major insights into metazoan evolution.Several studies85,90,105 have revealed the complexity and the

A. Bertucci et al. / Bioorg. Med. Chem. 21 (2013) 1437–1450 1449
vertebrate-likelihood of anthozoan gene repertoire. Genome andtranscriptome sequencing are now more accessible and willundoubtedly lead to new discoveries and a deeper understandingof coral physiology at the cellular and molecular level. This is animportant concern given the increasing diversity and severity ofthreats that coral reefs currently have to face.
Acknowledgments
This research is a part of the Centre Scientifique de Monaco Re-search Program, which is supported by the Government of Princi-pality of Monaco. This research was supported by a Marie CurieInternational Outgoing Fellowship to AM and a fellowship fromthe Centre Scientifique de Monaco to AB. The authors gratefullythank Gergely Torda for the polyp picture in Figure 2 and BenMason for his suggestions and his detailed correction of Englishgrammar.
References and notes
1. Steele, R. E.; David, C. N.; Technau, U. Trends Genet. 2011, 27, 7.2. Turk, T.; Kem, W. R. Toxicon 2009, 54, 1031.3. Edmunds, P. J.; Carpenter, R. C. Proc. Natl. Acad. Sci. U.S.A. 2001, 98, 5067.4. Wilkinson, C. In Status of Corals Reef of the World; Wilkinson, C., Ed.; Australian
Institute of Marine Science: Townsville Australia, 2004; Vol. 1, p 7.5. Douglas, A. E.; McAuley, P. J.; Davies, P. S. J. Zool. London 1993, 231, 175.6. Dubinsky, Z. Ecosystems of the world 25; Elsevier Science: Coral reefs, 1990.7. Muscatine, L.; Falkowski, P. G.; Porter, J. W.; Dubinsky, Z. Proc. R. Soc. B Biol. Sci.
1984, 222, 181.8. Tambutté, S.; Holcomb, M.; Ferrier-Pages, C.; Reynaud, S.; Tambutte, E.;
Zoccola, D.; Allemand, D. J. Exp. Mar. Biol. Ecol. 2011, 408, 58.9. Allemand, D.; Tambutte, E.; Zoccola, D.; Tambutte, S. In Coral Reefs: An
Ecosystem in Transition; Dubinsky, Z., Stambler, N., Eds.; Springer, 2011; p 119.10. Erez, J. Nature 1978, 273, 199.11. Furla, P.; Galgani, I.; Durand, I.; Allemand, D. J. Exp. Biol. 2000, 203, 3445.12. Goreau, T. F. Biol. Bull. 1959, 116, 59.13. Xu, Y.; Feng, L.; Jeffrey, P. D.; Shi, Y.; Morel, F. M. M. Nature 2008, 452, 56.14. Moya, A.; Tambutte, S.; Bertucci, A.; Tambutte, E.; Lotto, S.; Vullo, D.; Supuran,
C.; Allemand, D.; Zoccola, D. J. Biol. Chem. 2008, 283, 25475.15. Elleuche, S.; Poggeler, S. Microbiol.-SGM 2010, 156, 23.16. Supuran, C. T. Curr. Pharm. Des. 2008, 14, 603.17. Supuran, C. T.; Di Fiore, A.; Alterio, V.; Monti, S. M.; De Simone, G. Curr. Pharm.
Des. 2010, 16, 3246.18. Supuran, C. T. Nat. Rev. Drug Disc. 2008, 7, 168.19. Supuran, C. T. Mol. Diversity 2011, 15, 305.20. Pastorekova, S.; Parkkila, S.; Pastorek, J.; Supuran, C. T. J. Enzyme Inhib. Med.
