
Canine ACL fibroblast integrin expression and cell alignment in response to cyclic tensile strain in...
10
Canine ACL Fibroblast Integrin Expression and Cell Alignment in Response to Cyclic Tensile Strain in Three-Dimensional Collagen Gels D. Ross Henshaw, Erik Attia, Madhu Bhargava, Jo A. Hannafin Laboratory for Soft Tissue Research, Sports Medicine and Shoulder Service, The Hospital for Special Surgery, 535 East 70th Street, New York, New York 10021 Received 14 January 2005; accepted 3 August 2005 Published online 1 February 2006 in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/jor.20050 ABSTRACT: Tissue-engineered ligament substitutes have the potential to become an alternative graft source for ligament reconstruction. If this approach is to become viable, one must first understand and define the mechanisms responsible for creation, maintenance, and remodeling of the native anterior cruciate ligament. It is well accepted that mechanical load alters fibroblast phenotypic expression in a variety of cell sources; however, the mechanosensitive pathways responsible for alteration in matrix production, remodeling, and alignment are unknown. We hypothesize that cell surface integrins play a role in this mechanotransduction process, and as such respond to application of cyclic tensile load. Linear 3D collagen gels containing canine ACL fibroblasts were created in Flexercell Tissue-Train Culture Plates. Gels were untethered (control), tethered without external strain (tethered), or tethered and exposed to 2.5% cyclic strain for 2 h per day for 4 days (strain). Quantitation of a1, a5, and b1 integrin subunit was performed using flow cytometry. Cell and matrix alignment was studied using light, polarized light, and fluorescent microscopy. Expression of a5 and b1 integrin subunits was increased significantly in fibroblasts in tethered and strained 3D collagen gels compared with the control, unloaded constructs ( p < 0.05). These integrins are known to function as mechanotransducers in other tissues, implicating a similar role in mechanotransduction in ACL fibroblasts. Histologic analysis of the tethered and strained gels demonstrated a linear arrangement of cells and parallel collagen fibril architecture. In contrast, cell distribution and collagen alignment were disorganized in the control, unloaded gels. The alignment of cells and collagen in the 3D gels parallel to applied strain is similar to the in vivo state. These data add to our understanding of the behavior of ACL fibroblasts in vitro. The ability to manipulate signal transduction pathways may enhance our ability to engineer implantable ACL grafts or to modify ACL healing response. ß 2006 Orthopaedic Research Society. Published by Wiley Periodicals, Inc. J Orthop Res 24:481–490, 2006 Keywords: ACL; tissue engineering; strain; integrin; fibroblast; canine INTRODUCTION There are over 100,000 anterior cruciate ligament (ACL) reconstructions performed annually in the United States with clinical success rates reported from 75–97%. 1 The most common sources of auto- genous grafts are central third patellar tendon, hamstring tendon, and quadriceps tendon grafts. The morbidity associated with autografts includes anterior knee pain, patella fracture, and residual hamstring weakness. 2,3 The use of allograft tissues avoids these complications; however, there is an associated risk of disease transmission, graft rejection, and limited availability. 4 Tissue-engi- neered ligament replacements have the potential to provide an alternative graft source that could be readily available, subject to quality control, and could avoid the morbidity of the current graft options. Construction of viable and biomechani- cally equivalent ligament replacements requires a fundamental understanding of ACL biology including fibroblast matrix synthesis and remo- deling in response to the local mechanical envir- onment. JOURNAL OF ORTHOPAEDIC RESEARCH MARCH 2006 481 Correspondence to: J. Hannafin (Telephone: 212-606-1419, Fax: 212-327-1417; E-mail: hannafi[email protected]) ß 2006 Orthopaedic Research Society. Published by Wiley Periodicals, Inc.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Canine ACL fibroblast integrin expression and cell alignment in response to cyclic tensile strain in...

Canine ACL Fibroblast Integrin Expression and CellAlignment in Response to Cyclic Tensile Strain inThree-Dimensional Collagen Gels
D. Ross Henshaw, Erik Attia, Madhu Bhargava, Jo A. Hannafin
Laboratory for Soft Tissue Research, Sports Medicine and Shoulder Service, The Hospital for Special Surgery,535 East 70th Street, New York, New York 10021
Received 14 January 2005; accepted 3 August 2005
Published online 1 February 2006 in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/jor.20050
ABSTRACT: Tissue-engineered ligament substitutes have the potential to become an alternativegraft source for ligament reconstruction. If this approach is to become viable, one must firstunderstandanddefine themechanisms responsible for creation,maintenance, and remodelingof thenative anterior cruciate ligament. It is well accepted that mechanical load alters fibroblastphenotypic expression in a variety of cell sources; however, the mechanosensitive pathwaysresponsible for alteration in matrix production, remodeling, and alignment are unknown. Wehypothesize that cell surface integrins play a role in this mechanotransduction process, and as suchrespond to application of cyclic tensile load. Linear 3D collagen gels containing canine ACLfibroblasts were created in Flexercell Tissue-Train Culture Plates. Gels were untethered (control),tethered without external strain (tethered), or tethered and exposed to 2.5% cyclic strain for 2 h perday for 4 days (strain). Quantitation of a1, a5, and b1 integrin subunit was performed using flowcytometry. Cell and matrix alignment was studied using light, polarized light, and fluorescentmicroscopy. Expression of a5 and b1 integrin subunits was increased significantly in fibroblasts intethered and strained 3D collagen gels compared with the control, unloaded constructs (p<0.05).These integrins are known to function asmechanotransducers in other tissues, implicating a similarrole inmechanotransduction inACLfibroblasts.Histologic analysis of the tetheredand strained gelsdemonstrated a linear arrangement of cells and parallel collagen fibril architecture. In contrast, celldistribution and collagen alignment were disorganized in the control, unloaded gels. The alignmentof cells and collagen in the 3D gels parallel to applied strain is similar to the in vivo state. These dataadd to our understanding of the behavior of ACLfibroblasts in vitro. The ability tomanipulate signaltransduction pathways may enhance our ability to engineer implantable ACL grafts or to modifyACL healing response. � 2006 Orthopaedic Research Society. Published by Wiley Periodicals, Inc.
