
Brief treatment with an aromatase inhibitor during sex differentiation causes chromosomally female...
8
THE JOURNAL OF EXPERIMENTAL ZOOLOGY 270:255-262 (1994) Brief llreatment With an Aromatase Inhibitor During Sex Differentiation Causes Chromosomally Female Salmon to Develop as Normal, Functional Males FRANCESC PIFERRER, SILVIA ZANUY, MANUEL CARRILLO, IGOR I. SOLAR, ROBERT H. DEVLIN, AND EDWARD M. DONALDSON Department of Fisheries and Oceans, West Vancouver Laboratory, West Vancouver; British Columbia V7V lN6, Canada (RP, I.I.S., R.H.D., E.M.D.); and Consejo Superior de Investigaciones Cientificas, Instituto de Acuicultura de Torre de la Sal, 12595 Castell6n, Spain (El?, S.Z., M.C.) ABSTRACT Although many studies implicate sex steroids in the process of sexual differentia- tion, the exact role played by these substances in lower vertebrates, especially teleost fish, is not clear, since it is not conclusively known whether sex steroids are the cause or an early consequence of sex differentiation. By hormonal manipulation and specific crossings, it is possible to produce all- female stocks of the otherwise gonochoristic chinook salmon (Oncorhynchus tshawytscha). These all- female stocks are an excellent model to study the effects of steroids on sexual differentiation since the genetic sex is known before the actual differentiation of the embryonic gonads takes place. In the present study, we show that treatment with a nonsteroidal inhibitor of aromatase, the enzyme that catalyzes the conversion from androgens to estrogens, for only 2 hours when the gonads were bipotent, caused genetic females to develop into normal males. Furthermore, these males had testes which were indistinguishable both in size and in structure from those of genetic males, and completed all the stages of spermatogenesis.At 2 years of age, these males produced viable sperm capable of induc- ing normal embryonic development when used to fertilize eggs, with a resulting all-female progeny These results provide strong support for Ymamoto's theory (Yamamoto, '69) that androgens and estrogens are natural sex inducers in gonochoristic fish, and suggest that aromatase plays a pivotal role in the sex differentiation of salmon. Thus, by brief treatment with an aromatase inhibitor at a specific time in development, an organism was induced to develop a funtional, phenotypic sex differ- ent from its genetic sex. @ 1994 Wiley-Liss, Inc. The embryonic gonad is unique as the only or- gan that can differentiate in two distinct direc- tions, towards either a testis or an ovary (Gilbert, '91). The process of sexual differentiation begins with the development of gonia into either sper- matogonia or oogonia, and proceeds through ex- tensive morphological and functional changes that ultimately result in the establishment of the male or female germ line and the sex-specific, nongerm- inal accompanying tissue. While the process or sex differentiation is well described, the responsible mechanisms in vertebrates are complex and still not fully understood (Adkins-Regan, '87; Nagai, '92). In mammals, the gonadal sex is normally de- termined by the genetic sex. Thus, Sry, the gene responsible for testis development in mice, is lo- cated at the top of a gene regulatory cascade (Koopman et al., '90). Downstream target candi- dates for these genes include the anti-Mullerian hormone (AMH), responsible for the regression of 0 1994 WILEY-LISS, INC. the Miillerian ducts and inhibition of embryonic aromatase, and the steroidogenic enzymes (Vigier et al., '89; Koopman et al., '91). Thus, it has been long suggested that in mammals the enzymatic programming leading to sex differentiation is de- termined at a genetic level, and that the rates of formation of androgens and estrogens are charac- teristic of the specific sex (Milewich et al., '77). However, the role of AMH and sex steroids in the process of sex differentiation is far from clear, es- pecially in the context of vertebrate evolution. Problems that complicate these investigations in- clude the fact that the gonadal sex is strongly de- termined by the genetic sex-induced functional sex-reversal is difficult or impossible in higher ver- Received March 4, 1994; revision accepted June 3, 1994. Address reprint requests to Dr. Edward M. Donaldson, Depart- ment of Fisheries and Oceans, West Vancouver Laboratory, 4160 Ma- rine Drive, West Vancouver, B.C., V7V 1N6, Canada.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Brief treatment with an aromatase inhibitor during sex differentiation causes chromosomally female...

THE JOURNAL OF EXPERIMENTAL ZOOLOGY 270:255-262 (1994)
Brief llreatment With an Aromatase Inhibitor During Sex Differentiation Causes Chromosomally Female Salmon to Develop as Normal, Functional Males
FRANCESC PIFERRER, SILVIA ZANUY, MANUEL CARRILLO, IGOR I. SOLAR, ROBERT H. DEVLIN, AND EDWARD M. DONALDSON Department of Fisheries and Oceans, West Vancouver Laboratory, West Vancouver; British Columbia V7V lN6, Canada (RP, I.I.S., R.H.D., E.M.D.); and Consejo Superior de Investigaciones Cientificas, Instituto de Acuicultura de Torre de la Sal, 12595 Castell6n, Spain (El?, S.Z., M.C.)