Chem. 2004, 19, 199.21. Badger, M. R.; Price, G. D. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1994, 45, 369.22. Supuran, C. T. Curr. Pharm. Des. 2010, 16, 3233.23. Tashian, R. E.; Hewett-Emmett, D.; Carter, N.; Bergenhem, N. C. EXS 2000, 105.24. Wakefield, T. S.; Kempf, S. C. Biol. Bull. 2001, 200, 127.25. Bertucci, A.; Tambutte, S.; Supuran, C. T.; Allemand, D.; Zoccola, D. Mar.
Biotechnol. (NY) 2011, 13, 992.26. Moya, A.; Huisman, L.; Ball, E. E.; Hayward, D. C.; Grasso, L. C.; Chua, C. M.;
Woo, H. N.; Gattuso, J. P.; Foret, S.; Miller, D. J. Mol. Ecol. 2012, 21, 2440.27. Furla, P.; Richier, S.; Allemand, D. In Coral Reefs: An Ecosystem in Transition;
Dubinsky, Z., Stambler, N., Eds.; Springer: Po Box 17, 3300 Aa Dordrecht,Netherlands, 2011; p 187.
28. Roth, E.; Jeon, K.; Stacey, G. In Molecular Genetic of Plant–Microbe Interactions;Palacios, R. V. D., Ed.; The American Phytopatological Society: St Paul,Minnnesota, 1988; p 220.
29. Stumm, W.; Morgan, J. J. Aquatic Chemistry. An Introduction EmphasizingChemical Equilibria in Natural Waters, 2nd ed.; John Wiley & Sons: New York,1981.
30. Morse, D.; Salois, P.; Markovic, P.; Hastings, J. W. Science 1995, 268, 1622.31. Reinfelder, J. R. Annu. Rev. Mar. Sci. 2011, 3, 291.32. Giordano, M.; Beardall, J.; Raven, J. A. Annu. Rev. Plant Biol. 2005, 56, 99.33. Raven, J. A.; Giordano, M.; Beardall, J.; Maberly, S. C. Photosynth. Res. 2011, 109,
281.34. Badger, M. R.; Kaplan, A.; Berry, J. A. Plant Physiol. 1980, 66, 407.35. Aizawa, K.; Miyachi, S. FEMS Microbiol. Rev. 1986, 39, 215.36. Hayes, R. L.; Goreau, N. I. Biol. Bull. 1977, 152, 26.37. Land, L. S.; Lang, J. C.; Smith, B. N. Limnol. Oceanogr. 1975, 20, 283.38. Allemand, D.; Furla, P.; Bénazet-Tambutté, S. Can. J. Bot. 1998, 76, 925.39. Leggat, W.; Marendy, E. M.; Baillie, B.; Whitney, S. M.; Ludwig, M.; Badgaer, M.
R.; Yellowlees, D. Funct. Plant Biol. 2002, 29, 309.40. Furla, P.; Allemand, D.; Ferrier-Pages, C.; Shick, M. Integr. Comp. Biol. 2005, 45,
595.41. Dove, P. M.; De Yoreo, J. J.; Weiner, S. Biomineralization; Washington:
Mineralogical Society, Reviews in Mineralogy and Geochemistry, 2003.
42. Jackson, D. J.; Macis, L.; Reitner, J.; Degnan, B. M.; Worheide, G. Science 1893,2007, 316.
43. Mitsunaga, K.; Akasaka, K.; Shimada, H.; Fujino, Y.; Yasumasu, I.; Numanoi, H.Cell Differ. 1986, 18, 257.
44. Kakei, M.; Nakahara, H. Biochim. Biophys. Acta, Gen. Subj. 1996, 1289, 226.45. Miyamoto, H.; Miyoshi, F.; Kohno, J. Zool. Sci. 2005, 22, 311.46. Tohse, H.; Murayama, E.; Ohira, T.; Takagi, Y.; Nagasawa, H. Comp. Biochem.
