
A new coral carbonic anhydrase in Stylophora pistillata
11
ORIGINAL ARTICLE A New Coral Carbonic Anhydrase in Stylophora pistillata Anthony Bertucci & Sylvie Tambutté & Claudiu T. Supuran & Denis Allemand & Didier Zoccola Received: 13 July 2010 / Accepted: 10 January 2011 / Published online: 12 February 2011 # Springer Science+Business Media, LLC 2011 Abstract Scleractinian corals are of particular interest due to their ability to establish an intracellular mutualistic symbiosis with phototrophic dinoflagellates and to deposit high rates of calcium carbonate in their skeleton. Carbonic anhydrases have been shown to play a crucial role in both processes. In this study, we report the molecular cloning and characterization of a novel α-CA in the coral Stylophora pistillata. This enzyme shares homologies with the human isoform CA II and is referred to as STPCA-2. STPCA-2 is 35.2 kDa and possesses all key amino acids for catalytic activity. With a ratio between catalytic and Michaelis constants (k cat /K m ) of 8.3.10 7 M -1 s -1 is considered as highly active. Owing to its intracellular localisation in the oral endoderm and in the aboral tissue, we propose that STPCA-2 is involved in pH regulation and/or inorganic carbon delivery to symbiont and calcification. Keywords Scleractinian corals . Carbonic anhydrase . Symbiosis . Calcification . Intracelullar pH Introduction Carbonic anhydrases (CA, EC 4.2.1.1) are ubiquitous metalloenzymes that catalyse the reversible hydration of carbon dioxide into bicarbonate (CO 2 +H 2 O ⇆ HCO 3 - +H + ). The CA family is polyphyletic, with at least five classes: α, β, γ, ∂ and ζ. They are involved in numerous physiological processes from invertebrates to vertebrates like, for exam- ple, respiration with CO 2 /HCO 3 - transport and exchange between tissues and lungs, pH regulation, electrolyte secretion, calcification and also pathologic processes (for a review, see Supuran 2008 and Xu et al. 2008). Scleractinian corals have been of particular interest for research over the past two decades owing to their fundamental role in reef formation and maintenance of this highly diverse ecosystem in oligotrophic tropical regions. The ecological success of corals comes from two major features: the symbiotic association that many of them establish with photosynthetic dinoflagellates from the genus Symbiodinium (commonly named zooxanthellae), and the precipitation of an aragonitic calcium carbonate (CaCO 3 ) skeleton. Both these processes involve carbonic anhydrases. In seawater, where pH is about 8.2, most dissolved inorganic carbon (DIC) is under the form of bicarbonate (HCO 3 - ). Moreover, zooxanthellae are enclosed in a host- derived vacuole inside the endodermal cell layer. Thus Symbiodinium, as a broad array of marine algae, has developed CO 2 -concentrating mechanisms (CCM) in which CAs play a crucial role (for review see Giordano et al. 2005) to increase the CO 2 availability for DIC transport and/or photosynthesis. Thus, CCM allow to maintain high rate of carbon fixation. Studies on this role of CAs in DIC uptake for photosynthesis in symbiotic Cnidarians have mainly been performed in non-calcifying organisms such as the sea anemones Aiptasia pulchella (Weis 1993), Antho- pleura elegantissima (Weis and Reynolds 1999) or Anemo- nia viridis (Furla et al. 2000a). In reef-building corals, CCM also exist (Al-Moghrabi et al. 1996; Leggat et al. 1999; Furla et al. 2000a; Leggat et al. 2002), but CAs are particularly studied for their involvement in biomineraliza- A. Bertucci : S. Tambutté : D. Allemand : D. Zoccola (*) Centre Scientifique de Monaco, Avenue Saint Martin, 98000 Monaco, Monaco e-mail: [email protected] C. T. Supuran Dipartimento di Chimica, University of Florence, via della Lastruccia, 3, Rm. 188, Polo Scientifico, 50019 Sesto Fiorentino (Florence), Italy Mar Biotechnol (2011) 13:992–1002 DOI 10.1007/s10126-011-9363-x
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of A new coral carbonic anhydrase in Stylophora pistillata

ORIGINAL ARTICLE
A New Coral Carbonic Anhydrase in Stylophora pistillata
Anthony Bertucci & Sylvie Tambutté &
Claudiu T. Supuran & Denis Allemand & Didier Zoccola
Received: 13 July 2010 /Accepted: 10 January 2011 /Published online: 12 February 2011# Springer Science+Business Media, LLC 2011
Abstract Scleractinian corals are of particular interest due totheir ability to establish an intracellular mutualistic symbiosiswith phototrophic dinoflagellates and to deposit high rates ofcalcium carbonate in their skeleton. Carbonic anhydrases havebeen shown to play a crucial role in both processes. In thisstudy, we report the molecular cloning and characterization ofa novel α-CA in the coral Stylophora pistillata. This enzymeshares homologies with the human isoform CA II and isreferred to as STPCA-2. STPCA-2 is 35.2 kDa and possessesall key amino acids for catalytic activity. With a ratio betweencatalytic and Michaelis constants (kcat/Km) of 8.3.10
7 M−1s−1
is considered as highly active. Owing to its intracellularlocalisation in the oral endoderm and in the aboral tissue, wepropose that STPCA-2 is involved in pH regulation and/orinorganic carbon delivery to symbiont and calcification.
Keywords Scleractinian corals . Carbonic anhydrase .
Symbiosis . Calcification . Intracelullar pH
Introduction
Carbonic anhydrases (CA, EC 4.2.1.1) are ubiquitousmetalloenzymes that catalyse the reversible hydration ofcarbon dioxide into bicarbonate (CO2+H2O⇆HCO3
−+H+).
The CA family is polyphyletic, with at least five classes: α,β, γ, ∂ and ζ. They are involved in numerous physiologicalprocesses from invertebrates to vertebrates like, for exam-ple, respiration with CO2/HCO3
− transport and exchangebetween tissues and lungs, pH regulation, electrolytesecretion, calcification and also pathologic processes (fora review, see Supuran 2008 and Xu et al. 2008).
Scleractinian corals have been of particular interest forresearch over the past two decades owing to their fundamentalrole in reef formation and maintenance of this highly diverseecosystem in oligotrophic tropical regions. The ecologicalsuccess of corals comes from two major features: thesymbiotic association that many of them establish withphotosynthetic dinoflagellates from the genus Symbiodinium(commonly named zooxanthellae), and the precipitation ofan aragonitic calcium carbonate (CaCO3) skeleton. Boththese processes involve carbonic anhydrases.
In seawater, where pH is about 8.2, most dissolvedinorganic carbon (DIC) is under the form of bicarbonate(HCO3
−). Moreover, zooxanthellae are enclosed in a host-derived vacuole inside the endodermal cell layer. ThusSymbiodinium, as a broad array of marine algae, hasdeveloped CO2-concentrating mechanisms (CCM) in whichCAs play a crucial role (for review see Giordano et al.2005) to increase the CO2 availability for DIC transportand/or photosynthesis. Thus, CCM allow to maintain highrate of carbon fixation. Studies on this role of CAs in DICuptake for photosynthesis in symbiotic Cnidarians havemainly been performed in non-calcifying organisms such asthe sea anemones Aiptasia pulchella (Weis 1993), Antho-pleura elegantissima (Weis and Reynolds 1999) or Anemo-nia viridis (Furla et al. 2000a). In reef-building corals,CCM also exist (Al-Moghrabi et al. 1996; Leggat et al.1999; Furla et al. 2000a; Leggat et al. 2002), but CAs areparticularly studied for their involvement in biomineraliza-
A. Bertucci : S. Tambutté :D. Allemand :D. Zoccola (*)Centre Scientifique de Monaco,Avenue Saint Martin,98000 Monaco, Monacoe-mail: [email protected]
C. T. SupuranDipartimento di Chimica, University of Florence,via della Lastruccia, 3, Rm. 188, Polo Scientifico,50019 Sesto Fiorentino (Florence), Italy
Mar Biotechnol (2011) 13:992–1002DOI 10.1007/s10126-011-9363-x

tion since Goreau (1959) showed the reduction in calcifi-cation rates by CA-specific inhibitors. In these lastorganisms, CAs convert metabolic CO2 produced bycalcifying cells into HCO3
− that will be used for CaCO3
deposition (Isa and Yamazato 1984; Kingsley and Watabe1987; Tambutté et al. 1996; Furla et al. 2000b; Al-Horani etal. 2003b; Marshall and Clode 2003; Tambutté et al. 2007b;Moya et al. 2008b). CAs are also found in several othercalcifying cnidarians such as gorgonians (Lucas and Knapp1997) and alcyonarians (Rahman et al. 2008).