J Orthop Res 24:481–490, 2006
Keywords: ACL; tissue engineering; strain; integrin; fibroblast; canine
INTRODUCTION
There are over 100,000 anterior cruciate ligament(ACL) reconstructions performed annually in theUnited States with clinical success rates reportedfrom 75–97%.1 The most common sources of auto-genous grafts are central third patellar tendon,hamstring tendon, and quadriceps tendon grafts.Themorbidity associated with autografts includesanterior knee pain, patella fracture, and residual
hamstring weakness.2,3 The use of allograft tissuesavoids these complications; however, there isan associated risk of disease transmission, graftrejection, and limited availability.4 Tissue-engi-neered ligament replacements have the potentialto provide an alternative graft source that couldbe readily available, subject to quality control,and could avoid the morbidity of the current graftoptions. Construction of viable and biomechani-cally equivalent ligament replacements requiresa fundamental understanding of ACL biologyincluding fibroblast matrix synthesis and remo-deling in response to the local mechanical envir-onment.
JOURNAL OF ORTHOPAEDIC RESEARCH MARCH 2006 481
Correspondence to: J. Hannafin (Telephone: 212-606-1419,Fax: 212-327-1417; E-mail: [email protected])
� 2006 Orthopaedic Research Society. Published by Wiley Periodicals,Inc.

Ligament structure, function, response toinjury, and development depend on mechanicalstimuli. With daily activities, the ACL is subjectedto varying amounts of tensile strain, which iscrucial for ligament homeostasis.5–8 Clinically,the effect of load deprivation on ligament structurecan be seen following rupture of the human ACL.Midsubstance tears do not heal, and the majorityof the ligaments involute with retraction of thetorn ends. This phenomenon results from contrac-tion of the ligament stump by intrinsic myofibro-blasts,9 coupled with matrix metalloproteinasedigestion of the ligament matrix.10,11
Several studies have shown that collagen andmatrix degrading enzyme production are regu-lated in response to mechanical load.12–15 CanineACL fibroblasts increase collagen production anddecrease matrix degrading enzyme activity whensubjected to tensile strain, while stress depriva-tion results in decreased collagen synthesis andupregulation of MMPs.16 We hypothesize thattensile strain plays a homeostatic role in main-taining ligament fibroblast function.
Cell binding to extracellular matrix moleculesincluding laminin, collagen, and fibronectin ismediated by cell surface integrins. Followingattachment to the extracellular matrix, cell signal-ing occurs via transmembrane integrin dimerslinking the extracellular and intracellular environ-ment.17–20b1 and a5 subunits are present on rabbitACL and MCL fibroblasts, with increased expres-sion of these integrins in response to ACLinjury.21,22 The expression of these same integrinsubunits, a5 and b1, increases significantly whenexposed to tensile strain in monolayer culture.23,24
Although the resultswere significant, tensile strainapplication was performed in a monolayer culturesystem, which produced a nonuniform strain field.In response to the limitations of this system, webegan investigating the use of an in vitro linearthree-dimensional (3D) strain model, which per-mits application of a uniform strain across a 3Dcollagen gel. Using a 3D model allows histologicevaluation of fibroblast and collagen alignment.Wehypothesize that ACL cell surface integrins willincrease in response to cyclic tensile strain whencultured in a 3D gel matrix. We also hypothesizethat cell and collagen alignment will respond tostrainwithcreationofa linear, organizedstructure.