Physiol. B Biochem. Mol. Biol. 2006, 145, 257.47. Taylor, D. L. Endocytobiology 1983, 2, 689.48. Al-Horani, F. A.; Al-Moghrabi, S. M.; de Beer, D. J. Exp. Mar. Biol. Ecol. 2003, 288,
1.49. Venn, A.; Tambutte, E.; Holcomb, M.; Allemand, D.; Tambutte, S. PloS One
2011, 6.50. Zoccola, D.; Tambutté, E.; Kulhanek, E.; Puverel, S.; Scimeca, J. C.; Allemand,
D.; Tambutté, S. Biochim. Biophys. Acta 2004, 1663, 117.51. Truchot, J.-P. Comparative Aspects of Extracellular Acid–Base Balance; Springer-
Verlag, 1987.52. Sikes, C. S.; Wilbur, K. M. J. Phycol. 1980, 16, 433.53. Henry, R. P. Annu. Rev. Physiol. 1996, 58, 523.54. Weis, V. M.; Smith, G. J.; Muscatine, L. Mar. Biol. 1989, 100, 195.55. Graham, D.; Smillie, R. M. Aust. J. Plant Physiol. 1976, 3, 113.56. Kawaguti, S.; Sakumoto, D. Bull. Oceanographical Inst. Taïwan 1948, 65.57. Moya, A.; Tambutté, S.; Tambutté, E.; Zoccola, D.; Caminiti, N.; Allemand, D. J.
Exp. Biol. 2006, 209, 3413.58. Ferrier, J. M. Plant Physiol. 1980, 66, 1198.59. Weis, V. M. J. Exp. Mar. Biol. Ecol. 1993, 174, 209.60. Al-Moghrabi, S.; Goiran, C.; Allemand, D.; Speziale, N.; Jaubert, J. J. Exp. Mar.
Biol. Ecol. 1996, 199, 227.61. Bénazet-Tambutté, S.; Allemand, D.; Jaubert, J. Symbiosis 1996, 20, 199.62. Weis, V. M. Biol. Bull. 1991, 180, 496.63. Palmqvist, K.; Sjöberg, S.; Samuelsson, G. Plant Physiol. 1988, 87, 437.64. Furla, P.; Bénazet-Tambutté, S.; Jaubert, J.; Allemand, D. Am. J. Physiol. 1998,
274, R303.65. Furla, P.; Allemand, D.; Orsenigo, M. N. Am. J. Physiol. 2000, 278, R870.66. Tsuruoka, S.; Schwartz, G. J. Am. J. Physiol. 1998, 43, F139.67. Venn, A.; Tambutte, E.; Lotto, S.; Zoccola, D.; Allemand, D.; Tambutte, S. Proc.
Nat. Acad. Sci. U.S.A. 2009, 106, 16574.68. Lapointe, M.; MacKenzie, T. D. B.; Morse, D. Plant Physiol. 2008, 147, 1427.69. Baillie, B. K.; Yellowlees, D. Proc. R. Soc. London 1998, 265, 465.70. Leggat, W.; Badger, M. R.; Yellowlees, D. Plant Physiol. 1999, 121, 1247.71. Goiran, C.; Al-Moghrabi, S.; Allemand, D.; Jaubert, J. J. J. Exp. Mar. Biol. Ecol.
1996, 199, 207.72. Bertucci, A.; Tambutte, E.; Tambutte, S.; Allemand, D.; Zoccola, D. Proc. R. Soc.