In the literature, only five α-CAs have beendescribed in Scleractinian corals. Two atypicalmembrane-bound isozymes in Fungia scutaria, FCA-aand -b, that could play a role in the onset of calcificationin larvae (deBoer et al. 2006), a secreted one in Stylophorapistillata, STPCA, which plays a key role in biomineral-ization (Moya et al. 2008b; Bertucci et al. 2009a, b;Bertucci et al. 2010a; Bertucci et al. b) and two genesfound in Acropora millepora: A030-E11, expressed in thesepta where calcification is occurring to form adultstructures, and C007-E7, involved in the onset ofcalcification (Grasso et al. 2008). Here, we report thecloning, sequencing and characterization of a new α-CAin the Scleractinian coral Stylophora pistillata. Thisprotein shares similarities with the human cytoplasmicisoform II, which is known as the most efficient CAisozyme with a kcat=1,4.10
6 s−1 and one of the most activeenzyme with a kcat/Km=1,5.10
8 M−1s−1. This isoform isusually involved in intracellular pH regulation in differenthuman cell types (for a review, see Supuran 2008). As thisenzyme is localised intracellularly, we propose that thisnew coral CA, called STPCA-2, is likely to have the samerole in coral cells.
Materials and Methods
Biological Material Experiments were conducted in thelaboratory on the branching symbiotic Scleractinian coralStylophora pistillata (Esper 1797). Colonies were stored ina 300-L aquarium, supplied with seawater from theMediterranean Sea (exchange rate 2% per hour) undercontrolled conditions: semi-open circuit, temperature of26.0±0.2°C, salinity of 38.2 psu, light of 175 μmol photonsper square metre per second (using fluorescent tubesCustom Sea Life®) on a 12:12 photoperiod. Corals werefed three times weekly with a mix of Artemia salinanauplii, frozen adults of Artemia salina, and frozen krill.
RNA Extraction, cDNA Synthesis, PCR Amplification,Cloning and Sequencing Total RNAs extraction and cDNAsynthesis were performed as described previously (Moya etal. 2008a). Polymerase chain reaction (PCR) amplification
was carried out using Platinum Taq DNA polymerase(Invitrogen) and degenerate primers: forward (5′-CAGTTNCAYTTYCAYTGGGG-3′) designed on the con-served Q(L/F)HFHWG region, and reverse (5′-ACCRASCACWGCCARGCC-3′) designed on the conservedPDGLAVLG region. PCR amplification product was ligatedinto a plasmid vector (pGEM-T easy vector system II,Promega). DNA sequencing of three independent clones werecarried out on both strands with SP6 and T7 primer sequencesbyMacrogen Inc. To obtain the 5′ and 3′ ends of the sequence,rapid amplification of cDNA ends (RACE) experiments wereperformed using the 5′/3′ RACE Kit by Roche. Both endswere cloned in pGEM-T easy vector and sequenced asmentioned above. To know which partner of the symbiosisdoes this CA belong to, we performed a present/absent PCRassay on both cDNA and gDNA from either holobiont (coral+zooxanthellae) or cultured zooxanthellae using the followingprimers: 5′-GGCCACTTGCCCAGGAAACAGGG-3′ and5′-CGTCTACGGCTGTCACATTGGC-3′, respectively for-ward and reverse. Control genes are 36B4 (Moya et al.2008a) and RuBisCO, respectively for the animal and algalgene expression (for primers sequences, see Bertucci et al.2010a).
Sequence and Phylogenetic Analyses BLASTX analyseswere conducted on the NCBI server (Altschul et al. 1997)on GenBank database. Signal peptide presence wasdetermined using the server SignalP-3 (Emanuelsson et al.2007). Phosphorylation and glycosylation prediction anal-yses were performed on the Center for Biological SequenceAnalysis prediction server (Blom et al. 1999). Glycosyl-phosphatidylinositol (GPI) modification site prediction wasperformed using both the big-PI predictor (Eisenhaber et al.1999) and DGPI servers.
Phylogenetic trees were constructed with both themaximum likelihood (ML) and Bayesian methods in orderto check for concordance of the results. The alignments ofall amino acid sequences were performed with the Multalinserver (Corpet 1988). Based upon the amino acid align-ment, ML estimates of the topology and branch length wereobtained using PhyML v3.0 (Guindon and Gascuel 2003)with the WAG+I+G model of substitution as recommendedby alignment analysis with ProtTest v2.0 (Abascal et al.2005). Support for individual branches was inferred bybootstrap analyses (500 replicates). Further phylogeneticrelationships were investigated using Bayesian techniquesas implemented in the computer programme MrBayesv3.1.2 (Huelsenbeck and Ronquist 2001) starting from arandom tree, using the WAG+I+G model generating850,000 generations with sampling every 100 generations,and with four chains in order to obtain the final (consensus)tree and to determine the posterior probabilities at thedifferent nodes.
Mar Biotechnol (2011) 13:992–1002 993

Expression in CHO Cells and Protein Purification PCRproducts were prepared from oligo-dT reverse transcriptioncoral cDNAs and cloned into pIRES-DSRED mammalianexpression vector (Clontech, Inc.). PCR reaction wascarried out with the following primers: forward: (5′-CTAGCTAGCTAGACCATGTATTTTCTGTGGCTAATTTC-3′) and reverse (5′-TCCCCCGGGGGATCAGTGATGGTGATGGTGATGCGATCCTCTTTCCCGGCTAGAAAGGACAGAG-3′). The recombinant vector STPCA-2ORFHIS contains a chimeric sequence encoding the ORFof the STPCA-2 fused to a polyhistidine region at the C-terminus. After sequencing, the plasmid clone was intro-duced into Chinese hamster ovary (CHO) cells with theLipofectamine transfection reagent (Invitrogen). A positiveclone, obtained by limiting dilution, was used to produceSTPCA-2. When the CHO-STPCA-2 cells were 70–80%confluent, the culture medium was harvested 7 days afterand deposited onto Ni-NTA agarose beads (Qiagen) towhich the 6×His-Tag of the recombinant protein wouldbind. The beads were washed with washing buffer (20 mMimidazole, 300 mM NaCI, 50 mM NaH2PO4 and 0,05%Tween 20, pH 8.0), and the 6×His-Tagged protein waseluted with the elution buffer (200 mM imidazole, 300 mMNaCI, 50 mM NaH2P04 and 0,05% Tween 20, pH 8.0).Imidazole was removed by concentration of eluted proteinusing a Centricon Plus centrifugal filter unit (5,000 MWcutoff) before a washing step with phosphate-bufferedsaline (PBS). At the end, the retentate was collected, andprotein concentration was measured using a Qubit kit assay(Invitrogen).