MATERIALS AND METHODS
Cells were obtained by explant culture or collagenase(Worthington Biochemical Corporation, Lakewood, NJ)
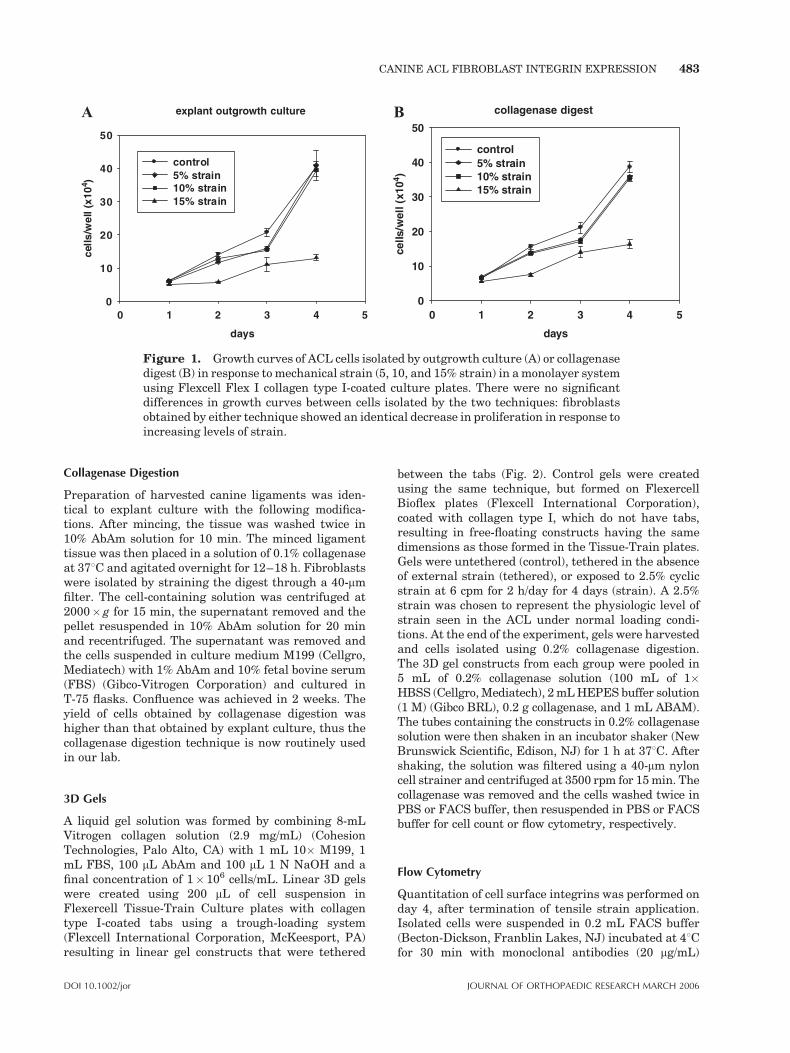
digestion from canine ACL tissue and cultured inmedium M199 (Cellgro, Mediatech, Herndon, VA) with1% antibiotic/antimycotic (AbAm; Cellgro, Mediatech)and 10% fetal bovine serum (FBS) (Gibco-VitrogenCorporation, Herndon) at 378C, 100% humidity and 5%CO2. We questioned whether the cells, which wereobtained by explant culture, were a subpopulation ofligament fibroblasts, which were more responsive tomechanical stimuli. In response to this question, weconducted experiments comparing cells derived byexplant culture to cells isolated by collagenase diges-tion. Cells derived by either technique demonstratedsimilar baseline growth kinetics. To determine whetherthe cells isolated by explant culture respond differentlyto mechanical strain, experiments were performed todetermine the effect of cyclic tensile strain on cellproliferation.24 Cells obtained by both of the aforemen-tioned techniques were exposed to tensile strain in aFLEXERCELL FX 2000 System using Flex I type Icollagen-coated plates (Flexcell International Corpora-tion, McKeesport, PA) at 5, 10, and 15% strain at 6 cpmfor 2 h followed by 22 h rest for 5 consecutive days. Theeffect of cyclic strain on the proliferation of fibroblastsisolated by the two procedures was very similar (Fig. 1).Cell proliferation in both cases was inhibited inresponse to strain; this inhibition was dependent onthe magnitude of strain (Fig. 1). In the early experi-ments, cells were obtained by outgrowth culture. Inlater experiments, cells were obtained by collagenasedigestion.
Explant Digestion Technique
Canine anterior cruciate ligaments (ACL) wereobtained from dogs sacrificed for other IACUCapproved studies at the Hospital for Special SurgeryofWeillMedical College of Cornell University within 6 hof sacrifice. The ligaments were harvested underaseptic conditions and placed in sterile medium(M199) containing 10% antibiotic–antimycotic solu-tion. A portion of the ligament at both the tibial andfemoral ends was trimmed and discarded. The outersynovial layer was removed via sharp dissection andthe ligaments were cut into 0.5–1-cm pieces and placedin media. The ligament segments were finely mincedinto 20–40 pieces and placed into T-75 flasks containingM199 medium containing 10% fetal calf serum and 1%AbAm solution. The flasks were incubated at 378C in ahumidified atmosphere of 5% CO2–95% air in a cellculture incubator. The cells migrated out of theexplants, attached to the flasks, and divided. Theexplant pieces were removed after 10–12 days. Aconfluent monolayer was routinely achieved in 2 weeks.Monolayers were then treated with 0.25% trypsin(Gibco-Vitrogen Corporation) then suspended in cellfreezing medium [90% fetal bovine serum (FBS) (Gibco-Vitrogen Corporation) and 10% Dimethyl sulfoxide(DMSO)] and cryopreserved in liquid nitrogen untilneeded for experiments.