B-Biol. Sci. 2010, 277, 87.73. Isa, Y.; Yamazato, K. Galaxea 1984, 3, 25.74. Tambutté, E.; Allemand, D.; Mueller, E.; Jaubert, J. J. Exp. Biol. 1996, 199, 1029.75. Al-Horani, F. A.; Al-Moghrabi, S. M.; de Beer, D. Mar. Biol. 2003, 142, 419.76. Marshall, A. T.; Clode, P. L. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2003,
136, 417.77. Romano, S. L.; Palumbi, S. R. J. Mol. Evol. 1997, 45, 397.78. Kingsley, R. J.; Watabe, N. J. Exp. Zool. 1987, 241, 171.79. Lucas, J. M.; Knapp, L. W. Mar. Biol. 1996, 126, 471.80. Rahman, M. A.; Isa, Y. J. Exp. Mar. Biol. Ecol. 2005, 321, 71.81. Rahman, M. A.; Isa, Y.; Takemura, A.; Uehara, T. Calcif. Tissue Int. 2006, 78, 178.82. Rahman, M. A.; Isa, Y.; Uehara, T. Mar. Biotechnol. (NY) 2006, 8, 415.83. Rahman, M. A.; Oomori, T.; Uehara, T. Mar. Biotechnol. (NY) 2008, 10, 31.84. Tambutté, S.; Tambutté, E.; Zoccola, D.; Caminiti, N.; Lotto, S.; Moya, S.;
Allemand, D.; Adkins, J. Mar. Biol. 2007, 151, 71.85. Technau, U.; Rudd, S.; Maxwell, P.; Gordon, P. M. K.; Saina, M.; Grasso, L. C.;
Hayward, D. C.; Sensen, C. W.; Saint, R.; Holstein, T. W.; Ball, E. E.; Miller, D. J.Trends Genet. 2005, 21, 633.
86. Supuran, C. T.; Scozzafava, A. Bioorg. Med. Chem. 2007, 15, 4336.87. Fasseas, M. K.; Tsikou, D.; Flemetakis, E.; Katinakis, P. Mol. Biol. Rep. 2011, 38,
1777.88. Weis, V. M.; Reynolds, W. S. Physiol. Biochem. Zool. 1999, 72, 307.89. Sabourault, C.; Ganot, P.; Deleury, E.; Allemand, D.; Furla, P. BMC Genomics
2009, 10, 333.90. Kortschak, R. D.; Samuel, G.; Saint, R.; Miller, D. J. Curr. Biol. 2003, 13, 2190.91. Meyer, E.; Aglyamova, G. V.; Wang, S.; Buchanan-Carter, J.; Abrego, D.;
Colbourne, J. K.; Willis, B. L.; Matz, M. V. BMC Genomics 2009, 10, 1.92. Traylor-Knowles, N.; Granger, B. R.; Lubinski, T. J.; Parikh, J. R.; Garamszegi, S.;
Xia, Y.; Marto, J. A.; Kaufman, L.; Finnerty, J. R. BMC Genomics 2011, 12, 585.93. Hewett-Emmett, D.; Tashian, R. E. Mol. Phylogenet. Evol. 1996, 5, 50.94. McDevitt, M. E.; Lambert, L. A. Biochim. Biophys. Acta 1854, 2011, 1814.95. Stanley, G. D., Jr.; Fautin, D. G. Science 1913, 2001, 291.96. DeBoer, M. L.; Krupp, D. A.; Weis, V. M. Biol. Bull. 2006, 211, 18.97. Grasso, L. C.; Maindonald, J.; Rudd, S.; Hayward, D. C.; Saint, R.; Miller, D. J.;
Ball, E. E. BMC Genomics 2008, 9, 540.98. Shinzato, C.; Shoguchi, E.; Kawashima, T.; Hamada, M.; Hisata, K.; Tanaka, M.;
Fujie, M.; Fujiwara, M.; Koyanagi, R.; Ikuta, T.; Fujiyama, A.; Miller, D. J.; Satoh,N. Nature 2011, 476, 320.
99. Ganot, P.; Moya, A.; Magnone, V.; Allemand, D.; Furla, P.; Sabourault, C. PLoSGenet. 2011, 7.
100. Aspatwar, A.; Tolvanen, M. E.; Parkkila, S. BMC Mol. Biol. 2010, 11, 25.101. Okamoto, N.; Fujikawa-Adachi, K.; Nishimori, I.; Taniuchi, K.; Onishi, S.
Biochim. Biophys. Acta 2001, 1518, 311.