Western Blotting Purified proteins were subjected to 12%SDS-PAGE and electroblotted onto a PVDF membrane.Blots were probed with the monoclonal mouse anti-Histidine Tag (Serotec; 1:1,000 dilution) and the STPCA-2 polyclonal antibody raised against the whole protein inrabbit (Eurogentec; 1:1,000 dilution). STPCA-2 was alsotreated overnight by N-glycosidase F (Roche AppliedScience) in 250 mM Tris, pH 8.0, 50 mM 2-mercaptoethanol at 37°C.
Immunolocalisation with Anti-STPCA-2 Apexes of colo-nies were prepared for immunolocalisation as describedpreviously by Puverel et al. 2005. Briefly, apexes of S.pistillata were fixed in 3% paraformaldehyde in S22buffer (450 mmol L−1 NaCl, 10 mmol L−1 KCl,58 mmol L− 1 MgCl2 , 10 mmol L− 1 CaCl2 ,100 mmol L−1 Hepes, pH 7.8) at 4°C overnight and thendecalcified using 0.5 mol L−1 ethylenediaminetetraaceticacid in Ca-free S22 at 4°C. They were then dehydrated inan ethanol series and embedded in Paraplast. Cross-sections (6-μm thick) were cut and mounted on silane-coated glass slides.
Immunolocalisation was performed according to theprotocol previously described by Puverel et al. (2005). Insummary, deparaffinized sections of tissues were incubatedfor 1 h in saturating medium [1% BSA, 0.2% teleosteangelatin, 0.05% Tween 20 in 0.05 mol L−1 PBS pH 7.4] atroom temperature (RT). The samples were then incubatedwith the anti-STPCA-2 or the pre-immune serum asprimary antibodies (1:100 dilution, 1 h at RT and overnightat 4°C in moist chambers). After rinsing in saturatingmedium, the samples were incubated with biotinylated anti-rabbit antibodies (Amersham 1:250 dilution, 1 h at RT) assecondary antibodies. After rinsing with PBS pH 7.4, thesamples were finally stained for 15 min with streptavidinAlexa Fluor 568 (Molecular Probes, 1:50 dilution) and 4′,6-diamidino-2-phenylindole, DAPI (Sigma, 2 μg mL−1).Samples were embedded in ProLong Antifade medium(Molecular Probes) and observed with a confocal laserscanning microscope (Leica, SP5).
CA Activity and Inhibition Assay An Applied Photophysicsstopped flow instrument has been used for assaying theCA-catalysed CO2 hydration activity (Khalifah 1971).Phenol red (at a concentration of 0.2 mM) has been usedas indicator, working at the maximum absorbance of557 nm, with 10 mM Hepes (pH 7.5) or Tris (pH 8.3) asbuffer, and 20 mM Na2SO4 or 20 mM NaCl (formaintaining constant the ionic strength), following theCA-catalysed CO2 hydration reaction for a period of 10–100 s. The CO2 concentrations ranged from 1.7 to 17 mMfor the determination of the kinetic parameters andinhibition constants. For the inhibitor at least six traces ofthe initial 5–10% of the reaction have been used fordetermining the initial velocity. The uncatalysed rates weredetermined in the same manner and subtracted from thetotal observed rates. Stock solutions of inhibitor (1 mM)were prepared in distilled–de-ionised water with 10–20%(v/v) DMSO (which is not inhibitory at these concentra-tions) and dilutions up to 0.01 nM were done thereafterwith distilled–de-ionised water. Inhibitor and enzymesolutions were preincubated together for 15 min at roomtemperature prior to assay, in order to allow for theformation of the enzyme–inhibitor complex. The inhibitionconstants were obtained by non-linear least squares meth-ods using PRISM 3, from Lineweaver–Burk plots, asreported earlier, and represent the mean from at least threedifferent determinations.
Results
Cloning and Sequencing The complete sequence is 1,417-bp long with a 951 nucleotides open reading frame
994 Mar Biotechnol (2011) 13:992–1002

beginning 87 nucleotides downstream from the 5′ end. Thissequence encodes a protein of 316 amino acids with acalculated molecular mass of about 35.2 kDa. Thenucleotide sequence is available on Genbank™ databaseby accession number EU532164. BLAST searches ofGenBank give best matches mainly with vertebrate cyto-solic isoforms CA II.
Presence/Absence PCR Assays To ensure wether our geneis of host or symbiont origin, we performed PCRamplification of a fragment of our gene both on genomicand complementary DNA using RuBisCO as a vegetalcontrol gene and 36B4 (Moya et al. 2008a) as an animalcontrol gene in holobiont (coral and symbiont) and culturedzooxanthellae (CZ). Figure 1 shows that the holobiontpossesses and expresses all the genes cited above. In CZ,only RuBisCO is present in gDNA and cDNA. Ourcarbonic anhydrase gene, as well as the animal controlgene, is not present in either the CZ genome or tran-scriptome. This assay proves that this CA belongs to theanimal partner and was named STPCA-2 for “STylophoraPistillata Carbonic Anhydrase isoform 2”. The gDNAfragments of STPCA-2 and 36B4 contain introns. Thesesequences are available on GenBank (accession numberHM748807 and HM748806).
Sequence Analysis A ClustalW alignment of STPCA-2 withhuman isoforms CA I, CA II, CA VI and coral isoformsFCA-a, FCA-b, A030-E11, C007-E7 and STPCA, respec-tively from F. scutaria, Acropora millepora and Stylophorapistillata is presented in Fig. 2. It should be noted thatSTPCA-2 appears very different from STPCA. Indeed theyare 35% identical and 48% homologous in nucleotide
sequence. All the 36 residues forming the active site(Hewett-Emmett and Tashian 1996) are present inSTPCA-2. A conserved domain search allowed us toidentify His113, His115 and His138 as the three residuesinvolved in zinc cofactor binding. This analysis alsoidentifies the other 33 amino acid residues that areimportant for enzymatic activity among which four are ofparticular relevance. The zinc-bound water molecule estab-lishes hydrogen bond interactions with the hydroxyl moietyof Thr252, which in turn is bridged to the carboxylatemoiety of Glu125. These interactions, called gatekeeper,enhance the nucleophilicity of the zinc-bound watermolecule and orient the substrate (CO2) in a favourablelocation for the nucleophilic attack. In order to regeneratethe active form of the enzyme, a proton transfer reactionfrom the active site to the environment takes place, which isassisted by the proton shuttle residue His82. Gln111 is alsoidentified as an important residue in the active site, thisresidue, corresponding to Gln92 in hCAII is involved in thebinding of many sulfonamide/sulfamate inhibitors. Thisprotein contains 16 putative phosphorylation sites (tenserines, one threonine and five tyrosine) and two N-glycosylation sites in positions 31 and 194. Prediction toolsindicate that the first 15 amino acids are a signal peptide.This feature seems to be in contradiction with thehomologies between STPCA-2 and the cytosolic α-CA II.These tools did not identify any GPI anchor or transmem-brane domains. One can also notice the presence of a largeinsert of 30 amino acids expanding between positions 212and 243.
Biochemical Characterization A Western blotting of therecombinant STPCA-2 labelled with a 6×His-Tag per-formed either with an anti-Histidine Tag or an anti-STPCA-2 specific antibody shows an apparent weight ofabout 45 kDa suggesting post-translational modifications.An N-glycosidase F treatment lowers this weight around35.2 kDa as predicted by the putative amino acids sequence(Fig. 3).
