
Alteraciones del acoplamiento excitación-contracción y la ...

UNIVERSIDAD NACIONAL DE SAN LUIS
FACULTAD DE QUÍMICA BIOQUÍMICA Y FARMACIA
Características de la contracción de células mioides
peritubulares de los túbulos seminíferos de testículo de rata
Trabajo de Tesis
Presentado por
Licenciado Darío Fernández
Para optar el grado académico de
Doctor en Bioquímica
Asesor Científico: Doctor Luis Alberto López
Co-asesor Científico: Doctor Juan Carlos Cavicchia
San Luis. República Argentina
2008

AGRADECIMIENTOS
La presente investigación representa un esfuerzo que culmina con
varios años de estudios y dedicación hacia la ciencia.
Agradezco al Dr. Luis López, por sus observaciones atentas, sus
críticas y sugerencias, sus investigaciones paralelas a las mías y a la
orientación personal y profesional, que imprimió a mi labor, para poder
comenzar a recorrer el camino de la ciencia.
Expreso mi cordial gratitud al Dr. Juan Carlos Cavicchia por su gran
aporte y co-asesoramiento científico.
Estoy muy agradecido de todo el personal del Instituto de Histología y
Embriología de la Facultad de Ciencias Médicas de la Universidad
Nacional de Cuyo.
También quiero agradecer a mis compañeros de trabajo del
Laboratorio de Citoesqueleto y Ciclo Celular, por los momentos
compartidos a lo largo del desarrollo de esta investigación.
Por último, agradezco a Diego Fernández por la colaboración que me
ha prestado en el desarrollo de imágenes y animaciones de la molécula
de miosina.
Darío Fernández
Mendoza, 2 008.

_____________________________________________________________________
_____________________________________________________________________ Índice General
Para Mario y Aurora
A mis Hermanos, Mario y Diego
A Cami, Joe y Timoteo

_____________________________________________________________________
_____________________________________________________________________ Índice General
ÍNDICE GENERAL
CAPÍTULO I 11
INTRODUCCIÓN 11
1. EL TESTÍCULO 12
1.1. Estructura del testículo 12
1.1.1. Túbulos seminíferos del testículo 13
1.1.2. Células mioides peritubulares 13
1.1.3. El epitelio seminífero 14
1.1.4. Desarrollo del túbulo seminífero 16
1.1.5. Desarrollo del epitelio seminífero 17
1.1.6. Espermatogénesis 21
1.1.7. El ciclo del epitelio seminífero 21
1.1.8. Organización del epitelio seminífero 24
1.1.9. Regulación de la contracción del túbulo seminífero 29
2. CARACTERÍSTICAS DE MIOSINA 31
2.1. Miosinas 31
2.1. Descubrimiento de Miosina 31
2.2. Estructura de la molécula de Miosina II 32
2.2.1. Estructura de la porción C-terminal de miosina II 35
2.3. Estructura de los filamentos gruesos de miosina II 35
2.3.1. Ensamble y desensamble de filamentos gruesos 37
2.3.2. Implicancia de la fosforilación de las cadenas livianas en la
formación del filamento 38

_____________________________________________________________________
_____________________________________________________________________ Índice General
2.3.3. Implicancia de la fosforilación de la cadena pesada en la
formación de filamentos de miosina II 40
3. PROPÓSITO DEL TRABAJO DE TESIS 41
CAPÍTULO II 42
CARACTERÍSTICAS DE LA MOLÉCULA DE MIOSINA DE CÉLULAS
MIOIDES PERITUBULARES DEL TESTÍCULO 42
1.1. Contenido de miosina en el túbulo seminífero de testículo 43
1.2. Purificación de miosina-CMP 47
1.2.1. Método agregación-desagregación 47
1.2. 2. Purificación de miosina-CMP en gradiente de sacarosa 49
1.2. 3. Purificación de miosina-CMP en columna de hidroxiapatita
50
2. CARACTERÍSTICAS PEPTÍDICAS DE LA CADENA PESADA DE
MIOSINA-CPM 52
2.1. Digestión Enzimática 52
2.2. Espectrometría de masas 53
2.2.1. Miosina-CMP de testículo de rata 53
2.2.2. Miosina-CMP de testículo de toro 58
3. PROPIEDADES DE SOLUBILIDAD DE MIOSINA-CMP 60
3.1. Solubilidad in vitro 60
3.1.1. Solubilidad de miosina de otros tejidos 61
3.2. Agregación in vitro 63
3.3. Desagregación in vitro 66

_____________________________________________________________________
_____________________________________________________________________ Índice General
4. LOCALIZACIÓN DE MIOSINA-CMP EN LOS TEJIDOS DEL TESTÍCULO
70
4.1 Células mioides peritubulares. 70
5. DISCUSIÓN DEL CAPÍTULO II 73
CAPÍTULO III 76
CARACTERÍSTICAS DE LAS CÉLULAS MIOIDES PERITUBULARES DEL
TESTÍCULO 76
1. IDENTIFICACIÓN DE CÉLULAS MIOIDES PERITUBULARES EN
TÚBULOS SEMINÍFEROS AISLADOS 77
1.1. Tinción con nitrato de plata 77
1.2. Contracción de las células mioides peritubulares 79
1.2.1. Diámetro de los túbulos seminíferos y tamaño de las células
mioides peritubulares 80
2. GRADO DE AGREGACIÓN DE MIOSINA-CMP 83
2.1. Efecto de endotelina-1 83
3. DISCUSIÓN DEL CAPÍTULO III 85
CAPÍTULO IV 86
ACTIVIDAD DE LAS CÉLULAS MIOIDES PERITUBULARES
RELACIONADA CON EL EPITELIO SEMINIFERO 86
1. DESARROLLO DEL TÚBULO SEMINÍFERO Y DE LAS CÉLULAS
MIOIDES QUE LO RODEAN 87
1.1. Diámetro del túbulo seminífero de testículos en desarrollo 87
1.2. Caracteres morfológicos de las células-MP en túbulos
seminíferos en desarrollo 89

_____________________________________________________________________
_____________________________________________________________________ Índice General
1.3. Isotipos de miosina-CMP en las células mioides peritubulares en
desarrollo 92
2. CARACTERÍSTICAS DE LA CONTRACCIÓN DE LOS SEGMENTOS DEL
EPITELIO SEMINÍFERO Y DE LAS CÉLULAS MIOIDES QUE LO RODEAN
94
2.1 Diámetro de distintos segmentos de túbulos seminíferos 95
2.2 Morfología de las células mioides peritubulares en los distintos
segmentos del túbulo seminífero 96
2.3. Isotipos de miosina-CMP en los distintos segmentos del túbulo
seminífero 99
3. DISCUSIÓN DEL CAPITULO IV 101
CAPÍTULO V 103
DISCUSION GENERAL Y CONCLUSIONES 104
1. Características de miosina-CMP 104
2. Contracción de las células MP y expresión de isotipos de miosina-
CMP 111
CAPÍTULO VI 117
MATERIALES Y METODOS 118
1. Animales y materiales biológicos 118
2. Aislamiento de túbulos seminíferos y de células del testículo 118
3. Visualización y medición de las células mioides peritubulares en el
túbulo seminífero 119
4. Tratamiento de los túbulos seminíferos con endotelina-1 120
5. Análisis de proteínas 120

_____________________________________________________________________
_____________________________________________________________________ Índice General
6. Obtención y purificación de proteínas 122
7. Digestión de proteínas con tripsina 125
8. Cuantificación de proteínas 125
9. Precipitación de proteínas 126
10. Agregación y desagregación de miosina 126
11. Análisis de datos 127
Bibliografía 129
Publicaciones relacionadas con los resultados presentados 141

_____________________________________________________________________
_____________________________________________________________________ Índice General
ABREVIATURAS
ASB: Albúmina de suero bovino.
ATP: Adenosina trifosfato.
Células MP: Células mioides peritubulares.
ET-1: endotelina-1.
E.S.M: error standart medio.
H: hora
kDa: kilo Dalton.
M: concentración molar.
mM: concentración milimolar.
mm (minúscula): milímetro.
µM: concentración micromolar.
nM: concentración nanomolar.
Min: minuto.
µm: micrómetro.
µm2: micrómetro cuadrado.
Miosina-CMP: Miosina proveniente de células mioides peritubulares.
PAGE-SDS: Electroforesis en gel de poliacrilamida con SDS.
PBS: bufer fosfato salino.
Pn: posnacimiento
SDS: Dodecil sulfato de sodio.
SMM-II: Miosina II de músculo liso.

_____________________________________________________________________
_____________________________________________________________________ Índice General
TS: Túbulo(s) seminífero(s).
Xg: unidad gravitatoria.

CAPÍTULO I
INTRODUCCIÓN

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 12
1. EL TESTÍCULO
1.1. Estructura del testículo
Los órganos reproductores masculinos, se clasifican en órganos
sexuales internos y externos. Los internos incluyen testículo,
epidídimo, conducto deferente y glándulas sexuales anexas, (estas
últimas incluyen vesícula seminal, próstata y glándula bulbouretal).
Los externos son pene y escroto.
El testículo, como se ilustra en la figura I.1. Está rodeado por
una gruesa cápsula de tejido conectivo, la túnica albugínea, que se
extiende hacia el interior del testículo. Dentro de las cavidades que
forma la túnica, se encuentran los túbulos seminíferos que son los
responsables de la producción y transporte de los espermatozoides.
Los túbulos seminíferos están separados por tejido intersticial
que contiene células de Leydig y vasos sanguíneos.

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 13
Figura I.1. Esquema general de la organización del testículo.
(Figura extraída de Geneser y col 2000)
1.1.1. Túbulos seminíferos del testículo
Los túbulos seminíferos son una serie de conductos que
contienen en su interior el epitelio seminífero y están rodeados por
células mioides peritubulares y tejido conectivo.
1.1.2. Células mioides peritubulares
Las células mioides peritubulares (células MP) son de origen
mesenquimático, representan el mayor componente celular de la pared
de los túbulos seminíferos y participan en la regulación paracrina de la
función testicular.

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 14
Estudios in vitro han demostrando que las células MP secretan
un número de sustancias, incluyendo componentes de la matriz
extracelular y factores de crecimiento, algunos de los cuales estimulan
en forma paracrina la secreción de las células de Sértoli
Las células MP son contráctiles, expresan proteínas de
verdaderas células musculares lisas tales como alfa-isoactina, y
miosina y forman prominentes haces de microfilamentos. En la rata
adulta los microfilamentos dentro de cada célula están orientados en
forma circular y longitudinal al eje mayor del túbulo seminífero.
La actividad contráctil de las células MP es responsable de la
contracción de los túbulos seminíferos para el transporte de
espermatozoides y fluido testicular, y por lo menos parcialmente,
para la liberación de espermatozoides durante la espermiación
(Virtanen y col., 1986); (Tung y Fritz, 1990).
Varios agonistas (endotelina-1, vasopresina, el factor de
crecimiento derivado de las plaquetas, oxitocina y prostaglandinas,
entre otros, regulan la contracción de las células MP (Rossi y col.,
2002)
1.1.3. El epitelio seminífero
El epitelio seminífero es un epitelio estratificado complejo
compuesto por dos tipos celulares básicos: células de Sertoli y células
espermatogénicas.
Las células de Sértoli, representan una población no proliferante
que se extienden desde la membrana basal hasta la luz del túbulo

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 15
seminífero. Tienen forma cilíndrica, con complejas prolongaciones
apicales y laterales que rodean las células espermatogénicas
adyacentes y llenan los espacios vacíos entre ellas.
Las células de Sértoli adyacentes están unidas entre sí por
complejos de unión que forman la barrera hematotesticular, que es
una barrera fisiológica que separa el tejido de las sustancias
transportadas por la sangre.
Las células de Sertoli desempeñan un papel esencial en el
desarrollo de las células espermatogénicas, ya que secretan líquidos o
restringen el movimiento de moléculas, creando micro ambientes
esenciales para el desarrollo y la diferenciación de las mismas.
Además las células de Sértoli median el movimiento a través del
epitelio seminífero de esteroides, metabolitos y sustancias nutritivas;
restringen la filtración de moléculas extracelulares al epitelio por medio
de los complejos de unión; fagocitan las células espermatogénicas en
degeneración y el exceso de citoplasma eliminado por las espermátides
en diferenciación; secretan proteínas ligadoras de andrógenos, que
concentran la testosterona dentro del epitelio seminífero y la parte
proximal de las vías de conducción; secretar varias sustancias
inhibidoras y estimuladoras que regulan la mitosis, meiosis, funciones
esteroidogénicas de las células de Leydig y la liberación de
gonadotropinas; controlan el movimiento de células espermatogénicas
en el epitelio seminífero y la liberación de los espermatozoides a la luz
del túbulo. (Brinster, 2007)

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 16
1.1.4. Desarrollo del túbulo seminífero
La más extensa y detallada investigación del desarrollo de los
túbulos seminíferos ha sido llevada a cabo en ratas de laboratorio. Los
túbulos fueron reconstruidos de cortes seriados de testículos,
comenzando con el día 16 de vida embrionaria y continuando hasta los
de ratas adultas.
En cada testículo fetal hay 20-31 cordones sexuales distintos
que están organizados en arcos en forma de C, apilados entre si como
el cartílago de la tráquea. Se pueden distinguir dos grupos de arcos;
unos localizados periféricamente, formando los cordones externos y
corriendo próximos a la túnica albugínea y los otros formados por
grupos mas pequeños alejados de la túnica albugínea. Los cordones
tienen solo dos conexiones a la rete testis. A medida que el
desarrollo de los cordones progresa, su crecimiento longitudinal causa
más ondulaciones y plegamiento de los arcos.
En el momento del nacimiento, los arcos están extensamente
plegados formando alrededor de 90 pequeñas circunvalaciones. El
desarrollo de los túbulos continúa por el crecimiento longitudinal de las
porciones entre las circunvalaciones sin variar el número de ellas. El
subsiguiente desarrollo de los túbulos seminíferos está dado por el
crecimiento en longitud y en diámetro. Como se muestra en la figura
I.2 el diámetro de los túbulos seminíferos tiene un crecimiento
sostenido hasta los 50 días postnacimiento (pn), donde adquiere la

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 17
medida de los túbulos seminíferos de los testículos adultos (Greep y
Astwood, 1975)
1.1.5. Desarrollo del epitelio seminífero
Los túbulos seminíferos en la rata fetal contienen dos tipos de
células. Las células de sostén (precursores de las células de Sértoli y
los gonocitos (células germinales primordiales). Los dos tipos celulares
están en activa división mitótica durante la vida fetal.
En la rata, en el momento de nacimiento, alrededor del 95 % de
los túbulos no presentan lumen, son cordones macizos. Entre los 10-
14 días pn, algunos cordones comienzan a mostrar pequeñas zonas
con lumen que van progresando marcadamente, hasta llegar al día 18
pn donde el 85 % de los cordones están abiertos. Para el día 30 pn
todos los cordones ya son verdaderos túbulos con lumen. El diámetro
de la luz de los túbulos crece lentamente hasta el día 20 pn y luego
rápidamente hasta el día 50 pn. (Ver figura I.3)
En el momento del nacimiento, las células de Sertoli no
presentan características especiales de su citoplasma y recién
comienzan a tener el aspecto de células maduras desde el día 16 pn.
Se observan transformaciones del citoplasma apical, denominadas
especializaciones ectoplásmicas (ES) y la formación de uniones
laterales entre células de Sértoli, formando una barrera tipo epitelial.
Para el día 22 pn las células de Sértoli presentan las características de
células maduras.

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 18
Hay evidencias, que en la rata, algunos gonocitos embrionarios
degeneran y mueren en los primeros días del nacimiento, pero los
restantes, proliferan por mitosis en el día 4 pn y forman las
espermatogonias tipo A que entrarán en el ciclo espermatogénico para
formar la bases para la definitiva línea de células germinales. En el día
15 pn las espermatogonias y los espermatocitos primarios están
presentes. Espermátides redondas se encuentran en el día 30 pn.
Espermátides elongadas comienzan a observarse en algunos túbulos
en el día 40 pn y en todos al día 50 pn. A los 50 pn se considera que el
primer ciclo espermatogénico se ha completado. Recién a los 60 días
pn el epidídimo está maduro y contiene un gran número de
espermatozoides (Greep y Astwood, 1975).

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 19
Figura I.2. Diámetro del túbulo seminíferos en ratas de varias edades
(Figura extraída de Greep y Astwood, 1975)

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 20
Las células mioides peritubulares se originan de células
precursoras mesenquimáticas en los primeros días posnatal y
presentan escaso desarrollo hasta el día 14 pn (Campagnolo y col.,
2001). En el día 16 pn comienza a desarrollarse microfilamentos
paralelos aislados (filamentos de actina) en forma de haces orientados
en forma circular al eje mayor del túbulo seminífero, y en el día 30 pn
comienzan a aparecen en adición a los circulares, haces que corren
longitudinalmente al eje mayor del túbulo. Por el día 40 pn, hay un
gran desarrollo de haces de microfilamentos formando una red con
orientación circular y longitudinal, que luego pierde intensidad en la
edad adulta (Maekawa y col., 1995).
Figura I.3. Diámetro del lumen tubular en ratas de varias edades
(Figura extraída de Russell y col. 1989)

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 21
1.1.6. Espermatogénesis
La organización de las células internas del túbulo seminífero ha
sido estudiada especialmente en ratas. (Leblond y Clermont, 1952. a y
b; Clermont, 1957). En menor grado en ratones (Oakberg, 1956 a y
b) y eventualmente en otras especies.
Se denomina espermatogénesis al proceso sumamente ordenado
y definido que envuelve el desarrollo del espermatozoide desde células
progenitoras que residen dentro del epitelio germinal del testículo.
Las células germinales maduras, las espermatogonias, se
dividen para formar las células meióticas o espermatocitos primarios
que al final de la división reduccional forman las células haploides
denominadas espermatides (espermatocinesis). Las espermátides se
convierten en espermatozoides por el resultado de una complicada
metamorfósis envolviendo dramáticas modificaciones estructuras del
núcleo, formación de nuevas organelas, y la adquisición de
mecanismos que le permiten una movilidad direccional independiente
(espermogénesis).
1.1.7. El ciclo del epitelio seminífero
Un detallado estudio de los cambios estructurales progresivos
que se producen en las espermátides en el curso de su desarrollo y en
la formación de los espermatozoides llevó a los investigadores a
describir en la rata 14 etapas bien definidas de la espermiogénesis.

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 22
Además se comprobó que cada sección transversal del túbulo
seminífero contiene espermátides en alguno de los 14 estados
mencionados formando una asociación específica con ciertas células
del resto del epitelio germinal. Estas evidencias llevó el concepto
original de ciclo espermatogénico a la idea de ciclo del epitelio
seminífero definido como “una serie de cambios en una determinada
área del epitelio seminífero ubicada entre dos áreas del mismo estado
de desarrollo (Leblond y Clermont, 1952 a y b)
En la rata el ciclo del epitelio seminífero consiste de 14 estados o
asociaciones celulares características como se muestra en la figuras I.4
y I.5. Tanto la formación de asociaciones específicas de células como
la secuencia de su aparición en una determinada área del túbulo
seminífero son eventos altamente sincronizados.
La duración del ciclo del epitelio seminífero en ratas depende de
la variedad. En las ratas Sherman el ciclo es de 12 días, en ratas
Sprague-Dawley el ciclo es 12,9 días, en ratas Wister 13 días y en
ratas Vandicoot 10 días.

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 23
Figura I. 4. Micrografía electrónica de una porción de la pared del
epitelio seminífero de un testículo que muestra una asociación celular
o estado, compuesto de espermatogonia, espermatocitos y dos
generaciones de espermátides. A la derecha son las células
individuales que constituyen una asociación celular
(Figura extraída de Russell y col. 1990).

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 24
Figura. I.5. Mapa del ciclo de la espermatogénesis de la rata, las
columnas verticales, están designadas con números romanos
representan las asociaciones celulares o estados. La progresión del
desarrollo de las células es seguida horizontalmente hasta el borde
derecho del mapa del ciclo, la progresión continúa de la izquierda del
mapa en fila, éste termina con formación del espermatozoide
(Figura extraída de Russell y col. 1990).
1.1.8. Organización del epitelio seminífero
Las asociaciones de las células se desarrollan y permanecen en
una región del túbulo seminífero, por ejemplo, el estado I se convertirá
finalmente en el estado II sin movimientos con respecto al eje
longitudinal del túbulo. Hay distintos ordenamientos de asociaciones
celulares a lo largo de la longitud del túbulo. Perey y col. 1961 han
examinado muy meticulosamente las secciones longitudinales de

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 25
túbulos de rata y describieron el patrón de la espermatogénesis a lo
largo de la longitud del túbulo y comprobaron que los segmentos de
los túbulos están formados de asociaciones de célula solas o estados.
Los segmentos como asociaciones de células son designados por
números romanos, por ejemplo, una porción longitudinal del túbulo
seminífero ocupada por el estado IV es llamado segmento IV. Figura
I.6. (Greep y col. 1975)
Figura I.6 Esquema de una sección longitudinal de un túbulo
seminífero mostrando los segmentos que son designados por
números romanos. La sección proximal es la mas cercana al rete.
Cuando el final de la serie es alcanzado (estado I), la serie comienza a
descender desde el estado XIV. Nótese que la medida de los
segmentos con diferentes designaciones numéricas romanas no son
iguales, como no así los segmentos que poseen la misma designación
numérica romana. (Figura extraída de Russell et al. 1990).
Un segmento es una porción del túbulo seminífero en su eje
longitudinal ocupado por una simple asociación de células.