1450 A. Bertucci et al. / Bioorg. Med. Chem. 21 (2013) 1437–1450
102. Taniuchi, K.; Nishimori, I.; Takeuchi, T.; Fujikawa-Adachi, K.; Ohtsuki, Y.;Onishi, S. Neuroscience 2002, 112, 93.
103. Hirota, J.; Ando, H.; Hamada, K.; Mikoshiba, K. Biochem. J. 2003, 372, 435.104. Miyamoto, H. Biochem. Genet. 2012, 50, 269.105. Putnam, N. H.; Srivastava, M.; Hellsten, U.; Dirks, B.; Chapman, J.; Salamov, A.;
Terry, A.; Shapiro, H.; Lindquist, E.; Kapitonov, V. V.; Jurka, J.; Genikhovich, G.;Grigoriev, I. V.; Lucas, S. M.; Steele, R. E.; Finnerty, J. R.; Technau, U.;Martindale, M. Q.; Rokhsar, D. S. Science 2007, 317, 86.
106. Pandolfi, J. M.; Connolly, S. R.; Marshall, D. J.; Cohen, A. L. Science 2011, 333,418.
107. De’ath, G.; Lough, J. M.; Fabricius, K. E. Science 2009, 323, 116.108. Hoegh-Guldberg, O.; Mumby, P. J.; Hooten, A. J.; Steneck, R. S.; Greenfield, P.;
Gomez, E.; Harvell, C. D.; Sale, P. F.; Edwards, A. J.; Caldeira, K.; Knowlton, N.;Eakin, C. M.; Iglesias-Prieto, R.; Muthiga, N.; Bradbury, R. H.; Dubi, A.;Hatziolos, M. E. Science 2007, 318, 1737.
109. Edge, S. E.; Morgan, M. B.; Gleason, D. F.; Snell, T. W. Mar. Pollut. Bull. 2005, 51,507.
110. Meyer, E.; Aglyamova, G. V.; Matz, M. V. Mol. Ecol. 2011, 20, 3599.111. Moya, A.; Ganot, P.; Furla, P.; Sabourault, C. Mol. Ecol. 2012, 21, 1158.112. Caldeira, K.; Wickett, M. E. Nature 2003, 425, 365.113. Langdon, C.; Atkinson, M. J. Geophys. Res.-Oceans 2005, 110.114. Orr, J. C.; Fabry, V. J.; Aumont, O.; Bopp, L.; Doney, S. C.; Feely, R. A.;
Gnanadesikan, A.; Gruber, N.; Ishida, A.; Joos, F.; Key, R. M.; Lindsay, K.;Maier-Reimer, E.; Matear, R.; Monfray, P.; Mouchet, A.; Najjar, R. G.; Plattner,G. K.; Rodgers, K. B.; Sabine, C. L.; Sarmiento, J. L.; Schlitzer, R.; Slater, R. D.;Totterdell, I. J.; Weirig, M. F.; Yamanaka, Y.; Yool, A. Nature 2005, 437, 681.
115. Kleypas, J. A.; Yates, K. K. Oceanography 2009, 22, 108.116. Houlbreque, F.; Rodolfo-Metalpa, R.; Jeffree, R.; Oberhansli, F.; Teyssie, J. L.;
Boisson, F.; Al-Trabeen, K.; Ferrier-Pages, C. Coral Reefs 2012, 31, 101.117. Reynaud, S.; Leclercq, N.; Romaine-Lioud, S.; Ferrier-Pages, C.; Jaubert, J.;
Gattuso, J. P. Global Change Biol. 2003, 9, 1660.118. Kaniewska, P.; Campbell, P. R.; Kline, D. I.; Rodriguez-Lanetty, M.; Miller, D. J.;
Dove, S.; Hoegh-Guldberg, O. PloS One 2012, 7, e34659.119. IPCC. In Contribution of Working Group I to the Fourth Assessment Report of
the Intergovernmental Panel on Climate Change; Solomon S, e. a., Ed.;Cambridge Univ Press: New York, 2007.