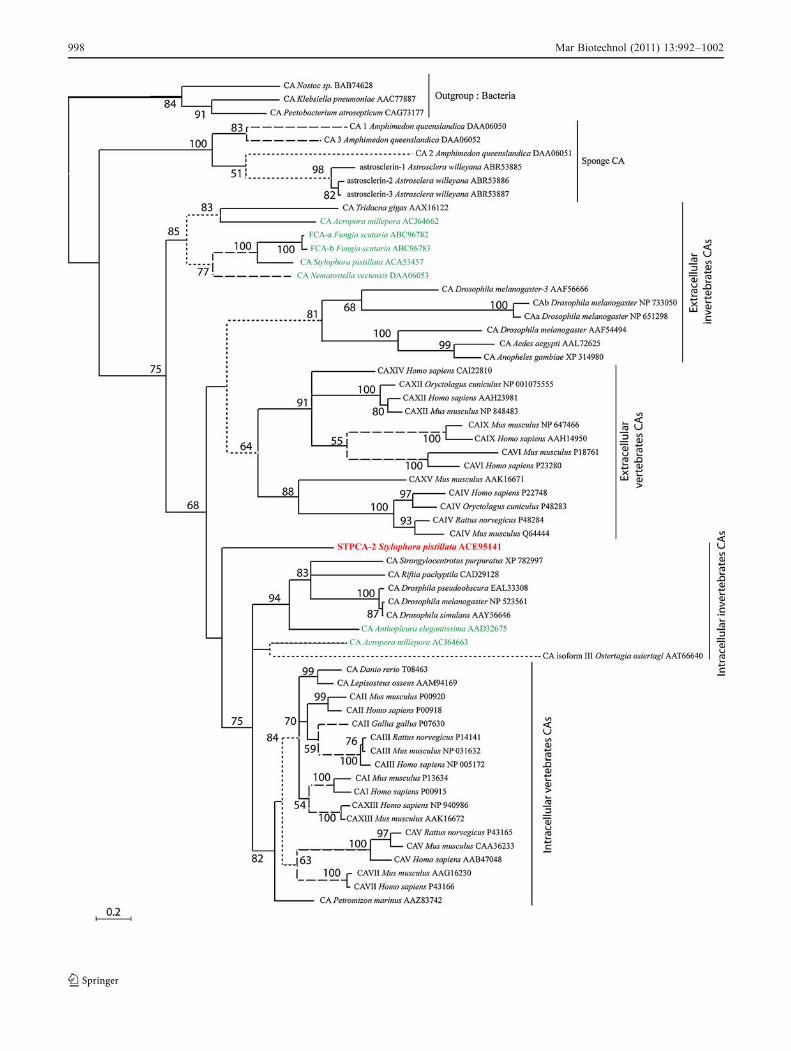
Phylogenetic Analyses To further characterise our se-quence, we compared it with 60 already published CAssequences from various animal taxa among which are sevenCnidarian sequences. PhyML and Bayesian analysis arecongruent. The resulting consensus tree separates the 61sequences into four major clusters (Fig. 4). The first clusterrepresents the bacteria outgroup. The second group containssponge CAs. This position of Sponges as an ancestral cladeis in accordance with the results of Jackson et al. (2007).The third and fourth clusters contain invertebrates andvertebrates sequences. “Extracellular” (i.e. transmembrane,secreted and membrane-bound) CAs are grouped in thethird cluster, whereas cytosolic and mitochondrial CAs are
Fig. 1 Presence and expression of STPCA-2 gene in holobiont andCZ. PCR products were deposited on 2.5% agarose gel. RuBisCO and36B4 are respectively used as algal and animal specific genes.Differences between gDNA and cDNA are due to the presence ofintron fragments; 1 kb and 25 bp are DNA ladders from Invitrogen
Mar Biotechnol (2011) 13:992–1002 995

in the fourth group called “intracellular”. We note that inthese two last clusters, vertebrates and invertebratessequences are well separated and Cnidarian sequences aregrouped in both clusters. STPCA-2 branches at the base ofthe intracellular isoforms group. The support value for thisbranching is higher in Bayesian analysis (100%) than inML analysis (>50%).
STPCA-2 Activity The kinetics parameters for the CO2
hydration by the purified recombinant STPCA-2 wasmeasured and compared to human isoform hCA I, hCA II,hCA III, hCA VA and VB, hCA VI, hCA IX and hCA XII(Supuran 2008) as well as the previous enzyme cloned inStylophora pistillata, STPCA (Moya et al. 2008b) (Table 1).STPCA-2 has a higher catalytic constant (5.6.105 s−1) thanthe previous coral enzyme, STPCA (3.1.105 s−1). With a
kcat/Km of 8.3.107 M−1s−1, STPCA-2 is among the mostefficient enzymes tested in this assay, similar to hCA IIand hCA VB. With a Ki of 74 nM, STPCA-2 is assensitive to acetazolamide (AZA) as the mitochondrialisoforms hCA VA and hCA VB (63 and 54 nM, respec-tively). STPCA-2 is less sensitive to AZA than thesecreted STPCA (16 nM).
Immunolocalisation Coral tissue sections were labelledwith anti-STPCA-2 antibody (Fig. 5). Figure 5a, b show atissue section of Stylophora pistillata incubated with pre-immune serum and anti-STPCA-2 antibody, respectively.Figure 5a is not different from natural autofluorescence ofcoral tissues. Magnifications on aboral (Fig. 5c) and oral(Fig. 5d) tissues indicate that STPCA-2 is mainly locatedinside aboral tissue and oral endoderm.
- - - - - - - - - - - - - - - - - - - - - M A S P D W G Y D D K N G - - - P E Q W S K L Y P I A N - G N N Q S P V D I K 35hCAI- - - - - - - - - - - - - - - - - - - - - - M S H H W G Y G K H N G - - - P E H W H K D F P I A K - G E R Q S P V D I D 34hCAII- - - - - - M R A L V L L L S L F L L G G Q A Q H V S D W T Y S E G A L D E A H W P Q H Y P A C G - G Q R Q S P I N L Q 53hCAVIM E L P R T I S W L F A M A A F F L K C T E S L G T W S Y R G P E S - - - P D T W A H H Y P D C A - G H K Q S P V N I V 56FCA-a F. scutariaM E L P R T I S W L F A M A A F F L K C T E S L G T W S Y R G P E S - - - P D T W A H H Y P D C A - G H K Q S P V N I V 56FCA-b F. scutariaM Q M A K F L L V A F V T A A V I S I T L G E D W S Y S V G A S D G - - - P E N W S G - - I C A S - G K K Q S P I D I L 54A030-E11 A. millepora- - - - - - - - - - - - - - - - - - - - - - - - M V W G Y D K E N G - - - P A N W Y K S C P I A F I G K R Q S P I D L I 33C007-E7 A. milleporaM K L S L F I S S L L A M I V A C P N L A E S A G S W T Y R D P E G - - - P D T W K H H Y K D C E - G H E Q S P I N I V 56STPCA S. pistillata- - - - - - - - - M Y F L W L I S M I A A A M C Q E Y G Y M S E Q G V P T P S N W S K V F P L C G - G K F Q S P I N I E 50STPCA-2 S. pistillata
T S E T K H D T S L K P I S V S - - - Y N P A T - - - A K E I I N V G H S F H V N F E D N D N R S V L K G G P F S - D S 88hCAIT H T A K Y D P S L K P L S V S - - - Y D Q A T - - - S L R I L N N G H A F N V E F D D S Q D K A V L K G G P L D - G T 87hCAIIR T K V R Y N P S L K G L N M T G - - Y E T Q A G - - E F P M V N N G H T V Q I S L P S T M R M T V A D G - - - - - T V 104hCAVIP K Q T V F D A G L A D L T I N - - - Y E P N V - - - L A K L E N N G H S I Q A S F L T G K - - S N I S G G G L P - S R 107FCA-a F. scutariaP K Q T V F D A G L A D L T I N - - - Y E P N V - - - L A K L E N N G H S I Q A S F L T G K - - S N I S G G G L P - S R 107FCA-b F. scutariaT K K T E Y D Y S L G S F T L T N Y K H A S P Q - - - N F K S F N N K H T L Q V S L P S D Y - - Y K V S G G N L P - G T 108A030-E11 A. milleporaD N E V K R D S S L E P L V V S - - - Y P P F S - - - K G K F L N N G H S V Q F V P D A S N N L S E L S G G P L A - K K 86C007-E7 A. milleporaP K D T F F E P G L A D L V V N - - - Y E K S V - - - S A K L F N N G H T V Q A T F L T G K - - S N I S G G N L T - S H 107STPCA S. pistillataT K K V K K K - S Y P D L K I S - - - F D N P C G R V T G E L L N A G H S P V V N I D S S K G G A K L S G G P L D C D E 106STPCA-2 S. pistillata
Y R L F Q F H F H W G - - S T N E H G S E H T V D G V K Y S A E L H V A H W N S A K Y S S L A E A A S - - K A D G L A V 144hCAIY R L I Q F H F H W G - - S L D G Q G S E H T V D K K K Y A A E L H L V H W N - T K Y G D F G K A V Q - - Q P D G L A V 142hCAIIY I A Q Q M H F H W G G A S S E I S G S E H T V D G I R H V I E I H I V H Y N - S K Y K S Y D I A Q D - - A P D G L A V 161hCAVIF Q T A Q L H F H W G - - S E N L R G S E H Q I N G R K Y P M E I H I V H Y N A E K Y P N A S T A M K - - K P D G L A V 163FCA-a F. scutariaF Q T A Q L H F H W G - - S E N L R G S E H Q I N G R K Y P M E I H I V H Y N A E K Y P N A S T A M K - - K P D G L A V 163FCA-b F. scutariaF T T V Q L H L H W G - - S D N T K G A E H G M D G M F Y P A E I H F V S F N - N K Y P N I S E S L S - - H S D G L A V 163A030-E11 A. milleporaY K F E Q F H F H W G - - D K D S E G S E H R V N G K M Y S A E L H F V H W D C E T H A S F P E T V K A N K R N G L C V 144C007-E7 A. milleporaF R A L Q M H F H W G - - S E N S R G S E H Q V G G R K F P L E I H I V H Y N A E K Y P S V S E A V D - - K G D G L A V 163STPCA S. pistillataY A L Q Q F H F H F G - - C E N S R G S E H L I D S Q A F P A Q L H L V F F N - K K Y E T F Q N A V D - - K P D G L A V 161STPCA-2 S. pistillata
