
Brachysira (Naviculales, Bacillariophyceae) in lowland waters from Colombia
18
This article was downloaded by: [Yimmy Montoya-Moreno] On: 10 April 2014, At: 12:09 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Diatom Research Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/tdia20 Brachysira (Naviculales, Bacillariophyceae) in lowland waters from Colombia Amelia A. Vouilloud a , Silvia E. Sala b , Marcela Núñez-Avellaneda c , Yimmy Montoya-Moreno c & Santiago R. Duque d a División Científica Ficología, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata Paseo del Bosque s/n, La Plata, Argentina b Instituto Amazónico de Investigaciones Científicas Sinchi, Leticia, Colombia c Grupo de Gestión y Modelación Ambiental Gaia, Universidad de Antioquia, Medellín, Colombia d Instituto Amazónico de Investigaciones, Sede Amazonia Universidad Nacional de Colombia, Leticia, Colombia Published online: 07 Dec 2013. To cite this article: Amelia A. Vouilloud, Silvia E. Sala, Marcela Núñez-Avellaneda, Yimmy Montoya-Moreno & Santiago R. Duque (2014) Brachysira (Naviculales, Bacillariophyceae) in lowland waters from Colombia, Diatom Research, 29:2, 147-163, DOI: 10.1080/0269249X.2013.866909 To link to this article: http://dx.doi.org/10.1080/0269249X.2013.866909 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http:// www.tandfonline.com/page/terms-and-conditions
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Brachysira (Naviculales, Bacillariophyceae) in lowland waters from Colombia

This article was downloaded by: [Yimmy Montoya-Moreno]On: 10 April 2014, At: 12:09Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Diatom ResearchPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tdia20
Brachysira (Naviculales, Bacillariophyceae) in lowlandwaters from ColombiaAmelia A. Vouillouda, Silvia E. Salab, Marcela Núñez-Avellanedac, Yimmy Montoya-Morenoc &Santiago R. Duqued
a División Científica Ficología, Facultad de Ciencias Naturales y Museo, Universidad Nacionalde La Plata Paseo del Bosque s/n, La Plata, Argentinab Instituto Amazónico de Investigaciones Científicas Sinchi, Leticia, Colombiac Grupo de Gestión y Modelación Ambiental Gaia, Universidad de Antioquia, Medellín,Colombiad Instituto Amazónico de Investigaciones, Sede Amazonia Universidad Nacional de Colombia,Leticia, ColombiaPublished online: 07 Dec 2013.
To cite this article: Amelia A. Vouilloud, Silvia E. Sala, Marcela Núñez-Avellaneda, Yimmy Montoya-Moreno & Santiago R.Duque (2014) Brachysira (Naviculales, Bacillariophyceae) in lowland waters from Colombia, Diatom Research, 29:2, 147-163,DOI: 10.1080/0269249X.2013.866909
To link to this article: http://dx.doi.org/10.1080/0269249X.2013.866909
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Diatom Research, 2014Vol. 29, No. 2, 147–163, http://dx.doi.org/10.1080/0269249X.2013.866909
Brachysira (Naviculales, Bacillariophyceae) in lowland waters from Colombia
AMELIA A. VOUILLOUD1∗, SILVIA E. SALA1, MARCELA NÚÑEZ-AVELLANEDA2,YIMMY MONTOYA-MORENO3 & SANTIAGO R. DUQUE4
1División Científica Ficología, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata Paseo del Bosque s/n, LaPlata, Argentina2Instituto Amazónico de Investigaciones Científicas Sinchi, Leticia, Colombia3Grupo de Gestión y Modelación Ambiental Gaia, Universidad de Antioquia, Medellín, Colombia4Instituto Amazónico de Investigaciones, Sede Amazonia Universidad Nacional de Colombia, Leticia, Colombia
This study focuses on the genus Brachysira Kützing from lowland waters as part of the diatom flora investigation of Colombia. Periphytonsamples were collected in lentic and lotic waterbodies from different Amazonian basins and the Ayapel swamp system in the northernAtlantic plain of Colombia. Diatoms were analysed with light and scanning electron microscopy. Two new taxa, B. guarrerai and B.huitotarum, are described. These species were differentiated from their allied taxa based on the combination of: the shape of the centralarea; the shape and number of the areolae that delimit the central area; stria position; presence or absence of papillae on the interstriae;and degree of silicification of the ribs that delimit the raphe. In addition, five other taxa are reported and described: B. microcephala(Grunow) Compère and B. subrostrata Lange-Bertalot are recorded for the first time from Colombia, while B. manfredii Lange-Bertalotwas previously found in the Sierra Nevada.The other two taxa were identified as B. aff. brebissonii Ross and B. aff. microcephala becausethe Colombian specimens differ slightly from the type material of these cosmopolitan species.
Keywords: Bacillariophyceae, Brachysira guarrerai, Brachysira huitotarum, Colombia, diatom taxonomy, neotropical
IntroductionThe genus Brachysira was erected by Kützing (1836)based on the marine species B. aponina Kützing. Kützing(1844) transferred it to the genus Navicula Bory. Round &Mann (1981) re-examined the type material of B. aponinaand redefined the genus considering also some species ofAnomoeoneis Pfitzer that they transferred to Brachysira.These authors defined the distinctive characters based onthe type species of the genus as: (1) raphe between twoprominent longitudinal ribs; (2) a prominent marginal ribat the valve face and mantle union; (3) another siliceouslongitudinal rib located between the marginal rib and theraphe; (4) mantle with a row of elongate areolae contin-uing around the valve apices. They also point out thatthe valve face is a plane and has transapically elongatedareolae closed by internal hymens. The external proximalraphe ends are expanded and terminal fissures are absent.Internally, the proximal raphe fissures are separated by asiliceous longitudinal thickening and distal ends finish insmall helictoglossae. Round et al. (1990) mentioned othercharacteristics including the presence of one plastid, exter-nal terminal T-shaped raphe ends, cingulum with opencopulae, valvocopula folded inwards at its junction withthe valve margin to form a long chamber and the moreadvalvar bands sometimes with one row of poroids. In
∗Corresponding author. Email: [email protected]
(Received 28 December 2012; accepted 11 November 2013)
their study of Brachysira species from oligotrophic lakesfrom Florida, Shayler & Siver (2004a) noted difficultiesin the identification of some species that are frequentlymentioned as cosmopolitan. They performed an exhaus-tive revision of the prominent taxa from the eastern UnitedStates and pointed out that other characters such as thenumber of areolae per stria, the proportion of the valvelength with striae composed of only one areola, the are-ola length and the presence of papillae on the interstriaeare important for distinguishing taxa. Lange-Bertalot &Moser (1994) analysed the genus in detail and described67 taxa, more than one third of them new to science, intheir study of neotropical taxa from Venezuela, Brazil andColombia. Subsequently, more new species and varietieshave been described and at present the genus comprises97 accepted taxa (Metzeltin & Lange-Bertalot 1998, 2007,Wolfe & Kling 2001, Shayler & Siver 2004a,b, Guiry &Guiry 2013). Although this is an extensively studied genusthere is still confusion over species identifications, partic-ularly those considered cosmopolitan, e.g., B. brebissoniiRoss, B. neoexilis Lange-Bertalot and B. microcephala(Grunow) Compère. Metzeltin & Lange-Bertalot (1998,2007) recorded several taxa from tropical habitats of SouthAmerica as Brachysira spp., which suggests that there arestill many undescribed taxa in the region.
© 2013 The International Society for Diatom Research
Dow
nloa
ded
by [
Yim
my
Mon
toya
-Mor
eno]
at 1
2:09
10
Apr
il 20
14

148 Vouilloud et al.
During a study of the diatom flora from different regionsof Colombia, both lowland and highland, only 16% ofthe 450 registered taxa were present in at least two ofthe studied areas (Sala et al. 1999, 2002a,b, 2008a,b, Sala& Ramírez 2008, Vouilloud et al. 2010, Montoya-Morenoet al. 2011, 2012). Waterbodies from Amazonia and theAyapel swamp system were most closely related in ouranalysis. As both regions are at low altitudes this similar-ity was attributed, among other factors, to strong altitudinalgradients at the tropics (S.E. Sala, unpubl. data).
Sixteen Brachysira taxa were found in Colombia(some reported as Anomoeoneis, Y. Montoya-Moreno, pers.comm.). During our study, Brachysira was only found inlowland waters of the Amazonian region and Ayapel, a sys-tem of swamps in the northern Atlantic plain of Colombia.The aim of this study is to analyse the fine valve mor-phology of neotropical Brachysira taxa, to compare themwith species described in the literature, and to improve theknowledge of this genus.
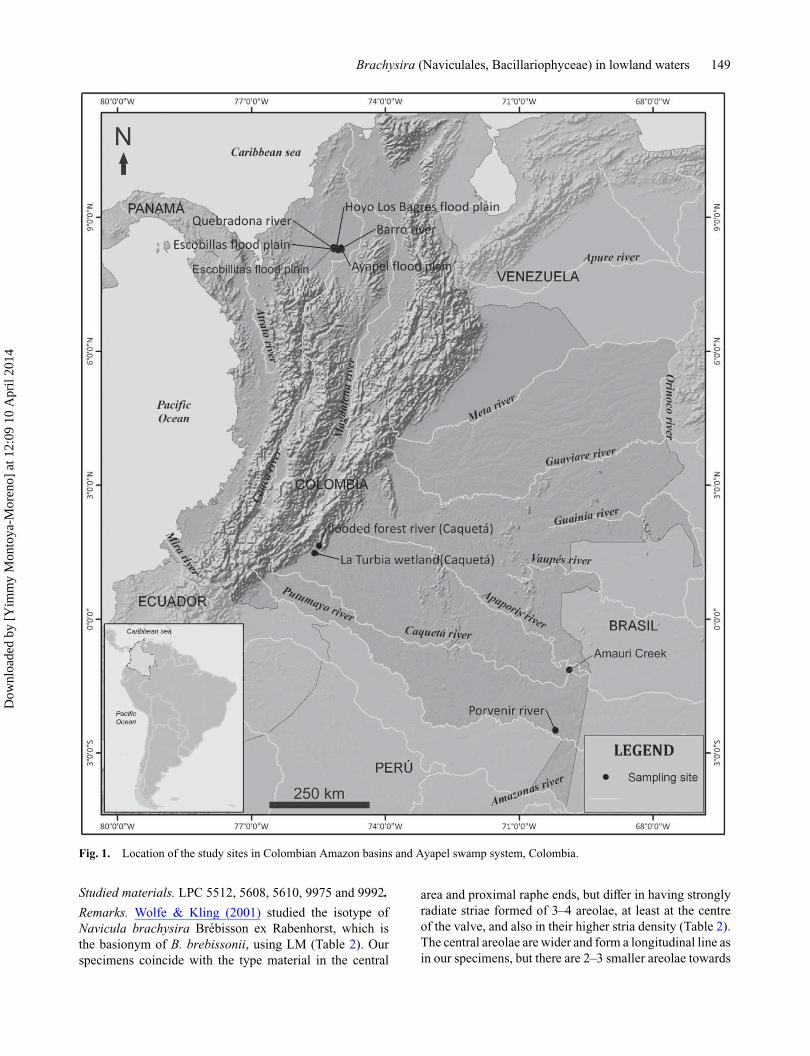
Materials and methodsSamples for this study were collected in the ColombianAmazon and in the Ayapel swamp system. In the formercase, lentic and lotic waterbodies of different basins weresampled in February 1991 and November 2003, and in thelatter during May 2006 and January 2009 (Fig. 1). Horizon-tal and vertical tows were made with a plankton net (meshsize = 24 μm) at the euphotic zone, and benthic diatomswere sampled by scraping macrophytes. At each samplingstation, water transparency (Secchi disc), temperature, pH,dissolved oxygen and conductivity were measured with aWTW 320 potentiometric probe (Table 1).
