
Neuropeptide coding of sympathetic preganglionic neurons; focus on adrenally projecting populations

Brain-Selective Overexpression of Angiotensin (AT1)Receptors Causes Enhanced Cardiovascular Sensitivity in
Transgenic MiceEric Lazartigues, Shannon M. Dunlay, Angela K. Loihl, Puspha Sinnayah, Julie A. Lang,
Joshua J. Espelund, Curt D. Sigmund, Robin L. Davisson
Abstract—To examine the physiological importance of brain angiotensin II type 1 (AT1) receptors, we developed a noveltransgenic mouse model with rat AT1a receptors targeted selectively to neurons of the central nervous system (CNS). Atransgene consisting of 2.8 kb of the rat neuron-specific enolase (NSE) 5� flanking region fused to a cDNA encodingthe full open-reading frame of the rat AT1a receptor was constructed and transgenic mice (NSE-AT1a) were generated.Two of six transgenic founder lines exhibited brain-selective expression of the transgene at either moderate or highlevels. Immunohistochemistry revealed widespread distribution of AT1 receptors in neurons throughout the CNS. Thisneuron-targeted overexpression of AT1a receptors resulted in enhanced cardiovascular responsiveness to intracerebro-ventricular (ICV) angiotensin II (Ang II) injection but not to other central pressor agents, demonstrating functionaloverexpression of the transgene in NSE-AT1a mice. Interestingly, baseline blood pressure (BP) was not elevated in eithertransgenic line. However, blockade of central AT1 receptors with ICV losartan caused significant falls in basal BP inNSE-AT1a mice but had no effect in nontransgenic controls. These results suggest that whereas there is an enhancedcontribution of central AT1 receptors to the maintenance of baseline BP in NSE-AT1a mice, particularly effectivebaroreflex buffering prevents hypertension in this model. Used both independently, and in conjunction with miceharboring gene-targeted deletions of AT1a receptors, this new model will permit quantitative and relevant investigationsof the role of central AT1a receptors in cardiovascular homeostasis in health and disease. (Circ Res. 2002;90:617-624.)
Key Words: renin-angiotensin system � blood pressure � hypertension � central nervous system � neurons
In addition to its classical role as an endocrine system,evidence suggests that all components of the renin-angio-
tensin system (RAS) exist in individual tissues,1–3 resulting inthe local synthesis, release, and action of angiotensin II (AngII). The brain RAS4–6 is hypothesized to contribute to normalregulation of blood pressure (BP) and volume homeostasisthrough its effects on sympathetic outflow7 and vasopressinsynthesis and release.8,9
Ang II type 1 (AT1) receptors are densely localized in severalregions of the central nervous system (CNS) involved in cardio-vascular regulation, including structures of the lamina terminalisof the forebrain,10–12 brain stem sites such as the nucleus of thesolitary tract, ventrolateral medulla,6,13,14 and hypothalamic para-ventricular and supraoptic nuclei.12 Several AT1-rich sites lack ablood-brain-barrier, eg, subfornical organ, organum vasculosumof the lamina terminalis, area postrema, and median preopticnucleus, thus allowing direct interfacing with circulating Ang II.Brain AT1 receptor activation by both locally and systemicallyderived Ang II is thought to modulate normal processing ofsensory input and neurohumoral outflow.15,16 Moreover, stud-
ies showing increased density or activity of AT1 receptors inthese cardiovascular control regions in various genetic andexperimental hypertension models have implicated centralAT1 receptors as causal in the development and maintenanceof hypertension.17–19
Classical physiological and pharmacological approacheshave demonstrated the importance of the brain RAS inregulating cardiovascular homeostasis. Recently developedtransgenic strategies now permit further dissection of theprecise mechanisms by which this occurs. Using gene target-ing, we recently examined the relative contribution of AT1
receptor subtypes, AT1a and AT1b, to central Ang II respons-es.20 Although these studies did clarify that Ang II–elicitedBP increase can be selectively ascribed to the AT1a subtype,whereas AT1b receptors are necessary for the drinking re-sponse, they did not allow us to definitively establish therelative roles of central versus peripheral AT1 receptorsbecause the genes were deleted globally.
The specific objective of this study was to develop a noveltransgenic model with CNS-targeted AT1 receptors, with the
Original received August 30, 2001; revision received January 23, 2002; accepted January 25, 2002.From the Department of Anatomy and Cell Biology (E.L., S.M.D., A.K.L., P.S., J.A.L., J.J.E., R.L.D.) and the Departments of Internal Medicine and
Physiology and Biophysics (C.D.S.), the University of Iowa College of Medicine, Iowa City, Iowa.Correspondence to Robin L. Davisson, PhD, Department of Anatomy and Cell Biology, 1-251 Bowen Science Bldg, University of Iowa College of
Medicine, Iowa City, IA 52242. E-mail [email protected]© 2002 American Heart Association, Inc.
Circulation Research is available at http://www.circresaha.org DOI: 10.1161/01.RES.0000012460.85923.F0
617 by guest on May 27, 2015http://circres.ahajournals.org/Downloaded from by guest on May 27, 2015http://circres.ahajournals.org/Downloaded from by guest on May 27, 2015http://circres.ahajournals.org/Downloaded from by guest on May 27, 2015http://circres.ahajournals.org/Downloaded from by guest on May 27, 2015http://circres.ahajournals.org/Downloaded from

overall goal of unraveling the relative contribution of brainversus peripheral vascular and extravascular AT1 receptors toBP regulation. We chose to overexpress the AT1a receptorbased on our earlier findings that this subtype, but not AT1b,is critical in mediating central Ang II BP responses.
Materials and Methods
Generation of Transgenic Mice andAnimal HusbandryA fusion transgene (NSE-AT1a) consisting of 4 kb of the ratneuron-specific enolase (NSE) 5� flanking region (kind gift of J.G.Sutcliffe, Scripps Research Institute, La Jolla, Calif)21–23 and acDNA encoding the full open-reading frame of the rat AT1a receptor(kind gift of K.E. Bernstein, Emory University, Atlanta, Ga)24 wasconstructed (Figure 1A). The transgene segment was obtained bydigestion with SalI, separated by agarose gel electrophoresis, andrecovered using the Qia-Quick gel purification kit (Qiagen). Trans-genic mice were generated by microinjection into fertilized C57BL/6JX�SJL/J (B6SJL F2) mouse embryos, and positive founders (seenext paragraph) were bred for study as described previously.25 Allmice were fed standard mouse chow (LM-485; Teklad PremierLaboratory Diets) and water ad libitum. Care of the mice used in theexperiments exceeded the standards set forth by the NIH in theirguidelines for the care and use of experimental animals. All
procedures were approved by the University Animal Care and UseCommittee at the University of Iowa.