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 26
La longitud de un segmento en particular es variable y sólo
puede ser expresado como una media de la longitud para una especie
en particular, la longitud media del segmento es proporcionalmente
más larga para estados con mas frecuencia observados.
El desarrollo sincrónico de las células dentro del segmento no es
completamente homogéneo, distintos criterios morfológicos pueden
ser usados para dividir varios estados en subsegmentos; Perey y col.
1961 destaca que el subsegmento es bruscamente demarcado a lo
largo de la longitud del túbulo (figura I.7). (Greep y col. 1975)
Figura I.7. Se muestran los subsegmentos en una sección longitudinal
del segmento XIV. El subsegmento XIVa representa la porción de la
asociación o estado XIV con figuras meióticas en meiosis I (metafase,
anafase y telofase). El subsegmento XIVb representa la porción del
estado XIV con espermatocitos secundarios y el subsegmento XIVc
muestra la porción del estado que contiene las figuras mitóticas en
meiosis II. (Figura extraída de Russell et al. 1990).
En secciones del túbulo seminífero, una asociación celular con
células características de un sub-segmento es llamada sub-estado. Los

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 27
sub-estados son designados con números romanos con un sufijo
alfabético ej. VIIa, XIVb.
Los segmentos están ordenados dentro del túbulo seminífero;
comenzando desde la rete testis (proximal), presentan un orden
descendiente, si por ejemplo el segmento XI es cercano a la rete
testis, entonces más distante a lo largo de túbulo se encontrará
primero el segmento X, IX, VII y VII, etc. Cuando el segmento I es
alcanzado, el siguiente segmento es el XIV, seguido por el XIII y el XII
y así sucesivamente de manera descendiente. Las uniones de los
segmentos consecutivos, son llamadas continuidad de orden
segmentaria. El patrón general de orden descendiente en una dirección
lejana a la rete testis es llamada orden segmentaria descendiente.
Figura I.8. Ocasionalmente las modulaciones ocurren a cortas
distancias a lo largo del túbulo invirtiendo el orden. Sin embargo a una
corta distancia de orden descendiente se observa lo contrario. Figura
I.8 (Russell y col. 1990)
Figura I.8 Una modulación en la pendiente de orden segmental.
Aunque el patrón de orden descendiente es evidente, una corta
porción del túbulo invierte la orden del segmento a un patrón
ascendente. (Figura extraída de Russell et al. 1990).
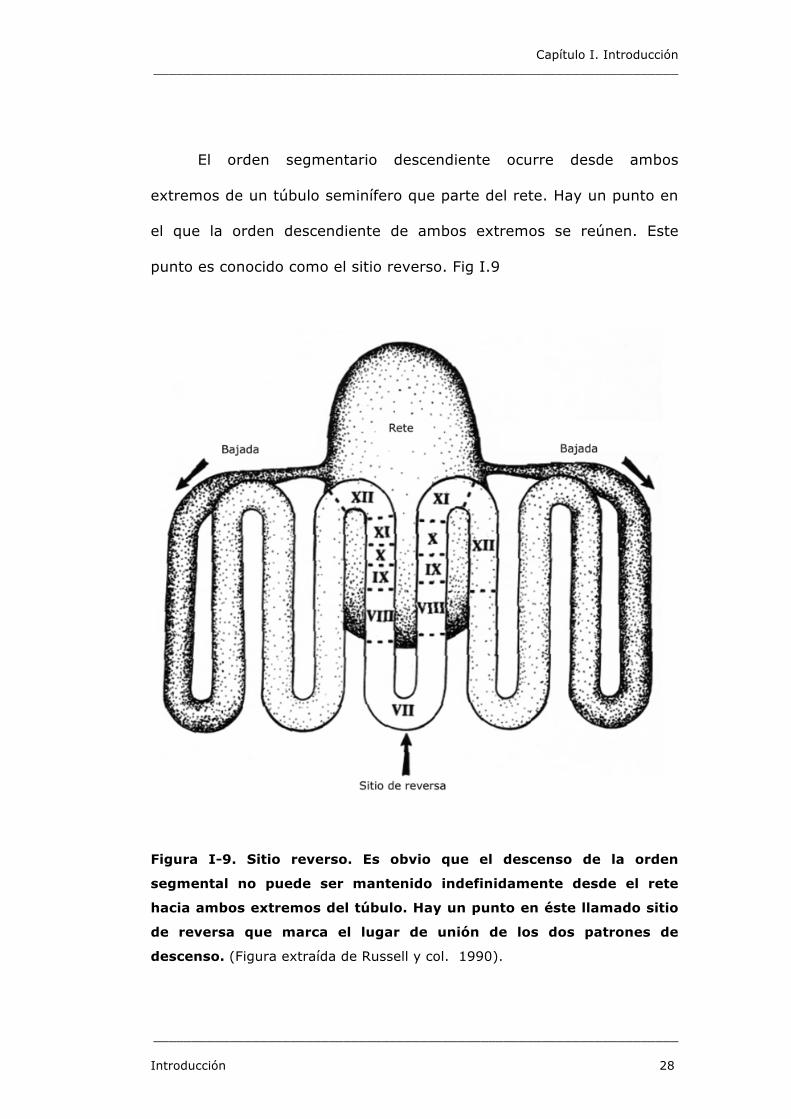
Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 28
El orden segmentario descendiente ocurre desde ambos
extremos de un túbulo seminífero que parte del rete. Hay un punto en
el que la orden descendiente de ambos extremos se reúnen. Este
punto es conocido como el sitio reverso. Fig I.9
Figura I-9. Sitio reverso. Es obvio que el descenso de la orden
segmental no puede ser mantenido indefinidamente desde el rete
hacia ambos extremos del túbulo. Hay un punto en éste llamado sitio
de reversa que marca el lugar de unión de los dos patrones de
descenso. (Figura extraída de Russell y col. 1990).

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 29
El sitio de reversa no es equidistante del rete a ambos lados del
túbulo. Si hay o no modulación, la orden descendiente del segmento
eventualmente guiará a una serie completa de segmentos. Una serie
completa a lo largo de la longitud del túbulo se define como una onda.
Figura I.9.
Una onda del epitelio seminífero es una serie completa de
segmentos, incluyendo algunas modulaciones que ocurren a lo largo de
túbulo.
Muchas ondas (eventualmente cerca de 24) están presentes en
un túbulo seminífero, éstas no son iguales en longitud varían de 1 a 5
cm. (Russell y col. 1990)
1.1.9. Regulación de la contracción del túbulo seminífero
Una ver terminada la espermiogénesis los espermatozoides se
desprenden de las células de Sertoli y son transportados a la rete
testis. En esta función participan en forma sincronizada las células de
Sértoli y las células MP.
El estimulante de la contracción de las células MP en el túbulo
seminífero es el péptido endotelina (ET), que es un potente
vasoconstrictor de origen epitelial secretado por las células de Sertoli
(Fantoni, 1993)
Las células MP de rata expresan dos tipos de receptores de alta
afinidad a ET, (receptores ET-A y ET-B) que controlan la contracción
celular a través de la movilización de calcio intracelular (Tripiciano,
1997).

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 30
Un aspecto central relacionado con el control de la
contractibilidad del túbulo seminífero se relaciona con la producción de
ET por las células de Sértoli.
ET es secretado como un precursor inactivo que es activado en
el espacio pericelular por un clivaje específico de la molécula por la
enzima convertidora de endotelina (ECE) (Opgenorth y col. 1992).
La ECE solo es expresada en las células MP donde en el epitelio
seminífero los espermatozoides han terminado su maduración y están
listos para ser liberados al lumen del túbulo, es decir los estadíos IX-XI
(Palombi, 2002).

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 31
2. CARACTERÍSTICAS DE MIOSINA
2.1. Miosinas
Un gran número de proteínas conocidas como miosinas
constituyen la parte motriz del sistema miosina-microfilamentos. Las
miosinas son proteínas motoras que convierten la energía química en
fuerza mecánica.
2.1. Descubrimiento de Miosina
A finales de siglo XIX, Kühne transformó la teoría de la
contracción muscular cuando observó un nematodo que nadaba
libremente dentro del tejido muscular (Kühne 1863). El concluyó que
el contenido del músculo no era sólido como se había pensado, y en
lugar de eso era una solución concentrada de “albúmina”. Encontró
que soluciones que contenían altas concentraciones salinas extraían
una sustancia del músculo a la que él llamó “miosina” (Kühne 1864).
Engelhardt y Ljubimowa en 1939 descubrieron que la miosina de
Kühne, poseía actividad ATPasa y era la fuente de energía para la
producción de fuerza durante la contracción muscular. Más tarde,
Albert Szent-Gyorgyi en 1942, demostró in vitro la formación de fibras
artificiales de miosinas y de otras proteínas a las que ellos llamaron
“actina” (debido a que activaban a miosina). Desde ese tiempo hasta
la actualidad trabajos bioquímicos y fisiológicos han contribuido al
entendimiento de la estructura básica de la molécula de miosina.
Además, se han encontrado diversas miosinas clasificándolas en dos

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 32
grupos: Miosinas convencionales a aquellas que forman filamentos y
comprenden a la familia de proteínas conocidas como miosinas II.
Miosinas no convencionales a las que no forman filamentos y tienen
una función distinta a la de la contracción muscular y se conocen las
miosinas I y III a XVII (Redowicz, 2007).
2.2. Estructura de la molécula de Miosina II
La molécula de miosina II es uno de los principales componentes
del sistema contráctil y comprende más del 50% de las proteínas
totales. Las moléculas de miosina II se organizan en forma de
filamentos gruesos y se convierten en elementos de transducción de
energía y desarrollo de fuerza. La familia de miosina II incluye a
miosinas del músculo esquelético y liso, de músculo cardíaco y
también de sistemas no musculares. Todas las moléculas de miosina II
tienen algunas propiedades comunes y otras específicas de cada tipo
celular.
La molécula de Miosina II es larga, asimétrica y de un peso
molecular de aproximadamente 450 kDa. Posee dos cabezas
globulares localizadas en el extremo amino (NH2) de la molécula,
seguida de una larga cola semejante a varas entrelazadas.
Cada molécula de miosina II es un hexámero compuesto por dos
cadenas pesadas de aproximadamente 205 kDa cada una, dos cadenas
livianas esenciales de 16 kDa a 20 kDa cada una y dos cadenas
livianas reguladoras de aproximadamente 16 kDa a 20 kDa de peso
molecular cada una. Las cadenas livianas regulatorias ( rLCM ),

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 33
participan en la regulación de la contracción del músculo por el calcio,
y las cadenas livianas esenciales ( eLCM ), regulan la actividad ATPasa
de la molécula. (Lehman, 1978; Szent-Györgyi, 1980; Wagner y
Stone. 1983).
Las cadenas pesadas son asimétricas y cada una contiene una
cabeza globular y una cola semejante a una vara. La porción globular
tiene actividad enzimática (hidroliza ATP) y sitios de interacción con
actina y con las cadenas livianas.
La porción de la cola es la responsable del ensamble de la
molécula en filamentos gruesos. En la Figura I.10 se observa una
molécula de miosina II con sus dominios.
Figura I.10. Características de la molécula de miosina II.
A. Esquema de la proteína mostrando las cadenas y dominios que la
componen.

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 34
B. Aspecto de la proteína observada por micrografías electrónicas
de alta resolución. (Figura extraída de Alberts y col. 2002)
Los dominios peptídicos de miosina II se pueden separar por
tratamientos proteolíticos controlados. La molécula de miosina II
puede digerirse con quimiotripsina y papaína. La digestión con
quimiotripsina (Prince y col. 1981), produce un corte dando dos
fragmentos, uno denominado meromiosina pesada HMM (Heavy Mero-
Myosin) y otro meromiosina liviana LMM (Light Mero-Myosin). La
digestión con papaína (Cohen y col. 1970) del fragmento HMM genera
dos nuevos subfragmentos, S1 (Subfragmento 1) y S2 (Subfragmento
2). S1 comprende la porción globular de la molécula y S2 el cuello. Ver
Figura I.11.
Figura I.11. Clivaje proteolítico de miosina II. El tratamiento de
miosina II con quimiotripsina genera dos fragmentos, llamados
meromiosina pesada (HMM), y meromiosina liviana (LMM). El

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 35
tratamiento del fragmento HMM con papaína genera dos fragmentos
más, S1 constituido por las dos cabezas globulares y S2 por el tallo
restante. (Figura extraída de Lodish y col. 2005)
2.2.1. Estructura de la porción C-terminal de miosina II
La porción carboxilo terminal del LMM de la cadena pesada de
miosina II es el responsable de las propiedades de agregación,
solubilización y forma el núcleo de los filamentos. El fragmento LMM
mide 156 nm de longitud, posee una conformación “coiled coil”, motivo
estructural encontrado en muchas proteínas, constituido por las largas
hélices alfa, con un patrón de aminoácidos hidrofóbicos que se repite.
Los aminoácidos hidrofóbicos se combinan con los de otras moléculas
de miosinas y forman los filamentos gruesos.
La información contenida en la secuencia primaria de las LMM es
importante para entender las interacciones que ocurren entre las
moléculas de miosina adyacentes en el filamento grueso.
La porción LMM tiene una secuencia de aminoácidos
característicos, posee un heptapéptido repetido (-a-b-c-d-e-f-g-),
donde los aminoácidos a y d son hidrofóbicos y forman una interfase
entre las hélices alfa de la proteína plegada. (Parry, 1981; McLachlan;
Ueno y Harrington. 1981 a y b 1981; Stewart M. 1982).
2.3. Estructura de los filamentos gruesos de miosina II
La molécula de miosina II forma dos tipos de filamentos
gruesos, los filamentos bipolares y los de polaridad lateral. Los
filamentos bipolares son característicos de miosina II de músculo

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 36
estriado y de músculo liso. Se forman por la nucleación de moléculas
de miosina II orientadas en forma antiparalela. Las colas se
entrecruzan y forman una región central desprovista de las cabezas,
denominada zona desnuda. (Craig y Megerman. 1977; Hinssen y col.
1978).
Los filamentos de polaridad lateral o side polar son
característicos de miosina II de músculo liso y de células no
musculares, se interpreta que se forman por la unión de dímeros
antiparalelos de miosina II. Cada nuevo dímero que se incorpora al
filamento se une evitando la cabeza de las moléculas vecinas.
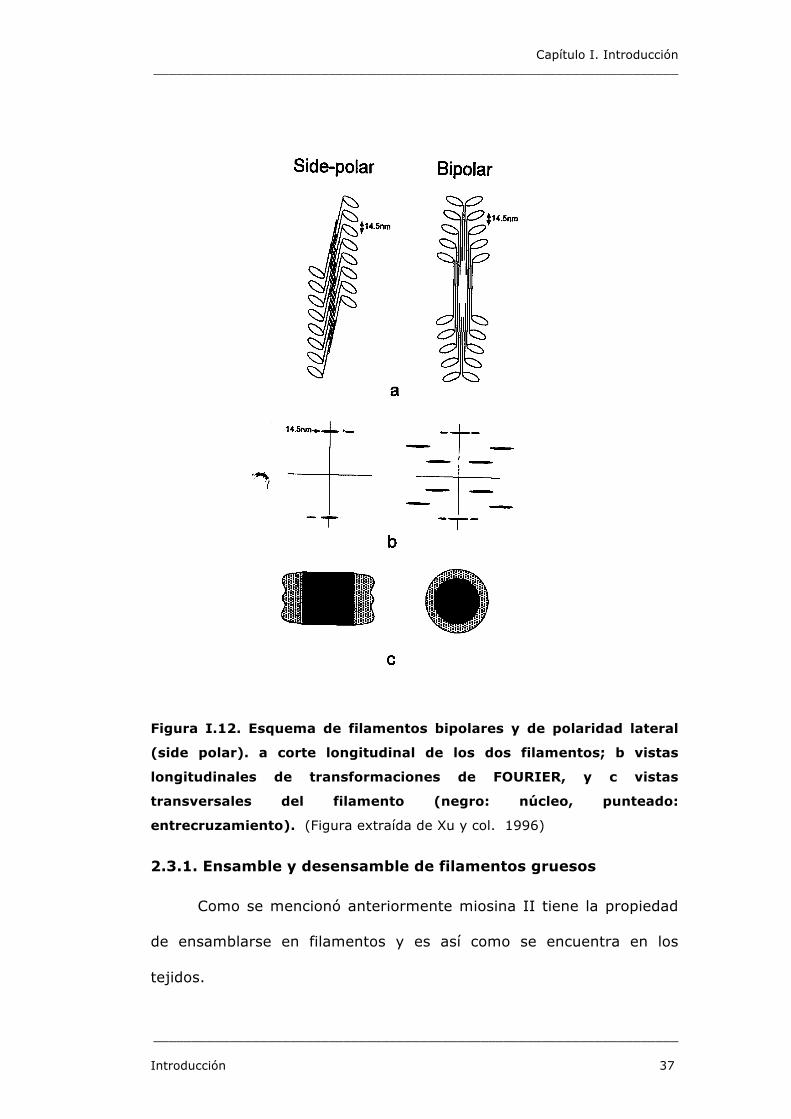
Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 37
Figura I.12. Esquema de filamentos bipolares y de polaridad lateral
(side polar). a corte longitudinal de los dos filamentos; b vistas
longitudinales de transformaciones de FOURIER, y c vistas
transversales del filamento (negro: núcleo, punteado:
entrecruzamiento). (Figura extraída de Xu y col. 1996)
2.3.1. Ensamble y desensamble de filamentos gruesos
Como se mencionó anteriormente miosina II tiene la propiedad
de ensamblarse en filamentos y es así como se encuentra en los
tejidos.

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 38
Los filamentos de todos los tejidos musculares se disocian al ser
expuestos a altas concentraciones salinas, aproximadamente 0,6 M de
NaCl, sugiriendo que las uniones entre las moléculas son
principalmente de naturaleza iónica (Szuchet. 1977; Emes y Rowe.
1978 a y b; Trinick y Cooper, 1980).
Estudios de microscopía electrónica demuestran que el
tratamiento con 160 mM de NaCl provoca el acortamiento de los
filamentos a un tamaño intermedio, con 170 mM de NaCl se obtienen
fragmentos más cortos y con 300 mM de NaCl la completa disociación.
Estos tres estados de desensamble demuestran que existen distintos
grados de empaquetamiento de la molécula de miosina II para formar
el filamento (Trinick y Cooper, 1980).
2.3.2. Implicancia de la fosforilación de las cadenas livianas en
la formación del filamento
La cadena liviana regulatoria de miosina II ( rMLC ), es
fosforilada por una quinasa denominada, quinasa de la cadena liviana
regulatoria (MLCK), y es dependiente del complejo calcio-calmodulina.
Asimismo la cadena liviana es defosforilada por una fosfatasa tipo I
(MYPT 1). (Onishi y col. 1978); (Scholey y col. 1980).
Cuando la cadena liviana está fosforilada, miosina II adquiere un
estado conformacional conocido como 6 S. Estudios por microscopía
electrónica demuestran que en estas condiciones la molécula tiene la
cola o porción helicoidal desplegada.

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 39
Si la cadena liviana regulatoria se defosforila, miosina II cambia su
conformación a un estado de 11 S y la cola de la molécula se pliega.
(Suzuki, y col. 1978; Trybus y col. 1982; Kendrick-Jones y col. 1982;
a, b y c).
La condición natural de miosina II en el tejido es en estado 6S,
es decir los sitios hidrofóbicos de las colas están expuestos y se
combinan en forma antiparalela con otras moléculas de miosina II.
(Craig y col. 1983).
Cuando miosina II se encuentra en estado 11 S, la molécula es incapaz
de formar filamentos y permanece soluble. Ver Figura I.11.
Figura I.13. Ensamblaje controlado de miosina II. ( A ) La
fosforilación controlada de una de las dos cadenas ligeras tiene al
menos dos efectos in vitro:provoca un cambio en la conformación de
la cabeza de miosina, exponiendo su lugar de unión a la actina, y
libera la cola de miosina de la “zona pegajosa” de la cabeza,
permitiendo que la molécula se ensamble formando cortos filamentos
bipolares. La enzima responsable de esta fosforilación es la quinasa
de la cadenaligera de la miosina. ( B ) Tinción negativa de filamentos
cortos de miosina II quehan sido inducidos a ensamblarse por

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 40
fosforilación de sus cadenas ligeras. (Figura extraída de Alberts y col.
2002)
2.3.3. Implicancia de la fosforilación de la cadena pesada en la
formación de filamentos de miosina II
El fragmento LMM de miosinas de músculo liso y de células no
musculares, es fosforilado en múltiples serinas cerca del extremo
carboxilo terminal por la proteína quinasa C (PKC) y la caseína quinasa
II, (Bresnick, 1999).
Las fosforilaciones en la porción LMM inhiben el ensamble de miosina II
en filamentos. Ver figura I.14.
Figura I.14. Secuencia de aminoácidos del extremo carboxilo terminal
de dos isoformas de miosina II. Los sitios marcados con asteriscos
son fosforilados por la PKC, y las puntas de flecha indican los sitios
fosforilados por la caseína quinasa II. (Figura extraída de Bresnick 1999)

Capítulo I. Introducción _____________________________________________________________________
_____________________________________________________________________ Introducción 41
3. PROPÓSITO DEL TRABAJO DE TESIS
El túbulo seminífero es una estructura compleja que contiene los
distintos tipos celulares que participan en la formación de los
espermatozoides y su posterior traslado a la rete testis. Es de nuestro
interés estudiar las características contráctiles de las células mioides
peritubulares del túbulo seminífero en el testículo de rata en desarrollo
y adulto.
Se aislará miosina de las células mioides peritubulares y se
analizará: la estructura peptídica, la solubilidad en medios de baja y
alta fuerza iónica y la participación en el proceso de contracción.
Se desarrollará un método de tinción indispensable para medir
los parámetros de las células MP en túbulos seminíferos aislados tanto
en reposo como estimulados a contraerse.
Por último se estudiará la relación de las características
morfológicas de las células MP con la expresión de isotipos de
miosina-CMP en túbulos seminíferos del testículo en desarrollo y en los
segmentos conteniendo los distintos estados del epitelio seminífero.

CAPÍTULO II
CARACTERÍSTICAS DE LA MOLÉCULA DE MIOSINA DE CÉLULAS
MIOIDES PERITUBULARES DEL TESTÍCULO

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 43
CARACTERÍSTICAS DE LA MOLÉCULA DE MIOSINA DE CÉLULAS
MIOIDES PERITUBULARES DEL TESTÍCULO
Introducción:
En este capítulo se analizará las característica de miosina de las
células mioides peritubulares del testículo, se determinará las
propiedades de la proteína tanto en su estructura peptídica como en la
solubilidad en medios de baja y alta fuerza iónica.
1. AISLACIÓN Y PURIFICACIÓN DE MIOSINA PRESENTE EN
LOS TÚBULOS SEMINÍFEROS
1.1. Contenido de miosina en el túbulo seminífero de testículo
Cuando se homogenizan testículos de rata desprovistos de la
túnica albugínea, en soluciones de baja fuerza iónica, se comprueba la
existencia de una proteína de 205 kDa. (Figura II. 1)

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 44
Figura II.1. Proteínas de un homogeneizado de testículo. SDS-PAGE
7,5% teñido con azul de Coomassie. C+: miosina de músculo liso purificada
parcialmente; Homogeneizado: homogeneizado de testículos de rata
sembrados en dos concentraciones. Las flechas en el interior de las figuras
indican la posición de 205 kDa. Las flechas a la izquierda de la figura indican
la posición de las proteínas de peso molecular conocido.
Cuando el homogeneizado se centrifuga a 100.000 xg por 60 minutos,
se comprueba la existencia de una proteína de 205 kDa (que
denominamos proteína 205) que se encuentra en su totalidad en el
sobrenadante. Si el sobrenadante es incubado 30 minutos a 20° C, la
proteína 205 se agrega y es posible sedimentarla por centrifugación.