Samples were fixed with 6–8% buffered formalin or 10%Lugol’s solution and treated to eliminate organic matter fol-lowing different methods: Hasle & Fryxell (1970), CEN(2003) and/or in a muffle furnace at 500◦C for 10 min. Sam-ples for light microscopy (LM) were mounted as permanentslides with Naphrax� and samples for scanning electronmicroscopy (SEM) were coated with gold–palladium ina Jeol FINE COAT ION SPUTTER JFC1100. Observa-tions were conducted with a Wild M20 and Leica DM2500 phase contrast LM and a Jeol JSM–6360L SEM.The terminology used is that proposed by Anonymous(1975) and Ross et al. (1979).
Unclean and clean samples and permanent slides weredeposited at the Herbarium of the División CientíficaFicología, Museo de Ciencias Naturales de La Plata, Uni-versidad Nacional de La Plata (UNLP) Argentina under thefollowing numbers:
LPC 5512: Amauri Creek, Departamento del Amazonas,Colombia. 11-1994. Collected by Santiago R. Duque.
LPC 5575: Porvenir River, Departamento del Amazonas,Colombia. 06-2001. Collected by Marcela Núñez-Avellaneda.
LPC 5608: Temporal Swamp La Turbia, Departamento deCaquetá, Colombia. 11-2003. Collected by MarcelaNúñez-Avellaneda.
LPC 5610: Inundated forest creek, Departamento deCaquetá, Colombia. 11-2003. Collected by MarcelaNúñez-Avellaneda.
LPC 9939: Permanent Swamp Escobillitas, Departamentode Córdoba, Colombia. 01-2009. Collected by YimmyMontoya-Moreno.
LPC 9942: Permanent Swamp Escobillitas, Departamentode Córdoba, Colombia. 04-2007. Collected by YimmyMontoya-Moreno.
LPC 9947: Quebradona River, Departamento de Córdoba,Colombia. 05-2007. Collected by Yimmy Montoya-Moreno.
LPC 9966: Permanent Swamp Escobillitas, Departamentode Córdoba, Colombia. 01-2009. Collected by YimmyMontoya-Moreno.
LPC 9975: Barro River, Departamento de Córdoba,Colombia. 09-2006. Collected by Yimmy Montoya-Moreno.
LPC 9984. Permanent Swamp Ayapel, Departamento deCórdoba, Colombia. 01-2009. Collected by YimmyMontoya-Moreno.
LPC 9992. Permanent Swamp Hoyo Los Bagres, Departa-mento de Córdoba, Colombia. 01-2009. Collected byYimmy Montoya-Moreno.
LPC 9999. Barro River, Departamento de Córdoba,Colombia. 01-2009. Collected by Yimmy Montoya-Moreno.
ResultsBrachysira aff. brebissonii Ross (Figs 2–6)Description. Valves lanceolate, valve face flat withrounded ends (Figs 2–4). Striae uniseriate, mostly radiate,only the last 2–3 striae near the apices are parallel, mostof them comprise 2, sometimes 3, transapically elongatedareolae at the valve ends, the last 5–6 striae at the valveapices contain only one areola (Figs 5–6). Areolae well-spaced, delimiting a wide hyaline, undulate longitudinalarea visible with LM, without anastomosing lines (Figs 2–3). A stria with small round areolae marks the position ofthe Voigt discontinuity (Fig. 5). Interstriae with irregularlyspaced papillae, 2–3 per interstria at the centre (Figs 5–6).Raphe simple, filiform, slightly sinuous externally, proxi-mal and distal ends slightly curved to the primary side ofthe valve (Figs 2–5). Proximal ends simple; terminal fis-sure T–shaped with curved branches (Fig. 6). Central areaasymmetrical, rounded axial area straight and very narrow(Figs 2–5). Mantle without papillae (Fig. 6).
Morphometric data (n = 6). Length: 17–20 μm; width:4.5–5.0 μm; length-to-width ratio: 3.8–4.0; 27–30 striae in10 μm; D(pre) (distance between the proximal raphe ends):1.5 μm.
Dow
nloa
ded
by [
Yim
my
Mon
toya
-Mor
eno]
at 1
2:09
10
Apr
il 20
14
Brachysira (Naviculales, Bacillariophyceae) in lowland waters 149
Fig. 1. Location of the study sites in Colombian Amazon basins and Ayapel swamp system, Colombia.
Studied materials. LPC 5512, 5608, 5610, 9975 and 9992.Remarks. Wolfe & Kling (2001) studied the isotype ofNavicula brachysira Brébisson ex Rabenhorst, which isthe basionym of B. brebissonii, using LM (Table 2). Ourspecimens coincide with the type material in the central
area and proximal raphe ends, but differ in having stronglyradiate striae formed of 3–4 areolae, at least at the centreof the valve, and also in their higher stria density (Table 2).The central areolae are wider and form a longitudinal line asin our specimens, but there are 2–3 smaller areolae towards
Dow
nloa
ded
by [
Yim
my
Mon
toya
-Mor
eno]
at 1
2:09
10
Apr
il 20
14
150V
ouilloudetal.
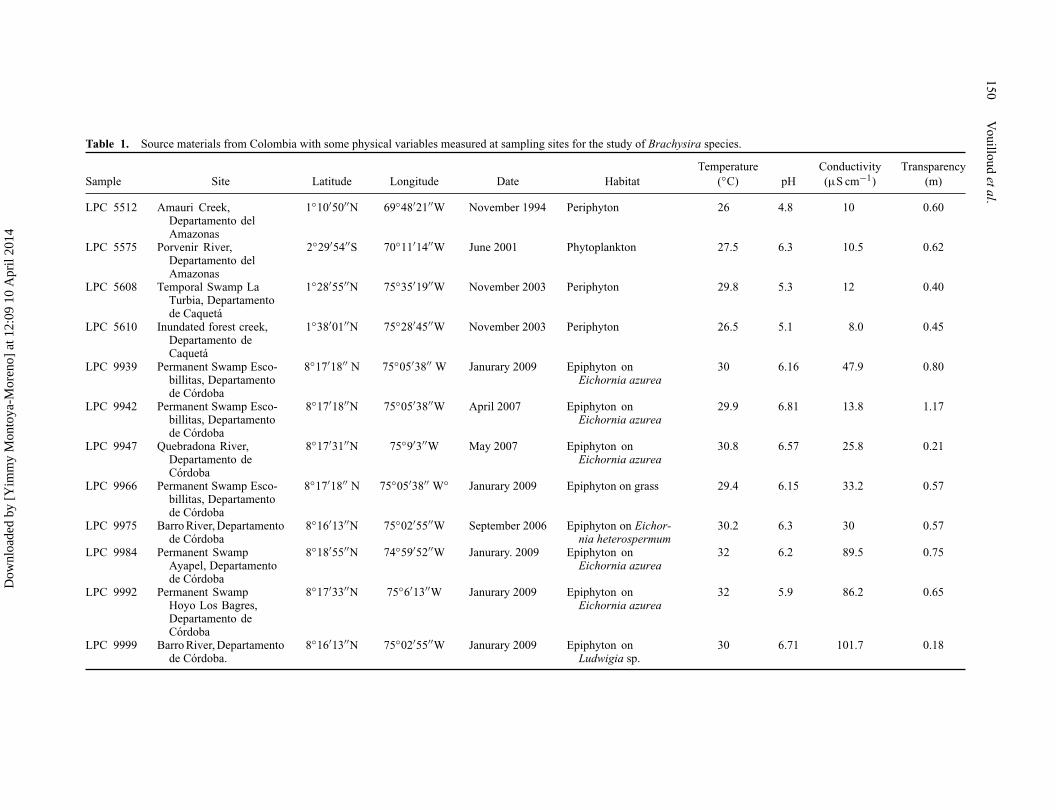
Table 1. Source materials from Colombia with some physical variables measured at sampling sites for the study of Brachysira species.