Analysis of Nucleic AcidsGenomic DNA was purified from tail biopsies and subjected to eitherSouthern blot analysis for identification of positive founders (Figure1A) or to PCR for identification of transgenic offspring, bothdescribed in detail previously.26,27 Transgene expression was ana-lyzed using RNase protection assay (RPA) and a cDNA probederived from a region of rat AT1a sequence with relatively lowernucleotide homology to the mouse sequence compared with the restof the gene (184–586 relative to transcription start site). The probewas PCR-amplified from rat genomic DNA, cloned into pCRII(Invitrogen), and single-stranded antisense RNA was generated byan in vitro transcription reaction containing T7 polymerase and�-32P[UTP]. To obtain sufficient RNA from peripheral neural tis-sues, ganglia and nerve bundles were widely harvested and com-bined from cervical, para-aortic, and extremity sites. RPAs wereperformed using the HybSpeed RPA kit (Ambion) following themanufacturer’s instructions and as described previously.27 Quantifi-cation was performed using a PhosphorImager SI system (MolecularDynamics) in conjunction with ImageQuant software (version 5.0).AT1a mRNA levels were normalized by dividing the AT1a signal bythe mouse �-actin signal.
Detection of AT1 Receptor ProteinAnalysis of AT1 receptor levels in brain of transgenic and nontrans-genic mice was performed by Western blot analysis as describedpreviously28 using a rabbit polyclonal antibody (Santa Cruz, sc-579,1:100 dilution). Intensities of the bands were analyzed by densitom-etry using ImageQuant software (see previous paragraph).
Immunofluorescent localization of AT1 receptor protein in brain oftransgenic mice (6084-2 n�5; 6085-2 n�5) and nontransgeniccontrols (n�5) was performed using a rabbit polyclonal antibodytargeted against the carboxyl terminus of the rat AT1a receptor(kindly provided by Dr M. McKinley, Howard Florey Institute,Parkville, Victoria, Australia). As described in detail previously,29
brains were perfused, postfixed overnight, and cryosectioned (30�m, coronal). Free-floating sections were subjected to immunofluo-rescence staining using the tyramide signal amplification method asdescribed previously30 and incubated with AT1 receptor antibody for48 hours (1:5000 dilution). In addition to specificity controls, adilution series for AT1 antibody was carried out (1:5000, 1:10 000,1:20 000), and comparable AT1 immunofluorescence was observedat all concentrations. Finally, primary antibody was omitted in someincubations (transgenic and control) to check for autofluorescence.Confocal microscopy (Zeiss LSM 510) was used to analyze immu-nohistochemical staining (488 nm for visualization of fluorescein).Anatomical analyses were performed with the aid of a mouse brainatlas.31
Physiological StudiesNSE-AT1a transgenics from two different founder lines (6085-2,n�9; 6084-2, n�7) and nontransgenic controls (n�12) were surgi-cally instrumented with intracerebroventricular (ICV) and left com-mon carotid arterial cannulae for central administration of drugs andmeasurement of cardiovascular parameters, respectively, as de-scribed previously.27 In the first set of experiments, the dose-dependent effects of Ang II (50 to 200 ng; 200 nL) were determined.Doses were given randomly, only two consecutive injections wereperformed in each mouse, and at least 30 minutes was allowedbetween injections. Baroreflex sensitivity was estimated using linearregression analysis for the central Ang II–induced cardiovascularchanges in heart rate and mean arterial pressure (�HR/�MAP) asdescribed previously.27,32 To confirm AT1 receptor involvement,mice (6084-2, n�5; 6085-2, n�5; nontransgenics, n�5) werepretreated ICV with either saline or the AT1 receptor antagonistlosartan (20 �g) followed 20 minutes later by ICV injection of thehighest dose of Ang II (200 ng). Finally, to determine the selectivityof the cardiovascular responses to Ang II, we also examined the
Figure 1. Structure of the NSE-AT1a transgene and detection oftransgenic founders. A, Fusion construct contains elementsessential to CNS-specific expression of the transgene, includingthe neuron-specific enolase (NSE) promoter, intron sequence toaid in processing/transport of the mRNA, and a polyA additionto target appropriate 3� end processing. The AT1a cDNA con-tains the full open-reading frame of the receptor. The probeused for Southern blot analysis is indicated. B, RepresentativeSouthern blot showing various NSE-AT1a transgenic founders,including the two lines used in physiological studies herein(6085-2 and 6084-2). Genomic DNA (10 �g) from tail biopsieswas digested with SacI, separated on 0.8% agarose gel, andhybridized to a random primer-labeled probe (indicated in panelA). The probe detects a 4.4-kb band diagnostic of the trans-gene. Relative transgene copy number can be estimatedbecause the probe used detects both mouse (M) AT1 receptorsubtypes (a and b) as well as the exogenous rat (R) AT1 recep-tor. Positive transgenic founders are indicated by asterisks (*).
618 Circulation Research March 22, 2002
by guest on May 27, 2015http://circres.ahajournals.org/Downloaded from

effects of another central pressor agent, the cholinergic agonistcarbachol (50 ng ICV), in a separate group of transgenic (6085-2,n�4; 6084-2, n�4) and nontransgenic (n�4) mice.
StatisticsData are expressed as mean�SEM. Data were analyzed by Student’st test or ANOVA (following Bartlett’s test of homogeneity ofvariance) followed by Newman-Keuls correction for multiple com-parisons. Statistical analyses were performed using Prism (version3.0) software package (GraphPad Software Inc).
ResultsSix NSE-AT1a transgenic founders (of 59 pups born) contain-ing varying copies of the transgene were identified by thepresence of a 4.4-kb hybridization band on Southern blots(Figure 1B). All six founders were successfully bred toestablish transgenic lines, with transmission of the transgeneto �50% of male and female offspring. By using RPA, threeseparate transgenic lines with differing levels of transgenemRNA in brain were identified (6084-2, 6055-1, and 6085-2)(Figures 2A and 2B). Line 6084-2 exhibited the highest levelof transgene expression, while lines 6085-2 and 6055-1exhibited more moderate levels.