ABSTRACT Although many studies implicate sex steroids in the process of sexual differentia- tion, the exact role played by these substances in lower vertebrates, especially teleost fish, is not clear, since it is not conclusively known whether sex steroids are the cause or an early consequence of sex differentiation. By hormonal manipulation and specific crossings, it is possible to produce all- female stocks of the otherwise gonochoristic chinook salmon (Oncorhynchus tshawytscha). These all- female stocks are an excellent model to study the effects of steroids on sexual differentiation since the genetic sex is known before the actual differentiation of the embryonic gonads takes place. In the present study, we show that treatment with a nonsteroidal inhibitor of aromatase, the enzyme that catalyzes the conversion from androgens to estrogens, for only 2 hours when the gonads were bipotent, caused genetic females to develop into normal males. Furthermore, these males had testes which were indistinguishable both in size and in structure from those of genetic males, and completed all the stages of spermatogenesis. At 2 years of age, these males produced viable sperm capable of induc- ing normal embryonic development when used to fertilize eggs, with a resulting all-female progeny These results provide strong support for Ymamoto's theory (Yamamoto, '69) that androgens and estrogens are natural sex inducers in gonochoristic fish, and suggest that aromatase plays a pivotal role in the sex differentiation of salmon. Thus, by brief treatment with an aromatase inhibitor at a specific time in development, an organism was induced to develop a funtional, phenotypic sex differ- ent from its genetic sex. @ 1994 Wiley-Liss, Inc.
The embryonic gonad is unique as the only or- gan that can differentiate in two distinct direc- tions, towards either a testis or an ovary (Gilbert, '91). The process of sexual differentiation begins with the development of gonia into either sper- matogonia or oogonia, and proceeds through ex- tensive morphological and functional changes that ultimately result in the establishment of the male or female germ line and the sex-specific, nongerm- inal accompanying tissue. While the process or sex differentiation is well described, the responsible mechanisms in vertebrates are complex and still not fully understood (Adkins-Regan, '87; Nagai, '92). In mammals, the gonadal sex is normally de- termined by the genetic sex. Thus, Sry, the gene responsible for testis development in mice, is lo- cated at the top of a gene regulatory cascade (Koopman et al., '90). Downstream target candi- dates for these genes include the anti-Mullerian hormone (AMH), responsible for the regression of 0 1994 WILEY-LISS, INC.
the Miillerian ducts and inhibition of embryonic aromatase, and the steroidogenic enzymes (Vigier et al., '89; Koopman et al., '91). Thus, it has been long suggested that in mammals the enzymatic programming leading to sex differentiation is de- termined at a genetic level, and that the rates of formation of androgens and estrogens are charac- teristic of the specific sex (Milewich et al., '77). However, the role of AMH and sex steroids in the process of sex differentiation is far from clear, es- pecially in the context of vertebrate evolution. Problems that complicate these investigations in- clude the fact that the gonadal sex is strongly de- termined by the genetic sex-induced functional sex-reversal is difficult or impossible in higher ver-
Received March 4, 1994; revision accepted June 3, 1994. Address reprint requests to Dr. Edward M. Donaldson, Depart-
ment of Fisheries and Oceans, West Vancouver Laboratory, 4160 Ma- rine Drive, West Vancouver, B.C., V7V 1N6, Canada.

256 F. PIFERRER ET AL.
tebrates (Nagai, '92 )-and that the secretion of sex steroids may be a consequence, rather than the cause, of sexual differentiation.
In contrast, the sex-determining mechanisms of lower vertebrates are diverse, labile, and easily influenced by environmental factors (Adkins- Regan, '87). In particular, teleost fish exhibit all the mechanisms of sex determination known in vertebrates (Price, '84). Moreover, the gonadal sex of some fishes can be reversed by sex hormones if applied early enough during development (Yama- moto, '58). These observations led to the postula- tion that, in teleosts, androgens and estrogens are the substances responsible for male and female sex differentiation, respectively (Yamamoto, '69, '75). Since then, this has resulted in the develop- ment of protocols for the masculinization and femi- nization of a large number of fish for experimental or economic purposes (Hunter and Donaldson, '83). However, a puzzling question that remains to be answered is whether the effects of exogenous sex steroids are due to their ability to mimic the ef- fects of the endogenous sex steroids. If this is proven to be the case, it would mean that sex ste- roids are natural sex inducers in fish. Conversely, the observed effects of exogenous sex steroids may be pharmacological.