I G V L M K V G - - E A N P K L Q K V L D A L Q A I K T K G K R A P F T - - N F D - - - P S T L L P S S L - - - - - - - 190hCAIL G I F L K V G - - S A K P G L Q K V V D V L D S I K T K G K S A D F T - - N F D - - - P R G L L P E S L - - - - - - - 188hCAIIL A A F V E V K N Y P E N T Y Y S N F I S H L A N I K Y P G Q R T T L T G - - L D - - - V Q D M L P R N L Q - - - - - - 210hCAVIL G I L V E L Q A - N D N P V L N S I T D M L E T I R Y K G - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - 192FCA-a F. scutariaL G I L V E L Q A - N D N P V L N S I T D M L E T I R Y K G - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - 192FCA-b F. scutariaL G V L L K I G A - S K N I H Y D K F L M E S R V P K E P G T N Y T M T I P S F P - - - L D P L L P A D R T - - - - - - 213A030-E11 A. milleporaL G F F L K V G - - A E N A A L K P I T D M L P K V K K A G S S E P L A I - E F D - - - M R S V M P K D L T - - - - - - 192C007-E7 A. milleporaL G I L V E L Q V - Q D N P V F D V M V D N L D K A R Y K G N - - E V I L P S L Q - - - P F S F L P H D I A - - - - - - 211STPCA S. pistillataL G V L I T A T C - P G N R V L G S F A K K L T K I I E E G A S A N V T - - A V D G I K L N Y L M P Y N N K Q G D E D E 218STPCA-2 S. pistillata
- - - - - - - - - - - - - - - - - - - - - - - - D F W T Y P G S L T H P P L Y E S V T W I I C K E S I S V S S E Q L A Q 226hCAI- - - - - - - - - - - - - - - - - - - - - - - - D Y W T Y P G S L T T P P L L E C V T W I V L K E P I S V S S E Q V L K 224hCAII- - - - - - - - - - - - - - - - - - - - - - - - H Y Y T Y H G S L T T P P C T E N V H W F V L A D F V K L S R T Q V W K 246hCAVI- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - 192FCA-a F. scutaria- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - 192FCA-b F. scutaria- - - - - - - - - - - - - - - - - - - - - - - - K Y F R Y N G S L T T P T C N E A V T W T V F K E A V E I S K A Q M D M 249A030-E11 A. millepora- - - - - - - - - - - - - - - - - - - - - - - - D Y Y T Y D G S L T T P P L L E C V K W I V F K E P L E V S A A Q M E A 228C007-E7 A. millepora- - - - - - - - - - - - - - - - - - - - - - - - Q Y Y T Y R G S L T T P G C F E S V Q W F V F N H T F P I S Q A Q L D K 247STPCA S. pistillataD D E D I A G D D P D A V E E E E V D A K K K I K Y Y T Y K G S L T T P P C Y E S V T W I V F K D K I K I S N T Q L K K 278STPCA-2 S. pistillata
F R S L - L S N V E G D N A V P M Q H N N R P T Q P L K G - - - - R T V R A S F 261hCAIF R K L - N F N G E G E P E E L M V D N W R P A Q P L K N - - - - R Q I K A S F K 260hCAIIL E N - - - - S L L D H R N K T I H N D Y R R T Q P L N - - - - H R V V E S N F P N Q E Y T L G S E F Q F Y L H K I E E 298hCAVI- - - - - - - - - - D T R - - - - - - - - - - - - - - - - - - - - L C - R A L V 201FCA-a F. scutaria- - - - - - - - - - D N R N F - - - - - - - - - - - - - - - - - - R C P K A L F I P S S R N Q P I L H L Q R L S H H R Y 224FCA-b F. scutariaL R G - - - - - L K M N A S N E I V D N Y R P V Q P L N G - - - - R T V K S S F M V I T D Q A I T L Q M S K V V F L P I 300A030-E11 A. milleporaF R - - - - - A V E N G N G E P L V G N Y R P P C P V H S - - - - R T V A A S F K 260C007-E7 A. milleporaF R D L F D S E K Q D T K K L P L V D N Y R P V Q P L Y G R S V S E A S N A L L F P V A R H Q T K L W I A W D S L M T R 307STPCA S. pistillataF R K L - K A Q Y G G A P G - L M C D N I R P V Q P L H K - - - - R K V Y S V L S S R E 316STPCA-2 S. pistillata
261hCAI260hCAII
I L D Y L R R A L N 308hCAVI201FCA-a F. scutaria
R 225FCA-b F. scutariaL F A A L F M L 308A030-E11 A. millepora
260C007-E7 A. milleporaQ Y F M K Q Q S I C A L Y Q P Q 323STPCA S. pistillata
316STPCA-2 S. pistillata
* * *
Fig. 2 ClustalW alignment of the deduced amino acid sequence ofSTPCA-2 with the human isoforms I (hCAI), II (hCAII), VI (hCAVI)and the coral enzymes FCA-a and FCA-b from F. scutaria, A030-E11and C007-E7 from A. millepora and STPCA from S. pistillata. Theamino acids are numbered on the right. Identical amino acids areshaded in black. >75%, >65% and 50 % identical amino acids are
shaded in dark, medium and light gray respectively. The three zinc-binding histidines are indicated by an asterisk. The active site residuesare indicated by inverted black triangles and potential N-glycosylationsites are indicated by black triangles. The signal peptides in N-terminus are indicated in bold
996 Mar Biotechnol (2011) 13:992–1002

Discussion
In this study, we report the cloning in the Scleractiniancoral Stylophora pistillata of a gene encoding a putativeprotein with high similarities with the α-class of CAs andmore particularly with the cytosolic isoform CAII. Thisprotein belongs to the animal partner (Fig. 1) and has thenbeen named STPCA-2 for “STylophora Pistillata CarbonicAnhydrase isoform 2”. The sequence analysis (Fig. 2)revealed that STPCA-2 possesses key amino acids residuessuch as the three zinc-binding histidines, the protonshuttle, the gatekeeper residues and other amino acidsinvolved in the hydrogen-bound network (Hewett-Emmett and Tashian 1996; McCall et al. 2000). Yet,Glu136 (Glu117 in human CAII) is substituted by a Gln inSTPCA-2. This modification could lower the hydrogen-bound network around the active site and alter STPCA-2activity. However, this enzyme appears to be highly activeand efficient (Table 1). With a kcat/Km of 8.3.107, this newcoral CA has, with the human mitochondrial isoform VB(kcat/Km=9.8.10
7), a catalytic efficiency close to hCA II.The lower sensitivity of STPCA-2 to acetazolamidecompared to the previous S. pistillata enzyme STPCA isan encouraging clue to renew inhibition assays in order tobetter study this novel CA in S. pistillata. Indeed thisapproach on STPCA (see Bertucci et al. 2009a, b) allowedus to find some specific interacting molecules thatpotentially inhibit or activate calcification. The same workperformed on STPCA-2 would probably identify suchmolecules and will permit specific in vivo experiments in
which it would be possible to aim at a specific enzyme andto identify its role. So far, the role of CAs in coralphysiology has been determined only by using the non-selective inhibitors AZA and/or ethoxzolamide (EZ). Theproblem is that the two main processes involving CAs,symbiosis and calcification, are strongly linked together(the phenomenon known as light-enhanced calcification isindisputably the best example), and the effects of AZAand EZ on calcification could as much be due to adisruption in carbon supply to photosynthesis as to a directeffect on the biomineralisation process.