Focusing on these 3 lines, we next determined whether thetransgene was indeed expressed selectively in brain. TotalRNA was purified from a wide variety of tissues of both maleand female transgenic and nontransgenic mice and wassubjected to RPA using the same transgene-specific probe. Arepresentative RPA of a positive NSE-AT1a (line 6085-2)mouse and a nontransgenic littermate is shown in Figures 2Cand 2D. High-level expression of the transgene was detectedin brain but in no other tissues examined in line 6085-2, and
the lack of expression in brain or any other tissues innontransgenic controls further confirmed selectivity of theprobe. Extremely low-level expression of rat AT1a receptorswas detected in the adrenal gland of 6085-2 mice, butinterestingly, the transgene was undetectable in peripheralneural tissue. This observation was confirmed using RT-PCR(data not shown). The same brain-restricted (and very low-level adrenal) expression pattern was observed in line 6084-2,and no sex-related differences in expression level or patternwere observed in either line. In contrast, line 6055-1 exhib-ited widespread expression of the transgene with varyinglevels between males and females, thus further studies wererestricted to lines 6084-2 and 6085-2.
To determine whether NSE-AT1a transgene expression wasalso reflected at the protein level, Western blot analysis wasperformed on brains from 6084-2 and 6085-2 mice andnontransgenic littermates. Because all available antibodiescross-react with both mouse and rat AT1 receptors, wecompared brain AT1 protein levels in transgenic mice withbaseline levels of the protein in nontransgenic controls. Asshown in Figure 3, both 6084-2 and 6085-2 mice exhibitenhanced AT1 receptor protein (�70-kDa band) in braincompared with nontransgenic animals, and the relative levelsare correlated to mRNA levels for these two lines (see Figure 2).
Distribution of transgene expression in brain of 6084-2 and6085-2 mice was determined using immunohistochemistryand confocal microscopy. The AT1 receptor antibody used forthese studies was specifically developed for use in brainimmunocytochemistry at light and electron microscopic lev-els.33 Although the antibody does not distinguish between
Figure 2. Brain-specific expression of the transgene. A, Representative RNase protection assay of rat AT1a transgene expression in 20�g of brain RNA from each of the positive transgenic founder lines and nontransgenic controls (non-trans.). Selectivity of the AT1a
probe for the transgene is indicated by the lack of bands in nontransgenics. B, Summary of transgenic AT1a mRNA levels (normalizedto �-actin signal) in different mice (n�3) from each line. Quantification was performed as described in Materials and Methods. Valuesare expressed as mean�SEM. C and D, Representative RNase protection assays of 20 �g of total tissue RNA isolated from NSE-AT1a
(line 6085-2) transgenic and nontransgenic mice. The rat AT1a and �-actin protected transcripts are indicated, as are the free probes.Rat kidney was used as a positive control. All panels are 24-hour exposures.
Lazartigues et al Brain-Selective AT1a Receptor Overexpression 619
by guest on May 27, 2015http://circres.ahajournals.org/Downloaded from

endogenous and transgenic (rat) AT1 receptors, a comparisonof distribution and fluorescence intensity between transgenicand nontransgenic mice allowed us to qualitatively estimaterelative levels of transgene expression. Widespread AT1
receptor immunofluorescence was detected throughout thebrain of both transgenic lines, with a similar distributionpattern in each line (see online data supplement available athttp://www.circresaha.org). This is in contrast to the ratherdiscrete localization of AT1 receptors to a few areas of thebrain, most notably components of the lamina terminalis ofthe forebrain, brain stem nuclei such as the nucleus of thesolitary tract, ventrolateral medulla, and hypothalamic areas,in nontransgenic mice and in rats using this antibody.33 Inaddition to a more widespread distribution, abundance of AT1
receptors was markedly elevated in both transgenic linescompared with controls and generally higher in 6084-2 versus6085-2 mice. This is illustrated in typical confocal images(Figures 4A through 4F) of two representative regions, thelateral hypothalamic area (left panels) and the supraopticnucleus (right panels), where AT1 receptor staining is highlyabundant in 6084-2 (Figures 4A and 4B), abundant in 6085-2(Figures 4C and 4D) and either undetectable or at low levelsin nontransgenic (Figures 4E and 4F) mice. Figures 4G and4H) demonstrates low levels of background staining insections from nontransgenics, indicating that AT1 immuno-staining observed in transgenic animals is not due to nonspe-cific activity of the antibody. A complete lack of immuno-fluorescence in sections (transgenic and controls; data notshown) incubated without the primary antibody further con-firms the specificity of staining.
Strict neuron-specific localization of the transgene in brainof NSE-AT1a mice is difficult to demonstrate because of thecross-reactivity of the AT1 antibody with both the transgeneand endogenous AT1 receptors that may be localized to glialcells.34–36 However, we performed morphological analyses toconfirm localization of AT1 immunostaining in neurons. Asseen in representative examples in Figures 4A, 4C, and 4E),AT1 staining was observed in cell bodies and fibers withmorphological characteristics typical of neurons. This neuro-nal pattern of staining was observed throughout the brain inboth transgenic lines.