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 45
La proteína 205 sedimentada puede ser nuevamente solubilizada por la
incubación con 4 mM ATP (Sobrenadante ATP). (Ver figura II.2).
Figura II 2. Solubilización de la proteína 205 por ATP. Proteína de 205
kDa en el sobrenadante ATP. SDS PAGE 7,5% teñido con azul de Coomassie
Se muestran las proteínas de tres experimentos. Las flechas indican la
posición de las proteínas de peso molecular conocido (PM).
Debido a que la proteína 205 tiene la misma movilidad
electroforética que la cadena pesada de miosina de músculo liso, se
procedió a analizar por inmunoensayos la capacidad de un anticuerpo

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 46
monoclonal anti-miosina II de músculo liso de aorta de reconocer a la
proteína. Para su comparación se utilizaron miosinas de músculo liso
de varios tejidos. Como se muestra en la figura II.3, el anticuerpo
reconoce a la proteína 205, de la misma manera que a otras miosinas
de músculo liso conocidas.
Actividad ATPasa: Se ha comprobado que la proteína de 205
kDa aislada en testículo de rata, es reconocida por un anticuerpo
monoclonal anti-miosina de músculo liso. Llama la atención que la
proteína 205 se encuentre soluble en medios de baja fuerza iónica,
siendo que las proteínas miosinas de músculo liso no son solubles en
esas condiciones. Para asegurarse de que se trata de miosina, la
proteína 205 fue sometida a un ensayo de actividad ATPasa. Se conoce
que solo las proteínas de la familia de las miosinas hidrolizan ATP en
medios de alta concentración de KCl (0,6 M) y EDTA (medio EDTA-K).
Los ensayos indicaron que las fracciones ricas en la proteína 205 kDa,
hidrolizaron ATP a 0,8 ± 0.01 µM ATP/min/mg de proteína en
coincidencia con la hidrólisis de ATP en medio EDTA-K de miosina de
músculo liso de estómago de pollo (McKenna y col. 1989).
Debido a que la proteína 205 coincide con la movilidad relativa
de la cadena pesada de miosina de músculo liso, es reconocida por un
anticuerpo anti-miosina de músculo liso y además fracciones
enriquecidas de la proteína hidrolizan ATP en medio EDTA-K,
mostrando una actividad de ATPasa propia de las proteínas miosina, se

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 47
procedió a denominar a la proteína 205 como miosina -CMP. (Más
adelante en el texto se explicará el motivo de la denominación).
Figura II.3. Reconocimiento de la proteína 205, con anticuerpo anti-
miosina II de músculo liso. A. SDS-PAGE 5% teñido con azul de
Coomassie. B. Inmunoensayo del gel. 1 y 2 citosol de testículo de rata en dos
concentraciones; 3-6, miosina de tejidos de rata:3 y 4: de aorta (edad
perinatal y adulta respectivamente); 5: útero, 6, cordón umbilical; 7 proteína
205 concentrada por agregación-desagregación. Las flechas indican las
posiciones de miosinas reconocidas en las bandas del gel y en la lámina de
nitrocelulosa.
1.2. Purificación de miosina-CMP
1.2.1. Método agregación-desagregación: En base a que miosina-
CMP está soluble en el citosol, se agrega a 20° C y se resuspende con
ATP, se desarrolló un método sencillo para poder aislarla. (Ver figura
II.4).
El método que llamamos de agregación-desagregación permite
obtener una fracción enriquecida de miosina-CMP, sin utilizar columnas

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 48
cromatográficas. Como se observa en la figura II.5, por este método
se obtienes muestras donde aproximadamente el 70 % de las
proteínas corresponde a miosina-CMP.
Figura II.4. Esquema del método de agregación-desagregación de
miosina-CPM
Diagrama esquemático del método utilizado para aislar miosina-CMP de
testículo de rata y toro.

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 49
Figura II.5. Aislamiento de miosina-CMP por el método de agregación-
desagregación. SDS-PAGE 7,5 % teñido con azul de coomassie. La flecha
indica la posición de miosina-CMP.
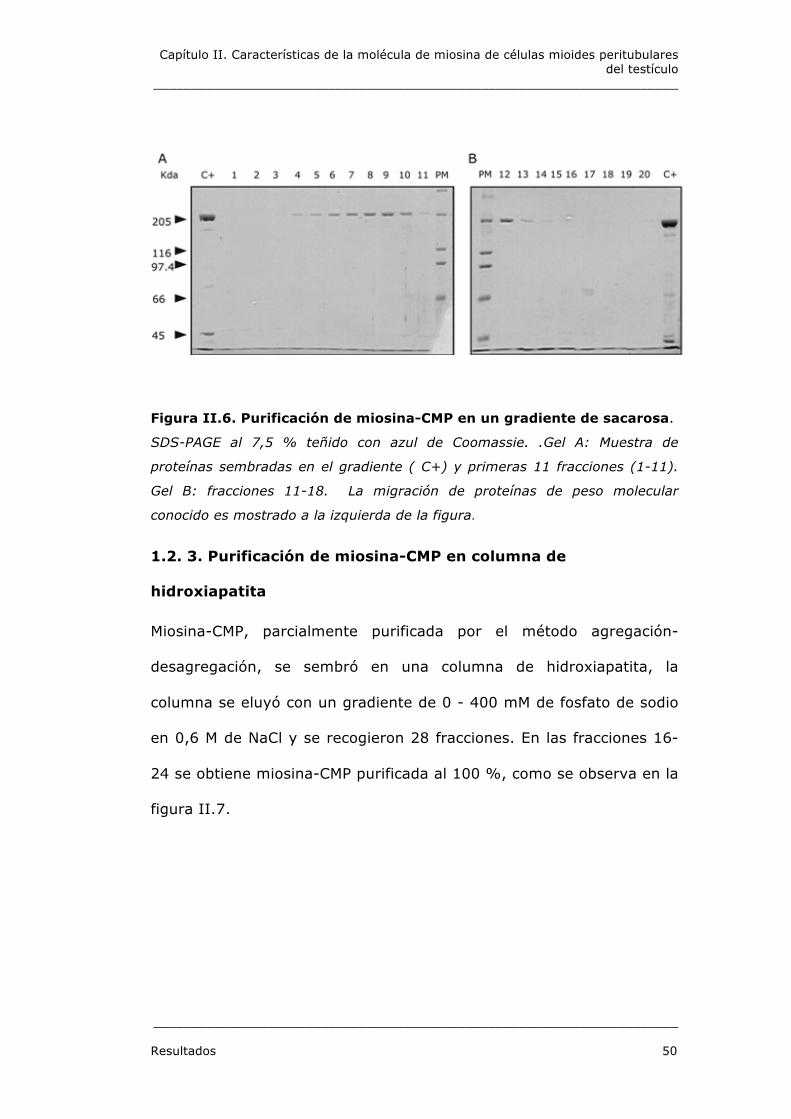
1.2. 2. Purificación de miosina-CMP en gradiente de sacarosa
Una vez puesta a punto la técnica para obtener una fracción
enriquecida de miosina-CMP, se utilizó esta preparación para purificar
la proteína. Para ello se carga la muestra en un gradiente de sacarosa
(5 %-20 %). El gradiente se centrifuga a 100.000 xg, 18 horas, 4 °C.
Se colectan 18 fracciones, que se numeran desde el fondo del tubo.
Las fracciones 7 a 11 contienen miosina-CMP altamente purificada,
aunque perdura una pequeña contaminación con proteínas de bajo
peso molecular. Ver figuras II.6.
Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 50
Figura II.6. Purificación de miosina-CMP en un gradiente de sacarosa.
SDS-PAGE al 7,5 % teñido con azul de Coomassie. .Gel A: Muestra de
proteínas sembradas en el gradiente ( C+) y primeras 11 fracciones (1-11).
Gel B: fracciones 11-18. La migración de proteínas de peso molecular
conocido es mostrado a la izquierda de la figura.
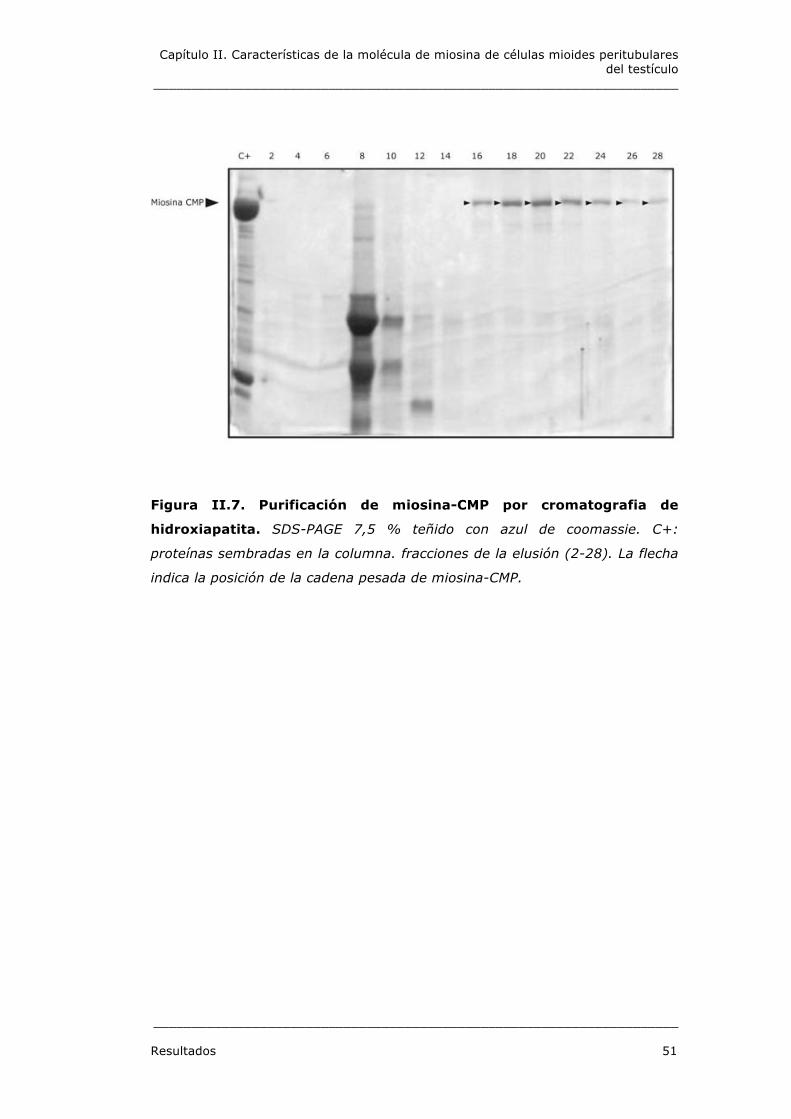
1.2. 3. Purificación de miosina-CMP en columna de
hidroxiapatita
Miosina-CMP, parcialmente purificada por el método agregación-
desagregación, se sembró en una columna de hidroxiapatita, la
columna se eluyó con un gradiente de 0 - 400 mM de fosfato de sodio
en 0,6 M de NaCl y se recogieron 28 fracciones. En las fracciones 16-
24 se obtiene miosina-CMP purificada al 100 %, como se observa en la
figura II.7.
Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 51
Figura II.7. Purificación de miosina-CMP por cromatografia de
hidroxiapatita. SDS-PAGE 7,5 % teñido con azul de coomassie. C+:
proteínas sembradas en la columna. fracciones de la elusión (2-28). La flecha
indica la posición de la cadena pesada de miosina-CMP.

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 52
2. CARACTERÍSTICAS PEPTÍDICAS DE LA CADENA PESADA DE
MIOSINA-CPM
2.1. Digestión Enzimática
Para conocer la composición peptídica de miosina-CMP se
procedió a realizar cortes de la proteína con tripsina a distintos
tiempos. Miosinas de músculo liso de la túnica albugínea y la arteria
espermática del testículo fueron digeridas en paralelo. Los péptidos de
la digestión, fueron analizados en un inmunoblotting con un anticuerpo
antimiosina de músculo liso (Figura II.8)
Figura II.8. Digestión de miosina con tripsina. Miosina de arteria
espermática (Art. Esp.), túnica Albuguinea (Tunica Alb.) y túbulos seminíferos
(miosina cmp) fueron digeridas con tripsina 20 y 40 min a 20 °C. A: SDS-
PAGE 7,5 % teñido con azul de Coomassie , B: inmunoensayo del gel con
anti-miosina de músculo liso. Las cabezas de flechas a la derecha de la figura
indican los péptidos producidos por la digestión de miosina CMP. La migración
de proteínas de peso molecular conocido es mostrado a la izquierda de la
figura.

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 53
La digestión con tripsina mostró que se producían péptidos de
distinto peso molecular de miosina-CMP con respecto a la miosina de
músculo liso de la túnica albugínea y de la arteria espermática Figura
II.8).
2.2. Espectrometría de masas
Debido a que miosina-CMP presenta un patrón de péptidos
distintos a los de otras proteínas de músculo liso (ver figura II.8), se
procedió a analizar la huella peptídica por espectrometría de masas.
Bandas de un gel al 7,5 % de la cadena pesada de miosina-CMP
de rata y toro, teñidas con azul de Coomassie fueron escindidas, y
analizadas por Maldi-TOF (MALDI: Matrix-Assisted Laser
Desorption/Ionization, desorción/ionización mediante láser asistida por
matriz). En la ionización, los analitos cocristalizados con una matriz
apropiada son convertidos en iones mediante la acción de un láser.
Esta fuente de ionización suele asociarse a un analizador de tiempo de
vuelo (TOF: Time-Of-Flight) en el que los iones se separan en función
de su relación masa-carga tras ser acelerados en un campo eléctrico, y
de allí se obtiene la huella peptídica de las masas de la proteína. La
comparación de los péptidos obtenidos fueron analizados utilizando el
programa MASCOT de Matrix science. Se determinó la búsqueda
considerando que miosina-CMP había sido digerida con tripsina y había
sido modificada por carboximetilación.
2.2.1. Miosina-CMP de testículo de rata: En un análisis de
amplio espectro se comprobó que con un “score” significativo de 95 %,

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 54
de los 65 péptidos de miosina-CMP analizados, 27 de ellos coincidían
con miosina de músculo liso de rata Rattus norvegicus (número de
acceso al banco de datos gi|34868040). Estos resultados indicaban
que los péptidos de miosina CMP que coincidían con los de músculo liso
de rata representaban el 18% del total de la proteína. (Ver figura II.9)
1 MAQKGQLSDD EKFLFVDKNF INSPMAQADW VAKKLVWVPS EKQGFEAASI
51 KEEKGDEVVV ELVENGKKVT VGKDDIQKMN PPKFSKVEDM AELTCLNEAS
101 VLHNLRERYF SGLIYTYSGL FCVVVNPYKH LPIYSEKIVD MYKGKKRHEM
151 PPHIYAIADT AYRSMLQDRE DQSILCTGES GAGKTENTKK VIQYLAVVAS
201 SHKGKKDSSI TGELEKQLLQ ANPILEAFGN AKTVKNDNSS RFGKFIRINF
251 DVTGYIVDLL EKSRAIRQAR DERTFHIFYY LIAGAKEKMR NDLLLESFNS
301 YTFLSNGFVP IPAAQDDEMF QETLEAMSIM GFSEEEQLAI LKVVSSVLQL
351 GNIVFKKERN TDQASMPDNT AAQKVCHLVG INVTDFTRAI LTPRIKVGRD
401 VVQKAQTKEQ ADFAIEALAK ATYERLFRWI LSRVNKALDK THRQGASFLG
451 ILDIAGFEIF EVNSFEQLCI NYTNEKLQQL FNHTMFILEQ EEYQREGIEW
501 NFIDFGLDLQ PCIELIERPN NPPGVLALLD EECWFPKATD KSFVEKLCSE
551 QGNHPKFQKP KQLKDKTEFS IIHYAGKVDY NASAWLTKNM DPLNDNVTSL
601 LNASSDKFVA DLWKDVDRIV GLDQMAKMTE SSLPSASKTK KGMFRTVGQL
651 YKEQLGKLMT TLRNTTPNFV RCIIPNHEKR SGKLDAFLVL EQLRCNGVLE
701 GIRICRQGFP NRIVFQEFRQ RYEILAANAI PKGFMDGKQA CILMIKALEL
751 DPNLYRIGQS KIFFRTGVLA HLEEERDLKI TDVIMAFQAM CRGYLARKAF
801 TKRQQQLTAM KVIQRNCAAY LKLRNWQWWR LFTKVKPLLQ VTRQEEEMQA
851 KEEEMQKIKE RQQKAESELK ELEQRHTQLA EEKTLLQEQL QAETELYAEA
901 EEMRVRLAAK KQELEEILHE MEARLEEEED RSQQLQAERK KMAQQMLKEE
951 KSRQELEKLK RKLEGDASDF HEQIADLQAQ IAELKMQLAK KEEELQAALA
1001 RLDEEITQKN NALKKIRELE GHVSDLQEDL DSERAARNKA EKQKRDLGEE

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 55
1051 LEALKTELED TLDSTATQQE LRAKREQEVT MLKKALDEET RSHEAQVQEM
1101 RQKHTQAVEE LTEQLEQFKR AKANLDKSKQ TLEKENADLA GELRVLGQAK
1151 QEVEHKKKKL EGQLQELQSK CSDGERARTE LSDKVHKLQN EVESVTGMLN
1201 EAEGKAIKLA KEVASLGSQL QDTQELLQEE TRQKLNVSTK LRQLEDERNS
1251 LQDQLDEEME AKQNLERHVS TLNIQGPYGS PLGLCGLFVQ LFPLLLDFLS
1301 WRKSVHFSGF HSWPLQLSDS KKKLQDLAST IEVMEEGKKR LQKEMEGLGQ
1351 QYEEKAAAYD KLEKTKNRLQ QELDDLVVDL DNQRQLVSNL EKKQKKFDQL
1401 LAEEKNISSK YADERDRAEA EAREKETKAL SLARALEEAL EAKEELERTN
1451 KMLKAEMEDL VSSKDDVGKN VHELEKSKRA LETQMEEMRT QLEELEDELQ
1501 ATEDAKLRLE VNMQALKGQF ERDLQARDEQ NEEKRRQLQR QLHEYETELE
1551 DERKQRALAA AAKKKLEGDL KDLELQADSA VKGREEAIKQ LRKLQAQMKD
1601 FQRELDDARA SRDEIFATSK ENEKKAKSLE AELMQLQEDL AAAERARKQA
1651 DLEKEELAEE LASSLSGRNT LQDEKRRLEA RIAQLEEELE EEQGNMEAMS
1701 DRVRKATLQA EQLSNELVTE RSAAQKNESA RQQLERQNKE LRSKLQEVEG
1751 AVKAKLKSTV AALEAKIVQL EEQIEQEARE KQAATKLLKQ KDKKLKEVLL
1801 QVEDERKMVE QYKEQAEKGN TKVKQLKRQL EEAEEESQRI NANRRKLQRE
1851 LDEATESNEA MGREVNALKS KLRRGNEASF VPSRRAGGRR VIENTDGSEE
1901 EMDARDSDFN GTKASE
Figura II.9. Comparación de los péptidos de miosina-CMP de rata.
Secuencia peptídica de miosina de músculo liso de rata (Rattus norvegicus
gi|3486804). Se muestra el resultado del análisis de 65 peptidos de miosina-
CMP, de los cuales, 27 (resaltados en rojo) coincidieron con los de miosina de
musculo liso y cubrieron el 18% de la proteína.