482 HENSHAW ET AL.
JOURNAL OF ORTHOPAEDIC RESEARCH MARCH 2006 DOI 10.1002/jor
Collagenase Digestion
Preparation of harvested canine ligaments was iden-tical to explant culture with the following modifica-tions. After mincing, the tissue was washed twice in10% AbAm solution for 10 min. The minced ligamenttissue was then placed in a solution of 0.1% collagenaseat 378C and agitated overnight for 12–18 h. Fibroblastswere isolated by straining the digest through a 40-mmfilter. The cell-containing solution was centrifuged at2000� g for 15 min, the supernatant removed and thepellet resuspended in 10% AbAm solution for 20 minand recentrifuged. The supernatant was removed andthe cells suspended in culture medium M199 (Cellgro,Mediatech) with 1% AbAm and 10% fetal bovine serum(FBS) (Gibco-Vitrogen Corporation) and cultured inT-75 flasks. Confluence was achieved in 2 weeks. Theyield of cells obtained by collagenase digestion washigher than that obtained by explant culture, thus thecollagenase digestion technique is now routinely usedin our lab.
3D Gels
A liquid gel solution was formed by combining 8-mLVitrogen collagen solution (2.9 mg/mL) (CohesionTechnologies, Palo Alto, CA) with 1 mL 10� M199, 1mL FBS, 100 mL AbAm and 100 mL 1 N NaOH and afinal concentration of 1� 106 cells/mL. Linear 3D gelswere created using 200 mL of cell suspension inFlexercell Tissue-Train Culture plates with collagentype I-coated tabs using a trough-loading system(Flexcell International Corporation, McKeesport, PA)resulting in linear gel constructs that were tethered
between the tabs (Fig. 2). Control gels were createdusing the same technique, but formed on FlexercellBioflex plates (Flexcell International Corporation),coated with collagen type I, which do not have tabs,resulting in free-floating constructs having the samedimensions as those formed in the Tissue-Train plates.Gels were untethered (control), tethered in the absenceof external strain (tethered), or exposed to 2.5% cyclicstrain at 6 cpm for 2 h/day for 4 days (strain). A 2.5%strain was chosen to represent the physiologic level ofstrain seen in the ACL under normal loading condi-tions. At the end of the experiment, gels were harvestedand cells isolated using 0.2% collagenase digestion.The 3D gel constructs from each group were pooled in5 mL of 0.2% collagenase solution (100 mL of 1�HBSS (Cellgro,Mediatech), 2mLHEPES buffer solution(1 M) (Gibco BRL), 0.2 g collagenase, and 1 mL ABAM).The tubes containing the constructs in 0.2% collagenasesolution were then shaken in an incubator shaker (NewBrunswick Scientific, Edison, NJ) for 1 h at 378C. Aftershaking, the solution was filtered using a 40-mm nyloncell strainer and centrifuged at 3500 rpm for 15min. Thecollagenase was removed and the cells washed twice inPBS or FACS buffer, then resuspended in PBS or FACSbuffer for cell count or flow cytometry, respectively.
Flow Cytometry
Quantitation of cell surface integrins was performed onday 4, after termination of tensile strain application.Isolated cells were suspended in 0.2 mL FACS buffer(Becton-Dickson, Franblin Lakes, NJ) incubated at 48Cfor 30 min with monoclonal antibodies (20 mg/mL)
A explant outgrowth culture
days
10 432 5
cells
/wel
l (x1
04 )
0
10
20
30
40
50
control 5% strain10% strain15% strain
collagenase digest
days
0 1 32 54
cells
/wel
l (x1
04 )
0
10
20
30
40
50
control 5% strain10% strain15% strain
B
Figure 1. Growth curves of ACL cells isolated by outgrowth culture (A) or collagenasedigest (B) in response tomechanical strain (5, 10, and 15% strain) in amonolayer systemusing Flexcell Flex I collagen type I-coated culture plates. There were no significantdifferences in growth curves between cells isolated by the two techniques: fibroblastsobtained by either technique showed an identical decrease in proliferation in response toincreasing levels of strain.