To demonstrate functional expression of exogenous AT1a recep-tors in NSE-AT1a transgenic mice, we examined the dose-dependenteffects of central acute administration of Ang II on MAP and HR inconscious freely moving mice. Resting MAP and HR values beforeAng II are summarized (see Table). The only significant alterationin basal cardiovascular values was a significant tachycardia ob-
served in 6084-2 mice. There were no significant differences inbaseline MAP between either transgenic line and nontransgeniclittermates, indicating that brain-selective overexpression of AT1a
receptors did not cause hypertension under conditions of basal RASactivation. However, both transgenic lines exhibited significantlyenhanced cardiovascular sensitivity to central Ang II compared withcontrols (Figure 5). In nontransgenics, Ang II caused typical pressorand bradycardic responses observed previously in mice21 and inmany other species,6,32,37 but the magnitude of these responses wasdramatically increased in both transgenic lines (Figure 6). This is incontrast to the equivalent cardiovascular effects (P�0.05 for allcomparisons) produced by the central pressor agent, carbachol,38 innontransgenic (�MAP, 14�1 mm Hg; �HR, �61�11 bpm, n�4),6084-2 (�MAP, 17�3 mm Hg; �HR, �74�21 bpm), and 6085-2(�MAP, 13�1 mm Hg; �HR, �73�27 bpm, n�4) mice. Inaddition, central Ang II–induced baroreflex responses were signif-icantly enhanced in both 6085-2 and 6084-2 (�3.62�0.46 and�6.3�0.47 bpm/mm Hg, respectively; P0.05) compared withnontransgenics (�1.98�0.28 bpm/mm Hg). To confirm that theenhanced cardiovascular sensitivity to central Ang II in NSE-AT1a
transgenics is indeed due to AT1 receptors and not some othermechanism, we examined the effects of losartan on Ang II–inducedcardiovascular responses in separate animals (Figure 6B). Interest-ingly, losartan itself caused a significant decrease in baseline MAPin both transgenic lines (6085-2: �MAP �18�2 mm Hg; 6084-2:�MAP �11�2 mm Hg; P0.01 versus control) but had no effecton MAP in nontransgenic mice (1�2 mm Hg). The Ang II–induced pressor and bradycardic responses were completely abol-ished by pretreatment with the AT1 antagonist in both transgeniclines and in controls, confirming that AT1 receptors are essential inmediating the Ang II–induced cardiovascular effects in all threegroups.
In addition to markedly different effects of central Ang IIon the magnitude of MAP and HR responses in NSE-AT1a
and control mice, the time course of these cardiovascularchanges differed between the groups. In control mice, theincrease in BP and decrease in HR were similar in duration(4.42�0.27 and 6.43�1.2 minutes, respectively), indicativeof activation of baroreflex mechanisms. In contrast, bothtransgenic lines exhibited much longer-lasting bradycardicresponses relative to the duration of the pressor responses.Whereas the Ang II–induced increases in BP were of signif-icantly longer duration in both transgenic lines comparedwith controls (6085-2: 7.47�0.39 minutes; 6084-2:6.73�0.88 minutes; P0.05), the bradycardic response per-sisted long after the BP had returned to baseline in these mice
Figure 3. AT1 receptor levels are elevated in brain ofNSE-AT1a mice. A, Representative Western blot analy-sis of 50-�g brain protein from NSE-AT1a transgenic(6084-2 and 6085-2) and nontransgenic mice, using arabbit polyclonal antibody that cross-reacts with bothmouse and rat AT1 receptors. Rat kidney and braintissues were used as positive controls for rat AT1
receptor protein. The immunoreactive AT1 band (�70kDa) is marked by an arrow. B, Summary of AT1
receptor levels in different mice (n�5) from eachtransgenic line and nontransgenic controls. Quantifi-cation was performed as described in Materials andMethods. Data are expressed as mean�SEM.***P0.001, **P0.01, and *P0.05.
620 Circulation Research March 22, 2002
by guest on May 27, 2015http://circres.ahajournals.org/Downloaded from

(6085-2: 22.71�3.2 minutes; 6084-2: 27.56�4.39 minutes;P0.001 versus control).
DiscussionIntrinsic tissue RAS, defined as tissue-based systems with thepotential for local Ang II production and activity, have beenhypothesized to play critical roles in cardiovascular regula-tion that are distinct from that mediated by the classicalsystemic RAS. However, the importance of this mechanismremains unresolved because of the difficulty in experimen-tally dissecting tissue and systemic RASs. The brain RAS isparticularly controversial in this regard because of the direct
interfacing of central Ang II receptors with circulating Ang IIat brain regions lacking a blood-brain-barrier. As an initialstep toward dissecting the relative physiological significanceof brain versus peripheral vascular and extravascular Ang IIreceptors, we report herein the development of a noveltransgenic mouse model with functional expression of exog-enous AT1a receptors selectively in the brain. Our long-termgoal is to utilize these mice to examine the physiologicalconsequences of overexpression of AT1a receptors selectivelyin brain of normal mice (with intact endogenous AT1 recep-tors) and in mice otherwise devoid of AT1a receptors throughbreeding onto the AT1a-null background.27,39
Of six transgenic founders harboring the rat AT1a receptorunder the control of the NSE promoter, two individuallyderived lines exhibited exogenous AT1a mRNA almost exclu-sively in the brain. No other tissues were positive for rat AT1a
mRNA except low-level expression in the adrenal gland,which was not surprising given that medullary chromaffin
Figure 4. NSE-AT1a mice exhibit enhanced AT1 receptor immu-nostaining in widespread regions of the brain. Confocal imagesof AT1 receptor immunoreactivity in representative brain regions,including the lateral hypothalamic area (left panels) and thesupraoptic nucleus (right panels) from 6084-2 (A and B), 6085-2(C and D), and nontransgenic (E and F) mice. Coronal sections(30 �m) were incubated with a primary antibody targeted to thecarboxyl terminus of the AT1a receptor. Specificity of the AT1
antibody was confirmed in control experiments in which sec-tions from nontransgenic (G) and 6084-2 transgenic (H) micewere coincubated in the same antibody bath. fx indicates fornix;LHt, lateral hypothalamus; opt, optic tract; PaLM, paraventricu-lar hypothalamic lateral magnocellular nucleus; PaMM, paraven-tricular hypothalamic medial magnocellular nucleus; PaMP,paraventricular hypothalamic medial parvicellular nucleus; andSON, supraoptic nucleus.
Figure 5. NSE-AT1a transgenics exhibit cardiovascular hyper-sensitivity to ICV Ang II. A typical example of the effects of AngII (200 ng, 200 nL) administered ICV on cardiovascularresponses in nontransgenic and NSE-AT1a (6085-2) transgenicmice. Mice were instrumented with ICV and carotid arterial can-nulae and allowed 4 days of recovery. Experiments were per-formed in conscious, freely moving animals. Arrows indicatetime of Ang II injection. PP indicates pulsatile pressure; MAP,mean arterial pressure; and HR, heart rate.
Baseline MAP and HR in NSE-AT1a Transgenic andNontransgenic Controls
Nontransgenic(n�12)
6085-2(n�9)
6084-2(n�7)
MAP, mm Hg 125�3 123�4 125�4
HR, bpm 626�14 607�29 703�14*
BP and HR were recorded in conscious, freely moving mice via a catheterinserted in the left common carotid artery. Mice were allowed 4 days ofrecovery. Values are expressed as mean�SEM.