The phylogenetic analysis (Fig. 4) confirms that the α-CA family expanded from a single ancestral gene throughseveral independent gene duplication events in spongesand eumetazoans as shown by Jackson et al. (2007). Theseauthors suggest that a putative signal peptide was presentin the ancestral α-CA that was either secreted ormembrane-bound. STPCA-2 possesses a signal peptide inthe N-terminus and branches at the base of intracellularenzymes. This could indicate that STPCA-2 is an ancestralisoform of α-CA as indicated by its position, whereasother Cnidarian CAs group well together inside theintracellular as well as in the extracellular clusters.Moreover, STPCA-2 possesses another atypical feature, a30-amino acid-long insert expanding between Lys212 andLys243 (Fig. 2). This insert does not affect the active siteof STPCA-2. Atypical CAs have already been described ininvertebrates. For instance, in pearl oysters, nacreinpossesses a split CA domain that may confer a regulatoryfunction to this organic matrix protein (Miyamoto et al.1996). In Scleractinian corals, deBoer et al. (2006) haveidentified, in F. scutaria larvae, two isoforms of CAhomologues that possess the three zinc-binding histidinesbut lack crucial amino acids for catalytic activity andexhibit a conserved ATP/GTP binding site. In the samemanner, the two CAs described in A. millepora (Grasso etal. 2008) possess a signal peptide, one of these (A030-E11, accession number ACJ64663) matches with humanCA II as well as STPCA-2. This feature seems to becommon in Cnidarians CAs.
In vertebrates, including Homo sapiens, the physiolog-ical functions of CAs have widely been investigated overthe last 70 years (Supuran 2008): hCA II is involved inrespiration, regulation of the acid/base homeostasy and inthe bone development and function, such as the differen-tiation of osteoclasts or the provision of acid for boneresorption in osteoclasts. In Fig. 5, we observe thatSTPCA-2 is mainly localised in oral endoderm and aboraltissue. As mentioned previously, the way the host suppliesDIC to its algal symbiont remains still poorly character-ised (Al-Moghrabi et al. 1996).
Furla and co-workers (2000a) proposed a model toexplain the mechanism of inorganic carbon uptake by
Fig. 3 Western blot: recombinant STPCA (see Moya et al. 2008b)and STPCA-2 are subjected to SDS-PAGE, electroblotted andincubated with anti-His Tag (lanes a, b and c) or anti-STPCA-2(lanes d and e). a, d Recombinant STPCA. b, e RecombinantSTPCA-2. c Recombinant STPCA-2 after N-glycosidase F treatment
Mar Biotechnol (2011) 13:992–1002 997
998 Mar Biotechnol (2011) 13:992–1002

Cnidarian oral tissue. This model includes two CAs. Thefirst is externally bound to the apical membrane of ectodermalcells and is coupled with a proton pump. This CA is used toconvert the external HCO3
− into CO2, which easily diffuseinto the cell. There, a second CA converts CO2 back toHCO3
− that will be transferred to the endodermal cellscontaining symbionts. This conversion also avoids the leakof inorganic carbon by diffusion of CO2 out of the cell andcan be considered as an inorganic carbon trapping mecha-nism. Our results show that STPCA-2 does not correspond toany of this two CA. This means that other isoforms exist andremain to be characterised in symbiotic Cnidarians.
In a recent paper, Venn et al. (2009) performed the firstintracellular pH (pHi) measurement in Cnidarians. Twoimportant results came out of this study. First, the pHi of S.pistillata endodermal cells containing symbiotic algae is of7.41±0.22 in the light. This result suggests that theintracellular DIC form is HCO3
−. Secondly, a more acidicarea (pH 6.0 or lower) surrounds zooxanthellae. It is notclear whether this more acidic area is associated with themembrane or with the medium surrounding the symbiont(called perisymbiotic space), but this acidification could beachieved by a plasma membrane P-type H+-ATPasebelonging to the zooxanthellae (Bertucci et al. 2010a). Insuch a condition, DIC form is predicted to be CO2. STPCA-2 is present in these symbiont-containing cells (Fig. 5d) and
could play different roles depending on the surrounding pH,i.e. DIC trapping under the form of bicarbonate in thecytosol and formation of CO2 near the photosynthetic algaeto supply photosynthesis.
Concerning calcification, it has been shown that 25%of DIC used for CaCO3 deposition comes from theseawater HCO3
−, whereas the major source (75%) is themetabolic CO2 (Erez 1978; Furla et al. 2000b) producedby the numerous mitochondrion present in aboral ecto-derm, also called calicoderm (Furla et al. 2000b; Tambuttéet al. 2007a). In this manner, external DIC must cross theaboral tissue where STPCA-2 is likely to ensure thetrapping of this DIC under the form HCO3
− prior to itstransport toward the calcifying cells through a way that isstill unknown. Finally, in these calcifying cells, even ifCO2 could diffuse toward the site of calcification (whereSTPCA is present, Moya et al. 2008b) a part of thisinternal DIC could be converted into HCO3
− intracellular-ly by STPCA-2. The presence of an intracellular CA in thecalicoderm was already suggested by Furla et al. (2000b).All these data are summarised in a model diagramshowing the movements of the different forms of DICand the localisation of STPCA and STPCA-2 enzymes incoral tissues (Fig. 6).
Our study confirms the importance of carbonicanhydrases in coral physiology. Owing to its intracellu-lar localisation and similarity with hCA II, we proposethat the present STPCA-2 is involved in pH regulationby trapping the DIC under the form of bicarbonate andhelps its delivery either to symbionts or to thecalcification site. Due to the presence of a signalpeptide, we cannot exclude the possibility thatSTPCA-2 is localised inside intracellular vesicles thatare not observable with confocal microscopy. Thus,further pharmacological and physiological studies willattempt to better characterise this enzyme and determineits role.
