
Alteraciones del acoplamiento excitación-contracción y la ...
135
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez. Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Alteraciones del acoplamiento excitación-contracción y la ...

Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Universitat d'Alacant Universidad de Alicante
Tesis Doctoral
Alteraciones del acoplamiento excitación-contracción y la función del factor de
crecimiento similar a la insulina-I en el deterioro del músculo esquelético con el
envejecimiento
Directores: Osvaldo Delbono Andrés Morales
UNIVERSITAT D'ALACANT C E D í P
o 1 OCT. 200/;
m....\ Niim
Estela González Rodríguez Alicante, septiembre, 2004
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Universitat d'Alacant Í ¿ ^ Universidad de Alicante
Departament de Fisiología, Genética i Microbiología Departamento de Fisiología, Genética y Microbiologí
Dr. Osvaldo Delbono, Catedrático del Departamento de Fisiología y Farmacología de Wake Forest * University Scliool of Medicine (Winston-Salem, Carolina del Norte) y Dr. Andrés Morales, Profesor Titular del Departamento de Fisiología, Genética y Microbiología de la Universidad de Alicante,
CERTIFICAN: Que el trabajo de investigación conducente a la obtención del grado de Doctor titulado: "Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento similar a la insulina-I en el deterioro del músculo esquelético con el envejecimiento", de la que es autora Dña. Estela González Rodríguez, ha sido realizado bajo su dirección en los Departamentos de Fisiología y Farmacología de Wake Forest University School of Medicine y de Fisiología, Genética y Microbiología de la Universidad de Alicante.
Y para que así conste, y surta los efectos oportunos, firman el presente certificado en Alicante, a dieciséis de septiembre de dos mil cuatro.
jiW^ áldo Delbono Andrés Morales
Tel, + 34 965 90 9494- Fax + 34 965 90 9569 Campus de Sant Vicent del Raspeig
Ap. 99. E-03080 Alacant e-mail; [email protected]
web: http.//www,ua.es/fgm
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

AGRADECIMIENTOS
Al Dr. Osvaldo Delbono, por toda la paciencia y la dedicación demostrada a lo
largo de estos años. Agradezco sobre todo, su cálida acogida, la oportunidad
de haber trabajado en su laboratorio y la comprensión que ha manifestado
siempre hacia mi falta de conocimientos e inexperiencia. Al Dr. Andrés
Morales por su apoyo incondicional y por todo el tiempo dedicado a este
trabajo. La pasión y dedicación de ambos por la ciencia me ha animado y me
ha provisto de ejemplos en los cuales modelar mi propia carrera científica.
Mis compañeros de trabajo, en el laboratorio del Dr. Delbono, se merecen un
reconocimiento muy especial; María-Laura Messí, Zhenlin Zheng, ZhongMin
Wang, HongQu Shan y Anthony Payne. Todos han sido y son unos
compañeros y amigos excelentes. Agradezco a Laura por llevar a cabo los
experimentos de SDS PAGE e inmunohistoquímica para la determinación del
tipo de fibras y a Zhenlin por las determinaciones de IGF-1 en músculo.
Mis familiares y amigos han llenado mi carrera de post-grado de memorias
maravillosas.
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

índice
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

ÍNDICE
Lista de abreviaturas v/
1. INTRODUCCIÓN
1.1. Estructura y función del músculo esquelético 2 1.2. Acoplamiento excitación-contracción en músculo esquelético 5 1.3. Subtipos de fibras musculares y tipos de contracción 8 1.4. Importancia de la pérdida de función muscular con la edad 8 1.5. Mecanismos responsables de la pérdida de la función muscular
con el envejecimiento 10 1.6. El desacoplamiento de EC como mecanismo responsable de
la debilidad muscular 15 1.7. La técnica de fibra única e intacta como excelente aproximación
experimental para determinar la fuerza específica del músculo esquelético 16
1.8. Eventos elementales de la liberación de Ca^*(EELC) -Chispas de Ca^^ del RS 17
1.9. El músculo esquelético y el factor de crecimiento similar a la insulina de tipo I (IGF-1) 19
2. OBJETIVOS 21
3. MATERIAL Y MÉTODOS
3.1. Disección y montaje de fibras únicas intactas 25 3.2. Registros de contracción de fibra única 26 3.3. Registros de contracción de los experimentos de fatiga 27 3.4. Experimentos de contracción de músculo entero 27 3.5. Registros de Ca " intracelular en fibras únicas intactas de FDB 28 3.6. Calibración del indicador Fluo 3-AM y cálculo de la
concentración intracelular de Ca^* 29 3.7. Análisis del tipo de fibra en músculo esquelético 31 3.8. Medida de la concentración de IGF-1 en músculo FDB 32 3.9. Preparación de las fibras musculares para el registro de
chispas de Ca^" 32 3.10. Imágenes de fluorescencia de las chispas Ca^* 33 3.11. Análisis de las imágenes de chispas de Ca "" 34 3.12. Composición de cadena pesada de miosina (CPM) 36 3.13. Preparación de las membranas e Immunoblots del RyR 36 3.14. Análisis estadístico 38
4. RESULTADOS
4.1. Efecto del envejecimiento sobre la fuerza específica de células únicas intactas de los músculos extensor común de los dedos (EDL) y soleo de ratón disminuye
III
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

4.1.1. La fuerza específica máxima en células de músculos rápidos y lentos 41
4.1.2. Cinética de contracción de fibras de EDL y soleo de ratones jóvenes, de mediana edad y viejos 45
4.2. índice de fatiga en fibras musculares del EDL y Soleo de ratones jóvenes y viejos
4.2.1. índice de fatiga en fibras musculares de EDL y soleo de ratones jóvenes y viejos 48
4.2.2. Curso temporal del índice de fatiga de fibras de EDL y soleo de ratones jóvenes y viejos 52
4.3. El factor de crecimiento similar a la insulina-I previene la disminución de Ca^^ intracelular y fuerza específica en fibras únicas intactas de músculo de ratones transgénicos
4.3.1. La expresión de IGF-1 en músculos FDB de ratones transgénicos 56
4.3.2. La fuerza específica de músculo FDB entero de ratones normales y lGF-1-transgénicos 57
4.3.3. Área de la sección transversal y fuerza específica máxima registrada en células únicas intactas del músculo FDB 58
4.3.4. Valores máximos de [Ca " ]! en fibras únicas intactas del músculo FDB 64
4.3.5. Efecto de la cafeína en la disminución de la fuerza específica y en el pico de [Ca^""]! con el envejecimiento 65
4.3.6. Tipo de fibras que componen el músculo FDB a distintas edades en ratones control y transgénicos
(IGF-1) 67
4.4. Eventos elementales de liberación de Ca^^: "chispas" y "ascuas" en fibras de músculo esquelético de ratones viejos
4.4.1. Eventos de liberación de Ca " en imágenes XY y de escan de línea 71
4.4.2. Efecto de los moduladores de RyRs en la frecuencia de las chispas de Ca^^ 73
4.4.3. Propiedades de los EELC identificados en imágenes X-Y o de escan de línea en fibras de ratones jóvenes y viejos 75
4.4.4. Eventos tipo "ascua" en imágenes de escan de línea en fibras de EDL de ratones jóvenes y viejos 80
4.4.5. Subtipos de fibras musculares y frecuencia de las chispas de Ca * 82
4.4.6. Función mitocondrial y frecuencia de las chispas de Ca "" en fibras musculares de ratones jóvenes y viejos 83
4.4.7. RyRI expression in EDL muscle from ageing mice 84
IV
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

g DISCUSIÓN
5.1. Efecto del envejecimiento sobre la fuerza específica de células únicas intactas de los músculos extensor común de los dedos (EDL) y soleo de ratón disminuye
5.1.1 La fuerza específica de fibras intactas de los músculos esquelético EDL y soleo de ratón disminuye con la edad 88
5.1.2. Mecanismos responsables de la disminución de fuerza específica en músculo esquelético con el envejecimiento 89
5.1.3. Cinética de contracción en fibras de EDL y soleo de ratones jóvenes, de mediana edad y viejos 91
5.2. índice de fatiga en fibras musculares del EDL y soleo de ratones jóvenes y viejos
5.2.1. Fatiga muscular en ratón y otras especies 94 5.2.2. Fatiga en fibras de los músculos EDL y soleo 95 5.2.3. Mecanismos responsables de la alteración del
funcionamiento neuromuscular con el envejecimiento 95
5.3. El factor de crecimiento similar a la insulina-I previene la disminución de Ca^* intracelular y fuerza específica en fibras únicas intactas de músculo de ratones transgénicos
5.3.1. IGF-1 previene la disminución de fuerza específica con el envejecimiento 98
5.3.2. Mecanismos responsables de la disminución de fuerza específica de fibras musculares con el envejecimiento 100
5.3.3. Mecanismos del efecto de IGF-1 en ratones IGF-1-transgénicos 102
5.4. Eventos elementales de liberación de Ca^*: "chispas" y "ascuas" en fibras de músculo esquelético de ratones viejos
5.4.1. EELC en músculo esquelético de ratón adultos 105 5.4.2. Efectos del envejecimiento en las propiedades
de los EELC 107
6. CONCLUSIONES 113
7 BIBLIOGRAFÍA 115
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Abreviaturas
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

LISTA DE ABREVIATURAS
AMM
ATP
AST
Ca *
CPM
CICR
DE
DEC
DHPR
DMM
EC
EEM
EDL
EELC
FE
FDB
IGF-I
MA
RDI
RS
RyR
SERCA
TMB
Anchura a la mitad del máximo
Adenosina trifosfato
Área de la sección transversal
Ion calcio
Cadena pesada de miosina
Liberación de Ca "" inducida por Ca ""
Desviación estándar
Desacoplamiento de excitación-contracción
Receptor de la dihidropiridina
Duración a la mitad del máximo
Excitación-contracción
Error estándar de la media
Extensor común de los dedos {Extensor digitorum longus)
Eventos elementales de liberación de calcio
Fuerza específica
Flexor corto de los dedos {Flexor digitorum brevis)
Factor de crecimiento similar a la insulina-I
Mediana edad
Región de interés
Retículo sarcoplasmático
Receptor de la rianodina
ATPasa de Ca "" del retículo endoplasmático
Tetrametilbenzidina
vn
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

^ Túbulos-T TT ^ Sensor de voltaje
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

1. Introducción
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Introducción
1. INTRODUCCIÓN
1.1, Estructura y función del músculo esquelético
Los músculos permiten a los animales convertir energía química, en forma de
ATP, en energía mecánica. Atendiendo a criterios morfofuncionales los músculos de los
vertebrados se pueden clasificar en dos tipos: i) músculo estriado, que presenta
estriaciones características producidas por grupos organizados de filamentos de actina
y miosina que forman los sarcomeres; dentro de este grupo se incluye el músculo
cardiaco, que forma parte de la pared del corazón y se caracteriza, entre otras cosas,
por carecer de innervación específica para cada fibra y generar potenciales de acción
de muy larga duración; y el músculo esquelético, en el que cada fibra recibe inervación
de una motoneurona y , como su nombre indica, está unido al esqueleto; y ii) músculo
liso, localizado en las paredes de los órganos huecos, excepto el corazón, y
caracterizado por carecer de las típicas estriaciones del músculo esquelético y generar
contracciones más lentas.
El músculo estriado esquelético está rodeado de una cubierta de tejido conectivo
denominado epimisio. Además, bandas de fibras musculares y las propias células
están delimitadas por tejido conectivo que constituye, respectivamente, el perimisio y el
endomisio. A través de este tejido fibroso discurren los vasos sanguíneos, linfáticos y
nervios musculares. Adicionalmente, el tejido conectivo en el músculo genera
estructuras más fuertes como los tendones, aponeurosis, rafes y capa reticular de la
dermis o periostio, que proveen soporte para la inserción y contracción del músculo.
Las fibras musculares esqueléticas son células multinucleadas cilindricas de una
gran longitud (pueden alcanzar varios centímetros) y un diámetro que puede variar
entre 5 y 100 |am. Las fibras musculares están rodeadas de una membrana
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Introducción
denominada sarcolenna y contienen miofibrillas formadas por las proteínas contráctiles
como la miosina, actina, tropomiosina, el complejo troponina y proteínas de soporte
estructural como la actinina y tinina. La organización espacial específica de los
filamentos contráctiles finos (constituidos por actina, tropomiosina y el complejo
troponina) y gruesos (formados por miosina) es responsable de la estriación cruzada
característica de los músculos estriados. Las diferencias en el índice de refracción de
distintas regiones de la fibra muscular determinan la alternancia de bandas claras,
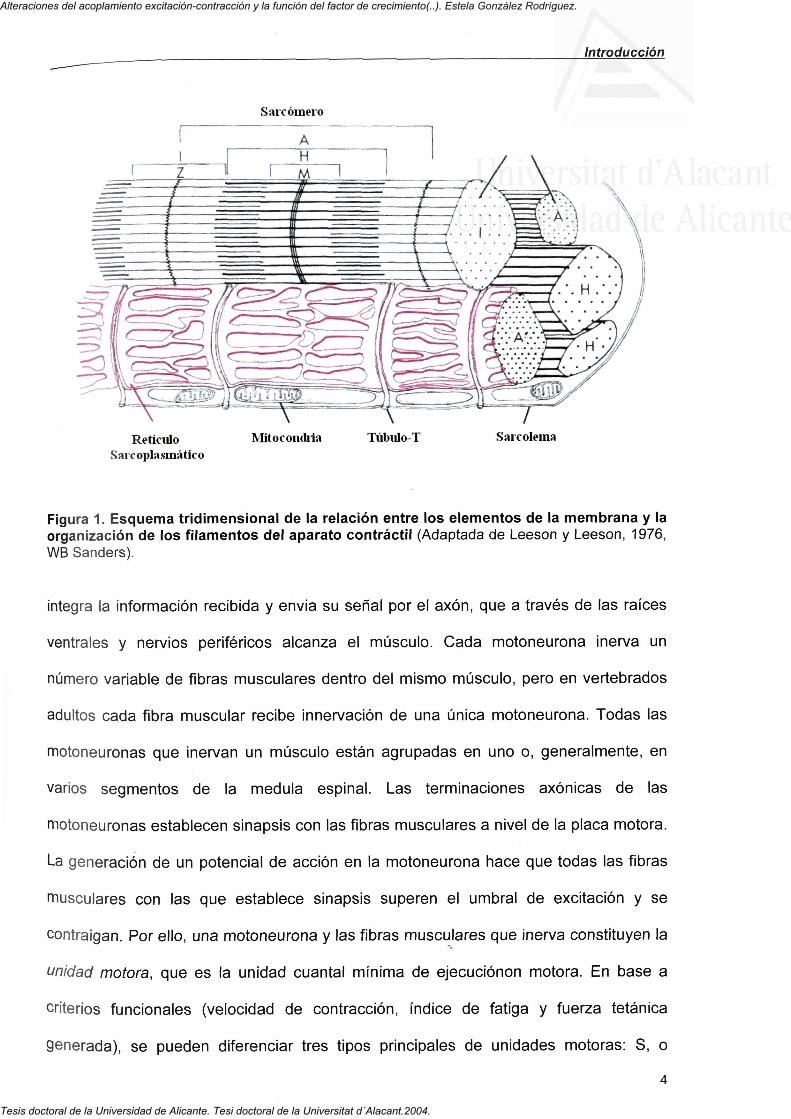
isótropas o bandas " I " y bandas oscuras, anisótropas o bandas "A". La figura 1 ilustra la
organización de las bandas I y A en la sección de una fibra única. Las líneas Z,
localizadas en el medio de las bandas I determinan el límite del sarcomere. Los
filamentos gruesos están formados por la miosina, proteína con capacidad de unirse a
la actina que posee una unidad catalítica con un sitio que hidroliza ATP, la fuente de
energía para la contracción muscular. Las moléculas de actina-G se polimerizan en
filamentos largos y finos formando una hélice con hendiduras donde se localizan las
moléculas de tropomiosina. Las moléculas de troponina se sitúan a intervalos regulares
a lo largo de las moléculas de tropomiosina. El complejo troponina tiene tres
componentes: troponina T que se une a los componentes de la tropomiosina; troponina
I que inhibe la interacción entre miosina y actina y troponina C que contiene 4 sitios
HLH para unión de Ca "", que es el responsable del inicio de la contracción muscular
(Loeser y Delbono, 2003).
1.1.1. La Unidad Motora
Las motoneuronas, localizadas a nivel de los núcleos motores troncoencefálicos
y en el asta ventral de la médula espinal, constituyen la vía final común para la
transmisión de señales de centros motores al músculo esquelético. La motoneurona
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.
Introducción
Sarcómero
Retículo Sarc opla siná tic o
Mítocondiia Tíibiüo-T Saicoleina
Figura 1. Esquema tridimensional de la relación entre los elementos de la membrana y la organización de los filamentos del aparato contráctil (Adaptada de Leeson y Leeson, 1976, WB Sanders).
integra la información recibida y envia su señal por el axon, que a través de las raíces
ventrales y nervios periféricos alcanza el músculo. Cada motoneurona inerva un
número variable de fibras musculares dentro del mismo músculo, pero en vertebrados
adultos cada fibra muscular recibe innervación de una única motoneurona. Todas las
motoneuronas que inervan un músculo están agrupadas en uno o, generalmente, en
varios segmentos de la medula espinal. Las terminaciones axónicas de las
motoneuronas establecen sinapsis con las fibras musculares a nivel de la placa motora.
La generación de un potencial de acción en la motoneurona hace que todas las fibras
musculares con las que establece sinapsis superen el umbral de excitación y se
contraigan. Por ello, una motoneurona y las fibras musculares que inerva constituyen la
unidad motora, que es la unidad cuantal mínima de ejecuciónon motora. En base a
criterios funcionales (velocidad de contracción, índice de fatiga y fuerza tetánica
generada), se pueden diferenciar tres tipos principales de unidades motoras: S, o
4
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Introducción
lentas; FR, o resistentes a fatiga; y FF, o de fatiga rápida. Las diferencias funcionales
(Je los distintos tipos de unidades motoras están basadas en diferentes propiedades
bioquímicas y estructurales de las fibras musculares que las constituyen (Loeser y
Delbono, 2003).
1.2. Acoplamiento excitación-contracción en músculo esquelético
La secuencia de eventos que tienen lugar para la conversión de una señal
eléctrica (potencial de acción) de la fibra muscular en desarrollo de fuerza se define
como acoplamiento de excitación-contracción (EC). El potencial de acción en la fibra
muscular se genera como consecuencia de la liberación de acetilcolina por el terminal
nervioso a nivel de la placa motora, unión del neurotransmisor a receptores nicotínicos
de acetilcolina en el elemento post-sináptico (membrana de la fibra muscular o
sarcolema), y aumento a este nivel de las conductancias al sodio y potasio
responsables de los potenciales sinápticos que, si superan el umbral de excitación,
generan la respuesta activa. El potencial de acción muscular es conducido a lo largo
del sarcolema y por medio de los túbulos-T (Fig. 2) se distribuye la excitación hasta el
interior de la fibra muscular. La transducción de la señal eléctrica en mecánica requiere
de la participación de una proteína localizada en la membrana de los túbulos T, el
receptor de dihidropiridina (DHPR). El DHPR es un canal de Ca "" dependiente del
voltaje, subtipo L, que al ser activado por la despolarización que acompaña al potencial
de acción sufre un cambio de conformación, asociado a movimientos de carga dentro
de la membrana y genera una corriente de Ca^* de entrada (Fig.3). Se cree que la
interacción física entre el DHPR y el receptor de la rianodina (RyR1), un canal de
liberación de Ca "" localizado en la membrana del retículo sarcoplasmático (RS), como
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Introducción
Motoneurona
Triada
Figura 2. Propagación del potencial de acción a lo largo del sarcolema y en los túbulos-T. (Adaptada de Heiny, J.A. 2001)
consecuencia del cambio conformacional del DHPR, produce la activación del RyR1 y
liberación masiva de Ca " de las cisternas terminales del RS (Schneider y Chandler,
1973). El Ca "" se une a la troponina C y esta por medio de la troponina T desplaza la
tropomiosina hacia un lado, exponiendo los sitios de unión de la actina y permitiendo la
formación de los puentes cruzados entre la miosina y los filamentos de actina, que
median el deslizamiento de los filamentos delgados sobre los gruesos y la generación
de fuerza (Looser y Delbono, 2003). Por todo esto, el Ca "" juega un papel muy
importante en el desarrollo de la fuerza muscular. La figura 3 representa los pasos del
proceso de acoplamiento EC en fibras musculares esqueléticas de mamíferos. La
corriente de Ca "" de entrada es debida a la activación de los DHPRs localizados en la
membrana de los túbulos-T. El movimiento de carga es el resultado del desplazamiento
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Introducción
Voltaje de membrana fvn
-80 mV
MoElamentos
Figura 3. Acoplamiento de excitación-contracción en músculo esquelético de mamíferos. La corriente de Ca^* entra a través del receptor de dihidropiridina (DHPR) localizado en la membrana de los túbulos T. El movimiento de carga es el resultado del movimiento de la unidad del DHPR que actúa como sensor de voltaje. La liberación de Ca "" del retículo sarcoplasmático (RS) ocurre a través del receptor de la rianodina (RyRI) y produce un aumento transitorio de Ca "" en el citoplasma. El Ca "" citoplasmático produce la contracción mediante la activación del ciclo de los puentes cruzados. Barras de calibración vertical: 5 nA, 10 ijMms"\ 5 fjM, y 200 kPa para la corriente de Ca "" y movimiento de carga, liberación de Ca * del RS, transitorio de Ca "" y fuerza, respectivamente (Payne y cois., 2004); adaptada de Melzer, W., A. Herrmann-Frank, y H. Luttgau. Biochim.Biophys.Acta 1241:59-116, 1995]
de la subunidad del DHPR que actúa como sensor de voltaje. La liberación de Ca^* del
RS ocurre a través de la isoforma muscular del receptor de la rianodina (RyRI), cuya
activación produce el aumento transitorio de la [Ca^*] mioplasmático (transitorio de
Ca^*). El incremento de la [Ca^^l resulta finalmente en la formación de los puentes
cruzados y la generación de fuerza.
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Introducción
1.3. Subtipos de fibras musculares y tipos de contracción
Se han identificado distintos tipos de fibras musculares mediante tinciones
inmunológicas o de ATPasa, basadas en la dependencia del pH de la actividad ATPasa
de la miosina, que se correlaciona con las propiedades antigénicas de la miosina en
musculo de contracción rápida y lenta (ver Tabla 1). De acuerdo con la clasificación
histoquímica, las fibras musculares están divididas en tipo I y tipo II (A, B y X) (Tabla 1).
Las fibras humanas se clasifican en rápidas y lentas en base a sus propiedades de
contracción en respuesta a una estimulación aislada (genera un único potencial de
acción). Las fibras rápidas, también denominadas tipo II, glicolíticas o blancas, se
contraen en menos de 10 ms y participan mayoritariamente en movimientos rápidos,
finos y precisos. Las fibras lentas, también denominadas tipo I, oxidativas o rojas,
muestran un tiempo de contracción de hasta 100 ms y participan en movimientos
sostenidos y menos precisos. En contra de lo pensado en el pasado, las fibras
musculares son estructuras dinámicas con una capacidad de adaptación excepcional.
Sus perfiles fenotípicos pueden ser afectados tanto por la inervación, el ejercicio,
sobrecarga mecánica e inactividad, hormonas y envejecimiento (Greenlund y Nair,
2003; Pette y Staron, 2001).
1.4. Importancia de la pérdida de la función muscular con la edad
La población de ancianos esta creciendo a nivel mundial. En la actualidad, en los
Estados Unidos solamente hay alrededor de 39 millones de individuos mayores de 65
años de edad y este número sigue creciendo, aumentando así la demanda y los gastos
del cuidado a largo plazo. En el año 2001, la administración del cuidado y la salud de
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Introducción
Tabla 1.
Clasificación histoquímica de las fibras musculares
Subtipos de Características
fibras funcionales e
musculares histoquímicas
Tinción de ATPasa
Tipo 1
Tipo HA
Tipo MB
Tipo IIX
Contracción
lenta, oxidativa
Contracción
rápida,
oxidativa,
glicolítica
Contracción
rápida,
glicolítica
Fetal
pH9.4
Clara
Oscura
Oscura
Oscura
pH4.6
Clara
Clara
Oscura
Oscura
pH4.3
Oscura
Clara
Oscura
Oscura
Adaptada de (Loeser y Delbono, 2003)
EEUU gastó aproxinnadamente 103 billones de dólares en residencias para la tercera
edad y este gasto aumentara hasta mas de 183 billones (un aumento del 77 %) en el
2010. Por lo tanto, el envejecimiento representa un tema muy relevante tanto desde el
punto de vista de la salud como socioeconómico. La asistencia a residencias de la
tercera edad es requerida con mayor frecuencia por ancianos que no son capaces de
cuidar de si mismos. Aunque la pérdida de independencia ocurre a varios niveles, la
disminución de fuerza muscular se considera una de las causas mas importantes que
produce fragilidad y discapacidad en los ancianos. La disminución de fuerza muscular
es responsable de caídas y fracturas que pueden agravar otros cuadros clínicos y
contribuyen enormemente a la discapacidad en los ancianos (Greenlund y Nair, 2003).
Así pues, a medida que la población envejece, se vuelve más importante comprender
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Introducción
(OS mecanismos responsables de la disminución de fuerza muscular para contribuir al
desarrollo de tratamientos e intervenciones que contrarresten la pérdida de función y
masa muscular con el envejecimiento, y mejoren así la calidad de vida de los ancianos.
1.5. Mecanismos responsables de la pérdida de la función muscular con el
enveiecimiento
A pesar de la importancia de la fuerza muscular en la prevención de
discapacidades, los mecanismos responsables de este fenómeno sólo se conocen
parcialmente. No obstante, en los últimos años se han realizado estudios a nivel
morfológico, celular y molecular de los mecanismos responsables de la pérdida de
función muscular, tanto en humanos como en modelos animales de envejecimiento.
Los factores que determinan alteraciones del músculo esquelético se pueden englobar
en cuatro grupos: 1) primariamente neurogénicos, 2) primariamente miogénicos, 3) la
combinación de alteraciones musculares y neurales y 4) mecanismos generales
relacionados con el músculo esquelético (Tabla 2) (Loeser y Delbono, 2003).
1.5.1. Mecanismos primariamente neurogénicos
Los mecanismos primariamente neurogénicos incluyen la reducción en el
número y/o tamaño de las motoneuronas espinales, y alteraciones en el flujo axonal y
en la unión neuromuscular. Cada uno de estos factores, individualmente o en
combinación, produce denervación muscular crónica y remodelado de las unidades
motoras. No sorprende, por tanto, que algunos ancianos presenten evidencias
electromiográficas de denervación muscular, sugiriendo alteraciones de las
motoneuronas que no se vinculan a la clásica enfermedad amiotrófica (esclerosis
10
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Introducción
lateral amiotrófica). La denervación muscular se asocia con la reinervación de fibras,
que se refleja en la agrupación de fibras musculares en secciones histológicas. Los
ciclos de denervación muscular asociados con la reinervación son los responsables de
la remodelación de las unidades motoras. En la actualidad no está claramente
establecida la importancia funcional de este remodelado de las unidades que se
produce con el envejecimiento. El remodelaje de motoneuronas que ocurre con el
envejecimiento involucra fundamentalmente la denervación de fibras rápidas y la
reinervación de estas fibras por colaterales axónicas de motoneuronas que inervan
fibras lentas. De esta forma, el remodelado de las unidades motoras se acompaña de
cambios en la distribución del tipo de fibras en músculos mixtos. A pesar de que la
reinervación de fibras musculares tiende a compensar la denervación, con el
envejecimiento, se produce una pérdida neta de fibras musculares indicando que el
grado de denervación de fibras musculares excede al de reinervación. No está claro si
la denervación muscular es debida a alteraciones en las motoneuronas, en los
terminales nerviosos, o en el transporte axonal. La velocidad de conducción de los
nervios periféricos no se afecta significativamente con el envejecimiento (Loeser y
Delbono, 2003), sugiriendo que con el envejecimiento no existen probablemente
alteraciones en la mielina o reducciones importantes en el calibre de los axones.
Aunque se ha sugerido que la denervación es un factor que contribuye a las
alteraciones del músculo esquelético observadas con el envejecimiento, el nivel de
denervación en músculos individuales y su efecto en músculos de humanos esta por
determinar.
1.5.2. Mecanismos primariamente miogénicos
Estudios previos han contemplado la hipótesis de que la pérdida de masa
11
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Introducción
Tabla 2
Patogenia de las Alteraciones del Músculo Esquelético con el Envejecimiento
I. Alteraciones primariamente neurogénicas
A. Motoneuronas espinales
1. Reducción en el número
2. Reducción de tamaño
B. Alteraciones del flujo axonal
C. Alteraciones de la transmisión neuromuscular
1. Disminución del número de terminales nerviosos
2. Reducción de la liberación de neurotransmisor
3. Disminución del número de receptores de Acetilcolina
II. Alteraciones primariamente miógénicas
A. Daño causado por contracción
B. Disminución de la estabilidad de los puentes de actina-miosina
C. Alteraciones de la señal de transducción (factores tróficos/resistencia a
hormonas)
D. Disminución de la proliferación de las células satélite
III. Combinación de factores
A. Músculos en suspensión
B. Desacoplamiento de excitación-contracción
iV. Mecanismos generales relacionados con el músculo esquelético
A. Estrés oxidative
B. Mutaciones del DNA mitocondrial
C. Vasculopatías relacionadas con el envejecimiento
Adaptada de Loeser y Delbono, 2003.
12
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Introducción
muscular puede ser responsable de la disminución de fuerza nriuscular. Sin embargo, la
disminución de fuerza con el envejecimiento es mayor que la disminución de masa
muscular en humanos (Brooks y Faulkner, 1988). Estudios in vitro sobre contractilidad
realizados en músculos de ratones y ratas han revelado una disminución de fuerza
específica (fuerza normalizada por el área de la sección transversal del músculo) con la
edad (Brooks y Faulkner, 1994a). Esto sugiere que la capacidad intrínseca del músculo
esquelético de generar fuerza esta alterada en mamíferos de edad avanzada, incluidos
los humanos (Delbono, 2002). Varios mecanismos primariamente miogénicos podrían
ser responsables de la disminución de fuerza específica con el envejecimiento, estos
incluyen: /) Alteraciones en la capacidad de recuperación después de daños inducidos
por contracciones (Brooks y Faulkner, 1990), //) disminución de la estabilidad de los
puentes de actina-miosina (Lowe y cois., 2002), iii) alteraciones en la transducción de la
señal en el músculo (acoplamiento de excitación-contracción) (Delbono, 2003) y iv)
disminución de la proliferación de las células satélite (Rennie y cois., 2004).
Las alteraciones producidas por la contracción han sido relacionadas con un
aumento de la fragilidad mecánica y una disminución de la capacidad restaurativa con
la edad, fenómeno que no ha sido bien caracterizado. Cuando los músculos
experimentan repetidos ciclos de contracción/relajación sufren un daño que se acentúa
de forma significativa en animales viejos. Además, los músculos de animales viejos se
recuperan más lentamente que los de animales jóvenes o incluso no llegan a
recuperarse de forma completa (Brooks y Faulkner, 1990).
Alteraciones que impiden la activación completa de la maquinaria contráctil para
desarrollar fuerza pueden contribuir a la disminución de fuerza específica. Existen
numerosos datos experimentales que indican que con el envejecimiento se produce
una disminución de fuerza específica en los distintos tipos de fibras musculares (I, lia,
y llb/llx), tanto de músculos humanos como de rata (Frontera y cois., 2000; Larsson y
13
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Introducción
cols., 1997; Li y Larsson, 1996; Lowe y cols., 2002; Thompson y Brown, 1999). La
observación de que fibras musculares carentes de membrana externa de músculos de
humanos ancianos y otros mamíferos pequeños desarrollan menos fuerza específica
que la de músculos de individuos jóvenes cuando son activadas a nivel máximo sugiere
que la capacidad intrínseca de generar fuerza del aparato miofibrilar está alterada
(Lowe y cois., 2002). Los experimentos realizados en células sin sarcolema no reflejan
las alteraciones en el proceso de acoplamiento excitación-contracción, ya que las fibras
carecen de membrana plasmática y, por ello, cualquier cambio en la fuerza contráctil
generada en fibras de animales viejos refleja directamente alteraciones del aparato
contráctil.
El proceso de acoplamiento de EC se ha demostrado que está alterado con el
envejecimiento. En 1995, Delbono y cois propusieron, por primera vez, que el
mecanismo de acoplamiento de EC esta alterado en músculo esquelético de mamíferos
viejos. Desde entonces se han descrito nuevas evidencias experimentales que apoyan
esta hipótesis (Ryan y cois., 2000; Delbono, 2003) (ver abajo).
Las fibras musculares de mamíferos adultos son células completamente
diferenciadas y; por ello, carecen de actividad mitótica para producir mionúcleos
adicionales cuando aumenta la síntesis de proteínas y el músculo se hipertrofia. El
incremento en el número de mionúcleos se obtiene por diferenciación de células
satélite, que están localizadas en las invaginaciones del sarcolema debajo de la lámina
basal. Dado que para alcanzar una hipertrofia muscular se requiere la activación de
estas células es posible que en la atrofia o en las alteraciones del proceso de
regeneración que sufren los músculos de sujetos de avanzada edad si que pueda estar
disminuida la capacidad proliferativa y el número de estas células satélite. (Decary y
cois., 1997; Renault y cois., 2002; Rennie y cois., 2004).
14
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