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 56
1 MSLWLTALES LQGSALSGTM QDRTQRQSLP TGQTDETDTQ GTKQVWILVP
51 VFLVRTVSCL RGGAWQSFQV ALLQRPCSFD NVAPESGDQM HVLRPHCGLW
101 DGEEGHHLLG GDEAREKNLF LAALLEEKVG EEVQRKRRIL TCRVWSVGSW
151 ASPESLRARF GGGTTGVLYL FGKLTGRLAC PRPDPKSGEK EEREGHVWKV
201 RVSPRCPRQA SLRPVAGAVS FPRLRAVCFQ WECPRPEVPD GPLLPLLGTS
251 YREDQSILCT GESGAGKTEN TKKVIQYLAV VASSHKGKKD TSITQIGGLE
301 TASGVTAPAI AGAVTETARL EALGGLGQAP LVAGGALLPA TDAGNPAFPS
351 RLWHNVLEHT ILRLSLRGSE RGFSLRRLSL RLMALLWHAS LPLLPAVIPE
401 FLPRALAPAH RCVLVHIPTD PSYGNFTRAQ DFHGGAPSEG AGGLWTASAA
451 FNLMNGAQAT CFEALFCSSV KTIPDVNKFQ GELEKQLLQA NPILEAFGNA
501 KTVKNDNSSR FGKFIRINFD VTGYIVGANI ETYLLEKSRA IRQARDERTF
551 HIFYYLIAGA KDKMKNDLLL EGFNNYTFLS NGFVPIPAAQ DDEMFQETVE
601 AMAIMGFSEE EQLSAQKVCH LMGINVTDFT RSILTPRIKV GRDVVQKAQT
651 KEQADFAIEA LAKATYERLF RWILSRVNKA LDKTHRQGAS FLGILDIAGF
701 EIFEDAHRAS PVSHTQVNSF EQLCINYTNE KLQQLFNHTM FILEQEEYQR
751 EGIEWNFIDF GLDLQPCIEL IERPNNPPGV LALLDEECWF PKATDKSFVE
801 KLCTEQGNHP KFQKPKQLKD KTEFSIIHYA GKVDYNASAW LTKNMDPLND
851 NVTSLLNASS DKFVADLWKD VDRIVGLDQM AKMTESSLPS ASKTKKGMFR
901 TVGQLYKEQL GKLMTTLRNT TPNFVRCIIP NHEKRSGKLD AFLVLEQLRC
951 NGVLEGIRIC RQGFPNRIVF QEFRQRYEIL AANAIPKGFM DGKQACILMI
1001 KALELDPNLY RIGQSKIFFR TGVLAHLEEE RDLKITDVIM AFQAMCRGYL
1051 ARKAFAKRQQ QLTAMKVIQR NCAAYLKLRN WQWWRLFTKP VCTRGLRLNW
1101 SWHQFFGPEK GPLLVRHVKP LLQVTRQEEE MQAKEDELQK TKERQQKAEN
1151 ELKELEQKHT QLAEEKNLLQ EQLQAETELY AEAEEMRVRL AAKKQELEEI
1201 LHEMEARLEE EEDRGQQLQA EKKKMAQQML DLEEQLEEEE AARQKLQLEK
1251 VTAEAKIKKL EDDILVMDDQ NNKLSKEGLS SGRLSGGLHL LRGPGFVAVR
1301 LKKEEKSRQE LEKLKRKLEG EASDFHEQIA DLQAQIAELK MQLAKKEEEL
1351 QAALGRPSLV YTCARSPFHW PLKHHLPEAG TTLLPLLRRV FFVNASLPFT

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 57
1401 LLLALRPPPG CSGPHTPRAL RNLSPHHPCV LQDGARVTGQ NHHLGCEPAW
1451 KTLLQSKRPA PPLDDEIAQK NNALKKIREL EGHISDLQED LDSERAARNK
1501 AEKQKRDLGE ELEALKTELE DTLDSTATQQ ELRAKREQEV TVLKKALDEE
1551 TRSHEAQVQE MRQKHTQAVE ELTEQLEQFK RAKANLDKNK QTLEKENADL
1601 AGELRVLNQA KQEVEHKKKK LEVQLQELQS KCSDGERARA ELNDKVHKLQ
1651 VTHFLWEPRN CRDQPHHGSG ISQSVDSHKT YHNEVESVTG MLNEAEGKAI
1701 KLAKDVASLG SQLQDTQELL QEETRQKLNV STKLRQLEDE RNSLQDQLDE
1751 EMEAKQNLER HISTLNIQLS DSKKKLQDFA STVEALEEGK KKFQKEIEGL
1801 TQQYEEKAAA YDKLEKTKNR LQQELDDLVV DLDNQRQLVS NLEKKQKKFD
1851 QLLAEEKSIS SKYADERDRA EAEAREKETK ALSLARALEE ALEAKEELER
1901 TNKMLKAEME DLVSSKDDVG KNVHELEKSK RALETQMEEM KTQLEELEDE
1951 LQATEDAKLR LEVNMQALKG QFERDLQARD EQNEEKRRQL QRQLHEYETE
2001 LEDERKQRAL AAAAKKKLEG DLKDLELQAD SAIKGREEAI KQLRKLQAQM
2051 KDFQRELEDA RASRDEIFAT AKENEKKAKS LEADLMQLQE DLAAAERARK
2101 QADLEKEELA EELASSVSGR NTLQDEKRRL EARIAQLEEE LEEEQGNMEA
2151 MSDRVRKATQ QAEQLNNELA TERSAAQKNE SARQQLERQN KELRSKLQEM
2201 EGAVKSKFKS TIAALEAKIA QLEEQVEQEA REKQATAKSL KQKDKKLKEV
2251 LLQVEDERKM AEQYKEQAEK GNAKVKQLKR QLEEAEEESQ RINANRRKLQ
2301 RELDEATESN EAMGREVNAL KSKLRRGNET AFVPSRRSGG RRVIENADGS
2351 DEEVDARDAD FNGTKASE
Figura II.10 Comparacion de los peptidos de miosina-CMP de toro.
Secuencia peptídica de miosina de músculo liso Myh11 de perro (Canis
familiaris gi|57088007). Se muestra el resultado del análisis de 65 peptidos
de miosina CMP de toro, de los cuales, 24 (resaltados en rojo) coinciden con
los de miosina Myh 11 y cubrieron el 13% de la proteína.

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 58
Además, los análisis de espestrometría de masas indicaron que
diez péptidos con masas entre 855 y 1277 no coincidían con los
peptidos de ninguna proteína conocida. (Ver tabla I).
2.2.2. Miosina-CMP de testículo de toro: Debido a que se
habían aislado y purificado miosina-CMP de testículo de toro, se
procedió también a analizar por Maldi-Tof. Utilizando el mismo
programa y el mismo banco de datos que para miosina CMP de rata, se
comprobó que con un score significativo de 81 %, de los 65 peptidos
analizados, 24 de ellos coincidían con miosina de músculo liso de
perro, Myh 11 Canis familiaris (número de acceso al banco de datos:
gi|57088007). Estos resultados indicaban que los peptidos de miosina-
CMP que coincidían con la proteína Myh 11 y que representaban el
13% del total de la proteína. (Ver figura 10).
Coincidentemente con miosina-CMP de rata, miosina-CMP de
toro presenta siete péptidos con masas entre 855 y 1670 que no
coinciden con los peptidos de ninguna proteína conocida. (Ver tabla
II.I).
Tabla II. I. Masa estimada de péptidos específicos de miosina-
CMP

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 59
MASA DE LOS PÉPTIDOS
MIOSINA-CMP DE RATA
MIOSINA-CMP DE TORO
1. 855,09
2. 861,10
3. 877,05
4. 1050,11
5. 1066,09
6. 1072,10
7. 1277,10
8. 1552,75
9. 1624.84
10. 1706.87
1. 855,09
2. 861,05
3. 864,20
4. 877,00
5. 1034,52
6. 1615,79
7. 1670,85

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 60
3. PROPIEDADES DE SOLUBILIDAD DE MIOSINA-CMP
3.1. Solubilidad in vitro
Como se mencionó anteriormente, miosina-CMP se encuentra
soluble cuando se homogeniza el testículo, de todas maneras no
conocemos la relación de miosina-CMP soluble y agregada. Para
determinar el porcentaje de miosina-CMP soluble realizamos el
siguiente ensayo. Homogenizamos túbulos seminíferos con baja y alta
fuerza iónica, purificamos miosina-CMP por el método de agregación-
desagregación y ulterior centrifugado sobre 20 % de sacarosa.
Determinamos la concentracion de miosina-CMP en S3. Como se indica
en la figura II.11, fue similar la cantidad de miosina-CMP obtenida con
y sin fuerza iónica. La cuantificación de las bandas de miosina-CMP por
comparación de bandas conocidas de albúmina indicó que se habían
extraído 14,8 ± 1,9 µg/g de miosina-CMP por el uso del medio sin
fuerza iónica y 15,8 ± 2,2 µg/g de miosina-CMP por el uso de fuerza
iónica (0,6 M NaCl). Estos resultados confirman que la mayoría de
miosina-CMP se encuentra soluble en el citoplasma de la célula.

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 61
Figura II.11. Extracción de miosina-CMP de testículo de rata con y sin
fuerza iónica. 5µl y 10 µl de S3 de la extracción de miosina-CMP sin fuerza
iónica (a-b) y con fuerza iónica (c-d). 1, 2 y 3 µg de ASB (e-g). La figura
muestra un gel de poliacrilamida al 7,5 %, teñido con azul de Coomassie.
La posición de miosina-CMP es mostrada a la izquierda de la figura y la de
albúmina (66 Kda) a la derecha.
3.1.1. Solubilidad de miosina de otros tejidos: Hemos
demostrado que miosina-CMP es una proteína que se asemeja a
miosina II de músculo liso, pero llama la atención que se encuentre
soluble en el citosol.
Nos preguntamos si existe miosina soluble en otros tejidos del
testículo. Para ello se comparó la solubilidad de miosina-CMP de los
túbulos seminíferos con la solubilidad de miosina de la arteria
seminífera y de la túnica albugínea. Los tres tejidos se trataron en
forma idéntica. Se homogenizaron con buffer PM, se centrifugaron, se
separó el sobrenadante (S1) y el sedimento se resuspendió con 0,65 M

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 62
de NaCl en PM, se centrifugó y se obtuvo un sobrenadante (S2). Al
analizar el contenido de miosina en S1 y S2 se observó que solo
miosina-CMP del túbulo seminífero se encontraba soluble en S1,
mientras que miosina de músculo liso de la arteria espermática y de la
túnica albugínea se encontraba en S2. Por la tanto en las condiciones
utilizadas, la miosina II de músculo liso se encuentra agregada
formando filamentos, como ya ha sido demostrado en numerosos
ensayos, en contraposición a miosina-CMP. Ver figura II.12.
Figura II.12. Grado de solubilidad de miosina de distintos tejidos.
Sobrenadante (S1) y sedimento resuspendido con fuerza iónica (S2) de
túbulos seminíferos (A), arteria espermática (B) y túnica albugínea (C). SDS-
PAGE al 7,5 % teñido con azul de coomassie (coomassie blue) y
reconocimiento de miosina de músculo liso en el blotting (anti-SMM II). La
migración de proteínas de peso molecular conocido es indicada a la izquierda
de la figura.

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 63
Estos resultados indican que si se trata de filamentos de miosina
II de músculo liso las condiciones de homogeneización con buffer PM
no alteran su estabilidad. Por lo tanto, en este momento se puede
inferir que la molécula de miosina-CMP tiene alguna diferencia
estructural que le impide formar filamentos en medios de baja fuerza
iónica.
3.2. Agregación in vitro
Debido a que hemos comprobado que más del 90 % de miosina-
CMP se encuentra soluble en los túbulos seminíferos, nos intereso
analizar el comportamiento que presenta in vitro cuando se la expone
a soluciones de distinta fuerza iónica. Además se observó el
comportamiento de miosina II de músculo liso y estriado. Como se ha
comprobado en otros laboratorios, a 4° C, miosina II de músculo liso y
estriado, dializadas en una solución sin NaCl, están completamente
agregadas. Mientras que en solución 100 mM de NaCl, 80 % de
miosina II de músculo estriado y el 79 % de miosina II de músculo liso
permanecen agregadas respectivamente. Con 150 mM y 300 mM, el
50 % y el 40 % de miosina II de músculo estriado permanecen
agregadas respectivamente (figura II.13)
Cuando se determina el grado de solubilidad de miosina-CMP se
comprueba que en diálisis con 0 mM de NaCl sólo el 10 % esta
agregada y ese porcentaje se mantiene con las otras soluciones de
NaCl probadas ( 100 - 300 mM ) (ver la figura II.13). El mismo grado
de solubilidad presenta miosina-CMP de toro sometida a las mismas

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 64
concentraciones de NaCl. Es decir, miosina-CMP tiene un alto grado de
solubilidad en soluciones de baja fuerza iónica a 4° C en contraste con
el comportamiento de miosina II de músculo liso y estriado. (Ver figura
II.13).
Figura II.13. Ensayo de agregación de distintos tipos de Miosina a
4°C. Soluciones conteniendo miosina-CMP de rata y toro, miosina II de
músculo estriado y liso, de aproximadamente 150 µg/ml fueron dializadas
por 12 horas en contra de 0-300 mM de NaCl a 4ºC. En el gráfico se muestra
el porcentaje soluble de cada tipo de miosina en las distintas concentraciones
de NaCl. Los datos presentados corresponden al promedio de dos
experimentos y la dispersión no es mayor a un 5 %.

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 65
Debido a que nosotros teníamos experiencia de que a 20 °C miosina-
CMP se agrega, repetimos el experimento anterior utilizando las
mismas soluciones de diálisis, pero efectuando el ensayo a 20° C.
(figura II. 14). Diálisis con 0 y 50 mM de NaCl muestra que el 100 %
de miosina-T está agregada, de la misma forma que miosina II de
músculo liso y estriado. Con diálisis de 100 mM de NaCl el 90 % de
miosina-T de rata y el 100 % de miosina-T de toro estan agregadas,
de manera similar a miosina II de músculo liso y estriado ( 89 % y 87
% respectivamente). Con 150 mM de NaCl los porcentajes observados
fueron 50 % y 20 % de agregación para miosina-T de rata y toro
respectivamente, y 75 % y 70 % para miosina II de músculo liso y
músculo estriado, respectivamente. Con 300 mM de NaCl todas las
miosinas probadas presentan un muy bajo grado de agregación (0 %
para miosina-CMP de rata y toro, 30 % y 20 % para miosina II de
músculo liso y estriado).
Por lo tanto, miosina-CMP en medio de baja fuerza iónica tiene
la particularidad de estar totalmente soluble a 4 °C y totalmente
agregada a 20 °C.
Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 66
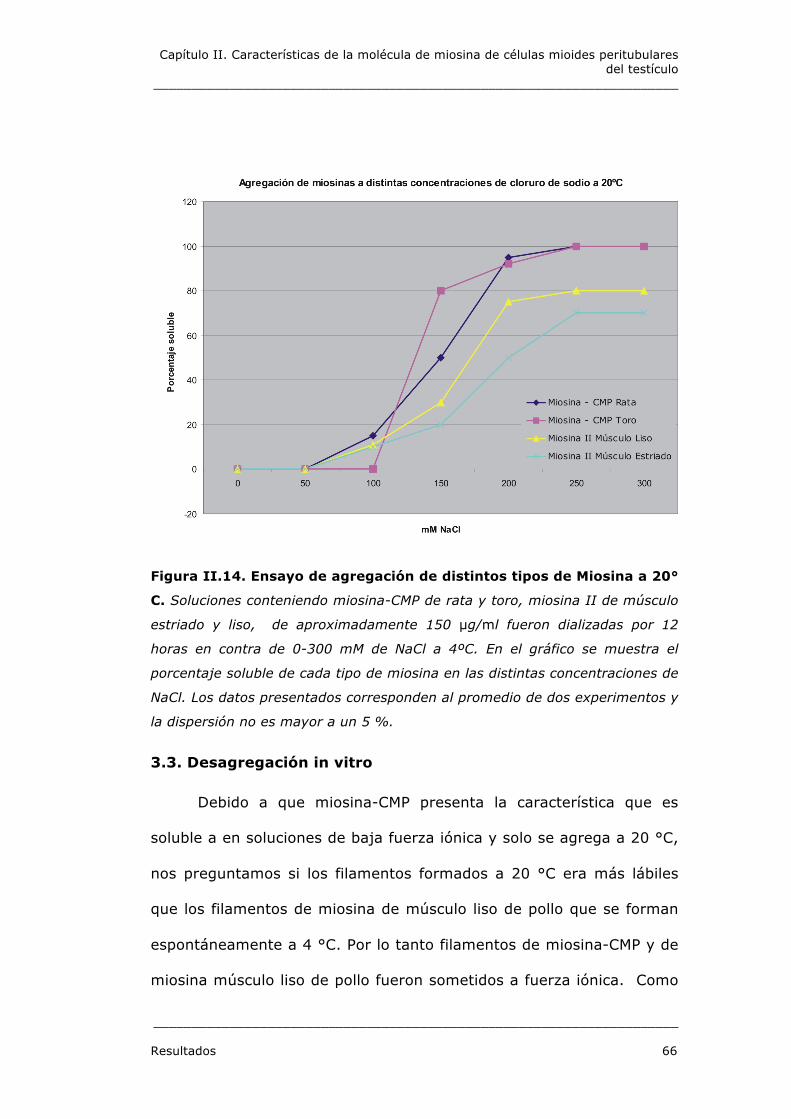
Figura II.14. Ensayo de agregación de distintos tipos de Miosina a 20°
C. Soluciones conteniendo miosina-CMP de rata y toro, miosina II de músculo
estriado y liso, de aproximadamente 150 µg/ml fueron dializadas por 12
horas en contra de 0-300 mM de NaCl a 4ºC. En el gráfico se muestra el
porcentaje soluble de cada tipo de miosina en las distintas concentraciones de
NaCl. Los datos presentados corresponden al promedio de dos experimentos y
la dispersión no es mayor a un 5 %.
3.3. Desagregación in vitro
Debido a que miosina-CMP presenta la característica que es
soluble a en soluciones de baja fuerza iónica y solo se agrega a 20 °C,
nos preguntamos si los filamentos formados a 20 °C era más lábiles
que los filamentos de miosina de músculo liso de pollo que se forman
espontáneamente a 4 °C. Por lo tanto filamentos de miosina-CMP y de
miosina músculo liso de pollo fueron sometidos a fuerza iónica. Como

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 67
la figura II.15 lo indica, es necesario 200 mM de NaCl para solubilizar
alrededor del 20 % de los filamentos de ambas miosinas, y 300 mM de
NaCl para solubilizar el 60 % de los filamentos de ambas miosinas. Por
lo tanto cuando se logran obtener las condiciones para polimerizar
miosina-CMP, los filamentos formados tienen la misma estabilidad a
fuerza iónica que los de miosina-músculo liso de pollo.
Figura II.15. Desagregación de filamentos de miosina por fuerza
iónica. Filamentos de miosina-CMP de rata y miosina II de liso, (formados de
aproximadamente 150 µg/ml de miosina, fueron incubados con 0-300 mM de
NaCl a 20 ºC. En el gráfico se muestra el porcentaje soluble de cada tipo de
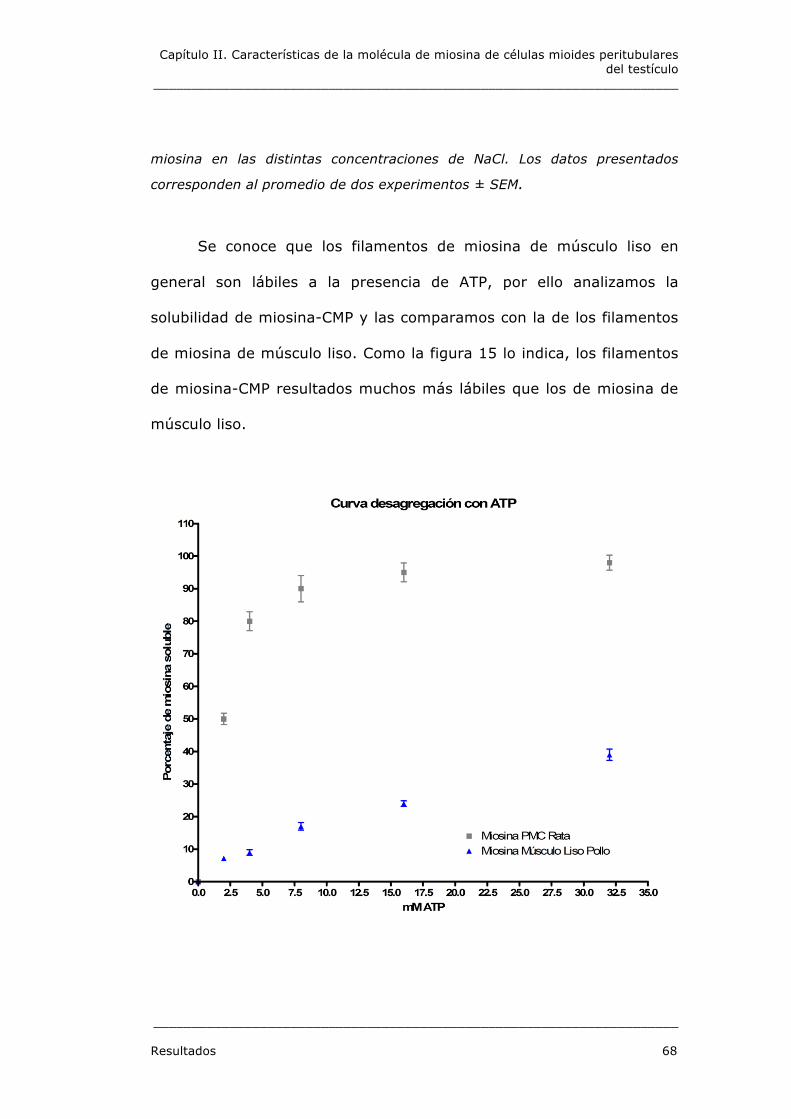
Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 68
miosina en las distintas concentraciones de NaCl. Los datos presentados
corresponden al promedio de dos experimentos ± SEM.
Se conoce que los filamentos de miosina de músculo liso en
general son lábiles a la presencia de ATP, por ello analizamos la
solubilidad de miosina-CMP y las comparamos con la de los filamentos
de miosina de músculo liso. Como la figura 15 lo indica, los filamentos
de miosina-CMP resultados muchos más lábiles que los de miosina de
músculo liso.

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 69
FIGURA II.16. Desagregación de filamentos de miosina por ATP.
Filamentos de miosina-CMP de rata y miosina II de músculo liso, (formados
de aproximadamente 150 µg/ml de miosina, fueron incubados con de 0-35
mM de ATP a 20 ºC. En el gráfico se muestra el porcentaje soluble de cada
tipo de miosina en las distintas concentraciones de ATP. Los datos
presentados corresponden al promedio de dos experimentos ± SEM.

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 70
4. LOCALIZACIÓN DE MIOSINA-CMP EN LOS TEJIDOS DEL
TESTÍCULO
4.1 Células mioides peritubulares.
Para analizar en que células del testículo se encuentra miosina-
CMP, testículos de rata desprovistos de la túnica albugínea y vasos
sanguíneos, fueron sometidos a dos tratamientos enzimáticos ( ver
sección Materiales y Métodos)
Se separaron los túbulos seminíferos del tejido intersticial por un
tratamiento suave con tripsina; y luego los túbulos seminíferos (TS)
fueron disgregados utilizando una solución de digestión que contiene
colagenasa y hialuronidasa (ver materiales y métodos), separando de
esta manera a las células mioides peritubulares (células MP) del resto
de las células del TS (TSd).
Para analizar el grado de desprendimiento de las células MP
durante el tratamiento enzimático del TS, en distintos momentos del
tratamiento, muestras de TS se tiñeron con nitrato de plata y se
observaron en una lupa.
Debido a que las células MP tienen la particularidad de ser las
únicas células del testículo que presentan reacción a fosfatasa alcalina
(Chapin y col. 1987; Palombi y Di Carlo, 1988; Anthony y Skinner,
1989), las fracciones celulares obtenidas del tratamiento enzimático
del TS fueron sometidas a la reacción de fosfatasa alcalina. Además en
la fracción de células MP se determinó el porcentaje de células con
reacción positiva a fosfatasa alcalina.

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 71
El anticuerpo anti-miosina detectó miosina en los túbulos seminíferos
intactos y en la fracción de las células mioides peritubulares Fig. 13 B.
Además el análisis de fosfatas alcalina en las las fracciones indicó un
alto porcentaje de actividad en la fracción de células MP. Por lo tanto,
con estos resultados podemos confirmar que la miosina encontrada en
el testículo proviene de las células MP como lo indica la figura 13 C.
C.
Fracciones TS TSd I CMP
Actividad de Fosfatasa
alcalina
( % del total)
100
16,7± 2,3
4,5± 2,1
70± 5,3
Figura II.16. Localización de miosina-CMP en el tejido testicular.
Túbulos seminíferos intactos (TS); túbulos seminíferos sometidos a la
digestión (TSd); intersticio (I); y fracción de células MP (CMP) fueron
analizados por SDS-PAGE al 7,5 % teñido con azul de Coomassie (A).
Inmunoensayo con anti-miosina (B) y actividad fosfatasa (C). Control positivo
para miosina-CMP(C+), La posición de la cadena pesada de miosina se indica

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 72
en B con una flecha. Las flechas a la izquierda de la figura indican la posición
de proteínas con peso molecular conocido. En C se indica el porcentaje de
actividad de fosfatasa alcalina recuperada en las fracciones, siendo el 100%
de la actividad el corresponde al de los túbulos seminíferos intactos.