Isozyme Activity level kcat (s−1) kcat/Km (M−1 s−1) Ki (acetazolamide) (nM)
hCA I Moderate 2.0.105 5.0.107 250
hCA II Very high 1.4.106 1.5.108 12
hCA III Very low 1.0.104 3.0.105 300,000
hCAVA Low 2.9.105 2.9.107 63
hCAVB High 9.5.105 9.8.107 54
hCAVI Moderate 3.4.105 4.9.107 11
hCA IX High 3.8.105 5.5.107 25
hCA XII Moderate 4.2.105 3.5.107 5.7
STPCA Moderate 3.1.105 4.6.107 16
STPCA-2 High 5.6.105 8.3.107 74
Table 1 Kinetic parameters forthe CO2 hydration reaction cat-alysed by the human carbonicanhydrases isoforms I, II, III,VA, VB, VI, IX and XII and theStylophora pistillata enzymesSTPCA and STPCA-2 at 20°Cand pH 7.5 in 10 mM Hepesbuffer, and their inhibition datawith AZA
Fig. 4 Phylogenetic analysis of STPCA-2 (indicated in bold red) and60 previously cloned CA sequences inferred from both ML andBayesian analyses. The results from ML bootstrap analysis are shownabove the branches, only values >50% are indicated. The full, dashedand dotted branches represent >95%, 75–95% and 50–75% Bayesianposterior probabilities, respectively. The accession numbers are fromthe protein sequence database of Entrez from the National Center ofBiotechnology Information. The branch lengths are proportional to thenumber of substitutions per site (see scale bar in the figure). OtherCnidarian sequences are indicated in green
�
Mar Biotechnol (2011) 13:992–1002 999
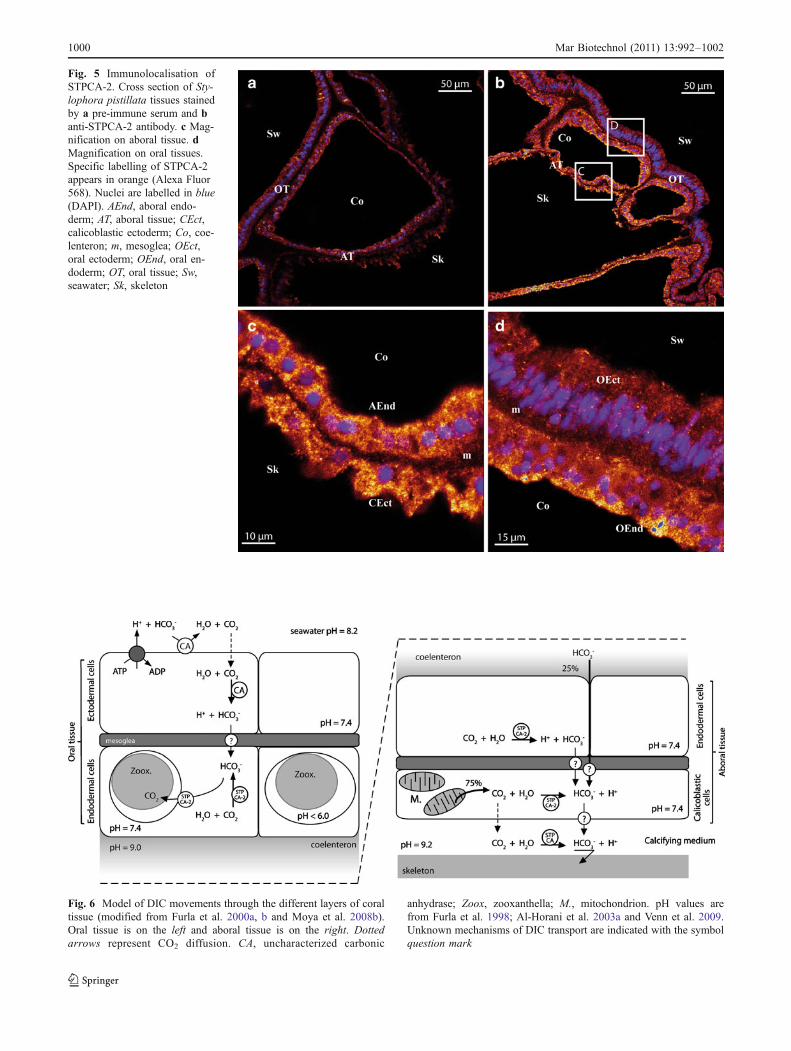
Fig. 6 Model of DIC movements through the different layers of coraltissue (modified from Furla et al. 2000a, b and Moya et al. 2008b).Oral tissue is on the left and aboral tissue is on the right. Dottedarrows represent CO2 diffusion. CA, uncharacterized carbonic
anhydrase; Zoox, zooxanthella; M., mitochondrion. pH values arefrom Furla et al. 1998; Al-Horani et al. 2003a and Venn et al. 2009.Unknown mechanisms of DIC transport are indicated with the symbolquestion mark
Fig. 5 Immunolocalisation ofSTPCA-2. Cross section of Sty-lophora pistillata tissues stainedby a pre-immune serum and banti-STPCA-2 antibody. c Mag-nification on aboral tissue. dMagnification on oral tissues.Specific labelling of STPCA-2appears in orange (Alexa Fluor568). Nuclei are labelled in blue(DAPI). AEnd, aboral endo-derm; AT, aboral tissue; CEct,calicoblastic ectoderm; Co, coe-lenteron; m, mesoglea; OEct,oral ectoderm; OEnd, oral en-doderm; OT, oral tissue; Sw,seawater; Sk, skeleton
1000 Mar Biotechnol (2011) 13:992–1002

Acknowledgments We are grateful to Nathalie Techer for hertechnical help and to Dominique Desgré for coral maintenance.
This study was conducted as part of the Centre Scientifique deMonaco Research Programme, supported by the Government of thePrincipality of Monaco. Anthony Bertucci was supported by afellowship from the Scientific Centre of Monaco.