In an attempt to clarify the contribution of the sex steroids in the process of sex differentiation, we examined the role of aromatase, the enzyme that catalyzes the conversion of androgens to es- trogens. We hypothesized that since this enzyme is located at the end of the gonadal steroidogenic pathway (Nagahama, '87)' aromatase activity may act as a regulatory switch in the process of sex differentiation. Since this investigation was initi- ated, recent studies employing aromatase inhibi- tors have revealed the importance of aromatase in the normal development of the female sex in amphibia (Yu et al., '931, reptiles (Lance and Bogart, '92), and birds (Elbrecht and Smith, '92). In these studies, however, treatment with aroma- tase inhibitors resulted in the masculinization of the ovary in tadpoles (adults were not examined), in the disruption of ovarian development but not masculinization in alligators, or in the production of male chickens that, however, were not fertile.
In this first study in fish, chinook salmon (Oncorhynchus tshawytscha) was used as a model because it is possible to obtain all-female stocks of this otherwise gonochoristic species, which has an XX femaleXY male mechanism of sex deter- mination (Hunter et al., '83) comparable to that of mammals. All-female stocks have been produced
by the fertilization of normal eggs with the sperm obtained from genetic females that previously had been sex-reversed into functional males by andro- gen treatment (Hunter et al., '83). In the absence of further manipulations, the resulting progeny is 100% genotypically and phenotypically female. Thus, both genetic and gonadal sex are known in advance, well before sexual differentiation takes place. In addition, sex-reversal studies have re- sulted in the development of protocols for the mas- culinization of all-female stocks of chinook salmon by using a variety of natural and synthetic an- drogens administered before sexual development (Piferrer et al., '93). Since the production of estro- gen has been identified in Pacific salmon to start precisely at the time of highest sensitivity to the action of exogenous steroids (Feist et al., 'go), this suggests that the production of estrogen and thus aromatase activity are relevant during this pe- riod. Thus, the all-female stock of chinook salmon provides an excellent model for studying the role of steroids in the process of sexual differentia- tion and, specifically, the role of the aromatase enzyme complex.
In the present study, we have used a nonsteroi- dal aromatase inhibitor for the elucidation of the role of aromatase in normal sex differentiation in this lower vertebrate. The fish treated with in- hibitor were examined at intervals until they reached 2 years of age, at which time a propor- tion of the males begin to mature in this species and complete their life cycle.
MATERIALS AND METHODS Animals
All-female chinook salmon (Oncorhynchus tsha- wytscha) fertilized eggs were obtained from a pri- vate farm (Sea Spring Salmon Ltd., Chemainus, BC) and transported to our laboratory. These eggs had been fertilized with sperm obtained from ge- netic female chinook salmon previously masculin- ized by androgen treatment (Hunter et al., '83). Eggs were placed in small plexiglass chambers fit- ted with 1 mm2 mesh, incubated at 10 & 0.5"C in aerated well water, and left to develop until hatch- ing was complete.
Experimental treatments Stock solutions of 17a-methyltestosterone (MT),
17a-methyldihydrotestosterone (MDHT) (Sigma Chemical Co., St. Louis, MO), and the nonsteroidal aromatase inhibitor 4-(5,6,7,8-tetrahydroimidazo [1,5-a] pyridin-5-yl) benzonitrile monohydro-

SEX DIFFERENTIATION IN SALMON 257
chloride (AI) (Ciba Geigy compound CGS 16949A; Ciba Geigy, Summit, NJ) were prepared in 95% ethanol. Aliquots of the ethanolic solutions were added t o 5 liters of well water to obtain a final concentration of 1 or 10 mgAiter. The concentra- tion of ethanol in water never exceeded 0.08%. Further details of this type of treatment have been described elsewhere (Piferrer et al., '93). Each ex- perimental group received only a single 2-hour im- mersion treatment in the solutions at 10°C. Under these conditions, drugs are readily absorbed mainly through the gills into the general circula- tion (Allen and Hunn, 86). Appropriate treatment timing is essential to obtain a high proportion of males especially when a single immersion is used. Thus all treatments were carried out 3 days after 50% hatching, when Pacific salmon embryos are highly sensitive to the action of exogenous ste- roids, when the gonads are still bipotent (Piferrer et al., '93), and when aromatase appears to be- come functional (Feist et al., '90). A control group was used to verify that all fish used in these ex- periments were homogametic females.
The young fish were transported to the Rosewall Creek Experimental Fish Hatchery (Fanny Bay, BC) and reared following standard salmonid aquaculture conditions. Mortalities were recorded daily in each experimental group. Fish were grown for 6 months before the first sampling, to allow sufficient time for the gonads to develop, hence permitting the detection of permanent effects of treatment. At about 1 year of age, fish were sampled again, transferred to seawater, and reared under natural conditions in floating net pens off the coast of Vancouver Island for a further year. Then, when fish had become 2-year-old maturing adults, they were sampled for the last time.