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 73
5. DISCUSIÓN DEL CAPÍTULO II
Los resultados indican que en el túbulo seminífero de la rata se
encuentra una proteína con movilidad relativa de 205 kDa. Dicha
proteína es reconocida por un anticuerpo monoclonal anti miosina de
músculo liso. Dicha proteína fue bautizada por nosotros miosina-CMP.
Miosina-CMP tiene la particularidad que se encuentra soluble en
el citosol del homogeneizado de túbulos seminíferos mientras que se
mantenga a 4 °C. Si la suspención se incuba a 37 °C por 30 min,
miosina-CMP se agrega en filamentos.
En base al comportamiento de miosina-CMP a 4 y 37 °C se
desarrolló en este laboratorio un método para purificar la proteína a
partir de un citosol de homogeneizado de tubulos seminíferos. Dicho
método, que consiste en agregar y desagregar la proteína en medios
de baja fuerza iónica, permite obtener una suspención de miosina-CMP
con un 90 % de pureza. Si este material es sometido a centrifugación
en gradiente de sacarosa, se obtienen fracciones de miosina-CMP con
100 % de pureza. Además, se obtuvieron fracciones puras de miosina-
CMP por cromatografía en columna de hidroxiapatita, a partir de
proteínas obtenidas por el método de agregación-desagregación.
Con miosina-CMP purificada se continuó caracterizando la
proteína. Se comprobó que miosina-CMP presenta gran capacidad para
hidrolizar ATP en medio EDTA-K, condiciones solo inherentes para
hidrolizar ATP por la familia de miosinas.

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 74
La digestión parcial de miosina-CMP con tripsina y la
comparación de bandas en electroforesis en geles de poliacrilamida,
permitió evidenciar que presentaba péptidos con movilidad relativa
distinta a la de miosina de músculo liso de arteria espermática y túnica
albugínea de rata.
Además, la composición peptídica de miosina-CMP fue analizada
utilizando espectrometría de masa, específicamente realizando el
análisis Maldi-Tof. Este análisis permitió dilucidar la composición
peptídica de miosina-CMP de testículo de rata y toro y se comprobó
que tenía una baja homología con miosina de músculo liso de rata y de
pollo respectivamente. Además miosina-CMP de rata y toro, mostró
masas de péptidos que entre 10 y 12 no presentaban homología con
ninguna proteína conocida y otros, que solo coincidían con peptidos de
proteínas muy alejados de la familia de proteínas.
Además en este capítulo se analizó la localización de miosina-
CMP y se comprobó que se encontraba en las células mioides
peritubulares del testículo.
Por lo tanto, los resultados obtenidos en este capítulo se pueden
resumir que se encontró una proteína de la familia de las proteínas,
que tiene la particularidad de no ensamblar en filamentos en medios
de baja fuerza iónica. Que miosina-CMP tiene parcial homología con
miosina de músculo liso conocida y que varios péptidos son inéditos.
Nos preguntamos sobre la naturaleza de la solubilidad de
miosina-CMP y en base a los resultados obtenidos comprobamos que la

Capítulo II. Características de la molécula de miosina de células mioides peritubulares del testículo
_____________________________________________________________________
_____________________________________________________________________ Resultados 75
temperatura puede producir agregación. Por otro lado miosina-CMP
purificada presenta la misma característica que la obtenida en el
citosol. Es decir solo se agrega por temperatura. Además
comprobamos que los filamentos formados de miosina-CMP son
estables como los de miosina de músculo liso conocidas. Solo que los
filamentos son más lábiles que los de miosina de músculo liso.
La falta de agregación de miosina-CMP es atribuida a diferencias
peptídicas con la de miosinas de músculo liso convencionales.
Nos preguntamos la función de miosina-CMP en las células
miodes peritubulares del testículo.
Es conocido que muchas miosinas no convencionales se
encuentran solubles en el citoplasma de las células y se les atribuye
función de transporte intracelular. Otras proteínas se encuentran
solubles en determinados momentos y se agregan en filamentos en
circunstancias que las células necesitan de la tracción de estas
proteínas motoras.
Por lo tanto en el próximo capítulo analizaremos la característica
de miosina-CMP en las células mioides peritubulares en desarrollo y
adultas en distintos tramos del túbulo seminífero. Además
relacionaremos la expresión de isotipos de miosina-CMP con la
morfología de las células MP tanto en reposo como en contracción.

CAPÍTULO III
CARACTERÍSTICAS DE LAS CÉLULAS MIOIDES PERITUBULARES DEL
TESTÍCULO

Capítulo III. Características de las células mioides peritubulares del testículo _____________________________________________________________________
_____________________________________________________________________ Resultados 77
CARACTERÍSTICAS DE LAS CÉLULAS MIOIDES PERITUBULARES
DEL TESTÍCULO
Introducción:
Se ha comprobado que las células mioides peritubulares
contienen miosina-CMP que se encuentra soluble en el citoplasma
(datos presentados en el Capitulo II ). Debido a ello nos preguntamos
si miosina-CMP participa activamente en el proceso de contracción.
En este capítulo se describe un método de tinción útil para medir los
parámetros de las células MP. Se determinan las dimensiones y radios
de las células MP en túbulos seminíferos aislados tanto en reposo como
contraídos, y se determina el estado de agregación de miosina-CMP en
túbulos seminíferos contraídos.
1. IDENTIFICACIÓN DE CÉLULAS MIOIDES PERITUBULARES EN
TÚBULOS SEMINÍFEROS AISLADOS
1.1. Tinción con nitrato de plata
Rutinariamente, para observar las células MP en los TS se utiliza
microscopía electrónica de Barrido (Palombi y col. 1992). Esta técnica,
es laboriosa y permite la visualización de porciones discretas de los
segmentos.
Debido a que necesitábamos analizar las células MP en
segmentos completos de TS utilizamos la técnica de tinción con nitrato
de plata, que nos permitió analizar en forma sencilla gran número de
células MP.
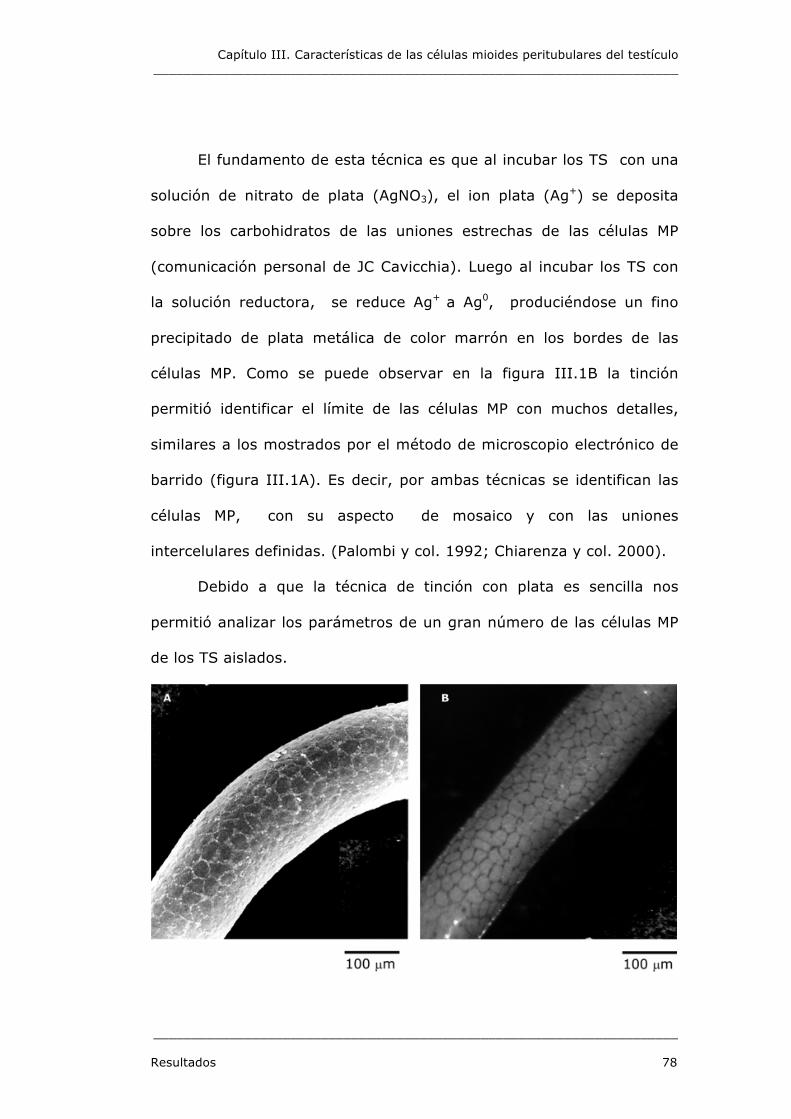
Capítulo III. Características de las células mioides peritubulares del testículo _____________________________________________________________________
_____________________________________________________________________ Resultados 78
El fundamento de esta técnica es que al incubar los TS con una
solución de nitrato de plata (AgNO3), el ion plata (Ag+) se deposita
sobre los carbohidratos de las uniones estrechas de las células MP
(comunicación personal de JC Cavicchia). Luego al incubar los TS con
la solución reductora, se reduce Ag+ a Ag0, produciéndose un fino
precipitado de plata metálica de color marrón en los bordes de las
células MP. Como se puede observar en la figura III.1B la tinción
permitió identificar el límite de las células MP con muchos detalles,
similares a los mostrados por el método de microscopio electrónico de
barrido (figura III.1A). Es decir, por ambas técnicas se identifican las
células MP, con su aspecto de mosaico y con las uniones
intercelulares definidas. (Palombi y col. 1992; Chiarenza y col. 2000).
Debido a que la técnica de tinción con plata es sencilla nos
permitió analizar los parámetros de un gran número de las células MP
de los TS aislados.

Capítulo III. Características de las células mioides peritubulares del testículo _____________________________________________________________________
_____________________________________________________________________ Resultados 79
Figura III.1. Detalles de la pared del túbulo seminífero: A: Microscopía
electrónica de barrido. B: Tinción con nitrato de plata.
1.2. Contracción de las células mioides peritubulares
Se conoce que las células MP tienen la posibilidad de contraerse por
acción de hormonas tales como, oxitocina, angiotensina II y Endotelina
I (ET-1) (Tripiciano y col. 1996; Tripiciano y col. 1997).
Figura III.2. Túbulo seminífero estimulado con Endotelina-1. La flecha
negra indica las células contraídas por endotelina-1 y las flechas blancas los
espermatozoides expulsados.
Debido a que ET-1 es una hormona secretada por las células de
Sértoli, consideramos que era útil para analizar los parámetros de
contracción de las células MP en TS aislados. Para ello los TS del
testículo de rata de 90 días fueron aislados en medio MEM a 32oC, una
fracción fue usada como control y la otra tratada con 50 nM de ET-1

Capítulo III. Características de las células mioides peritubulares del testículo _____________________________________________________________________
_____________________________________________________________________ Resultados 80
por 20 segundos. Los TS fueron fijados con 3% de glutaraldehído y
teñidos con nitrato de plata.
Al observar en la lupa los TS tratados con ET-1 se comprobó que
presentaban un movimiento ondulatorio y expulsaban espermatozoides
por los extremos. (Figura III.2), signos que indican que ET-1 produce
la contracción del TS.
Figura III.3. Túbulos seminíferos teñidos con nitrato de plata. Túbulos
seminíferos de ratas 90 días. A. Control; B. Tratados con 50 nM de ET-1 por
20 seguntos.
1.2.1. Diámetro de los túbulos seminíferos y tamaño de las
células mioides peritubulares
El diámetro del TS control fue de 235.25 ± 6.19 µm, mientras
que el de los TS tratados con ET-1 presentó segmentos de 200.83 ±
5.13 µm y de 251.67 ± 7.89 µm. (Figura III.4).

Capítulo III. Características de las células mioides peritubulares del testículo _____________________________________________________________________
_____________________________________________________________________ Resultados 81
La superficie de las células MP de los TS controles fue de 810.48
± 185.8 µm2 y el de la las células de TS tratados 539.36 ± 144.7 µm2
(figura III.5).
El largo del eje x e y de las células MP controles fue de 34.79 ±
5 µm y 26.1 ± 5.5 µm respectivamente. Mientras que el eje x e y de
las células MP de TS tratados fue de 28.15 ± 3.5 µm y 26.7 ± 4.5 µm
respectivamente (Figura III.5).
Para poder interpretar en que forma se contraen las células MP,
calculamos la relación entre los ejes x e y antes y después de la
contracción. Las mediciones mostraron una relación x/y de 1.24 en el
control y de 0.84 en el tratado. Estos resultados indican que las células
MP al contraerse, mayormente reducen el eje x, es decir aquel
longitudinal al eje mayor del TS.
Figura III.4. Diámetro de los túbulos seminíferos. A: controles y B:
tratados con ET-1.

Capítulo III. Características de las células mioides peritubulares del testículo _____________________________________________________________________
_____________________________________________________________________ Resultados 82
Figura III.5. Tamaño de las células mioides peritubulares. A. Célula de
TS control. B. Células de TS tratado con ET-1.C. Superposición de la figura A
sobre la B

Capítulo III. Características de las células mioides peritubulares del testículo _____________________________________________________________________
_____________________________________________________________________ Resultados 83
2. GRADO DE AGREGACIÓN DE MIOSINA-CMP
2.1. Efecto de endotelina-1
En el capítulo II se mostró que miosina-CMP de las células MP,
es un isotipo de miosina de músculo liso que tiene la particularidad de
ser soluble en soluciones con fuerza iónica similar a la fisiológica (50-
100 mM de NaCl). Además se mostró que miosina-CMP se ensamblaba
en filamentos a 37 °C.
Debido a que miosina-CMP, bajo ciertas condiciones, se
ensambla in vitro a 37 °C, pensamos que podría participar en la
contracción de las células MP. Como ya se mostró arriba, ET-1 produce
la contracción de cada célula MP del TS. Para analizar la distribución de
miosina-CMP, túbulos seminíferos incubados con ET-1, por 20
segundos, fueron inmediatamente homogeneizados y analizados para
determinar la distribución de miosina-CMP en el sobrenadante y
sedimento.
En TS controles (no tratados con ET-1) 85 ± 5% de miosina-
CMP se encuentra soluble, mientras que en los TS contraídos con ET-1,
solo el 40 ± 6 % queda en el sobrenadante. (Ver figura III.6). De
estos resultados se desprende que miosina-CMP es capaz de
ensamblarse en filamentos cuando la célula MP se contrae.

Capítulo III. Características de las células mioides peritubulares del testículo _____________________________________________________________________
_____________________________________________________________________ Resultados 84
Figura III.6. Agregación de miosina-CMP por ET-1. Miosina-CMP en
sobrenadante y sedimento de túbulos seminíferos controles (A) y tratados con
50 nM de ET-1 por 20 segundos (B), purificada por el método agregación-
desagregación. 3 µg de sobrenadante y 8 µg del sedimento fueron analizados
por SDS-PAGE al 7.5% y teñido con azul de coomassie de. La intensidad de
las bandas proteicas se analizó con el software Scion Image.

Capítulo III. Características de las células mioides peritubulares del testículo _____________________________________________________________________
_____________________________________________________________________ Resultados 85
3. DISCUSIÓN DEL CAPÍTULO III
Los resultados presentados en este capítulo indican que se
desarrolló una técnica sencilla de tinción con nitrato de plata, que
permitió determinar las características morfológicas de las células MP
en la pared del TS.
Asimismo, por el tratamiento con ET-1 se logró, a juzgar por el
movimiento de los TS y por la expulsión de espermatozoides, la
contracción de los TS aislados.
La tinción de los TS con nitrato de plata permitió dilucidar
claramente el límite de cada célula MP y determinar sus dimensiones.
Se comprobó que las células MP tienen una superficie de
alrededor 810 µm2 que se reduce un 33,5 % por acción de ET-1. El eje
x de las células MP es de alrededor 34 µm y se reduce en 9 µm por el
efecto de ET-1, mientras que el eje y de las células MP es de alrededor
26 µm y no cambia por acción de ET-1; indicando que la célula se
contrae acortando el eje x. El acortamiento del eje x podría inducir un
incremento del diámetro del TS, que podría justificar el hallazgo
experimental de encontrar sectores de TS tratados con ET-1 con un
diámetro mayor al del control.
El análisis del grado la distribución de miosina-CMP entre soluble
y agregada permitió comprobar que una significativa cantidad de
miosina-CMP se agrega en filamentos cuando los TS se contraen, lo
que indica que miosina-CMP participa activamente en la contracción de
las células MP.

CAPÍTULO IV
ACTIVIDAD DE LAS CÉLULAS MIOIDES PERITUBULARES
RELACIONADA CON EL EPITELIO SEMINIFERO

Capítulo IV. Actividad de las células mioides peritubulares relacionada con el epitelio seminífero
_____________________________________________________________________
_____________________________________________________________________ Resultados 87
ACTIVIDAD DE LAS CÉLULAS MIOIDES PERITUBULARES
RELACIONADA CON EL EPITELIO SEMINIFERO
Introducción:
Ya se ha mostrado en este trabajo que las células MP tienen un
tipo particular de miosina que denominamos miosina-CMP. Miosina-
CMP se encuentra soluble en el citoplasma y es capaz de formar
filamentos en el momento que las células MP se contraen.
Además se ha desarrollado un método para identificar las células
MP en los TS, tinción con plata, que permite el análisis morfométrico
de un gran número de células.
En este capítulo se analizan las características morfológicas de las
células MP y la expresión de isotipos de miosina-CMP en los TS del
testículo en desarrollo y en los segmentos del TS relacionados con los
estados del epitelio seminífero del testículo de rata adulta.
1. DESARROLLO DEL TÚBULO SEMINÍFERO Y DE LAS CÉLULAS
MIOIDES QUE LO RODEAN
Desde el primer día de nacimiento, los TS del testículo
comienzan a madurar y van cambiando su morfología. Nos
preguntamos en que momento comienzan a tener capacidad de
contraerse, como es la morfología de las células MP y que isotipos de
miosina-CMP expresan.
1.1. Diámetro del túbulo seminífero de testículos en desarrollo
En los primeros días postnatal (pn), el testículo presenta
esbozos de los TS, pero en realidad son cordones sólidos. Recién en el

Capítulo IV. Actividad de las células mioides peritubulares relacionada con el epitelio seminífero
_____________________________________________________________________
_____________________________________________________________________ Resultados 88
día 10 pn, algunos cordones muestran lumen y recién en el día 30 pn
todos los túbulos presentan lumen. (Ver capitulo I).
Se ha comprobado que el diámetro del TS va creciendo
sostenidamente desde el nacimiento hasta día 40 pn (capítulo I).
Mediciones propias nos indican la dinámica de crecimiento de los
TS en los primeros 30 días pn. Como la figura IV.1 lo indica desde el
día 14 pn al 18 pn el diámetro crece de 60 a 120 µm, luego del día 18
al 24 pn, crece solo de 120 µm a 150 µm, retomando luego el ritmo de
crecimiento de los primeros días desde el día 24 pn hasta el 30 pn.
Figura IV.1. Diámetro de los túbulos seminíferos de ratas en desarrollo. Se
determinó el diámetro de los túbulos seminíferos en cortes

Capítulo IV. Actividad de las células mioides peritubulares relacionada con el epitelio seminífero
_____________________________________________________________________
_____________________________________________________________________ Resultados 89
histológicos de testículos de rata. Los datos son el valor medio ± SEM
de tres ratas.
1.2. Caracteres morfológicos de las células-MP en túbulos
seminíferos en desarrollo
Analizamos las células MP en los primeros días de desarrollo
postnatal, en el día 16 pn las células muestran una superficie de 136,2
µm2 con un diámetro en el eje x de 10,6 µm y en el y de 12,1 µm. En
el día 20 pn la superficie de las células MP ya es de 412,9 µm2 siendo
el diámetro del eje x e y de 19,2 µm y 21,5 µm respectivamente.
Además en el día 30 pn el área de las células MP es de 481,6 µm2 y el
diámetro en el eje x e y es 22,7 µm y 23,0 µm respectivamente.
(Figura IV.2 y tabla IV.1)
Capítulo IV. Actividad de las células mioides peritubulares relacionada con el epitelio seminífero
_____________________________________________________________________
_____________________________________________________________________ Resultados 90
FIGURA IV.2. Túbulos seminíferos de ratas en desarrollo. Tinción con
nitrato de plata. A-B de 16 días control y tratados. C-D de 20 días
control y tratado y E-F de 30 días control y tratado. Las barras indican
200 µm.

Capítulo IV. Actividad de las células mioides peritubulares relacionada con el epitelio seminífero
_____________________________________________________________________
_____________________________________________________________________ Resultados 91
TABLA IV.1. Área y diámetros de las células MP en ratas en desarrollo
AREA
(µm2 ±ESM)
Diámetro (x)
(µm ±ESM)
Diámetro (y)
(µm ±ESM)
EDAD (días pn) control tratado control tratado control tratado
16
136,2
±10,2
139,1
±8,5
10,6
±0,5
11,9
±0,3
12,1
±0,14
11,7
±0,61
20
412,9
±10,1
220,3
±11,1
**
19,2
±0,35
14,3
±0,71
**
21,5
±0,12
16,6
±0,21
**
30
481,6
±24,1
343,6
±18,4
**
22,7
±1,5
19,3
±1,4
**
23,0
±2,2
18,5
±1,4
**
** p < 0,01
Nos preguntamos si las células MP en los primeros días de
desarrollo postnatal responden al tratamiento con ET-1. Como se
puede observar en la figura IV y tabla IV.1, ET-1 no indujo cambios ni
el área ni en los diámetros de los x e y en las células de 16 días pn. En
cambio ET-1 produjo una reducción del alrededor del 50 % del área de
las células del día 20 pn y una significativa disminución del diámetro
del los dos ejes (x, y). Además ET-1 en células de 30 días pn produjo
una reducción de alrededor del 30 % en el área y una significativa
reducción en el diámetro solo del eje y.

