
Transcranial magnetic stimulation studies in Alzheimer's disease

Brain insulin and insulin receptors 423J Neural Transm (1998) 105: 423–438
Brain insulin and insulin receptors in aging andsporadic Alzheimer’s disease
L. Frölich1,2, D. Blum-Degen2, H.-G. Bernstein3, S. Engelsberger2,J. Humrich2, S. Laufer2, D. Muschner2, A. Thalheimer2, A. Türk2,
S. Hoyer4, R. Zöchling5, K. W. Boissl5, K. Jellinger6, and P. Riederer2
1 Department of Psychiatry I, University of Frankfurt/Main, 2 Department ofPsychiatry, University of Würzburg, 3 Department of Psychiatry, University of
Magdeburg, 4 Department of Pathochemistry and General Neurochemistry,University of Heidelberg, Federal Republic of Germany
5 NÖ Landesnervenklinik, Mauer/Amstetten, and 6 L. Boltzmann Instituteof Clinical Neurobiology, Vienna, Austria
Accepted November 11, 1997; received August 4, 1997
Summary. The search for the causes of neurodegenerative disorders is a majortheme in brain research. Acquired disturbances of several aspects of cellularmetabolism appear pathologically important in sporadic Alzheimer’s disease(SDAT). Among these brain glucose utilisation is reduced in the early stagesof the disease and the regulatory enzymes important for glucose metabolismare reduced. In the brain, insulin, insulin-like growth factors and their recep-tors regulate glucose metabolism and promote neuronal growth. To detectchanges in the functional activity of the brain insulin neuromodulatory systemof SDAT patients, we determined the concentrations of insulin and c-peptideas well as insulin receptor binding and IGF-I receptor binding in severalregions of postmortem brain cortex during aging and Alzheimer’s disease.Additionally, we performed immunohistochemical staining with antibodiesagainst insulin in neocortical brain areas in SDAT and controls. We show forthe first time that insulin and c-peptide concentration in the brain are corre-lated and decrease with aging, as do brain insulin receptor densities. Weakinsulin-immunoreactivity could be demonstrated histochemically in pyrami-dal neurons of controls, whereas in SDAT a stronger insulin-immunoreac-tivity was found. On a biochemical level, insulin and c-peptide levels werereduced compared to middle-aged controls, but were unchanged compared toage-matched controls. Brain insulin receptor densities in SDAT were de-creased compared to middle-aged controls, but increased in comparison toage-matched controls. IGF-I receptor densities were unchanged in aging andin SDAT. Tyrosine kinase activity, a signal transduction mechanism commonto both receptor systems, was reduced in SDAT in comparison to middle-aged

424 L. Frölich et al.
and age-matched control groups. These data are consistent with aneurotrophic role of insulin in the human brain and a disturbance of insulinsignal transduction in SDAT brain and favor the hypothesis that insulindependent functions may be of pathogenetic relevance in sporadic SDAT.
Keywords: Insulin, insulin receptor, Alzheimer’s disease, aging.
Introduction
Alzheimer’s disease (SDAT) is one of the most common neurodegenerativedisorders in aging. The disease is characterized by abnormalities in severalneurotransmitter systems (Gsell et al., 1993). Disturbances in the cholinergic,serotoninergic, noradrenergic and dopaminergic systems are affectedin SDAT. These transmitter changes correspond inversely to the neurotrans-mitter changes in Parkinson’s disease (Riederer et al., 1990). Additionally,several neuropeptides and amino acid neurotransmitters, e.g. glutamate,are also affected (Ferrier et al., 1993; Beal et al., 1985; Higgins and Mufson,1989; Chalmers et al., 1990). A loss of cholinergic neurons in the basal fore-brain is predominant and correlates with the volume of cortical gray matter(Cullen et al., 1997). These neurochemical changes may relate to abnormali-ties in glucose metabolism in vivo (Hoyer, 1986). Consistent reductions in theregional cerebral metabolic rate of glucose have been identified using positronemission tomography (Fukuyama et al., 1994; Haxby et al., 1986; Herholz,1995). The diminished glucose utilisation was not attributed to an insuffi-cient supply of glucose to the brain, but rather to diminished glucose break-down in brain tissue (Hoyer et al., 1988, 1991). Correspondingly, the activitiesof number of enzymes involved in glycolytic and oxidative glucose metabo-lism are reduced in post-mortem brain tissue of SDAT (Bowen et al., 1976;Iwangoff et al., 1980; Perry et al., 1980; Gibson et al., 1988; Frölich et al., 1997).These reductions appear to be more pronounced than the reductions inbiochemical variables previously related to brain atrophy (Bowen et al.,1979).
The existence of both insulin and insulin receptors in the CNS is wellestablished (Baskin et al., 1988; Potau et al., 1991; Wozniak et al., 1993).During protein processing insulin is released from proinsulin by proteolyticcleavage; C-peptide is also released in equimolar amounts (Steiner et al.,1985). Insulin was first localized in the CNS of rats by immunohistochemicalstaining (Havrankova et al., 1978b). Later insulin mRNA was demonstratedin various brain areas, suggesting the peptide is synthesized in the brain(Young, 1986; Marks et al., 1990; Devaskar et al., 1994). In spite of this, theorigin of brain insulin remains controversial. In the human brain, insulinregulates enzymes of cerebral glucose metabolism (Hoyer et al., 1993) viaspecific high-affinity insulin receptors (Havrankova et al., 1978a), which differfrom peripheral insulin receptors only in the amount of glycosylation (Baskinet al., 1988; Wozniak et al., 1993; de Pablo and de la Rosa, 1995). Additionally,insulin binds to insulin-like growth factor I receptors and, via these receptors,possibly exerts more generalized trophic effects on neuronal cells and inter-acts with cholinergic neurotransmission (Calissano et al., 1993; Quirion et al.,

Brain insulin and insulin receptors 425
1991; Rotwein, 1991; Kyriakis et al., 1987). In addition to its regulatory func-tion in the maintenance of brain glucose metabolism and its trophic actions,insulin regulates food intake (Wozniak et al., 1993). The binding of insulin toits receptor is followed by an autophosphorylation of tyrosine residues at the�-chain of the insulin receptor resulting in a subsequent activation of theintrinsic tyrosine kinase, which phosphorylates the first known endogenoussubstrate IRS-1 (insulin-receptor-substrate-1) (Sun et al., 1993). Phosphory-lated IRS-1 transfers the signal to a wide spectrum of cellular signal transduc-tion pathways, which are not yet fully understood (for review see White andKahn, 1994).