CANINE ACL FIBROBLAST INTEGRIN EXPRESSION 483
DOI 10.1002/jor JOURNAL OF ORTHOPAEDIC RESEARCH MARCH 2006
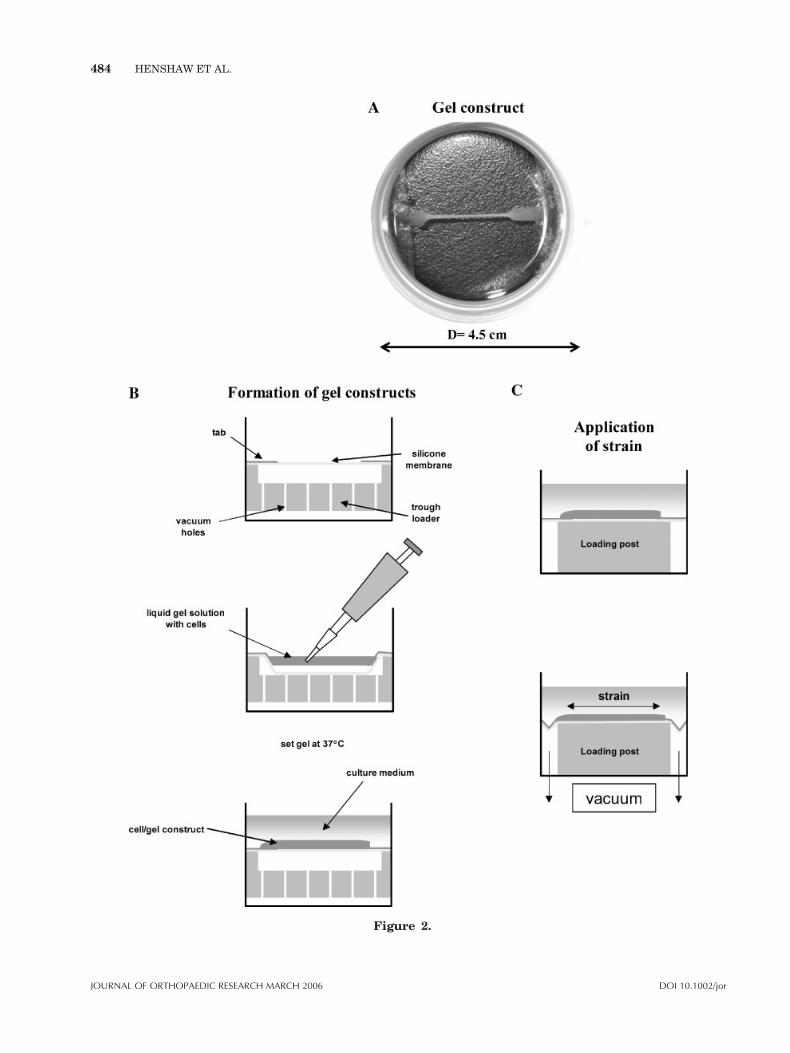
Figure 2.
484 HENSHAW ET AL.
JOURNAL OF ORTHOPAEDIC RESEARCH MARCH 2006 DOI 10.1002/jor

raised against extracellular domains of human a1 (cat.#1599 clone: HP2B6 Immunotech) and a5 (cat. #0771clone: SAM1 Immunotech), and b1 (cat. #05-232 clone:DE9, Upstate Biotech, Cleveland, OH) integrin sub-units. The cells were then washed three times withFACS buffer, resuspended in 0.2 mL of FACS buffer,and incubated with R-phycoerythrin conjugated goatantimouse IgG (Caltag Laboratories, Burlingame, CA)at 48C for 30 min. The cells were again washed threetimes with FACS buffer, suspended in 0.2 mL of FACSbuffer, and analyzed for fluorescence using a FACScanflow cytometer and CellQuest software (Becton-Dick-inson Immunocytometry Systems).
Univariate histogram displays were used to comparefluorescence intensity measurements to determine thebinding of different antibodies to cells in each treatmentgroup. The background fluorescence signal for cellsstained with isotype control was defined by markerregion 1 (M1) and fluorescence signal greater than thiscontrol was quantitated in region 2 (M2). Statisticalanalysis using CellQuest software was performed todetermine the relative distribution of cells in gate M1and M2 and the mean fluorescence intensity (MFI) forthepositive population inM2.TheMFI of 10,000 cells foreach treatment was recorded and the expression ofintegrin receptor is shown as a percent change fromcontrol values. There were a total of six experimentsperformed for a5 integrin subunit (a5) and b1 integrinsubunit (b1) andfiveexperiments fora1 integrin subunit(a1) analysis. Paired t-tests were performed usingstatistical analysis software from Sigma Plot (SPSSScience).
Histology
Gel constructs were fixed in 10% formalin, dehydrated,paraffin imbedded, and 5 mm sections created. Thesesections were stained with hematoxylin and eosin forstandard light microscopy or were utilized for fluor-escent microscopy. Fluorescence staining for f-actinwas performed using rhodamine phalloidin with aDAPI nucleic acid stain counterstain (MolecularProbes, Eugene, OR). Light and polarized light micro-scopy was used to qualitatively assess cell alignmentand collagen orientation. Four specimens from thecontrol and strained groups were examined. Experi-enced orthopedic microscopists performed the micro-scopic analysis.
RESULTS
Cell and Collagen Alignment
Histologic assessment of H&E-stained samples inthe control gels showed random cell distribution(Fig. 3A). In contrast, examination of the strainedgels demonstrated a linear distribution of cells(Fig. 3B). Control gels evaluated under polarizedlight for collagen alignment, showed a randomarrangement of collagen in the matrix, whilestrained gels showed a more linear arrangementof collagen (Fig. 4A and B).
Fluorescent staining of the actin cytoskeletonshowed a random arrangement of actin in thecontrol gels (Fig. 5A). Tethered and strainedconstructs had amore linear arrangement of actinparallel to the direction of restraint or appliedstrain (Fig. 5B and C).