Capítulo IV. Actividad de las células mioides peritubulares relacionada con el epitelio seminífero
_____________________________________________________________________
_____________________________________________________________________ Resultados 92
1.3. Isotipos de miosina-CMP en las células mioides
peritubulares en desarrollo
Si se analiza la movilidad electroforética de miosina-CMP
proveniente de testículos de rata adulta, en geles de 4,5 %, se
detectan dos bandas de 205 y 200 kDa que denominamos isotipos de
miosina-CMP A y B respectivamente. Las dos bandas son reconocidas
por los anticuerpos anti-miosina de músculo liso. Comprobamos que en
el testículo adulto, el 40 % de miosina-CMP es del isotipo A y 60 %
del isotipo B. (Figura IV.3).
Figura IV. 3. Isotipos de miosina-CMP en testículo de rata adulta. Gel
SDS-PAGE al 4,5 %, teñido con azul de coomasie.
Se analizó la expresión de los isotipos A y B de miosina-CMP en
túbulos seminíferos de testículo de rata en desarrollo. Para ello se
purificó miosina-CMP de testículos de rata de 12 a 43 días de edad y
cada muestra se analizó por electroforesis.
Los resultados indican que en los túbulos seminíferos de ratas
desde el día 12 de edad van expresando más cantidad del isotipo B

Capítulo IV. Actividad de las células mioides peritubulares relacionada con el epitelio seminífero
_____________________________________________________________________
_____________________________________________________________________ Resultados 93
que del A hasta llegar al día 27 donde expresan 70 % del isotipo B y
solo 30 % del A (Figuras IV.4 y 5).
Figura IV.4. Isotipos miosina-CMP A y B de túbulos seminíferos de
ratas en desarrollo (12-43 días de nacimiento). SDS-PAGE al 4,5 %
teñido con azul de coomassie. Se muestra un experimento típico.
Llamativamente, desde el día 27 pn comienza a decrecer la
expresión del isotipo B y por ende a expresarse el A, llegando al día 35
donde se expresan ambos isotipos en el mismo porcentaje, 50 %.
Luego desde el día 35 pn, nuevamente comienza a expresarse mayor
cantidad del isotipo B, 60%, manteniéndose esa relación hasta los
túbulos seminíferos de los testículos de rata adulta.
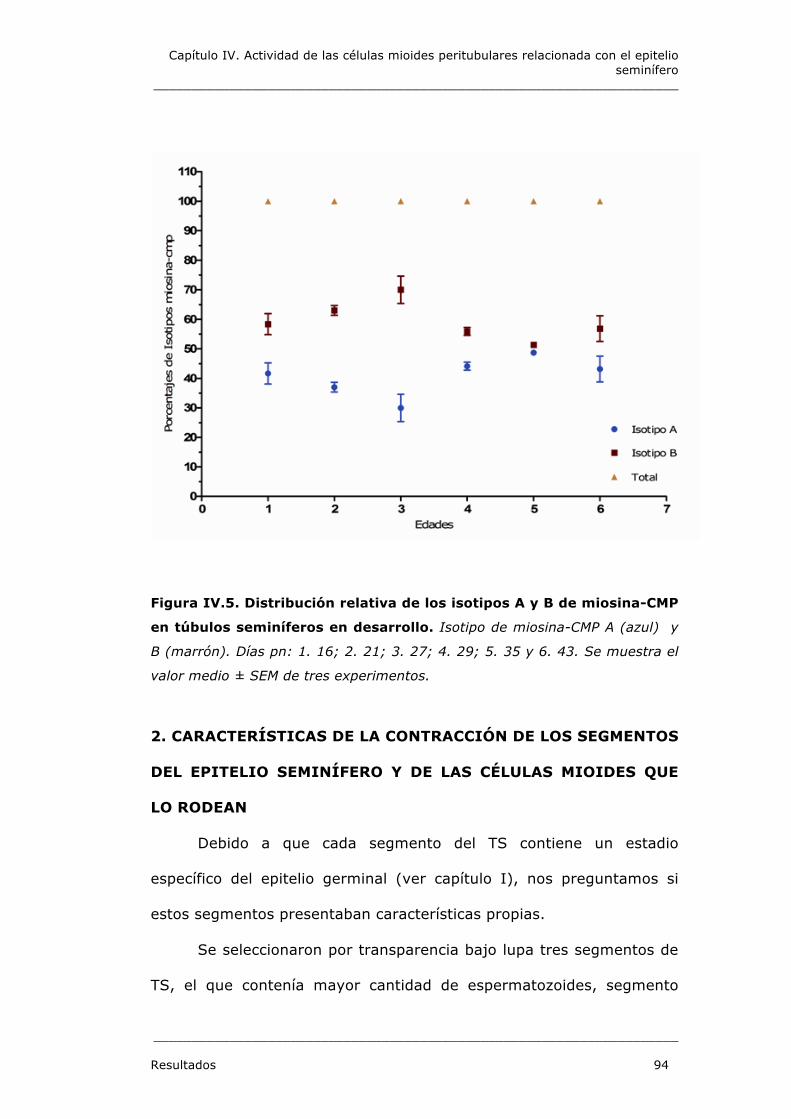
Capítulo IV. Actividad de las células mioides peritubulares relacionada con el epitelio seminífero
_____________________________________________________________________
_____________________________________________________________________ Resultados 94
Figura IV.5. Distribución relativa de los isotipos A y B de miosina-CMP
en túbulos seminíferos en desarrollo. Isotipo de miosina-CMP A (azul) y
B (marrón). Días pn: 1. 16; 2. 21; 3. 27; 4. 29; 5. 35 y 6. 43. Se muestra el
valor medio ± SEM de tres experimentos.
2. CARACTERÍSTICAS DE LA CONTRACCIÓN DE LOS SEGMENTOS
DEL EPITELIO SEMINÍFERO Y DE LAS CÉLULAS MIOIDES QUE
LO RODEAN
Debido a que cada segmento del TS contiene un estadio
específico del epitelio germinal (ver capítulo I), nos preguntamos si
estos segmentos presentaban características propias.
Se seleccionaron por transparencia bajo lupa tres segmentos de
TS, el que contenía mayor cantidad de espermatozoides, segmento

Capítulo IV. Actividad de las células mioides peritubulares relacionada con el epitelio seminífero
_____________________________________________________________________
_____________________________________________________________________ Resultados 95
VII-VIII; el segmento inmediato al VII-VIII que no contenía
espermatozoides, segmento IX-XIV; y el inmediato al segmento VII-
VIII que contenía relativa cantidad de espermatozoides, segmento I-
VI. Cada segmento fue tratado con y sin ET-1 y fue analizado el
diámetro del TS, la morfología de las células MP y la expresión de
isotipos de miosina-CMP.
2.1 Diámetro de distintos segmentos de túbulos seminíferos
El diámetro del segmento I-VI fue de alrededor 230 µm y por el
tratamiento con endotelina incrementó su diámetro a 250 µm. El
segmento IX-XIV tiene un diámetro de 200 µm y por el tratamiento
con ET-1 no sufre ningún aumento significativo en su diámetro. En
cambio el segmento VII-VIII con un diámetro de 200 µm cambió a
uno de 280 µm por el tratamiento con ET-1. (Figuras IV.6 y 7).

Capítulo IV. Actividad de las células mioides peritubulares relacionada con el epitelio seminífero
_____________________________________________________________________
_____________________________________________________________________ Resultados 96
Figura IV.6. Diámetro de distintos segmentos del túbulos seminífero.
Se midieron los segmentos I-VI, VII-VIII y IX-XIV antes (controles) y después
del tratamiento con ET-1 (tratados). Se muestra el valor medio ± SEM de tres
experimentos.
2.2 Morfología de las células mioides peritubulares en los
distintos segmentos del túbulo seminífero
Se determinó el área y los diámetros de los ejes x e y de las
células MP de los tres segmentos del TS.
El área de las células MP del segmento I-VI fue de 1141 µm2 y
por el tratamiento con endotelina-1 (ET-1) se redujo a 995 µm2

Capítulo IV. Actividad de las células mioides peritubulares relacionada con el epitelio seminífero
_____________________________________________________________________
_____________________________________________________________________ Resultados 97
produciendo un cambio del 12,8 %. El diámetro en los ejes x-y fueron
de 38,9 µm -31,7 µm respectivamente y después del tratamiento con
ET-1 34,1 µm -31,1 µm respectivamente mostrando una reducción
significativa en diámetro del eje x. (Figura IV.7 y tabla IV.2)
Figura IV.7. Túbulos seminíferos en estadíos I –VI. Tinción con plata. A:
Control, B: Tratado con ET-1 durante 20 segundos.
El área de las células MP del segmento VII-VIII fue de 964 µm2
y por el tratamiento con ET-1 se redujo a 769 µm2 produciendo un
cambio del 20,2 %. El diámetro en los ejes x-y fueron de 35,9 µm -
30,3 µm respectivamente y después del tratamiento con ET-1 28,3 µm
- 30,0 µm respectivamente mostrando una reducción significativa solo
en el diámetro del eje x. (Figura IV.8 y tabla IV.2).

Capítulo IV. Actividad de las células mioides peritubulares relacionada con el epitelio seminífero
_____________________________________________________________________
_____________________________________________________________________ Resultados 98
Figura: IV.8. Túbulos seminíferos en estadío VII-VIII. Tinción con plata. A:
Control, B: Tratado con ET-1 durante 20 segundos.
Mientras que las células MP del segmento IX-XIV tienen un área
938 µm2 que no se modifico por el tratamiento con E-1. Por
consiguiente no se modificaron el diámetro en el eje x e y que fue de
34,8 µm y 29,5 µm respectivamente. (Figura IV.9 y tabla IV.2)
Figura IV.9. Túbulos seminíferos en estadío IX-XIV. Tinción con plata.
A: Control, B: Tratado con ET-1 durante 20 segundos.

Capítulo IV. Actividad de las células mioides peritubulares relacionada con el epitelio seminífero
_____________________________________________________________________
_____________________________________________________________________ Resultados 99
Tabla IV.2. Áreas y diámetros de las células MP en el estadío I-VI, VII-
VIII y IX-XIV.
AREA
(µm2 ±ESM)
Diámetro (x)
(µm ±ESM)
Diámetro (y)
(µm ±ESM)
ESTADIOS control tratado control tratado control tratado
I-VI
1141,2
±55,1
995,3
±67,2
**
38,9
±0,8
34,1
±0,9
**
31,7
±0,8
31,1
±0,9
VII-VIII
964,9
±59,1
769,7
±81,2
**
35,9
±0,8
28,3
±1,6
**
30,3
±1,2
30,0
±0,7
IX-XIV
938,9
±64,3
970,2
±67,2
34,8
±0,9
35,3
±0,8
29,5
±0,7
29,8
±0,8
** p < 0,01
2.3. Isotipos de miosina-CMP en los distintos segmentos del
túbulo seminífero
En los tres segmentos del TS se purificó miosina-CMP y se
analizó el grado de expresión de los isotipos A y B.
Como ya se había mostrado en la figura IV.3 en las células MP
del conjunto de los TS del testículo adulto se expresan 40 % del isotipo
A y 60 % del isotipo B. Cuando se analiza la expresión de los isotipos A
y en el segmento I-VI de los TS se observa que la expresión es 40 %
de A y 60 % de B y la misma expresión se encuentra en el segmento
IX-XIV. Mientras que en el estadio VII-VIII se expresa 75 % del A y
solo 25 % del B. De estos resultados se desprende que hay

Capítulo IV. Actividad de las células mioides peritubulares relacionada con el epitelio seminífero
_____________________________________________________________________
_____________________________________________________________________ Resultados 100
preponderancia en la expresión del isotipo B en el segmento IX-XIV del
TS.
Figura IV.10. Distribución relativa de los isotipos de miosina-CMP en
segmentos de túbulos seminíferos. Isotipos de miosina-CMP A (azul) y B
(rojo) en los segmentos del túbulo seminífero conteniendo los estados I-VI,
VII-VIII y IX-XIV del epitelio seminífero y en el túbulo seminífero entero,
control. Se muestra el valor medio ± S.E.M de tres experimentos. Inserto en
la parte superior de la figura se muestra un ejemplo típico de las bandas A y B
de miosina-CMP en los tres segmento del túbulo seminífero y en el control.
Tinción con azul de coomassie.

Capítulo IV. Actividad de las células mioides peritubulares relacionada con el epitelio seminífero
_____________________________________________________________________
_____________________________________________________________________ Resultados 101
3. DISCUSIÓN DEL CAPITULO IV
Los resultados presentados indican que los TS presentan desde el
día 14 pn un marcado crecimiento en diámetro que se mantiene hasta
el día 18 pn, luego se aprecia un enlentecimiento en el ritmo de
crecimiento hasta el día 18, donde comienza a crecer en forma
sostenida.
Durante el desarrollo del TS las células MP presentan cambios
notables. A los 16 días pn las células MP tienen un diámetro de 136
µm2 que se incrementa a 412 µm2 en el día 20 pn. Además desde el
día 20 pn las células MP manifiestan la capacidad de contraerse por
estímulo con ET-1.
Junto con el cambio morfológico de las células MP y su capacidad
de contraerse se aprecian cambios en la expresión de los isotipos de
miosina-CMP. En el día 16 pn se manifiesta el 40 % del isotipo A y 60
% del isotipo B. Mientras que en el día 21 pn se observa incremento en
la expresión del isotipo B, 65%, y en el día 27 pn al valor máximo de
70%.
En una segunda parte de éste capítulo se muestra que las
células MP de los distintos segmentos del TS presentan características
propias.
Las células MP del segmento I-VI son de mayor área y se
reducen un 12 % al contraerse. Las células MP del segmento VII-VIII
se contraen fuertemente y reducen su área un 20 %. En cambio las

Capítulo IV. Actividad de las células mioides peritubulares relacionada con el epitelio seminífero
_____________________________________________________________________
_____________________________________________________________________ Resultados 102
células MP del segmneto IX-XIV no cambian su área al ser estimuladas
a contraerse, interpretándose que no se contraen.
Miosina–CMP además presenta particularidades de expresión en
las distintas porciones del TS. En el segmento I-VI 40 % corresponde
al isotipo A y 60% al isotipo B, mientras que en el segmento VII-VIII,
25 % corresponde al isotipo A y 75 % corresponde al isotipo B. En el
segmento IX-XIV se restablece la proporción 40 % isotipo A, 60 %
isotipo B.
Es interesante destacar que la expresión del isotipo B de
miosina-CMP está relacionada con la mayor actividad de contracción de
las células MP tanto en el desarrollo como en el testículo adulto.

CAPÍTULO V
DISCUSIÓN GENERAL Y CONCLUSIONES

Discusión General _____________________________________________________________________
_____________________________________________________________________ Discusión 104
DISCUSION GENERAL Y CONCLUSIONES
1. Características de miosina-CMP
En este trabajo se muestra que una proteína de testículo de rata
que denominamos miosina-CMP tiene movilidad electroforética de 205
kDa (cadena pesada) y de 22 y 17 kDa (cadenas livianas), presenta
características que la permiten clasificar dentro de la familia de las
miosinas. La cadena pesada presenta una secuencia de aminoácidos
con parcial homología a la cadena pesada de miosina II de músculo
liso y además es reconocida con anticuerpos monoclonales anti-
miosina II de músculo liso. El perfil de péptidos de la cadena pesada
presenta diferencias con miosina II de la arteria espermática y de la
túnica albugínea.
Miosina-CMP se encuentra en las células MP de los TS del
testículo de rata, basándonos en los siguientes criterios. Se aísla de las
células que revisten los TS y la remoción de dichas células deja
desprovisto al TS de miosina-CMP.
El 90 % de miosina-CMP se encuentra soluble (sin formar
filamentos) en el citoplasma celular. Este comportamiento se puso en
evidencia al recuperar miosina-CMP en el sobrenadante de
homogeneizados de TS con soluciones de baja fuerza iónica.
Miosina-CMP, en medios de baja fuerza iónica, permanece
soluble a 4°C y se agrega y forma filamentos a 20 °C.

Discusión General _____________________________________________________________________
_____________________________________________________________________ Discusión 105
Los filamentos formados in vitro de miosina-CMP tienen la
misma estabilidad a fuerza iónica que los filamentos de miosina II de
músculo liso, pero mayor labilidad a ATP.
De los resultados detallados arriba llama la atención el siguiente
hecho, miosina-CMP presenta muchas de las características de la
familia de moléculas de miosina II de músculo liso, excepto que se
encuentra soluble en el citosol de las células y no forma filamentos in
vitro a 4°C.
En principio, el comportamiento de miosina-CMP puede atribuirse
a algunas de las siguientes razones: Desfosforilación de sitios de las
cadenas livianas o fosforilación de sitios de la cadena pesada. Falta de
dominios peptídicos en el extremo carboxilo terminal.
Desfosforilación de la cadena liviana. Las moléculas de miosina
II de músculo liso deben mantenerse fosforiladas para ensamblarse en
filamentos. La falta de fosforilaciones hace soluble a las moléculas de
miosina. (Ver Capitulo I sección 2.3.2).
Fosforilación de las cadenas pesadas. Se ha comprobado que en
las células no musculares, en ciertas circunstancias, miosina II se
solubiliza por la fosforilación de la cadena pesas (ver Capitulo I sección
2.3.3) y una ulterior defosforilación permite que forme filamentos
nuevamente.
Dominios peptídicos. Hodge y col. 1992, han propuesto que la
función de la porción no helicolidal de la cola de miosina II es la de
promover o potenciar el ensamble mediante la unión a sitios

Discusión General _____________________________________________________________________
_____________________________________________________________________ Discusión 106
específicos de las moléculas vecinas. La ausencia o mutación de esta
pieza provoca la acumulación de una conformación intermedia que no
es apta para el ensamble en filamentos.
En base al comportamiento de otras miosinas II en los sistemas
descriptos, se puede interpretar que miosina-CMP en el citoplasma de
las células MP tiene desfosforilada la cadena liviana o fosforilada la
cadena pesada, de tal modo que se mantiene soluble.
Si tenemos en cuenta el comportamiento de miosina-CMP in
vitro, que a 4° C está soluble, mientras que a 20° C se agrega, sin
que medien reacciones de fosforilación/desfosforilación, nos lleva a
considerar poco probables las hipótesis anteriores y plantear que
miosina-CMP carece de dominios de ensamble o que no están
expuestos.
Es de destacar, que el cambio de solubilidad de miosina-CMP
por la temperatura sólo se produce in vitro, y es probable que esté
reflejando un comportamiento que se repite en el citoplasma celular
activado por otros factores, tales como proteínas.
Se conocen proteínas como “kinase-related protein ” (KPR),
telokin y proteína de 38 kDa que confieren estabilidad a miosina II en
condiciones donde la concentración de ATP es elevada y los niveles de
fosforilación de las cadenas livianas es escasa ,(Shirinsky y col., 1993;
Okagaki y col., 2000). KRP previene que miosina II adopte
conformación 10 S porque compite con al región de miosina II donde
la cola se pliega (Olney y col., 1996; Silver y col., 1997).

Discusión General _____________________________________________________________________
_____________________________________________________________________ Discusión 107
¿ Forma miosina-CMP filamentos en las células mioides perituburales?.
Una vez que comprobamos que la proteína soluble miosina-CMP
proviene de las células MP, que es un isotipo de miosina II de músculo
liso, que presenta mayor solubilidad in vitro, que se ensambla in vitro
en filamentos por tratamiento térmico, nos preguntamos si miosina-
CMP en el citoplasma de la célula se ensambla y participa en la
contracción de la célula.
Para contestar esta pregunta era necesario contar con un
método sencillo y reproducible que nos permitiera observar y medir las
células MP en la pared de los TS para corroborar su contracción. El
intercambio de ideas con el Dr. Juan C. Cavicchia nos llevó a ensayar
con la tinción con plata de los TS. De esta manera se puso a punto un
método que nos permitió en pocos minutos observar en lupa
estereoscópica las células MP con todos sus detalles.
El primer paso fue analizar in vitro la contracción del TS con ET-
1, una hormona que participa activamente en el testículo en la
contracción del TS. (Fantoni y col. 1993).
Utilizando ET-1 en segmentos aislados de TS, se observó que
producía la expulsión de espermatozoides por los extremos y cambios
en la sección de los TS. El TS en conjunto presentó un diámetro de
235 µm y cuando se lo trató con ET-1, ciertos segmentos presentaron
un diámetro significativamente mayor, 251 µm.
Las mediciones morfológicas de las células MP en el testículo
adulto indicaron que tienen el aspecto hexagonal con un área de 810

Discusión General _____________________________________________________________________
_____________________________________________________________________ Discusión 108
µm2 que se reduce a 539 µm2 por el tratamiento con ET-1. Presenta un
eje x (longitudinal al eje mayor del TS) de 34 µm que es mayor al eje
y (perpendicular al eje mayor del TS) de 28 µm. Sólo en los
segmentos donde el TS aumenta de diámetro se observaron células MP
con cambios morfológicos. Nos llamó la atención que en la contracción
de la célula MP, el eje x es el que se reduce, mientras que el eje y no
cambia, indicando que las células MP presentan una organización del
citoesqueleto que le permite contraerse en forma anisodiamétrica.
Habiendo comprobado en forma morfológica la contracción de
las células MP, nos preguntamos si miosina-CMP participa en el
mecanismo de contracción. Para ello, a TS aislados antes y después de
la contracción se les analizó la distribución de miosina-CMP.
Se comprobó que durante la contracción de las células MP, una
significante cantidad de miosina-CMP faltaba en el sobrenadante y se
encontraba agregada como filamentos, interpretándose que miosina-
CMP participa activamente en la contracción de las células.
De estos resultados se infiere que miosina-CMP participa dentro
de la célula en el sistema contráctil. Se encuentra soluble cuando está
inactiva, forma filamentos en respuesta a un estímulo y vuelve a
solubilizarse después de la contracción.
Existen antecedentes relacionados con un estado cíclico de
solubilidad de miosina en las células. Las células en división, disponen
de un mecanismo por el cuál solubilizan miosina II citoplasmática, la
trasladan, la ensamblan en filamentos y forman el anillo contráctil de

Discusión General _____________________________________________________________________
_____________________________________________________________________ Discusión 109
la citocinesis. Este es un modelo de desensamble-ensamble llevado a
cabo por la fosforilación-desfosforilación de la cadena pesada de
miosina II. (Burgess, 2005)
En células satélites del hígado, que participan en la contracción de
los sinusoiedes hepáticos, la fosforilación de las cadenas livianas de
miosina II induce la formación de filamentos en el momento de la
contracción. Este modelo de desensamble-ensamble de miosina II es
llevado a cabo por la desfosforilación-fosforilación de las cadenas
livianas (Saab y col. 2002).
Volviendo a miosina-CMP en las células MP, de acuerdo a los
resultados obtenidos in vitro, no sería el estado de fosforilación el que
determina la solubilidad de miosina-CMP, sino proteínas que facilitarían
el ensamble de la proteína en filamentos, por lo tanto miosina-CMP
alternaría entre el estado soluble o agregado dependiendo de proteínas
que estabilizaran la unión entre miosinas.
¿Cual es la ventaja de disponer de un sistema de ensamble-
desensamble de miosina? Las moléculas de miosina son verdaderos
motores celulares que generan la tracción mecánica de la célula. En
las células del músculo estriado y cardíaco las moléculas de miosina
son parte de la unidad de contracción, el sarcómero, en forma de
filamentos gruesos constituidos cada uno por mas de 50 moléculas de
miosina, que generan contracción al desplazar los microfilamentos.
Tanto en las células de músculo estriado como cardíaco, los
sarcómeros permanecen armados toda la vida de la célula, que en