In Alzheimer’s disease, it is posited that defects in regulation of glucosemetabolism, i.e. resulting from changes in CNS insulin receptor function orsubsequent insulin signal transduction, might be an early contributing event tothe onset of SDAT (Blum-Degen et al., 1995; Henneberg and Hoyer, 1995;Hoyer, 1996).
In an experimental study on human post-mortem brain tissue we haveinvestigated whether the immunoreactivity of insulin and c-peptide in thebrain change with aging and, whether the respective levels in SDAT differfrom those in two control groups, a) a middle-aged adult group and b) an age-matched elderly group. We have also investigated, whether the densities andaffinities of the insulin and insulin-like growth factor I receptors change withaging and whether the receptor densities in SDAT differ from controls. Theactivity of tyrosine kinase was measured using ELISA and related to normalaging, these values were compared with those of SDAT and control individu-als. Finally, we have investigated whether the immunohistochemical stainingpattern of neocortical areas with antibodies against insulin differs qualita-tively between SDAT and age-matched controls.
Material and methods
Patients
A clinical diagnosis of dementia was made according to DSM-IIIR and included a mentalstatus examination. All dementia patients had been hospitalized for long periods untildeath and all were severely demented. A full medical examination of each patient wasperformed to exclude metabolic causes of dementia. The patients fulfilled the diagnosticcriteria of the NINCDS/ADRDA for probable SDAT (McKhann et al., 1984). Controlpatients without a history of neurological or psychiatric disease were hospitalized anddied from somatic disorders. The sample consisted of 17 patients with SDAT (mean age:79.8 � 2.0, age range: 67–91 years) and 21 controls (mean age: 64.5 � 5.1), age range: 21–93 years). From the total number of controls, two groups were selected: a) a middle-agedgroup of 8 subjects (mean age: 39.1 � 5.6, age range: 21–62 years) and b) an age-matchedgroup of 13 subjects (mean age: 80.1 � 2.5, age range: 68–93). Sex distribution was 8/9(male/female) in the SDAT group and 10/11 in the control group. Post mortem delay was24.7 � 4.1 hours for the SDAT group, 37.8 � 3.7 for the middle-aged controls, and 25.4� 4.0 hours for the age-matched control group (range 4 to 50 hours). Brain lactate contentwas 20.7 � 1.5 mg/dl in the SDAT group, 14.1 � 3.2 mg/dl in the middle-aged controls, and21.7 � 1.4 mg/dl in the age-matched control group. All brains were stored in the German-Austrian Reference Center for Neurodegenerative Disorders at the Department ofPsychiatry, University of Würzburg.

426 L. Frölich et al.
A second set of brains from the Department of Psychiatry, University of Magdeburgwere obtained for immunocytochemical analysis of insulin-containing neurons. For theseinvestigations, autopsied neocortical brain specimens were obtained from clinically de-mented and neuropathologically confirmed patients with dementia of Alzheimer type andneurologically normal subjects. The sample consisted of 5 patients with SDAT (mean age:72.4 � 5.1, age range: 65–78 years) and 4 controls (mean age: 73.8 � 5.1, age range: 69–81 years). Sex distribution was 2/3 (male/female) in the SDAT group and 1/3 in thecontrol group. Post mortem delay was 10.4 � 4.1 hours for the SDAT group, and 11.3 �4.0 hours for the age-matched control group (range 9 to 13 hours).
Human brain tissue
Brains were removed at autopsy and divided by mid-sagittal section following the guide-lines for human brain dissection (Pope, 1982) and in accordance with the guidelines set bythe responsible ethics committees (University of Würzburg and University of Magdeburg,respectively). One hemisphere was fixed in buffered formaldehyde solution for histologi-cal examination. The final diagnosis of SDAT was established histologically by the pres-ence of numerous plaques and tangles in the cerebral isocortex and hippocampus in theabsence of considerable vascular lesions. In control brains, the macroscopic examinationof the brain did not show any pathological changes, and the histological examination ofthe cerebral cortex and the hippocampus did not show more senile plaques and tanglesthan those attributable to age (Khachaturian, 1985; Tierney et al., 1988).
For neurochemical analysis, one hemisphere was deep-frozen in dry ice at autopsyand then stored at �80°C. The frozen hemispheres were partially thawed to �10°C fordissection. Brain tissue from the four cortical lobes was obtained using an anatomical atlas(Nieuwenhuys et al., 1990) and followed a standard procedure (Gsell et al., 1993).
For immunocytochemistry, samples from prefrontal gyrus, pre- and postcentral gyrusand angular gyrus were removed from the brains as quickly as possible.
Immunohistochemical analysis
The samples were cut into small pieces of approximately 1cm3. These blocks were rinsedwith phosphate-buffered saline (PBS, pH 7.4, 0.05M) and fixed by immersion in ice-coldparaformaldehyde solution (4% in PBS). After a fixation time of 24–72 hours, the brainblocks were embedded in paraffin and cut into 6µm serial sections. Each fifth section wasstained for hematoxylin and eosin. The cases were coded and randomized over thedifferent experimental protocols.
Dewaxed sections were subjected to immunohistochemistry using a monoclonal anti-body against insulin raised in Balb/c mice (ascites) as described earlier (Dorn et al., 1983).This monoclonal anti-insulin antibody recognizes specifically the polypeptide in humanbrain tissue (Dorn et al., 1984). The antibody (protein concentration in the ascites: 10mg/ml) was used as primary immunoreagent at a dilution of 1 :100 in PBS. Sections wereincubated overnight at 4°C in a moist chamber. After repeated rinsing in PBS the sectionswere incubated with goat anti-mouse IgG serum (SIFIN), rinsed again and treated withmonoclonal peroxidase-antiperoxidase complex (mouse, DAKO). 3,3�-diaminobenzidineserved as chromogene to visualize the immunoreaction. Controls involved the omission ofthe primary antibody or its substitution by either buffer or ascites. All control sectionswere free of immunostaining.
Biochemical analysis
Approximately 500mg–2000 mg of the frozen tissue was pulverized in a porcelain mortarunder liquid N2. For radioimmuno-assays, this tissue was homogenized at 4°C in a motor-driven glass/teflon potter (1000/min, 40 up and down strokes) in 9 volumes of mediumcontaining 10 mM MOPS (pH 7.6), 120mM NaCl, 1 mM EDTA, 0.1 mM benzethonium
Brain insulin and insulin receptors 427
chloride, 1mM benzamidine and 0.1% trasylol. For radioligand binding assays, this tissuewas homogenized in MOPS/sucrose buffer and washed twice. The homogenates werestored at �80°C and assayed within 8 weeks.