introducción
Otros mecanismos generales relacionados con el músculo esquelético son: daño
oxidative (Weindruch, 1995), mutaciones en el DNA de la mitocondria (Hoopes, 2002) y
vasculopatías relacionadas con el envejecimiento (Rogers y Evans, 1993).
1.6. El desacoplamíento de EC como mecanismo responsable de la
debilidad muscular
Aunque se ha propuesto que son varios los mecanismos que causan la
disminución de fuerza específica (ver arriba Mecanismos primariamente miogénicos),
este trabajo esta enfocado en los mecanismos que alteran el acoplamiento EC.
La hipótesis del desacoplamiento EC sugiere que con el envejecimiento se
produce una reducción en el número y/o función de los DHPR y RyRI, lo que
DISMINUCIÓN DE LA EXPRESIÓN DE DHPR Y RyRI EN MÚSCULO ESQUELÉTICO
^ '
DISME^ÍUCIÓN DEL NÚMERO DE DHPR EN EL
SARCOLEMAYRyRl OR RyRI
y r DESACOPLAMÍENTO ENTRE DHPR-RyRl
^ r REDUCCIÓN DE LA
LIBERACIÓN DE Ca^^ DEL RETÍCULO
SARCOPLÁSMICO
^ r
DISMESrUCIÓN DE LA FUERZA ESPECÍFICA
Fulgura 4. Esquema de la hipótesis del desacoplamiento de excitación-contracción con el envejecimiento (Adaptada de Delbono, 2002).
15
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Introducción
disminuye la liberación de Ca^* del RS (retículo sarcoplasmático) y, como
consecuencia, la fuerza específica (Delbono y cois., 1995) (Fig.4).
Para estudiar los cambios que se producen con el envejecimiento en el
acoplamiento EC se ha determinado en fibras musculares de animales jóvenes y viejos
los siguientes parámetros: el movimiento de carga, la corriente de Ca " del DHPR y el
transitorio de Ca^* mioplasmático. Las principales conclusiones aportadas por estos
estudios son: i) El número de DHPR y DHPR/RyR1 en el músculo extensor común de
los dedos (EDL) disminuye con el envejecimiento (Renganathan y cois., 1997a; Ryan y
cois., 2000); ii) el movimiento de carga y la corriente de Ca "" del DHPR disminuyen con
la edad en fibras del músculo cuadríceps de humanos y en el flexor corto de los dedos
(FDB) (Delbono y cois., 1995; Wang y cois., 2000); iii) El incremento transitorio de la
[Ca ""]; disminuye con la edad (Delbono y cois., 1995; Wang y cois., 2000). El
porcentaje de reducción de la liberación de Ca^* intracelular (transitorio de Ca^*) es
similar al de la disminución del número de DHPR y el movimiento de carga.
1.7. La técnica de la fibra única e intacta como excelente aproximación
experimental para determinar la fuerza específica del músculo esquelético
La preparación de fibra única e intacta ha mostrado ser muy útil para determinar
la fuerza específica en músculo esquelético (Lannergren y Westerbiad, 1987). La fibra
única tiene todos los elementos necesarios para reproducir la contracción muscular,
manteniendo el sistema de túbulos T, sarcolema y retículo sarcoplasmático intactos. De
esta forma se pueden realizar estudios funcionales que relacionen cambios controlados
en el voltaje con valores de fuerza específica y cambios en la [Ca^^ji, mediados por la
liberación de Ca "" del RS. En fibras musculares de ratones adultos jóvenes, pero no de
16
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Introducción
ratones viejos, se han obtenido registros simultáneos de la fuerza específica y de la
[Ca^*], (Lannergren y Westerbiad, 1987).
El RS desempeña un papel importante en la regulación del Ca "" mioplasmático.
La magnitud del incremento de la [Ca^""]! libre durante la contracción es el mayor
determinante del desarrollo de fuerza y depende de varios factores, siendo uno de ellos
el contenido de Ca^* del RS. Una menor cantidad de Ca " en el RS podría ser la causa
de la disminución de fuerza específica con la edad. Aunque experimentos de fijación de
voltaje en células humanas han demostrado que existe una cantidad extra de Ca ""
disponible en el RS en fibras de ratones viejos, se desconoce todavía si este Ca^* está
también disponible en fibras únicas musculares durante la contracción evocada por un
estímulo eléctrico. Además, si esta cantidad de Ca "" extra pudiera ser liberada, no se
sabe si el resultado sería un aumento del desarrollo de fuerza en fibras de ratones
viejos. La cafeína es un activador muy potente del RyRI y se ha utilizado ampliamente
para aumentar la liberación de Ca " del RS. Experimentos con cafeína en células
únicas de músculo de ratones jóvenes y viejos podrían proveer información muy útil en
lo que al contenido de Ca^* del RS se refiere.
1.8. Eventos elementales de liberación de Ca^^fEELC) - (Chispas de Ca^ )
delRS
Los eventos elementales de liberación de Ca^* del RS, también denominados
chispas de Ca " , son fenómenos discretos y localizados que surgen de la apertura de
un grupo pequeño de canales-RyR1s de liberación de Ca " . El concepto de que es
necesaria una elevación espacial homogénea del Ca^* citoplasmático para el
reconocimiento de una señal de Ca^* ha sido totalmente abandonado con el
descubrimiento de las chispas de Ca^*. Las chispas de Ca^* presumiblemente ocurren
17
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Introducción
en tal alta frecuencia y número durante la despolarización de la fibra muscular, que los
eventos individuales son difíciles de distinguir durante el transitorio de Ca^''. El
desarrollo de condiciones experimentales bajo las cuales las chispas de Ca "" pueden
ser observadas y caracterizadas ha constituido un enorme avance en el estudio del
proceso de liberación de Ca "" del RS (Schneider y Ward, 2002). Las chispas de Ca ""
son capaces de elevar localmente la [Ca " ]; a valores muy altos (10-100 pM), mientras
que el aumento global de la [Ca ""]! es de menos de 2 nM (Jaggar y cois., 2000). Las
chispas de Ca "" se detectaron por primera vez con imágenes de microscopia confocal
en células musculares cardíacas (Cheng y cois., 1993) y más tarde en una variedad de
tejidos incluyendo el músculo esquelético de anfibios y mamíferos (Fig. 5) (Klein y cois.,
1996) (Shirokova y cois., 1998; Tsugorka y cois., 1995).
Figura 5. Chispas de Ca ^ registradas en una fibra del músculo EDL de un ratón joven. Las imágenes corresponden a una serie de 50 imágenes tomadas a una velocidad de 3 s/imagen con una resolución espacial de 512x512 píxeles y tamaño de pixel de 0.134 pm.
Las chispas de Ca "" han sido estudiadas para entender el proceso celular de la
liberación de Ca "" y su control, ya que aportan información sobre la apertura de los
RyRIs (dispersión espacial, amplitud y duración) en células vivas. El análisis de las
propiedades de las chispas ayudara a conocer mejor el proceso de acoplamiento EC y
a comprender las alteraciones que experimenta este proceso en estados
patofisiológicos como la hipertrofia cardíaca., paro cardíaco e hipertermia maligna
(Cheng y cois., 1999). Por ejemplo, se ha demostrado que las chispas de Ca ""
espontáneas están significativamente alteradas en la disfunción del músculo
18
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Introducción
esquelético asociada a la insuficiencia cardíaca congestiva (Ward y cois., 2003). El
estudio de las chispas de Ca^* también esta siendo utilizado para determinar algunos
aspectos de la regulación metabólica de la liberación de Ca "" por el RyR1 (Isaeva y
Shirokova, 2003a). Esta aproximación experimental es de gran relevancia ya que
existen evidencias que indican que la modulación y función de la actividad del RyR1
esta alterada con la edad (Margreth y cois., 1999). Hasta ahora no se ha realizado
ningún estudio sobre chispas de Ca^* en músculo esquelético de mamíferos viejos.
1.9. El músculo esquelético y el factor de crecimiento similar a la insulina
detipoKIGF-D
IGF-1 es un peptide estructuralmente similar a la proinsulina que tiene como
función principal el producir el crecimiento y diferenciación del músculo esquelético
(Florini y cois., 1996). Con la edad, la producción y actividad del eje de la hormona del
crecimiento/ IGF-1 disminuye lo cual se ha asociado con el aumento de los procesos
catabólicos (Lamberts y cois., 1997) que llevan a la pérdida de masa muscular y fuerza.
La expresión de IGF-1 en fibras musculares, mediante la infección de un adenovirus,
previene la pérdida de masa y fuerza muscular con el envejecimiento mediante la
estimulación de la regeneración del músculo a través de la activación de las células
satélite (Barton-Davis y cois., 1998). Además, la expresión de la isoforma muscular de
IGF-1 produjo hipertrofia sostenida y regeneración en músculo esquelético de animales
viejos (Musaro y cois., 2001). La expresión de IGF-1 dirigida específicamente al
músculo esquelético puede prevenir algunos aspectos de las alteraciones en el
mecanismo de acoplamiento de EC, como por ejemplo: i) IGF-1 previene la disminución
del número de DHPRs con el envejecimiento (Renganathan y cois., 1998)
(Renganathan y cois., 1997b); ii) protege de la disminución del movimiento de carga del
19
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Introducción
DHPR y de la liberación de Ca "" intracelular en fibras del músculo FDB de ratones
viejos (Wang y cois., 2002); iii) puede estimular la transcripción de la subunidad a1S
del DHPRR en células musculares mediante la acción sobre el factor de transcripción
CREB (Zheng y cois., 2002a); este proceso ocurre mediante un mecanismo que
involucra a las proteínas calcioneurina y calmodulina cinasa (Zheng y cois., 2004); y iv)
protege de alteraciones en el área pre y post sináptica de la unión neuromuscular
(Messi y Delbono, 2003). Sin embargo, no existe información sobre el efecto de IGF-1
en la eficiencia del proceso de contracción muscular o la capacidad generadora de
fuerza específica. El impacto de los cambios arriba mencionados sobre la contractilidad
de fibras únicas necesita ser investigado mediante la medida de la fuerza específica y
el Ca " intracelular en fibras únicas de músculos de animales IGF-1- transgénicos.
20
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

2. Objetivos
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Objetivos
2. Objetivos
A pesar de la importancia de la conservación de la fuerza muscular en la
prevención de la discapacidad física, los mecanismos biológicos responsables del
deterioro funcional y estructural del músculo esquelético asociados al envejecimiento se
conocen sólo parcialmente. Se desconoce si la debilidad muscular es consecuencia de
alteraciones en las propiedades intrínsecas de las fibras musculares, y/o de un mayor
nivel de fatiga de las mismas, que conduce a un menor desarrollo de fuerza durante el
proceso de envejecimiento. Uno de los eventos cruciales del proceso que lleva a la
contracción muscular (el acoplamiento de excitación-contracción-EC) es la liberación de
Ca^* del retículo sarcoplasmático (RS). No se sabe si una alteración en el proceso de
liberación de Ca^* en células de ratones viejos es responsable de la disminución de
fuerza específica. Además, la debilidad muscular esta asociada a la disminución de los
niveles circulantes del factor de crecimiento similar a la insulina (IGF-1), que ocurre
durante el envejecimiento. Aunque se ha demostrado que IGF-1 puede restaurar
algunos aspectos del acoplamiento de EC durante la edad avanzada no se sabe si este
factor puede restaurar la fuerza muscular específica. Por todo ello, en este trabajo se
estudiara si la debilidad muscular que ocurre con el envejecimiento es una
consecuencia de alteraciones en las propiedades intrínsecas de las fibras musculares,
y/o de un mayor nivel de fatiga de las mismas. Además, estudiaremos, utilizando la
técnica de contracción de fibra única e intacta, si la disminución de la fuerza específica
esta asociada a la disminución de la liberación de Ca^* del RS, y si esta disminución se
puede prevenir mediante la expresión transgénica de IGF-1 en el músculo esquelético.
Para ello:
22
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Objetivos
1) Se medirán las propiedades contráctiles de fibras únicas del músculo
esquelético lento y rápido de ratones jóvenes, de edad mediana y viejos, utilizando la
técnica de contracción de fibra única intacta, para determinar si la disminución de
fuerza específica con el envejecimiento es el resultado de alteraciones en las fibras que
componen el músculo.
2) Se determinará el nivel de fatiga de fibras únicas del músculo esquelético de
ratones jóvenes y viejos para saber si la resistencia muscular contribuye a una
limitación de la fuerza desarrollada por el músculo esquelético de mamíferos de edad
avanzada.
3) De existir cambios en los niveles de fuerza específica con la edad se
estudiará si la disminución de la fuerza específica que ocurre con el envejecimiento
esta asociada a una disminución de la liberación de Ca "" del RS. Para ello, mediremos,
simultáneamente, los niveles de Ca "" intracelular y la fuerza especifica de fibras
musculares esqueléticas de ratones jóvenes y viejos.
4) De existir una disminución en la liberación de Ca "" del RS se estudiará la
fuerza específica y la liberación de Ca^* en fibras musculares de ratones transgénicos-
lGF-1 para determinar si la expresión transgénica de IGF-1 en músculo esquelético
puede prevenir la disminución de la liberación de Ca "" del RS y/o la fuerza específica.
5) Por último, y en el caso de que exista una disminución en la liberación de
Ca "" del RS se caracterizarán los eventos elementales de la liberación de Ca "" (EELC-
0 chispas de Ca "") en fibras musculares de ratones jóvenes y viejos. El estudio de las
chispas de Ca^* nos permitirá averiguar si existen alteraciones en los RyR1s que
contribuyen al proceso de desacoplamiento de EC.
23
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

