
Utilisation of carrion by Silphidae (Coleoptera) on lowland heathland, England.
38
1 1 Utilisation of carrion by Silphidae (Coleoptera) on lowland heathland, England. 2 R. I. M. Guy* 3 4 * Corresponding Author: [email protected] 5 6 Abstract: 7 1. The aims of this study were a) to identify Silphidae species utilising carrion in heathland habitats, b) to 8 determine if their use of different stages of heathland vegetation (open areas and mature heather 9 specifically) differed and c) to record and analyse the colonisation order of the species observed. 10 2. Heathland research frequently focuses on specialist species restricted to these habitats, while more 11 generalist species are frequently omitted. A gap in the published literature was observed regarding 12 invertebrate communities associated with carrion in heathland habitats and this study aimed to begin to 13 fill that gap by focussing on Silphidae, a group of beetles with many necrophagous species. 14 3. Silphidae were sampled using pit-fall traps at 32 sites baited with carrion (Rabbit carcases) split equally 15 between the two vegetation conditions. Each site was visited 3 times over the course of a 6 day period to 16 observe changes in species assemblage over the course of decomposition. 17 4. 940 Silphidae were found, from 10 species. Overall there was not a significant difference in abundance 18 between vegetation types; on an individual species level a majority of species were shown to be 19 significantly more abundant in heather areas. Heather hosted a more diverse range of species. Significant 20 differences were also observed in species succession of carrion. 21 5. These results fill gaps in Silphidae ecological understanding, especially how habitat features are used. 22 Some of the data may be of use in the field of forensic entomology. Perhaps most importantly portions of 23 Silphidae ecology which are unknown or poorly understood have been identified and suggestions for 24 further study made. 25 26 Key Words: Necrophagous Coleoptera, Vegetation structure, Entomofaunal Succession, Heathland, 27 Preferential Habitat Selection. 28
Transcript of Utilisation of carrion by Silphidae (Coleoptera) on lowland heathland, England.

1
1
Utilisation of carrion by Silphidae (Coleoptera) on lowland heathland, England. 2
R. I. M. Guy* 3
4
* Corresponding Author: [email protected] 5
6
Abstract: 7
1. The aims of this study were a) to identify Silphidae species utilising carrion in heathland habitats, b) to 8
determine if their use of different stages of heathland vegetation (open areas and mature heather 9
specifically) differed and c) to record and analyse the colonisation order of the species observed. 10
2. Heathland research frequently focuses on specialist species restricted to these habitats, while more 11
generalist species are frequently omitted. A gap in the published literature was observed regarding 12
invertebrate communities associated with carrion in heathland habitats and this study aimed to begin to 13
fill that gap by focussing on Silphidae, a group of beetles with many necrophagous species. 14
3. Silphidae were sampled using pit-fall traps at 32 sites baited with carrion (Rabbit carcases) split equally 15
between the two vegetation conditions. Each site was visited 3 times over the course of a 6 day period to 16
observe changes in species assemblage over the course of decomposition. 17
4. 940 Silphidae were found, from 10 species. Overall there was not a significant difference in abundance 18
between vegetation types; on an individual species level a majority of species were shown to be 19
significantly more abundant in heather areas. Heather hosted a more diverse range of species. Significant 20
differences were also observed in species succession of carrion. 21
5. These results fill gaps in Silphidae ecological understanding, especially how habitat features are used. 22
Some of the data may be of use in the field of forensic entomology. Perhaps most importantly portions of 23
Silphidae ecology which are unknown or poorly understood have been identified and suggestions for 24
further study made. 25
26
Key Words: Necrophagous Coleoptera, Vegetation structure, Entomofaunal Succession, Heathland, 27
Preferential Habitat Selection. 28