Quantification of insulin and the corresponding c-peptide was done with commer-cially available RIA-kits (radio adsorbent assay, manufactured by Biermann Diagnostics,Bad Nauheim, Germany).
Radioligand binding assays were performed as “cold” saturation assays with 125J-insulin or 125J-IGF-I, respectively. 10�10M tracer were displaced by increasing amounts (9steps) of cold ligand, unspecific binding was determined by 10�7M cold ligand. Separationof bound and free ligand was achieved by centrifugation. Preliminary estimations ofbinding parameters (Bmax and KD values) for both receptors were calculated with the PCsoftware “EBDA”. Final values were calculated with the PC software “LIGAND”
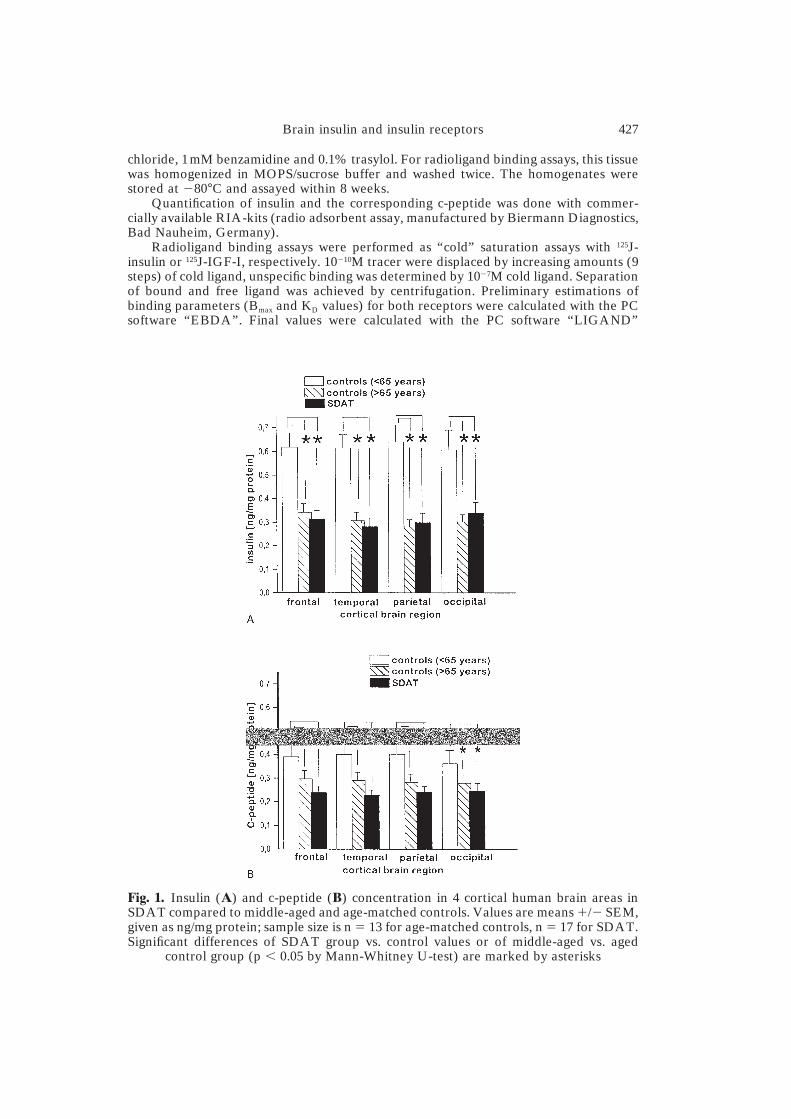
Fig. 1. Insulin (A) and c-peptide (B) concentration in 4 cortical human brain areas inSDAT compared to middle-aged and age-matched controls. Values are means �/� SEM,given as ng/mg protein; sample size is n � 13 for age-matched controls, n � 17 for SDAT.Significant differences of SDAT group vs. control values or of middle-aged vs. aged
control group (p � 0.05 by Mann-Whitney U-test) are marked by asterisks
428 L. Frölich et al.
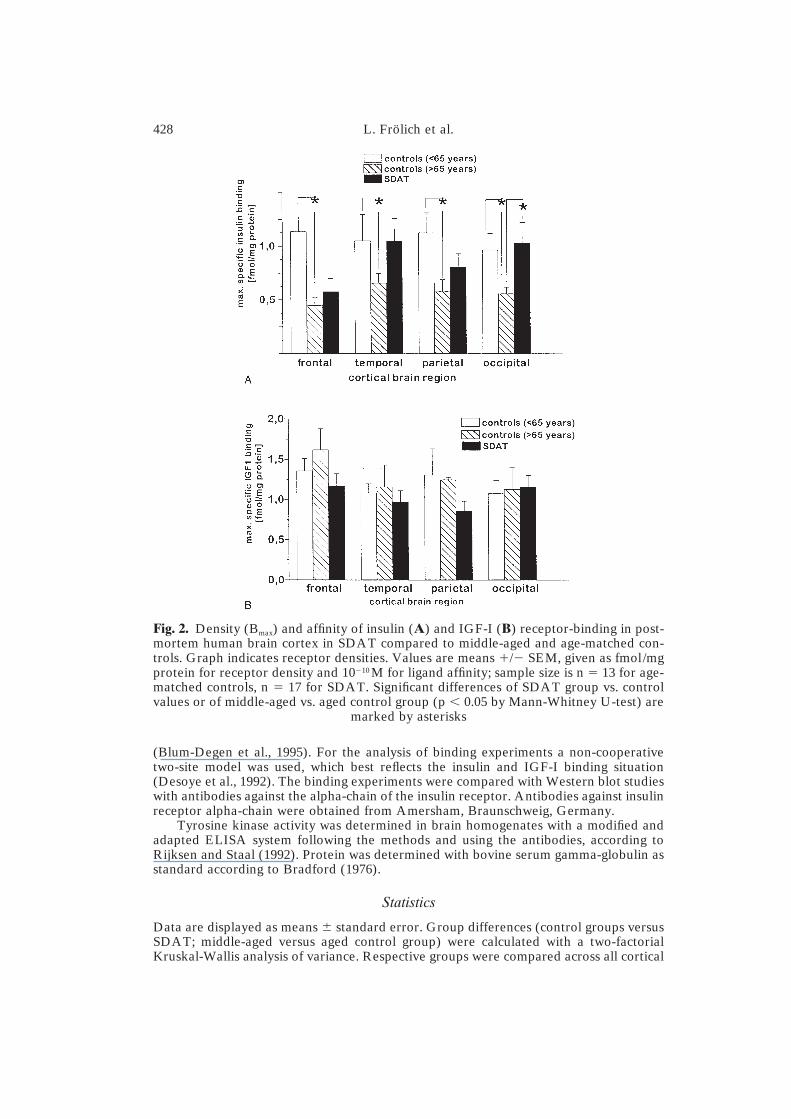
Fig. 2. Density (Bmax) and affinity of insulin (A) and IGF-I (B) receptor-binding in post-mortem human brain cortex in SDAT compared to middle-aged and age-matched con-trols. Graph indicates receptor densities. Values are means �/� SEM, given as fmol/mgprotein for receptor density and 10�10 M for ligand affinity; sample size is n � 13 for age-matched controls, n � 17 for SDAT. Significant differences of SDAT group vs. controlvalues or of middle-aged vs. aged control group (p � 0.05 by Mann-Whitney U-test) are
marked by asterisks
(Blum-Degen et al., 1995). For the analysis of binding experiments a non-cooperativetwo-site model was used, which best reflects the insulin and IGF-I binding situation(Desoye et al., 1992). The binding experiments were compared with Western blot studieswith antibodies against the alpha-chain of the insulin receptor. Antibodies against insulinreceptor alpha-chain were obtained from Amersham, Braunschweig, Germany.