3. Material y Métodos
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Material y Métodos
3. MATERIAL Y MÉTODOS
3.1. Disección v montaje de fibras únicas e intactas
Mediante disección se aislaron fibras únicas e intactas de los músculos EDL y
soleo de ratones de 2 a 6 (joven; n=17) de 12 a16 (edad mediana; n= 7) y de 20-24
meses de edad (viejo; n=15) de cepa DBA (colonia del Instituto Nacional de Harían-
Instituto Nacional del envejecimiento-NIA) o FVB (nuestra colonia). También se aislaron
fibras del músculo FDB de ratones de 2-4 meses de edad de cepa DBA o FVB. Ambas
cepas se han utilizado en estudios previos como modelo de animal de envejecimiento
(Bakker y cois., 1997a; Renganathan y cois., 1998). Los procedimientos con animales
siguieron un protocolo previamente aprobado por el comité del cuidado y uso de
animales de la Universidad de Wake Forest (Winston-Salem, NC, USA). Los animales
se sacrificaron mediante dislocación cervical y los músculos se disecaron de ambas
patas posteriores. Las fibras únicas fueron disecadas bajo estereoscopio (30-50X),
poniendo especial atención en dejar la menor cantidad posible de restos celulares de
células vecinas. Para permitir un montaje estable y seguro de la fibra, la conexión de la
fibra con el tejido conectivo y los tendones se mantuvo intacta. Los tendones fueron
fijados con unos micro-clips hechos de papel de aluminio y la preparación transferida a
la cámara de registro. Las fibras musculares fueron montadas entre un transductor de
fuerza (403 A Aurora Scientific) y un microposicionador que permitió la regulación de la
longitud de la fibra. Las fibras fueron perfundidas continuamente con la solución de
registro (concentraciones expresadas en mM: NaCI 121, KCI 5, CaCb 1.8, MgCb 0.5,
NaH2P04 0.4, NaHCOs 24, glucosa 5.5) aireada continuamente con una mezcla de 5%
CO2 y 95% O2 para conseguir un pH de 7.4. Para el registro se ajustó la longitud de las
fibras musculares de tal forma que pudieran desarrollar la fuerza máxima en respuesta
25
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Material y Métodos
a la estimulación eléctrica.
3.2. Registros de contracción de fibra única
Las fibras se estimularon directamente mediante un campo eléctrico generado
entre dos electrodos de platino conectados a un estimulador con unidad de aislamiento
eléctrico. Trenes de pulsos rectangulares de frecuencia variable, entre 5 y 100 Hz, de
una duración de 350 ms se aplicaron mediante estimulación eléctrica con un intervalo
de 5 minutos para determinar la fuerza máxima específica. Trenes de duración más
larga fueron necesarios para conseguir la fuerza máxima específica en células del
músculo soleo. Para la adquisición de datos se utilizó un ordenador personal, un
convertidor analógico digital (Digidata 1200, Axon Instruments, Foster City, CA) y el
programa de adquisición y análisis PCLAMP (Axon Instruments). El diámetro de la
célula fue medido en la cámara de registro a una magnificación de 200-400X por dos
investigadores y la sección transversal calculada como n(cl/2)^, donde d es el diámetro
de la célula.
Un experimento típico se desarrolló de la siguiente manera: La fibra muscular
intacta fue montada en la cámara de registro y estirada hasta alcanzar la longitud
óptima (Lo) (longitud que permite el desarrollo de la fuerza máxima) mediante
estimulación breve. Después de obtener Lo, las fibras se dejaron descansar entre 5-7
min. Seguidamente se estudió la relación fuerza-frecuencia mediante la aplicación de
trenes de pulsos eléctricos de 350 ms de duración a diferentes frecuencias, que
variaron, de forma creciente, entre 5 y 200 Hz. Cuando dos frecuencias consecutivas
desarrollaron la misma fuerza, la frecuencia siguiente no fue aplicada, con el fin de
reducir la duración del experimento.
26
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Material y Métodos
3.3. Registros de contracción de los experimentos de fatiga
Se utilizaron dos protocolos para el estudio de fatiga, llamados de intervalo corto
(IC) y de intervalo largo (IL). Ambos protocolos fueron estudiados a la frecuencia que
evocó la fuerza máxima (ver capítulo I). Los trenes de pulsos fueron de 350 ms de
duración y el intervalo entre trenes fue de 1 ó 3.65 s para los protocolos IC o IL,
respectivamente; estos protocolos se aplicaron durante 5 y 10 min. para IC y IL,
respectivamente. Las células de los músculos EDL fueron de contracción rápida,
mientras que las del soleo de contracción lenta. Para determinar si una célula es de
contracción rápida o lenta se midieron varios parámetros: tiempo al pico de contracción,
tiempo de relajación media y duración de la estimulación necesaria para desarrollar la
fuerza máxima.
3.4. Experimentos de contracción de músculo entero
Para los experimentos de contracción de músculo entero la parte distal del
músculo FDB fue disecado de ambas patas traseras del ratón. El músculo fue montado
entre un transductor de fuerza 404A (Aurora Scientific) (complianza: 0.1 pm mN-1,
frecuencia de resonancia: 2 KHz) y un microposicionador ajustable para permitir el
control de la posición y la longitud del músculo. El músculo se perfundió continuamente
por medio de una bomba peristáltica, de forma similar a como se ha descrito
previamente en la sección de experimentos de fibra única.
Los registros de contracción y el análisis de experimentos de músculo entero
siguieron la misma técnica descrita para los experimentos de fibra única con algunas
modificaciones. Para producir la contracción máxima a diferentes frecuencias se
aplicaron normalmente trenes de pulsos de 500 ms. En ocasiones, se aplicaron trenes
27
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Material y Métodos
de mayor duración, cuando la contracción no alcanzó un nivel estable de meseta
durante el estimulo. El diámetro del músculo se midió en la cámara de registro a través
de un microscopio a una magnificación de 30X o con preparaciones histológicas de
secciones transversales. Estas últimas fueron adquiridas digitalmente y el área de las
secciones se midió usando el programa Isee (Inovision, Durhan, NC, USA). El área de
la sección transversal (AST) de cada músculo se midió trazando una región de interés
(RDI) en la imagen digital a lo largo del borde de varias secciones de cada músculo. El
área se calculó en base a las dimensiones de altura y anchura de cada pixel y al
número total de píxeles en la RDI.
3.5. Registros de Ca ^ intracelular en fibras únicas intactas de FDB
La [Ca^*]i de una fibra muscular se registró simultáneamente con la fuerza
específica, usando el indicador de fluorescencia fluo-3-AM (Molecular Probes, Eugene,
OR, USA). Las fibras se incubaron con 5 f M fluo 3-AM por 30-40 min usando una
concentración preparada de fluo 3-AM de 1 mM en DMSO, lo que permitió una muy
baja concentración de DMSO en la cámara de registro durante la incubación (<0.5%).
Después de la incubación las células se perfundieron con solución de registro durante 5
min antes de empezar el protocolo de contracción. Para los registros de fluorescencia
la fibra se iluminó con un rayo láser de longitud de onda 488 nm. El rayo láser pasó a
través de un módulo de sean OZ (Noran Instruments Confocal System, Middieton, Wl,
USA) y a través de un objetivo Fluar 20 X (Zeiss, Oberkochen, Germany) montado en
un microscopio invertido (Axiovert S100 2TV, Zeiss) antes de iluminar la preparación.
La fluorescencia emitida fue recogida por el objetivo y dirigida al módulo de sean OZ,
en la configuración "sin banda" y a través del filtro de emisión de longitud de onda de
500 nm antes de ser recogida por el fotomultiplicador y digitalizada (Fig. 6). El control
28
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Material y Métodos
de los equipos, la adquisición de las imágenes y el procesado se hicieron a través del
programa Intervision (Noran Instruments) en un ordenador Silicon Graphics 02
(Mountain View, Ca, USA).
Para el análisis de datos, varias RDI fueron seleccionadas de cada célula y los
valores de la emisión máxima de fluorescencia fueron utilizados para las
comparaciones estadísticas.
Objetivo
jjlaiio de foco
Figura 6. Esquema del microscopio utilizado para los registros de fluorescencia
3.6. Calibración del indicador Fluo 3-AM y cálculo de la concentración
jntracelular de Ca ^
Para cuantificar la [Ca^*]¡, las señales fluorescentes se transformaron en
concentraciones de Ca "". Para ello, se calibró en fibras musculares la fluorescencia
29
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Material y Métodos
emitida por fluo S-AIVI a distintas concentraciones de Ca^*. Aunque la constante de
disociación, Kd, es una propiedad del indicador de Ca "*" y por ello tiene un valor
relativamente constante, se conoce que puede ser afectada por diversos factores como
el pH, la viscosidad y la fuerza iónica, entre otros. Debido a esto, medimos la Kd del
indicador fluo 3-AM para Ca^* en células disociadas de músculo FDB. Los músculos
FDB de ratones jóvenes fueron incubados con colagenasa a una concentración de
2mg/mL (Sigma, St. Louis, MO, USA) en una solución de disociación que contenía 155
mM aspartate de cesio, 5 mM aspartate de magnesio y 10 mM HEPES (pH 7.4 con
CsOH) (Beam y Franzini-Armstrong, 1997), durante 3h a 37 C. Las fibras musculares
se disociaron usando pipetas Pasteur de diferentes tamaños de punta. Seguidamente,
las fibras fueron incubadas con fluo 3-AM (5 p-M) durante 30-40 min. Tras la incubación,
se lavó el fluo 3-AM y las células fueron expuestas a soluciones estandarizadas de
diferentes concentraciones de Ca^*. La concentración de Ca "" en las soluciones
estandarizadas se calculó de acuerdo con el procedimiento previamente descrito (Tsien
y Pozzan, 1989). En estas condiciones se midió la fluorescencia basal. Después, las
fibras se permeabilizaron con el detergente saponina a una concentración de 0.01%
para permitir la entrada de Ca^* y se midió la fluorescencia máxima alcanzada antes de
que las células se movieran o su forma se distorsionara. Los valores correspondientes
a cada punto experimental se obtuvieron de la media de al menos 4 células. La
fluorescencia a diferentes niveles de Ca "" se normalizó con la fluorescencia máxima y
los valores de la media se ajustaron a la siguiente ecuación: y= 1/(1+(Kd/[Ca^''])) (Wang
y cois., 1999a). La Kd de fluo 3-AM, medida en estas condiciones, fue 708 nM. La
[Ca^*]¡ se calculó usando la siguiente ecuación: [Ca^*] = Kd (F-Fmin)/(Fmax-F) (Tsien y
Pozzan, 1989), donde Kd es la constante de disociación; F el valor de fluorescencia de
pico registrado en cada célula; Fmax, la fluorescencia máxima, que se midió al final del
experimento en cada célula, perfundiendo solución Ringer con 0.01% de saponina; y
30
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Material y Métodos
Fmin, la fluorescencia en condiciones de reposo, que se midió en un grupo de células
equilibradas con 2 mM BAPTA AM. Este procedimiento dio resultados similares a los
obtenidos en células individuales disecadas manualmente y que no habían sido
incubadas con BAPTA-AM.
3.7. Análisis del tipo de fibra en músculo esquelético
Después de los registros de contracción, los músculos FDB se recogieron de la
cámara de registro, se estiraron a una longitud aproximada a la U, se sumergieron en
medio OCT (Tissue-Tek, Torrance, CA, USA), y se congelaron rápidamente en 2 -
metilbutano (Fisher Scientific, Pittsburgh, PA, USA) enfriado en hielo seco a -40 C.
Los músculos se conservaron a -80 C hasta ser usados posteriormente. Las muestras
congeladas se seccionaron en un criostato (Leica CM3000, Nussioch, Germany) a 21
C. Las secciones (10 ¡am) se guardaron a 4 C hasta ser procesadas para
inmunohistoquímica. Para la identificación de los tipos de fibras, las secciones
musculares fueron expuestas a anticuerpos primarios NCL-MHCs (tipo I), NCL-MHCf
(rápidas totales) (Novocastra, Newcastle upon Tyne, UK), A4.74 (type IIA) (Alexis
Biochemicals, San Diego, CA, USA), o BF-F3 (tipo MB) (ATCC, Rockville, MD, USA) a
una dilución de 1:20 en PBS. Como anticuerpo secundario se usó un conjugado de
conejo anti-ratón fluoresceína isotiocianato (FITC) a una dilución de 1:100. El tipo IIX se
obtuvo restando las fibras de tipo HA y MB al número total de fibras rápidas detectado
con el anticuerpo NCL-MHCf (Novocastra). Como en FDB no se detectaron fibras de
tipo MB, el músculo EDL fue usado como control positivo para el anticuerpo BF-F3. La
fluorescencia de las secciones fue adquirida usando un microscopio invertido (Axiovert
100, Zeiss) y un sistema de imagen con una cámara CCD PXL-EEV-37 (Photometries,
Tucson, AZ, USA). El programa Isee de Inovision se utilizó en una estación de trabajo
31
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Material y Métodos
Silicon Graphics O2 para el procesado de las imágenes.
3.8. Medida de la concentración de IGF-1 en músculo FDB
Para medir la concentración de IGF-1 humano (hlGF-1) en los músculos FDB se
utilizó el kit Active IGF-1 Elisa (DSL-10-5600, Diagnostic System Laboratories, Inc.,
Webster, TX, USA). Los músculos individuales de animales control y transgénicos se
congelaron en nitrógeno líquido y se guardaron a -80 C para su uso posterior. Los
músculos fueron descongelados en solución de ensayo (1 mg de músculo en 10 |JL de
solución de ensayo), homogeneizados, sonicados y centrifugados durante 10 min a
12000g. El sobrenadante y el estándar se añadieron en los pozos y se incubaron con el
complejo anticuerpo-enzima durante 2 h. Después de lavar este complejo, se
añadieron, secuencialmente, la solución de cromógeno tetrametilbenzidina (TMB) y la
solución de parado. La absorbancia de cada pozo se determinó a 450 nm en un lector
de microplaca AB Biosystem. Para la normalización, la concentración de proteína en
los sobrenadantes se midió usando el ensayo de proteína BCA (Pierce, Rockford, IL,
USA). El límite de detección de hlGF-1 fue 0.03 ng (g proteína)"\
3.9. Preparación de las fibras musculares para el registro de chispas de
Caf:
Los músculos EDL de ambas patas traseras fueron disecados de ratones
jóvenes (3-6 meses de edad; n= 7) y viejos (21-24, n= 10) de cepa FVB, DBA y CB6F1.
Los restos de vasos sanguíneos, nervios y el tejido conectivo que rodea al músculo
fueron eliminados y los músculos se trataron con colagenasa 2 mg/ml y 10% suero
bovino fetal durante 10-15 min en una solución que contenía (en mM): ácido aspártico
32
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Material y Métodos
155, aspartato de magnesio 5 mM y Hepes 10, pH 7.4 ajustado con CsOH (Beam y
Franzini-Armstrong, 1997). Los músculos fueros transferidos después a una solución
relajante que contenía (in mM): ácido glutámico 140, Hepes 10, glucosa 5, EGTA 1,
MgCb 10, CaCl2 0.3, pH 7 (ajustado con KOH) y segmentos de células individuales
fueron disecados cuidadosamente y fijados en una posición ligeramente estirada (a
longitud de sarcomere de 2.5-3 )j,m) al fondo de una cámara de Lucite de 200 ML- Las
fibras se permeabilizaron con saponina 0.01% durante 2 min en la solución relajante.
Las células se incubaron después con fluo-4 en una solución interna que contenía (en
mM): glutamato de sodio, 140; Hepes, 10; glucosa, 5; EGTA, 0.5; NaaATP, 5;
fosfocreatina-Na2, 5; MgCb, 5.5, CaCb, 0.9 y fluo-4 0.08 (Molecular Probes, Eugene,
OR, USA), pH: 7 (Kirsch y cois., 2001). Los productos químicos utilizados se diluyeron
en solución interna el día del experimento a partir de las siguientes soluciones
concentradas: rianodina, 10 mM (Calbiochem, La Jolla, CA, USA), Imperatoxina-A, 10
I M (Alomone Labs, Jerusalem, Israel) diluidas en agua destilada. La solución
concentrada de cafeína (100 mM) (Sigma, ST. Louis, MO, USA) se preparó en solución
interna el día del experimento. Las soluciones de Carbonyl cyanide p-
(Trifluoromethoxy) phenylhydrazone (FCCP) (Sigma, ST. Louis, MO, USA) (39 mM) y
ologomicina (Sigma, St. Louis, MO, USA) (10 mM) se prepararon en etanol 95 y 100 %,
respectivamente.
2+ 3.10. Imágenes de fluorescencia de las chispas Ca
La cámara experimental se montó en la base de un microscopio invertido
Axiovert 100 (Zeiss, Oberkochen, Alemania) equipado con un sistema de láser confocal
(Radiance 2100 K1, Bio-Rad Laboratories, Hercules, CA, USA). Las fibras se
observaron a través de un objetivo C-Apochromat de inmersión en agua de 40X y
33
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Material y Métodos
apertura 1.2 (Zeiss) usando un láser de Criptón-Argón de longitud de onda 488 nm. La
fluorescencia emitida se midió a 528 ± 25 nm. Para la mayoría de los experimentos la
intensidad del láser se mantuvo entre el 6-12% con un filtro de densidad neutra. Se
tomaron dos tipos de imágenes: de cuadro completo xy y de escan de línea xt. Las
imágenes xy tuvieron una dimensión 512x512 píxeles, siendo el tamaño del pixel de
0;134 \xm, en ambas direcciones, xe y, donde imágenes consecutivas se tomaron cada
1.2 y 3.2 s. La fibra se orientó de forma paralela al eje x de escaneo. Se registraron
series de 50 imágenes en 3-5 sitios diferentes en cada fibra. Las imágenes de escan de
línea se adquirieron de 512 píxeles (0.134 pm/pixel) en la dirección x y 1000 píxeles (2
ms/línea) en la dirección t. Se escaneó la totalidad del diámetro de la célula a intervalos
de 20 píxeles. Para la adquisición de imágenes se utilizó el programa Radiance
LaserSharp 2000 sofware (Bio-RAD).
3.11. Análisis de las imágenes de chispas de Ca ^
La detección de los eventos espontáneos de la liberación de Ca " (EELC) se
llevó a cabo de dos formas: visualmente por dos investigadores y automáticamente
usando una versión modificada del programa descrito por Cheng y cois, (1999). Este
programa utiliza IDL 5.6 (Interactive Data Language, Research Systems Inc., Boulder,
CO., USA). El Dr. Sandor Gyorke (Dpt. de Fisiología, Texas, Tech University Health
and Sciences Center) cedió generosamente una versión modificada del programa. El
número de eventos por imagen se utilizó para el cálculo de los parámetros estadísticos
de la frecuencia de eventos en las series xyt (Kirsch y cois., 2001). La fluorescencia de
base (Fo) se determinó calculando la fluorescencia media de la imagen sin procesar
excluyendo posibles eventos (áreas de píxeles continuos que exhibieron una
fluorescencia > 1.5 veces la desviación estándar (DE) por encima de la fluorescencia
34
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.
Material y Métodos
media de la fibra). La normalización de regiones de la imagen que contenían EELC se
llevó a cabo calculando el cociente F{x,y) o F{x,t) entre Fo(x) (Kirsch y cois., 2001). Las
regiones de posibles eventos locales de Ca "" se identificaron en las imágenes AF/Fo
como regiones continuas de píxeles con intensidad de fluorescencia > 2 DE por encima
de la fluorescencia media (Chun y cois., 2003). Las imágenes de fluorescencia
normalizadas fueron filtradas con un filtro kernel de 3 píxeles de tamaño. De las
imágenes de fluorescencia normalizadas y filtradas que contenían EELC se
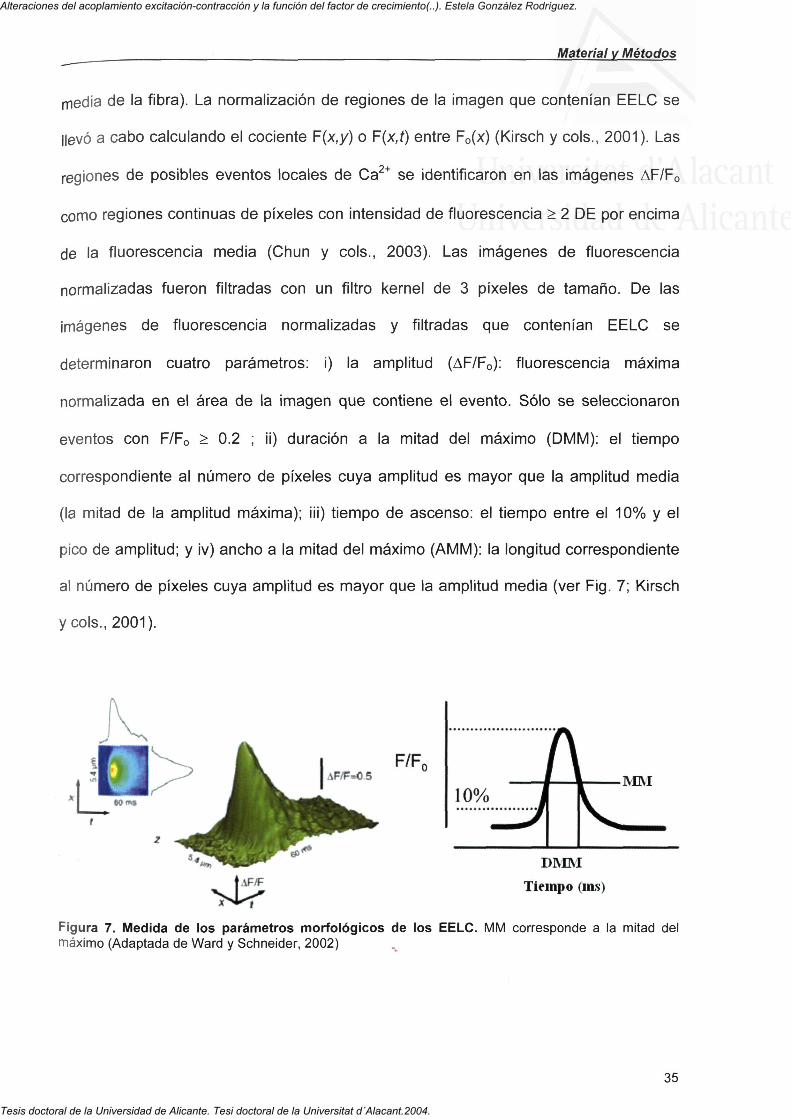
determinaron cuatro parámetros: i) la amplitud (AF/Fo): fluorescencia máxima
normalizada en el área de la imagen que contiene el evento. Sólo se seleccionaron
eventos con F/Fo > 0.2 ; ii) duración a la mitad del máximo (DMM): el tiempo
correspondiente al número de píxeles cuya amplitud es mayor que la amplitud media
(la mitad de la amplitud máxima); iii) tiempo de ascenso: el tiempo entre el 10% y el
pico de amplitud; y iv) ancho a la mitad del máximo (AMM): la longitud correspondiente
al número de píxeles cuya amplitud es mayor que la amplitud media (ver Fig. 7; Kirsch
y cois., 2001).
AF/F=0.5 F/F.
DMM
Tiempo (ins)
Figura 7. Medida de los parámetros morfológicos de los EELC. MM corresponde a la mitad del máximo (Adaptada de Ward y Schneider, 2002)
35
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Material y Métodos
3.12. Composición de cadena pesada de miosina (CPM)
La composición de CPM de las fibras se determinó mediante gel de
electroforesis de poliacrilamida (SDS-PAGE), como se ha descrito previamente (Giulian
y cois., 1983; Messi y Delbono, 2003) con algunas modificaciones. En breve, las fibras
únicas musculares se disolvieron en 40 n\ de una solución que contenía: TRIS, 62.5
mM, 1% de azul de bromofenol, 15% de glicerol, 5% betamercaptoetanol e inhibidor de
proteasas (1 tableta/10 mL Complete Mini: roche Diagnostic, Mannheim, Germany),
después del experimento de registro de chispas de Ca "* y se guardaron a -80 C.
Todas las muestras se utilizaron para la electroforesis entre 1 y 4 semanas después de
los registros de Ca "". El día del experimento, las muestras se sonicaron en hielo
durante 2 min con un desmembranador sónico (Modelo 60, Fisher Scientific).
Posteriormente, se incubaron durante 5 min a 95 C. Debido a la ausencia de
standards comercialmente disponibles en el mercado, un grupo de músculos que
contenía los cuatro tipos de MCP (I, Ha, lib y llx) o el diafragma de rata fueron utilizados
como referencias estándar. La concentración de acrilamida fue de 4 % en el gel de
apilamiento y de 6% en el gel de corrido. La matriz del gel incluyó glicerol al 30%. Para
la detección de la MCP se utilizó el kit Silver stain plus (Bio-Rad).
3.13. Preparación de las membranas e Inmunobiots del RvR
Las fracciones de membrana se prepararon de 12 músculos EDL disecados de 3
ratones jóvenes y 3 viejos. Para cada experimento se agruparon las membranas de dos
músculos y se realizaron 3 Western blots con muestras de ratones jóvenes y viejos. Un
grupo mixto de músculos de las extremidades traseras de rana Northern leopard se
utilizó como referencia positiva para ambos, RyRa y RyR[3, que son los homólogos de
36
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Material y Métodos
RyR1 y RyR3 (Chun y cois., 2003). Los músculos EDL se homogeneizaron
manualmente en 150 |jl de solución de homogeneización enfriada en hielo (20 mM
Hepes, pH 7.4, 250 mM sucrosa, y un mezcla de inhibidor de proteasas, Roche) con
pipetas de diferentes tamaños de punta. Las fracciones de membrana se extrajeron
añadiendo 150 pl de solución de extracción (5 mM NaPOa, 75 mM NaCI, 1% Triton-X
100, 1% deoxicolato, 0.1% SDS), que también contenía el inhibidor de proteasas, y se
centrifugaron a 10000 g durante 15 min. Las proteínas en el sobrenadante se
separaron en geles SDS-PAGE 4% para la separación del RyR (Zheng y cois., 2002a;
Chun y cois., 2003) y se transfirieron a membranas de difluoruro de polivinilideno -
PVDF (Hybond-P, Amersham Biosciences) en la solución de transferencia (12 mM
TRIS-base, 92 mM glicina, 0.02 % SDS y 20% metanol) a 25 V durante la noche a 4°C.
En cada línea se cargó la misma cantidad de proteína (40pg). Analizamos la expresión
de la ATPasa de calcio del retículo endoplásmico de fibras rápidas (SERCA-1), cuya
expresión no cambia con el envejecimiento, como control interno para el cargado del
gel (Ferrington y cois., 1997). Las membranas PVDF se bloquearon durante 3 h en una
solución de bloqueo que contenía: 5% de leche desnatada (Carnation) en 150 mM
NaCI, 10 mM Tris.HCI, pH 7.4, 0.1% Tween 20 (Tris buffer sodium Tween, TBST) a
temperatura ambiente. Las membranas se Incubaron después con el anticuerpo
monoclonal de RyR (Developmental Studies Hybridoma Bank, University of Iowa)
diluido a 1:5000 en solución de bloqueo a temperatura ambiente durante 1h, y se
lavaron posteriormente 3 veces en TBST sin Tween-20. Después se incubaron durante
1h con el anticuerpo secundario anti-ratón conjugado con peroxidasa, diluido a 1:10000
en leche en polvo 5% TBST (Amersham Biosciences). Posteriormente, las membranas
se lavaron tres veces durante 5 min cada vez en TBS, 0.05% Tween 20 y una vez en
TBS. Finalmente, las membranas se incubaron en el reactivo de quimioluminiscencia
(ECL) (Pierce, Rockford, IL) y se visualizaron en película de rayos X.
37
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Material y Métodos
3.14. Análisis estadístico
Todos los valores han sido expresados como la media ± el error estándar de la
media (ESM) o la desviación estándar (DS) con el número de observaciones (n).
El análisis estadístico para los experimentos de fibra única de EDL , soleo y FDB
se ha desarrollado con el test del análisis de la varianza (ANOVA) seguido de los tests
de BONFERRONI o DUNN o Student-Newman-Keuls dependiendo de la distribución de
los datos. Se aceptó como límite de significación un valor de p<0.05.
Para comparar los resultados de los experimentos de fatiga se utilizó el análisis
de la varianza (ANOVA) seguido de los test de Bonferroni o DUNN, y las pruebas CHI-
cuadrado y Fisher, de acuerdo con la distribución de los datos.
Los experimentos de las chispas de Ca " se compararon utilizando los test
paramétricos Student t-test, paired t-test y el test de Mann Whitney Rank sum.
38
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

4. Resultados
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

CAPITULO I:
EFECTO DEL ENVEJECIMIENTO SOBRE LA FUERZA ESPECIFICA DE CÉLULAS
ÚNICAS E INTACTAS DE LOS MÚSCULOS EXTENSOR COMÚN DE LOS DEDOS
(EDL) Y SOLEO DE RA TON
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
4.1.1. La fuerza específica máxima en células de músculos rápidos v lentos
Se determinó la fuerza específica de fibras aisladas de los músculos EDL y
soleo en respuesta a una estimulación única (twitch) o prolongada (tétano), utilizando
los métodos previamente descritos. Debido a que éste es el primer estudio sobre las
propiedades contráctiles de fibras únicas de los músculos EDL y soleo, nos propusimos
determinar la reproducibilidad de la fuerza máxima con el tiempo. Para ello, calculamos
el cociente entre la fuerza máxima o tetánica al final del experimento y la fuerza
máxima al principio del experimento. Los resultados demuestran que la disminución de
fuerza no fue significativa durante un periodo de tiempo de 2-3 h. Los coeficientes
calculados para las fibras del músculo EDL de ratones jóvenes y viejos son,
respectivamente, 0.9 ± 0.2 y 0.99 ± 0.08 (P= 0.66); para las fibras del músculo soleo
0.95 ± 0.09 y 0.91 ± 0.05 (P= 0.74) y para las fibras del músculo FDB de ratones
jóvenes 0.92 ± 0.08. No se encontraron diferencias significativas entre edades o células
de diferentes músculos. Estos resultados demuestran que la fuerza tetánica en células
únicas intactas de músculo esquelético de ratón es estable durante un periodo de 2 ó 3
h y que la disminución de la fuerza durante el transcurso del experimento fue menor del
10% para todos los grupos. El análisis del diámetro y la sección muscular transversal
demostró que las fibras del soleo de ratones viejos son significativamente más
pequeñas que las fibras de músculo de ratón joven o de mediana edad y que las fibras
del músculo EDL de ratón viejo son más pequeñas que las fibras de ratones de
mediana edad (ver TABLA 3). La fuerza específica máxima se midió en fibras de FDB,
EDL y soleo de animales jóvenes. Los valores de fuerza específica de fibras de
distintos músculos de ratones de la misma edad no fueron significativamente
diferentes.
41
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.
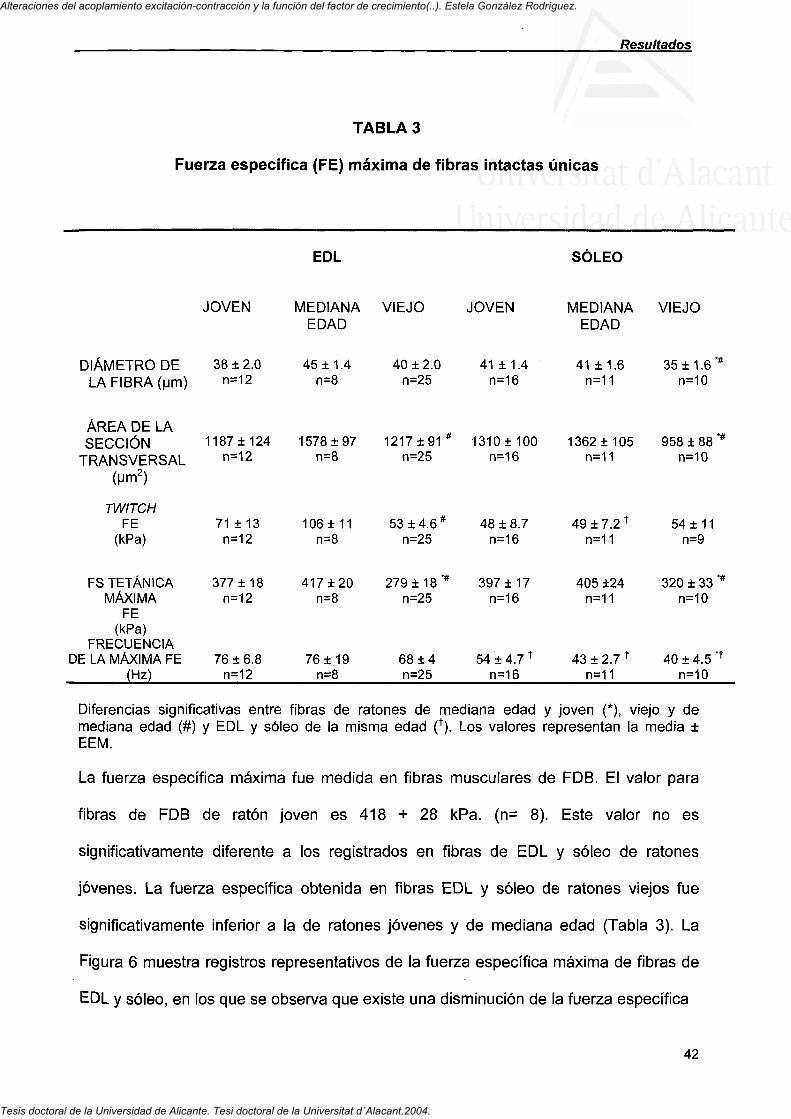
Resultados
TABLA 3
Fuerza especifica (FE) máxima de fibras intactas únicas
EDL SOLEO
DIÁMETRO DE LA FIBRA (|jm)
ÁREA DE LA SECCIÓN
TRANSVERSAL (Ijm^)
TWITCH FE
(kPa)
FS TETÁNICA MÁXIMA
FE (kPa)
FRECUENCIA DE LA MÁXIMA FE
(Hz)
JOVEN
38 ±2.0 n=12
1187 + 124 n=12
71 ±13 n=12
377 ±18 n=12
76 ± 6.8 n=12
MEDIANA EDAD
45 ±1.4 n=8
1578 ±97 n=8
106±11 n=8
417±20 n=8
76 ±19 n=8
VIEJO
40 ±2.0 n=25
1217±91 * n=25
53 ±4 .6* n=25
279 ±18** n=25
68 ±4 n=25
JOVEN
41 ±1.4 n=16
1310±100 n=16
48 ± 8.7 n=16
397 ±17 n=16
54 ± 4.7 ^ n=16
MEDIANA EDAD
41 ±1.6 n=11
1362 ±105 n=11
49 ± 7.2 ^ n=11
405 ±24 n=11
43±2.7'f n=11
VIEJO
35 ±1.6** n=10
958 ± 88 ** n=10
54 ±11 n=9
320 ± 33 ** n=10
40 ± 4.5 '^ n=10
Diferencias significativas entre fibras de ratones de mediana edad y joven (*), viejo y de mediana edad (#) y EDL y soleo de la misma edad C*"). Los valores representan la media ± EEM.
La fuerza específica máxima fue medida en fibras musculares de FDB. El valor para
fibras de FDB de ratón joven es 418 + 28 kPa. (n= 8). Este valor no es
significativamente diferente a los registrados en fibras de EDL y soleo de ratones
jóvenes. La fuerza específica obtenida en fibras EDL y soleo de ratones viejos fue
significativamente inferior a la de ratones jóvenes y de mediana edad (Tabla 3). La
Figura 6 muestra registros representativos de la fuerza específica máxima de fibras de
EDL y soleo, en los que se observa que existe una disminución de la fuerza específica
42
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.
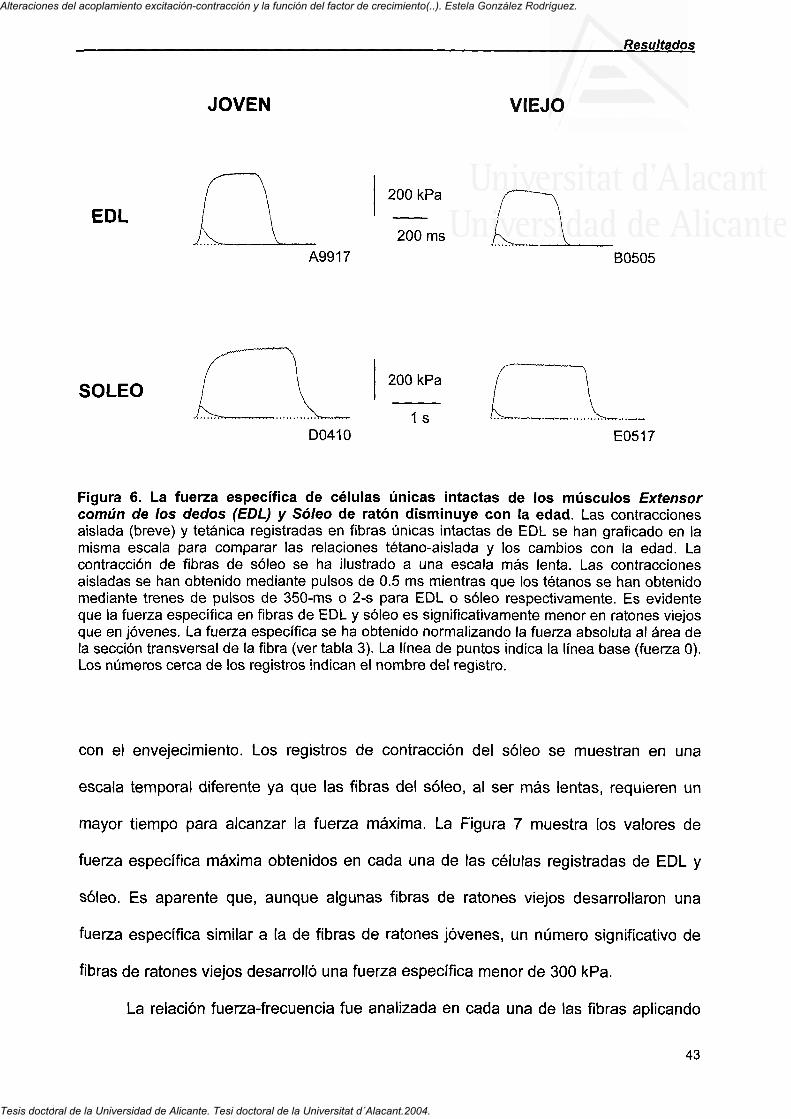
Resultados
JOVEN VIEJO
EDL 200 kPa
200 ms A9917 B0505
SOLEO 200 kPa
1 s D0410 E0517
Figura 6. La fuerza específica de células únicas intactas de los músculos Extensor común de los dedos (EDL) y Soleo de ratón disminuye con la edad. Las contracciones aislada (breve) y tetánica registradas en fibras únicas intactas de EDL se tian graficado en la misma escala para comparar las relaciones tétano-aislada y los cambios con la edad. La contracción de fibras de soleo se ha ilustrado a una escala más lenta. Las contracciones aisladas se lian obtenido mediante pulsos de 0.5 ms mientras que los tétanos se han obtenido mediante trenes de pulsos de 350-ms o 2-s para EDL o soleo respectivamente. Es evidente que la fuerza específica en fibras de EDL y soleo es significativamente menor en ratones viejos que en jóvenes. La fuerza específica se ha obtenido normalizando la fuerza absoluta al área de la sección transversal de la fibra (ver tabla 3). La línea de puntos indica la línea base (fuerza 0). Los números cerca de los registros indican el nombre del registro.
con el envejecimiento. Los registros de contracción del soleo se muestran en una
escala temporal diferente ya que las fibras del soleo, al ser más lentas, requieren un
mayor tiempo para alcanzar la fuerza máxima. La Figura 7 muestra los valores de
fuerza específica máxima obtenidos en cada una de las células registradas de EDL y
soleo. Es aparente que, aunque algunas fibras de ratones viejos desarrollaron una
fuerza específica similar a la de fibras de ratones jóvenes, un número significativo de
fibras de ratones viejos desarrolló una fuerza específica menor de 300 kPa.
La relación fuerza-frecuencia fue analizada en cada una de las fibras aplicando
43
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
un tren de pulsos (350 ms duración) a las siguientes frecuencias: 5, 10, 20, 30, 40, 50,
75, 100, 150 y 200 Hz. La Figura 8 muestra la relación fuerza- frecuencia de las fibras
de EDL y soleo. La frecuencia necesaria para alcanzar la fuerza máxima fue menor
500 1
03
?¿. 400 CD
E X
>CD
E CD O
'c >CD CD UJ
300
200
100
O
o
o o
D D
O
O
B
D
D
D
B B B
m
H S
H
S B
S S S
~~i r ME V
EDL
J ME V
SOLEO
Figura 7. Fuerza tetánica máxima específica desarrollada por fibras únicas e intactas de EDL y soleo de diferentes edades. Todas las fibras de EDL y soleo incluidas en este estudio han sido agrupadas de acuerdo con la distribución de la edad de los ratones en joven(J), mediana edad (ME) y viejo (V). Cada punto en el gráfico representa una sola medida de fuerza específica (FE). El asterisco (*) indica diferencias significativas con valores de otras edades del mismo músculo
en las fibras del soleo que en las de EDL. Cuando comparamos la curva fuerza-
frecuencia para cada uno de los tipos de fibras (rápidas o lentas) vemos que fibras de
ratones viejos necesitaron frecuencias de estímulo similares a las de ratones jóvenes
para desarrollar la máxima fuerza (Fig. 8).
44
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
'o' CL
n (D N i _
0) 13 LL
1,0 1
0,8
U,b J
0,4 j
0,2
EDL
J,,,^
• • •
Joven Mediana edad Viejo
- Joven Mediana Edad Viejo
25 50 75 100
SOLEO
1,0
o ü . CL
en ü CD 3 LL
0,8 •
0,6 •
0,4 •
0,2
,.4--í
25 50 75
Frecuencia (Hz)
100
Figura 8. Relación Frecuencia-Fuerza en fibras únicas intactas de los músculos EDL y soleo . Fuerza tetánica (relación (P)/fuerza tetánica máxima (Po)) - frecuencia de fibras de EDL y soleo de ratones jóvenes, de mediana edad y viejos. Las fibras de contracción lenta (soleo) alcanzan máxima fuerza a frecuencias menores que fibras de contracción rápida (EDL). La frecuencia a la que se obtiene la fuerza tetánica máxima se muestra en la Tabla 3. Los datos representan media ± EEM. Los signos (*) y (#) indican diferencias significativas entre fibras de ratones de mediana edad y viejos, respectivamente, y fibras de ratones jóvenes.
4.1.2. Cinética de contracción de fibras de EDL v soleo de ratones jóvenes,
de mediana edad y viejos
máxima- y
El tiempo al pico - definido como el tiempo necesario para desarrollar la fuerza
el tiempo de relajación media -tiempo desde el pico hasta el punto de
45
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
relajación correspondiente a la mitad de la máxima fuerza- se determinaron para las
contracciones aisladas de las fibras de EDL y soleo (Tabla 4). El tiempo al pico fue más
corto en fibras de EDL que de soleo de ratones jóvenes, de mediana edad y viejos.
Este parámetro fue más corto, aunque no significativamente, en fibras de soleo de
ratones de mediana edad comparado con las fibras de ratones jóvenes. El tiempo al
pico medido en fibras de soleo de ratones viejos no fue significativamente diferente del
de fibras de ratones jóvenes y de mediana edad. El tiempo medio de relajación fue
significativamente mas corto en fibras de EDL que de soleo de ratones jóvenes, de
mediana edad y viejos, sin embargo la comparación entre grupos de diferente edad no
mostró ninguna diferencia significativa para EDL o soleo. Esto indica que no se
observaron cambios significativos con la edad en el tiempo de relajación media.
TABLA 4
Cinética de contracción en fibras únicas intactas
EDL SOLEO
JOVEN MEDIANA VIEJO JOVEN MEDIANA VIEJO EDAD EDAD
n=11 n=8 n=25 n=12 n=10 n=7 Tiempo al pico(ms) 33 ±6.1 27 ±1.3 30 ±1.5 96 ± 7.5 + 73 ± 5.8 " 89 ±18 +
Tiempo medio de 62 ±12 60 ± 6.5 66 ± 8.2 130 ±14+ 170 ±18+ 170 ±39 + relajación
íms] Diferencias significativas entre fibras de EDL y soleo de ratones de la misma edad (+). Los valores indican la media ± EEM
46
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