2
1. Introduction 29
Lowland Heathland 30
The area of lowland heathland in the UK has declined significantly over the last few centuries (Webb, 31
1989; Rackham, 2003); despite this UK heathland areas represents 20% of remnant European 32
heathland (JNCC, Undated). While a range of heathland types are recognised, lowland heathland is 33
generally considered to be a semi-natural mosaic of sub-climax vegetation stages, historically 34
prevented from succeeding to scrub woodland by human influences such as livestock grazing, peat-35
cutting and fuel wood harvesting (Webb, 1989; Gardner, 1991; Rackham, 2003). Dwindling commercial 36
use for these areas led to habitat degradation and a resulting loss of specialist species diversity 37
(Symes & Day, 2003; Cuesta et al., 2006), with many heathland sites destroyed through conversion to 38
agriculture or commercial forestry plantation (Rackham, 2003; Lin et al., 2007). Remnant areas are 39
often fragmented and isolated and many have deteriorated through lack of appropriate 40
management making these areas conservation priorities (Symes & Day, 2003). Statutory protection 41
has been granted for heathland across Europe and the UK. Five types of lowland heathland are listed 42
in Annex 1 of the European Habitats Directive (1992) and in the UK lowland heath was a BAP priority 43
habitat with 67% of sites designated as Sites of Special Scientific Interest (SSSI) (JNCC, Undated). 44
Current management methods are well established and include conservation grazing, scrub and 45
invasive species removal, rotational heather cutting or burning and turf stripping, among others 46
(Sutherland & Hill, 1995; Symes & Day, 2003). The heterogeneity these methods promote is essential in 47
providing the range of niches and habitat features occupied by heathland specialists (Kirby, 2001; 48
Symes & Day, 2003). Substantial guidance is available from governmental bodies and NGO’s alike on 49
appropriate management of these areas (e.g. Buglife, Undated; JNCC, Undated; Symes & Day, 2003 (RSPB); 50
Anon., 2013 (Buglife)). Projects to restore or recreate favourable conditions on former heathland sites, 51
now degraded or destroyed, are achieved by managing or manipulating soil conditions and 52
vegetation succession (Aerts et al. 1995; Van den Berg et al., 2003; Hawley et al., 2008). 53

3
Heathland is sometimes considered a species poor habitat (Gardner, 1991). Webb & Hopkins (1984a) 54
suggest that high invertebrate species diversity indicates habitat deterioration (also Symes & Day, 55
2003). Specialist heathland species - including plant, invertebrate, bird and reptile species - often 56
have restricted distributions or specialist habitat requirements (Kirby, 2001; Symes & Day, 2003). While 57
heathland communities may be lower in species diversity than other habitat specific communities 58
the restricted range and precise requirements of many of these species makes the conservation of 59
their habitat essential (Webb, 1989; Symes & Day, 2003; Lin et al., 2007). 60
Heathland is of particular importance to invertebrates: of 133 BAP priority species found on 61
heathland in the UK, 82 are invertebrates and of these 47 are restricted or very restricted to 62
heathland (Webb et al., 2010). While UK BAP’s were superseded in 2010, the priority species lists are 63
still used to inform devolved conservation priority planning (JNCC, 2014). Kirby (1994) lists 133 Red 64
Data Book species and 210 scarce (notable) species of invertebrates associated with lowland 65
heathland. High quality heathland provides a diverse range of niches catering for an equally diverse 66
range of associated specialist species; many studies into heathland ecology and management focus 67
on the ecology and conservation of these species (e.g. Webb & Hopkins, 1984a; Gardner, 1991; Blake et 68
al., 2003; Dennis, 2003; Schirmel & Buchholz, 2011). While some of these studies appear repetitive they 69
nevertheless provide significant evidence for the importance of heterogeneity in heathland 70
environments (Price, 2003; Schirmel, 2010; Schirmel & Buchholz, 2011). 71
Habitat characteristics which are of significance to heathland invertebrates include: 72
- heterogeneous vegetation structure, 73
- areas of exposed soil (especially sandy soils), 74
- warm micro-climates associated with bare ground or very short swards, 75
- nectar resources from characteristic heathland flora, 76
- complex architectural structure of shrub and scrub species, 77
- soil moisture, 78

4
- the gradients between extremes in the above features. 79
(Buglife, Undated; JNCC, Undated; Gardner, 1991; Usher 1992; Kirby, 2001; Symes & Day, 2003; Kirby, 2004; 80
Cuesta et al., 2006; Schirmel & Buchholz, 2011; Anon., 2013). 81
In contrast to upland heather moors, few lowland heathland species rely on heather as a food 82
resource (Kirby, 2001), although many species utilise it to some degree. In heathland settings it is 83
often of greater importance as an architectural feature, particularly to web spinning spiders (Webb & 84
Hopkins, 1984a). The effectiveness of different heathland management techniques, for the 85
conservation of favourable habitat conditions and heathland species, has been widely studied (Usher, 86
1992; Dennis, 2003; Lin et al., 2007; Schirmel, 2010). 87
Many of the above studies are focussed upon the same groups of invertebrates: ground beetles 88
(Carabidae: Coleoptera) and spiders (Araneae) being the most common and frequently studied 89
together (Webb & Hopkins, 1984ab; Gardner, 1991; Blake et al., 2003; Cuesta et al., 2006; Lin et al., 2007; 90
Schirmel & Buchholz, 2011). Aside from the specialist fauna of heathlands, they also provide important 91
resources for generalist species (Kirby, 2001; Symes & Day, 2003). Nectar resources are used by a 92
diverse range of species, especially at times of year when they are restricted elsewhere (Anon., 2013). 93
Some species will disperse onto heathland from surrounding habitats to exploit resources (Webb et 94
al., 1984). High densities of invertebrates, both generalist and specialist, attract predatory species 95
which are found in abundance (Kirby, 2001). 96
These generalist species receive little attention in heathland specific research and management 97
guidance: with such a diverse specialist fauna this prioritisation is understandable. The nature of 98
heathland habitats, with complex mosaics of different vegetation structures, can provide a useful 99
experimental setting in which to study the habits of generalist species. The complex succession of 100
heathland vegetation frequently produces succession gradients: Schirmel & Buchholz (2011) used 101
this graduating vegetation community to their advantage by studying invertebrates in five different 102