Tyrosine kinase activity was determined in brain homogenates with a modified andadapted ELISA system following the methods and using the antibodies, according toRijksen and Staal (1992). Protein was determined with bovine serum gamma-globulin asstandard according to Bradford (1976).
Statistics
Data are displayed as means � standard error. Group differences (control groups versusSDAT; middle-aged versus aged control group) were calculated with a two-factorialKruskal-Wallis analysis of variance. Respective groups were compared across all cortical

Brain insulin and insulin receptors 429
Fig. 3. Tyrosine kinase (TrK) activity in various post-mortem human brain areas inSDAT compared to middle-aged and age-matched controls. Values are means �/� SEM,given as U/mg homogenate for TrK activity; sample size is n � 13 for age-matchedcontrols, n � 17 for SDAT. Significant differences of SDAT group vs. control values or ofmiddle-aged vs. aged control group (p � 0.05 by Mann-Whitney U-test) are marked by
asterisks
brain regions for overall significant differences. Subsequently, Mann-Whitney U-testswere used for calculating separate group differences for each of the four cortical brainregions. A significance level of p � 0.05 was accepted. Calculations were performed withthe SPSS-PC package.
Results
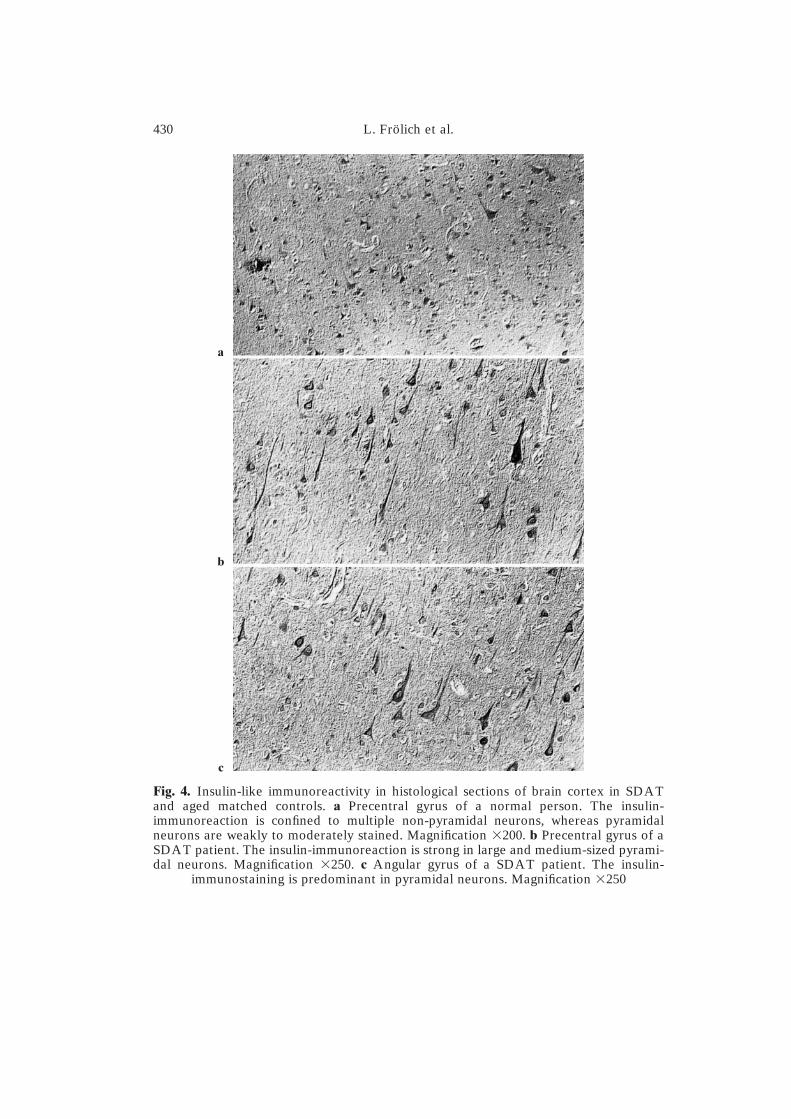
In the normal brains, insulin immunoreactivity could be detected in manypyramidal and non-pyramidal neurons. The intensity of immunostaining wasweak. The reaction product was perinuclearly distributed. Insulin-containingcell processes were observed in a few layer V pyramidal neurons (Fig. 4a).In SDAT brains, layers III and V pyramidal neurons were characterized bystrong insulin immunoreactivity labeling perikarya and many dendrites (Fig.4b), while non-pyramidal neurons were only weakly stained. There was noincrease in the total amount of immunoreactive neurons in SDAT patients incomparison with normal subjects. Glial and endothelial cells were always freeof immunoreaction (Fig. 4c). Very few neuritic plaques were found to reactwith the insulin antibody.
Western blot studies for the insulin receptor demonstrated the existenceof insulin receptor alpha-chain in the post-mortem human brain, both innormal and Alzheimer-diseased brains (Fig. 5).