Sampling The response of the sexually differentiating
salmonids was evaluated by investigating the go- nadal morphology of fish from each group. At 6, 14, and 18 months of age, 20 to 50 fish, depend- ing on sampling date, were randomly taken from each group, sacrificed in water containing a le- thal dose of 2-phenoxyethanol (Syndel Laborato- ries, Vancouver, BC), and measured (fork length to 0.1 cm) and weighed (to 0.01 8). Cross-sections were obtained from a point caudal to the pectoral fin, fixed in 4% paraformaldehyde, dehydrated in ethanol, and embedded in paraffin. Sections (5 pm) were stained in hematoxylin-eosin. A third sampling of about 50 additional fish per group was carried out when fish were 2 years old. In this
case, fish gonads were also exposed by laparatomy and examined visually in toto and weighed to de- termine the gonadosomatic index (GSI) as the pro- portion of gonadal weight to body weight. Thus, with the successive samplings, a total of about 100 fish were examined from each group for their gonadal condition.
Fertilization tests Sperm was obained from 2-year-old genetic fe-
males sex-reversed into males by AI treatment. For fertilization, 2 ml of sperm was mixed thor- oughly with 100 ml of pooled eggs from various females for 1 min. Water was added for 2 min and then fertilized eggs were rinsed and incubated at 10 2 1°C. Percent ocular pigmentation and per- cent hatcing were calculated to determine whether the sperm used was capable of inducing nor- mal embryonic development. The phenotypic sex of the resulting progency was determined by histological examination after 4 months, as de- scribed above.
DNA analysis Liver, gonads, and gametes were collected from
2-year-old individuals and stored frozen at -70°C. Genomic DNA was obtained from these tissues as described earlier (Devlin et al., '91). Briefly, chopped tissue was digested in proteinase K buffer (10 mM Tris, 10 mM ethylenediaminetetraacetic acid [EDTA], 1% sodium dodecyl sulphate [SDSI, 200 pg proteinase Wml, pH 8.0) at 37°C overnight. Excess protein was precipitated by adding NaCl to 1.5 M, and the samples were centrifuged at 100,OOOg for 10 min. The supernatant was ex- tracted once with phenol-chloroform-isoamyl alco- hol and precipitated with ethanol. The precipitate was then washed with 70% ethanol, centrifuged, dried briefly under vacuum, and dissolved in TE buffer (10 mM Tris, 1 mM EDTA, pH 8.0).
Genetic sex was identified using a polymerase chain reaction (PCR) test for a Y-chromosome linked growth hormone pseudogene sequence that has been described previously (Du et al., '93).
RESULTS An all-female stock of chinook salmon was
treated with an aromatase inhibitor (A11 in a single 2-hour immersion during the critical period of gonadal bipotentiality to test whether it would cause genetic females to develop into males. His- tological examination of the control group at 6 months of age revealed the presence of 100% females. In contrast, 22% of the AI-treated fish

258 F. PIFERRER ET AL.
TABLE 1. Effect of aromatase inhibitor; aromatizable and nonaromatizable androgens on the induction of male
phenotype i n genetic female salmon'
Dose Number Percent Treatment (mgAiter) sexed masculinization2
62 0 Control - A1 1 29 0 A1 10 27 22 MDHT 10 33 91 MT 10 30 23 MT+AI 10+1 32 97 MT+AI 10+10 31 98
'Female chinook salmon were exposed to the nonsteroidal aromatase inhibitor (AI), the aromatizable androgen 17a-methyltestosterone (MT), or the nonaromatizable androgen 17a-methyldihydrotes- tosterone (MDHT). Administration of the compounds alone or in com- bination was carried out 3 days after hatching, when the gonads are bipotent, through a single 2-hour immersion treatment. 'Percent masculinization calculated according to Piferrer et al. ('93).
developed as males (Table 1). Histological ex- amination showed that at this age, control females had ovaries filled with synchronously developing oocytes in the perinucleolar state (Fig. 1B). Con- versely, sex-reversed females treated with A1 had gonads (Fig. 1C) indistinguishable from testes of normal XY males of the same age (Fig. 1D).
To further characterize the effects ofAI, we took advantage of recent observations relating the ca- pacity of some adrogens to be aromatized into es- trogen with their masculinizing potency (Piferrer and Donaldson, '91; Piferrer et al., '93). Thus, the nonaromatizable androgen 17a-methyldihydro- testosterone (MDHT) at doses of 400 pgAiter and higher, consistently results in essentially 100% masculinization in chinook salmon. In contrast, aromatizable l7a-methyltestosterone (MT) causes 100% masculinization at 400 pg/liter but with pro- gressively higher doses smaller percentages of males are obtained. Since the only structural dif- ference between MDHT and MT is the presence of the 5a-reduction in the former, which confers resistance to aromatase, these findings suggest that at higher concentrations MT is aromatized into estrogen, thus reducing the number of males produced. Further proof of the ability of MT at high doses to be aromatized into estrogen comes from several experiments, including one where MT but not MDHT has been shown to be able to elicit specific estrogenic responses such as the induc- tion of' vitellogenesis (Hori et al., '79). Therefore, we used the aromatizable MT administered alone or in combination with AI, whereas nonaroma- tizable MDHT was used to confirm that chinook salmon embryos at this age are fully responsive
to brief androgen treatment. Table 1 shows that the masculinization obtained with MDHT was 91%, but only 23% after MT treatment. However, coadministration of MT with A1 resulted in the production of close to 100% males, suggesting that the AI had prevented the aromatization of MT and thus reestablished its full androgenic potency.