5
stages of succession on the same site along a succession gradient and demonstrated significantly 103
different invertebrate assemblages in different stages of vegetation. 104
Carrion Beetles (Silphidae) 105
Heathlands have a close association, both historically and currently, with animals both wild and 106
domestic (Rackham, 2001; Dennis, 2003). It is therefore reasonable to hypothesise that heathlands 107
would support invertebrate communities associated with animals, including dung and carrion. 108
Several publications have alluded to the invertebrate communities associated with both carrion and 109
dung in heathlands being ‘rich’ (Kirby, 2001 pg. 77; see also Buglife, Undated) without reference to 110
published research or any description of these communities. Studies into these communities are not 111
readily available, if they have been conducted at all. 112
This research gap provides an opportunity to engage in original research into relatively well-known 113
species in well-known habitats. The chosen group for this study is Silphidae (Coleoptera), commonly 114
known as carrion beetles. Studies into this group most commonly fall into one of two categories: 1) 115
necrophagous (carrion feeding) species in this group are included in forensic entomology research; 116
2) the social behaviour of Nicrophorinae, a sub-family of Silphidae. 117
Necrophagous invertebrates are useful in forensic investigations for determining approximate time 118
of death or minimum post-mortem interval (PMImin) (Byrd & Castner, 2010; Huffman & Wallace, 2012). 119
One primary method for the calculation of this period is entomofaunal succession - the predictable 120
sequence in which different species colonise carrion (Dekeirsschieter et al., 2011b; Huffman & Wallace, 121
2012), this is widely studied for a range of invertebrate groups (e.g. Watson & Carlton, 2003; Watson & 122
Carlton, 2005; Sharanowski et al., 2008). Different species of necrophagous invertebrate specialise in 123
colonising carrion at different stages of the decomposition process: thus through a comparison of 124
observed species assemblages with the behavioural habits of the species found it is possible to 125
determine, in association with other recorded environmental variables, an approximate PMImin 126

6
(Kulshretha & Satpathy, 2001; Midgley et al., 2010). In this field Diptera are far more widely studied 127
because they are considered to provide a more accurate estimation of PMImin due to faster 128
colonisation, greater exhibition of niche specialisation and a greater range of species (Huffman & 129
Wallace, 2012), but Coleoptera can provide useful information, particularly in later stages of 130
decomposition (Kulshretha & Satpathy, 2001). Existing studies must be applied with caution however, 131
as geographical, seasonal and environmental variables can all result in significantly different species 132
assemblages and colonisation patterns (Chapman & Sankey, 1955; Grassberger & Frank, 2004; 133
Dekeirsschieter et al., 2011b). 134
These geographical variations are reasonably well understood, but finer scale differences between 135
or within specific habitats are less well studied. Some studies comparing necrophagous Coleoptera 136
communities in different habitats have been conducted, however the habitat parameters are often 137
very broad and poorly defined (e.g. ‘woodland, agricultural and urban areas’ compared by Dekeirsschieter 138
et al. (2011a)). Some studies have been conducted with greater focus on different types of a certain 139
habitat (one example is different types of woodland as studied by Matuszewski et al. (2008)), but literature 140
of this nature in the UK is lacking. The above studies show significant variation in species diversity 141
and abundance. 142
The other common area of Silphidae study is the behavioural ecology of Nicrophorinae (a sub-family 143
within Silphidae often known as ‘Burying beetles’) which exhibit co-operative breeding in the form of 144
bi-parental care for their larvae, a behaviour very rare in Coleoptera species (Dekeirsschieter et al., 145
2011b). 146
While not all Silphidae specialise in carrion feeding, the Nicrophorinae as well as a number of 147
Silphinae species (another subfamily) require carrion to compete their breeding cycle (Dekeirsschieter 148
et al., 2011b; Ray et al., 2014). This association takes two forms: direct carrion feeding and predation of 149
other invertebrates feeding on the carrion. Some species exhibit both these behaviours depending 150
on the stage of the life cycle (Chapman & Sankey, 1955; Dekeirsschieter et al., 2011b). In the UK there are 151