*P0.05, vs nontransgenic controls.
Lazartigues et al Brain-Selective AT1a Receptor Overexpression 621
by guest on May 27, 2015http://circres.ahajournals.org/Downloaded from

cells stain positive for NSE and are of neuroectodermalorigin.40 We chose the NSE promoter because NSE is adistinct isoform of the glycolytic enzyme enolase found onlyin terminally differentiated neurons and neuroendocrine cells,whereas a different enolase isozyme (non–neuronal enolase)is expressed in neuroblasts and all other non–neuronal cells ofthe CNS.40 Hence, NSE-driven transgenes are reported to beneuron-selective.21,22,41 Although we could not rule out thepossibility of AT1a transgene expression in astrocytes or othernon–neuronal cells, our morphological analyses suggest sig-nificantly enhanced AT1 receptor staining in neurons ofNSE-AT1a mice.
Consistent with the finding that NSE is one of the mostabundantly expressed and widely distributed proteins in theadult mammalian brain,42 NSE-AT1a transgenics exhibitedAT1a immunostaining at high levels throughout most brainregions. The losartan-sensitive enhanced cardiovascular re-sponsiveness of both transgenic lines to acute central Ang IIis presumably the functional consequence of overexpressionof AT1a receptors observed in cardiovascular control regionsof these mice. These data provide important evidence forfunctional expression of the transgene. Interestingly, the AT1a
expression level is not directly correlated with cardiovascularsensitivity in the two transgenic lines. This may be interestingwith regard to differential regulation and/or neural circuitryassociated with transgene expression in these two differenttransgenic lines. The mechanism of enhanced cardiovascularsensitivity to central Ang II in NSE-AT1a mice is the subjectof ongoing investigations.
Overexpression of AT1a receptors in brain stem sites knownto regulate sympathetic outflow43 and in hypothalamic nucleiinvolved in vasopressin synthesis and release44,45 raises the
possibility that one or both of these mechanisms is involvedin the hyperresponsiveness of NSE-AT1a mice. The relativeeffect of systemically versus centrally administered Ang II inthe context of AT1a overexpression observed in circumven-tricular organs of these mice is also of considerable ongoinginterest.
Interestingly, despite increased Ang II–elicited pressor andbradycardic responses, NSE-AT1a transgenics are not hyper-tensive at rest. At first, this may seem to contradict earlierheld notions that increased levels of AT1 receptor protein,mRNA, or binding activity found in central cardiovascularcontrol regions of various genetic and experimental models ofhypertension were responsible for the increased BP in thesemodels.17,18 However, the present data and earlier findingscan be reconciled by postulating that increased brain AT1
receptor levels alone do not cause centrally mediated hyper-tension, but activation of the RAS, ie, increased production ofAng II, either centrally or peripherally, is also required. Weare currently investigating this possibility by examining theeffects of sustained RAS activation on baseline BP inNSE-AT1a mice using radiotelemetry.
Another explanation for normal resting BP in mice withfunctional overexpression of brain AT1a receptors is thatbaroreflex activity is particularly effective in maintaining BPhomeostasis in the face of increased receptor numbers. This issupported by our finding that NSE-AT1a mice exhibit signif-icant depressor responses to ICV losartan, whereas centralblockade of AT1 receptors has no effect on baseline BP incontrol mice. Furthermore, centrally induced baroreflex re-sponses seem enhanced in both transgenic lines comparedwith controls as suggested by our baroreflex estimation.Taken together, these results suggest that although there is anenhanced contribution of central AT1 receptors to the main-tenance of baseline BP in NSE-AT1a transgenic mice, effec-tive baroreflex buffering prevents hypertension in this model.However, a complete understanding of baroreflex function inthis model will require direct investigation of HR changeselicited over a range of centrally and peripherally inducedchanges in BP.32
Analysis of baroreflex function in NSE-AT1a transgenics isalso of interest given the persistent bradycardia observed inthese mice long after return of BP to baseline on ICV Ang II.Indeed, the temporal profile of this response suggests that atleast a portion of the Ang II–induced bradycardia is notdirectly the result of baroreflex activity. One possible mech-anism may involve the well-known sympathoinhibitory ef-fects of vasopressin via actions at the area postrema.46 It isplausible that overexpression of AT1a receptors in NSE-AT1a
mice leads to enhanced Ang II–induced release of vasopres-sin, which in turn acts on area postrema neurons that interactwith other hindbrain sites known to mediate sympathoinhibi-tion. Alternatively, the long-lasting Ang II–elicited bradycar-dia may involve direct activation of areas involved in HRcontrol, eg, the nucleus ambiguus, ventrolateral medulla, andnucleus tractus solitarius. Indeed, these areas exhibit high-level overexpression of AT1 receptors in the NSE-AT1a
transgenic mice.The functional consequences of ectopic and noncardiovas-
cular region–localized expression of AT1a receptors in brain
Figure 6. Cardiovascular hypersensitivity in NSE-AT1a mice isprevented by losartan. A, Summary of the effects of Ang II (50,100, 200 ng; 200 nL) administered ICV on MAP and HR in non-transgenic (open bars, n�12), 6084-2 (shaded bars, n�7), and6085-2 (solid bars, n�9) transgenic mice. B, Effects of Ang II(200 ng, ICV) on MAP and HR after pretreatment with eithersaline or the AT1 receptor antagonist losartan were examined inseparate nontransgenic (n�5), 6084-2 (n�5), and 6085-2 (n�5)transgenic mice. Data are expressed as absolute change(�mean�SEM). ***P0.001, **P0.01, and *P0.05 vs non-transgenics; †P0.01 vs saline-treated.
622 Circulation Research March 22, 2002
by guest on May 27, 2015http://circres.ahajournals.org/Downloaded from

of NSE-AT1a transgenic mice remain to be determined.Central AT1 receptors have been implicated in a wide varietyof responses that are apparently independent of BP regula-tion, including anxiety,47 exploratory behavior,48 dopamineformation and release,49 and development, plasticity, andmaturation of the brain.50 This, coupled with the widespreaddistribution of AT1 receptors in brain of NSE-AT1a transgen-ics, makes these mice potentially useful models for exploringother noncardiovascular-related functions of AT1 receptors.