CAPITULO II:
ÍNDICE DE FATIGA EN FIBRAS MUSCULARES DEL EDL Y SOLEO DE RATONES
JÓVENES Y VIEJOS
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
4.2.1. índice de fatiga en fibras musculares de EDL y soleo de ratones
¿gxynes V viejos
Hemos estudiado el índice de fatiga en la resistencia muscular de fibras de los
músculos EDL y soleo de ratón de distintas edades. El índice de fatiga, definido como
el cociente entre la fuerza máxima desarrollada en la última contracción tetánica y la
primera, se midió usando los protocolos de intervalo corto (IC) y largo (IL). El protocolo
IC produjo más fatiga que el protocolo IL. La figura 9 muestra la secuencia de
contracciones tetánicas en respuesta a los protocolos IL (A) y IC (B) en una célula del
músculo EDL de un ratón joven (5 meses de edad). El índice de fatiga fue de 0.78 y
0.28 para los protocolos IL e IC, respectivamente.
2,5 min
200 kPa
1 mío
Figura 9. Curso temporal de la fatiga en una célula intacta de EDL en respuesta a dos protocolos de estimulación diferentes. Registros típicos de fatiga de una fibra de EDL estimulada con el protocolo de intervalo largo (A) y corto (B) durante 10 y 5 min, respectivamente. Cada contracción tetánica aparece como una barra vertical. La fuerza ha sido normalizada con respecto al área de la sección transversal. El intervalo entre (A) y (B) fue de 5 min. Obsérvese la recuperación completa de la fuerza entre (A) y (B).
48
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.
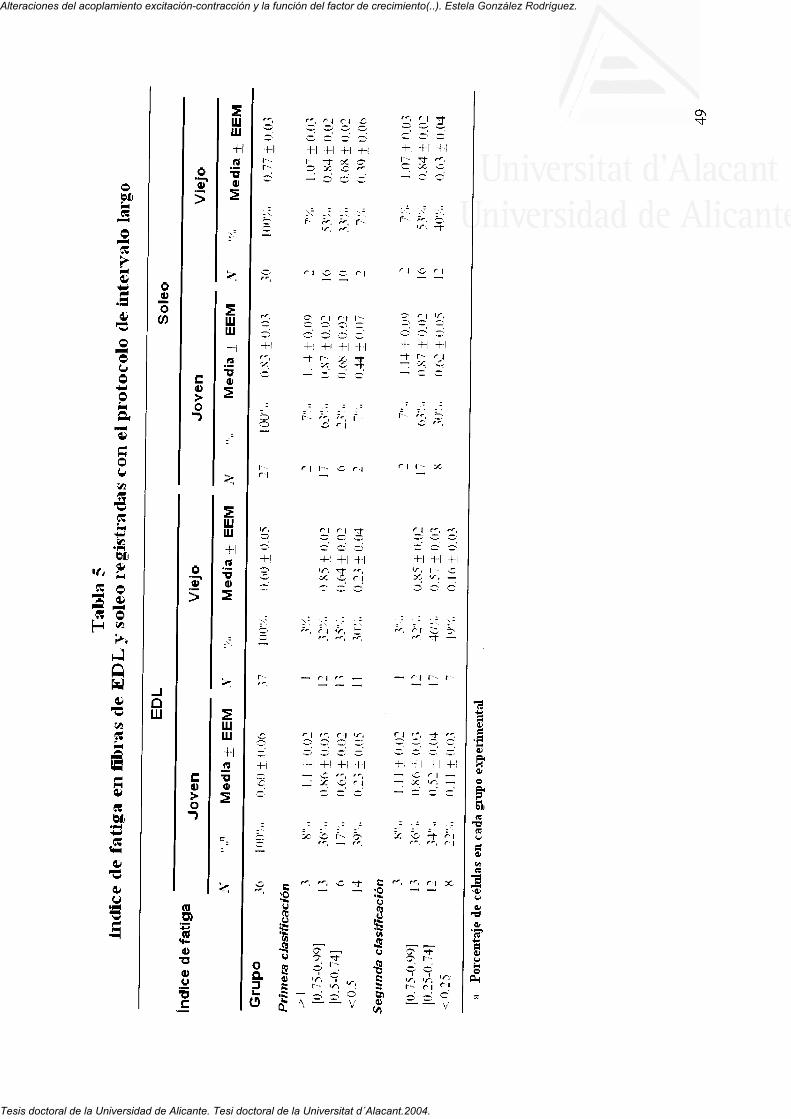
o
I
I o (J o
o
Vi
« o
Q
a;
;«
i c «
•¿
U
I
c > O
I
Q UJ
C
> O
0 3r >
s UJ m +1 2 •5 01 S
lU
•o «
lU
m +1 .2 (U
LU UJ
I •a
I
+1 i- •
r-
i--r I
+1
O
a O
'O
w ÍS Ai
5
+1 +1 +1 +1 I - Tf X r-~ X. vC ''-,
r-- r*-. r*-. !=••-
^1 O r. <-1
£ : CJ C-.' í "
+1 +1 +1 +! •t r- X "t
. >c. o -I-
1 - " , ' ^ I -O r I
r I I-- sC- r I
+1 +1 +1 ' ^ > ^ ' • ' ' ,
X sO r i
' . '' I 'r, C
<- I ' ' , r I i-r,
4-1 +1 +1 +1
+1 4-1 4-1 I - ^ ' • ' ,
C X sC
r- r', i
4-1 4-1 +\ -t r- r| -^ X o
4-1 4-1 4-1 >r, 1 - o se ly. —
4-1 4-1 +1 4-1 — -c ^ I — — X ly, ^
X >c- f- b' X o •+ i'I
I / ' . r -
U
8
u
a>
(A
r- r\ < I
I a
I
o
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

o u
C
I I •a
u
t o a> o
CO
lU
+1 .2
2
o ID
+1
c 0) > O
m m +1 .2
r_\ r_\ r_)
+-I +1 +1
r- r- r-•rt ''1 r I
-fl +1 +1 r- >r, c , X. C Tf
+1 +1
r- jc
r I '-^
+1 +1 r- —'
O
!?
C!
•I-i
© 4>
o
Q
—' C" i>l r 1
I
m lU
+1 .2 01
+1 H +1
— X
T —
tí)
Oí
a
Q LU
C 0) > o
lU
+1 .2
ot
+1
a
a I o» 0>
c
o a O
+1 X
c '0 ' s
••^
•5 u 2 s _ t J N
Q. •
2
r^,
, C> í?
r— — ^
t r ,
, , ' ^ •
I'--
1
—.'
JC
1'--
' •Tt
o
''
e-
«0 • u s •ft <A
•S u
c 9) 0) oo
2;
r'",
, , C--• ?
,-—-,
t - •
^ - i -r--' Y r 1
.'
_
r ¡ O '
'
íJ;
5i
5
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.
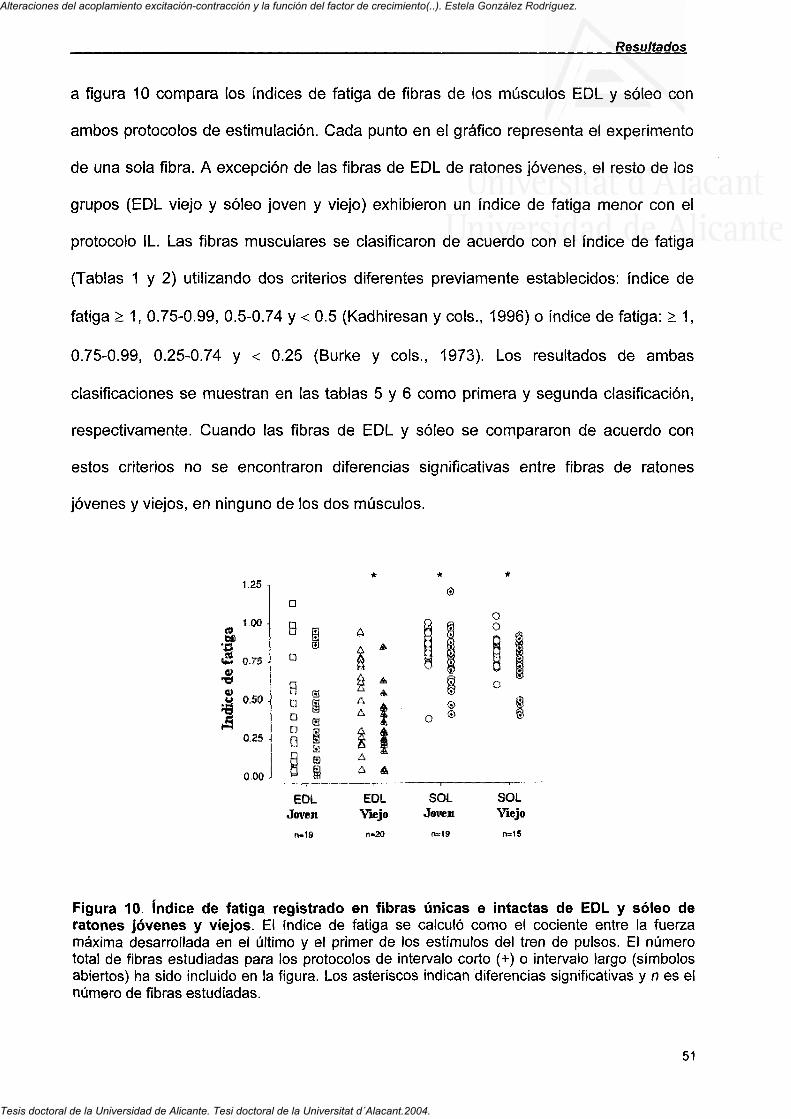
Resultados
a figura 10 compara los índices de fatiga de fibras de los músculos EDL y soleo con
ambos protocolos de estimulación. Cada punto en el gráfico representa el experimento
de una sola fibra. A excepción de las fibras de EDL de ratones jóvenes, el resto de los
grupos (EDL viejo y soleo joven y viejo) exhibieron un índice de fatiga menor con el
protocolo IL. Las fibras musculares se clasificaron de acuerdo con el índice de fatiga
(Tablas 1 y 2) utilizando dos criterios diferentes previamente establecidos: índice de
fatiga > 1, 0.75-0.99, 0.5-0.74 y < 0.5 (Kadhiresan y cois., 1996) o índice de fatiga: > 1,
0.75-0.99, 0.25-0.74 y < 0.25 (Burke y cois., 1973). Los resultados de ambas
clasificaciones se muestran en las tablas 5 y 6 como primera y segunda clasificación,
respectivamente. Cuando las fibras de EDL y soleo se compararon de acuerdo con
estos criterios no se encontraron diferencias significativas entre fibras de ratones
jóvenes y viejos, en ninguno de los dos músculos.
1.25
_ 100 (O
3 Tí
Oí
a 0,50 -j
0.75
0.25
0,00
D
B
a
a D • B
a i a
E B
EDL Joven
n.1S
A A
A
A
EDL Viejo
n«20
I
SOL Joven
tet9
O
o
SOL Viejo
n=tí
Figura 10. índice de fatiga registrado en fibras únicas e intactas de EDL y soleo de ratones Jóvenes y viejos. El índice de fatiga se calculó como el cociente entre la fuerza máxima desarrollada en el último y el primer de los estímulos del tren de pulsos. El número total de fibras estudiadas para los protocolos de intervalo corto (+) o intervalo largo (símbolos abiertos) ha sido incluido en la figura. Los asteriscos indican diferencias significativas y n es el número de fibras estudiadas.
51
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
El índice de fatiga no mostró correlación alguna con la fuerza específica
registrada en cada una de las fibras. En concreto, la correlación índice de fatiga-fuerza
específica máxima para células de EDL y soleo de ratones jóvenes y viejos tuvo
valores r de 0.148, 0.0015, 0.046 y 0.22, respectivamente (P < 0.05).
4.2.2. Curso temporal del índice de fatiga de fibras de EDL y soleo de
ratones jóvenes y viejos
Para determinar si la disminución de fuerza con el tiempo era diferente con la
edad, el índice de fatiga fue medido cada minuto, durante un total de 10 minutos
usando los protocolos IL y IC. La figura 11 muestra el cambio del índice de fatiga con el
tiempo de fibras de EDL (A) y soleo (B) usando el protocolo IL. Es obvio que las células
del músculo soleo se fatigaron menos que las de EDL y que el índice de fatiga en
células de ratones viejos es similar al de células de ratones jóvenes, en ambos
músculos. El curso temporal del índice de fatiga para fibras de EDL (A) y soleo (B)
usando el protocolo IC se muestra en la figura 12. Como puede observarse, el índice
de fatiga producido por el protocolo IC fue marcadamente superior al obtenido con el
protocolo IL (Figs. 1-3). También puede observarse que la resistencia muscular de
fibras de ratones jóvenes y viejos no mostró diferencias significativas. En conclusión,
estos experimentos muestran que ni el cociente entre el primer y último estímulo
tetánico (índice de fatiga) ni la evolución temporal de este difieren en fibras de los
músculos EDL y soleo de ratones jóvenes y viejos.
52
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.
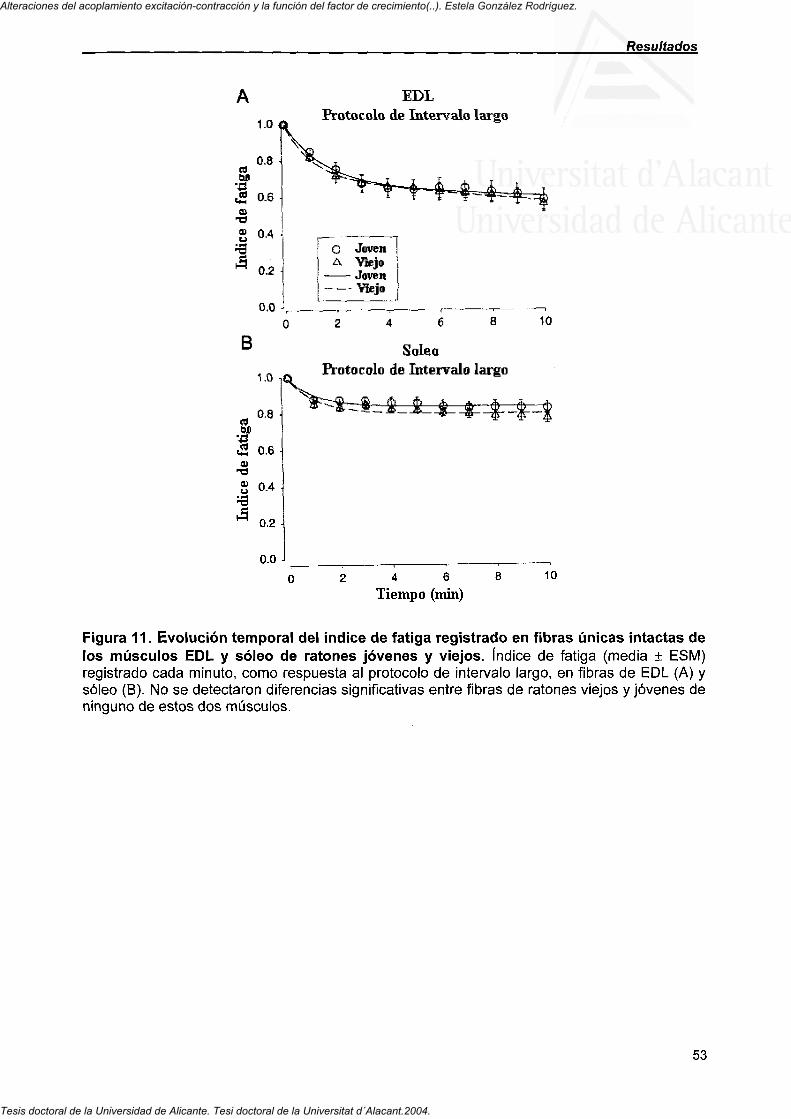
Resultados
a A
B
0,2
0.0
1.0
^ 0,8 -j bu
<^ 0.6
S 0.4 ía
^ 0.2
0.0
EDL Protocolo de Intervalo largo
O Joven A Viejo
Joven l^jo
2 4 6 8
Soleo Protocolo de Intervalo largo
10
2 4 6 8
Tiempo (min) 10
Figura 11. Evolución temporal del índice de fatiga registrado en fibras únicas intactas de los músculos EDL y soleo de ratones jóvenes y viejos. índice de fatiga (media ± ESÍVI) registrado cada minuto, como respuesta al protocolo de intervalo largo, en fibras de EDL (A) y soleo (B). No se detectaron diferencias significativas entre fibras de ratones viejos y jóvenes de ninguno de estos dos músculos.
53
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.
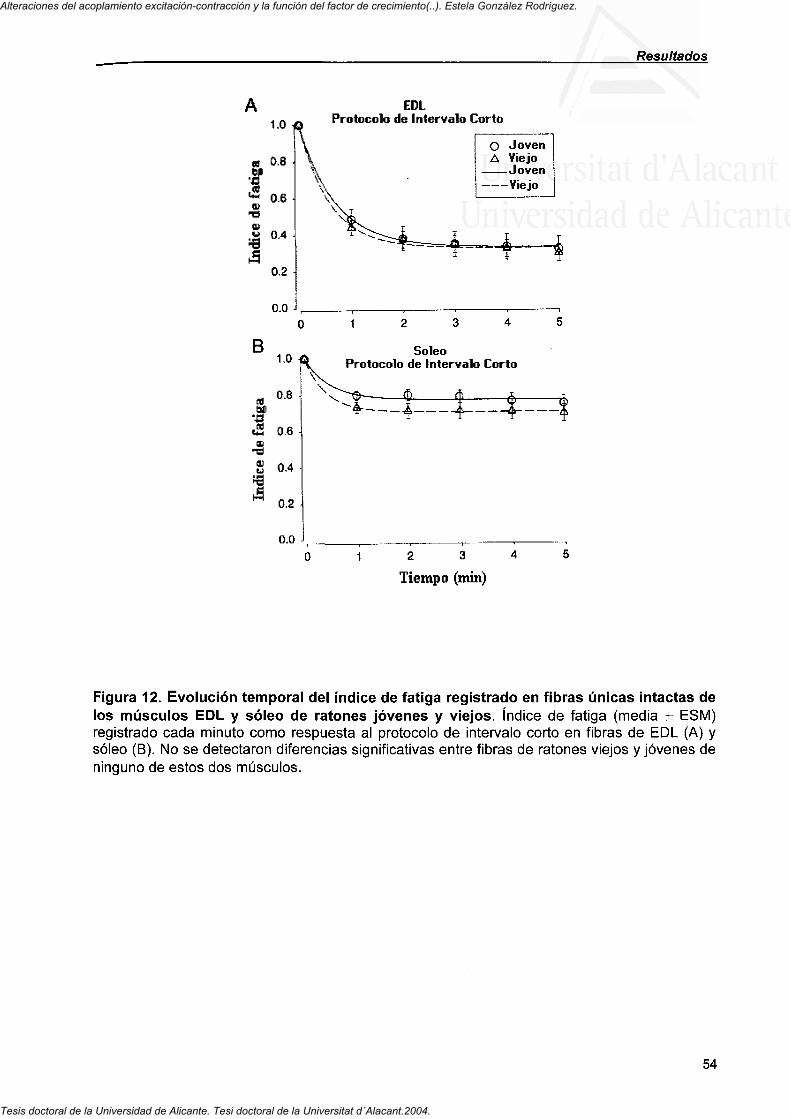
Resultados
EDL Protocolo de Intervalo Corto
O Joven A Viejo
Joven Viejo
' " ^ T ^ ^ = -'y = '~- ffl •
B 1,0
^ 0.6 m
ni
0.2
0.0
\ V
Soleo Protocolo de Intervalo Corto
4 L 4- zf—4:3
2 3 4
Tiempo (min)
Figura 12. Evolución temporal del índice de fatiga registrado en fibras únicas intactas de ios músculos EDL y soleo de ratones jóvenes y viejos. índice de fatiga (media + ESM) registrado cada minuto como respuesta al protocolo de intervalo corto en fibras de EDL (A) y soleo (B). No se detectaron diferencias significativas entre fibras de ratones viejos y jóvenes de ninguno de estos dos músculos.
54
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