On the biochemical level, both insulin and c-peptide could be demon-strated reliably in human post-mortem brain tissue. There were no significantregional differences in the insulin concentration between the four corticalbrain regions. Insulin and c-peptide levels, however, were correlated signifi-cantly with each other (r � 0.294, p � 0.001, data not shown). The variabilityof insulin and c-peptide between the regions of an individual brain (coefficientof variation for insulin: 25.2% in SDAT, 23.7% for controls, coefficient of
430 L. Frölich et al.
Fig. 4. Insulin-like immunoreactivity in histological sections of brain cortex in SDATand aged matched controls. a Precentral gyrus of a normal person. The insulin-immunoreaction is confined to multiple non-pyramidal neurons, whereas pyramidalneurons are weakly to moderately stained. Magnification �200. b Precentral gyrus of aSDAT patient. The insulin-immunoreaction is strong in large and medium-sized pyrami-dal neurons. Magnification �250. c Angular gyrus of a SDAT patient. The insulin-
immunostaining is predominant in pyramidal neurons. Magnification �250
a
b
c

Brain insulin and insulin receptors 431
Fig. 5. Western blot analysis of the alpha-subunit of insulin receptor (molecular weight:appr. 130kD) in the human brain. Data show various regions from one control brainwithout neuropsychiatric disorders (upper part) and one Alzheimer brain (lower part).Both films document a band correponding to the molecular weight of the alpha-subunit of
the insulin receptor in all brain regions except from white matter
variation for c-peptide: 20.1% in SDAT, 15.6% in controls, data not shown)was much lower than the variability between the different brains (coefficientof variation for insulin: 42.7% in SDAT, 35.3% for controls, coefficient ofvariation for c-peptide: 39.7% in SDAT, 41.3% in controls, data not shown),indicating that the insulin content in an individual brain reflects a biologicalconstant. In normal aging, both insulin and c-peptide decreased significantlyin all cortical brain regions with advancing age, which is demonstrated by areduction of the mean values of the middle-aged versus the aged controlgroup by 31–45% (depending on the cortical brain region) for insulin and 23–

432 L. Frölich et al.
30% (depending on the cortical brain region) for c-peptide (Fig. 1a and b).Insulin receptor densities decreased with advancing age only in frontal(�49%) and parietal (�38%) cortical brain regions, but not in temporal(100%) and occipital (101%) brain cortices (Fig. 2a) without significantchanges in ligand affinity (data not shown). These age-related differenceswere statistically significant for the frontal and parietal cortex. In contrast, thestructurally related IGF-I receptor did not show decreased receptor densitieswith aging (Fig. 2b) nor changes in ligand affinity (data not shown), whichdemonstrated some specificity of the changes in the insulin/insulin receptorsystem. Tyrosine kinase activity, the common signal transduction systemfor both insulin and IGF-I receptors, also decreased with aging, which isdemonstrated by significant reductions of the mean values of the middle-agedversus the aged control group by 31–58% (depending on the brain region)(Fig. 3).
In senile dementia of Alzheimer type, insulin and c-peptide levels in allfour regions of brain cortex showed a significant reduction compared tomiddle-aged controls (p � 0.05 in all cortical brain regions) (Fig. 1a and b),but did not differ significantly from the age-matched control group.
Insulin receptor densities were unchanged in SDAT, when compared tothe middle-aged control group, but increased compared to the age-matchedcontrols, most pronounced and significant in the occipital cortex (p � 0.05)(Fig. 2a). Insulin receptor ligand affinities did no differ significantly from theircontrol values (data not shown). These changes may indicate a compensatoryup-regulation of receptor number in SDAT, possibly as a response toimpaired signal transduction. Neither IGF-I receptor densities (Fig. 2b)nor IGF-1 receptor ligand affinities (data not shown) differed from theirrespective control values.
Tyrosine kinase activity in SDAT was reduced in all cortical regionsstudied, compared to both control groups. This loss of activity was significantin temporal and occipital cortex (p � 0.05) in comparison to the middle-agedcontrol group, and significant for occipital cortex only (p � 0.05) in compari-son to the age-matched control group (Fig. 3).
Discussion
Insulin and c-peptide are derived from a common precursor, proinsulin, fromwhich these peptides are released in equimolar amounts by proteolytic cleav-age (Polonsky and Rubenstein 1984). Only insulin has biological activity andis degraded by proteases. The most important among those proteases, insulindegrading enzyme, also degrades other peptides, e.g. the �-amyloid peptide(Kurochkin and Goto, 1994). It has been demonstrated by cell culture andanimal experiments that insulin in the brain potently affects neuronal glucosemetabolism and cell differentiation (Knusel et al., 1990; Puro and Agardh,1984; Mill et al., 1985; Henneberg and Hoyer, 1994; Hoyer et al., 1993) via themitochondrial citric acid cycle (Bessman et al., 1986). Our data provide fur-ther evidence for a biological role of insulin in the brain. The qualitativeimmunohistochemical studies indicate a regional distribution and cellular

Brain insulin and insulin receptors 433
localization of insulin related material in the cortical neurons of normal sub-jects, confirming earlier observations of insulin in brain neurons (Dorn et al.,1984). C-peptide was present in amounts comparable to those of insulin inbrain homogenates, suggesting the expression of proinsulin in the brain, fromwhich c-peptide and insulin are produced. Uptake of c-peptide from the bloodappears highly unlikely because of the lack of uptake systems for c-peptide atthe blood-brain barrier. Furthermore, our data showed that insulin correlatedsignificantly with c-peptide, also suggesting the production of their commonprecursor in the brain. Finally, there is a lower within sample variabilitythan between sample variability of insulin levels. These four findings togethersuggest that the insulin level in the brain are compartmented and represent abiological constant, consistent with its role of a neurotrophic factor. Majorregional differences in insulin concentration between the four corticalareas investigated were not evident, suggesting a homogeneous distribution ofinsulin in the brain cortex on a biochemical level. According to our immuno-histochemical data this may be attributed to pyramidal neurons of layers IIIand V.
The large differences between middle-aged controls and age-matchedcontrols demonstrated that insulin and c-peptide levels, as well as the densityof insulin receptors, decrease with advancing age. These data will be discussedin detail in a separate publication (Frölich et al., in preparation). Duringhuman ontogenesis, insulin receptors have also been shown to decrease(Potau et al., 1991). Our data show further that a structurally related receptorsystem, the IGF-I receptor, does not decrease during aging. This indicates thatthe changes of the insulin receptor system do not merely reflect an unspecificcellular loss during aging, but indeed may be due to a specific susceptibilityand may thus have biological significance.