In summary, we have developed a useful transgenic modelthat exhibits functional overexpression of AT1a receptorsselectively in the brain. Used both independently, and inconjunction with mice harboring gene-targeted deletions ofAT1a receptors, this new model will permit quantitative andrelevant investigations of the role of central AT1a receptors incardiovascular homeostasis in health and disease.
AcknowledgmentsThe work described herein was funded by grants from the NIH(HL14388-30 and HL63887-02). E. Lazartigues is funded by apostdoctoral fellowship (20572Z) from the American Heart Associ-ation. The authors would like to thank M. Cassell (University ofIowa) for assistance with neuroanatomical analyses and valuablediscussions and Merck and DuPont Laboratories for providinglosartan. Transgenic mice were generated and maintained at theUniversity of Iowa Transgenic Animal Facility, which is supportedin part by the College of Medicine and the Diabetes and Endocri-nology Research Center. DNA sequencing was performed at theUniversity of Iowa DNA Core Facility.
References1. Phillips MI, Speakman EA, Kimura B. Levels of angiotensin and
molecular biology of the tissue renin angiotensin systems. Regul Pept.1993;43:1–20.
2. Lee MA, Bohm M, Paul M, Ganten D. Tissue renin-angiotensin systems:their role in cardiovascular disease. Circulation. 1993;87:7–13.
3. Ganong WF. Origin of the angiotensin II secreted by cells. Proc Soc ExpBiol Med. 1994;205:213–219.
4. Lippoldt A, Paul M, Fuxe K, Ganten D. The brain renin-angiotensinsystem: molecular mechanisms of cell to cell interactions. Clin ExpHypertens. 1995;17:251–266.
5. Muratani H, Teruya H, Sesoko S, Takishita S, Fukiyama K. Brain angio-tensin and circulatory control. Clin Exp Pharmacol Physiol. 1996;23:458–464.
6. Phillips MI, Sumners C. Angiotensin II in central nervous system phys-iology. Regul Pept. 1998;78:1–11.
7. Scroop GC, Lowe RD. Efferent pathways of the cardiovascular responseto vertebral artery infusions of angiotensin in the dog. Clin Sci (Colch).1969;37:605–619.
8. Unger T, Rascher W, Schuster C, Pavlovitch R, Schomig A, Dietz R,Ganten D. Central blood pressure effects of substance P and angiotensinII: role of the sympathetic nervous system and vasopressin. EurJ Pharmacol. 1981;71:33–42.
9. Paul M, Bader M, Steckelings UM, Voigtlander T, Ganten D. Therenin-angiotensin system in the brain: localization and functional signif-icance. Arzneimittelforschung. 1993;43:207–213.
10. Saavedra JM, Correa FM, Kurihara M, Shigematsu K. Increased numberof angiotensin II receptors in the subfornical organ of spontaneouslyhypertensive rats. J Hypertens. 1986;4:S27–S30.
11. McKinley MJ, Badoer E, Vivas L, Oldfield BJ. Comparison of c-fosexpression in the lamina terminalis of conscious rats after intravenous orintracerebroventricular angiotensin. Brain Res Bull. 1995;37:131–137.
12. Lenkei Z, Corvol P, Llorens-Cortes C. The angiotensin receptor subtypeAT1A predominates in rat forebrain areas involved in blood pressure, bodyfluid homeostasis and neuroendocrine control. Brain Res. 1995;30:53–60.
13. Muratani H, Ferrario CM, Averill DB. Ventrolateral medulla in sponta-neously hypertensive rats: role of angiotensin II. Am J Physiol. 1993;264:R388–R395.
14. Lind RW, Swanson LW, Ganten D. Organization of angiotensin II immu-noreactive cells and fibers in the rat central nervous system: an immu-nohistochemical study. Neuroendocrinology. 1985;40:2–24.
15. Keil LC, Summy-Long J, Severs WB. Release of vasopressin by angio-tensin II. Endocrinology. 1975;96:1063–1065.
16. Ito S, Sved AF. Blockade of angiotensin receptors in rat rostral ventro-lateral medulla removes excitatory vasomotor tone. Am J Physiol. 1996;270:R1317–R1323.
17. Gutkind JS, Kurihara M, Saavedra JM. Increased angiotensin II receptorsin brain nuclei of DOCA-salt hypertensive rats. Am J Physiol. 1988;255:H646–H650.
18. Gutkind JS, Kurihara M, Castren E, Saavedra JM. Increased concen-tration of angiotensin II binding sites in selected brain areas of sponta-neously hypertensive rats. J Hypertens. 1988;6:79–84.
19. Nazarali AJ, Gutkind JS, Correa FM, Saavedra JM. Enalapril decreasesangiotensin II receptors in subfornical organ of SHR. Am J Physiol.1989;256:H1609–H1614.
20. Davisson RL, Oliverio MI, Coffman TM, Sigmund CD. Divergentfunctions of angiotensin II receptor isoforms in the brain. J Clin Invest.2000;106:103–106.
21. Forss-Petter S, Danielson PA, Catsicas S, Battenberg E, Price J,Nerenberg M, Sutcliffe JG. Transgenic mice expressing �-galactosidasein mature neurons under neuron-specific enolase promoter control.Neuron. 1990;5:187–197.
22. Race RE, Priola SA, Bessen RA, Ernst D, Dockter J, Rall GF, Mucke L,Chesebro B, Oldstone MBA. Neuron-specific expression of a hamsterprion protein minigene in transgenic mice induces susceptibility tohamster scrapie agent. Neuron. 1995;15:1183–1191.
23. Mucke L, Masliah E, Johnson WB, Ruppe MD, Alford M, RockensteinEM, Forss-Petter S, Pietropaolo M, Mallory M, Abraham CR. Synap-totrophic effects of human amyloid � protein precursors in the cortex oftransgenic mice. Brain Res. 1994;666:151–167.
24. Murphy TJ, Alexander RW, Griendling KK, Runge MS, Bernstein KE.Isolation of a cDNA encoding the vascular type-1 angiotensin II receptor.Nature. 1991;351:233–236.
25. Sigmund CD. Major approaches for generating and analyzing transgenicmice: an overview. Hypertension. 1993;22:599–607.