Discusión General _____________________________________________________________________
_____________________________________________________________________ Discusión 110
muchos casos coincide con la vida del organismo (Landsverk y Epstein.
2005).
En cambio, en las células de músculo liso, la unidad de
contracción es más laxa, aunque los filamentos de miosina se
mantienen armados durante toda la vida de la célula Landsverk y
Epstein. 2005.)
En las células no musculares, miosina sigue siendo la unidad de
contracción, pero la organización de los filamentos de miosina es
temporal y depende de la actividad de la célula. De esa manera,
cuando una célula se desplaza sobre un sustrato, continuamente esta
reorganizando los filamentos de miosina y de actina, y en muchos
casos las moléculas de miosinas cambian de estado, de solubles pasan
a formar filamentos y luego a ser solubles nuevamente (Conrad y col.
1995).
Las células MP presentan características de células de músculo
liso, pero de acuerdo a nuestros resultados miosina-CMP, que es la
principal proteína con característica de miosina que presenta la célula,
se mantiene soluble y solo se ensambla en filamentos por el estímulo
de la contracción.
En este aspecto miosina-CMP se comporta como una miosina de
células no musculares, y podría estar indicando la plasticidad que
presenta el citoesqueleto para cambiar en cada instante la forma de la
célula MP. El hecho de que hemos encontrado células MP con distinta
morfología en los segmentos del TS con diferentes estados del epitelio

Discusión General _____________________________________________________________________
_____________________________________________________________________ Discusión 111
seminífero podría confirmar la hipótesis de un citoesqueleto
sumamente dinámico.
2. Contracción de las células MP y expresión de isotipos de
miosina-CMP
En el desarrollo del testículo. En el testículo de rata miosina-CMP
se expresa en gran cantidad en las células MP y participa en la
contracción de las mismas provocando la reducción del área celular y
modificando la forma de la célula (se reduce en mayor proporción el
eje x).
Cuándo se analiza la movilidad electroforética de la cadena
pesada de miosina-CMP se observan dos bandas que corresponden al
isotipo A (205 kDa) y B (200 kDa).
Desde el momento del nacimiento el TS presenta profundos
cambios morfológicos, primero aparece el lumen y luego va creciendo
en diámetro y longitud (Ver CAPII). Observamos que los TS, desde el
día 14 pn al día 18, duplican el diámetro, luego desde el día 18 pn al
24 el crecimiento en diámetro se enlentese y nuevamente retoma el
ritmo de crecimiento en el día 24 pn.
De la misma manera las células MP en los TS aumentan tres
veces el área desde el día 16 pn al 20 pero luego conservan las
mismas dimensiones hasta el día 30 pn.
Con respecto a miosina-CMP, Las células MP del día 16 pn
expresan 40 % del isotipo A y 60 % del isotipo B. Luego las células del

Discusión General _____________________________________________________________________
_____________________________________________________________________ Discusión 112
día 21 pn al día 24 van expresando mayor cantidad del isotipo B,
llegando al día 24 pn a expresar el 70 %.
Es conocido que las células MP comienzan a producen
microfilamentos circulares del citoesqueleto desde el día 16 pn y en el
día 30 pn aparecer en adición a los circulares, haces que corren
longitudinales al eje mayor del TS. (Ver Cap. I Sección 1.1.5).
Por lo tanto en este período 16-20 días pn, donde las células MP
cambian significativamente el área el citoesqueleto de actina se
reorganiza profundamente de la misma forma que se incrementa la
expresión del isotipo B de miosina-CMP.
Concomitantemente con la reorganización del citoesqueleto, las
células MP comienzan a tener una marcada actividad contráctil que se
pone de manifiesto el día 20 pn.
¿Es necesaria la expresión elevada del isotipo B de miosina-CMP para
que la célula se contraiga apropiadamente? Es conocido que dos
isotipos de la cadena pesada de miosina II de músculo liso se
encuentran en las células de músculo liso y son conocidos como SM1
(204 kDa) y SM2 (200 kDa). SM1 presenta un extremo C-terminal con
9 aminócidos más que SM2. En la rata, SM2 aparece mas tarde en el
desarrollo embrionario y es expresado a elevados niveles en células
diferenciadas de músculo liso. (Nagai R y col. 1989; Loukianov y col.
1997).
Debido a los antecedentes descriptos y teniendo en cuenta que
SM2 es equivalente al isotipo B de miosina-CMP, podemos inferir que

Discusión General _____________________________________________________________________
_____________________________________________________________________ Discusión 113
en el desarrollo postnacimiento de las células MP, el isotipo B de
miosina-CMP se expresa en el momento de mayor diferenciación de las
células MP y es posible que sea fundamental para el desarrollo
morfológico y contráctil de la célula.
En los segmentos del túbulo seminífero conteniendo las células
del ciclo espermatogénico. Hemos comprobado que en el testículo de
rata adulta la dinámica de contracción de los segmentos del TS está
relaciona a los estados del epitelio seminífero que contienen. Así, en el
segmento I-VI la contracción es discreta si se compara con el
segmento VII-VIII y la desaparece en el segmento IX-XIV.
Por otro lado la expresión de los isotipos de miosina-CMP
también esta relacionada con el estado contráctil de las células MP, en
el segmento I-VI y el IX-XIV donde la contracción es escasa o nula se
expresa 40 % del isotipo A y 60 % del isotipo B. En cambio en el
segmento VII-VIII el isotipo A se expresa solo un 22 % y el B el 78 %.
En conclusión, en el segmento VII-VIII, donde los
espermatozoides están maduros y deben ser transportados a la rete
testis, el TS se contrae más activamente, las células MP cambian mas
profundamente la morfología por la contracción y producen una
elevada expresión del isotipo B de miosina-CMP.
Nuestros resultados coinciden con experimentos realizados en
otro laboratorio, donde demuestran que la estimulación con oxitocina
produce el mayor efecto contráctil en el segmento VII-VIII del TS.
(Harris y Nicholson 1998).

Discusión General _____________________________________________________________________
_____________________________________________________________________ Discusión 114
En los TS de testículos de rata adulta, la espermatogénesis es
un proceso constante, donde las células germinales están
continuamente desarrollándose desde las espermatogonias tipo A. La
reestructuración de la citoarquitectura ocurre en discretos estados del
epitelio seminífero, correlacionado con la translocación de clones de
células germinales en meiosis temprana desde el compartimiento basal
al luminal. (Fantoni y col. 1993). En este caso los testículos están bajo
cambios que rememoran algunos de los que ocurren en otros órganos
durante el desarrollo fetal. Es conocido que el efecto morfogenético de
las hormonas esta ligado a la interacción del epitelio con las células del
estroma.
Las principales células tipo somáticas del túbulo seminífero son
las células MP (mesenquimaticas) y las células epiteliales de Sertoli.
Resultados de co-cultivos in vitro realizados por Palombi y col. 2002
demostraron que las células de Sertoli y las células MP interaccionan
entre sí llevando a la formación de la lámina basal y una arquitectura
similar a la de TS. La presencia de células MP además resulta en un
incremento de la producción de “androgen binding protein” (ABP) por
las células de Sertoli. Proteínas liberadas por las células MP estimulan
la producción de ABP y transferrina por las células de Sertoli. (Palombi
y col. 2002). Las células de Sertoli producen y secretan ET-1 que
activan a las células MP que contienen los receptores ETA y ETB para
ET-1 (Fantoni y col. 1993; Tripiciano y col. 1997). Es conocido que la
expresión cíclica de la enzima convertidora de ET-1 (ECE) por las

Discusión General _____________________________________________________________________
_____________________________________________________________________ Discusión 115
células de Sertoli regula funcionalmente la contracción del TS. Se ha
comprobado que las células de Sertoli solo en el estadio IX-X del
epitelio seminífero secretan ECE, por ende aportan ET-1 a las células
MP vecinas para contraerse. (Tripiciano y col. 1999).
¿ Las células MP presentan un ciclo de diferenciación en sincronía con
el epitelio espermático?. Durante el desarrollo del testículo hemos visto
que las células MP se transforman profundamente desde el día 16 pn,
cambiando su tamaño, morfología, organización de los microfilamentos
y expresión de isotipos miosina-CMP.
En el testículo de rata adulta, las células MP de los segmentos
del TS que contienen a las células de los distintos estados del epitelio
seminífero muestran distinto comportamiento en cuanto a su tamaño,
grado de contracción y expresión de isotipos de miosina-CMP. Se
interpreta que en el estado VII-VIII, donde los espermatozoides deben
ser transportados a la rete testis, es donde más activas se encuentran
las células MP, actividad que luego pierden cuando los
espermatozoides han dejado el lumen del TS.
Hay muchas evidencias en la bibliografía de la íntima relación de
la célula de Sertoli con la célula MP, de tal modo que la célula de
Sertoli determina cuándo se activan y contraen las célula MP. Pero
para que la célula MP sea sensible a ET-1, debe encontrarse en el
segmento VII-VIII del epitelio seminífero, de lo contrario es refractaria.
A lo largo de este trabajo hemos mostrado la aparición de
rasgos diferenciales de la célula MP en el estado VII-VIII como la

Discusión General _____________________________________________________________________
_____________________________________________________________________ Discusión 116
mayor expresión del isotipo B de miosina PM e incremento de la
capacidad contráctil, por lo que interpretamos que de la misma
manera como madura la célula MP en el desarrollo del testículo, las
células MP se van diferenciado a lo largo de cada ciclo del epitelio
seminífero para ser totalmente activas en el estado VII-VIII.

CAPÍTULO VI
MATERIALES Y MÉTODOS

Capítulo VI. Materiales y Métodos _____________________________________________________________________
_____________________________________________________________________ Materiales y Métodos 118
MATERIALES Y METODOS
1. Animales y materiales biológicos
Se utilizaron ratas Wistar (Charles River) criadas en el
animalario del IHEM y fueron alimentadas ad libitum hasta que se
sacrificaron por decapitación cervical. Los animales fueron mantenidos
de acuerdo con la guía para el cuidado y uso de animales de
laboratorio del NIH (USA). Los testículos de toro y los estómagos
musculares de pollo fueron obtenidos en un matadero de la ciudad.
2. Aislamiento de túbulos seminíferos y de células del testículo
Aislamiento de túbulos seminíferos y segmentos: Testículos de
rata desprovistos de la túnica albugínea y de la arteria seminífera se
digirieron con 1 % de tripsina en buffer fosfato, 5 minutos, a 25 ° C y
se centrifugaron en centrífuga de mesa. En el sedimento quedaron los
túbulos seminíferos y en el sobrenadante las células del intersticio.
Segmentos de túbulos seminíferos con distintos estados seminíferos.
Los túbulos seminíferos en una caja de petri, se colocaron en la platina
de la lupa. Se eligieron segmentos conteniendo la mayor cantidad de
espermatozoides (segmento VI-VII) y se seccionaron con una tijera.
Además se secciono la porción que seguía al segmento VI-VIII que no
contenía espermatozoides, segmento IX-XIV, y la porción que contenía
escasos espermatozoides, segmento I-V. Los tres segmentos fueron
colocados en recipientes distintos y utilizados para los experimentos.

Capítulo VI. Materiales y Métodos _____________________________________________________________________
_____________________________________________________________________ Materiales y Métodos 119
Células mioides peritubulares: Los túbulos seminíferos desprovistos de
las células del intersticio se digieren con 0,2 mg/ml de colagenasa,
0,02 mg/ml de hialuronidasa, 2% de ácido etilendiamintetraacético
(EDTA), 1 mg/ml de albúmina, 4,5 mg/ml de HEPES, en buffer PBS
(Phosphate Buffer Saline), por 90 minutos a 34° C. La digestión se
centrifugó y en el sobrenadante quedan las células mioides. El
desprendido de las células mioides se monitoreó bajo lupa en túbulos
teñidos con nitrato de plata.
3. Visualización y medición de las células mioides peritubulares
en el túbulo seminífero
Tinción con Cloruro de Plata: Túbulos seminíferos o segmentos
con distintos estados del epitelio seminal fueron fijados con
glutaraldehído al 3% e incubados con cloruro de plata al 1% en H2O,
10 min. Se lavaron con H2O y se incubaron con solución reductora (30
% sucrosa, 0,4 M acido nítrico, 30 % alcohol etílico) por 20 min. Las
células MP fueron visualizadas con lupa estereoscópica a 20 X y
fotografíadas o gravadas para su análisis. En cada experimento una
regla graduada de referencia fue gravada en paralelo con las muestras.
El diámetro del TS y el área y los ejes x (longitudinal al eje mayor del
TS) e y (perpendicular al largo del TS) de las células MP fueron
cuantificados con el software Scion Image.
Microscopía electrónica de barrido: Los túbulos seminíferos se fijaron
con 3% de glutaraldehído en buffer cacodilato, postfijazo con 1% de
OsO4 en buffer Zetterquist, y deshidratados en 25%-100% de acetona.

Capítulo VI. Materiales y Métodos _____________________________________________________________________
_____________________________________________________________________ Materiales y Métodos 120
Luego del punto crítico con acetona se los bañó con oro y los túbulos
seminíferos se analizaron en un microscopio de barrido.
4. Tratamiento de los túbulos seminíferos con endotelina-1
Túbulos seminíferos aislados o segmentos conteniendo distintos
estados germinales fueron incubados con 0-50 nM ET-1 en medio MEM
20 segundos y las células MP fueron visualizadas por tinción con
cloruro de plata.
5. Análisis de proteínas
Electroforesis en geles de poliacrilamida. Fueron utilizados geles
de poliacrilamida 4-7,5 % con dodecil sulfato de sodio (SDS) (SDS-
PAGE), bajo condiciones de reducción. Las proteínas se corrieron en
paralelo con marcadores de peso molecular. La corrida electroforética
se efectuó de 2-10 mA.
Transferencia de proteínas: Luego de corrido el gel, las proteínas
se transfirieron a lámina de nitrocelulosa utilizando el siguiente buffer
de transferencia (0,02 M de tris-base, 0,15 M de glicina, 20 % de
alcohol metílico y 0,1 % de SDS y una corriente de 250 mA. Las
proteínas transferidas se analizaron con colorante Ponceau S.
Inmunoensayo: Se bloqueó la lámina de nitrocelulosa con 0,1 %
de albúmina de suero bovino (ASB) en buffer TTBS (0,3 M NaCl, 0,1 M
tris, 0,05 M tween 20 y 0,01 % N3Na), 90 min a 20° C. Se Lavó con
buffer TTBS e se incubó dos horas con el primer anticuerpo diluido
disuelto en 1 % ASB en TTBS. Lavar con buffer TTBS e incubar con el
segundo anticuerpo, conjugado con fosfatasa alcalina disuelto en 1 %

Capítulo VI. Materiales y Métodos _____________________________________________________________________
_____________________________________________________________________ Materiales y Métodos 121
ASB en TTBS, una hora. Lavar con TTBS y realizar la reacción de la
fosfatasa alcalina.
Reación de fosfatasa alcalina: Se incubó la lámina de
nitrocelulosa en 30 ml de solución AP (2 mg/ml de phenazine
methosulfate, con trazas de nitroblue tetrazolium y 5-Bromo-4-chloro-
3-indolyl phosphate en medio alcalino a pH 8,8). Se detuvo la reacción
por lavados con agua destilada.
Fosfatasa alcalina en suspención de células: Se utilizó el kit de
laboratorios Wiener, se les adiciona sustrato buffer al blanco, al estándar
y a la muestra, se incuban 10 minutos a 37° C y se agrega el reactivo
color. Se lee el color de la reacción en el espectrofotómetro a 520 nm.
ATPasa. La actividad K+-EDTA-ATPasa se midió en 0.2 mg/ml de
miosina purificada en 300 µl de 15 mM imidazole.ClH, pH 7.5, 0.6 M
KCl, 5mM EDTA, 1mM ATP a 20 °C. La reacción comienza por la
adición de miosina, se remueve en varios puntos de la reacción una
cantidad mínima, se lo lleva a una concentración de 0,4 N de ácido
percloroacético para detener la reacción, antes de comenzar con el
análisis espectroscópico con verde de malaquita a 600nm (Facemyer y
Cremo, 1992; Cremo y col. 1995).
Análisis de péptidos por espectroscopía de masa. Identificación
huella peptídica: Tras su localización, las bandas de interés se
recortaron de forma manual minimizando la cantidad de gel y se
digirieron in-situ en modo automático (empleando un robot-digestor de
Bruker), con tripsina utilizando el protocolo descrito por (Shevchenko y

Capítulo VI. Materiales y Métodos _____________________________________________________________________
_____________________________________________________________________ Materiales y Métodos 122
col. 1996) con algunas modificaciones (variable dependiendo del
método de tinción utilizado). El sobrenadante de la digestión (que
contiene los péptidos) se acidificó con trifluoroacético (TFA) (0.1%
concentración final) y se secó en centrífuga “speedvac” y se
resuspendió en 5 ml de TA (TFA 0.1% Acetonitrilo 33%). Una pequeña
alícuota (0.5 ml) se depositó en una placa “Anchor-chip” (Bruker)
empleando DHB (2,5-Dihydroxybenzoic acid) como matriz a una
concentración de 5 g/l mediante el método “fast evaporation”. La
placa se introdujo en un espectrómetro de masas de tipo MALDI-TOF
(matriz-assisted laser desorption ionisation/time-of-flight), modelo
Autoflex de Bruker equipado con reflector. Los espectros de masas
obtenidos se utilizaron como huella peptídica para la identificación de
proteínas en las bases de datos utilizando los motores de búsqueda
(Mascot, Profound) accesibles en la red.
6. Obtención y purificación de proteínas
Miosina II de la túnica albugínea y de la arteria seminífera del
testículo: la túnica albugínea o la arteria seminífera de testículos de
rata se congela con nitrógeno líquido, se pulveriza en un mortero y se
resuspende con buffer PM (250 mM PIPES, 5 mM de sulfato de
magnesio y 10 mM de EDTA).
Miosina II de músculo liso: Miosina de músculo liso (MII-ML) se
extrajo de estómago de pollo por modificación del método de Ikebe y
Hartshorne, 1995. El estómago muscular de pollo recién faenado se
homgeneizó en una picadora de carne en buffer A (10mM NaPO4 pH

Capítulo VI. Materiales y Métodos _____________________________________________________________________
_____________________________________________________________________ Materiales y Métodos 123
6,5, 0,3mM KCl, 1,5mM EGTA, 5mM MgCl2, 1mM ATP). Se diluyó la
muestra con 4 volúmenes de agua destilada y luego se filtró con lana
de vidrio (para eliminar lípidos). A la muestra obtenida desde el
filtrado se le agregó 10 volúmenes de agua destilada y se incubó toda
la noche a 4°C. Posteriormente se realizó una centrifugación de 15
minutos a 5.000 r.p.m. Se resuspendió el sedimento, conteniendo los
filamentos de miosina, con buffer B (0,6 M de KPO4 pH 6,5, 0,025 M
EGTA, 10 M KCl ) y se dializó 12 horas en contra de buffer C ( 0,025 M
de KPO 4, 0,6 M de KCl, 0,01 M de EGTA, 0,001 M de DTT). Se agregó
a la suspención 1 volumen de agua destilada y se centrifugó a 14.000
r.p.m. durante 30 minutos. El sobrenadante conteniendo miosina II se
conservó en 50 % glicerol a – 20 °C hasta su uso.
Obtención miosina-CMP por el método de agregación-
desagregación: Testículos de rata o de toro sin túnica albugínea ni
arteria espermática, se homogenizaron en medio PMEE (35 mM de
Pipes, 5 mM de MgSO4, 1 mM de EGTA, 0,5 mM de EDTA, pH 7,4) en
una relación 1:1. El homogenizado se centrifugó a 100.000 xg, 1 h a
4° C. El sobrenadante (S1) se incubó 30 min a 20° C y se centrifugó a
100.000g por 30 minutos sobre sacarosa al 20 % en PMEE. El
sedimento (P2) se lavó con PMEE y se centrifugó nuevamente. El
nuevo sedimento limpio (P2) se resuspendió en una solución de 4 mM
ATP en PMEE, se centrifugó y el sobrenadante (S3) se guardó a -20° C
hasta su uso.