Temperature Conductivity TransparencySample Site Latitude Longitude Date Habitat (◦C) pH (μS cm−1) (m)
LPC 5512 Amauri Creek,Departamento delAmazonas
1◦10′50′′N 69◦48′21′′W November 1994 Periphyton 26 4.8 10 0.60
LPC 5575 Porvenir River,Departamento delAmazonas
2◦29′54′′S 70◦11′14′′W June 2001 Phytoplankton 27.5 6.3 10.5 0.62
LPC 5608 Temporal Swamp LaTurbia, Departamentode Caquetá
1◦28′55′′N 75◦35′19′′W November 2003 Periphyton 29.8 5.3 12 0.40
LPC 5610 Inundated forest creek,Departamento deCaquetá
1◦38′01′′N 75◦28′45′′W November 2003 Periphyton 26.5 5.1 8.0 0.45
LPC 9939 Permanent Swamp Esco-billitas, Departamentode Córdoba
8◦17′18′′ N 75◦05′38′′ W Janurary 2009 Epiphyton onEichornia azurea
30 6.16 47.9 0.80
LPC 9942 Permanent Swamp Esco-billitas, Departamentode Córdoba
8◦17′18′′N 75◦05′38′′W April 2007 Epiphyton onEichornia azurea
29.9 6.81 13.8 1.17
LPC 9947 Quebradona River,Departamento deCórdoba
8◦17′31′′N 75◦9′3′′W May 2007 Epiphyton onEichornia azurea
30.8 6.57 25.8 0.21
LPC 9966 Permanent Swamp Esco-billitas, Departamentode Córdoba
8◦17′18′′ N 75◦05′38′′ W◦ Janurary 2009 Epiphyton on grass 29.4 6.15 33.2 0.57
LPC 9975 Barro River, Departamentode Córdoba
8◦16′13′′N 75◦02′55′′W September 2006 Epiphyton on Eichor-nia heterospermum
30.2 6.3 30 0.57
LPC 9984 Permanent SwampAyapel, Departamentode Córdoba
8◦18′55′′N 74◦59′52′′W Janurary. 2009 Epiphyton onEichornia azurea
32 6.2 89.5 0.75
LPC 9992 Permanent SwampHoyo Los Bagres,Departamento deCórdoba
8◦17′33′′N 75◦6′13′′W Janurary 2009 Epiphyton onEichornia azurea
32 5.9 86.2 0.65
LPC 9999 Barro River, Departamentode Córdoba.
8◦16′13′′N 75◦02′55′′W Janurary 2009 Epiphyton onLudwigia sp.
30 6.71 101.7 0.18
Dow
nloa
ded
by [
Yim
my
Mon
toya
-Mor
eno]
at 1
2:09
10
Apr
il 20
14

Brachysira (Naviculales, Bacillariophyceae) in lowland waters 151
Figs 2–14. Specimens of Brachysira aff. brebissonii and B. guarrerai sp. nov., LM (Figs 2–3, 7–9) and SEM (Figs 4–6, 10–14). Figs2–6. Brachysira aff. brebissonii. Figs 2–3. Valves of different size. Fig. 4. Whole valve in external view. Figs 5–6. Detail of valve inexternal view showing small papillae on the interstriae, T-shaped distal raphe end and Voigt discontinuity (arrow). Fig. 2. Specimen ofBarro River (Ayapel). Fig. 3. Specimen of Temporal Swamp La Turbia (Amazonia). Fig. 4. Specimen of Amauri Creek (Amazonia). Figs7–14. Brachysira guarrerai sp. nov. Figs 7–9. Valves showing size range variation. Fig. 9. Holotype specimen. Fig. 10. Whole valve inexternal view. Fig. 11. Whole valve in internal view. Fig. 12. Detail of external valve face, note ribs slightly prominent, lack of papillaeand T-shaped distal raphe end. Fig. 13. Internal view of the valve centre. Fig. 14. Internal view of the valve end. Scale bars = 5 μm (Figs2–4, 7–11), 2 μm (Figs 5, 12–14), 1 μm (Fig. 6).
Dow
nloa
ded
by [
Yim
my
Mon
toya
-Mor
eno]
at 1
2:09
10
Apr
il 20
14

152 Vouilloud et al.
the apices. A few specimens were found making it insuffi-cient to decide whether our material and that described andillustrated in the literature unequivocally correspond to B.brebissonii.
However, the specimens from Colombia recorded asB. brebissonii ssp. brebissonii in Lange-Bertalot & Moser(1994, pl. 44, Figs 1–10) differ from the type material in thesame way as ours.
Ecology and distribution. This species was recordedin periphyton samples collected from swamps, streamsand rivers of the Amazonian region and Ayapel system,with high water temperature (26–32◦C), acid pH (4.8–6.3),very low to low conductivity (8–86 μ S cm−1) and lowtransparency (0.40–0.65 m).
Brachysira guarrerai Vouilloud, Sala &Núñez-Avellaneda (Figs 7–14)Description. Valves lanceolate, slightly asymmetrical withrespect to the transapical axis, with rostrate to subcapi-tate ends (Figs 7–10). Valve face flat without a prominentmarginal rib at the valve face and mantle junction. Uniseri-ate striae, parallel to slightly radiate at valve centre andradiate over the rest of the valve face. Striae with 2–3transapically elongated areolae, the last 10–12 with onlyone areola (Figs 10 and 12). The areolae delimiting thecentral area are longer on the primary side of the valve.1–2 longitudinal undulate and anastomosing hyaline linesdelimited by the irregularly spaced areolae are visible withLM. Interstriae without papillae (Fig. 12). Areolae closedwith internal hymens (Figs 11 and 13–14). Raphe simple,filiform, slightly sinuous, bordered by slightly thickenedsiliceous ribs (Figs 10 and 12). External proximal rapheends straight, slightly expanded; terminal fissure T–shaped,with curved branches (Fig. 12). Internally proximal rapheends punctiform (Fig. 13); distal raphe ends straight, helic-toglossae small placed near the apices (Fig. 14). Centralarea elliptical, asymmetrical. Axial area narrow and straight(Figs 7–10).
Morphometric data (n = 24). Length: 19–38 μm;width: 5.5–7.0 μm; length-to-width ratio: 3.6–5.4; 26–32striae in 10 μm; D(pre): 0.8 μm.
Holotype. Slide LPC 5575 (1), División CientíficaFicología, Facultad de Ciencias Naturales y Museo(UNLP), La Plata, Argentina. Holotype specimen here des-ignated Finder: R 44.1 (The England Finder™ Graticule,model 5–7) and illustrated in Fig. 9.
Type locality. Río Porvenir, Putumayo River Basin,Colombia.
Etymology. The specific epithet refers to Dr SebastiánA. Guarrera, Professor Emeritus at the UNLP, Argentina, apioneer of freshwater algal studies in South America.
Studied materials. The species was only found in thetype locality (LPC 5575).
Taxonomic remarks. Brachysira guarrerai is similar toB. garrensis (Krammer & Lange-Bertalot) Lange-Bertalot& Moser, with prominent axial and marginal ribs and with-out papillae (Table 2). But the latter taxon differs in valveoutline, being rhomboid to lanceolate, with undifferentiatedto slightly protracted ends, and having radiate and moredensely arranged striae at valve centre.
The specimens collected in Venezuela and recorded asB. cf. garrensis in Lange-Bertalot & Moser (1994) coincidein morphological and morphometrical characteristics withB. guarrerai (Table 2). Based on these similarities, they areconspecific.
Ecology and distribution. This species was onlyrecorded in the Amazon Region, in the Porvenir Riverwith high water temperature (27.5◦C), slightly acid pH(6.3), very low conductivity (10.5 μ S cm−1) and low trans-parency (0.62 m).
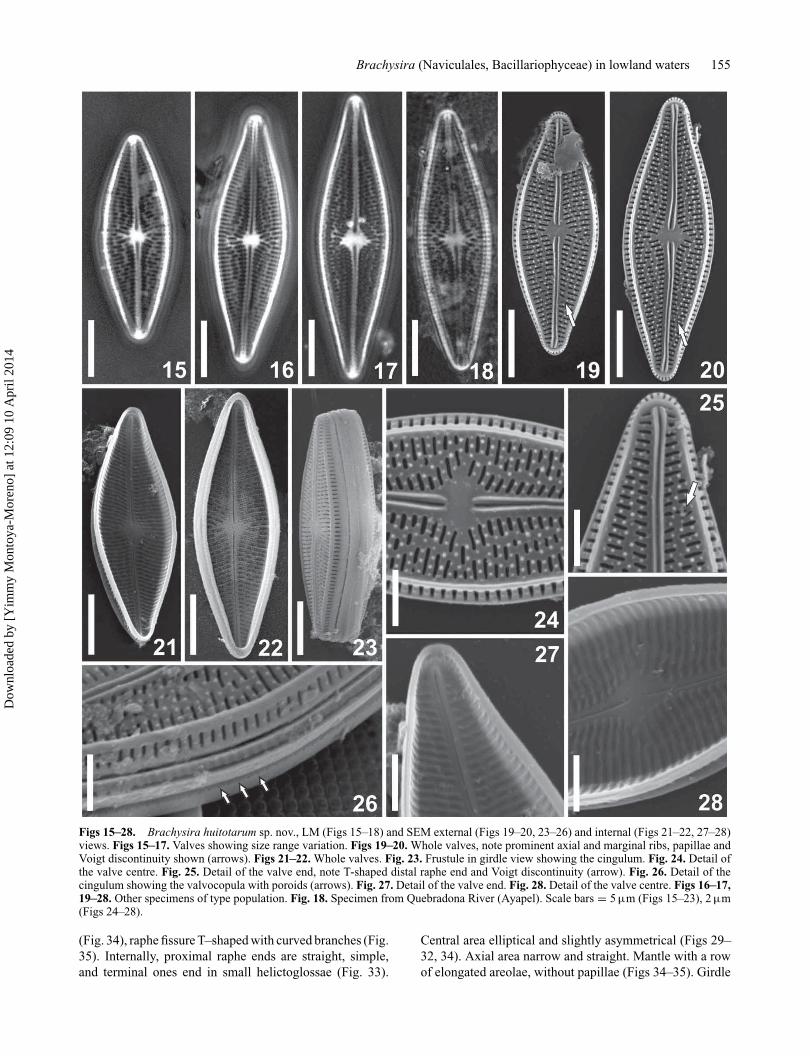
Brachysira huitotarum Vouilloud, Sala &Núñez-Avellaneda (Figs 15–28)Description. Valves broadly lanceolate to rhomboid, valveface flat, with rounded to subrostrate apices, prominentmarginal ribs at valve face and mantle junction (Figs 15–20,23, 26). Uniseriate striae, strongly radiate at the centre witha shortened stria inserted on each side, moderately radiateover the rest of the valve (Figs 15–24). Most of the striaeon the valve face are composed of 3 areoale, except at cen-tre where they are formed by 2 areolae plus the shortenedstria which has one areola, and at apices, striae are formedby one transapically elongated areola (Figs 19–20, 24–25).The distribution of the areolae delimits sinuous longitudi-nal lines, one at the valve centre that divides in two towardsthe apices and then anastomoses at the apex (Figs 15–20).Areolae closed by internal hymens (Figs 21–22, 27–28).Voigt discontinuity revealed by the greater separation ofthe striae or by irregular areolae (Fig. 25, arrow). Inters-triae with small, irregularly spaced papillae, 5–6 papillaeper interstria at the centre (Fig. 24). Raphe simple, filiform,externally slightly sinuous, proximal and distal ends curvedto the primary valve side, flanked by prominent siliceousribs (Figs 19–20, 24–25). Proximal raphe ends simple (Fig.24); terminal fissure T–shaped with curved branches (Fig.25); internally proximal raphe ends are curved to the pri-mary side of the valve (Fig. 28); distal raphe ends straight,helictoglossae very small, near the apices (Fig. 27). Centralarea rhomboid, slightly irregular and asymmetrical (Figs15–20, 24). Axial area narrow, straight, narrower on theprimary side. The mantle is inclined with respect to thevalve face, so that the elongated areolae on the mantle arevisible in valve view, coincident with each stria (Figs 23,26). The mantle also has papillae. Girdle with 4 copulae,valvocopula with a row of small poroids (Fig. 26, arrows).