CAPITULO III:
EL FACTOR DE CRECIMIENTO SIMILAR A LA INSULINA-I PREVIENE LA
DISMINUCIÓN DE Ca^* INTRACELULAR Y FUERZA ESPECÍFICA EN FIBRAS
ÚNICAS E INTACTAS DE MÚSCULOS DE RATONES TRANSGÉNICOS
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
4.3.1. La expresión de IGF-1 en músculos FDB de ratones transgénícos
Diversos grupos han demostrado que la expresión de hlGF-1 en ratones
transgénicos S1S2 es sostenida (Coleman y cois., 1995; Renganathan y cois., 1998).
El alto nivel de expresión transgénica resulto en concentraciones de IGF-1 en músculo
esquelético de por lo menos 47 la de músculos normales (Coleman y cois., 1995). Sin
embargo, la expresión de IGF-1 en músculos específicos no se conoce. Existen
evidencias de que el nivel de expresión de IGF-1 en animales transgénicos varia de
músculo a músculo (Musaro y cois., 2001). Por todo esto, determinamos la
concentración de hlGF-1 en el músculo FDB y la comparamos con la del EDL (otro
músculo rápido de la pata trasera) de ratones jóvenes y viejos controles y transgénicos.
Las medidas se tomaron en músculos diferentes a los usados en los experimentos de
contracción y fluorescencia. Para hacer estas determinaciones, cuatro o cinco
músculos de dos o tres ratones se utilizaron simultáneamente y los resultados se
normalizaron a la cantidad de proteína muscular. La concentración de hlGF-1 en los
músculos FDB y EDL de ratones jóvenes y viejos normales estuvo por debajo del nivel
de detección. En ratones transgénicos, el valor medio de tres determinaciones de la
concentración de hlGF-1 en ratones jóvenes y viejos fueron, para los músculos FDB:
255 + 18 and 58 ± 4, y en músculos EDL : 250 ± 22 y 144 ± 16 ng (mg proteína) para
los músculos FDB y EDL, respectivamente. En conclusión, la expresión de hlGF-1 en
ratones transgénicos jóvenes es similar en los músculos FDB y EDL y que, en ambos
músculos, disminuye con la edad.
56
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
4.3.2. La fuerza específica de músculo FDB entero de ratones normales y
IGF-1 -transgénicos
La fuerza específica se midió en músculos FDB de ratones jóvenes (n=6) y
viejos (n=10) control y jóvenes (n=8) y viejos (n=8) transgénicos. Ambas fuerzas, de
contracción aislada y tétano, se normalizaron con respecto al área de la sección
transversal (AST) de la fibra. Los valores de fuerza especifica tetánica registrados en
estos experimentos son similares a los descritos previamente para los músculos EDL y
cuadríceps (Fitts y cois., 1984) (Eddinger y cois., 1986), pero inferiores a los referidos
para los músculos EDL y soleo (Brooks y cois., 1988, Barton-Davis y cois., 1998). Una
posible explicación podría ser que un número significativo de fibras no desarrolla la
fuerza máxima debido a la compleja orientación e inserción del tendón del músculo
FDB.
C5
160
140
120
J 100 S 80 'é 60 es N 40
É 20 ^
o
Contracción aislada Tétano
1 Jo\'en Joven Viejo Viejo
üansgéiüco taansgénico
Figura 13. Fuerza específica máxima de músculos FDB de ratones controles e IGF-1-transgénicos jóvenes y viejos. La fuerza de contracción tetánica disminuyó significativamente en músculos de ratones viejos control comparado con ratones jóvenes control e IGF-1 jóvenes y viejos transgénicos (P<0.05). No se detectaron diferencias significativas entre jóvenes control o transgénicos y viejos transgénicos. La fuerza específica de contracción aislada no fue significativamente diferente entre grupos. *, significativamente diferente. Los valores representan la media + E.S.M.
57
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
FDB. La figura 13 muestra los valores correspondientes a estos cuatro grupos
experimentales. La fuerza específica de la contracción aislada en músculos FDB no
mostró diferencias significativas entre los cuatro grupos. La fuerza específica tetánica
disminuyó significativamente en los ratones viejos (92 ± 6 kPa) comparada con la de
ratones jóvenes control (135 ± 17 kPa) y transgénicos (126 ± 8 kPa) (P < 0.05). El
porcentaje de disminución de fuerza específica con la edad fue del 32 % y es
consistente con el de publicaciones previas en músculo rápido EDL de ratones y ratas
(ver debajo). Los valores de fuerza especifica obtenidos para ratones jóvenes control y
transgénicos no fueron significativamente distintos. La fuerza específica registrada en
ratones viejos transgénicos (114 ± 7 kPa) no fue significativamente distinta a la
registrada en ratones jóvenes control o transgénicos, pero fue significativamente mayor
que la registrada en ratones viejos control. Estos resultados indican que la expresión
transgénica de IGF-1 en músculo esquelético previene la disminución de la fuerza
específica máxima del músculo entero que ocurre con el envejecimiento.
Estudios previos de nuestro laboratorio han demostrado que el pico intracelular
de Ca "" disminuye en fibras musculares de ratones viejos medido con la técnica de
fijación de voltaje (Wang y cois., 2000). Para examinar si la edad y/o el IGF-1 modifican
el pico intracelular de [Ca ""]; en fibras contráctiles, registramos simultáneamente la
contracción y la concentración de Ca "" intracelular en fibras únicas intactas del músculo
FDB de ratones jóvenes y viejos controles e IGF-1-transgénicos.
4.3.3. Área de la sección transversal v fuerza especifica máxima registrada
en células únicas intactas del músculo FDB
El análisis del área de la sección transversal de las fibras del músculo FDB
58
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
mostró la existencia de atrofia con el envejecimiento, proceso que fue prevenido con la
expresión transgénica de IGF-1. El diámetro y el AST de las fibras de ratones viejos
control fue: 23 ± 0.9 ^m y 436 ± 55 )Lim , respectivamente (n = 20). Estos valores fueron
significativamente menores que los registrados en fibras de ratones jóvenes control ( 29
+ 1.2 y 671 ± 41 i m , n = 17), jóvenes transgénicos ( 36 ± 2.3 y 1054 fam , n= 9) y
viejos transgénicos (31 ± 1.8 ^m y 781 ± 82 |u.m , n= 14). También, el diámetro y el AST
de las fibras de ratones transgénicos jóvenes fueron significativamente mayores
comparados con los de ratones jóvenes control, pero no los valores de ratones viejos
transgénicos.
La curva frecuencia-fuerza fue representada con los datos de la fuerza
desarrollada a diferentes frecuencias, de menor a mayor. La curva frecuencia-fuerza se
registró para la mayoría de las fibras usando trenes de 350 ms de duración y frecuencia
variable. Esta curva nos permitió encontrar la frecuencia a la que se alcanzó la fuerza
máxima de cada fibra. La secuencia de frecuencias utilizadas fue 5, 10, 20, 50, 75 y
100 Hz. La estimulación de la fibras se interrumpió cuando la fuerza específica
desarrollada a dos frecuencias consecutivas fue la misma. La mayoría de las fibras
alcanzó la fuerza máxima a 75 o 100 Hz. En algunos casos se necesitó una
estimulación mas prolongada (400 ms) para alcanzar la fuerza máxima. La figura 14A
muestra la curva fuerza-frecuencia de fibras de FDB de ratones jóvenes y viejos,
controles y transgénicos. Los valores se han normalizado a la fuerza máxima de cada
célula. No se encontraron diferencias significativas en la fuerza específica de la
contracción aislada entre los cuatro grupos experimentales. Estos resultados indican
que ni la edad ni la expresión de IGF-1 ejercen un efecto significativo en la frecuencia
necesaria para desarrollar la fuerza máxima. La figura 15 muestra registros
representativos de la fuerza de contracción aislada (Aa) y tetánica (Ba) de fibras únicas
de FDB. La fuerza especifica de la contracción aislada no se modificó
59
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
significativamente con la edad o la expresión transgénica de IGF-1; sin embargo, la
disminución de la fuerza específica máxima tetánica registrada en fibras de ratones
viejos control no se observó en ratones viejos transgénicos. La figura 16A compara la
A
o
e
o fe
+
U
1.0
'S 0.8
0.6
0.4
0.2
1,0
Joven Joven Tg Viejo Viejo Tg Joven Joven Tg Viejo Viejo Tg
20 40 60 80 100
- I 0.8
0.6 T / ;
0,4
0,2
i
20 40 60 80
Frecuencia (Hz)
100
Figura 14. Relación fuerza-frecuencia y [Ca *]i-frecuencia en células únicas intactas de músculos FDB de ratones controles e IGF-1 transgénicos, Jóvenes y viejos. La fuerza (A) y concentración mioplásmica de Ca "" (B) han sido normalizados por el valor máximo de cada fibra. Los datos representan la media ± E.S.M. Los resultados han sido ajustados a una función exponencial simple. No existen diferencias significativas entre los diferentes grupos.
60
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.
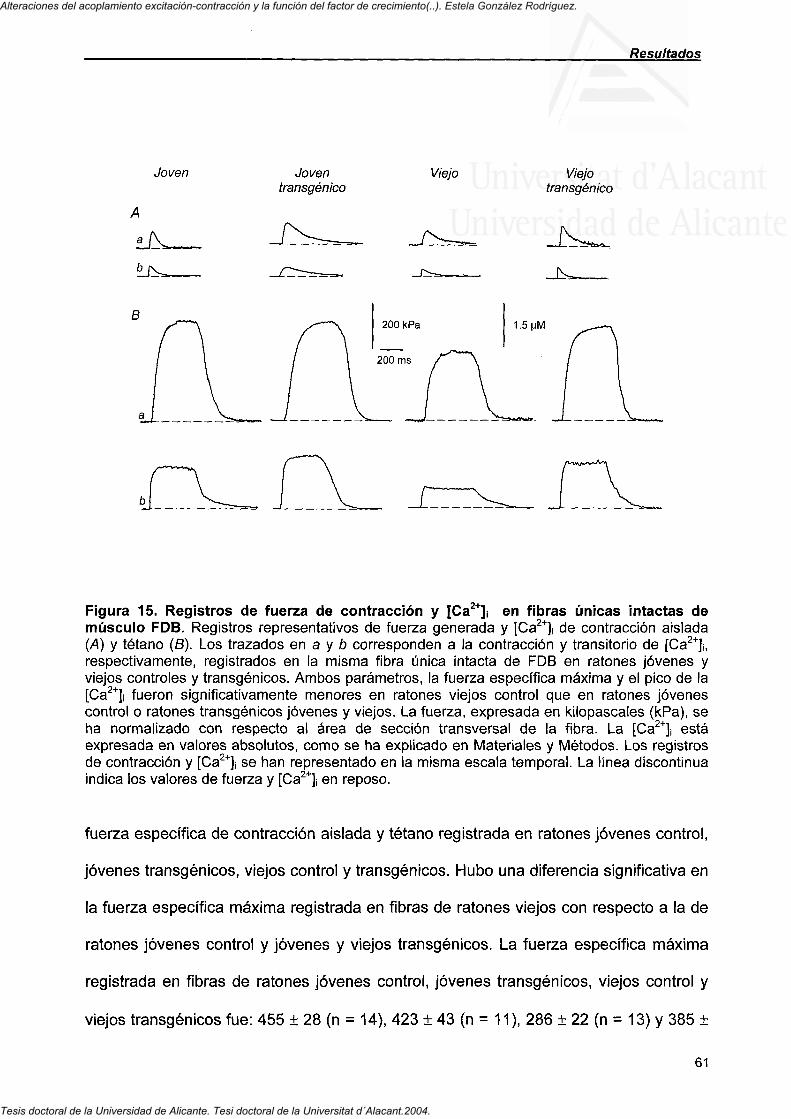
Resultados
Joven Joven transgénico
Viejo
^JV_
Viejo transgénico
Figura 15. Registros de fuerza de contracción y [Ca^ ]¡ en fibras únicas intactas de músculo FDB. Registros representativos de fuerza generada y [Ca ""]! de contracción aislada (A) y tétano (fí). Los trazados en a y ib corresponden a la contracción y transitorio de [Ca ""]!, respectivamente, registrados en la misma fibra única intacta de FDB en ratones jóvenes y viejos controles y transgénicos. Ambos parámetros, la fuerza específica máxima y el pico de la [Ca ""]! fueron significativamente menores en ratones viejos control que en ratones jóvenes control o ratones transgénicos jóvenes y viejos. La fuerza, expresada en kilopascales (kPa), se ha normalizado con respecto al área de sección transversal de la fibra. La [Ca ""]! está expresada en valores absolutos, como se ha explicado en IVIateriales y Métodos. Los registros de contracción y [Ca ^ ji se han representado en la misma escala temporal. La línea discontinua indica los valores de fuerza y [Ca^ ji en reposo.
fuerza específica de contracción aislada y tétano registrada en ratones jóvenes control,
jóvenes transgénicos, viejos control y transgénicos. Hubo una diferencia significativa en
la fuerza específica máxima registrada en fibras de ratones viejos con respecto a la de
ratones jóvenes control y jóvenes y viejos transgénicos. La fuerza específica máxima
registrada en fibras de ratones jóvenes control, jóvenes transgénicos, viejos control y
viejos transgénicos fue: 455 ± 28 (n = 14), 423 ± 43 (n = 11), 286 + 22 (n = 13) y 385 ±
61
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.
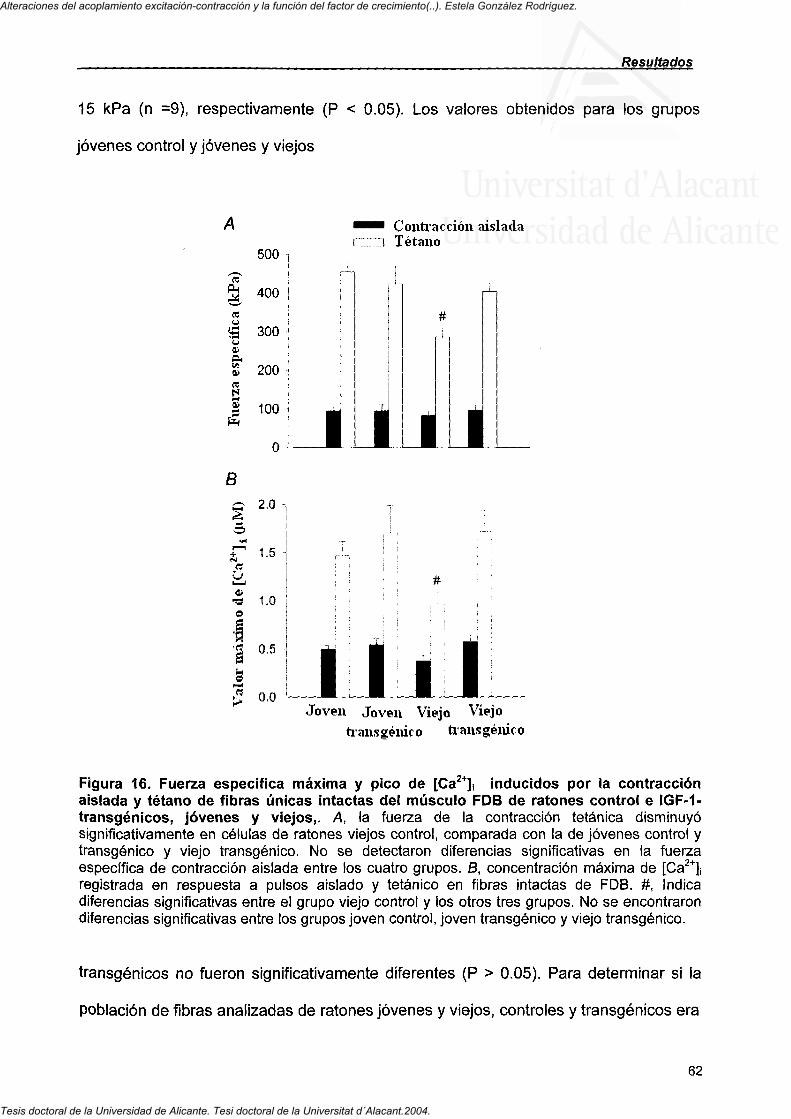
Resultados
15 kPa (n =9), respectivamente (P < 0.05). Los valores obtenidos para los grupos
jóvenes control y jóvenes y viejos
f^
B S- 2,0
+
o
>
1.5
•« 1.0 o
I tí
0,5
0.0
• • • • Contiacción ai,slada I : ::J Tétano
ouu -
400
300
100
n
J -] _ í t
j.lilJU
lili Joven Joven Viejo Viejo
tian.s£émco ti an-sgénico
Figura 16. Fuerza específica máxima y pico de [Ca^ ]¡ inducidos por la contracción aislada y tétano de fibras únicas intactas del músculo FDB de ratones control e IGF-1-transgénícos, jóvenes y viejos,. A, la fuerza de la contracción tetánica disminuyó significativamente en células de ratones viejos control, comparada con la de jóvenes control y transgénico y viejo transgénico. No se detectaron diferencias significativas en la fuerza específica de contracción aislada entre los cuatro grupos. B, concentración máxima de [Ca ""], registrada en respuesta a pulsos aislado y tetánico en fibras intactas de FDB. #, Indica diferencias significativas entre el grupo viejo control y los otros tres grupos. No se encontraron diferencias significativas entre los grupos joven control, joven transgénico y viejo transgénico.
transgénicos no fueron significativamente diferentes (P > 0.05). Para determinar si la
población de fibras analizadas de ratones jóvenes y viejos, controles y transgénicos era
62
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
diferente, analizamos la velocidad de generación de fuerza (dP/df) normalizada por la
fuerza máxima, el tiempo al pico y el tiempo medio de relajación de la contracción
aislada (Tabla 7). El análisis estadístico no mostró ninguna diferencia significativa entre
los cuatro grupos para ninguno de los tres parámetros medidos. Para determinar el
fenotipo de las fibras estudiadas, investigamos la expresión de cadena pesada de
miosina (CPM) (ver debajo).
Tabla Vil
Cinética del Ca ^ intracelular y la contracción twitch en fibras únicas e
intactas del músculo FOB
Joven Joven Viejo Viejo control transgénico control transgénico
dP/dt(ms-') Normalizado
Tiempo al pico (ms)
Tiempo de relajación
media (ms)
dCa^Vdt (pM/ms)
0.035 ± 0.004
38 ±2
68 ± 6
0.05 ±0.01
0.033 ± 0.004
43 ± 1
71 ± 5
0.06 ± 0.01
0.038 ± 0.007
45 ± 3
87 ± 7
0.06 ±0.01
0.037 ± 0.006
42 ± 4.5
81 ±11
0.07 ± 0.02
Los valores se han expresado como la media ± EEM de 14-17 fibras por grupo. El análisis estadístico entre grupos no mostró diferencias significativas (P > 0.05). dP/dt es la velocidad de desarrollo de fuerza y se ha normalizado a la fuerza de contracción aislada. dCa^Vdt es la velocidad de liberación de Ca "".
63
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
4.3.4. Valores máximos de fCa ""!! en fibras únicas intactas del músculo
FPB
Los incrementos transitorios de la [Ca^ ]¡ se midieron simultáneamente con la
contracción usando el indicador de Ca "" fluorescente fluo 3-AM. La figura 15Ab y Bb
muestra transitorios de Ca "" intracelular en respuesta a estimulaciones breves y
prolongadas, respectivamente, en fibras intactas de músculo FDB de ratones jóvenes y
viejos control y transgénicos. Los valores pico de la [Ca^""]! registrada en ratones
transgénicos viejos no fue significativamente diferente a la registrada en ratones
jóvenes y viejos control y jóvenes transgénicos en respuesta a estimulaciones aisladas.
El valor de pico de la [Ca^^Ji fue significativamente menor en fibras de ratones viejos
control comparado con ratones jóvenes control y transgénicos, en respuesta a
estimulaciones prolongadas (tétano), y que esta disminución se pudo prevenir con la
expresión transgénica de IGF-1.
Para determinar si la edad y/o IGF-1 modifican la curva frecuencia-íCa^""], ,se
midió la [Ca^""]! en respuesta a diferentes frecuencias de estimulación en fibras de FDB
de ratones jóvenes y viejos, controles y transgénicos (Fig. 28). Los valores de [Ca^^Ji
son inferiores al 100% porque la concentración de Ca " ha sido normalizada por el
valor máximo alcanzado en cada fibra, que no siempre se obtuvo a 100Hz (en algunas
células se alcanzó a 75 Hz). Los valores normalizados de [Ca^*] no mostraron
diferencias en la frecuencia de estímulo necesaria para alcanzar el pico intracelular de
Ca^* para ninguno de los grupos experimentales, en correspondencia con la curva
frecuencia-fuerza mostrada en la figura 14A. No se detectaron diferencias significativas
en los valores de pico de [Ca^*]; de fibras de ratones jóvenes control y transgénicos en
respuesta a la estimulación tetánica (Fig. 43). Sin embargo, los valores obtenidos en
64
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
fibras de ratones viejos controles en respuesta a la estimulación tetánica fueron
significativamente menores que los registrados en los otros tres grupos. Entre los
grupos joven control y transgénico y viejo transgénico no se encontraron diferencias
significativas en este parámetro (P>0.05). Los valores de pico intracelular de [Ca^*]¡ en
respuesta a un estímulo prolongado fueron 1.47 ± 0.15, 1.70 ± 0.29, 0.97 ± 0.13 y 1.7 ±
0.22 |a.M, para fibras de ratón joven control, joven transgénico, viejo control y viejo
transgénico, respectivamente. Los valores de los transitorios de [Ca^*]¡ evocados por
una a estimulación aislada fueron: 0.50 ± 0.04, 0.54 ± 0.07, 0.38 ± 0.06 y 0.57 + 0.07
laM, para fibras de ratón joven control, joven transgénico, viejo control y viejo
transgénico, respectivamente. Estos valores no son significativamente diferentes. La
velocidad de cambio de la [Ca^^Ji fue calculada en experimentos individuales como la
primera derivada de la señal del transitorio de Ca^* en respuesta a una estimulación
aislada. Este parámetro, d[Ca^*]i, representa la liberación de Ca "" del RS por pulsos
breves (Melzer y cois., 1987). La velocidad de aumento en [Ca^""]! (díCa^""]! /dt) no fue
significativamente diferente entre los cuatro grupos (P > 0.05, Tabla 7). La velocidad
de disminución de [Ca^^Ji no se midió debido a la cinética lenta de la disociación del
complejo fluo 3-03 "*".
4.3,5. Efecto de la cafeína en la disminución de la fuerza específica y en el
pico de rCa^ liCon el envejecimiento
La figura 17A muestra el efecto potenciador de la cafeína sobre la fuerza
tetánica máxima de fibras de ratones jóvenes control (n = 8), jóvenes transgénicos (n =
8), viejos control (n = 15) y viejos transgénico (n = 7). La fuerza especifica máxima y los
valores de pico de [Ca^""]! fueron medidos simultáneamente en 7-8 fibras. La diferencia
65
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
en el número de experimentos refleja la inclusión en los análisis de fibras en las que
sólo uno de los dos parámetros fue medido, o la exclusión de aquellas en las que la
medida no fue precisa debido a razones técnicas. La potenciación inducida por cafeína
es consistente con el aumento de liberación de Ca "" inducido por ella, en células únicas
observado con la técnica de fijación de voltaje (Delbono y cois., 1995). Los valores de
fuerza específica obtenidos en fibras de ratones jóvenes y viejos control expuestas a
cafeína fue significativamente diferente (P < 0.05).
, _ K
w (J
a> P<
« y a>
| i (
700 1
600
500
400 •
3ÜÜ
200 •
100
D
B
a
3.0
2.5
2.0
^ 1.5
I -° S 0.5
« 0.0
Contiol Cafeína
Joven Joven Viejo Viejo
tianssémco tiansgéiuco
Figura 17. Efecto de la cafeína en la fuerza específica máxima y en la [Ca ]¡ de fibras únicas intactas de ratones controles y transgénicos, jóvenes y viejos. La fuerza específica (A) y el pico de Y^^^\ (B) antes y después de la aplicación de cafeína 5 mM (efecto máximo tras 30 min de incubación). * y # , indican diferencias significativas en el mismo grupo antes y después de cafeína y entre ratones viejos control y el resto de los grupos antes de cafeína, respectivamente. **, indica diferencias significativas entre fibras de ratones jóvenes y viejos tratados con cafeína.
66
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
Sin embargo, no se detectaron diferencias significativas en las fibras de ratones
jóvenes antes de la exposición a cafeína y las fibras de ratones viejos después del
tratamiento con cafeína. Además, no se encontraron diferencias significativas entre
fibras de los grupos joven control y transgénico y viejo transgénico después de la
exposición a cafeína. En resumen, estos resultados muestran que la cafeína puede
restaurar la fuerza especifica de células de FDB de ratones viejos al nivel de las fibras
de ratones jóvenes antes de la exposición a cafeína; sin embargo, persiste un deficit
cuando las fibras de ambos grupos son comparadas después del tratamiento con
cafeína.
La relación fuerza-frecuencia de fibras de ratones jóvenes y viejos, control y
transgénicos alcanzó la meseta durante el tétano, lo que sugiere que se alcanzó la
activación máxima. Para determinar si una disminución de la liberación de Ca "" es la
causa responsable de la pérdida de fuerza observada en células de ratones viejos, se
estudió el efecto de la cafeína sobre los valores máximos de [Ca^*]i . La figura 17B
representa el efecto de la cafeína sobre la [Ca^""]!, medido en fibras únicas intactas de
FDB de ratones jóvenes control (n = 9), jóvenes transgénicos (n = 7), viejos control (n =
8) y viejos transgénico (n = 7). El efecto máximo de la cafeína se obtuvo siempre en los
primeros 30 minutos. Como se puede observar, la cafeína aumentó significativamente
los valores de pico de [Ca^""]! en los cuatro grupos experimentales, aunque el efecto
fue más notorio en fibras de ratones viejos control que en el resto de los grupos. Las
diferencias no fueron estadísticamente significativas.
4.3.6. Tipo de fibras que componen el músculo FDB a distintas edades en
ratones control y transgénicos (IGF-1)
Para determinar si la disminución de fuerza específica registrada en músculo
67
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
entero y fibras únicas de FDB está causada por cambios en la composición del tipo de
fibra, se analizó la composición de la cadena pesada de miosina (CPM), usando la
técnica de inmunohistoquímica descrita anteriormente (ver Material y Métodos). La
figura 18 muestra secciones discontinuas en serie del músculo FDB analizado
mediante inmunohistoquímica con anticuerpos en contra de los tipos de fibra I, lia y lib.
Esta técnica nos permitió cuantificar el porcentaje relativo de los subtipos de fibra en el
músculo FDB. La tabla 8 muestra el análisis estadístico de la caracterización del tipo de
fibra de 4-5 músculos de los usados para los experimentos de contracción
Jo\'en Jo"ven
ti í»ii?; fféiüí o ^ lej o tiniis^éiuío
Tipo I
Tipo n b
Tipo Ha
Figura 18. Composición del músculo FDB en tipo de fibras con la edad y la expresión transgénica de IGF-1. Secciones en serie representativas de músculos FDB marcados inmunohistoquímicamente con los anticuerpos de fibras de tipo I, lia, lib de ratones jóvenes y viejos control y IGF-1-transgénicos. La cuantificación de los porcentajes relativo de cada tipo de fibras se muestra en la Tabla 8.
en este trabajo. El número total de fibras por músculo no fue significativamente
diferente entre los grupos, por lo que la determinación de la proporción relativa de
subtipos de fibras fue hecha en base a un número comparable, independientemente de
la edad o el genotipo considerado. Las fibras que expresan CPM del tipo lib no se
detectaron en ninguno de los cuatro grupos. La isoforma o tipo de MHC predominante
68
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.
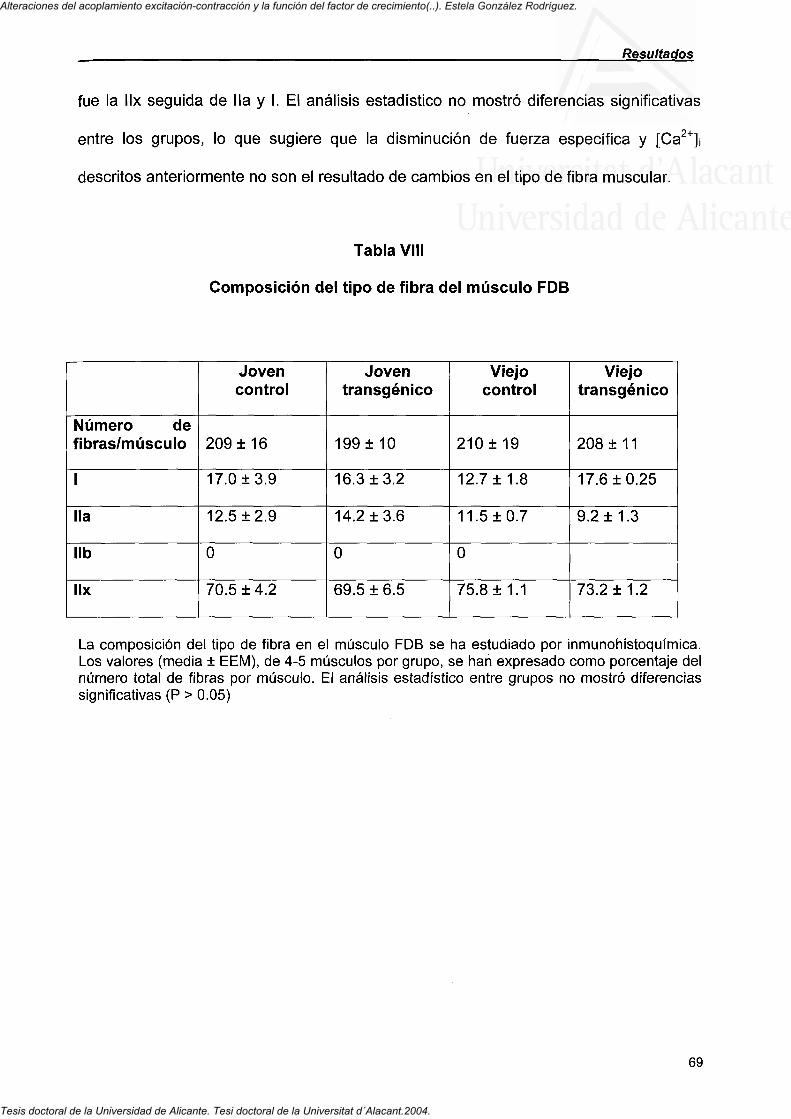
Resultados
fue la llx seguida de lia y I. El análisis estadístico no mostró diferencias significativas
entre los grupos, lo que sugiere que la disminución de fuerza específica y [Ca ""]]
descritos anteriormente no son el resultado de cambios en el tipo de fibra muscular.
Tabla VIII
Composición del tipo de fibra del músculo FDB
Número de fibras/músculo
1
lia
lib
llx
Joven control
209 ±16
17.0 ±3.9
12.5 ±2.9
0
70.5 ±4.2
Joven transgénico
199 ±10
16.3 ±3.2
14.2 ±3.6
0
69.5 ± 6.5
Viejo control
210±19
12.7 ±1.8
11.5 ±0.7
0
75.8 ±1.1
Viejo transgénico
208 ± 11
17.6 ±0.25
9.2 ±1.3
73.2 ±1.2
La composición del tipo de fibra en el músculo FDB se ha estudiado por inmunohistoquímica. Los valores (media ± EEM), de 4-5 músculos por grupo, se han expresado como porcentaje del número total de fibras por músculo. El análisis estadístico entre grupos no mostró diferencias significativas (P > 0.05)
69
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