Integrin Expression
The percentage of cells expressing a1, a5, and b1did not change between control, tethered, andstrained groups; however, the amount of a5 andb1 expressed by each cell varied in response totether and cyclic strain conditions.
a1 was expressed in 42� 20% of fibroblasts andwas independent of treatment group. TheMFI wasnot affected by the tether or cyclic strain condition(N¼ 5). In contrast, 93� 8% of cells expressed a5and 87� 20% of cells expressed b1 and the MFI ofeach subunit increased in response to strain. Theabsolute MFI varies from experiment to experi-ment; thus, for thepurposeofcomparisonthefluore-scent intensity for a1, a5, and b1 from each groupwas reported relative to the control values. MFI ofboth a5 and b1 was increased significantly in fibro-blasts in tethered and cyclically strained 3D colla-gen gels compared with controls (N¼ 6) (Fig. 6Aand B). a5 expression was increased 41� 24%(p< 0.007) over controls in the tethered group and48� 21% (p< 0.003) over control in the cyclicallystrained group. There was no significant differencein a5 expression between the tethered and strained
Figure 2. Flexercell Tissue Train Culture Plates (Flexcell International Corporation)with a tethered gel construct (A). Constructs are created using a trough loading systemwith 200 mL Vitrogen with 1� 105 fibroblasts/200 mL. Tethered and strained constructsattach to collagen-coated tabs at each end (B). Strained constructs are subjected to 2.5%uniaxial strain at 6 cpm 2 h per day for 4 days (C). Tethered constructs remain attachedat either end without load and control constructs are plated without tabs and allowed tofloat free.
CANINE ACL FIBROBLAST INTEGRIN EXPRESSION 485
DOI 10.1002/jor JOURNAL OF ORTHOPAEDIC RESEARCH MARCH 2006

groups. MFI of b1 also increased significantlyin response to tethered and strained conditions.For the tethered group, b1 increased 20� 11%(p< 0.006) compared with controls and 24� 15%(p< 0.01) in the strained 3D constructs comparedwith the controls. In contrast, for a1, there was nochange in MFI between control, tethered, andcyclically strained groups (n¼ 5) (Fig. 6C).
DISCUSSION
The 3D culture of ACL fibroblasts collagen gelspermits the study of fibroblast biology in an
environment where spatial orientation of cellsmay be more similar to an in vivo state comparedwith traditional monolayer culture systems. Itmust be noted, however, that although the 3Dorientation of the cells mimics that seen in anin vivo state, there are still many differences.Fibroblasts in vivo are embedded in a muchdenser matrix with a higher collagen concentra-tion and a somewhat limited vascular supply,which may result in lower levels of cell exposureto cytokines and growth factors than that seen ingel culture. This model is superior to a monolayersystem in that it provides a more uniform loadingenvironment and affords the ability to perform 2D
Figure 3. H&E staining of control (A) and strained (B) constructs (�200, originalmagnification). The constructs exposed to 2.5% cyclic tensile strain demonstrate linearalignment of the fibroblasts and collagen matrix. The unstrained constructs have curledup and demonstrate a disorganized appearance.
Figure 4. Polarized light microscopy of control (A) and strained (B) ACL constructs(�200, original magnification). The polarized light demonstrates parallel alignment ofnewly deposited collagen in strained gels in contrast to the random orientation seen inthe control.
486 HENSHAW ET AL.
JOURNAL OF ORTHOPAEDIC RESEARCH MARCH 2006 DOI 10.1002/jor

and 3D histologic analysis of the tissue. In thisstudy, the alignment of cells along the collagenmatrix reflects a response of the cellular environ-ment to the applied strain. Both cyclicallystrained and tethered constructs demonstrated
Figure 5. The effect of cyclic tensile strain on orienta-tion of f-actin filaments (�100, original magnification).Control constructs (A, B) show random distribution ofF-actin filaments and loaded constructs demonstratelinear orientation of the cytoskeletal filaments in thedirection of applied strain (arrow).
Figure 6. Effect of cyclic strain on a5 (A), b1 (B), anda1 (C) expression in ACL Fibroblast shown as percentincrease over control. In both tethered and loadedconstructs a5 and b1 mean fluorescence intensity, ameasure of receptor concentration per cell, increasedsignificantly over control values. a1 expression did notsignificantly change between groups (C).
CANINE ACL FIBROBLAST INTEGRIN EXPRESSION 487
DOI 10.1002/jor JOURNAL OF ORTHOPAEDIC RESEARCH MARCH 2006

collagen fiber alignment parallel to the long axisof the construct. Cell alignment and f-actindistribution followed the same pattern, with cellsand f-actin fibers aligned parallel to the long axisof the construct when tethered or cyclicallystrained. In contrast, free gels had random cell,f-actin and collagen alignment, further support-ing the importance of tensile strain for ligamentstructural organization. This finding shows thatmechanical stimuli are important for ligamenthomeostasis, and for development of a longitudi-nal, linear architecture.