26. Davisson RL, Kim HS, Krege JH, Lager DJ, Smithies O, Sigmund CD.Complementation of reduced survival, hypotension, and renal abnor-malities in angiotensinogen-deficient mice by the human renin and humanangiotensinogen genes. J Clin Invest. 1997;99:1258–1264.
27. Davisson RL, Yang GY, Beltz TG, Cassell MD, Johnson AK, SigmundCD. The brain renin-angiotensin system contributes to the hypertension inmice containing both the human renin and human angiotensinogentransgenes. Circ Res. 1998;83:1047–1058.
28. Stec DE, Davisson RL, Haskell RE, Davidson BL, Sigmund CD. Efficientliver-specific deletion of a floxed human angiotensinogen transgene byadenoviral delivery of Cre recombinase in vivo. J Biol Chem. 1999;274:21285–21290.
29. Yang G, Gray TS, Sigmund CD, Cassell MD. The angiotensinogen geneis expressed in both astrocytes and neurons in murine central nervoussystem. Brain Res. 1999;817:123–131.
30. Adams JC. Biotin amplification of biotin and horseradish peroxidasesignals in histochemical stains. J Histochem Cytochem. 1992;40:1457–1463.
31. Franklin KBJ, Paxinos G. The Mouse Brain in Stereotaxic Coordinates.San Diego, Calif: Academic Press, Inc; 1997.
32. Barron KW, Trapani AJ, Gordon FJ, Brody MJ. Baroreceptor denervationprofoundly enhances cardiovascular responses to central angiotensin II.Am J Physiol. 1989;257:H314–H323.
33. Giles ME, Fernley RT, Nakamura Y, Moeller I, Aldred P, Ferraro T,Penschow JD, McKinley MJ, Oldfield BJ. Characterization of a specificantibody to the rat angiotensin II AT1 receptor. J Histochem Cytochem.1999;47:507–516.
34. Leung KH, Chang RS, Lotti VJ, Roscoe WA, Smith RD, TimmermansPB, Chiu AT. AT1 receptors mediate the release of prostaglandins inporcine smooth muscle cells and rat astrocytes. Am J Hypertens. 1992;5:648–656.
35. Rydzewski B, Zelezna B, Tang W, Sumners C, Raizada MK. AngiotensinII stimulation of plasminogen activator inhibitor-1 gene expression inastroglial cells from the brain. Endocrinology. 1992;130:1255–1262.
36. Tallant EA, Diz DI, Ferrario CM. Identification of AT1 receptors oncultured astrocytes. Adv Exp Med Biol. 1996;396:121–129.
37. Bunnemann B, Fuxe K, Ganten D. The renin-angiotensin system in thebrain: an update. Regul Pept. 1993;46:487–509.
Lazartigues et al Brain-Selective AT1a Receptor Overexpression 623
by guest on May 27, 2015http://circres.ahajournals.org/Downloaded from

38. Buccafusco JJ. The role of central cholinergic neurons in the regulation ofblood pressure and in experimental hypertension. Physiol Rev. 1996;48:179–211.
39. Ito M, Oliverio MI, Mannon PJ, Best CF, Maeda N, Smithies O, CoffmanTM. Regulation of blood pressure by the type 1A angiotensin II receptorgene. Proc Natl Acad Sci U S A. 1995;92:3521–3525.
40. Schmechel D, Marangos PJ, Brightman M. Neuron-specific enolase is amolecular marker for peripheral and central neuroendocrine cells. Nature.1978;276:834–836.
41. Fattori E, Lazzaro D, Musiani P, Modesti A, Alonzi T, Ciliberto G. IL-6expression in neurons of transgenic mice causes reactive astrocytosis andincrease in ramified microglial cells but no neuronal damage. EurJ Neurosci. 1995;7:2441–2449.
42. Sakimura K, Araki K, Kushiya E, Takahashi Y. Partial purification andcharacterization of messenger RNA coding 14-3-2 protein from rat brain.J Neurochem. 1982;39:366–370.
43. Tagawa T, Dampney AL. AT1 receptors mediate excitatory inputs torostral ventrolateral medulla pressor neurons from hypothalamus. Hyper-tension. 1999;34:1301–1307.
44. Toney GM, Porter JP. Functional role of brain AT1 and AT2 receptors inthe central angiotensin II pressor response. Brain Res. 1993;603:57–63.
45. Meng H, Wielbo D, Gyurko R, Phillips MI. Antisense oligonucleotide toAT1 receptor mRNA inhibits central angiotensin induced thirst and vaso-pressin. Regul Pept. 1994;54:543–551.
46. Bishop VS, Hay M. Involvement of the area postrema in the regulation ofsympathetic outflow to the cardiovascular system. Front Neuroendo-crinol. 1993;14:57–75.
47. Castren E, Saavedra JM. Repeated stress increases the density of angio-tensin II binding sites in rat paraventricular nucleus and subfornicalorgan. Endocrinology. 1988;122:370–372.
48. Saavedra JM. Emerging features of brain angiotensin receptors. RegulPept. 1999;85:31–45.
49. Johren O, Sanvitto GL, Egidy G, Saavedra JM. Angiotensin II AT1A
receptor mRNA expression is induced by estrogen-progesterone in dopa-minergic neurons of the female rat arcuate nucleus. J Neurosci. 1997;17:8283–8292.
50. Johren O, Saavedra JM. Expression of AT1A and AT1B angiotensin IIreceptor messenger RNA in forebrain of 2-wk-old rats. Am J Physiol.1996;271:E104–E112.