We observed that the masculinization of genetic females by AI treatment of sexually undifferenti- ated embryos was permanent, and that the males obtained were reproductively normal. Periodical examinations showed that the proportion of mas- culinized females remained essentially the same at 14, 18, or 24 months of age and that no differ- ences in survival existed between the control and the AI-treated groups. Furthermore, A1 treatment did not affect the somatic growth: control 55.0 fi 12.5 g vs. AI-treated (10 mg) 57.5 fi 11.0 g (mean 2 SEMI at 14 months of age.
Sexual maturation of normal chinook salmon males occurs at 2, 3, or 4 years, whereas females have a similar pattern but mature 1 year later on average (Donaldson and Benfey, '87). As in all other species of Pacific salmon, chinook die after the first sexual maturation and spawning. In our experiments after 2 years of rearing, the control females maintained their silver bright external appearance (Fig. 1A) and had ovaries containing large oocytes (Fig. 1E). In contrast, 40% of the AI-treated genetic females that developed as males matured, a proportion that exceeds the per- centage of chinook salmon genetic males that first attain sexual maturation at 2 years of age. The skin of these maturing males darkened (Fig. lA), and internal examination revealed mature testes which were normal both in shape and in size, filled with seminiferous tubules containing spermatozoa (Fig. 1F). In addition, the proportion of gonadal weight to body weight (i.e., the gonadosomatic in- dex) of these A1 masculinized females was not sig- nificantly different from that of genetic males of the same age (7.9 2 1.5% vs. 8.2 2 2.5%).
To confirm that treatment with aromatase in- hibitor renders genetic females into functional males capable of successful completion of sper- matogenesis, the sperm of 2 AI-treated females was obtained and used to fertilize eggs from nor- mal females. Both fertilization and embryonic de- velopment rates were normal (survival 91.7% to ocular pigmentation; 73.7% to hatching). Further- more, histological examination of the resulting progenies consistently showed the presence of 100% females (n = 61). Finally, the males used for the fertilization tests were sacrificed and ge-
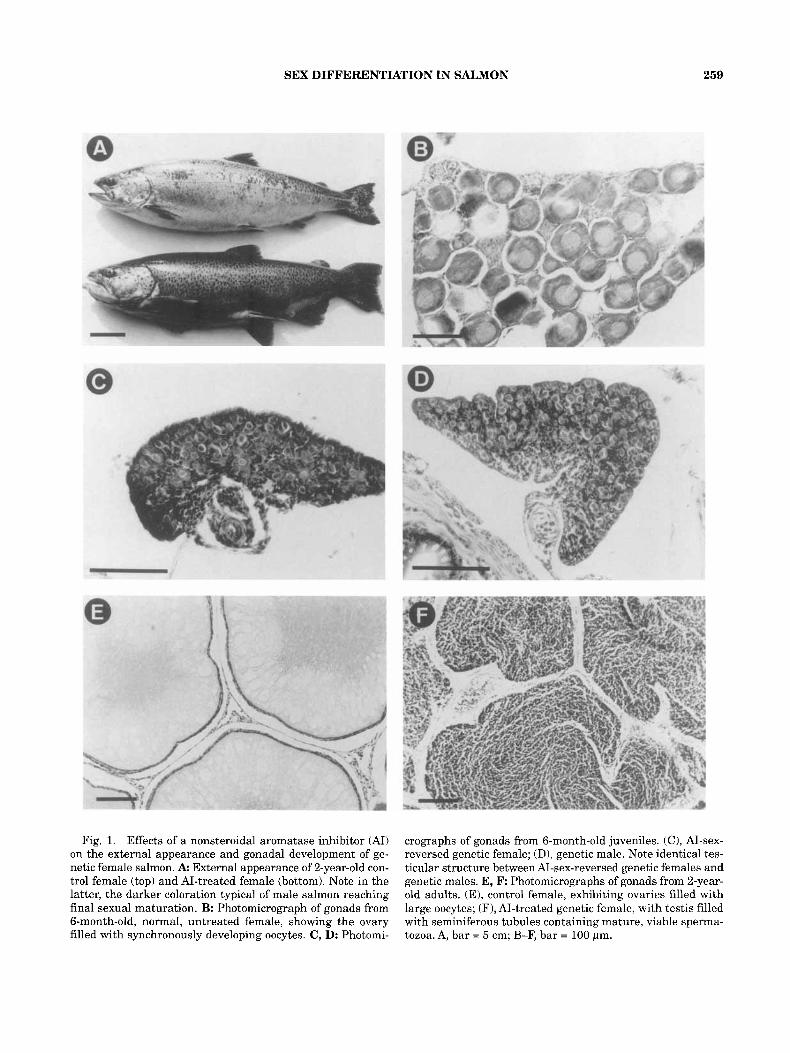
SEX DIFFERENTIATION IN SALMON 259
Fig. 1. Effects of a nonsteroidal aromatase inhibitor (AI) on the external appearance and gonadal development of ge- netic female salmon. A External appearance of 2-year-old con- trol female (top) and AI-treated female (bottom). Note in the latter, the darker coloration typical of male salmon reaching final sexual maturation. B: Photomicrograph of gonads from 6-month-old, normal, untreated female, showing the ovary filled with synchronously developing oocytes. C, D: Photomi-
crographs of gonads from 6-month-old juveniles. (C), AI-sex- reversed genetic female; (D), genetic male. Note identical tes- ticular structure between AI-sex-reversed genetic females and genetic males. E, F: Photomicrographs of gonads from 2-year- old adults. (E), control female, exhibiting ovaries filled with large oocytes; (F), AI-treated genetic female, with testis filled with seminiferous tubules containing mature, viable sperma- tozoa. A, bar = 5 cm; B-F, bar = 100 pm.

260 F. PIFERRER ET AL.
nomic DNA was prepared from either sperm or whole testes. These DNA samples were used to determine the genetic sex of these fish using a PCR test that detects a Y-chromosomal growth hormone pseudogene in chinook salmon (Du et al., '93). Determination of chromosomal sex confirmed that the males that had matured and produced sperm in the AI-treated groups were indeed ge- netic females (Fig. 2).