7
21 species of Silphidae (Booth, 2012). Of these 16 are known to be associated with carrion: all 7 152
Nicrophorinae and 9 Silphinae (Dekeirsschieter et al., 2011b). 153
Carrion is a rich albeit ephemeral resource and as such many organisms utilise it (Putman, 1983; 154
Wilson & Wilkovich, 2011), although many do so only as opportunists (Moreno-Opo & Margalida, 2013). 155
Seasonality is a significant factor in determining competition for carrion resources: during colder 156
seasons when invertebrates are inactive or less active, and in some geographical regions throughout 157
the year, vertebrate scavengers, primarily mammalian and avian, account for the majority of carrion 158
consumption (by biomass) (Putman, 1983; DeVault et al., 2004; DeVault et al., 2011). In warmer seasons 159
when necrophagous-specialist invertebrate species are active they contribute significantly to the 160
consumption of carrion, effectively competing with vertebrate scavengers (Putman, 1983). Some 161
estimates place invertebrate utilisation of carrion as high as 85% (Ray et al., 2014), with one study 162
(having excluded vertebrate scavengers) recording a 44kg carcase being consumed solely by 163
invertebrate and microbial activity in 3 weeks in a temperate environment (Grassberger & Frank, 164
2004). With this level of potential competition, both inter- and intra-specific, the ability to reduce 165
competitive pressure is an important mechanism (Milne & Milne, 1976; Putman, 1983) and provides 166
Nicrophorinae an important advantage in the utilisation of small vertebrate carcasses. 167
Decomposition of carrion progresses through four stages: Fresh, Bloated, Decay and Dry (Putman, 168
1983; Tantawi et al., 1996), although some consider the Decay stage in two separate stages - Active 169
and Advanced (Matuszewski et al., 2008; Castro et al., 2013), and count five stages overall. The onset of 170
these stages is affected by a huge range of environmental, climatic, faunal and bio-chemical factors 171
and as such is not consistent (Chapman & Sankey, 1955; Putman, 1983; Tantawi et al., 1996). Different 172
invertebrate species specialise in utilising carrion at different stages of the process which leads to 173
the observed entomofaunal succession described previously (Dekeirsschieter et al., 2011b). The two 174
subfamilies utilise carrion resources slightly differently, this will be discussed in detail in the context 175
of the study results. 176

8
Research Aims 177
The overall purpose of this study was to begin to describe a species groups (Silphidae) use of a 178
specialist resource (carrion) in a specific habitat (lowland heathland). With no previous work in this 179
area it is hoped that the findings of this study may act as a base point for further study into Silphidae 180
in a range of habitats to fill the research gaps into their ecology identified above. 181
The aims of this specific research project were: 182
1. Record Silphidae species which are present and their abundance. 183
2. Compare how Silphidae use areas of bare ground and mature heather. 184
3. Record the sequence of entomofaunal succession. 185