CAPITULO IV:
EVENTOS ELEMENTALES DE LIBERACIÓN DE Ca^\ " CHISPAS" Y "ASCUAS",
EN FIBRAS DE MÚSCULO ESQUELÉTICO DE RA TONES VIEJOS
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
RESULTADOS
En este estudio hemos investigado si el envejecimiento produce cambios en la
frecuencia, cinética y/o morfología de un gran número de EELC registrados en fibras
peladas químicamente de músculo EDL de ratones jóvenes y viejos. Para ello, hemos
registrado EELC en los modos x-y y escan de línea, hemos analizado la composición
de CPM de las fibras utilizadas en los registros de fluorescencia, y hemos analizado la
expresión de RyR1 y RyR3 en músculo EDL con immunoblots. Además, hemos
investigado la influencia de la función mitocondrial en la generación de EELC en fibras
de EDL de ratones jóvenes y viejos.
.2+ 4.4.1. Eventos de liberación de Ca en imágenes XY y de escan de línea
Para analizar los EELC en el modo x-y hemos escaneado sistemáticamente varios
lugares (3-5 de acuerdo con la longitud de la fibra) de fibras de EDL orientadas
paralelamente a la dirección de escaneo x, y hemos registrado series de 50 imágenes
con un intervalo de 1.2 o 3.2 s por imagen. La figura 1A y B muestra imágenes x-y
registradas en fibras de EDL de un ratón joven y viejo, respectivamente, adquiridas en
una secuencia de 50 imágenes a intervalo de 3.2 s por imagen. Se detectó un menor
número de eventos en fibras de ratones viejos comparados con ratones jóvenes (ver
debajo para los estadísticos). También estudiamos fibras de EDL de grupos de la
misma edad en el modo de escan de línea. Las fibras se escanearon a lo largo de todo
el diámetro, a intervalos de 20 píxeles.
71
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
JOVíl l Vifjo
Figura 19. Chispas y "ascuas" de Ca " espontáneas de fibras permeabilizadas químicamente de EDL de ratones jóvenes y viejos. Ay B corresponden a imágenes XY de fluorescencia normalizada de una serie xyt de 50 imágenes (intervalo 3.2 s), que muestran chispas de Ca " de un ratón joven y uno viejo, respectivamente. C y D ilustran eventos representativos de liberación de Ca "" (resolución de escan de 2 ms/línea) en una imagen xí y el curso temporal correspondiente de fluorescencia. E y F muestran estelas (DTMM > 30 ms) registradas en el modo xt. La barra de calibración AF/Fo indica 1 para todos los eventos excepto para el evento de la fila superior del panel C, donde AF/Fo es 2.
La figura 19C y D nnuestra imágenes de AF/Fo de fluorescencia, y el correspondiente
curso temporal de las chispas de Ca^"" registradas en las imágenes de escan de línea
en una fibra de ratón viejo y joven. Los eventos denominados "ascuas", definidos como
aquellos que muestran una DTMM > 30 ms (Chun y cois, 2003) también en se
observaron en fibras de ratones de ambas edades. La figura 19 E y F muestra
imágenes de escan de línea representativas y el curso temporal de la intensidad
72
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
fluorescente de Ca " registrado en el centro del evento, en ratones jóvenes y viejos,
respectivamente.
El número total de eventos (EELC) analizado fue de 455 y 587 en el modo x-y y
64 y 49 en el modo de escan de línea en 14 y 22 fibras de EDL de ratones jóvenes y
viejos, respectivamente. El número tan bajo de eventos caracterizados en el modo de
escan de línea se debe a la escasa ocurrencia de eventos y, por tanto, a la baja
probabilidad de poner la línea de escan a través de un evento antes de que ocurra
(Chun y cois., 2003).
4.4.2. Efecto de los moduladores de RyRs en la frecuencia de las chispas
de Ca^*
Para definir la fuente de los EELC en nuestra preparación y en las condiciones
de nuestros registros, hemos analizado los efectos de agentes farmacológicos bien
caracterizados, como la rianodina, imperatoxina-A y cafeína, en la frecuencia de los
EELC en series xyt. Rianodina e imperatoxina-A inducen estados de subconductancia
prolongada de los RyR (Rousseau y cois, 1987; Tripathy y cois, 1998). La figura 20A
ilustra imágenes de dos fibras musculares de ratón joven (arriba) y viejo (debajo)
mostrando un aumento significativo de la frecuencia de EELC (panel del medio)
después de la aplicación de rianodina 500 nM. El aumento de frecuencia fue de 11, 9 y
13.5 veces con respecto al control en tres fibras de ratones jóvenes y 11 y 4 veces en
dos fibras de ratones viejos. Registramos el aumento máximo en la frecuencia entre 5-
30 segundos después de la aplicación de rianodina (Fig. 2A, b). El típico efecto dual de
la rianodina sobre el RyR se manifestó entre 30-60 s después de la aplicación (fig. 2A,
c), cuando toda la actividad de los RyR fue completamente bloqueada en cada una de
73
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
J o \ e i i
C'oiihol Kiiiuodiitii í5-50s)
B
Rimt04liiiii yiQ-6Qff)
O 2
•i \r/F
Jo\ei i
YUjo
(•'oiiliol Iinpernroxiiiii-A íLOs-4 iniíi)
Jo'veii
Mejo
C'oiifiol Cirfeüiii (5-60fi)
Figura 20. Efecto de la rianodina (500 nM), Imperatoxina-A (20 nM) y cafeína (0.5 mM) en la frecuencia de las chispas de Ca^*. Efecto de 500 nM de rianodina (A), 20 nM de Imperatoxina-A (6) y 0.5 mM de cafeína (C) en la frecuencia de los EELC. Se observó un aumento significativo en la frecuencia de eventos en fibras de ratones jóvenes y viejos con los tres fármacos. La actividad de las chispas desapareció completamente después de 20-40 s de la aplicación de rianodina en todas las células registradas {A.c). Las imágenes representan un área de 45 x 23 ^m.
74
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
las células registradas. La figura 20B Ilustra fibras representativas de un ratón joven y
otro viejo en las que la frecuencia de los EELC aumento en respuesta a la aplicación de
imperatoxina-A 20 nM. El aumento en la frecuencia después de la aplicación de
imperatoxJna-A 20 nM fue de 22, 2 y 3 veces, con respecto al control, en tres fibras de
ratones jóvenes y de 11.5 y 4 en 2 fibras de ratones viejos. Estos datos indican que los
RyRs median la generación de los EELC en nuestra preparación. En algunas otras
células de ambos grupos, joven y viejo, observamos una liberación de Ca^* masiva,
demostrada por el aumento repentino de la fluorescencia global después de la
aplicación de rianodina 500 nM o imperatoxina-A 20 nM. De forma similar, cafeína 0.5
mM aumento la frecuencia de los eventos en 3.6 y 3.5 veces, con respecto al control,
en dos fibras de ratones jóvenes y en 4, 2.3, 3.5 y 10 veces en dos fibras de ratones
viejos. Concentraciones de cafeína de 1 mM o mayores indujeron una aumento de la
fluorescencia global que no permitió la identificación de los EELC en fibras de ratones
jóvenes y viejos.
4.4.3. Propiedades de los EELC identificados en imágenes X-Y o de escan
de línea en fibras de ratones [óvenes y viejos
El análisis de la amplitud y AMM de los ELLC, en el modo x-y y de los eventos
que duraron menos de 30 ms en el modo de escan de línea, están incluidos en la figura
21 y la tabla 9. La figura 21 muestra los histogramas de la distribución de la amplitud y
AMM de todos los eventos registrados en imágenes x-y en fibras de ratones jóvenes y
viejos. La amplitud (media y DS) en fibras de ratones jóvenes fue de 1.2 ± 0.9 y 1.0 ±
0.5 en fibras de ratones viejos (Tabla 9). Se puede observar en la distribución de los
valores de amplitud un rango más amplio en el grupo joven comparado con el grupo
75
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
Tabla IX
Propiedades espaciales de las chispas de Ca ^ identificadas en imágenes XY y
escan de línea
Amplitud AMM (AF/Fo) (Mm)
DMM Tiempo de Velocidad de (ms) ascenso ascenso
10%-Pico (ms"") íms)
N
Imágenes de escan de línea
Joven 1.2 ±0.9 1.8 ±1.2 18 ±7 15±7 0.14±0.11 30
Viejo 1.0 ±0.6 1.6 ±0.7 19 ±6 23 ±10* 0.07 ± 0.04* 19
Amplitud AMM (AF/Fo) (pm)
DMM Tiempo de Velocidad de (ms) ascenso ascenso
10%-Pico (ms"") (ms)
Imágenes XY
N
Joven 1.2 ±0.9 1.6 ±0.8 455
Viejo 1.0 ±0.5* 1.7 ±0.7 587
Eventos de DMM < 30 ms. Los parámetros se han definido en la sección de Material y Métodos. Los valores expresan la media ± DE, siendo N el número de eventos. El asterisco indica diferencias significativamente diferentes (P <0.05).
viejo. El rango de valores fue de 0.20-6.74 AF/Fo y una mediana de 1.0 AF/Fo y de
0.24-3.39 AF/Fo y con una mediana de 0.88 AF/Fo para jóvenes y viejos,
respectivamente. Aunque las fibras de ratones viejos exhibieron un mayor número de
eventos centrados en 0.9 AF/Fo comparado con los de ratones jóvenes, estas últimas
76
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.
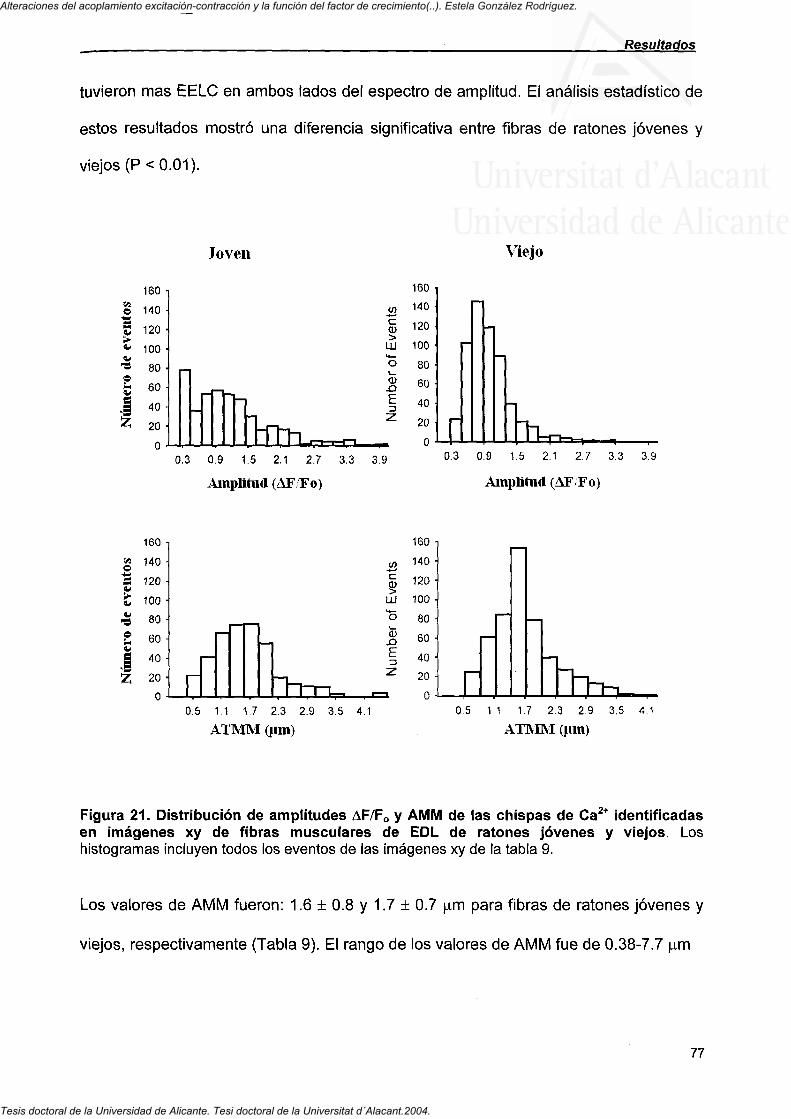
Resultados
tuvieron mas EELC en ambos lados del espectro de amplitud. El análisis estadístico de
estos resultados mostró una diferencia significativa entre fibras de ratones jóvenes y
viejos (P< 0.01).
Joven Viejo
160 1
140
120 •
100 •
80 •
60 -
40
20
O Ittn 0.3 0.9 1.5 2.1 2-7
Amplitud (ATTo)
160 1
c >
uu
E
3.3 3.9
0.5 1.1 1.7 2.3 2.9
ATIMM (íun) 3.5 4.1
IbU •
140
120
100 •
80 •
60 •
40 •
20
T~
r-
r 0.3 0.9
- 1
1 rTT—»—ta. _
1.5 2.1 2 7 3 3 3.9
Amplitud (AT To)
160 1 ___
140 •
120 •
100 T
80 H
60 '
40 -
20 -
—1
"h^ 0.5 11 1.7 2.3 2.9 3.5 4 1
ATM M (pm)
Figura 21. Distribución de amplitudes AF/Fo y AlVIlVI de las chispas de Ca ^ identificadas en imágenes xy de fibras musculares de EDL de ratones jóvenes y viejos. Los histogramas incluyen tocios los eventos de las imágenes xy de la tabla 9.
Los valores de AMM fueron: 1.6 ± 0.8 y 1.7 ± 0.7 jxm para fibras de ratones jóvenes y
viejos, respectivamente (Tabla 9). El rango de los valores de AMM fue de 0.38-7.7 ¡am
77
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
I mediana 1.57 i m, y de 0.54-6.2 nm y mediana 1.60 ^im, para el grupo joven y viejo,
espectivamente. No se detectaron diferencias significativas entres fibras de ratones
jóvenes y viejos en este parámetro.
En la modalidad de escan de línea se registraron 64 y 49 eventos en fibras de
ratones jóvenes y viejos, respectivamente. De estos, 30 y 19 EELC exhibieron una
duración de < 30 ms, en viejos y jóvenes, respectivamente, y se consideraron, por ello,
chispas de Ca " . La amplitud y cinética de los eventos considerados como "ascuas" se
describen en el próximo apartado. La amplitud y la AMM, medidas en los EELC
registrados en el modo de escan de línea, no fueron significativamente diferentes a los
registrados en imágenes x-y (Tabla 9). Aunque la amplitud de las chispas de Ca ""
medidas en el modo escan de línea fue menor en el grupo viejo comparado con el
joven, la diferencia no fue significativa. Esto se debe probablemente al bajo número de
eventos, comparado con los analizados para AF/Fo en imágenes x-y, que no permitió
una potencia suficiente para el análisis estadístico. También se midió la DMM, el
tiempo de ascenso (10%-pico) y la velocidad de ascenso en los eventos de escan de
línea. Los valores de DMM para fibras de ratones jóvenes y viejos fue: 18 ± 7 ms
(rango de 5-28) y 19 + 6 ms (rango de 10-30), respectivamente. El tiempo de ascenso
fue de 15 + 7 (rango 5-28 ms) y 23 ± 10 (7-40 ms), para fibras de ratones jóvenes y
viejos, respectivamente (P < 0.05). Las
medidas de la velocidad de elevación fueron de (en ms"^): 0.14 ± 0.11 (rango 0.04-0.4)
y 0.07 ± 0.04 (rango: 0.03-0.14), respectivamente (Tabla 9), la diferencia entre los dos
grupos en este parámetro también fue significativamente diferente (P < 0.05). En
resumen, las chispas registradas en fibras de EDL de ratones jóvenes y viejos
exhibieron diferencias significativas en amplitud, tiempo de elevación y velocidad de
elevación, pero no en el resto de parámetros.
78
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.
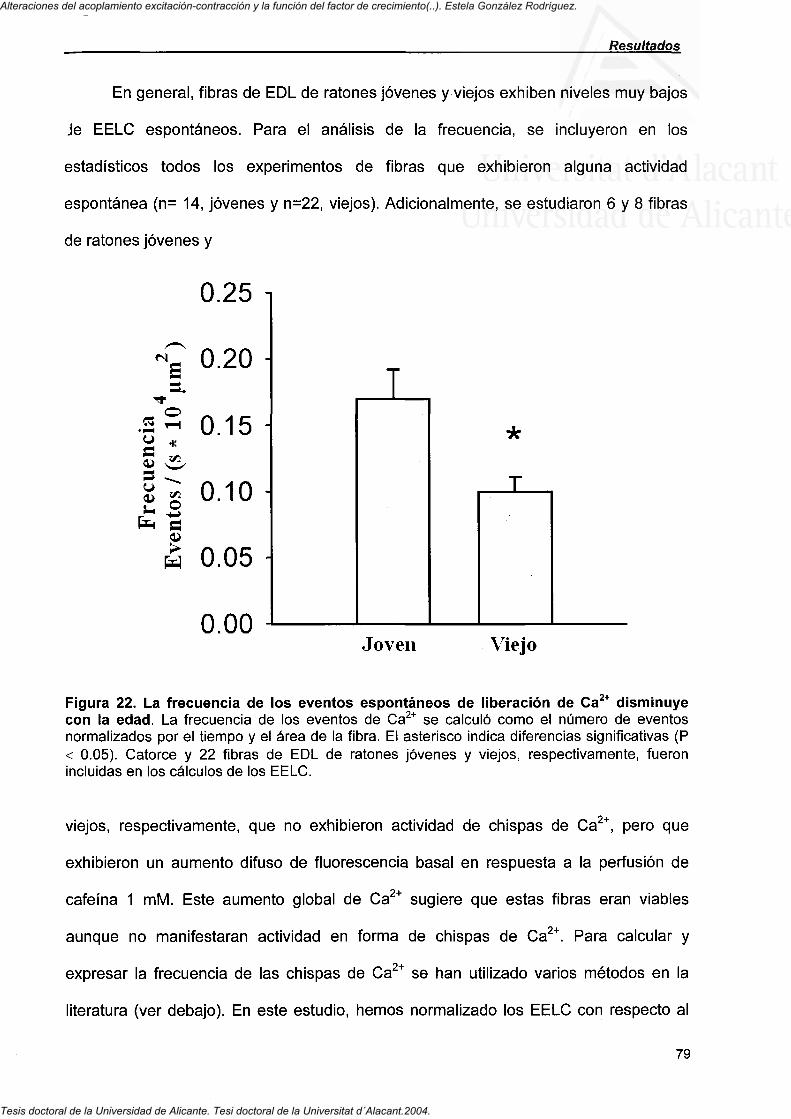
Resultados
En general, fibras de EDL de ratones jóvenes y viejos exliiben niveles muy bajos
Je EELC espontáneos. Para el análisis de la frecuencia, se incluyeron en los
estadísticos todos los experimentos de fibras que exhibieron alguna actividad
espontánea (n= 14, jóvenes y n=22, viejos). Adicionalmente, se estudiaron 6 y 8 fibras
de ratones jóvenes y
0.25 1
-C 0.20 -• I
.5S 0.15 -
r ^ fe SI > fe
0.10 -
0.05 -
0.00 Joven Viejo
Figura 22. La frecuencia de los eventos espontáneos de liberación de Ca ^ disminuye con la edad. La frecuencia de los eventos de Ca * se calculó como el número de eventos normalizados por el tiempo y el área de la fibra. El asterisco indica diferencias significativas (P < 0.05). Catorce y 22 fibras de EDL de ratones jóvenes y viejos, respectivamente, fueron incluidas en los cálculos de los EELC.
viejos, respectivamente, que no exhibieron actividad de chispas de Ca " , pero que
exhibieron un aumento difuso de fluorescencia basal en respuesta a la perfusión de
cafeína 1 mM. Este aumento global de Ca^* sugiere que estas fibras eran viables
aunque no manifestaran actividad en forma de chispas de Ca^*. Para calcular y
expresar la frecuencia de las chispas de Ca "" se han utilizado varios métodos en la
literatura (ver debajo). En este estudio, hemos normalizado los EELC con respecto al
79
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
espacio (área de la fibra, en nm^) y tiempo (s). Como media, se detecto sólo un evento
cada cinco o diez imágenes en el modo de adquisición x-y, en fibras de ratones jóvenes
y viejos, respectivamente. La frecuencia de EELC de todas las fibras de ratones viejos
y jóvenes analizadas fue (media ± DE, ESM): 0.17 + 0.17 (0.02) chispas/(s*10Vm^) y
0.10 ± 0.10 (0.01) (rango: 0.01 a 0.57) para el grupo joven y viejo, respectivamente (ver
debajo). El tiempo total de registro en fibras de ratones jóvenes y viejos fue de 142 y
243 min, respectivamente. La figura 22 muestra la media y el ES de los eventos
registrados en fibras de ratones jóvenes y viejos.
También se calculo la frecuencia como eventos / imagen para comparar
nuestros resultados con publicaciones previas. Registros en fibras de ratones viejos
exhibieron una frecuencia de 0.12 + 0.12 (media ± DE) rango: 0.02-0.73;
aproximadamente 1 chispa/8 imágenes) que es significativamente menor que en fibras
de EDL de ratones de 14 días de edad (P14) (1 chispa / imagen) (Chun y cois., 2003) y
en fibras de ratones jóvenes registradas en este trabajo (0.19 ± 0.19; rango 0.02 -1.14,
aproximadamente 1 chispa/5 imágenes). En conclusión, las fibras de EDL de ratones
viejos exhibieron una frecuencia de EELC menor que la de ratones jóvenes, diferencia
que fue estadísticamente significativa (P < 0.05).
4.4.4. Eventos tipo "ascua" en imágenes de escan de línea en fibras de EDL
de ratones jóvenes v viejos
La figura 19E y F ilustra eventos de tipo "ascua", de duraciones de 148 ms y 83
ms para el grupo joven y viejo, respectivamente. Las "ascuas" constituyeron 53 y 61 %
del número total de eventos registrados en células de ratones jóvenes y viejos,
respectivamente, en imágenes de escan de línea. El análisis de la amplitud, AMM,
80
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
DMMyelt iempo de elevación de estos eventos se presenta en la Tabla 10. La
Tabla X
Propiedades espaciales de los eventos tipo "ascua" de Ca^^ identificados en imágenes de escan de línea
Amplitud ATMM DTMM Tiempo de N ascenso
(AF/Fo) (pm) (ms) 10%-Pico (ms)
Joven 1.04 ±0.59 1.95 ±0.63 83 ± 62 49 ± 66 34
Viejo 0.77 ±0.50* 2.05 ± 0.63 115 ±84 56 ±48 30
Las "ascuas" de Ca " se han definido como aquellos eventos con DTMM > 30 ms. Los parámetros se han definido en Material y Métodos. Los valores se han expresado como la media ± DE, siendo N el número de eventos. Las "ascuas" constituyeron 57 y 63% del número total de eventos registrados en imágenes de escan de línea.
distribución de los parámetros morfológicos fue muy similar para ambos grupos. La
amplitud de las ascuas tuvo un rango de 0.29-2.59 AF/Fo. en células de ratones
jóvenes, y de 0.21-2.15 AF/Fo en células de ratones viejos. El rango para AMM fue de
0.80-3.4 |um y 0.94-3.2 )am en fibras de ratones jóvenes y viejos, respectivamente.
DMM fue de 31 a 312 ms en células del grupo joven y de 35 a 360 ms en células del
grupo viejo. La amplitud media de las chispas registradas en células de ratones viejos
fue significativamente menor que la registrada en ratones jóvenes. Aunque se
observaron eventos de una duración más larga y de velocidad de ascenso menor mas
frecuentemente en fibras de ratones viejos, no se detectaron diferencias significativas
para estos parámetros entre ambos grupos.
81
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
4.4.5. Subtipos de fibras musculares y frecuencia de las chispas de Ca ^
Estudios en varios tipos celulares sugieren que las mitocondrias son importantes
reguladores fisiológicos de las señales de Ca^* intracelular (Babcock y cois., 1997) y
que pueden jugar un papel en el secuestro de Ca^* (Ramesh y cois., 1998; Gunter y
cois, 2004), modificando asi la frecuencia de las chispas de Ca "" (Isaeva y Shirokova,
2003a). Un cambio con la edad en la composición del tipo de fibras musculares en
músculo EDL hacia isoformas más lentas (Messi y Delbono, 2003), que presentan un
mayor contenido de mitocondrias, llevaría a una disminución de la frecuencia de las
chispas de Ca^*. Para determinar si la reducción en la frecuencia y amplitud de las
chispas de Ca " , en fibras de ratones viejos, resulta de cambios en tipos de fibras
musculares se analizó la composición de un subgrupo de fibras del EDL usadas para la
detección de chispas de Ca "". Se analizaron 9 y 10 fibras de ratones jóvenes y viejos,
respectivamente. La figura 23 ilustra la composición de las isoformas de MPC de un
subgrupo de 3 fibras de cada grupo. Los números en las bandas indican la frecuencia
de las chispas expresada como número de chispas/ (s*10'*|im^) específicamente para
cada fibra. Las demás fibras también expresaron solamente la isoforma de CPM lib y
exhibieron una frecuencia de eventos muy dispar, aunque dentro del rango de
frecuencias mostrado en la figura 22. Estos resultados muestran que una diferencia del
doble en la frecuencia de los EELC no está asociada con cambios en subtipos de fibra
muscular. Se desconoce si el contenido de mitocondrias cambia con la edad dentro de
un subtipo de fibra específico.
82
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
0.05 0.14 0.08 0.07 0.04 0.04
m k Á m
lia =^ l lx — lib
V V V
Figura 23. Separación de las isoformas de cadena pesada de miosina (CPIVI) de las fibras de EDL registradas para los eventos de liberación de Ca^*. Gel de electroforesis de poliacrilamida (SDS-PAGE) mostrando las isoformas de CPM en fibras de EDL de ratones jóvenes (J) y viejos (V). Los números indican la frecuencia de las chispas (chispas / s-IO" |jm^) como se describe en la fig. 22. La línea de la derecha es el patrón de referencia.
4.4.6. Función mitocondrial y frecuencia de las chispas de Ca " en fibras
musculares de ratones jóvenes v viejos
Un estudio reciente en fibras de EDL de ratas jóvenes sugiere que la frecuencia
de EELC en células permeabilizadas depende en gran manera del nivel de actividad de
las mitocondrias, ya que la supresión de la actividad mitocondrial aumenta, de forma
reversible, la frecuencia de las chispas de Ca " (Isaeva y Shirokova, 2003b). En el
siguiente grupo de experimentos, hemos analizado si la reducción en la frecuencia y
amplitud de las chispas de Ca^"" registradas en células de ratones viejos es
consecuencia de un aumento de la actividad de las mitocondrias, probablemente
asociado con un aumento en la cantidad de mitocondria s en las fibras musculares con
el envejecimiento. Para ello, hemos usado un desacoplador de mitocondrias, el
carbonil-cianuro de p (trifluorometoxi) fenilhidrazona (FCCP, 25. |am), y oligomicina (2.5
l^m), un inhibidor de la ATP sintasa (Isaeva y cois., 2003a). El efecto de FCCP y
83
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Resultados
oligomicina fue analizado en 10 y 8 células de ratones jóvenes y viejos,
respectivamente. La combinación de FCCP y oligomicina aumentó la frecuencia de las
chispas de Ca^* en 6 y 3 células de las fibras de los grupos jóvenes y viejos,
respectivamente. El efecto de FCCP y oligomicina sobre las chispas de Ca^* fue muy
breve y transitorio, desapareciendo después de 1-4 min de incubación. El aumento en
la frecuencia de las chispas de Ca " fue de 2.4 + 0.42 y 2.6 ± 0.70 veces para fibras de
ratones jóvenes y viejos, respectivamente. No se detectaron diferencias significativas
entre el efecto en fibras de ratones viejos comparados con jóvenes (P < 0.05). Cuatro
de las 10 y 5 de las 8 fibras, de ratones jóvenes y viejos, respectivamente, no
mostraron ningún aumento en la frecuencia de las chispas de Ca " , en repuesta al
envenenamiento de las mitocondrias. Estos resultados indican que la diferencia en la
amplitud y frecuencia de las chispas de Ca "" registrada en este estudio no se explica
debido a cambios en la actividad mitocondrial en fibras de EDL de ratones viejos.
4.4.7. La expresión del receptor de la rianodina tipo 1 (RyRD en músculos
EDL de ratones viejos
El número de RyRI puede influenciar la frecuencia y amplitud de los EELC. Por
ello, el análisis de la expresión de RyR1 en el músculo EDL de ratón es relevante.
Hemos investigado la expresión de esta proteína en músculos de ratones jóvenes y
viejos utilizando inmunoblots. La figura 24 muestra un experimento típico con la banda
específica para RyRI en ausencia de inmunoreactividad para RyR3 en ambos, ratones
jóvenes y viejos. Subsecuentemente, las membranas se incubaron con el anticuerpo
específica anti-SERCAl, el control interno de cargado. El cociente joven/viejo para la
84
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.
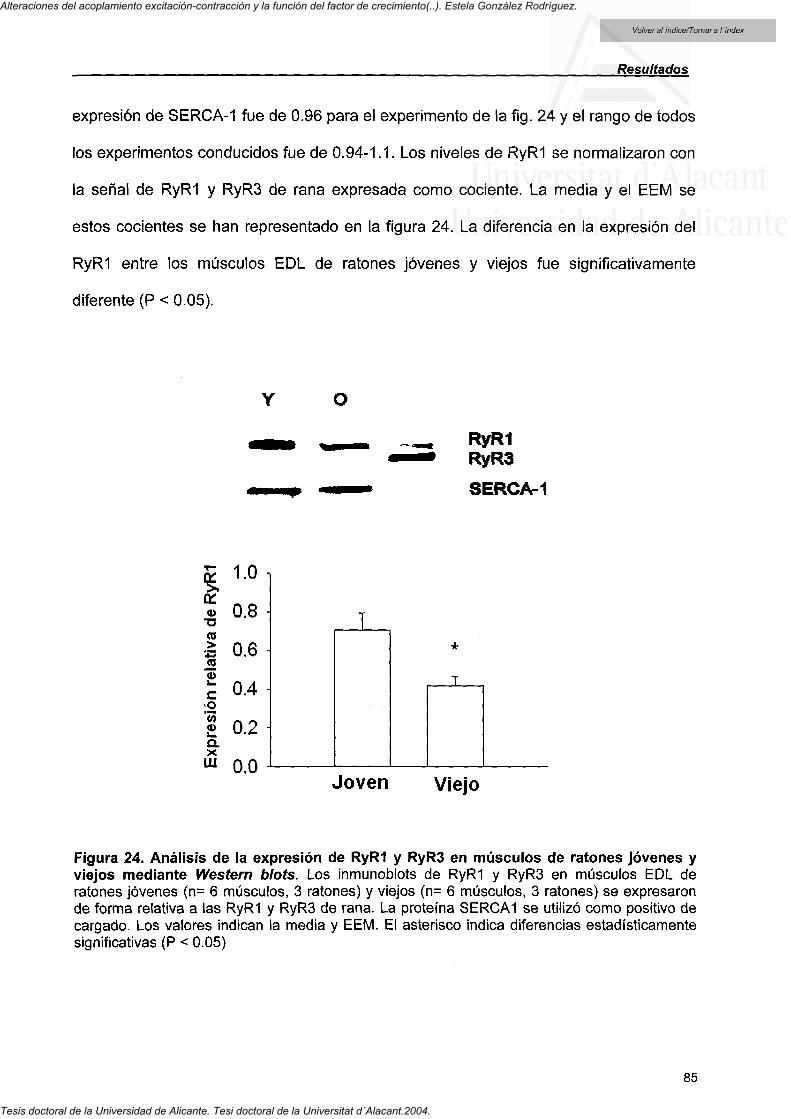
Resultados
expresión de SERCA-1 fue de 0.96 para el experimento de la fig. 24 y el rango de todos
los experimentos conducidos fue de 0.94-1.1. Los niveles de RyRI se normalizaron con
la señal de RyRI y RyR3 de rana expresada como cociente. La media y el EEM se
estos cocientes se han representado en la figura 24. La diferencia en la expresión del
RyRI entre los músculos EDL de ratones jóvenes y viejos fue significativamente
diferente (P < 0.05).
^ RyRI RyR3 SERCA-1
>. OH
(O
>
C O
« i _ Q. X m
1 .\J
0.8
0.6
0.4
0.2
n n .
T *
T
Joven Viejo
Figura 24. Análisis de la expresión de RyRI y RyR3 en músculos de ratones jóvenes y viejos mediante Western blots. Los inmunoblots de RyRI y RyR3 en músculos EDL de ratones jóvenes (n= 6 músculos, 3 ratones) y viejos (n= 6 músculos, 3 ratones) se expresaron de forma relativa a las RyR1 y RyR3 de rana. La proteína SERCA1 se utilizó como positivo de cargado. Los valores Indican la media y EEM. El asterisco indica diferencias estadísticamente significativas (P < 0.05)
85
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

5. Discusión
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

CAPÍTULO I
EFECTO DEL ENVEJECIMIENTO SOBRE LA FUERZA ESPECÍFICA DE CÉLULAS
ÚNICAS E INTACTAS DE LOS MÚSCULOS EXTENSOR COMÚN DE LOS DEDOS
(EDL) Y SOLEO DE RATÓN
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
En el capítulo I de este trabajo se determina, por primera vez, la fuerza
específica generada por fibras de músculo esquelético EDL y soleo y los cambios que
se producen con la edad. El análisis de la fuerza isométrica máxima de contracción nos
permite concluir que la fuerza específica máxima registrada en fibras únicas intactas de
FDB, EDL y soleo de ratones de la misma edad no es significativamente diferente.
También encontramos que la fuerza máxima específica generada por fibras de EDL y
soleo disminuye significativamente con la edad.
5.1.1. La fuerza específica de fibras intactas de los músculos esqueléticos
EDL V soleo de ratón disminuye con la edad
Nuestros resultados indican que la fuerza máxima desarrollada por fibras
intactas de EDL y soleo disminuye con la edad. La fuerza específica registrada en
fibras de los músculos EDL y soleo de ratones de edad joven y mediana es similar a la
registrada en fibras de FDB de ratones jóvenes en este trabajo (418 + 28 kPa) y a la
publicada para fibras de FDB de ratón en la literatura (391 + 59 kPa; rango: 300-480
kPa) (Lannergren y Westerbiad, 1987). La fuerza específica de fibras intactas en este
trabajo es mayor que la registrada en músculo entero o en un pequeño grupo (banda)
de fibras. La discrepancia se podría explicar por la sobrestimación del área de la
sección transversal o por la contribución heterogénea de las fibras musculares al
desarrollo de fuerza en preparaciones multifibrilares. La falta de una contribución
homogénea al desarrollo de fuerza podría deberse a diferencias en la orientación de las
fibras y/o de la longitud óptima (Sugi y Tsuchiya, 1998). Hasta la fecha, no se ha
publicado ningún estudio sobre la fuerza específica de células intactas de músculo
esquelético FDB, EDL o soleo de ratón viejo; por eso, la discusión estará centrada en la
comparación de nuestros resultados con aquellos registrados en preparaciones de
88
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
multifibra publicados en la literatura. La fuerza específica registrada aquí en fibras de
EDL y soleo mostró una disminución significativa en ratones viejos comparados con
ratones jóvenes y de mediana edad. Estos resultados son similares a los publicados
para músculo entero por diversos grupos (Larsson y Edstróm, 1986; Brooks y cois.,
1988). Existen, sin embargo, otros estudios que contradicen nuestros resultados. Por
ejemplo, se ha descrito, utilizando dos técnicas diferentes (bandas de fibras y fibras
peladas-sin membrana externa) que la fuerza específica de soleo es mayor en
animales viejos que en jóvenes, mientras que ninguna diferencia fue observada en
EDL (Eddinger y cois., 1986).
No se encontraron cambios significativos entre la relación fuerza especifica-
frecuencia de estimulación cuando se compararon fibras de EDL de animales de
mediana edad y viejos. Aunque esta diferencia fue significativa para fibras de soleo
debería notarse que existe una tendencia a desarrollar más fuerza a frecuencias
submáximas en fibras de ratones de mediana edad comparado con ratones jóvenes
(Brooks y cois., 1988).
5.1.2. Mecanismos responsables de la disminución de fuerza específica en
músculo esquelético con el envejecimiento
Factores primariamente neurogénicos, miogénicos y otros factores descritos
previamente pueden contribuir a la disminución de fuerza específica con el
envejecimiento (Looser y Delbono, 2003). En este trabajo registramos las alteraciones
en la capacidad de generar fuerza específica en fibras musculares aisladas en las que
la contracción ha sido generada mediante la despolarización del sarcolema o
membrana externa. Nuestro laboratorio ha propuesto que alteraciones en el
mecanismo de EC son responsables, en parte, de la disminución de fuerza específica
89
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
con la edad (Delbono y cois., 1995; Delbono y cois., 1997). Las siguientes evidencias
experimentales apoyan este concepto: i) El número de receptores de DHPR y RyR
disminuye con el envejecimiento en músculo de rata (Renganathan y cois., 1997a) y de
ratón (Renganathan y cois., 1998); ii) la reducción del número de receptores está
asociada con la reducción del movimiento de carga y el pico máximo de Ca ""
intracelular en fibras rápidas de músculo humano (Delbono y cois., 1995) y iii) más
recientemente, con la técnica de fijación de voltaje, hemos demostrado que el
movimiento de carga y el Ca^* intracelular disminuyen en fibras únicas de músculo FDB
de ratones viejos comparado con fibras de ratones jóvenes (Wang y cois., 2000).
Aparte de estos mecanismos, no se descarta que una disminución en la densidad de
proteínas contráctiles de las fibras musculares contribuya a la disminución de fuerza
con el envejecimiento. También se ha propuesto que cambios en tipos de fibra
muscular contribuyen al debilitamiento del músculo con la edad en roedores (Cácela y
cois., 1979) y humanos (Lexell, 1995). Algunos de estos estudios se han llevado a cabo
en voluntarios con varios estados nutricionales y en roedores endogámicos que
muestran una muy baja prevalencia de patologías causadas por el envejecimiento.
Estudios en ratas Fisher 344 y ratones C57BL/6 no muestran cambios en tipos de fibra
asociados al envejecimiento (Florini y cois., 1989; Phillips y cois., 1993). El hecho de
que fibras de los músculos FDB, EDL y soleo de ratones jóvenes y de mediana edad
desarrollen la misma fuerza específica no apoya la posibilidad de que cambios en tipos
de fibra y composición de miosina sean responsables de la disminución de fuerza
específica registrada en este trabajo. No obstante, se ha demostrado que existe una
disminución de fibras de tipo II, la isoforma de miosina de cadena pesada, o un cambio
de fibras de tipo IIB a IIX (Larsson y cois., 1993; Barton-Davis y cois., 1998). Este
fenómeno puede ser relevante para la cinética de contracción pero no en términos de
generación de fuerza máxima.
90
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
La relación fuerza isométrica-pCa medida en fibras peladas no muestra
diferencias significativas entre ratones viejos y jóvenes (Brooks y cois., 1994a), lo cual
sugiere que la sensitividad al Ca^* de las miofibrillas no juega un papel importante en el
envejecimiento. Sin embargo, hay datos que sugieren la existencia de alteraciones con
el envejecimiento a nivel de los puentes cruzados (Phillips y cois., 1993). Estudios en
humanos también muestran inconsistencias en términos de fuerza específica
desarrollada por grupos diferentes de músculos en los ancianos (Hakkinen y cois.,
1996; Kent-Braun y Ng., 1999; Lynch y cois., 1999). La dificultad en la medida del área
de la sección transversal en estos experimentos "in situ" podría explicar las
discrepancias.
5.1.3. Cinética de contracción en fibras de EDL y soleo de ratones jóvenes,
de mediana edad y viejos
La cinética de la contracción única (twitch) registrada en fibras de EDL es similar
a la registrada en células únicas intactas de FDB (Lannergren y Westerbiad, 1987),
después de corregir por el cambio de temperatura. En fibras de FDB, se obtuvieron
valores de 14 y 15 ms como tiempos de contracción y 7-8 y 6-7 ms de tiempos de
relajación media, ambos a 35 C (Lannergren y Westerbiad, 1987). Considerando
valores de Q10 de 3.2 y 4 ms para la contracción y relajación media, respectivamente,
los valores que hemos obtenido en nuestros experimentos están en concordancia con
los publicados en la literatura (Lannergren y Westerbiad, 1987).
No encontramos cambios significativos con la edad en los tiempos de pico y
relajación media de contracción aislada de las fibras de EDL y soleo, a pesar de los
cambios registrados en este parámetro para músculos enteros de soleo y gastrocnemio
(Klitgaard y cois., 1989; Larsson y Salviati, 1989; Narayanan y cois., 1996). Otros
91
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
grupos tampoco detectaron una relajación más lenta en fibras de los músculos EDL,
tibial anterior, soleo y vasto lateral superficial del cuadríceps (Fitts y cois., 1984;
Larsson y Edstróm, 1986; Brooks y cois., 1988). Estas discrepancias se pueden
explicar debido a diferencias entre las preparaciones y condiciones experimentales. A
partir de los resultados obtenidos en estos experimentos podemos concluir: (i) las fibras
únicas e intactas, deprivadas de influencia nerviosa, desarrollan menos fuerza en
edades tardías; (ii) la disminución de la fuerza del músculo entero con la edad puede
ser el resultado de diferentes factores, siendo uno de ellos la alteración de la capacidad
de generar fuerza de las fibras únicas; (iii) alteraciones en la capacidad mecánica de
animales senescentes son consecuencia de una disminución de la fuerza específica
desarrollada por fibras rápidas y lentas; (iv) aunque no hemos examinado los efectos
de la composición muscular durante el envejecimiento, los resultados de fibras únicas
intactas coinciden con muchos de los estudios en músculo entero sugiriendo que las
alteraciones en las mismas fibras contribuyen de forma significativa al cambio
registrado en músculo entero.
92
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