DISCUSSION The main purpose of the present study was to
determine whether exposure to an aromatase in- hibitor could influence the development of the phe- notype by provoking male development in fish known to be genetic females. We also set out to study whether these males would complete all stages of spermatogenesis and produce viable sperm, i.e., t o see whether these males were nor- mal from a reproductive standpoint. Because all chinook salmon used in this study were XX, the appearance of any male in the groups treated with androgen or A1 provides definite proof for the de- velopment of a gonadal sex different from the ge- netic sex. By using a new, potent nonsteroidal aromatase inhibitor, we obtained evidence suggest- ing that aromatase may play a role in the process of sex differentiation in salmon.
K b M F 1 2 C 1.0-
0.5-
0.3-
Fig. 2. Determination of chromosomal sex to show that males that matured and produced sperm at 2 years of age were indeed genetic females. Genomic DNA w-as extracted from liver (lane F), sperm (lanes M and 2), or whole testis (lane 1) of 2-year-old adults, and a polymerase chain reac- tion test for a Y-chromosomal locus (Du et al., '93) was used to determine genetic sex. The sex test identifies a male-spe- cific band of 0.3 kb in size, as well as several other non-sex specific bands. Lane Kb, molecular weight standards; lane M, control male; lane F, control female; lane 1, mature, AI-treated XX female/phenotypic male 1; lane 2, mature, AI-treated XX femaleiphenotypic male 2; lane C, no tem- plate control. Molecular weights are shown in kb in the left margin.
Previous experiments indicated that androgens such as MDHT are capable of causing an all-fe- male stock of chinook salmon to develop into males after a single 2-hour immersion treatment after hatching, at doses ranging from 0.4 to 10 mg/li- ter. Thus, in this first study in fish we used A1 at either 1 or 10 mg/liter. The lower dose of A1 was ineffective but the higher dose resulted in the pro- duction of 22% males. It is possible that the ab- sence of masculinization in the remainder of the treated salmon was the result of the short time during which A1 was administered, and that bet- ter timing, higher doses, or longer treatments would result in the masculinization of all the in- dividuals. We intend to investigate this in a fu- ture study
Treatment with MDHT and MT confirmed pre- vious results which showed that MDHT is capable of sustained masculinization even when applied at high doses, whereas MT is not (Piferrer et al., '93). The observation that combination of MT treatment with A1 resulted in essentially 100% males indicates that A1 had prevented the con- version of MT into estrogen. The reason for the additive effect of 10 mg MT/liter plus 1 mg AT/ liter is not known. However, the proportion of males after MT treatment alone was even lower than was expected. Thus, it is possible that A1 at 1 mg/ml is not sufficient to induce masculiniza- tion on its own but that it is sufficient to prevent aromatization of MT. Regarding specificity, one possibility would be that A1 exerted antiestrogenic or androgenic effects, but this seems unlikely be- cause A1 has no affinity for either androgen or estrogen receptors in mammals, and is highly spe- cific for aromatase (Steele et al., '87). The same would be expected in salmon since teleost sex ste- roid receptors are very similar to those of mam- mals (Pasmanik and Callard, '88). In addition, nonaromatizable MDHT was more potent in terms of promoting male sex differentiation than AT, which presumably blocked estrogen production by inhib- iting the aromatization of endogenous androgens.