References
Abascal F, Zardoya R, Posada D (2005) ProtTest: selection of best-fitmodels of protein evolution. Bioinformatics 21:2104–2105
Al-Horani FA, Al-Moghrabi SM, de Beer D (2003a) Microsensorstudy of photosynthesis and calcification in the scleractiniancoral, Galaxea fascicularis: active internal carbon cycle. J ExpMar Biol Ecol 288:1–15
Al-Horani FA, Al-Moghrabi SM, de Beer D (2003b) The mechanismof calcification and its relation to photosynthesis and respirationin the scleractinian coral Galaxea fascicularis. Mar Biol142:419–426
Al-Moghrabi S, Goiran C, Allemand D, Speziale N, Jaubert J (1996)Inorganic carbon uptake for photosynthesis by the symbioticcoral-dinoflagellate association II. Mechanisms for bicarbonateuptake. J Exp Mar Biol Ecol 199:227–248
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W,Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a newgeneration of protein database search programs. Nucleic AcidsRes 25:3389–3402
Bertucci A, Innocenti A, Zoccola D, Scozzafava A, Allemand D,Tambutté S, Supuran CT (2009a) Carbonic anhydrase inhibitors:inhibition studies of a coral secretory isoform with inorganicanions. Bioorg Med Chem Lett 19:650–653
Bertucci A, Innocenti A, Zoccola D, Scozzafava A, Tambutté S,Supuran CT (2009b) Carbonic anhydrase inhibitors. Inhibitionstudies of a coral secretory isoform by sulfonamides. Bioorg MedChem 17:5054–5058
Bertucci A, Tambutté E, Tambutté S, Allemand D, Zoccola D (2010a)Symbiosis-dependent gene expression in coral-dinoflagellateassociation: cloning and characterization of a P-type H+-ATPasegene. Proc R Soc B 277:87–95
Bertucci A, Zoccola D, Tambutté S, Vullo D, Supuran CT (2010b)Carbonic anhydrases activators. the first activation study of acoral secretory isoform with amino acids and amines. BioorgMed Chem 18:2300–2303
Blom N, Gammeltoft S, Brunak S (1999) Sequence and structure-based prediction of eukaryotic protein phosphorylation sites. JMol Biol 294:1351–1362
Corpet F (1988) Multiple sequence alignment with hierarchicalclustering. Nucleic Acids Res 16:10881–10890
deBoer ML, Krupp DA, Weis VM (2006) Two atypical carbonicanhydrase homologs from the planula larva of the scleractiniancoral Fungia scutaria. Biol Bull 211:18–30
Eisenhaber B, Bork P, Eisenhaber F (1999) Prediction of potentialGPI-modification sites in proprotein sequences. J Mol Biol292:741–758
Emanuelsson O, Brunak S, von Heijne G, Nielsen H (2007) Locatingproteins in the cell using TargetP, SignalP and related tools. NatProtoc 2:953–971
Erez J (1978) Vital effect on stable-isotope composition seen inforaminifera and coral skeletons. Nature 273:199–202
Furla P, Bénazet-Tambutté S, Jaubert J, Allemand D (1998) Functionalpolarity of the tentacle of the sea anemone Anemonia viridis: rolein inorganic carbon acquisition. Am J Physiol 274:R303–R310
Furla P, Allemand D, Orsenigo M (2000a) Involvement of H+-ATPaseand carbonic anhydrase in inorganic carbon uptake for endosym-
biont photosynthesis. Am J Physiol Regul Integr Comp Physiol278:R870–R881
Furla P, Galgani I, Durand I, Allemand D (2000b) Sources andmechanisms of inorganic carbon transport for coral calcificationand photosynthesis. J Exp Biol 203:3445–3457
Giordano M, Beardall J, Raven JA (2005) CO2 concentratingmechanisms in algae: mechanisms, environmental modulation,and evolution. Annu Rev Plant Biol 56:99–131
Goreau TF (1959) The physiology of skeleton formation in corals. I.A method for measuring the rate of calcium deposition by coralsunder different conditions. Biol Bull, Mar Biol Lab, Woods Hole116:59–75
Grasso L, Maindonald J, Rudd S, Hayward DC, Saint R, Miller DJ,Ball EE (2008) Microarray analysis identities candidate genes forkey roles in coral development. BMC Genomics 9:540
Guindon S, Gascuel O (2003) A simple, fast, and accurate algorithmto estimate large phylogenies by maximum likelihood. Syst Biol52:696–704
Hewett-Emmett D, Tashian RE (1996) Functional diversity, conser-vation, and convergence in the evolution of the alpha-, beta-, andgamma-carbonic anhydrase gene families. Mol Phylogenet Evol5:50–77
Huelsenbeck, Ronquist (2001) MRBAYES: Bayesian inference ofphylogenetic trees. Bioinformatics 17:754–755
Isa Y, Yamazato K (1984) The distribution of carbonic anhydrase in astaghorn coral Acropora hebes (Dana). Galaxea 3:25–36
Jackson DJ, Macis L, Reitner J, Degnan BM, Worheide G (2007)Sponge paleogenomics reveals an ancient role for carbonicanhydrase in skeletogenesis. Science 316:1893–1895
Khalifah RG (1971) The carbon dioxide hydration activity of carbonicanhydrase. I. Stop-flow kinetic studies on the native humanisoenzymes B and C. J Biol Chem 246:2561–2573
Kingsley RJ, Watabe N (1987) Role of carbonic anhydrase in calcificationin the gorgonian Leptogorgia virgulata. J Exp Zool 241:171–180
Leggat W, Badger MR, Yellowlees D (1999) Evidence for aninorganic carbon-concentrating mechanism in the symbioticdinoflagellate Symbiodinium sp. Plant Physiol 121:1247–1255
Leggat W, Marendy EM, Baillie B, Whitney SM, Ludwig M, BadgerMR, Yellowlees D (2002) Dinoflagellate symbioses: strategiesand adaptations for the acquisition and fixation of inorganiccarbon. Funct Plant Biol 29:309–322
Lucas JM, Knapp LW (1997) A physiological evaluation of carbonsources for calcification in the octocoral Leptogorgia virgulata(Lamarck). J Exp Biol 200:2653–2662
Marshall AT, Clode PL (2003) Light-regulated Ca2+ uptake and O2
secretion at the surface of a scleractinian coral Galaxeafascicularis. Comp Biochem Physiol A Mol Integr Physiol136:417–426
McCall KA, Huang C, Fierke CA (2000) Function and mechanism ofzinc metalloenzymes. J Nutr 130:1437S–1446S
Miyamoto H, Miyashita T, Okushima M, Nakano S, Morita T,Matsushiro A (1996) A carbonic anhydrase from the nacreouslayer in oyster pearls. Proc Natl Acad Sci USA 93:9656–9660
Moya A, Tambutté S, Beranger G, Gaume B, Scimeca JC, AllemandD, Zoccola D (2008a) Cloning and use of a coral 36B4 gene tostudy the differential expression of coral genes between light anddark conditions. Mar Biotechnol NY 10:653–663
Moya A, Tambutté S, Bertucci A, Tambutté E, Lotto S, Vullo D,Supuran CT, Allemand D, Zoccola D (2008b) Carbonic anhy-drase in the scleractinian coral Stylophora pistillata: character-ization, localization, and role in biomineralization. J Biol Chem283:25475–25484
Puverel S, Tambutté E, Zoccola D, Domart-Coulon I, Bouchot A,Lotto S, Allemand D, Tambutté S (2005) Antibodies against theorganic matrix in scleractinians: a new tool to study coralbiomineralization. Coral Reefs 24:149–156
Mar Biotechnol (2011) 13:992–1002 1001

Rahman MA, Oomori T, Uehara T (2008) Carbonic anhydrase incalcified endoskeleton: novel activity in biocalcification inalcyonarian. Mar Biotechnol (NY) 10:31–38
Supuran CT (2008) Carbonic anhydrases: novel therapeutic applicationsfor inhibitors and activators. Nat Rev Drug Discov 7:168–181
Tambutté E, Allemand D, Mueller E, Jaubert J (1996) A compart-mental approach to the mechanism of calcification in hermatypiccorals. J Exp Biol 199:1029–1041
Tambutté E, Allemand D, Zoccola D, Meibom A, Lotto S, Caminiti N,Tambutté S (2007a) Observations of the tissue-skeleton interface inthe scleractinian coral Stylophora pistillata. Coral Reefs 26:517–529
Tambutté S, Tambutté E, Zoccola D, Caminiti N, Lotto S, Moya S,Allemand D, Adkins J (2007b) Characterization and role ofcarbonic anhydrase in the calcification process of the azooxan-thellate coral Tubastrea aurea. Mar Biol 151:71–83
Venn AA, Tambutté E, Lotto S, Zoccola D, Allemand D, TambuttéS (2009) Imaging intracellular pH in a reef coral andsymbiotic anemone. Proc Natl Acad Sci USA 106:16574–16579
Weis VM (1993) Effect of dissolved inorganic carbon concentrationon the photosynthesis of the symbiotic sea anemone Aiptasiapulchella Carlgren: role of carbonic anhydrase. J Exp Mar BiolEcol 174:209–225
Weis VM, Reynolds WS (1999) Carbonic anhydrase expression andsynthesis in the sea anemone Anthopleura elegantissima areenhanced by the presence of dinoflagellate symbionts. PhysiolBiochem Zool 72:307–316
Xu Y, Feng L, Jeffrey PD, Shi Y, Morel FM (2008) Structure andmetal exchange in the cadmium carbonic anhydrase of marinediatoms. Nature 452:56–61
1002 Mar Biotechnol (2011) 13:992–1002




