The immunohistochemical reaction pattern of insulin in SDAT patientswas similar to that observed in L-ornithine decarboxylase (ODC) immunore-activity in neocortical neurons of demented individuals (Bernstein andMüller, 1995). Since the polyamine-generating enzyme ODC is strongly acti-vated by insulin (Yang et al., 1981; Lau et al., 1987), it is possible that theobserved qualitative impression of an increased cellular insulin-immunoreac-tivity and the noted increase in the ODC/polyamine system (Morrison et al.,1993; Bernstein Müller, 1995) in discrete neocortical nerve cells of SDATpatients are interconnected events reflecting restorative processes operatingon neurotrophic factors. Interestingly, immunohistochemical studies on insu-lin receptors in the substantia nigra have shown a reduced density of thesereceptors in Parkinson’s disease (Moroo et al., 1933), which suggests a differ-ential involvement in neurodegenerative disorders.
In our biochemical studies, samples with histopathologically confirmedAlzheimer’s disease were compared with two control groups, a middle-agedgroup and an age-matched group without neuropsychiatric disorders. Becauseof a close matching of both groups with respect to several important factors,i.e. sex, post-mortem delay, storage time of the brain tissue, and lactatecontent of the brain as a measure of the agonal period any major artifactsregarding our data may be excluded. Since the aging brain is at risk for SDAT

434 L. Frölich et al.
(Hoyer, 1996), i.e. may indeed have reached a ‘subclinical’ threshold of thedisease process leading to clinical SDAT, it is interesting to compare caseswith proven SDAT not only to such age-matched control cases, but alsothe middle-aged cases, which may be regarded as not yet having reached thethreshold for ‘“subclinical” (presymptomatic) SDAT. Compared with thismiddle-aged control group, a dramatic decrease of insulin and c-peptideconcentration could be found in SDAT brain, which was not evident incomparison to age-matched controls. The qualitative impression from thehistopatological data of an increased amount of insulin in SDAT com-pared to the age-matched controls, was not reflected in the biochemical insu-lin measurements of insulin. If this discrepancy was due to a lower sensitivityof the biochemical measurements, which utilized brain homogenates or dueto differences in the sensitivity of the antibodies employed, cannot be deter-mined at present, because of the lack of quantitative histopathological data.
With respect to insulin receptors in SDAT in comparison to the middle-aged control group, there was a slight decrease in receptor densities in frontaland parietal cortices only. In contrast there were no changes in temporal andoccipital cortices. These SDAT-related changes were less pronounced thanthe decrease in insulin receptors seen when the middle-aged control groupwas compared with the aged controls. With respect to insulin receptors inSDAT compared with the age-matched controls, we have shown for the firsttime that the number of insulin receptors are increased in all regions of braincortex, with a significant increase in the occipital cortex. It is suggested thatthis finding is due to an up-regulation of the insulin receptor system in SDAT,counteracting a general loss of insulin receptors with normal aging, possiblybecause of an impaired signal transduction system in SDAT. Tyrosine kinaseactivity represents the major effector system for insulin and IGF-1 receptorsand is reduced in all brain regions in SDAT compared with middle-agedcontrols. In the occipital cortex tyrosine kinase activity is also reduced com-pared with age-matched controls, as previously reported (Shapiro et al., 1991),reflecting an impaired signal transduction.
We have confirmed that IGF-I receptor densities in dementia ofAlzheimer type remain constant, both in comparison with a middle-agedcontrol group and in comparison with an age-matched control group, asshown earlier (De Kayser et al., 1994; Crews et al., 1992). IGF-I receptors arestructurally closely related to insulin receptors (Rotwein, 1991; Becker andRoth, 1990) and exert stimulating effects on cholinergic and dopaminergicneurons in culture (Araujo et al., 1989; Knusel et al., 1990; Quirion et al.,1991). The face that this neurotrophic receptor system is unchanged in SDAT,in contrast to insulin receptors, provides further evidence for a specific in-volvement of brain insulin receptors in the pathogenesis of SDAT.
In conclusion, our data are compatible with the hypothesis that the knowndisturbances in glucose metabolism are, at least in part, due to a reducedactivity of the insulin receptor dependent signal transduction in Alzheimerdisease brain most likely at the level of one of the kinase/phosphatases in-volved (Kyriakis et al., 1992). Thus, this abnormality can be assumed to be ofpivotal significance in the early pathogenesis of SDAT.

Brain insulin and insulin receptors 435
Acknowledgements
This work was supported by a grant from BIOMED I Nr. BMH 1-CT 94-1563.
References
Araujo DM, Lapchak PA, Collier B, Chabot JG, Quirion R (1989) Insulin-like growthfactor-I (somatomedin-C) receptors in the rat brain: distribution and interaction withthe hippocampal cholinergic system. Brain Res 484: 130–138
Baskin DG, Wilcox BJ, Figlewicz DP, Dorsa DM (1988) Insulin and insulin-like growthfactors in the CNS. Trends Neurosci 11: 107–111
Beal MF, Uhl G, Mazurek MF, Kowall N, Martin GB (1986) Somatostatin: alterations inthe central nervous system in neurological disorders. In: Martin GB, Barchas JD(eds) Neuropeptides in neurological and psychiatric disease. Raven, New York, pp215–257
Becker AB, Roth RA (1990) Insulin receptor structure and function in normal andpathological conditions. Ann Rev Med 41: 99–115
Bernstein H-G, Müller M (1995) Increased immunostaining for L-ornithine decarboxy-lase occurs in neocortical neurons of Alzheimer’s disease patients. Neurosci Lett 186:123–126
Bessmann SP, Mohan C, Zaidise I (1986) Intracellular site of insulin action: mitochon-drial Krebs cycle. Biochemistry 83: 5067–5070
Blum-Degen D, Frölich L, Hoyer S, Riederer P (1995) Altered brain glucose meta-bolism as a cause of neurodegenerative disorders? J Neural Transm [Suppl] 46: 139–147
Bowen DM, Smith AB, White P, Davison AN, Goodhart MJ, Curzon G, Iwangoff P,Mayer-Ruge W (1976) Neurotransmitter- related enzymes and indices of hypoxia insenile dementia and other abiotrophies. Brain 99: 459–496
Bowen DM, White P, Spillane JA, Goodhart MJ, Curzon G, Iwangoff P, Mayer-Ruge W,Davison AN (1979) Accelerated ageing or selective neuronal loss as an importantcause of dementia? Lancet i: 11–14
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgramquantities of proteins utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Calissano P, Ciotti MT, Battistini L, Zona C, Angelini A, Merlo D, Mercanti D (1993)Recombinant insulin-like growth factor I exerts a trophic action and confersglutamate sensitivity on glutamate-resistant cerebellar cells. Proc Natl Acad Sci USA90: 8752–8756
Chalmers DT, Dewar D, Graham DI, Brooks DN, McCulloch J (1990) Differentialalterations of cortical glutamatergic binding sites in senile dementia of the Alzheimertype. Proc Natl Acad Sci USA 87: 1352–1356
Crews FT, McElhaney R, Freund G, Ballinger WEJ, Raizada MK (1992) Insulin-likegrowth factor I receptor binding in brains of Alzheimer’s and alcoholic patients.J Neurochem 58: 1205–1210
Cullen KM, Halliday GM, Double KL, Brooks WS, Creasey H, Broe GA (1997) Cell lossin the nucleus basalis is related to regional cortical atrophy in Alzheimer’s disease.Neuroscience 78: 641–652
DeKayser J, Wilczak N, Goosens A (1994) Insulin-like growth factor-1 receptor densitiesin human frontal cortex and white matter during aging, in Alzheimer’s disease, and inHuntington’s disease. Neurosci Lett 172: 93–96
dePablo F, de la Rosa E (1995) The developing CNS: a scenario for the action ofproinsulin, insulin and insulin-like growth factors. Trends Neurosci 18: 143–150
Desoye G, Schmon B, Gmoser G, Friedl H, Urdl W, Weiss PAM (1992a) Insulin bindingto erythrocytes of nonpregnant women: a reevaluation, underlining the importance ofbody weight even in nonobese subjects. Clin Chim Acta 207: 57–71