Examination of AI-treated fish at 2 years of age confirmed that they were genetic females (molecu- lar sex identification and progeny test) which had developed into phenotypic males indistinguishable from regular males in appearance, growth, gonado- somatic index, and rate of maturity Thus, the ef- fects of A1 treatment during sex differentiation are permanent.
In chickens, treatment with an aromatase in- hibitor resulted in the masculinization of genetic females, although only 50% developed as perma-

SEX DIFFERENTIATION IN SALMON 261
nent males. Furthermore, the eggs produced by sexually mature females exposed to these males were not fertilized (Elbrecht and Smith, '92). In alligators, the same aromatase inhibitor used in this study prevented normal ovarian development but did not masculinize females (Lance and Bogart, '92). These results suggest that other regu- latory components may be necessary for normal male sexual differentiation and successful comple- tion of spermatogenesis in higher vertebrates. In contrast, after a brief treatment during a very spe- cific time in development, AI caused female sal- mon to develop as permanent and fertile males, implying that aromatase function and thus the conversion of androgen into estrogen may be nec- essary for female sex differentiation, and that the balance between androgens and estrogens can de- termine which of the two gonadal sexes develops, regardless of genetic sex. Moreover, contrary to the mode of action of steroidal aromatase inhibi- tors such as 4-hydroxyandrostenedione, which cause long-term, irreversible inactivation of aromatase enzyme activity (Brodie et al., '861, the aromatase inhibitor used in the present study binds reversibly to aromatase (Steele et al., '87). This further suggests a role for this enzyme in sex differentiation since, as A1 was applied for only a short time, it is safe to assume that the block- age was of brief duration.
Thus, it appears that during the course of ver- tebrate evolution new levels of complexity have emerged in the processes controlling sex differen- tiation. Our results clearly illustrate the lability of gonadal sex in a gonochoristic lower vertebrate. In addition, they provide evidence to support Yamamoto's theory (Yamamoto, '69) that andro- gens and estrogens are natural sex inducers in gonochoristic fish, and suggest that aromatase may play a pivotal role in the process of sex dif- ferentiation.
ACKNOWLEDGMENTS
We thank Sea Spring Salmon Ltd. for the sup- ply of all-female chinook salmon fertilized eggs, Dr. R. Love11 (Ciba-Geigy, Summit, NJ) for kindly providing the aromatase inhibitor, and R. Withler, K. Malange, A. Solmie, W. Hagen, F. Garcia, I. Baker, and the staff at the Rosewall Creek Ex- perimental Fish Hatchery for their assistance. This study was supported by a NATO Collabora- tive Research Grant CRG 890958 to M.C. and E.M.D. F.P. was the recipient of a NATO post- doctoral fellowship.
LITERATURE CITED Adkins-Regan, E. (1987) Hormones and sexual differentia-
tion. In: Hormones and Reproduction in Fishes, Amphib- ians and Reptiles. D.O. Norris and R.E. Jones, eds. Plenum Press, New York, pp. 1-29.
Allen, J.L., and J.B. Hunn (1986) Fate and distribution stud- ies of some drugs used in aquaculture. Vet. Human TOXicol.,
Brodie, A.M.H., L.-Y Wing, l? Goss, M. Dowsett, and R.C. Coombes (1986) Aromatase inhibitors and the treatment of breast cancer. J. Steroid Riochem., 24:91-97.
Devlin, R.H., B.K. McNeil, T.D.D. Groves, and E.M. Donaldson (1991) Isolation of a Y-chromosomal DNA probe capable of determining genetic sex in chinook salmon (Oncorhynchus tshaaytscha). Can. J. Fish. Aquat. Sci., 48:1606-1612.
Donaldson, E.M., and T.J. Benfey (1987) Current status of induced sex manipulation. In: Proc. Third Int. Symp. Reprod. Physiol. Fish. D.R. Idler, L.W. Crim, and J.M. Walsh, eds. Memorial University, St. John's, Newfoundland, pp.
Du, S.J., R.H. Devlin, and C.L. Hew (1993) Genomic struc- ture of growth hormone genes in chinook salmon (On- corhynchus tshawytscha): Presence of two functional genes, GH-I and GH-11, and a male-specific pseudogene, GH-y. DNA Cell Biol., 12:739-751.
Elbrecht, A., and R.G. Smith (1992) Aromatase enzyme ac- tivity and sex determination in chickens. Science, 255: 467-470.