120. Vidal-Dupiol, J.; Zoccola, D.; Tambutté, E.; Grunau, C.; Cosseau, C.; Smith, K.M.; Freitag, M.; Adjeroud, M.; Dheilly, N. M.; Allemand, D.; Mitta, G.;Tambutté, S. Unpublished results.
121. Dietz, T. H.; Wilcox, S. J.; Byrne, R. A.; Silverman, H. Can. J. Zool. -Rev. Can. Zool.1997, 75, 697.
122. Whiteley, N. M.; Scott, J. L.; Breeze, S. J.; McCann, L. J. Exp. Biol. 2001, 204,1003.
123. Wilson, J. M.; Whiteley, N. M.; Randall, D. J. Physiol. Biochem. Zool. 2002, 75,237.
124. Gilbert, A. L.; Guzman, H. M. Mar. Poll. Bull. 2001, 42, 742.125. Bielmyer, G. K.; Grosell, M.; Bhagooli, R.; Baker, A. C.; Langdon, C.; Gillette, P.;
Capo, T. R. Aquat. Toxicol. 2010, 97, 125.126. Bertucci, A.; Innocenti, A.; Scozzafava, A.; Tambutte, S.; Zoccola, D.; Supuran,
C. T. Bioorg. Med. Chem. Lett. 2011, 21, 710.127. Supuran, C. T.; Scozzafava, A. Carbonic Anhydrase – Its Inhibitors and Activators;
CRC Press: Boca Raton, FL, 2004.128. Innocenti, A.; Hall, R. A.; Schlicker, C.; Scozzafava, A.; Steegborn, C.;
Muhlschlegel, F. A.; Supuran, C. T. Bioorg. Med. Chem. 2009, 17, 4503.129. Bertucci, A.; Innocenti, A.; Zoccola, D.; Scozzafava, A.; Allemand, D.; Tambutte,
S.; Supuran, C. T. Bioorg. Med. Chem. Lett. 2009, 19, 650.130. Bertucci, A.; Innocenti, A.; Zoccola, D.; Scozzafava, A.; Tambutte, S.; Supuran,
C. T. Bioorg. Med. Chem. 2009, 17, 5054.131. Tambutté, E.; Tambutté, S.; Segonds, N.; Zoccola, D.; Venn, A.; Erez, J.;
Allemand, D. Proc. R. Soc. B-Biol. Sci. 2012, 279, 19.132. Blamart, D.; Rollion-Bard, C.; Meibom, A.; Cuif, J. P.; Juillet-Leclerc, A.;
Dauphin, Y. Geochem. Geophy. Geosy. 2007, 8.133. Reynaud, S.; Hemming, N. G.; Juillet-Leclerc, A.; Gattuso, J. P. Coral Reefs 2004,
23, 539.134. Holcomb, M.; Cohen, A. L.; Gabitov, R. I.; Hutter, J. L. Geochim. Cosmochim. Acta
2009, 73, 4166.135. Nyman, P. O. Biochim. Biophys. Acta 1961, 52, 1.136. Gilmour, K. M. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2010, 157, 193.137. Guindon, S.; Dufayard, J. F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O.
Syst. Biol. 2010, 59, 307.138. Abascal, F.; Zardoya, R.; Posada, D. Bioinformatics 2005, 21, 2104.139. Bertucci, A.; Zoccola, D.; Tambutte, S.; Vullo, D.; Supuran, C. T. Bioorg. Med.
Chem. 2010, 18, 2300.











![MANGROVES. SEAGRASSES AND CORALS [A simple layman-type field guide]](https://static.fdokumen.com/doc/165x107/6321610a0c12e1161503c4a8/mangroves-seagrasses-and-corals-a-simple-layman-type-field-guide.jpg)