CAPITULO II:
ÍNDICE DE FATIGA EN FIBRAS MUSCULARES DEL EDL Y SOLEO DE RATONES
JÓVENES Y VIEJOS
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
Las principales conclusiones de este estudio son: a) la resistencia isométrica de
fibras intactas de los músculos EDL y soleo no cambia de forma significativa con la
edad; b) el acortamiento del intervalo inter-tetánico produjo más fatiga que con
intervalos largos; sin embargo, no se detectaron cambios con la edad ni en fibras de
EDL ni de soleo, utilizando el mismo protocolo; c) el análisis de la fatiga, basado en dos
criterios diferentes, no mostró diferencia significativas con la edad en fibras de EDL o
soleo; y d) la evolución temporal del índice de fatiga, estudiado en un periodo de 5-10
min, no mostró diferencias en fibras de EDL o soleo disecadas de ratones jóvenes o
viejos.
5.2.1. Fatiga muscular en ratón y otras especies
El índice de fatiga y la evolución temporal de la resistencia isométrica en fibras
musculares de ratón de contracción rápida o lenta no cambiaron significativamente con
la edad. Datos previos obtenidos en músculo entero no han mostrado resultados
consistentes. Mientras que algunos trabajos en mamíferos viejos indican que no hay
cambios o disminuye en la resistencia muscular (Fitts y cois., 1984; Carlsen y Walsh,
1987; McCarter y McGee, 1987; Zhang y Kelsen, 1990; Lawler y cois., 1997), otros
estudios refieren ausencia de cambios (Larsson y Karisson, 1978) o incluso, aumentos
(Petrofsky y Lind, 1975). Los resultados de Págala y cois. (1998) muestran que la fatiga
del músculo entero no cambia o aumenta en EDL y soleo, respectivamente, con la
edad. La discrepancia de los datos en soleo se puede explicar debido al cambio en
tipos de fibras muscular que ocurre con el envejecimiento en el músculo entero.
94
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
5.2.2. Fatiga en fibras de los músculos EDL y soleo
Todas las fibras de los músculos EDL y soleo incluidas en este estudio fueron
rápidas o lentas, respectivamente, de acuerdo con los criterios definidos anteriormente
(ver Material y Métodos).
Los experimentos de fatiga en fibras de EDL y soleo de ratones jóvenes y viejos
mostraron un rango de valores muy amplio. La heterogeneidad de las fibras musculares
(de tipo I, HA, IIB y IIX) podría explicar, al menos en parte, la variabilidad de los
resultados, dado que en este estudio no hemos distinguido los diferentes tipos de fibras
rápidas. Estudios previos realizados en unidades motoras muestran una variabilidad
similar en la resistencia muscular a la fatiga de fibras rápidas (Burke y cois., 1973;
Kadhiresan y cois., 1996).
5.2.3. Mecanismos responsables de la alteración del funcionamiento
neuromuscular con el envejecimiento
Los mecanismos responsables de la disminución en la capacidad motora en
animales viejos no se conocen de forma clara. Se ha sugerido que alteraciones en la
excitabilidad de la vía corticoespinai, en la velocidad de conducción de motoneuronas y
en la eficacia de la transmisión sináptica en la unión neuromuscular o en la
contractilidad del músculo esquelético podrían ser responsables (véase Ross y cois.,
1997; Loeser y Delbono, 2003). La atrofia muscular asociada al desuso es un
condicionante mayor de la disminución de fuerza con la edad, sin embargo, esto no
parece explicar del todo la disminución de la capacidad motora asociada al
envejecimiento en mamíferos (Loeser y Delbono, 2003). Se ha publicado que la fuerza
específica máxima desarrollada por células únicas de EDL y soleo disminuye con la
95
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
edad (González y cois., 2000). Este estudio apoya el concepto de que la disminución
de la fuerza específica con la edad y la fatiga no están relacionados. Los mecanismos
intrínsecos responsables de la disminución de la capacidad generadora de fuerza con
el envejecimiento se conocen sólo parcialmente. Se han mencionado varios factores,
siendo el desacoplamiento de excitación-contracción (DEC) uno de ellos (Delbono y
cois., 1997). DEC ha sido demostrado en humanos y en modelos animales de
envejecimiento (Wang y cois., 2000). No obstante, nuestros resultados no descartan la
posibilidad de que existan, adicionalmente, alteraciones en el mecanismo de
transmisión sináptica que afecten la activación post-sináptica.
Los experimentos de este estudio fueron realizados a temperatura ambiente.
Wineinger y cois. (1998) en un estudio realizado en ratones de dos (joven) y 15
("adultos") meses de edad mostraron que músculos enteros de ratones de la cepa
C10BL/10SnJ "adultos" se fatigan más que los músculos de ratones jóvenes a
temperaturas más altas (35 C comparado con 20 C). La dependencia de la
temperatura en estos cambios es un efecto de la maduración, no del envejecimiento,
en la fatigabilidad muscular. Se desconoce si la temperatura afecta de forma diferente a
la resistencia muscular durante el envejecimiento. Si esto se confirmara, podría indicar
la existencia de diferencias metabólicas entre músculos de ratones jóvenes y viejos.
En este estudio hemos usado dos protocolos de alta frecuencia de estimulación.
La aplicación de bajas frecuencias podría haber ayudado a determinar si existen
cambios de otro tipo. Sin embargo, hemos concentrado este trabajo en si las fibras
musculares rápidas o lentas de ratones jóvenes y viejos desarrollan un nivel de fatiga
diferente aplicando dos protocolos de estimulación previamente utilizados. Queda
pendiente el estudio del efecto de varias frecuencias de estimulación (Edwards y cois.,
1977; Westerbiad y cois., 1993) en diferentes sub-tipos de fibras de mamíferos de
distintas edades, determinando las tensiones generadas y los cambios en la [Ca " ];.
96
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

CAPITULO III:
EL FACTOR DE CRECIMIENTO SIMILAR A LA INSULINA-I PREVIENE LA
DISMINUCIÓN DE CA^* INTRACELULAR Y FUERZA ESPECÍFICA EN FIBRAS
ÚNICAS E INTACTAS DE MÚSCULO DE RATONES TRANSGÉNICOS
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
Los resultados del presente estudio apoyan la hipótesis de que la expresión
transgénica de IGF-1 en músculo esquelético previene la disminución de [Ca^""]; que
ocurre durante la contracción con el envejecimiento y, subsecuentemente, la
disminución de la fuerza específica. El efecto del IGF-1 no esta asociado a un cambio
en el subtipo de fibra del músculo de ratón.
5.3.1. IGF-1 previene la disminución de fuerza específica con el
envejecimiento
En este trabajo hemos investigado el efecto de la expresión transgénica
sostenida de IGF-1 en músculo esquelético sobre la fuerza específica y la [Ca^*]¡ en
fibras únicas intactas. Nuestra hipótesis predecía que la expresión transgénica de IGF-
1 previene la disminución de fuerza específica secundariamente a aumentar el pico de
incremento de [Ca " ]¡ y corrige otras posibles alteraciones (por ejemplo, la disminución
de fuerza por puente cruzado). Los efectos de la expresión transgénica de IGF-1 sobre
la masa de músculo esquelético han sido publicados previamente en ratón y rata. La
expresión de IGF-1 en células previene la disminución de masa y fuerza muscular
asociada al envejecimiento. Este efecto se asume que ocurre mediante un mecanismo
de estimulación de la regeneración del músculo y la preservación de las fibras de tipo
IIB a través de la activación, mediante IGF-1, de células satélite (Barton-Davis y cois.,
1998). Se ha descrito que ratones transgénicos que expresan IGF-1 en músculo
esquelético exhiben hipertrofia muscular; sin embargo, no existe ningún dato publicado
en relación con la fuerza muscular que desarrollan a edades tardías (Musaro y cois.,
2001).
Nuestros resultados muestran que la disminución de fuerza específica que
ocurre con el envejecimiento en ratones FVB (la cepa usada para crear la cepa S1S2-
98
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
IGF-1); González y cois., 2000) se puede prevenir con la expresión transgénica de
hlGF-1, y que esto no parece ser el resultado de un cambio en la composición
muscular del tipo de fibra. Aunque la composición muscular de tipos de fibras fue
analizado en músculo entero, no es probable que las diferencias en los valores de
[Ca^*]j y fuerza específica se deban a cambios en el fenotipo de las fibras. Prueba de
ello es la ausencia de cambios significativos en la velocidad de contracción y relajación
y en la liberación de Ca "" intracelular con la expresión transgénica de IGF-1.
La disminución observada en la sección transversal de la fibra en ausencia de
cambios en el tipo de fibra, sugiere que el cambio de tipo de fibras y/o la denervación
de fibras, observada en músculos rápidos de rata (Larsson y cois., 1991), ocurre más
tarde en el músculo FDB del ratón que en músculos de otras especies. Se debe tener
en cuenta que el músculo FDB es comparativamente más lento que el EDL, debido a la
carencia de fibras de tipo lib (Allen y cols., 1993). El efecto significativo del IGF-1 sobre
el área transversal del músculo es consistente con datos previos (Coleman y cois.,
1995; Musaro y cois., 2001). La atrofia muscular registrada en músculos de animales
viejos control junto con la ausencia de diferencias significativas en el área de la sección
transversal entre ratones jóvenes control y viejos transgénicos sugiere que IGF-1
preserva el trofismo muscular y favorece la regeneración en todas las edades,
probablemente a través de un efecto en la activación de las células satélite (Barton-
Davis y cois., 1998). En nuestro estudio, el fenómeno de apoptosis no parece ser muy
marcado dado que el número de fibras por músculo fue similar en los cuatro grupos
considerados.
En este trabajo hemos observado una disminución en la fuerza específica del
músculo FDB del 32 % con el envejecimiento. Este valor es similar al registrado en
músculo FDB de rata (Carlsen y Waisfi, 1987) y EDL de ratón (Brooks y cois., 1988). El
grado de disminución de la fuerza específica obtenido en este trabajo es consistente
99
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
con estudios previos realizados en fibras únicas e intactas de los músculos EDL y
soleo, que mostraron disminuciones del 26 % y 20 %, respectivamente (González y
cois., 2000). No encontramos diferencias significativas en la fuerza específica de
músculo entero de ratones jóvenes control y ratones viejos transgénicos. Estos
resultados están en acuerdo con los registrados previamente en ratones en los que se
expreso IGF-1 por medio de un adenovirus (Barton-Davis y cols., 1998), o mediante un
mecanismo transgénico (Renganathan y cois., 1998), y sugieren que altas
concentraciones de IGF-1 en el músculo mejora el desarrollo de fuerza de los puentes
cruzados además de incrementar la masa muscular.
5.3.2. Mecanismos responsables de la disminución de fuerza específica de
fibras musculares con el envejecimiento
Se ha propuesto que una alteración en el mecanismo de activación del
sarcolema y la liberación de Ca " del RS es la causa de la disminución de fuerza
específica asociada al envejecimiento (Delbono y cois., 1995; Delbono, y cois., 1997).
Para examinar esta hipótesis hemos investigado si las alteraciones en la elevación de
la [Ca ""]; inducida por estimulación eléctrica produce una reducción de fuerza
específica en fibras del músculo FDB de ratones viejos. El valor del pico de [Ca^^ji
registrado en fibras de FDB de ratones jóvenes control en respuesta a una estimulación
breve está en el rango previamente descrito (Westerbiad y Alien, 1991; Carrol! y cois.,
1995), aunque es algo menor que el referido por algunos grupos (Bakker y cois.,
1997b). La ausencia de diferencias significativas entre ratones jóvenes control y
transgénicos es consistente con los experimentos de fijación de voltaje en células de
FDB en los que la [Ca^ ji fue medida con un indicador de Ca " de afinidad relativamente
baja (Wang y cois., 2002). La [Ca^^ji máxima registrada en fibras de ratones viejos fue
100
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
significativamente menor que la registrada en fibras de ratones jóvenes. La disminución
del pico intracelular de Ca^* (~ 34%) es similar a la disminución de fuerza específica en
fibras únicas (37%) y músculo entero (32%). Esta disminución fue menor que la
registrada anteriormente en células FDB con fijación de voltaje (~48%) (Wang y cois.,
2000). Para determinar si la alteración en la liberación de Ca " es responsable del
déficit de fuerza, se estudio el efecto de la cafeína sobre la fuerza máxima de fibras
musculares. La potenciación inducida por cafeína en la fuerza máxima de células
únicas es consistente con el aumento de la liberación de Ca^* inducido por cafeína en
células musculares registradas con la técnica de fijación de voltaje (Delbono, y cois.,
1995). Se podría argüir que el tétano no representa la condición de activación máxima
de la fibra, ya que cafeína aumenta la [Ca^ ]¡ y la fuerza específica en células intactas
del músculo FDB de ratón (Alien y Westerbiad, 1995). De acuerdo con esta evidencia
experimental, sería de esperar un mayor aumento en la [Ca ""]; después de alcanzar la
máxima liberación de Ca "* del RS evocada por estimulación eléctrica. Los
experimentos con cafeína en fibras de ratones jóvenes y viejos control y transgénicos
apoyan la conclusión de que existe una cantidad extra de Ca "" en el retículo
sarcoplasmático que está disponible en respuesta a estimulación eléctrica máxima.
Esto favorece la idea de que el agotamiento de Ca "" del retículo sarcoplasmático no
explica la disminución de la liberación de Ca "" que ocurre en fibras de ratones viejos.
Por el contrario, la disminución del número de DHPR y RyRI (Renganathan y cois.,
1997a), el aumento de RyRs desacoplados a DHPRs (Renganathan y cois., 1998), y la
posible alteración de la probabilidad de apertura del RyR asociada a alteraciones en la
regulación del canal (Damiani y cois., 1996), son algunas de las explicaciones que
justifican que fibras de ratones viejos no utilicen la cantidad de Ca "" extra para
compensar la disminución de la liberación de Ca "" dependiente de voltaje. Aunque el
valor máximo de la [Ca^*]¡ intracelular se recupera en su totalidad después de la
101
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
aplicación de cafeína, todavía persiste un déficit en la fuerza específica en células de
ratones viejos. Por esto, la existencia de otros mecanismos que contribuyan a la
disminución de la fuerza con el envejecimiento deben ser considerados. Alteraciones
de la maquinaria contráctil pueden contribuir a la disminución de fuerza específica con
la edad. La disminución de la fuerza específica de fibras tipo I, Na y llb/llx de músculos
de rata y humano ha sido previamente publicada (Li y Larsson, 1996; Larsson y cois.,
1997; Lowe y cois., 2002). La observación de que células peladas de humanos de edad
avanzada activadas de forma máxima y células de mamíferos pequeños desarrollan
menos fuerza específica que células musculares de individuos jóvenes sugiere que la
capacidad intrínseca del aparato miofibrilar está alterada con el envejecimiento,
además de las alteraciones en el acoplamiento de excitación-contracción. Ambos, la
activación submáxima del aparato contráctil y la pérdida de capacidad intrínseca para
desarrollar fuerza podrían cooperar en la determinación de la pérdida de fuerza con el
envejecimiento.
5.3.3. Mecanismos del efecto de IGF-1 en ratones IGF-1-transqénicos
La expresión local sostenida de IGF-1 previene la disminución del pico de la
[Ca^^ji durante la contracción y la fuerza específica que ocurre con el envejecimiento en
células de FDB de ratones S1S2. Esto sugiere que IGF-1 previene alteraciones en el
mecanismo de acoplamiento de excitación-contracción, además de en otros posibles
mecanismos (como la disminución de fuerza de los puentes cruzados) involucrados en
la disminución de fuerza con el envejecimiento. El pico de la [Ca^*]¡ registrado en fibras
de ratones transgénicos fue significativamente mayor que el registrado en fibras
ratones viejos control, pero similar al de fibras de ratones jóvenes control. IGF-1
aumenta la expresión de la subunidad a lS del DHPR (DHPRais) en cultivo primario de
102
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
músculo (Renganathan y cois., 1998; Wang y cois., 1999) y en músculos de ratones
adultos jóvenes y viejos, en concordancia con los datos previos sobreexpresando
durante toda la vida IGF-1 en músculo (10-15 veces mayor que los correspondientes
controles) (Renganathan y cois., 1998). El aumento de las subunidades a l del DHPR
es la consecuencia de un aumento de la trascripción genética (Zheng y cois., 2002a).
El mecanismo de acción de IGF-1 en el aumento del tamaño fibrilar parece involucrar la
acción de la calcineurina y el factor GATA (Musaro y cois., 1999; Semsarian y cois.,
1999). Estudios realizados en nuestro laboratorio han demostrado que el mecanismo
de la expresión de DHPRais es complejo (Zheng y cois., 2002a y b). Por esto, los
mecanismos por los cuales IGF-1 actúa para causar hipertrofia muscular y el control de
las moléculas involucradas en la dinámica de la [Ca^'*']\ no son completamente
idénticos. Se desconoce todavía si la isoforma muscular de IGF-1 (Owino y cois., 2001)
también contribuye en la prevención de la disminución de la expresión de estos canales
y la fuerza específica con la edad.
103
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

CAPITULO IV:
EVENTOS ELEMENTALES DE LIBERACIÓN DE Ca^* CHISPAS Y ASCUAS EN
FIBRAS DE MÚSCULO ESQUELÉTICO DE RATONES VIEJOS
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
El hecho de que músculos de animales senescentes no alcance los niveles
intracelulares de Ca^* registrados en células de músculos de ratones jóvenes en
células registradas con la técnica de voltage-clamp (Wang y cois, 2000) y en células
contráctiles (González y cois, 2003) nos ha llevado a analizar la generación de EELC
espontáneos en fibras musculares de animales jóvenes y viejos. En este trabajo,
hemos encontrado una disminución en la amplitud y la frecuencia de los EELC en
células de ratones viejos comparadas con jóvenes. Estas alteraciones no parecen estar
asociadas a cambios de la función mitocondrial o el tipo de célula. Además, la
expresión de RyR1 disminuye en músculo EDL de ratón viejo.
5.4.1. EELC en músculo esquelético de ratón adultos
El análisis farmacológico inicial nos permitió determinar que la fuente de los
EELC registrados en este trabajo es la permeabilidad de Ca " a través del RyR1 en
fibras de ratones jóvenes y viejos. Tres agentes que han demostrado tener un efecto
específico sobre el RyR1 han sido probados (rianodina, imperatoxina-A y cafeína).
Estos tres compuestos aumentaron la frecuencia de los EELC.
Subsecuentemente, comparamos nuestros resultados con los registrados en
otros estudios de músculo esquelético de adulto (Conkiin y cois., 1999a; Kirsch y cois.,
2001; Uttenweiler y cois, 2002) o embriónico/neonatal (Conkiin y cois., 1999b; Kirsch y
cois, 2001; Chun y cois., 2003; Isaeva y Shirokova., 2003a; Zhou y cois, 2003 y
Szentesi y cois, 2004). Hasta ahora, que nosotros sepamos, no se han realizado
estudios de EELC en músculo esquelético de mamíferos viejos. Además, aunque las
propiedades funcionales de los RyRs de mamíferos jóvenes se han estudiado de forma
extensiva en canales reconstituidos en membranas lipídicas artificiales (Fill y Copello,
2002; Meissner, 2002), no existe información, en este respecto, de músculo esquelético
105
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
de mamíferos viejos.
La amplitud, AMIVl, DMM y tiempo de ascenso de las chispas registradas aquí
son similares a los registrados en fibras intactas de FDB (Conckiin y cois., 1999a) y
fibras permeabilizadas de EDL (Kirsch y cois., 2001) de ratón. El tiempo de ascenso
más corto descrito para fibras de EDL de ratón neonato (P14) (Chun y cois, 2003),
comparado con los valores registrados en este trabajo, se podría explicar por la
existencia de la isoforma RyR3 en esta última preparación, ya que esta isoforma de
RyR no se ha detectado en ratón joven-adulto o viejo. La frecuencia, en nuestra
preparación, es similar a la registrada por Chun y cois (2003) en fibras durante el
periodo postnatal (1 chispa/ 5 imágenes). Sin embargo, hay algunas discrepancias con
otras publicaciones. Se han registrado 1-4 chispas / imagen o 455 eventos registrados
en 5 fibras permeabilizadas (Kirsch y col., 2001) o 220 eventos en 26 células intactas
(Conkiin y cois., 1999b). La frecuencia de las chispas en el presente estudio (0.19 ±
0.19 eventos / imagen; rango 0.02-1.14) es similar a la registrada en el periodo
postnatal temprano (Chun y cois., 2003) pero menor que en fibras de adulto (Kirsch y
col, 2001). Diferencias en los procedimientos empleados para la detección de los EELC
podrían explicar, al menos en parte, estas discrepancias. Además, en este estudio,
varias células no exhibieron ninguna actividad de EELC aunque se registró un aumento
global de fluorescencia con la aplicación de 1 mM de cafeína. Desafortunadamente,
estos resultados no se pueden comparar con los de otros grupos porque no se ha
publicado previamente el número de células viables funcionales que no exhiben EELC.
Las "ascuas", definidas como aquellos eventos de duración mayor de 30 ms,
constituyeron un 53 % de todos los eventos de escan de línea; si usamos el criterio de
50 ms, las "ascuas" representan el 38 %, valor que es similar al 33 % registrado en
experimentos previos (Kirsch y cois., 2001). Diferencias en la detección y análisis de
los eventos podría explicar estar diferencia ya que los procedimientos experimentales.
106
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
incluida la composición de la solución interna, son similares.
Los experimentos realizados en fibras de músculo esquelético de rata son más
numerosos y por tanto la caracterización de las propiedades de los EELC es más
completa. La amplitud media de las chispas en este estudio es similar a (Shirokova y
cois., 2003; Conkiin y cois., 1999a; Szentesi y cois., 2004) o más pequeña que (Kirsch
y cois., 2001; Uttenweiler y cois., 2002) la registrada en publicaciones previas, pero en
general dentro del rango de los valores de la literatura. Nuestros valores de AMM, DMM
y tiempo de ascenso son similares a los valores más pequeños registrados en rata; sin
embargo, existe una gran dispersión en los valores dados para estos parámetros
(Uttenweiler y cois., 2002; Zhou y cois., 2003; Csernoch y cois., 2004 y Szentesi y cois.,
2004), fenómeno que podría resultar de diferencias en las condiciones de registro. Si la
frecuencia se calculase en número de eventos / fibra, el valor registrado en este trabajo
es significativamente menor que el registrado en rata. Podría ser que la regulación del
DHPR en la liberación de Ca^* del RyR esté controlada de una forma más estricta en
ratón que en rata, lo cual podría llevar a una probabilidad más baja en la ocurrencia de
los eventos de Ca "". Sin embargo, estudios sobre la relación DHPR / RyR que registran
valores de 0.91 (Renganathan y col, 1998) y 0.95 (Renganathan y col, 1997a) en
músculo EDL de ratón y rata, respectivamente, apuntan en contra de esta posibilidad.
5.4.2. Efectos del envejecimiento en las propiedades de los EELC
El análisis de las amplitudes de los EELC de fibras de ratones jóvenes y viejos
reveló diferencias significativas en la liberación de Ca "". Las fibras de ratón viejo,
comparadas con las de ratones jóvenes, exhibieron un mayor número de eventos más
pequeños. El amplio rango de amplitudes indica que las chispas de Ca^^ son
polimórficas, como sucede en células de corazón (Shen y cois., 2004), más que
107
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
estereotipadas o de fenómeno todo-o-nada. La disminución con la edad del RNAm
(Zheng y cois, 2002b) y la proteína (registrada aquí) en músculo EDL de ratón, sugiere
la existencia de alteraciones en la regulación del RyR1 (Damiani y cois., 1996). Estas
alteraciones podrían justificar las diferencias en la liberación de Ca "" mediante la
influencia en el número de unidades de liberación y en la probabilidad de apertura del
canal. Aunque la similitud del valor DMM encontrada entre ambos grupos contradice
esta posibilidad, el aumento en el tiempo de ascenso y la disminución de la frecuencia
de los EELC en fibras de ratones viejos es consistente con esta hipótesis.
Las diferencias con la edad en la frecuencia de los ELLC registrada en este
trabajo no se justifican por alteraciones en la regulación del Ca "", Mg "" o ATP del
RyR1, basado en las características de la preparación (célula permeabilizada o
químicamente pelada) y el ambiente controlado de iones y de nucleotides. En la
literatura hay datos que indican que existe un efecto del Ca^* luminal en la frecuencia y
tiempo de apertura de RyR2 en células de corazón (Lukyanenko y cois, 1996).
Además, se ha descrito que un lazo del RyRI del lumen esta involucrado en la
actividad del canal en músculo esquelético (Gao y cois, 2000). Se desconoce si el
contenido de Ca " del RS cambia con la edad. Información con respecto a este punto
podría aportar más información relevante sobre los mecanismos de regulación del
RyRI por el Ca "" del RS. En este estudio se muestra evidencia directa de que la
expresión del RyR1 disminuye con la edad. Sin embargo, se carece de datos
referentes a cambios en la regulación del RyRI; si bien los que existen son algo
controversiales. Nuestra hipótesis es que cambios en el nivel de expresión y/o función
de ciertas proteínas, presumiblemente involucradas en el proceso de acoplamiento de
EC, resultan en alteraciones en las propiedades de apertura del RyRI con el
envejecimiento. El estado metabólico y de oxidación-reducción del RyRI puede
modular la actividad (Sun y cois., 2003) y frecuencia de las chispas de Ca^* en ratas
108
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
adultas-jóvenes. Sin embargo, el efecto en fibras musculares de mamíferos viejos se
desconoce. La interacción con calmodulina (Smith y cois. 1989), FKBP12, y/o
FKBP12.6 (Qi y cois, 1998) juega un papel relevante en la función de canal único, sin
embargo, no existe ninguna información sobre músculos de mamíferos viejos sobre
estos temas. La diferencia en la frecuencia de EELC con la edad no esta asociada a
cambios en la isoforma de los RyR de acuerdo con la ausencia de RyR3 en los análisis
de inmunoblot de músculos EDL de ratón joven y viejo que presentamos aquí.
La contribución de un menor número de RyRI en la generación de EELC se
reflejaría en la dimensión espacial de las chispas (AMM), pero esta variable no es
estadísticamente significativa entre fibras de ratones jóvenes y viejos. Se desconoce si
la reducción del número de RyRI ocurre de forma homogénea o en forma de "parche"
(no homogénea). Supuestamente, un déficit más homogéneo tendría influencias en la
amplitud y AMM. Un déficit en forma de "parche" podría tener un efecto más parcial en
la morfometría de los EELC permitiendo la ocurrencia de ciertos números de eventos
normales. Esto resultaría en un rango más amplio en la distribución de valores de AMM
en fibras de ratones viejos, fenómeno que no es aparente en el histograma de
frecuencias de AMM. Sobre la base de estos resultados, proponemos que un déficit de
forma homogéneo, durante la edad avanzada, determina una menor amplitud de los
EELC. La velocidad de ascenso de las chispas de Ca "", (parámetro que indica el flujo
de liberación de Ca ""), que se correlaciona con la amplitud de la chispa (Shen y cois.,
2004), fue significativamente diferente entre grupos, consistente con el descubrimiento
de que un menor número de RyRI contribuye a una menor amplitud de EELC en
animales viejos. Una correlación negativa entre el flujo de liberación de Ca "" (velocidad
de ascenso) y la duración de la liberación de Ca^* (tiempo de ascenso) se ha descrito
en células de corazón (r= -0.43), lo que da soporte a la idea de que una corriente de
liberación más fuerte termina la chispa más temprano (Shen y cois, 2004). El análisis
109
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
Je la correlación de estas dos variables en este trabajo esta de acuerdo con estudios
previos y provee una correlación negativa para fibras de ambos grupos: joven (r =-0.53)
y viejo (r= -0.50). Estos datos indican la preservación del mecanismos de terminación
de liberación de Ca " de RS en fibras de ratones viejos.
Aunque las ascuas no se han generado mediante la despolarización de la
membrana en este estudio, el examen de los EELC en el modo escan de línea nos
permite concluir que la amplitud de estos eventos es significativamente menor en fibras
de ratones viejos comparados con jóvenes. El estudio de la activación por voltaje de los
EELC permitirá determinar de una forma más apropiada si alteraciones en la función
del RyR1 o la interacción DHPR-RyR1 contribuyen a la disminución del pico global de
la liberación de Ca "" que ocurre con el envejecimiento (Wang y cois., 2000; González y
cois, 2003). La disminución con la edad del cociente DHPR/RyR1 descrita previamente
(Renganathan cois., 1997; Renganathan y cois., 1998), podría dejar un mayor número
de RyRI desacoplados de los DHPR y por ello sin contribuir a la generación de
"ascuas" Aparentemente, las fibras de rata responden a la despolarización del
sarcolema con ascuas, y la suma de estos eventos permite la reconstrucción de la
señal global de fluorescencia de Ca " , jugando así un papel crítico en el acoplamiento
de excitación-contracción (Csernoch y cois., 2004). El aumento de RyRs desacoplados
en ratones viejos haría que el acoplamiento EC sea más dependiente de la liberación
de Ca^* inducida por Ca " calcio (mecanismo conocido como CICR), la cual resultaría
en un aumento de la frecuencia de las chispas de Ca ""; sin embargo, el efecto de la
disminución en la expresión de los RyRs podría contrarrestar e incluso sobrepasar este
efecto en músculos de ratones viejos.
La homogeneidad de la población de células de ratones jóvenes y viejos
utilizados en este trabajo contradice aparentemente la noción de que el número de
fibras tipo lib disminuye con el envejecimiento (Larsson y cois., 1991; Messi y Delbono,
110
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Discusión
2003). Más del 80 % de las fibras de EDL de ratón son tipo lib y la disminución
registrada con el envejecimiento es aproximadamente del 20% (Messi y Delbono,
2003). Para nuestros experimentos hemos disociado fibras de la superficie del músculo
de forma sistemática, y como las fibras lib están distribuidas homogéneamente en todo
el músculo, aunque predominantemente localizadas en la periferia (datos no
mostrados), la probabilidad de disecar fibras de tipo lib en músculos de ratones viejos
es bastante alta.
Las fibras del músculo esquelético son ricas en mitocondrias y se ha demostrado
que estas organelas están localizadas en cercana proximidad al retículo
sarcoplasmático (Ogata y Yamasaki, 1985), actuando presumiblemente como
sumideros de Ca "" en el citoplasma cuando hay exceso de Ca^* (Ramesh y cois., 1998;
Gunter y cois., 2004). Estos estudios sugieren que cambios en la actividad de las
mitocondrias podrían causar cambios en la frecuencia y/o propiedades de los EELC.
Con el envejecimiento la función mitocondrial disminuye o no cambia (Trounce y cois.,
1989; Boffoli y cois., 1994; Rasmussen y cois., 2003). Con el objetivo de identificar los
mecanismos responsables de la disminución de la frecuencia de los EELC, estudiamos
la función de las mitocondrias en fibras de ratones jóvenes y viejos. Nuestros
experimentos de inhibición de la función mitocondrial, junto con la detección de una
variación considerable en la frecuencia de las chispas, dentro de un mismo tipo de
subtipo de fibra, no sugieren un papel relevante de las mitocondrias en las alteraciones
de la generación de los EELC y sus propiedades durante el envejecimiento.
111
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