Feist, G., C.B. Schreck, M.S. Fitzpatrick, and J.M. Redding (1990) Sex steroid profiles of coho salmon (Oncorhynchus kisutch) during early development and sexual differentia- tion. Gen. Comp. Endocrinol., 80:299-313.
Gilbert, S.F. (1991) Developmental Biology (3rd ed.) Sinauer Associates, Inc.. Sunderland, MA, 891 pp.
Hori, S.H., T. Kodama, and K. Tanahashi (1979) Induction of vitellogenin synthesis in goldfish by massive doses of an- drogens. Gen. Comp. Endocrinol., 37:306-320.
Hunter, G.A., and E.M. Donaldson (1983) Hormonal sex con- trol and its application t o fish culture. In: Fish Physiology, Vol. IX, Part B. W.S. Hoar, D.J. Randall, and E.M. Don- aldson, eds. Academic Press, New York, pp. 223-303.
Hunter, G.A., E.M. Donaldson, J. Stoss, and 1.1. Baker (1983) Production of monosex female groups of chinook salmon (Oncorhynchus tshawytscha) by the fertilization of normal ova with sperm from sex-reversed females. Aquaculture,
Koopman, F!, A. Munsterberg, B. Capel, N. Vivian, and R. Lovell-Badge (1990) Expression of a candidate sex-deter- mining gene during mouse testis differentiation. Nature, 348:450452.
Koopman, F!, J. Gubbay, N. Vivian, P. Goodfellow, and R. Lovell-Badge (1991) Male development of chromosomally female mice transgenic for Sry. Nature, 351:117-121.
Lance, VA., and M.H. Bogart (1991) Disruption of ovarian development in alligator embryos treated with an aromatase inhibitor. Gem Comp. Endocrinol., 86:59-71.
Milewich, L., EW. George, and J.D. Wilson (1977) Estrogen formation by the ovary of the rabbit embryo. Endocrinol-
Nagahama, Y. (1987) Gonadotropin action on gametogenesis and steroidogenesis in teleost gonads. Zool. Sci., 4:209-222.
Nagai, Y. (1992) Primary sex determination in mammals. Zool.
Pasmanik, M., and G.V Callard (1988) A high abundance re-
28:21-24.
108-119.
33:355-364.
ogy, 100:187-196.
Sci., 9:475-498.

262 F. PIFERRER ET AL.
ceptor in goldfish brain: Characteristics and seasonal changes. Endocrinolom, 123:1162-1171.
Piferrer, F., and E.M. Donaldson (1991) Dosage-dependent changes in the effect of aromatizable and nonaromatizable androgens on the resulting phenotype of coho salmon (Oncorhynchus kzsutch). Fish Physiol. Biochem., 9:145-150.
Piferrer, F., 1.1. Baker, and E.M. Donaldson (1993) Effects of natural, synthetic, aromatizable, and nonaromatizable an- drogens in inducing male sex differentiation in genotypic female chinook salmon (Oncorhynchus tshawytscha). Gen. Comp. Endocrinol., 91:59-65.
Price, D.J. (1984) Genetics of sex determination in fishes. A brief review. In: Fish Reproduction: Strategies and Tactics. G.W. Potts, and R.J. Wootton, eds. Academic Press, Lon- don, pp. 77-89.
Steele, R.E., L.B. Mellor, WK. Sawyer, J.M. Wasvary, and L.J. Browne (1987) In vitro and in vivo studies demonstrating potent and selective estrogen inhibition by the nonsteroi- dal aromatase inhibitor CGS 16949A. Steroids, 50:147-161.
Vigier, B., M.G. Forest, B. Eychenne, J. Bezard, 0. Garrigou, I? Robel, and M. Josso (1989) Anti-Mullerian hormone pro- duces endocrine sex reversal of fetal ovaries. Proc. Natl. Acad. Sci. U.S.A., 86:3684-3688.
Yamarnoto, T. (1958) Artificial induction of functional sex re- versal in genotypic females of the medaka (Oryzias Zatipes).
Yamamoto, T. (1969) Sex differentiation. In: Fish Physiology, Vol. 3. W.S. Hoar and D.J. Randall, eds. Academic Press, New York, pp. 117-175.
Yamamoto, T. (1975) The medaka, Oryzias Zatipes, and the guppy, Lebisles reticularis. In: Handbook of Genetics 4: Ver- tebrates of Genetic Interest. R.C. King, ed. Plenum Press, New York, pp. 133-149.
Yu, N.W., C.Y. Hsu, H.H. Ku, L.T. Chang, and H.W. Liu (1993) Gonadal differentiation and secretions of estra- diol and testosterone of ovaries of Rana catesbiana tad- poles treated with 4-hydroxyandrostenedione. J. Exp.
J. EXP. ZOO^., 137:227-260.
ZOO^., 265:252-257.




