624 Circulation Research March 22, 2002
by guest on May 27, 2015http://circres.ahajournals.org/Downloaded from
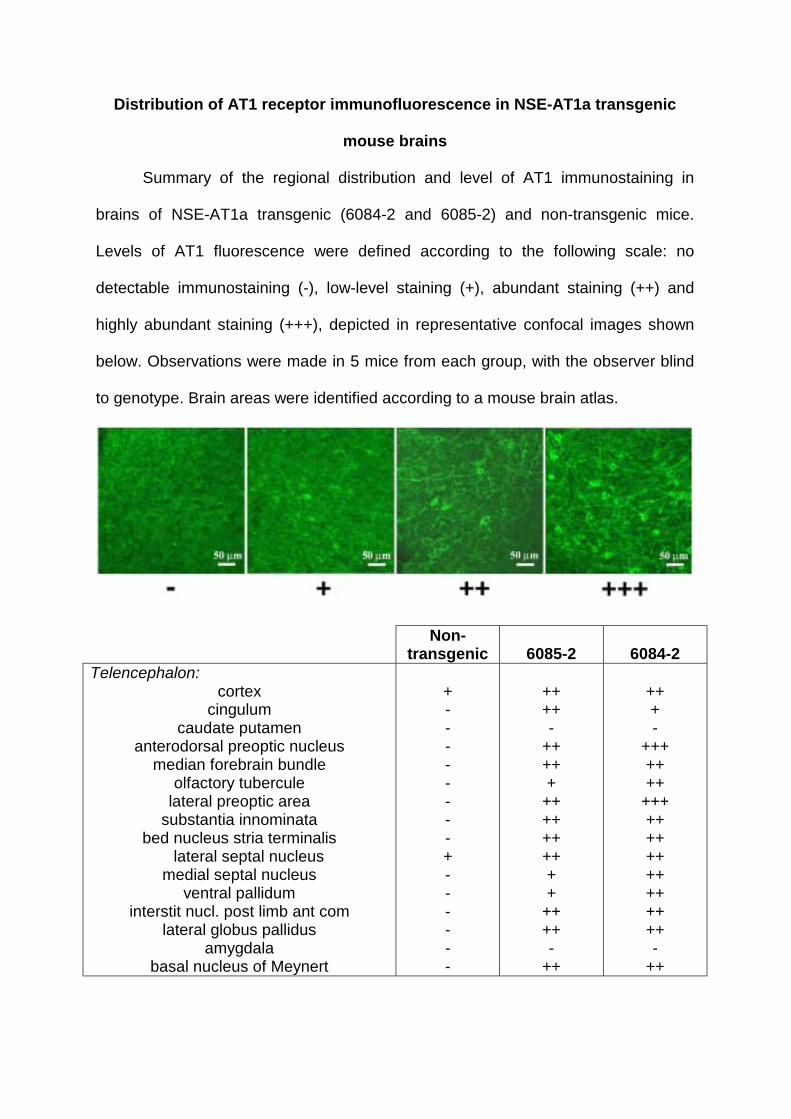
Distribution of AT1 receptor immunofluorescence in NSE-AT1a transgenic
mouse brains
Summary of the regional distribution and level of AT1 immunostaining in
brains of NSE-AT1a transgenic (6084-2 and 6085-2) and non-transgenic mice.
Levels of AT1 fluorescence were defined according to the following scale: no
detectable immunostaining (-), low-level staining (+), abundant staining (++) and
highly abundant staining (+++), depicted in representative confocal images shown
below. Observations were made in 5 mice from each group, with the observer blind
to genotype. Brain areas were identified according to a mouse brain atlas.
Non-transgenic 6085-2 6084-2
Telencephalon: cortex
cingulum caudate putamen
anterodorsal preoptic nucleus median forebrain bundle
olfactory tubercule lateral preoptic area
substantia innominata bed nucleus stria terminalis
lateral septal nucleus medial septal nucleus
ventral pallidum interstit nucl. post limb ant com
lateral globus pallidus amygdala
basal nucleus of Meynert
+ - - - - - - - - + - - - - - -
++ ++ -
++ ++ +
++ ++ ++ ++ + +
++ ++ -
++
++ + -
+++ ++ ++
+++ ++ ++ ++ ++ ++ ++ ++ -
++

Diencephalon: anteroventral periventricular nucleus
ventromedial preoptic nucleus paraventricular thalamic nucl., ventral
suprachiasmatic nucleus medial accessory optic tract reticular thalamic nucleus
mediodorsal thalamic nucleus reuniens thalamic nucleus
paravent. hypothalamic nucl., magnocell paravent. hypothalamic nucl., parvicell periventricular hypothalamic nucleus
lateral hypothalamic area lateroanterior hypothalamic nucleus
arcuate hypothalamic nucleus supraoptic nucleus
central medial thalamic nucleus hypocampus
- + + + - - - + + + - + + + - + +
+++ +++ +++ +++ +++ ++ + +
++ +++
+ ++ ++ ++ ++ ++ +
+++ +++ +++ +++ +++ ++
+++ ++
+++ +++ +++ +++ +++ ++
+++ ++ +
Pons-Midbrain: dorsal raphe nucleus
median longitudinal fasciculus pontine reticular nucleus median raphe nucleus
principal sensory trigeminal nucleus parvicellular nucleus trigeminal nerve
motor root trigeminal nerve periolivary nuclei
rostral periolivary region nucleus lateral lemniscus
laterodorsal tegmental nucleus subpeduncular tegmental nucleus
cerebellum
+ - + + - - - - - - + - -
++ ++ + +
++ ++ ++ + + + + + -
++ +++ ++ ++ ++ ++ ++ ++ ++ ++
+++ +++
-
Medulla: nucleus tractus solitary
dorsomedial spinal trigeminal nucleus ventral cochlear nucleus paratrigeminal nucleus
dorsal motor nucleus vagus hypoglossal nucleus
nucleus of Roller raphe magnus raphe obscurus
rostroventrolateral medulla caudal ventrolateral medulla
reticular nuclei (intermediate, lateral) nucleus ambiguus
+ - + - - - - + + + + - +
+++ +++ ++ ++ ++ +
++ ++ ++
+++ ++ ++
+++
+++ ++
+++ ++
+++ ++ ++ ++ ++
+++ ++ ++ ++

Circumventricular organs: subfornical organ
vascular organ lamina terminalis median preoptic area
median eminence area postrema
+ + + + +
+++ +++ ++
+++ ++
+++ +++ +++ +++ ++

J. Espelund, Curt D. Sigmund and Robin L. DavissonEric Lazartigues, Shannon M. Dunlay, Angela K. Loihl, Puspha Sinnayah, Julie A. Lang, Joshua
Cardiovascular Sensitivity in Transgenic Mice) Receptors Causes Enhanced1Brain-Selective Overexpression of Angiotensin (AT
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2002 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.0000012460.85923.F02002;90:617-624; originally published online February 7, 2002;Circ Res.
http://circres.ahajournals.org/content/90/5/617World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2002/03/19/90.5.617.DC1.htmlData Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on May 27, 2015http://circres.ahajournals.org/Downloaded from
Copyright © 2022 FDOKUMEN




