Capítulo VI. Materiales y Métodos _____________________________________________________________________
_____________________________________________________________________ Materiales y Métodos 124
Purificación de miosina-CMP en gradiente de sacarosa: Las
proteínas obtenidas por el método de agregación-desagregación se
sembraron en la parte superior de un gradiente de sacarosa (5 % - 20
% en PMEE), se centrifugaron a 150.000 xg 8 h a 4° C. Se colectaron
fracciones desde el fondo del tubo y se analizaron en SDS-PAGE al 7,5
%.
Purificación de miosina-CMP en columna de hidroxiapatita:
Miosina-CMP de rata o de toro aislada parcialmente por el método de
agregación-desagregación, se cargaron en una columna de
hidroxiapatita en buffer 0,6 M de NaCl en PM (Pipes-Magnesio), se
purgó la columna con el mismo buffer, y se eluyeron las proteínas con
un gradiente continuo de 0-400 mM de fosfato de sodio conteniendo
0,6 M de NaCl, pH 7,4. Se colectaron las fracciones, se precipitaron las
proteínas con ácido tricloroacético (TCA) y se analizaron por SDS-PAGE
al 7,5 %.
Purificación de miosina-CPM en geles de poliacrilamida:
Muestras de miosina-cmp se corrieron en SDS-PAGE al 7,5 %, se
cortaron la bandas de miosina-CMP y se enviaron a analizar o se
solubilizaron del gel con H2O2 20 volúmenes, por doce horas a 60° C.
Anticuerpos policlonales anti miosina-CPM: Se inyectaron
conejos por vía subcutánea con una suspención de miosina-CMP
purificada por electroforesis con adyuvante de Freund completo.
Después de un mes se obtuvo suero del conejo que se conservó en
solución de glicerol al 50 % a 70° C. Prueba del anticuerpo: La

Capítulo VI. Materiales y Métodos _____________________________________________________________________
_____________________________________________________________________ Materiales y Métodos 125
actividad y especificidad de los anticuerpos obtenidos se probaron por
inmunoensayo.
7. Digestión de proteínas con tripsina
Suspensiones de miosina fueron incubadas en relación 200/1
(peso/peso) con tripsina por el tiempo decidido a 25° C. La digestión
se detuvo con 0,1 µg/ml de inhibidor de tripsina.
8. Cuantificación de proteínas
Método de Lowry: Se tomaron alícuotas de las muestras a
medir, se le adicionó reactivo alcalino de Lowry, se incubaron 15 m a
22 °C. Se adicionó reactivo de Folin, se incubaron 20-45 m y se
leyeron en el espectrofotómetro a 500 nm.
Utilización de geles: Geles de poliacrilamida al 7,5% fueron
sembrados con las siguientes concentraciones Albúmina sérica bovina
(0,5–4 µg), corridos y teñidos con Azul de Coomassie R-250. Los geles
obtenidos fueron fotografiados con una cámara digital Kodak y
grabados en un sofware propio sistema EDAS. Las fotografías se
analizaron con el programa SCION IMAGE y se determinó el área de
las bandas de SAB con los datos de la densidad óptica de cada una de
las concentraciones, para evaluar la lineabilidad. En base a los valores
obtenidos se confeccionó una curva de densidad de área de las bandas
versus la concentración de ASB. Cuantificación de miosina: Se
sembraron geles de poliacriamida al 7,5% con muestras de miosina de
concentración desconocida y SAB con concentraciones de 0,5- 2 µg.
Los datos de densidad óptica de las bandas de miosina obtenidos con

Capítulo VI. Materiales y Métodos _____________________________________________________________________
_____________________________________________________________________ Materiales y Métodos 126
SCION IMAGE se extrapolaron a los de la curva confeccionada con los
correspondientes patrones de Albúmina.
9. Precipitación de proteínas
A una suspensión de proteínas conteniendo 5 % de ácido tricloro
acético en concentración final, se incubaron una hora a 0° C, se
centrifuga a 15.000 rpm por 30 minutos a 4° C. El sedimento se
resuspendió con sample buffer y se ajustó a pH 7.
10. Agregación y desagregación de miosina
Agregación in vitro: Suspensiones de miosina de distintos
orígenes en una concentración de 150 µg/ml se dializaron por 12 horas
a 4° C o a 20° C en soluciones de 0 - 300 mM de NaCl en buffer
fosfato (25 mM de NaPO4, 2 mM de MgCl2, 300 mM de NaCl a pH 7,5).
Después de la diálisis las muestras se centrifugaron a 30.000 rpm
sobre 20 % de sacarosa en buffer fosfato. Las proteínas del sedimento
y del sobrenadante de cada muestra se analizaron por SDS-PAGE al
7,5 % y las bandas de miosina se cuantificaron en el gel.
Agregación in vivo: Túbulos seminíferos, túnica albugínea y
arteria espermática de rata se homogenizaron en relación peso en
volumen 1:1 con buffer PM con y sin 0,6M NaCl a 0° C. Las proteínas
obtenidas fueron analizadas por SDS-PAGE al 7,5% y la cantidad de
miosina fue estimada por inmunoensayo.
Desagregación por fuerza iónica o ATP. Filamentos de miosina,
formados de aproximadamente 150 µg/ml de miosina, fueron
incubados con 0-300 mM de NaCl a 20 ºC o con 0-35 mM de ATP 30

Capítulo VI. Materiales y Métodos _____________________________________________________________________
_____________________________________________________________________ Materiales y Métodos 127
min a 20 °C. Las muestras se centrifugaron a 30.000 rpm sobre 20 %
de sacarosa en buffer fosfato. Las proteínas del sedimento y del
sobrenadante de cada muestra se analizaron por SDS-PAGE al 7,5 % y
las bandas de miosina se cuantificaron en el gel.
11. Análisis de datos
Los datos fueron analizados estadísticamente con el programa
Excel y Prism y se expresaron como la media ± error estandart medio
(E.S.M) de n experimentos.

BIBLIOGRAFÍA CITADA

Bibliografía _____________________________________________________________________
_____________________________________________________________________ Bibliografía Citada 129
Bibliografía
- Alberts, B.; D.B.J. Lewis; M.R.K. Roberts and J.D. Watson. Molecular
biology of the cell. Cuarta edición, Garland Publishing, Inc. 2002
- Anthony, C. T. and M. K. Skinner. Cytochemical and biochemical
characterization of testicular peritubular myoid cells. Biol.Reprod. 40
(4):811-823, 1989.
- Bresnick, A. R. Molecular mechanisms of nonmuscle myosin-II
regulation. Curr.Opin.Cell Biol. 11 (1):26-33, 1999.
- Brinster, R. L. Male germline stem cells: from mice to men. Science
316 (5823):404-405, 2007.
- Burgess, D. R. Cytokinesis: new roles for myosin. Curr.Biol. 15
(8):R310-R311, 2005.
- Campagnolo, L.; M. A. Russo, A. Puglianiello, A. Favale, and G.
Siracusa. Mesenchymal cell precursors of peritubular smooth muscle
cells of the mouse testis can be identified by the presence of the p75
neurotrophin receptor. Biol.Reprod. 64 (2):464-472, 2001.
- Chapin,R.E., J.L. Phelps, B. E. Miller, and T. J. Gray. Alkaline
phosphatase histochemistry discriminates peritubular cells in primary
rat testicular cell culture. J.Androl 8 (3):155-161, 1987.
- Chiarenza, C., A. Filippini, A. Tripiciano, E. Beccari, and F. Palombi.
Platelet-derived growth factor-BB stimulates hypertrophy of peritubular
smooth muscle cells from rat testis in primary cultures. Endocrinology
141 (8):2971-2981, 2000.

Bibliografía _____________________________________________________________________
_____________________________________________________________________ Bibliografía Citada 130
- Clermont, Y and B. Perey. Quantitative study of the cell population of
the seminiferous tubules in immature rats. Am.J.Anat. 100 (2):241-
267, 1957.
- Clermont Y. and B. Perey. The stages of the cycle of the seminiferous
epithelium of the rat: practical definitions in PA-Schiff-hematoxylin and
hematoxylin-eosin stained sections. Rev.Can.Biol. 16 (4):451-462,
1957.
- Cohen, C., S. Lowey, R. G. Harrison, J. Kendrick-Jones, and A. G.
Szent-Gyorgyi. Segments from myosin rods. J.Mol.Biol. 47 (3):605-
609, 1970.
- Conrad, A. H., T. Jaffredo, and G. W. Conrad. Differential localization
of cytoplasmic myosin II isoforms A and B in avian interphase and
dividing embryonic and immortalized cardiomyocytes and other cell
types in vitro. Cell Motil.Cytoskeleton 31 (2):93-112, 1995.
- Craig R. and J. Megerman. Assembly of smooth muscle myosin into
side-polar filaments. J.Cell Biol. 75 (3):990-996, 1977.
- Craig R., R. Smith, and J. Kendrick-Jones. Light-chain
phosphorylation controls the conformation of vertebrate non-muscle
and smooth muscle myosin molecules. Nature 302 (5907):436-439,
1983.
- Cremo, C. R., J. R. Sellers, and K. C. Facemyer. Two heads are
required for phosphorylation-dependent regulation of smooth muscle
myosin. J.Biol.Chem. 270 (5):2171-2175, 1995.

Bibliografía _____________________________________________________________________
_____________________________________________________________________ Bibliografía Citada 131
- a.Emes C. H. and A. J. Rowe. Hydrodynamic studies on the self
association of vertebrate skeletal muscle myosin. Biochim.Biophys.Acta
537 (1):110-124, 1978.
- b. Emes C. H. and A. J. Rowe. Frictional properties and molecular
weight of native and synthetic myosin filaments from vertebrate
skeletal muscle. Biochim.Biophys.Acta 537 (1):125-144, 1978.
- Engelhardt,W.A. and M.N. Ljubimowa. Myosine and
adenosinetriphosphatase. Nature 144: 668-669. 1939.
- Facemyer K. C. and C. R. Cremo. A new method to specifically label
thiophosphorylatable proteins with extrinsic probes. Labeling of serine-
19 of the regulatory light chain of smooth muscle myosin.
Bioconjug.Chem. 3 (5):408-413, 1992.
- Fantoni, G., P. L. Morris, G. Forti, G. B. Vannelli, C. Orlando, T.
Barni, R. Sestini, G. Danza, and M. Maggi. Endothelin-1: a new
autocrine/paracrine factor in rat testis. Am.J.Physiol 265 (2 Pt
1):E267-E274, 1993.
- Filippini, A., A. Tripiciano, F. Palombi, A. Teti, R. Paniccia, M.
Stefanini, and E. Ziparo. Rat testicular myoid cells respond to
endothelin: characterization of binding and signal transduction
pathway. Endocrinology 133 (4):1789-1796, 1993.
- Filippini, A., A. Tripiciano, C. Peluso, A. R. Morena, Ziparo M.
Stefanini, and F. Palombi. ECE-1 is cyclically expressed in Sertoli cells
and mediates the spatiotemporal control of tubular contractility.
Andrologia 32 (1):61, 2000.

Bibliografía _____________________________________________________________________
_____________________________________________________________________ Bibliografía Citada 132
- Greep, R.O. and E.B. Astwood Handbook of Physiology Section 7:
Endocrinology. Editor. American Physiology Society, Washinton D.C.
USA 1975.
- Harris G.C. and H.D. Nicholson. Stage-related differences in rat
seminiferous tubule contractility in vitro and their response to
oxytocin. J.Endocrinol. 157 (2):251-257, 1998.
- Hinssen, H., J. D'Haese, J. V. Small, and A. Sobieszek. Mode of
filament assembly of myosins from muscle and nonmuscle cells.
J.Ultrastruct.Res. 64 (3):282-302, 1978.
- Hodge, T. P., R. Cross, and J. Kendrick-Jones. Role of the COOH-
terminal nonhelical tailpiece in the assembly of a vertebrate nonmuscle
myosin rod. J.Cell Biol. 118 (5):1085-1095, 1992.
- a. Kendrick-Jones, J., R. Jakes, P. Tooth, R. Craig, and J. Scholey.
Role of the myosin light chains in the regulation of contractile activity.
Soc.Gen.Physiol Ser. 37:255-272, 1982.
- b. Kendrick-Jones, J., P. Tooth, K. A. Taylor, and J. M. Scholey.
Regulation of myosin-filament assembly by light-chain
phosphorylation. Cold Spring Harb.Symp.Quant.Biol. 46 Pt 2:929-938,
1982.
- c. Kendrick-Jones, J., K. A. Taylor, and J. M. Scholey.
Phosphorylation of nonmuscle myosin and stabilization of thick
filament structure. Methods Enzymol. 85 Pt B:364-370, 1982.
- Kühne, W. Untersuchungen über das Protoplasma und die
Contractilität. Leipzig:von Wilhelm Engelmann. 1864

Bibliografía _____________________________________________________________________
_____________________________________________________________________ Bibliografía Citada 133
- Landsverk M. L. and H. F. Epstein. Genetic analysis of myosin II
assembly and organization in model organisms. Cell Mol.Life Sci. 62
(19-20):2270-2282, 2005.
- a.Leblond C. P. and Y. Clermont. Definition of the stages of the cycle
of the seminiferous epithelium in the rat. Ann.N.Y.Acad.Sci. 55
(4):548-573, 1952.
- b.Leblond C. P. and Y. Clermont. Spermiogenesis of rat, mouse,
hamster and guinea pig as revealed by the periodic acid-fuchsin
sulfurous acid technique. Am.J.Anat. 90 (2):167-215, 1952.
- Lehman W. Thick-filament-linked calcium regulation in vertebrate
striated muscle. Nature 274 (5666):80-81, 1978.
- Lodish, H.; A. Berk; S.L. Zipursky; P. Matsudaira; D. Baltimore and J.
Darnell. Biología Celular y Molecular. Editorial Panamericana 2002.
- Loukianov, E., T. Loukianova, and M. Periasamy. Myosin heavy chain
isoforms in smooth muscle. Comp Biochem.Physiol B Biochem.Mol.Biol.
117 (1):13-18, 1997.
- Maekawa, M., H. Kazama, K. Kamimura, and T. Nagano. Changes in
the arrangement of actin filaments in myoid cells and Sertoli cells of
rat testes during postnatal development and after experimental
cryptorchidism. Anat.Rec. 241 (1):59-69, 1995.
- McKenna, N.M; Y.L. Wang and M.E. Konkel. Formation and
movement of myosin-containing structures in living fibroblasts. J.Cell
Biol. 109:1163-1172 1989.

Bibliografía _____________________________________________________________________
_____________________________________________________________________ Bibliografía Citada 134
- McLachlan A. D. and M. Stewart. Periodic charge distribution in the
intermediate filament proteins desmin and vimentin. J.Mol.Biol. 162
(3):693-698, 1982.
- Nagai,R., Kuro-o M, P. Babij, and M. Periasamy. Identification of two
types of smooth muscle myosin heavy chain isoforms by cDNA cloning
and immunoblot analysis. J.Biol.Chem. 264 (17):9734-9737, 1989.
- a. Oakberg, E. F. Duration of spermatogenesis in the mouse and
timing of stages of the cycle of the seminiferous epithelium.
Am.J.Anat. 99 (3):507-516, 1956.
- Okagaki, T., Nakamira, A., Suzuki,T., Ohmi, K. and Kohama, K.
Assembly of smooth muscle myosin by the 38 k protein, a homologue
of a subunit of pre-mRNA splicing factor-2. J.Cell Biol. 148: 653-663.
2000
- b. Oakberg, E. F. A description of spermiogenesis in the mouse and
its use in analysis of the cycle of the seminiferous epithelium and germ
cell renewal. Am.J.Anat. 99 (3):391-413, 1956.
- Olney, J.J., J.R. Sellers, and C.R. Cremo. Structure and function of
the 10s conformation of smooth muscle myosin. J.Biol.Chem. 271:
20375-20384. 1996.
- Onishi, H., Y. Yamada, M. Ikebe, H. Suzuki, and S. Watanabe. The
ATPase reaction in the steady state and in the initial burst catalyzed by
chicken gizzard myosin in 0.6 M KCl1. J.Biochem.(Tokyo) 83 (1):129-
135, 1978.

Bibliografía _____________________________________________________________________
_____________________________________________________________________ Bibliografía Citada 135
- Opgenorth, T.J., J.R. Wu-Wong, and K. Shiosaki. Endothelin-
converting enzymes. FASEB J. 6 (9):2653-2659, 1992.
- Palombi F. and Carlo C. Di. Alkaline phosphatase is a marker for
myoid cells in cultures of rat peritubular and tubular tissue.
Biol.Reprod. 39 (5):1101-1109, 1988.
- Palombi, F., D. Farini, M. Salanova, Grossi S. de, and M. Stefanini.
Development and cytodifferentiation of peritubular myoid cells in the
rat testis. Anat.Rec. 233 (1):32-40, 1992.
- Palombi, F., A. Filippini, and C. Chiarenza. Cell-cell interactions in the
local control of seminiferous tubule contractility. Contraception 65
(4):289-291, 2002.
- Parry, D. A. Structure of rabbit skeletal myosin. Analysis of the amino
acid sequences of two fragments from the rod region. J.Mol.Biol. 153
(2):459-464, 1981.
- Perey, B., Y.Clermont and C.P. Leblond. The wave of the
seminiferous epithelium in the rat. Am.J. Anat. 108: 47-78. 1961
- Prince, H. P., H. R. Trayer, G. D. Henry, I. P. Trayer, D. C. Dalgarno,
B. A. Levine, P. D. Cary, and C. Turner. Proton nuclear-magnetic-
resonance spectroscopy of myosin subfragment 1 isoenzymes.
Eur.J.Biochem. 121 (1):213-219, 1981.
- Redowicz, M. J. Unconventional myosins in muscle. Eur.J.Cell Biol. 86
(9):549-558, 2007.
- Rossi, F., A. Ferraresi, P. Romagni, L. Silvestroni, and V. Santiemma.
Angiotensin II stimulates contraction and growth of testicular

Bibliografía _____________________________________________________________________
_____________________________________________________________________ Bibliografía Citada 136
peritubular myoid cells in vitro. Endocrinology 143 (8):3096-3104,
2002.
- Russell, L.D., A. Bartke and J.C. Goh. Postnatal development of the
Sertoli cell barrier, tubular lumen, and cytoskeleton and Sertoli and
myoid cells in the rat, and their relationship to tubular fluid secretion
and flow. Am.J. Anat 184:179-189, 1989
- Russell, L.D., R.A. Ettlin; A.P. Sinhahikim and E.D. Clegg. Histological
and Histopathological Evaluation of the testis. Cache River Press. 1990
- Saab, S., S. P. Tam, B. N. Tran, A. C. Melton, P. Tangkijvanich, H.
Wong, and H. F. Yee, Jr. Myosin mediates contractile force generation
by hepatic stellate cells in response to endothelin-1. J.Biomed.Sci. 9 (6
Pt 2):607-612, 2002.
- Scholey, J.M., K. A. Taylor, and J. Kendrick-Jones. Regulation of non-
muscle myosin assembly by calmodulin-dependent light chain kinase.
Nature 287 (5779):233-235, 1980.
- Shevchenko, A., M. Wilm, O. Vorm, O. N. Jensen, A. V.
Podtelejnikov, G. Neubauer, A. Shevchenko, P. Mortensen, and M.
Mann. A strategy for identifying gel-separated proteins in sequence
databases by MS alone. Biochem.Soc.Trans. 24 (3):893-896, 1996.
- Shirinsky, V.P., A.V. Vorotnikov, K.G. Birukov, A.K. Nanaev, M.
Collinge, T.J. Lukas, J.R. Sellers and D.M. Watterson. A kinase-related
protein stabilizes unphosphorylated smooth muscle myosin
minifilaments in the presence of ATP. J.Biol.Chem. 268: 16578-16583.
1993

Bibliografía _____________________________________________________________________
_____________________________________________________________________ Bibliografía Citada 137
- Silver, D.L., A.V. Vorotnikov, D.M. Watterson, V.P. Shirinsky, and
J.R. Sellers. Sites of interaction between kinase-related protein and
smooth muscle myosin. J.Biol.Chem. 272: 25353-25359. 1997.
- Suzuki, H., H. Onishi, K. Takahashi, and S. Watanabe. Structure and
function of chicken gizzard myosin. J.Biochem.(Tokyo) 84 (6):1529-
1542, 1978.
- Szent-Gyorgyi, A. Studies from the Institute of Medical Chemistry,
University Szeged, vol. I-III. New York: Karger 1942.
Szuchet, S. Effect of purification procedures on the self-association of
myosin at high ionic strength. Arch.Biochem.Biophys. 180 (2):493-
503, 1977.
- Trinick, J. and J.Cooper. Sequential disassembly of vertebrate muscle
thick filaments. J.Mol.Biol. 141 (3):315-321, 1980.
- Tripiciano, A., A. Filippini, Q. Giustiniani, and F. Palombi. Direct
visualization of rat peritubular myoid cell contraction in response to
endothelin. Biol.Reprod. 55 (1):25-31, 1996.
- Tripiciano, A., F. Palombi, E. Ziparo, and A. Filippini. Dual control of
seminiferous tubule contractility mediated by ETA and ETB endothelin
receptor subtypes. FASEB J. 11 (4):276-286, 1997.
- Tripiciano, A., A. Filippini, F. Ballarini, and F. Palombi. Contractile
response of peritubular myoid cells to prostaglandin F2alpha. Mol.Cell
Endocrinol. 138 (1-2):143-150, 1998.
- Tripiciano, A., C. Peluso, A. R. Morena, F. Palombi, M. Stefanini, E.
Ziparo, M. Yanagisawa, and A. Filippini. Cyclic expression of

Bibliografía _____________________________________________________________________
_____________________________________________________________________ Bibliografía Citada 138
endothelin-converting enzyme-1 mediates the functional regulation of
seminiferous tubule contraction. J.Cell Biol. 145 (5):1027-1038, 1999.
- Trybus, K.M., T.W. Huiatt, and S. Lowey. A bent monomeric
conformation of myosin from smooth muscle. Proc.Natl.Acad.Sci.U.S.A
79 (20):6151-6155, 1982.
- Tung, P. S. and I. B. Fritz. Characterization of rat testicular
peritubular myoid cells in culture: alpha-smooth muscle isoactin is a
specific differentiation marker. Biol.Reprod. 42 (2):351-365, 1990.
- Tung, P. S. and I. B. Fritz. Characterization of rat testicular
peritubular myoid cells in culture: alpha-smooth muscle isoactin is a
specific differentiation marker. Biol.Reprod. 42 (2):351-365, 1990.
- a.Ueno H. and W. F. Harrington. Conformational transition in the
myosin hinge upon activation of muscle. Proc.Natl.Acad.Sci.U.S.A 78
(10):6101-6105, 1981.
- b.Ueno H. and W. F. Harrington. Cross-bridge movement and the
conformational state of the myosin hinge in skeletal muscle. J.Mol.Biol.
149 (4):619-640, 1981.
- Virtanen, I., M. Kallajoki, O. Narvanen, J. Paranko, L. E. Thornell, M.
Miettinen, and V. P. Lehto. Peritubular myoid cells of human and rat
testis are smooth muscle cells that contain desmin-type intermediate
filaments. Anat.Rec. 215 (1):10-20, 1986.
- Wagner P.D. and D.B. Stone. Myosin heavy chain-light chain
recombinations and interactions between the two classes of light
chains. J.Biol.Chem. 258 (14):8876-8882, 1983.

Bibliografía _____________________________________________________________________
_____________________________________________________________________ Bibliografía Citada 139
- Wagner P.D. and D.B.Stone. Calcium-sensitive binding of heavy
meromyosin to regulated actin requires light chain 2 and the head-tail
junction. Biochemistry 22 (6):1334-1342, 1983.
- Xu, J. Q., B. A. Harder, P. Uman, and R. Craig. Myosin filament
structure in vertebrate smooth muscle. J.Cell Biol. 134 (1):53-66,
1996.

PUBLICACIONES

Publicaciones _____________________________________________________________________
_____________________________________________________________________ Publicaciones 141
Publicaciones relacionadas con los resultados presentados
- Identification and characterization of myosin from testicular
peritubular myoid cells. Darío Fernández, Laura Gómez, Eduardo
Callegari y Luis A López. Manuscrito enviado
- Isotipos de miosina-CMP en células mioides peritubulares en el
desarrollo del testículo. En redacción
- Relación de los isotipos de miosina-CMP en la contracción de las
células MP en los estadíos de diferenciación del epitelio
seminífero. En redacción.
Copyright © 2022 FDOKUMEN




