Morphometric data (n = 28). Length: 17.0–28.5 μm;width: 5.2–7.5 μm; length-to-width ratio: 2.7–3.8; 25–30
Dow
nloa
ded
by [
Yim
my
Mon
toya
-Mor
eno]
at 1
2:09
10
Apr
il 20
14
Brachysira
(Naviculales,B
acillariophyceae)inlow
landw
aters153
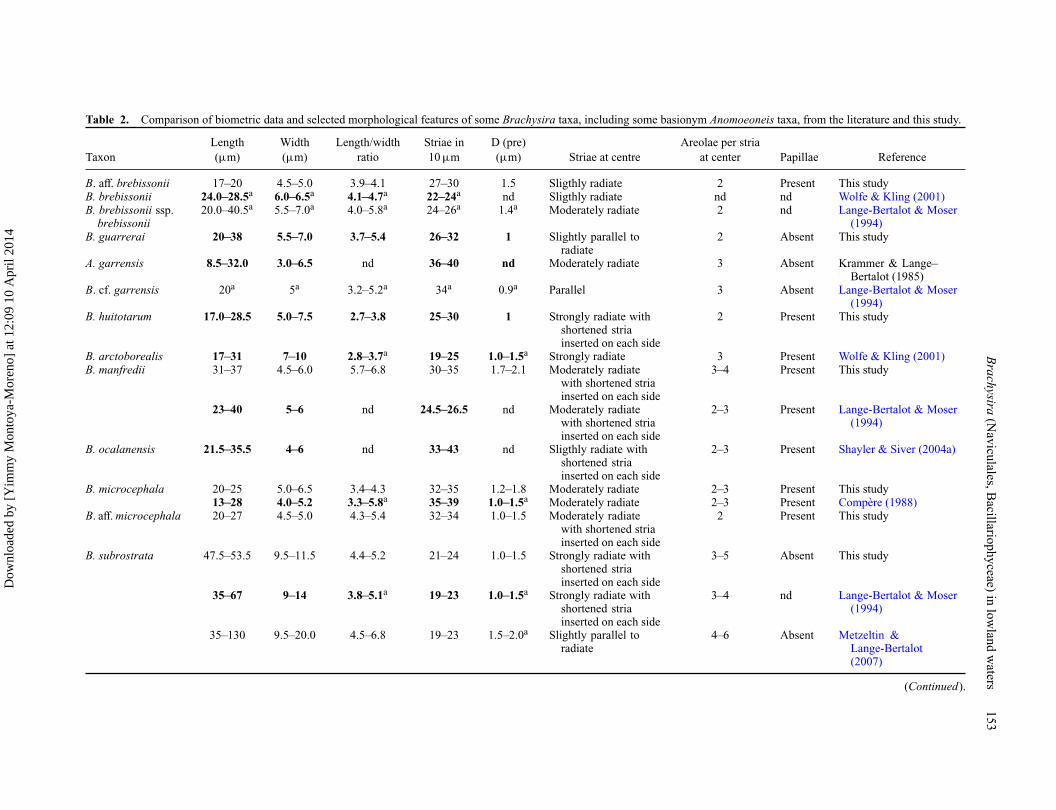
Table 2. Comparison of biometric data and selected morphological features of some Brachysira taxa, including some basionym Anomoeoneis taxa, from the literature and this study.
Length Width Length/width Striae in D (pre) Areolae per striaTaxon (μm) (μm) ratio 10 μm (μm) Striae at centre at center Papillae Reference
B. aff. brebissonii 17–20 4.5–5.0 3.9–4.1 27–30 1.5 Sligthly radiate 2 Present This studyB. brebissonii 24.0–28.5a 6.0–6.5a 4.1–4.7a 22–24a nd Sligthly radiate nd nd Wolfe & Kling (2001)B. brebissonii ssp.
brebissonii20.0–40.5a 5.5–7.0a 4.0–5.8a 24–26a 1.4a Moderately radiate 2 nd Lange-Bertalot & Moser
(1994)B. guarrerai 20–38 5.5–7.0 3.7–5.4 26–32 1 Slightly parallel to
radiate2 Absent This study
A. garrensis 8.5–32.0 3.0–6.5 nd 36–40 nd Moderately radiate 3 Absent Krammer & Lange–Bertalot (1985)
B. cf. garrensis 20a 5a 3.2–5.2a 34a 0.9a Parallel 3 Absent Lange-Bertalot & Moser(1994)
B. huitotarum 17.0–28.5 5.0–7.5 2.7–3.8 25–30 1 Strongly radiate withshortened striainserted on each side
2 Present This study
B. arctoborealis 17–31 7–10 2.8–3.7a 19–25 1.0–1.5a Strongly radiate 3 Present Wolfe & Kling (2001)B. manfredii 31–37 4.5–6.0 5.7–6.8 30–35 1.7–2.1 Moderately radiate
with shortened striainserted on each side
3–4 Present This study
23–40 5–6 nd 24.5–26.5 nd Moderately radiatewith shortened striainserted on each side
2–3 Present Lange-Bertalot & Moser(1994)
B. ocalanensis 21.5–35.5 4–6 nd 33–43 nd Sligthly radiate withshortened striainserted on each side
2–3 Present Shayler & Siver (2004a)
B. microcephala 20–25 5.0–6.5 3.4–4.3 32–35 1.2–1.8 Moderately radiate 2–3 Present This study13–28 4.0–5.2 3.3–5.8a 35–39 1.0–1.5a Moderately radiate 2–3 Present Compère (1988)
B. aff. microcephala 20–27 4.5–5.0 4.3–5.4 32–34 1.0–1.5 Moderately radiatewith shortened striainserted on each side
2 Present This study
B. subrostrata 47.5–53.5 9.5–11.5 4.4–5.2 21–24 1.0–1.5 Strongly radiate withshortened striainserted on each side
3–5 Absent This study
35–67 9–14 3.8–5.1a 19–23 1.0–1.5a Strongly radiate withshortened striainserted on each side
3–4 nd Lange-Bertalot & Moser(1994)
35–130 9.5–20.0 4.5–6.8 19–23 1.5–2.0a Slightly parallel toradiate
4–6 Absent Metzeltin &Lange-Bertalot(2007)
(Continued).
Dow
nloa
ded
by [
Yim
my
Mon
toya
-Mor
eno]
at 1
2:09
10
Apr
il 20
14
154 Vouilloud et al.
Tab
le2.
Con
tinue
d.
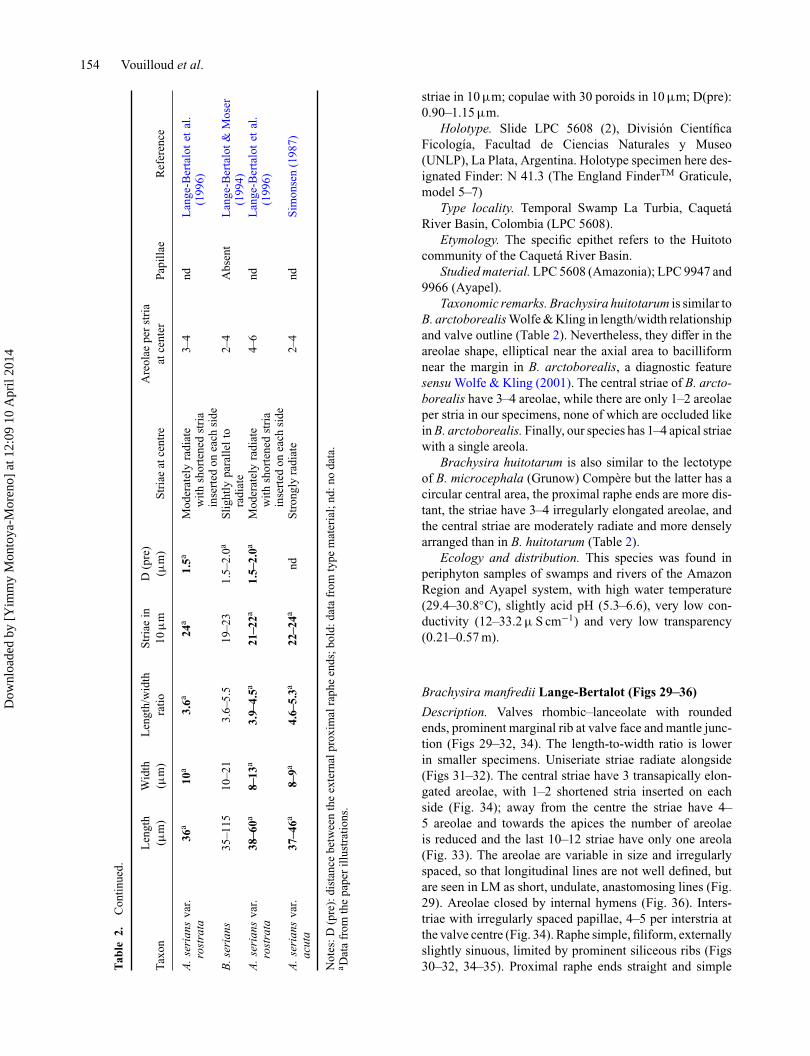
Len
gth
Wid
thL
engt
h/w
idth
Stri
aein
D(p
re)
Are
olae
pers
tria
Taxo
n(μ
m)
(μm
)ra
tio10
μm
(μm
)St
riae
atce
ntre
atce
nter
Papi
llae
Ref
eren
ce
A.s
eria
nsva
r.ro
stra
ta36
a10
a3.
6a24
a1.
5aM
oder
atel
yra
diat
ew
ithsh
orte
ned
stri
ain
sert
edon
each
side
3–4
ndL
ange
-Ber
talo
tet
al.
(199
6)
B.s
eria
ns35
–115
10–2
13.
6–5.