In response to 2.5% strain, both a5 and b1expression increased significantly over controlgels. Interestingly, the tethered ligament con-structs, showed significant increases in integrinexpression when compared to untethered controlgels. We hypothesize that the cells in tetheredconstructs generate an internal ‘‘autotension’’ viacytoskeletal and adhesion proteins that results inincreased integrin expression. The mechanism forthis response is not known, but may involveproduction and realignment of f-actin filaments,upregulation of a-smooth muscle actin or inter-cellular signaling by the fibroblasts. a-Smoothmuscle actin containing cells have been demon-strated in human ACL tissue following ligamentdisruption and may play a role in ‘‘cellularautotensioning’’ and ultimate retraction of tornACL ligament stumps.9
The application of tensile strain did not have asignificant additive effect on a5 and b1 expressionwhen comparing the tethered and strained 3Dconstructs. We had initially expected that tensilestrain would have an additive effect on integrinexpression when comparing strained and tetheredgels, but this was not observed in the presentstudy. It is possible that amount of applied strainwas too low to generate a significant response, oralternatively that the density of the cells in thecollagen gel allowed significant mechanical forcesto be created by the ‘‘cellular autotension.’’ A dose–response relationship between strain and integrinexpression in 3D constructs needs to be deter-mined.
In contrast to a5 and b1, a1 expression did notchange in response to load, despite the observationthat approximately 50% of fibroblasts in all groupsexpressed a1 (Fig. 5C). The MFI did not change incontrol, tether, or strain conditions, implying thatexpression of this integrin subunit is not respon-sive to the cyclic tensile strain in the limited rangetested. a1 is known to combine with b1 creating aheterodimer that binds collagen.22 This hetero-
dimer mediates collagen matrix reorganization ininjured vascular smoothmuscle cells.25 The role ofa1 in canine ACL fibroblasts is unknown, but thesubunit may also be involved in collagen binding.However, in these experiments the expressionappears to be independent of mechanical strain atthe levels tested.
Integrin subunits are known to transmitmechanical signals in many cell populations bylinking the extracellular matrix to the cell cytos-keleton and ultimately altering DNA transcrip-tion. We have previously shown that cellssubjected to tensile strain in monolayer cultureincrease type I collagen production and decreasesynthesis ofmatrix degrading enzymes.9Althoughthese data need to be replicated in 3D constructs,the response of integrin subunits to strain in bothmonolayer and 3D culture suggests that they playa role in ACL fibroblast mechanotransduction.
Four pathways for integrin signal transduct-ion have been identified in other cell types.11
These include activation of: (1) the integrin–cytoskeleton–intermediate filament FAK path-way, (2) stress activated ion channels, (3) stressactivated protein kinases, and (4) TGF-b-mediatedparacrine and autocrine mechanisms. Selectiveinhibition and stimulation of these pathwayswill help confirm the role of cell surface integrinsin fibroblast mechanotransduction and ligamenthomeostasis. Ultimately, the ability to control ormanipulate these signal transduction pathwaysmay enhance our ability to regulate fibroblastbiosynthetic function, improve our ability toengineer implantable ACL grafts, or improveligament primary healing.
Although cell alignment in response to strainhas been shown in other cell types, this is the firstreport showing alterations in integrin subunitexpression, cell alignment, and histology of ACLfibroblasts subjected to mechanical strain in a 3Dgel system.Using a 3Dmodel, this study replicatesour previous observations made using monolayerculture, which indicated that tensile strain causesan increase in integrin expression in ACL fibro-blasts. The use of 3D collagen gels permits a moreuniform distribution of strain, and thus changes inintegrin expression are representative of a moreuniform population of cells in the gel.26 The 3Dconstruct more closely represents the in vivoligament, and allows histochemical evaluation ofthe tissue architecture in response to mechanicalstrain and enhanced fibroblast biosynthetic activ-ity. This 3Dmodel allows one to combine biochem-ical and histologic data and provides a viable
488 HENSHAW ET AL.
JOURNAL OF ORTHOPAEDIC RESEARCH MARCH 2006 DOI 10.1002/jor

technique for the study of fibroblast response toalterations in biochemical andmechanical stimuli.
ACKNOWLEDGMENTS
The authors thank Lilly Ying for her assistance withthe histologic tissue preparation. We acknowledge theintellectual support of Peter Torzilli, PhD, and RussellWarren, MD. We additionally acknowledge the philan-thropic support provided by the Institute for SportsMedicine Research, the Soft Tissue Research Fund, andthe Surgeon in Chief Fund, Hospital for SpecialSurgery. This investigation was conducted in a facilityconducted with support from Research FacilitiesImprovement Grant Number CO6-RR12538-01 fromthe National Center for Research Resources, NationalInstitutes of Health.
REFERENCES
1. National Center for Health Statistics DftNHCS.1996. Ambulatory and inpatient procedures in theUnited States, 1996. Vital Health Stat 139:1–119.
2. Kartus J, Movin T, Karlsson J. 2001. Donor-sitemorbidity and anterior knee problems after ante-rior cruciate ligament reconstruction using auto-grafts. Arthroscopy 17:971–980.
3. Tashiro T, Kurosawa H, Kawakami A, et al. 2003.Influence of medial hamstring tendon harvest onknee flexor strength after anterior cruciate liga-ment reconstruction. A detailed evaluation withcomparison of single- and double-tendon harvest.Am J Sports Med 31:522–529.