6- Conclusiones
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Conclusiones
6. CONCLUSIONES
1) La fuerza específica de células únicas e intactas de los músculos
extensor común de los dedos (EDL) y soleo de ratón disminuye con el
envejecimiento
2) La fuerza tetánica específica máxima desarrollada por fibras únicas
intactas de EDL (músculo rápido) y de soleo (músculo lento) no es
significativamente diferente.
3) El estudio de la relación frecuencia de estímulo-fuerza específica de
fibras únicas de ratones de diferentes edades no fue significativamente
diferente para ninguno de los dos músculos (EDL y soleo).
4) El análisis de los parámetros de la cinética de contracción considerados
(tiempo al pico y tiempo medio de relajación) no cambiaron significativamente
con la edad en los músculos EDL o soleo.
5) No se observaron diferencias significativas con la edad en el índice de
fatiga de las fibras de EDL y soleo.
6) El índice de fatiga no mostró correlación alguna con la fuerza específica
registrada en fibras de EDL o soleo.
7) El curso temporal del índice de fatiga fue similar en células de ratones de
diferentes edades tanto en el músculo EDL como en el soleo.
8) La disminución de la liberación de Ca ' del retículo sarcoplasmático está
asociada a la disminución de fuerza específica que ocurre con el
envejecimiento.
9) Un agotamiento de las reservas de Ca "" del retículo sarcoplasmático no
explica la disminución de la liberación de Ca "" que ocurre en fibras de ratones
viejos.
113
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Conclusiones
10) El IGF-1 previene la disminución de fuerza específica con el
envejecimiento.
11) Las fibras de ratón viejo, comparadas con las de ratones jóvenes,
muestran un menor número de eventos elementales de liberación de Ca "" y,
además, de menor amplitud.
12) La reducción de la frecuencia y la amplitud de los eventos elementales
de liberación de Ca "" (EELC) con la edad está asociada a una disminución de
la expresión del receptor de la rianodina tipo 1 (RyRI).
13) Las alteraciones de los EELC de fibras de ratones viejos no parecen ser
mediadas por cambios en la función mitocondrial o en el tipo celular.
114
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

7. Bibliografía
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Bibliografía
BIBLIOGRAFÍA
Allen DG, Duty S y Westerbiad H (1993). Metabolic changes in muscle during exercise; their effects on muscle function. Proc Aust Physiol Pharmacol Soc 24, 65-75.
Allen DG y Westerbiad H (1995). The effects of caffeine on intracellular calcium, force and the rate of relaxation of mouse skeletal muscle. J Physiol (Lond) 487, 331-342.
Ashley CC, Mulligan IP y Lea TJ (1991). Ca ^ and activation mechanisms in skeletal muscle. Quat Rev Biophys 24, 1-73.
Babcock DF, Herrington J, Goodwin PC, Park YB y Hille B (1997). Mitochondrial participation in the intracellular Ca^^ network. J Cell Biol 136, 833-844.
Bakker AC, Joosten LA, Arntz OJ, Helsen MM, Bendele AM, Van de Loo, FA y Van den Berg WB (1997a). Prevention of murine collagen-induced arthritis in the knee and ipsilateral paw by local expression of human interleukin-1 receptor antagonist protein in the knee. Arth and Rheum 40, 893-900.
Bakker AJ, Head SI y Stephenson DG (1997b). Time course of calcium transients derived from Fura-2 fluorescence measurements in single fast twitch fibres of adult mice and rat myotubes developing in primary culture. Cell Calcium 21, 359-364.
Barton-Davis ER, Shoturma Dl, Musaro A, Rosenthal N y Sweeney HL (1998). Viral mediated expression of insulin-like growth factor I blocks the aging-related loss of skeletal muslce function. Proc Natl Acad Sc/95,15603-15607.
Beam KG y Franzini-Armstrong C (1997). Functional and structural approaches to the study of excitation-contraction coupling. Meth Cell Biol. 52, 283-306.
Boffoli D, Scacco SC, Vergari R, Solarino G, Santacroce G y Papa S (1994). Decline with age of the respiratory chain activity in human skeletal muscle. Biochim Biophys >Acfa 1226, 73-82.
Brooks SV y Faulkner JA (1988). Contractile properties of skeletal muscles from young, adult and aged mice. J Physiol 404, 71-82.
Brooks SV y Faulkner JA (1990). Contraction-induced injury: recovery of skeletal muscles in young and old mice. Am J Physiol 258, C436-442.
Brooks SV y Faulkner JA (1994a). Isometric, shortening, and lengthening contractions of muscle fiber segments from adult and old mice. Am J Physiol 267, C507-513.
Brooks SV y Faulkner JA (1994b) Skeletal muscle weakness in old age: underlying mechanisms. Med Sci Sports Exerc 26, 432-439.
Burke RE, Levine DN, Tsairis P y Zajac FEI (1973). Physiological types and histochemical profiles in motor units of the cat gastrocnemius. J Physiol 234, 723-748.
Caccia M R, Harris J B y Johnson M A (1979). Morphology and physiology of skeletal muscle in aging rodents. Muscle Nen/e 2, 202-221.
116
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Bibliografía
Carlsen RC y Walsh DA (1987). Decrease in force production and appearance of alpha-adrenergic mediated contracture in aging rat sl^eletal muscle. Pflugers Arch 408, 224-230.
Carroll SL, Klein MG y Schneider MF (1995). Calcium transients in intact rat skeletal muscle fibers in agarose gel. Am J Physiol 269, C28-34.
Cheng H, Lederer WJ, y Cannell MB (1993). Calcium sparks: elementary events underlying excitation-contraction coupling in heart muscle. Science 262, 740-744.
Cheng H, Song LS, Shirokova N, Gonzalez A, Lakatta EG, Rios E y Stern MD (1999). Amplitude distribution of calcium sparks in confocal images: theory and studies with an automatic detection method. Biophys J 76, 606-617.
Chun LG, Ward CW y Schneider MF (2003). Ca "" sparks are initiated by Ca^^ entry in embryonic mouse skeletal muscle and decrease in frequency postnatally. Am J Physiol Cell Physiol 285, C686-697.
Coleman ME, DeMayo F, Yin KC, Lee HM, Geske R, Montgomery C y Schwartz RJ (1995). Myogenic vector expression of insulin-like growth factor I stimulates muscle cell differentiation and myofiber hypertrophy in transgenic mice. J Biol. Chem 270, 12109-12116.
Conklin MW, Barone V, Sorrentino V y Coronado R (1999a). Contribution of ryanodine receptor type 3 to Ca "" sparks in embryonic mouse skeletal muscle. Biophys J 77, 1394-1403.
Conklin, MW, Powers, P, Gregg, RG y Coronado, R (1999b). Ca "" sparks in embryonic mouse skeletal muscle selectively deficient in dihydropyridine receptor alphalS or betala subunits. Biophys J 76, 657-669.
Csernoch L, Zhou J, Stern MD, Brum G y Rios E (2004). The elementary events of Ca "" release elicited by membrane depolarization in mammalian muscle. J Physiol 557, 43-48.
Damiani E, Larsson L y Margareth A (1996). Age-related abnormalities in regulation of the ryanodine receptor in rat fast-twitch muscle. Cell Calcium 19, 15-27.
Decary S, Mouly V, Hamida CB, Sautet A, Barbet JP y Butler-Browne GS (1997). Replicative potential and telomere length in human skeletal muscle: implications for satellite cell-mediated gene therapy. Hum Gene TherQ, 1429-1438.
Delbono O (2002). Molecular mechanisms and therapeutics of the deficit in specific force in ageing skeletal muscle. Biogerontology 3,265-270.
Delbono O (2003). Neural control of aging skeletal muscle. Aging Cell 2, 21-29.
Delbono O, O'Rourke KS y Ettinger WH (1995). Excitation-calcium release uncoupling in aged single human skeletal muscle fibers. J Memb Biol 148,211-222.
117
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Bibliografía
Delbono O, Renganathan M y Messi ML (1997). Excitation-Ca^* release-contraction coupling in single aged human skeletal muscle fiber. Muscle Nerve 5, S88-92.
Eddinger TJ, Cassens RG y Moss RL (1986). Mechanical and histochemical characterization of skeletal muscles from senescent rats. Am J Physiol 251, C421-430.
Edwards RHT, Hill DK, Jones DA y Merton PA (1977). Fatigue of long duration in human skeletal muscle after exercise. J Physiol (Lond) 272, 769-778.
Ferrington DA, Jones TE, Qin Z, Miller-Schyler M, Squier TC y Bigelow DJ (1997). Decrease conformational stability of the sarcoplasmic reticulum Ca-ATPasa in aged skeletal muscle. Biochim BiophysActa 1330, 233-247.
Fill M y Copello JA (2002). Ryanodine receptor calcium release channels. Physiol Rev 82, 893-922.
Fitts RH, Troup JP, Witzmann FA y Holloszy JO (1984). The effect of aging and exercise on skeletal muscle function. Mech Age Devil, 161-172.
Florin JR y Ewton DZ (1989). Skeletal muscle fiber types and myosin ATPase activity do not change with age or growth hormone administration. J Gerontol Biol Sci 44, B110-B117.
Florini JR, Ewton DZ, Magri KA y Mangiacapra FJ (1993). IGFs and muscle differentiation. [Review]. Adv Exp Med Biol 343, 319-326.
Florini JR, Ewton DZ y Coolican SA (1996). Growth hormone and insulin growth factor system in myogenesis. [Review] Endocrine 17,481-517.
Frontera WR, Suh D, Krivickas LS, Hughes VA, Goldstein R y Roubenoff R (2000). Skeletal muscle fiber quality in older men and women. Am J Physiol Cell Physiol. 279, C611-618.
Gao L, Balshaw D, Xu L, Tripathy A, Xin C y Meissner G (2000). Evidence for a role of the lumenal M3-M4 loop in skeletal muscle Ca " release channel (ryanodine receptor) activity and conductance. Biophys J79, 828-840. Giulian GG, Moss RL y Greaser M (1983). Improved methodology for analysis and quantitation of proteins on one dimensional silver-stained slab gels. Anal Biochem 129, 277-287.
González E, Messi ML y Delbono O (2000). The specific force of single intact mouse extensor digitorum longus (EDL) and soleus mouse muscle fibers declines with aging. J MembrBiolMQ, 175-183.
González E, Messi ML, Zheng Z y Delbono O (2003). Insulin-like growth factor-1 prevents age-related decrease in specific force and intracellular Ca^* in single intact muscle fibres from transgenic mice. J Physiol 552, 833-844.
Greenlund LJ y Nair KS (2003). Sarcopenia-consequences, mechanisms, and potential therapies. Mech Ageing Dev 124,287-299.
118
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Bibliografía
Gunter TE, Yule DI, Gunter KK, Ellseev RA y Salter JD (2004). Calcium and mitochondria. FEBS Lett 567, 96-102.
Hakkinen K, Kraemer WJ, Kallinen M, Linnamo V, Pastinen UM y Newton RU (1996). Bilateral and unilateral neuromuscular function and muscle cross-sectional area in middle-aged and elderly men and women. J Gerontol Biol Sci 51, B21-29.
Hoopes LL (2002). Error catastrophe in mutant mitochondria. Sci Aging Knowledge Environ. 2002, 6.
Isaeva EV y Shirokova N (2003a). Metabolic regulation of Ca "" release in permeabilized mammalian skeletal muscle fibres. J Physiol (Lond) 547, 453-462.
Isaeva EV y Shirokova N (2003b). Comparison of local calcium signals in slow and fast twitch mammalian muscle fibers. Biopiiys Soc Annual Meeting Abs, 1881-Pos.
Jaggar JH, Porter VA, Lederer WJ y Nelson MT (2000). Calcium sparks in smooth muscle. Am J Physiol Cell Physiol 278, C235-256.
Kadhiresan, V.A., Hassett, C.A. y Faulkner, J.A., 1996. Properties of single motor units in medial gastrocnemius muscles of adult y old rats. J Physiol 493, 543-552
Kent-Braun JA y Ng AV (1999). Specific strength and voluntary muscle activation in young and elderly women and men. J AppI Physiol 87, 22-29.
Kirsch WG, Uttenweiler D y Fink RH (2001). Spark- and ember-like elementary Ca "" release events in skinned fibres of adult mammalian skeletal muscle. J Physiol 537, 379-389.
Klein MG, Cheng H, Santana LF, Jiang YH, Lederer WJ y Schneider MF (1996). Two mechanisms of quantized calcium release in skeletal muscle. Nature 379, 455-458.
Klitgaard H, Marc R, Brunet A, Vyewalle H y Monod H (1989). Contractile properties of old rat muscles: effect of increased use. J AppI Physiol 67,1401-1418.
Lamberts SW, Van den Beld AW y Van der Leiy AJ (1997). The endocrinology of aging. Sc/ence 278, 419-424.
Lannergren J y Westerblad H (1987). The temperature dependence of isometric contractions of single, intact fibres dissected from a mouse foot muscle. J Physiol 390, 285-293.
Lannergren J y Westerblad H (1991). Force decline due to fatigue and intracellular acidification in isolated fibers from mouse skeletal muscle. J Physiol 434, 307-322.
Larsson L y Karlsson J (1978). Isometric and dynamic endurance as a function of age and skeletal muscle charactehstics. Acta Physiol. Scy. 104, 129-136.
Larsson L y Edstróm L (1986). Effects of age on enzyme-histochemical fiber spectra and contractile properties of fast- and slow-twitch skeletal muscles in the rat. J Neurol Sc/76, 69-89.
119
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Bibliografía
Larsson, L y Salviati, G. (1989). Effects of age on calcium transport activity of sarcoplasmic reticulum in fast- and slow-twitch rat muscle fibers. J Physiol 419, 253-264.
Larsson L, Ansved T, Edstrom L, Gorza L y Schiaffino S (1991). Effects of age on physiological, immunohistochemical and biochemical properties of fast-twitch single motor units in the rat. J Physiol (Lond) 443, 257-275.
Larsson L, Biral D, Campione M y Schiaffino S (1993). An age-related type MB to MX myosin heavy chain switching in rat skeletal muscle. Acta Physiol Scy. 147, 227-234.
Larsson L, Li X y Frontera WR (1997). Effects of aging on shortening velocity and myosin isoform composition in single human skeletal muscle cells. Am J Physiol 272, C638-649.
Lawler JM, Cline, CC, Hu Z y Coast JR (1997). Effect of oxidant challenge on contractile function of the aging rat diaphragm. Am J Physiol 272, E201-E207.
Leeson CR y Leeson TS (1976). Histology.Ed: WB Syers.
Lexell J (1995). Human aging, muscle mass, and fiber type composition. J Gerontol Biol Sci 50, 11-16.
Li X y Larsson L (1996). Maximum shortening velocity and myosin isoforms in single muscle fibers from young and old rats. Am J Physiol 270, C352-360.
Looser RF y Delbono O (2003). The musculoskeletal and joint system. In Principles of Geriatric Medicine and Gerontology. Eds: Hazzard WR, Blass JP, Halter JB, Ouslyer JG yTineti, ME. McGraw-Hill, New York. 1097-1111.
Lowe DA, Surek JT, Thomas DD y Thompson LV (2001). Electron paramagnetic resonance reveals age-related myosin structural changes in rat skeletal muscle fibers. Am J Physiol Cell Physiol 280, C540-547.
Lowe DA, Thomas DD y Thompson LV (2002). Force generation, but not myosin ATPase activity, declines with age in rat muscle fibers. Am J Physiol Cell Physiol 283, C187-192.
Lukyanenko V, Gyorke I y Gyorke S (1996). Regulation of calcium release by calcium inside the sarcoplasmic reticulum in ventricular myocytes. Pflugers Arch 432, 1047-1054.
Lynch NA, Metter EJ, Lindle RS, Fozard JL, Tobin JD, Roy TA, Fleg JL y Hurley BF (1999). Muscle quality I. Age-associated differences between arm and leg muscle groups. J AppI Physiol 86, 188-194.
Margreth A, Damiani E y Bortoloso E (1999). Sarcoplasmic reticulum in aged skeletal muscle. Acta Physiol Scy 167, 331-338.
McCarter R y McGee J (1987). Influence of nutrition and aging on the composition and function of rat skeletal muscle. J Gerontol 42, 432-441.
120
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Bibliografía
Meissner G (2002). Regulation of mammalian ryanodine receptors. Front Biosci 7, 2072-2080.
Melzer W, Herrmann-Frank A y Luttgau HC (1995). The role of Ca ^* ions in excitation-contraction coupling of skeletal muscle fibres. Biochim Biophys Acta 1241, 59-116.
Messi ML y Delbono O (2003). Target-derived trophic effect on skeletal muscle innervation in senescent mice. J A/eurosc/23,1351-1359.
Musaro A, McCullagh KJA, Naya FJ, Olson EN y Rosenthal N (1999). IGF-1 induces skeletal myocyte hypertrophy through calcineurin in association with GATA-2 y NF-ATcl. Nature 400, 581-585.
Musaro A, McCullagh KJ, Paul A, Houghton L, Dobrowolny G, Molinaro M, Barton-Davis ER, Sweeney HL y Rosenthal N (2001). Localized IGF-1 transgene expression sustains hypertrophy and regeneration in senescent skeletal muscle. Nature Genetics 27,195-200.
Narayanan N, Jones DL, Xu A y Yu JC (1996). Effects of aging on sarcoplasmic reticulum function and contraction duration in skeletal muscles of the rat. Am J Physiol 271,01032-1040.
Ogata T y YamasakI Y (1985). Scanning electron-microscopic studies on the three-dimensional structure of mitochondria in the mammalian red, white and intermediate muscle fibers. Cell Tissue Res 241, 251-256.
Owino W, Yang SY y Goldspink G (2001). Age-related loss of skeletal muscle function and the inability to express the autocrine form of insulin-like growth factor-1 (MGF) in response to mechanical overload. FEBS Lett 505, 259-263.
Págala MK, Ravindran K, Namba T, Grob D (1998). Skeletal muscle fatigue and physical endurance of young and old mice. Muscle Nerve 21, 1729-1739.
Payne AM, Gonzalez E, Zheng Z, Wang ZM, Messi ML y Delbono O (2004). Cardiaclike excitation-contraction coupling in aging skeletal muscle. Exp Biol Ann Meeting 850, 3.
Petrofsky JS y Lind AR (1975). Aging, isometric strength and endurance, and cardiovascular responses to static effort. J AppI Physiol 38, 91-95.
Pette D y Staron RS (2001). Transitions of muscle fiber phenotypic profiles. Histochem Ce//8/0/115, 359-372.
Phillips SK, Wiseman RW, Woledge RC y Kushmerick MJ (1993). Neither changes in phosphorus metabolite levels nor myosin isoforms can explain the weakness in aged mouse muscle. J Physiol 463, 157-167.
Qi Y, Ogunbunmi EM, Freund EA, Timerman AP y Fleischer S (1998). FK-binding protein is associated with the ryanodine receptor of skeletal muscle in vertebrate animals. J Biol Chem 273, 34813-34819.
121
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Bibliografía
Ramesh V, Sharma VK, Sheu SS y Franzini-Armstrong C (1998). Structural proximity of mitochondria to calcium release units in rat ventricular myocardium may suggest a role in Ca^* sequestration. Ann N Y Acad Sci 853, 341-344.
Rasmussen UF, Krustrup P, Kjaer M y Rasmussen HN (2003). Experimental evidence against the mitochondrial theory of aging. A study of isolated human skeletal muscle mitochondria. Exp Gerontol 38, 877-886.
Renault V, Rollan E, Thornell LE, Mouly V y Butler-Browne G (2002). Distribution of satellite cells in the human vastus lateralis muscle during aging. Exp Gerontol 37, 1513-1514.
Renganathan M, Messi ML y Delbono O (1997a). Dihydropyridine receptor-ryanodine receptor uncoupling in aged skeletal muscle. J Memb Biol 157, 247-253.
Renganathan M, Messi ML, Schwartz R y Delbono O (1997b). Overexpression of hlGF-1 exclusively in skeletal muscle increases the number of dihydropyridine receptors in adult transgenic mice. FEBS Left 417, 13-16.
Renganathan M, Messi ML y Delbono O (1998). Overexpression of IGF-1 exclusively in skeletal muscle prevents age-related decline in the number of dihydropyridine receptors. J Biol Chem 273, 28845-28851.
Rennie MJ, Wackerhage H, Spangenburg EE y Booth FW (2004). Control of the size of the human muscle mass. Annu Rev Physiol 66, 799-828.
Rogers MA y Evans WJ (1993). Changes in skeletal muscle with aging: effects of exercise training. Exerc Sport Sci Rev 21, 65-102.
Roos MR, Rice CL y Vyervoort AA (1997). Age-related changes in motor unit function. Muscle Nerve 20, 679-690.
Rousseau E, Smith JS y Meissner G (1987). Ryanodine modifies conductance and gating behavior of single Ca "" release channel. Am J Physiol 253, 364-368.
Ryan M, Carison BM y Ohiendieck K (2000). Oligomeric status of the dihydropyridine receptor in aged skeletal muscle. Mol Cell Biol Res Commun 4, 224-229.
Schneider MF y ChandLer WK (1973). Voltage dependent charge movement of skeletal muscle: a possible step in excitation-contraction coupling. Nature 242, 244-246.
Schneider MF y Ward CW (2002). Initiation and termination of calcium sparks in skeletal muscle. Frani 6/osc/ 7,212-222.
Semsarian C, Wu MJ, Ju YK, Marciniec T, Yeoh T, Allen DG, Harvey RP y Graham RM (1999). Skeletal muscle hypertrophy is mediated by a Ca^"'-dependent calcineurin signaling pathway. Nature 400, 576-581.
Shen JX, Wang S, Song LS, Han T y Cheng H (2004). Polymorphism of Ca^* sparks evoked from in-focus Ca release units in cardiac myocytes. Blophys J 86, 182-90.
122
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Bibliografía
Shirokova N, García J y Rios E (1998). Local calcium release in mammalian skeletal musc\e. J Physiol 5^2, 377-384.
Smith JS, Rousseau E y Meissner G (1989). Calmodulin modulation of single sarcoplasmic reticulum Ca -release channels from cardiac and skeletal muscle. Circ Res 64, 352-359.
Sugi H y Tsuchiya T (1998). Muscle mechanics I: Intact single muscle fibers. In Current Methods in muscle physiology. Advantages, Problems and Limitations. Oxford University Press, New York, 3-31.
Sun J, Xu L, Eu JP, Stamler JS y Meissner G (2003). Nitric oxide, NOC-12, and S-nitrosoglutathione modulate the skeletal muscle calcium release channel/ryanodine receptor by different mechanisms. An allosteric function for O2 in S-nitrosylation of the channel. J Biol Chem 278, 8184-8189.
Szentesi P, Szappanos H, Szegedi C, Gonczi M, Jona I, Cseri J, Kovacs L y Csernoch L (2004). Altered elementary calcium release events and enhanced calcium release by thymol in rat skeletal muscle. Biopliys J 86, 1436-1453.
Thompson LV y Brown M (1999). Age-related changes in contractile properties of single skeletal fibers from the soleus muscle. J AppI Physiol 86, 881-886.
Tripathy A, Resch W, Xu L, Valdivia HH y Meissner G (1998). Imperatoxin A induces subconductance states in Ca^* release channels (ryanodine receptors) of cardiac and skeletal muscle. J Gen Physiol 111, 679-690.
Trounce I, Byrne E y Marzuki S (1989). Decline in skeletal muscle mitochondrial respiratory chain function: possible factor in ageing. Lancet 1, 637-639.
Tsien RW y Pozzan T (1989). Measurement of cytosolic free Ca "" with quin2: practical aspects. Methods Enzymol 172, 230-262.
Tsugorka A, Rios E y Blatter LA (1995). Imaging elementary events of calcium release in skeletal muscle cells. Science 269,1723-1726.
Uttenweiler D, Kirsch WG, Schulzke E, Both M y Fink RH (2002). Model-based analysis of elementary Ca^* release events in skinned mammalian skeletal muscle fibres. Eur BiophysJ 31, 331-340.
Wang ZM, Messi ML y Delbono O (1999a). Patch-clamp recording of charge movement, Ca^* current and Ca transients in adult skeletal muscle fibers. Biophys J 77, 2709-2716.
Wang ZM, Messi ML, Renganathan M y Delbono O (1999b). Insulin-like growth factor-1 enhances rat skeletal muscle L-type Ca "" channel function by activating gene expression. J Physiol (Lond) 516, 331-341.
Wang ZM, Messi ML y Delbono O (2000). L-type Ca^* channel charge movement y intracellular Ca^* in skeletal muscle fillers from aging mice. Biophys J 78, 1947-1954.
123
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.

Bibliografía
Wang ZM, Messi ML y Delbono O (2002). Sustained overexpression of IGF-1 prevents age-dependent decrease in cliarge movement and intracellular calcium in mouse skeletal muscle. B/op/7ys J 82,1338-1344.
Ward CW, Reiken S, Marks AR, Marty I, Vassort G y Lacampagne A (2003). Defects in ryanodine receptor calcium release in skeletal muscle from post-myocardial infarct rats. FasebJM, 1517-1519.
Weindruch R (1995). Interventions based on the possibility that oxidative stress contributes to sarcopenia. J Gerontol A Biol Sci Med Sci 50, 157-161.
Westerblad H y Allen DG (1991). Changes of myoplasmic calcium concentration during fatigue in single mouse muscle fibers. J Gen Physiol 98, 615-635.
Westerblad H, Duty S, Allen DG (1993). Intracellular calcium concentration during low-frequency fatigue in isolated single fibers of mouse skeletal muscle. J AppI Physiol 75 (1), 382-388.
Wineinger MA, Walsh SA y Abresch RT (1998). The effect of age and temperature on MDX muscle fatigue. Muscle Nerve 21, 1075-1077.
Zhang Y y Kelsen SG (1990). Effects of aging on diaphragm contractile function in golden hamsters. Am Rev Respir Dis 142, 1396-1401.
Zheng Z, Wang ZM y Delbono O (2001a). Age-dependent IGF-1 regulation of gene transcription of Ca "" channels in skeletal muscle. Mech Aging Dev 122, 373-384.
Zheng Z, Wang ZM y Delbono O (2001b). IGF-1 increases transcriptional activity of the skeletal muscle DHPR alpha-1 subunit by acting on CREB element of the promoter region. In 31st Annual Meeting, ed. Neurosci S F 519.11.
Zheng Z, Wang ZM y Delbono O (2002a). Insulin-like growth factor-1 increases skeletal muscle DHPR alpha IS transcriptional activity by acting on the cAMP-response element-binding protein element of the promoter region. J Biol Chem 277, 50535-50542.
Zheng Z, Wang ZM y Delbono O (2002b). Charge movement y transcription regulation of L-type calcium channel alpha-1 S in skeletal muscle cells. J Physiol (Lond) 540, 397-409.
Zheng Z, Wang ZM y Delbono O (2004). Ca "" calmodulin kinase y calcineurin mediate IGF-1-induced skeletal muscle dihydropyridine receptor alpha(IS) transcription. J MembrBionQJ, 101-112.
Zhou J, Brum G, Gonzalez A, Launikonis BS, Stern MD y Rios E (2003). Ca "" sparks and embers of mammalian muscle. Properties of the sources. J Gen Physiol 122, 95-114.
124
Alteraciones del acoplamiento excitación-contracción y la función del factor de crecimiento(..). Estela González Rodríguez.
Tesis doctoral de la Universidad de Alicante. Tesi doctoral de la Universitat d´Alacant.2004.




