519
–23
1.5–
2.0a
Slig
htly
para
llelt
ora
diat
e2–
4A
bsen
tL
ange
-Ber
talo
t&M
oser
(199
4)A
.ser
ians
var.
rost
rata
38–6
0a8–
13a
3.9–
4.5a
21–2
2a1.
5–2.
0aM
oder
atel
yra
diat
ew
ithsh
orte
ned
stri
ain
sert
edon
each
side
4–6
ndL
ange
-Ber
talo
tet
al.
(199
6)
A.s
eria
nsva
r.ac
uta
37–4
6a8–
9a4.
6–5.
3a22
–24a
ndSt
rong
lyra
diat
e2–
4nd
Sim
onse
n(1
987)
Not
es:D
(pre
):di
stan
cebe
twee
nth
eex
tern
alpr
oxim
alra
phe
ends
;bol
d:da
tafr
omty
pem
ater
ial;
nd:n
oda
ta.
a Dat
afr
omth
epa
peri
llust
ratio
ns.
striae in 10 μm; copulae with 30 poroids in 10 μm; D(pre):0.90–1.15 μm.
Holotype. Slide LPC 5608 (2), División CientíficaFicología, Facultad de Ciencias Naturales y Museo(UNLP), La Plata, Argentina. Holotype specimen here des-ignated Finder: N 41.3 (The England FinderTM Graticule,model 5–7)
Type locality. Temporal Swamp La Turbia, CaquetáRiver Basin, Colombia (LPC 5608).
Etymology. The specific epithet refers to the Huitotocommunity of the Caquetá River Basin.
Studied material. LPC 5608 (Amazonia); LPC 9947 and9966 (Ayapel).
Taxonomic remarks. Brachysira huitotarum is similar toB. arctoborealis Wolfe & Kling in length/width relationshipand valve outline (Table 2). Nevertheless, they differ in theareolae shape, elliptical near the axial area to bacilliformnear the margin in B. arctoborealis, a diagnostic featuresensu Wolfe & Kling (2001). The central striae of B. arcto-borealis have 3–4 areolae, while there are only 1–2 areolaeper stria in our specimens, none of which are occluded likein B. arctoborealis. Finally, our species has 1–4 apical striaewith a single areola.
Brachysira huitotarum is also similar to the lectotypeof B. microcephala (Grunow) Compère but the latter has acircular central area, the proximal raphe ends are more dis-tant, the striae have 3–4 irregularly elongated areolae, andthe central striae are moderately radiate and more denselyarranged than in B. huitotarum (Table 2).
Ecology and distribution. This species was found inperiphyton samples of swamps and rivers of the AmazonRegion and Ayapel system, with high water temperature(29.4–30.8◦C), slightly acid pH (5.3–6.6), very low con-ductivity (12–33.2 μ S cm−1) and very low transparency(0.21–0.57 m).
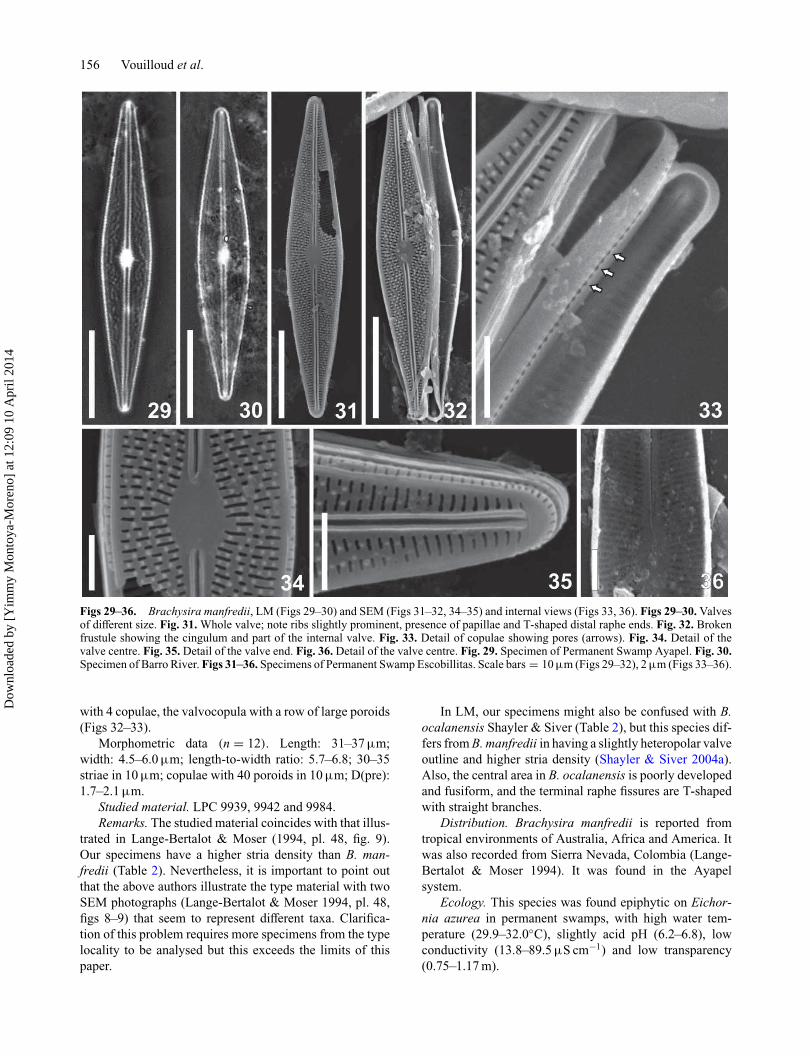
Brachysira manfredii Lange-Bertalot (Figs 29–36)Description. Valves rhombic–lanceolate with roundedends, prominent marginal rib at valve face and mantle junc-tion (Figs 29–32, 34). The length-to-width ratio is lowerin smaller specimens. Uniseriate striae radiate alongside(Figs 31–32). The central striae have 3 transapically elon-gated areolae, with 1–2 shortened stria inserted on eachside (Fig. 34); away from the centre the striae have 4–5 areolae and towards the apices the number of areolaeis reduced and the last 10–12 striae have only one areola(Fig. 33). The areolae are variable in size and irregularlyspaced, so that longitudinal lines are not well defined, butare seen in LM as short, undulate, anastomosing lines (Fig.29). Areolae closed by internal hymens (Fig. 36). Inters-triae with irregularly spaced papillae, 4–5 per interstria atthe valve centre (Fig. 34). Raphe simple, filiform, externallyslightly sinuous, limited by prominent siliceous ribs (Figs30–32, 34–35). Proximal raphe ends straight and simple
Dow
nloa
ded
by [
Yim
my
Mon
toya
-Mor
eno]
at 1
2:09
10
Apr
il 20
14
Brachysira (Naviculales, Bacillariophyceae) in lowland waters 155
Figs 15–28. Brachysira huitotarum sp. nov., LM (Figs 15–18) and SEM external (Figs 19–20, 23–26) and internal (Figs 21–22, 27–28)views. Figs 15–17. Valves showing size range variation. Figs 19–20. Whole valves, note prominent axial and marginal ribs, papillae andVoigt discontinuity shown (arrows). Figs 21–22. Whole valves. Fig. 23. Frustule in girdle view showing the cingulum. Fig. 24. Detail ofthe valve centre. Fig. 25. Detail of the valve end, note T-shaped distal raphe end and Voigt discontinuity (arrow). Fig. 26. Detail of thecingulum showing the valvocopula with poroids (arrows). Fig. 27. Detail of the valve end. Fig. 28. Detail of the valve centre. Figs 16–17,19–28. Other specimens of type population. Fig. 18. Specimen from Quebradona River (Ayapel). Scale bars = 5 μm (Figs 15–23), 2 μm(Figs 24–28).
(Fig. 34), raphe fissure T–shaped with curved branches (Fig.35). Internally, proximal raphe ends are straight, simple,and terminal ones end in small helictoglossae (Fig. 33).
Central area elliptical and slightly asymmetrical (Figs 29–32, 34). Axial area narrow and straight. Mantle with a rowof elongated areolae, without papillae (Figs 34–35). Girdle
Dow
nloa
ded
by [
Yim
my
Mon
toya
-Mor
eno]
at 1
2:09
10
Apr
il 20
14
156 Vouilloud et al.
Figs 29–36. Brachysira manfredii, LM (Figs 29–30) and SEM (Figs 31–32, 34–35) and internal views (Figs 33, 36). Figs 29–30. Valvesof different size. Fig. 31. Whole valve; note ribs slightly prominent, presence of papillae and T-shaped distal raphe ends. Fig. 32. Brokenfrustule showing the cingulum and part of the internal valve. Fig. 33. Detail of copulae showing pores (arrows). Fig. 34. Detail of thevalve centre. Fig. 35. Detail of the valve end. Fig. 36. Detail of the valve centre. Fig. 29. Specimen of Permanent Swamp Ayapel. Fig. 30.Specimen of Barro River. Figs 31–36. Specimens of Permanent Swamp Escobillitas. Scale bars = 10 μm (Figs 29–32), 2 μm (Figs 33–36).
with 4 copulae, the valvocopula with a row of large poroids(Figs 32–33).
Morphometric data (n = 12). Length: 31–37 μm;width: 4.5–6.0 μm; length-to-width ratio: 5.7–6.8; 30–35striae in 10 μm; copulae with 40 poroids in 10 μm; D(pre):1.7–2.1 μm.
Studied material. LPC 9939, 9942 and 9984.Remarks. The studied material coincides with that illus-
trated in Lange-Bertalot & Moser (1994, pl. 48, fig. 9).Our specimens have a higher stria density than B. man-fredii (Table 2). Nevertheless, it is important to point outthat the above authors illustrate the type material with twoSEM photographs (Lange-Bertalot & Moser 1994, pl. 48,figs 8–9) that seem to represent different taxa. Clarifica-tion of this problem requires more specimens from the typelocality to be analysed but this exceeds the limits of thispaper.
In LM, our specimens might also be confused with B.ocalanensis Shayler & Siver (Table 2), but this species dif-fers from B. manfredii in having a slightly heteropolar valveoutline and higher stria density (Shayler & Siver 2004a).Also, the central area in B. ocalanensis is poorly developedand fusiform, and the terminal raphe fissures are T-shapedwith straight branches.
Distribution. Brachysira manfredii is reported fromtropical environments of Australia, Africa and America. Itwas also recorded from Sierra Nevada, Colombia (Lange-Bertalot & Moser 1994). It was found in the Ayapelsystem.