436 L. Frölich et al.
Devaskar SU, Giddings SJ, Rajakumar PA, Carnaghi LR, Menon RK, Zahm DS (1994)Insulin gene expression and insulin synthesis in mammalian neuronal cells. J BiolChem 269: 8445–8454
Dorn A, Ziegler M, Bernstein H-G, Dietz H, Rinne A (1983) Introducing a monoclonalantibody to insulin: the islets of Langerhans as a model for immunohistochemistry.Acta Histochem 73: 293–295
Dorn A, Ziegler M, Bernstein H-G, Dietz H, Rinne A (1984) Concerning the presence ofinsulin-related peptide in the human brain: an immunohistochemical reinvestigationby use of monoclonal insulin antibodies. Acta Histochem 74: 81–84
Ferrier IN, Cross AJ, Johnson JA, Roberts GW, Crow TJ, Corsellis JA, Lee YC,O’Shaughnessy D, Adrian TE, McGregor GP, et al (1983) Neuropeptides inAlzheimer type dementia. J Neurol Sci 62: 159–170
Frölich L, Weinmueller M, Barth N, Goetz N, Dirr A, Gsell W, Jellinger K, Beckmann H,Riederer P (1997) (r)-, but not (s)-alpha lipoic acid stimulates deficient brain pyruvatedehydrogenase complex in vascular dementia, but not in Alzheimer dementia. ArchGerontol Geriatr (in press)
Fukuyama H, Ogawa M, Yamauchi H, Yamaguchi S, Kimura J, Yonekura Y, Konishi J(1994) Altered cerebral energy metabolism in Alzheimer’s disease: a PET study.J Nucl Med 35: 1–6
Gibson GE, Sheu KF, Blass JP, Baker A, Carlson KC, Harding B, Perrino P (1988)Reduced activities of thiamine-dependent enzymes in the brains and peripheraltissues of patients with Alzheimer’s disease. Arch Neurol 45: 836–840
Gsell W, Moll G, Sofic E, Riederer P (1993) Cholinergic and monoaminergic neurotrans-mitter systems in patients with Alzheimer’s disease and senile dementia of Alzheimertype: a critical evaluation. In: Maurer K (ed) Dementias: neurochemistry, neuro-pathology, neuroimaging, neuropsychology and genetics. Vieweg, Braunschweig,pp 25–51
Gsell W, Lange KW, Reuffer R, Heckers S, Heinsen H, Senitz D, Jellinger K, RansmayrG, Wichart I, Vock R, Beckmann H, Riederer P (1993) How to run a brain bank.A report from the Austro-German brain bank. J Neural Transm [Suppl] 39: 31–70
Havrankova J, Roth J (1978a) Insulin receptors are widely distributed in the centralnervous system of the rat. Nature 272: 827–829
Havrankova J, Schmechel D, Roth J, Brownstein M (1978b) Identification of insulin in ratbrain. Proc Natl Acad Sci USA 75: 5737–5741
Haxby JV, Grady CL, Duara R, Schlageter N, Berg G, Rapoport SI (1986) Neocorticalmetabolic abnormalities precede nonmemory cognitive defects in early Alzheimer’stype dementia. Arch Neurol 43: 882–885
Henneberg N, Hoyer S (1994) Short-term or long-term intracerebroventricular (i.c.v.)infusion of insulin exhibits a discrete anabolic effect on cerebral energy metabolismin the rat. Neurosci Lett 175: 153–156
Henneberg N, Hoyer S (1995) Desensitisation of the neuronal insulin receptor: a newapproach in the etiopathogenesis of late-onset sporadic dementia of the Alzheimertype. Arch Gerontol Geriatr 21: 63–74
Herholz K (1995) FDG PET and differential diagnosis of dementia. Alzheimer Dis AssocDisord 9: 6–16
Higgins GA, Mufson EJ (1989) NGF receptor gene expression is decreased in the nucleusbasalis in Alzheimer’s disease. Exp Neurol 106: 222–236
Hoyer S (1986) Senile dementia and Alzheimer’s disease. Brain blood flow and metabo-lism. Prog Neuropsychopharmacol Biol Psychiatry 10: 447–478
Hoyer S (1996) Oxidative metabolism deficiencies in brains of patients with Alzheimer’sdisease. Acta Neurol Scand [Suppl 165]: 18–24
Hoyer S, Oesterreich K, Wagner O (1988) Glucose metabolism as the site of the primaryabnormality in early-onset dementia of Alzheimer type? J Neurol 235: 143–148
Hoyer S, Nitsch RM, Oesterreich K (1991) Predominant abnormality in cerebral glucoseutilisation in late-onset dementia of the Alzheimer type: a cross-sectional comparison

Brain insulin and insulin receptors 437
against advanced late-onset and incipient early-onset cases. J Neural Transm [P-DSect] 3: 1–14
Hoyer S, Prem L, Sorbi S, Amaducci L (1993) Stimulation of glycolytic key enzymes incerebral cortex by insulin. Neuroreport 4: 991–993
Iwangoff P, Armbruster R, Enz A, Mayer-Ruge W (1980) Glycolytic enzymes fromhuman autoptic brain cortex: normal aged and demented cases. Mech Ageing Dev 14:203
Khachaturian Z (1985) Diagnosis of Alzheimer’s disease. Arch Neurol 42: 1097–1105Knusel B, Michel PP, Schwaber JS, Hefti F (1990) Selective and nonselective stimulation
of central cholinergic and dopaminergic development in vitro by nerve growth factor,basic fibroblast growth factor, epidermal growth factor, insulin and the insulin-likegrowth factors I and II. J Neurosci 10: 558–570
Kurochkin IV, Goto S (1994) Alzheimer’s �-amyloid peptide specifically interacts withand is degraded by insulin degrading enzyme. FEBS Lett 345: 33–37