4. Strickland SM, MacGillivray JD, Warren RF. 2003.Anterior cruciate ligament reconstruction withallograft tendons. Orthop Clin North Am 34:41–47.
5. Boorman RS, Shrive NG, Frank CB. 1998. Immo-bilization increases the vulnerability of rabbitmedial collateral ligament autografts to creep.J Orthop Res 16:682–689.
6. Newton PO, Woo SL, MacKenna DA, et al. 1995.Immobilization of the knee joint alters the mechan-ical and ultrastructural properties of the rabbitanterior cruciate ligament. J Orthop Res 13:191–200.
7. Urschel JD, Scott PG, Williams HT. 1988. Theeffect of mechanical stress on soft and hard tissuerepair; a review. Br J Plast Surg 41:182–186.
8. Woo SL, Gomez MA, Sites TJ, et al. 1987. Thebiomechanical and morphological changes in themedial collateral ligament of the rabbit afterimmobilization and remobilization. J Bone JointSurg Am 69:1200–1211.
9. Murray MM, Spector M. 1999. Fibroblast distribu-tion in the anteromedial bundle of the human
anterior cruciate ligament: the presence of alpha-smoothmuscle actin-positive cells. J Orthop Res 17:18–27.
10. Amiel D, Billings E Jr, Harwood FL. 1990.Collagenase activity in anterior cruciate ligament:protective role of the synovial sheath. J ApplPhysiol 69:902–906.
11. Amiel D, Ishizue KK, Harwood FL, et al. 1989.Injury of the anterior cruciate ligament: the role ofcollagenase in ligament degeneration. J Orthop Res7:486–493.
12. Banes AJ, Tsuzaki M, Yamamoto J, et al. 1995.Mechanoreception at the cellular level: the detec-tion, interpretation, and diversity of responsesto mechanical signals. Biochem Cell Biol 73:349–365.
13. Bolcato-Bellemin AL, Elkaim R, Abehsera A, et al.2000. Expression of mRNAs encoding for alphaand beta integrin subunits, MMPs, and TIMPs instretched human periodontal ligament and gingivalfibroblasts. J Dent Res 79:1712–1716.
14. Palejwala S, Stein DE,Weiss G, et al. 2001. Relaxinpositively regulates matrix metalloproteinaseexpression in human lower uterine segment fibro-blasts using a tyrosine kinase signaling pathway.Endocrinology 142:3405–3413.
15. Parsons M, Kessler E, Laurent GJ, et al. 1999.Mechanical load enhances procollagen process-ing in dermal fibroblasts by regulating levels ofprocollagen C-proteinase. Exp Cell Res 252:319–331.
16. Attia EA, Bhargavva M, Hannafin JA. 2004. Theeffect of strain on collagen synthesis by ACLfibroblasts in vitro. In: International Symposiumof Ligament and Tendons Annual Meeting, SanFrancisco, CA.
17. Clark EA, Brugge JS. 1995. Integrins and signaltransduction pathways: the road taken. Science268:233–239.
18. Juliano RL, Haskill S. 1993. Signal transductionfrom the extracellular matrix. J Cell Biol 120:577–585.
19. Sung KL, Whittemore DE, Yang L, et al. 1996.Signal pathways and ligament cell adhesiveness.J Orthop Res 14:729–735.
20. Wang N, Butler JP, Ingber DE. 1993. Mechano-transduction across the cell surface and throughthe cytoskeleton. Science 260:1124–1127.
21. Gesink DS, Pacheco HO, Kuiper SD, et al. 1992.Immunohistochemical localization of beta 1-integ-rins in anterior cruciate and medial collateralligaments of human and rabbit. J Orthop Res 10:596–599.
22. Schreck PJ, Kitabayashi LR, Amiel D, et al. 1995.Integrin display increases in the wounded rabbitmedial collateral ligament but not the woundedanterior cruciate ligament. J Orthop Res 13:174–183.
CANINE ACL FIBROBLAST INTEGRIN EXPRESSION 489
DOI 10.1002/jor JOURNAL OF ORTHOPAEDIC RESEARCH MARCH 2006

23. Attia EA, BhargavaM, Hannafin JA. 2004. Effect ofcyclic tensile strain on ACL fibroblast proliferationintegrin expression and F-actin alignment in 3Dcollagen gels. In: Orthopaedic Research SocietyAnnual Meeting, San Francisco, CA.
24. Bhargava MM, Beavis AJ, Edberg JC, et al. 1999.Differential expression of integrin subunits in canineknee ligament fibroblasts. J Orthop Res 17:748–754.
25. Gotwals PJ, Chi-Rosso G, Lindner V, et al. 1996.The alpha1beta1 integrin is expressed duringneointima formation in rat arteries and mediatescollagen matrix reorganization. J Clin Invest 97:2469–2477.
26. Grinnell F. 2003. Fibroblast biology in threedimensional collagen matrices. Trends Cell Biol13:264–269.
490 HENSHAW ET AL.
JOURNAL OF ORTHOPAEDIC RESEARCH MARCH 2006 DOI 10.1002/jor




