Ecology. This species was found epiphytic on Eichor-nia azurea in permanent swamps, with high water tem-perature (29.9–32.0◦C), slightly acid pH (6.2–6.8), lowconductivity (13.8–89.5 μS cm−1) and low transparency(0.75–1.17 m).
Dow
nloa
ded
by [
Yim
my
Mon
toya
-Mor
eno]
at 1
2:09
10
Apr
il 20
14

Brachysira (Naviculales, Bacillariophyceae) in lowland waters 157
Figs 37–42. Brachysira microcephala, LM (Figs 37–38) and SEM external views (Figs 39–42). Figs 37–38. Valves of different size. Fig.39. Whole valve. Fig. 40. Frustule showing ribs slightly prominent, presence of papillae and T-shaped distal raphe end. Fig. 41. Detail ofthe valve centre. Fig. 42. Detail of the valve end. Figs 37, 39. Specimens from Permanent Swamp Escobillitas. Figs 38, 40–42. Specimensfrom Permanent Swamp Ayapel. Scale bars = 5 μm (Figs 37–40), 2 μm (Figs 41–42).
Brachysira microcephala (Grunow) Compère (Figs37–42)Description. Valves broadly rhombic–lanceolate, valveface flat with rounded to rostrate ends (Figs 37–40).Uniseriate striae, radiate throughout the valve, with oneinterposed stria at each side of the valve centre (Figs 39–41). Striae with 3 transapically elongated areolae (Fig.41), two towards the poles, the 10–12 apical striae withonly one elongated areola (Fig. 42). Areolae irregularlyspaced, delimiting a hyaline longitudinal line that follows
the outline of the axial and central areas, with 1–2undulate and anastomosing short lines visible at valvecentre with LM (Figs 37–38). Interstriae with irregularlyspaced papillae, 4–5 at the centre of each interstria (Figs40–41). Raphe simple, filiform, externally slightly sinu-ous (Figs 39–40), proximal ends slightly curved, simple(Fig. 41); raphe fissure T-shaped with curved branches(Fig. 42). Central area small irregular and asymmetrical.Axial area narrow and straight. Mantle without papillae(Fig. 40).
Dow
nloa
ded
by [
Yim
my
Mon
toya
-Mor
eno]
at 1
2:09
10
Apr
il 20
14

158 Vouilloud et al.
Morphometric data (n = 17). Length: 20–25 μm;width: 5.0–6.5 μm; length-to-width ratio: 3.4–4.3; 32–35striae in 10 μm; D(pre): 1.2–1.8 μm.
Studied material. LPC 9939, 9947 and 9992.Remarks. The studied specimens agree with the lec-
totype of B. microcephala described and illustrated byCompère (1988), except in their lower stria density andbroader valves (Table 2).
Distribution. This species, considered cosmopolitan, isrecorded for the first time in Colombia, from the Ayapelsystem.
Ecology. This species was found epiphytic on Eichor-nia azurea in swamps and rivers, with high watertemperature (30–32◦C), slightly acid pH (5.9–6.6), lowconductivity (25.8–86.2 μ S cm−1) and low transparency(0.21–0.80 mm).
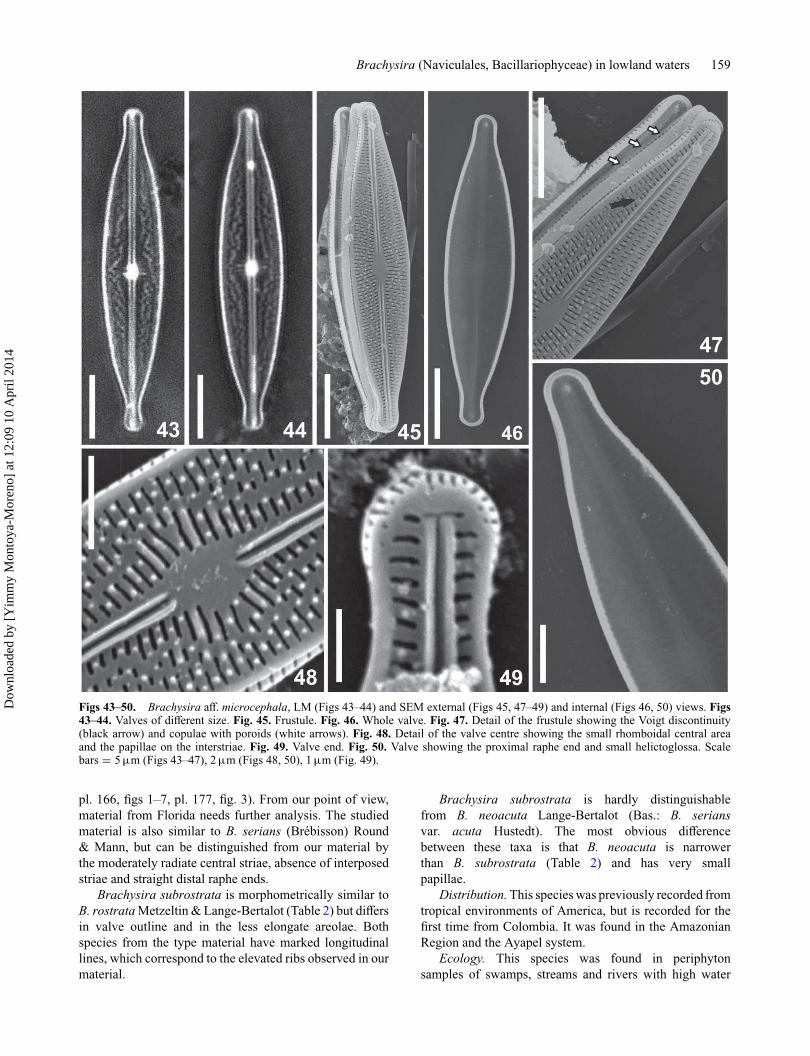
Brachysira aff. microcephala (Grunow) Compère (Figs43–50)Description. Valves lanceolate with capitate ends, valveface flat (Figs 43–46). Mantle with one row of elongatedareolae, aligned with the valve face striae (Fig. 47). Striaeuniseriate, slightly radiate at valve centre with 1–2 inter-posed striae on each side (Figs 45, 47), slightly radialtowards the ends where they are parallel. Voigt disconti-nuity revealed by a short stria that marks off the change instria position on the secondary valve side (Fig. 47, blackarrow). Areolae round to transapically elongate, irregularlyspaced, frequently 2 per stria at valve centre (Figs 47–48),then 3–4 smaller areolae per stria, two towards the apicesand finally, one elongate areola in the last 8–10 striae (Figs47–49). Areolae closed by internal hymens (Figs 46, 50).Sinuous, anastomosing hyaline longitudinal lines are visiblein LM (Figs 43–44). Interstriae with irregular spaced spinesor papillae, 3–5 at valve centre (Fig. 48). Raphe simple,externally slightly sinuous with straight, simple proximalends, T–shaped distal ends (Figs 47–49); internally prox-imal ends simple, distal ends straight, helictoglossae nearthe apices and very small, (Figs 46, 50). Central area cir-cular, asymmetric, delimited by longer areolae that othersin the same stria; axial area narrow, straight, narrower onthe secondary side (Fig. 45, 47). Girdle with 4 copulae,valvocopula with a row of poroids (Fig. 47, white arrows).
Morphometric data (n = 21). Length: 20–27 μm;width: 4.5–5.0 μm, length-to-width ratio: 4.3–5.4; 32–34striae in 10 μm; D(pre): 1.0–1.7 μm; copulae with 21–23poroids in 10 μm.
Studied materials. LPC 5512 and 5608 (Amazonia).Remarks. The studied specimens coincide with the type
material of B. microcephala in dimensions and valve struc-ture, but differ in valve outline (lanceolate with capitateto subcapitate ends), lower stria density and central striaecomposed of 2 areolae. Although these differences are insuf-ficient to consider the Amazonian specimens as a variety,
it is important to point out that these characters remainconstant within the analysed population and well differen-tiated from populations collected from Ayapel that agreewith the type of the species.
Distribution. This species was only found in the AmazonRegion.
Ecology. This species was found in periphyton sam-ples of streams and swamps with high water temperature(26.0–29.8◦C), acid pH (4.8–5.3), very low conductivity(10–12 μ S cm−1) and low transparency (0.4–0.6 m).
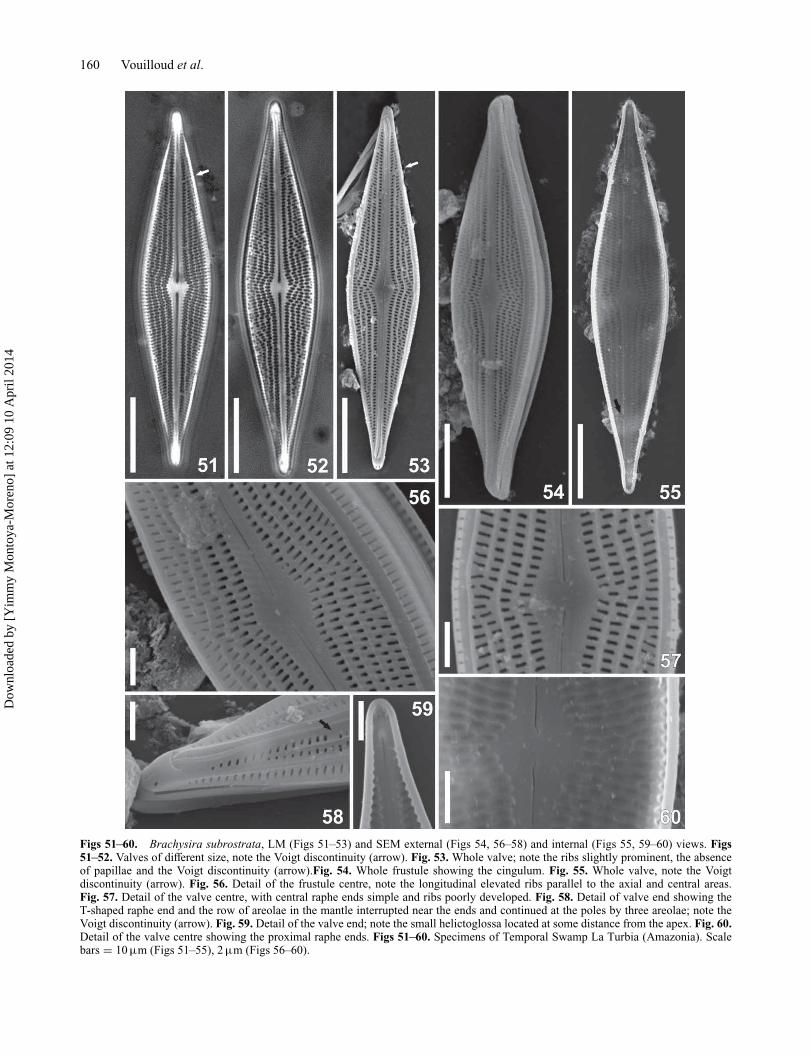
Brachysira subrostrata Lange-Bertalot (Figs 51–60)Description. Valves rhombic with rostrate ends. Mantlewith one row of elongate areolae aligned with the striae.Uniseriate striae strongly radiate at the valve centre with 2–3 interposed striae, moderately radial throughout the rest ofthe valve. Areolae circular to transapically elongate, more orless irregularly spaced, internally closed by hymens. Striaewith 4–6 areolae at valve centre. Interstriae without papillae.Voigt discontinuity marked by the absence of an areola onthe secondary side of the valve. Elevated longitudinal ribsparallel to the axial and central areas, three at the valve cen-tre, one at the valve ends. Central area small, rhomboidal,irregular; axial area moderately broad and straight. Exter-nally, raphe straight, bordered by two ribs that are poorlydeveloped near the proximal ends but prominent towardsthe apices; central and distal raphe ends curved gently tothe primary valve side; central raphe ends simple, termi-nal fissures Tshaped with curved branches. Internally raphestraight with proximal ends simple and straight and distalones ending in small helictoglossae at some distance fromthe apices. Mantle without papillae, with a row of elongateareolae that is interrupted near the ends but continued with3 areolae around the poles. Girdle bands without pores.