Kyriakis JM, Hausman RE, Peterson SW (1987) Insulin stimulates choline acetyltrans-ferase activity in cultured embryonic chicken retina neurons. Proc Natl Acad Sci USA84: 7463–7467
Lau C, Bartolome JV, Bartolome MB, Slotkin TA (1987) Central and sympathoadrenalresponses to insulin in adult and neonatal rats. Dev Brain Res 36: 277–280
Marks JL, Porte D Jr, Stahl WL, Baskin DG (1990) Localization of insulin receptormRNA in rat brain by in situ hybridization. Endocrinology 127: 3234–3236
Massague J, Czech MP (1982) The subunit structure of two distinct receptors for insulinlike growth factor I and II and their relationship to the insulin receptor. J Biol Chem257: 5038–5045
McKhann G, Drachman D, Folstein M, Katzman R, Price D, Stadlan E (1984) Clinicaldiagnosis of Alzheimer’s disease: report of the NINCDS-ADRDA work group underthe auspices of department of health and human services task force on Alzheimer’sdisease. Neurology 34: 939–944
Mill JF, Chao MV, Ishi DN (1985) Insulin, insulin-like growth factor II, and nerve growthfactor effects on tubulin mRNA levels and neurite formation. Proc Natl Acad Sci 82:7126–7130
Moroo I, Yamada T, Makino H, Tooyama I, McGeer PL, McGeer EG, Hirayama K(1993) Loss of insulin receptor immunoreactivity from the substantia nigra parscompacta neurons in Parkinson’s disease. Acta Neuropathol 87: 343–348
Morrison LD, Bergeron C, Kish SJ (1993) Brain S-adenosylmethionine decarboxylaseactivity is increased in different areas in Alzheimer’s disease. Neurosci Lett 154: 141–144
Nieuwenhuys R, Voogd J, van Huijzen C (1990) The human central nervous system.A synopsis and atlas. Springer, Berlin Heidelberg New York Tokyo
Perry EK, Perry RH, Tomlinson BE, Blessed G, Gibson PH (1980) Coenzyme A-acety-lating enzymes in Alzheimer’s disease: possible cholinergic “compartment” of pyru-vate dehydrogenase. Neurosci Lett 18: 105–110
Polonsky KS, Rubenstein AH (1984) C-peptide as a measure of the secretion an hepaticextraction of insulin; pitfalls and limitations. Diabetes 33: 486–493
Pope A (1982) General introduction. Human brain dissection. NINCDS Publications,Baltimore
Potau N, Escofet MA, Martinez MC (1991) Ontogenesis of insulin receptors in humancerebral cortex. J Endocrinol Invest 14: 53–58
Puro D, Agardh E (1984) Insulin-mediated regulation of neuronal maturation. Science225: 1170–1172
Quirion R, Araujo DM, Lapehak PA, Seto D, Chabot JG (1991) Growth factors andlymphokines: modulators of cholinergic neuronal activity. Can J Neurol Sci 18: 390–393
Riederer P, Sofic E, Moll G, Freyberger A, Wichart I, Gsell W, Jellinger K, HebenstreitG, Youdim MBH (1990) Dementia of Alzheimer type and Parkinson’s disease:

438 L. Frölich et al.
neurochemical overlaps and specific differences. In: Dostert P, Riederer P, Strolin-Benedetti M, Roncucci R (eds) Early markers in Parkinson’s and Alzheimer’s dis-ease. Springer, Wien New York, pp 221–232
Rijksen G, Staal GEJ (1992) Methodologies for the detection and nonradioactive assay ofprotein-tyrosine kinase activities in human brain and gliomas. Neuroprotocols 1: 201–206
Rotwein P (1991) Structure, evolution, expression and regulation of insulin-like growthfactors I and II. Growth Factors 5: 3–18
Shapiro IP, Masliah E, Saitoh T (1991) Altered protein tyrosine phosphorylation inAlzheimer’s disease. J Neurochem 56: 1154–1162
Steiner DF, Chan SJ, Welsh JM, Kwok SCM (1985) Structure and evolution of the insulingene. Ann Rev Genet 19: 463–469
Sun XJ, Rothenberg P, Kahn CR, Backer JM, Araki E, Wilden PA, Cahill DA, GoldsteinBJ, White MF (1991) Structure of the insulin receptor substrate IRS-1 defines aunique signal transduction protein. Nature 352: 73–77
Tierney MC, Fisher RH, Lewis AJ, Zorzitto ML, Snow WG, Reid DW, Nieuwstraten P(1988) The NINCDS-ADRDA work group criteria for the clinical diagnosis of prob-able Alzheimer’s disease: a clinicopathological study of 57 cases. Neurology 38: 359–364
White MF, Kahn CR (1994) The insulin signalling system. J Biol Chem 269: 1–4Wozniak M, Rydzewski B, Baker SP, Raizada MK (1993) The cellular and physiological
actions of insulin in the central nervous system. Neurochem Int 22: 1–10Wozniak M, Rydzewski B, Baker SP, Raizada MK (1993) The cellular and physiological
actions of insulin in the central nervous system. Neurochem Int 22: 1–10Yang JW, Raizada MK, Fellows RE (1981) Effect of insulin on cultured rat brain cells:
Stimulation of ornithine decarboxylase activity. J Neurochem 36: 1050–1070Young WS (1986) Periventricular hypothalamic cells in the rat brain contain insulin
mRNA. Neuropeptides 8: 93–97
Authors’ address: PD Dr. L. Frölich, Department of Psychiatry I, University ofFrankfurt/Main, Heinrich-Hofmann-Strasse 10, D-60528 Frankfurt am Main, FederalRepublic of Germany.
438 L. Frölich et al.: Brain insulin and insulin receptors
Copyright © 2022 FDOKUMEN




