Morphometric data (n = 10). Length: 47.5–53.5 μm,width: 9.5–11.5 μm, length-to-width ratio: 4.4–5.2; 21–24striae in 10 μm; D(pre): 1.2–1.4 μm (externally) and 2.2 μm(internally).
Studied material. LPC 5608 and 5610 (Amazonia);LPC 9992 and 9999 (Ayapel).
Remarks. Brachysira subrostrata was erected byLange-Bertalot & Moser (1994) based on Anomoeoneisserians var. rostrata Krasske, which was collected froma bog in Brazil (Lange-Bertalot et al. 1996, Krasske Col-lection, preparation D IV 125). Although the type materialwas not analysed with SEM, our specimens are consid-ered to correspond to this taxon because morphometric data(Table 2) and characters visible with LM coincide withthe description given by Lange-Bertalot & Moser (1994).Metzeltin & Lange-Bertalot (2007) illustrate specimensfrom Florida (USA) as B. subrostrata, thereby expandingthe size range (Table 2). The latter specimens differ from thetype as they have slightly radiate striae at the centre, with-out interposed striae (cf., Lange-Bertalot & Moser 1994,
Dow
nloa
ded
by [
Yim
my
Mon
toya
-Mor
eno]
at 1
2:09
10
Apr
il 20
14
Brachysira (Naviculales, Bacillariophyceae) in lowland waters 159
Figs 43–50. Brachysira aff. microcephala, LM (Figs 43–44) and SEM external (Figs 45, 47–49) and internal (Figs 46, 50) views. Figs43–44. Valves of different size. Fig. 45. Frustule. Fig. 46. Whole valve. Fig. 47. Detail of the frustule showing the Voigt discontinuity(black arrow) and copulae with poroids (white arrows). Fig. 48. Detail of the valve centre showing the small rhomboidal central areaand the papillae on the interstriae. Fig. 49. Valve end. Fig. 50. Valve showing the proximal raphe end and small helictoglossa. Scalebars = 5 μm (Figs 43–47), 2 μm (Figs 48, 50), 1 μm (Fig. 49).
pl. 166, figs 1–7, pl. 177, fig. 3). From our point of view,material from Florida needs further analysis. The studiedmaterial is also similar to B. serians (Brébisson) Round& Mann, but can be distinguished from our material bythe moderately radiate central striae, absence of interposedstriae and straight distal raphe ends.
Brachysira subrostrata is morphometrically similar toB. rostrata Metzeltin & Lange-Bertalot (Table 2) but differsin valve outline and in the less elongate areolae. Bothspecies from the type material have marked longitudinallines, which correspond to the elevated ribs observed in ourmaterial.
Brachysira subrostrata is hardly distinguishablefrom B. neoacuta Lange-Bertalot (Bas.: B. seriansvar. acuta Hustedt). The most obvious differencebetween these taxa is that B. neoacuta is narrowerthan B. subrostrata (Table 2) and has very smallpapillae.
Distribution. This species was previously recorded fromtropical environments of America, but is recorded for thefirst time from Colombia. It was found in the AmazonianRegion and the Ayapel system.
Ecology. This species was found in periphytonsamples of swamps, streams and rivers with high water
Dow
nloa
ded
by [
Yim
my
Mon
toya
-Mor
eno]
at 1
2:09
10
Apr
il 20
14
160 Vouilloud et al.
Figs 51–60. Brachysira subrostrata, LM (Figs 51–53) and SEM external (Figs 54, 56–58) and internal (Figs 55, 59–60) views. Figs51–52. Valves of different size, note the Voigt discontinuity (arrow). Fig. 53. Whole valve; note the ribs slightly prominent, the absenceof papillae and the Voigt discontinuity (arrow).Fig. 54. Whole frustule showing the cingulum. Fig. 55. Whole valve, note the Voigtdiscontinuity (arrow). Fig. 56. Detail of the frustule centre, note the longitudinal elevated ribs parallel to the axial and central areas.Fig. 57. Detail of the valve centre, with central raphe ends simple and ribs poorly developed. Fig. 58. Detail of valve end showing theT-shaped raphe end and the row of areolae in the mantle interrupted near the ends and continued at the poles by three areolae; note theVoigt discontinuity (arrow). Fig. 59. Detail of the valve end; note the small helictoglossa located at some distance from the apex. Fig. 60.Detail of the valve centre showing the proximal raphe ends. Figs 51–60. Specimens of Temporal Swamp La Turbia (Amazonia). Scalebars = 10 μm (Figs 51–55), 2 μm (Figs 56–60).
Dow
nloa
ded
by [
Yim
my
Mon
toya
-Mor
eno]
at 1
2:09
10
Apr
il 20
14

Brachysira (Naviculales, Bacillariophyceae) in lowland waters 161
temperature (26.5–32.0◦C), slightly acid pH (5.1–6.7),variable conductivity (8–102 μ S cm−1) and low trans-parency (0.18–0.65 m).
Discussion and conclusionsDuring this study of plankton and periphyton from lenticand lotic waterbodies of Colombian Amazonia, and froma system of shallow swamps in northeastern Colombia,11 morphotypes within the genus Brachysira were found.Two new taxa, B. guarrerai and B. huitotarum, weredescribed and the morphology and affinities of B. man-fredii, B. microcephala, B. subrostrata, B. aff. brebissoniiand B. aff. microcephala were discussed. The other fourmorphotypes are not included here, although they seem tobe new taxa. However, they were very scarce and morespecimens should be examined to confirm this.
Brachysira guarrerai and B. huitotarum were differenti-ated from their allies on the basis of: the shape of the centralarea, the shape and number of the areolae that delimit thecentral area, stria position, presence or absence of papil-lae on the interstriae and degree of silicification of the ribsthat delimit the raphe. Brachysira guarrerai differs from B.garrensis in its elliptical, asymmetrical central area, parallelstriae with 2–3 areolae at the valve centre, longer areolaebordering the central area on the primary valve side, oneor two undulate, anastomosing hyaline longitudinal linesdelimited by irregularly spaced areolae, interstriae withoutpapillae and raphe with slightly thickened siliceous ribs.Brachysira huitotarum differs from B. arctoborealis andB. microcephala in its rhombic, slightly irregular, asym-metric central area, strongly radiate striae at valve centrewith an interposed stria, striae around the central area withtwo areolae, towards the apices with three areolae (the cen-tral shorter) interposed with striae comprising two areolae,and with sinuous hyaline longitudinal lines, single at thevalve centre dividing in two towards the apices and thenanastomosing at the very end.
To avoid generating more confusion within the genus,our discussion is based on the comparison of the studiedspecimens with illustrations of type material, but this wasnot always possible as the latter is sometimes mountedin an inappropriate medium and/or is poorly illustrated.Brachysira brebissonii is one of the most widely distributedtaxa of the genus. Although Wolfe & Kling (2001) studiedthe isotype of Navicula brachysira (= B. brebissonii), thespecies concept in the literature is confusing and severaltaxa have been misidentified as this species. The specimenillustrated in Lange-Bertalot & Moser (1994, pl. 41, fig. 3)is the one that corresponds to the isotype shown by Wolfe &Kling (2001). Nevertheless, this problem has not been dis-cussed or even mentioned in recent publications, e.g., thenomenclatural analysis of N. brachysira and its synonymyto B. brebissonii sensu Lange-Bertalot & Moser in relationto specimens from Ocala National Forest (Shayler & Siver
2004a). Our specimens coincide with those from Colom-bia described as B. brebissonii morphotype brebissonii inLange-Bertalot & Moser (1994). Both differ from the typematerial in having strongly radiate central striae with 3–4areolae and in their stria density. Although the central are-olae are wider and form a longitudinal line, as in the typematerial, there are 2–3 smaller areolae towards the apicesin our specimens. Furthermore, only a few specimens werefound and we, therefore, consider it more appropriate to des-ignate them B. cf. brebissonii than to recognize a new taxon.
Brachysira microcephala is another taxon that isdescribed as cosmopolitan and recorded from various envi-ronments. The type material of this species was studiedin detail by Compère (1988). Material collected in Ayapelagrees with the lectotype, but specimens from Amazoniadiffer from it in outline and stria density, and have centralstriae composed of two areolae. They are therefore recordedas B. aff. microcephala until populations from more sites canbe analysed with SEM.
Brachyira huitotarum, B. aff. brebissonii and B. micro-cephala might easily be confused under LM, but they differin the shape of the central area, shape and number of areolaeper stria, characteristics of the central striae and longitudinallines (compare Figs 2–5; 15–17, 24; 37–39; 41). Brachysiraguarrerai and B. aff. microcephala are similar in valve out-line, but the former is broader, has more acute apices andparallel central striae.
Although Round et al. (1990), Compère (1988),Lange-Bertalot & Moser (1994), Wolfe & Kling (2001) andShayler & Siver (2004a, b) broaden the limits of the genus,it is necessary to consider other characters if all the speciesthat the genus currently comprises are included. Round &Mann (1981) stated that the girdle is composed of severalcopulae without pores. Later Round et al. (1990) pointedout that advalvar bands sometimes have one row of poroids.Brachysira huitotarum and B. aff. microcephala have a rowof pores on the valvocopula. In addition to the valve out-line, length, width, length-to-width ratio and stria density,Shayler & Siver (2004a, b) pointed out that the number ofareolae per stria, areola size and the presence of papillaeor spines are important characters. Features of the centralarea are also considered by us to be important, particularlythe distribution, size and shape of the central striae and thedistance between the proximal raphe ends.
Considering geographic distribution, B. brebissonii andB. microcephala are described as cosmopolitan, B. man-fredii was found in Australia, Africa and America, and B.subrostrata only in the neotropics. As for other diatomspecies, the apparent cosmopolitan distribution of someBrachysira spp. may be due to misidentifications, and/orthe lack of morphological and comparative studies of pop-ulations from distant areas. This problem has risen in otherstudies of other freshwater species from the same regions(Sala et al. 1993, 1998). Undoubtedly comparing popu-lations from America and other parts of the world withmolecular tools will provide additional evidence to help
Dow
nloa
ded
by [
Yim
my
Mon
toya
-Mor
eno]
at 1
2:09
10
Apr
il 20
14

162 Vouilloud et al.
understand the taxonomy and biogeography of neotropicaldiatoms.
Brachysira spp. were found in both lotic and lentic waterbodies in the study area. All sites in which the species werefound are oligotrophic, with acid to circumneutral pH, verylow or low conductivity and high turbidity and temperature.Brachysira brebissonii seems to have a wide distributionin Colombia, as previously reported by Lange-Bertalot &Moser (1994), Donato (2001), Martínez & Donato (2003)and Vélez et al. (2005 as A. brachysira), but consideringthe confusion over the identity of the species, these recordsshould be revisited. Brachysira guarrerai and B. aff. micro-cephala were found only in the Amazon Region, whereas B.manfredii and B. microcephala were only recorded from theAyapel system. Brachysira aff. brebissonii, B. huitotarumand B. subrostrata were found in both geographic areas.Although the studied sites have similar physical and chem-ical characteristics, differences in the diatom flora betweenthe two regions were also observed in a study that consid-ered the geographic distribution of diatoms in Colombia(S.E. Sala, unpubl. data).
AcknowledgementsThe authors wish to thank the Instituto Amazónico de Investi-gaciones Científicas Sinchi, Red Latinoamericana de Botánica,Convención Ramsar and the Instituto Nacional de Desarrollo(INADE) de Perú. We also want to specially thank Luc Ector forhis generosity in assisting with this work and Sonia Sua from theSinchi Institute who kindly prepared the map of the study area.
ReferencesAnonymous. 1975. Proposals for a standardization of diatom ter-
minology and diagnosis. Beihefte zur Nova Hedwigia 53:323–354.
CEN [Comité européen de normalisation]. 2003. Water quality –Guidance standard for the routine sampling and pretreatmentof benthic diatoms from rivers. – EN 13946, 2003. ComitéEuropéen de Normalisation, Geneva.
Compère P. 1988. Brachysira microcephala (Grunow) Com-père nom correct de “Navicula exilis” Grunow 1860 nonKützing 1844. Mémoires de la Société Royale de Botaniquede Belgique 10: 9–11.
Donato J.Ch. 2001. Fitoplancton de los lagos andinos del Norte deSuramérica (Colombia): Composición y Factores. ColecciónJorge Álvarez Lleras, 19. Academia Colombiana de CienciasExactas, Físicas y Naturales. Bogotá. 231pp.
Guiry, M.D. & Guiry, G.M. 2013. AlgaeBase. World-Wide Elec-tronic Publication, National University Of Ireland, Galway.Available from: http://www.algaebase.org [Accessed 13September 2013].
Hasle, G.R. & Fryxell, G.A. 1970. Diatoms: cleaning and mount-ing for light and electron microscopy. Transactions of theAmerican Microscopical Society 89: 469–474.
Kützing F.T. 1836. Algarum aquae dulcis germanicarum DecasXVI. pp. Halis Saxonum [Halle]: in commissis C.A.Schwetschkii et fil. 25 pp. Halle.
Kützing F.T. 1844. Die Kieselschaligen Bacillarien oderDiatomeen. Nordhausen: zu finden bei W. Köhne.152 pp.Nordhausen.
Lange-Bertalot H. & Moser G. 1994. Brachysira. Monogra-phie der Gattung. Wichtige Indikator Species für dasGewässer Monitoring und Naviculadicta nov. gen. Biblio-theca Diatomologica 29: 1–212.
Lange-Bertalot H., Külbst K., Lauser T., Nörpel-Schempp M.& Willmann M. 1996. Diatom taxa introduced by GeorgKrasske. Documentation and revision. Iconographia Diato-mologica 3: 1–358.
Martínez L.F. & Donato J. 2003. Efectos del caudal sobre la colo-nización de algas en un río de alta montaña tropical (Boyacá,Colombia). Caldasia 25: 337–354.
Metzeltin D. & Lange-Bertalot H. 1998. Tropical diatoms of SouthAmerica I. Iconographia Diatomologica 5: 1–695.
Metzeltin D. & Lange-Bertalot H. 2007. Tropical diatoms ofthe South America II. Iconographia Diatomologica 18:1–877.
Montoya-Moreno Y., Sala S., Vouilloud A.A. & Aguirre N.2011. Capartogramma crucicula (Grunow ex Cleve) Ross,un nuevo registro del género para Colombia. UniversitasScientiarum 16: 70–76.
Montoya-Moreno, Y., Sala, S.E., Vouilloud, A.A. & Aguirre, N.2012. Diatomeas (Bacillariophyta) perifíticas del complejocenagoso de Ayapel, Colombia. I. Caldasia 34: 457–474.
Ross R., Cox E.J., Karayeva D.G., Mann D.G., Paddock T.B.B.,Simonsen R. & Sims P.A. 1979. An amended terminologyfor the siliceous components of the diatom cell. Beihefte zurNova Hedwigia 64: 513–533.
Round F.E. & Mann D.G. 1981. The diatom genus Brachysira 1.Typification and separation from Anomoeoneis. Archiv fürProtistenkunde 124: 221–231.
Round F.E., Crawford R.M. & Mann D.G. 1990. The diatoms. biol-ogy and morphology of the genera. Cambridge UniversityPress, Cambridge. 747 pp.
Sala S.E. & Ramirez J.J. 2008. Cyclotella katiana nov. sp.(Bacillariophyceae) from La Reina Swamp, Parque Nacionallos Katíos, Colombia. Diatom Research 23: 147–157.
Sala S.E., Guerrero J.M. & Ferrario M.E. 1993. Redefinition ofReimeria sinuata and recognition of Reimeria uniseriata nov.spec. Diatom Research 8: 439–446.
Sala S.E., Sar S.E. & Ferrario M.E. 1998. Review of the materialsmentioned in Argentina as Amphora coffeaeformis. DiatomResearch 13: 323–336.
Sala S., Duque S.R., Núñez-Avellaneda M. & Lamaro A.A. 1999.Nuevos registros de diatomeas (Bacillariophyceae) de laAmazonia Colombiana. Caldasia 21: 26–37.
Sala S., Duque S.R., Núñez-Avellaneda M. & Lamaro A.A. 2002a.Diatoms from the Colombian Amazonia. Cryptogamie, Algo-logie 23: 75–99.
Sala S., Duque S.R., Núñez-Avellaneda, M. & Lamaro A.A.2002b. Diatoms from the Colombian Amazon: some speciesof the genus Eunotia (Bacillaryophyceae). Acta Amazonica32: 589–603.
Sala S.E., Núñez-Avellaneda M. & Vouilloud A.A. 2008a. Ultra-structure of the frustule of Urosolenia species from theColombian and Peruvian amazon: U . delicatissima nov.spec., U . amazonica nov. spec. and U . braunii (Hustedt).Diatom Research 23: 159–169.
Dow
nloa
ded
by [
Yim
my
Mon
toya
-Mor
eno]
at 1
2:09
10
Apr
il 20
14

Brachysira (Naviculales, Bacillariophyceae) in lowland waters 163
Sala S.E., Ramirez J.J. & Plata Diaz Y. 2008b. Diatoms fromlentic and lotic systems in Antioquia, Chocó and San-tander Departments in Colombia. Revista de Biología Tropi-cal/International Journal of Tropical Biology 56: 1159–1178.
Shayler H.A. & Siver P.A. 2004a. Biodiversity of the genusBrachysira in the Ocala National Forest, Florida, U.S.A.In: Proceedings of the seventeenth international diatomsymposium (Ed. by M. Poulin), pp. 309–333. BiopressLimited, Bristol.
Shayler H.A. & Siver P.A. 2004b. Description of a new species ofthe diatom genus Brachysira (Bacillariophyta), Brachysiragravida sp. nov. from the Ocala National Forest, Florida,U.S.A. Nova Hedwigia 78: 399–409.
Simonsen R. 1987. Atlas and catalogue of the diatom types ofFriedrich Hustedt. J. 3 volumes. Cramer. Stuttgart. 525 pp.,722 pls.
Vélez M.I., Berrío J.C., Hooghiemstra H. & Metcalfe S. 2005.Integrated diatom–pollen based Holocene environmentalreconstruction of Lake Las Margaritas, eastern savannahs ofColombia. The Holocene 15: 1184–1198.
Vouilloud A.A., Sala S.E., Núñez-Avellaneda M. & Duque S.R.2010. Diatoms from the Colombian and Peruvian Ama-zon: the genera Encyonema, Encyonopsis and Gomphonema(Cymbellales, Bacillariophyceae). Revista de BiologíaTropical/International Journal of Tropical Biology 58:45–62.
Wolfe A.P. & Kling H.J. 2001. A consideration of some NorthAmerican soft-water Brachysira taxa and description ofB. arctoborealis sp. nov. In: Lange-Bertalot-Festschrift:Studies on diatoms (Ed. by R. Jahn, J.P. Kociolek,A. Witkowski and P. Compère), pp. 243–264. Koeltz,Koenigstein.
Dow
nloa
ded
by [
Yim
my
Mon
toya
-Mor
eno]
at 1
2:09
10
Apr
il 20
14




