9
2. Materials and Methods 186
Study Site 187
The study was conducted on 90 hectares of SSSI designated heathland (Site of Special Scientific Interest 188
- Sutton & Hollesley Heaths; study site comprising blocks 11 & 12) located within the Suffolk Coast and 189
Heaths Area of Outstanding Natural Beauty (AONB), England (52.074’N, 1.388’E). The area is 190
representative of lowland dry heath (as described by Symes & Day (2003)), with acidic, sandy soil and 191
vegetation dominated by Calluna vulgaris, although unusually lacking in significant quantities of Ulex 192
europeaus (pers. comm.: B. Calversbert). The site is managed as a nature reserve with conservation 193
grazing by sheep and ponies alongside periodic heather cutting and bracken and scrub control 194
operations (pers. comm.: B. Calversbert). Natural grazing by Rabbit Oryctolagus cuniculus, and Fallow 195
Deer Dama dama and Muntjac Muntiacus reevesii (although only rabbits are present in high density) 196
is deemed beneficial in maintaining the desired complex mosaic of vegetation structure across the 197
site. 198
Open areas and areas dominated by mature heather were 199
chosen as two stages of heathland vegetation succession to 200
allow comparison for the research aims to be achieved (see 201
Box 2.1). Within the study area suitable sites (n = 32; see Fig 202
2.1) were identified using a combination of remote sensing 203
(satellite imagery: Google Maps) and site visits to ground truth 204
the satellite imagery (as a result of natural succession and 205
habitat management work the distribution of different 206
vegetation structures varies from year to year; satellite imagery was 2 years old so site visits were 207
essential). 208
Due to the nature of the site, placement of samples presented a problem. The site is used 209
extensively by recreational visitors as a result of open access rights, with dog walkers especially 210
Box 2.1 - Vegetation Type Description
Mature Heather - Vegetation dominated
by mature heather plants to a minimum
height of 25cm.
Bait point placement criteria:
≥ 80% ground cover mature heather with
height ≥ 25cm within 3m of bait point.
Open Ground - Bare soil or very short
sward height (≤5cm). Maintained through
grazing and / or management work.
Bait point placement criteria:
≥ 80% bare ground or close cropped sward
(≤ 5cm) within 3m of bait point.

10
Fig 2.1 - Study site and
individual bait point sites
common. It was decided that bait points would not be sited within obvious view of the paths to 211
reduce the risk of disturbance by members of the public and dogs. Bracken dominated the 212
vegetation in large areas of the site, further restricting where bait points could be located. Further, 213
due to the research aim to compare specific stages of heathland vegetation, large areas of the site 214
did not meet the required conditions (see Box 2.1). This precluded truly random site location. In an 215
attempt to gather representative data without ‘cherry picking’ sites, areas which met the two 216
vegetation conditions were identified and the total suitable area divided into 18 sites (two of which 217
remained unused due to bait shortages). Within open areas, traps were located by roughly dividing 218
the block into an approximate 10 x 10 grid with the bait site being placed in a randomly selected grid 219
square. In mature heather areas the exact location of each bait point was physically limited to a 220
point where the bait and exclusion cage could be installed (this usually required a space between 221
heather plants). Due to the psuedoreplication exhibited by these site selection methods (Hurlbert, 222
1984), care must be taken in applying the results of this analysis to other sites. 223

11
Carcase
Fig 2.2 - Pitfall trap arrangement, and
example of a bait point among heather
prior to the addition of a rabbit carcase.
Data Collection Methodology 224
Data collection was achieved using carrion bait (Rabbit 225
carcases: sourced from local pest control operation) with 226
pitfall traps. Bait was enclosed within wire exclusion cages 227
(460 x 330 x 200mm) to prevent disturbance by vertebrate 228
scavengers (Chick, 2008; Dekeirsschieter et al., 2011a). Mesh 229
size was approx. 30mm; the largest target species 230
maximum size is approx. 35mm length (Nicrophorus 231
humator), as such this mesh size should not have excluded 232
any target species from entering (Dekeirsschieter et al., 233
2011b). Four pitfall traps (plastic cups: approx. 50mm 234
diameter, 80mm deep) per bait point were located inside 235
the exclusion cage. Traps were orientated around the carcase with one each at the head and tail 236
ends, above the back and beneath the belly (see Fig 2.2) (based on Dekeirsschieter et al., 2011a). Each 237
bait point was checked, samples removed and traps refilled if necessary three times at 48 hr 238
intervals (+/- 2 hr) from initial placement of each carcase with the final check 6 days after initial 239
placement. Pilot studies showed that this time interval allowed most carcases to decompose to the 240
final ‘dry’ stage of decomposition (Tantawi et al., 1996). Putman (1983) showed that Silphidae were 241
most common during the ‘bloated’ and ‘decay’ stages with none found during the dry stage so this 242
was considered sufficient time for the purposes of this study. Due to the short interval between trap 243
checks no killing agent was used; traps contained water and a few drops of detergent only. The 244
content of all four traps at each bait point was considered as a single sample. Identification of 245
samples was achieved using a number of keys (Hackston, 2009; Wright, 2009ab, Hackston, 2012). 246
Due to the size of the species being dealt with no specialist equipment was required for 247
identification, with only a hand lenses being used to confirm diagnostic features in the smaller 248
species. 249

12
Data collection was undertaken over a three week period in August 2014. For the first two weeks 250
twelve bait points were run simultaneously - six per vegetation type; for the final week eight bait 251
points were used - four for each vegetation type totalling 32 total sample sites, 16 of each veg type 252
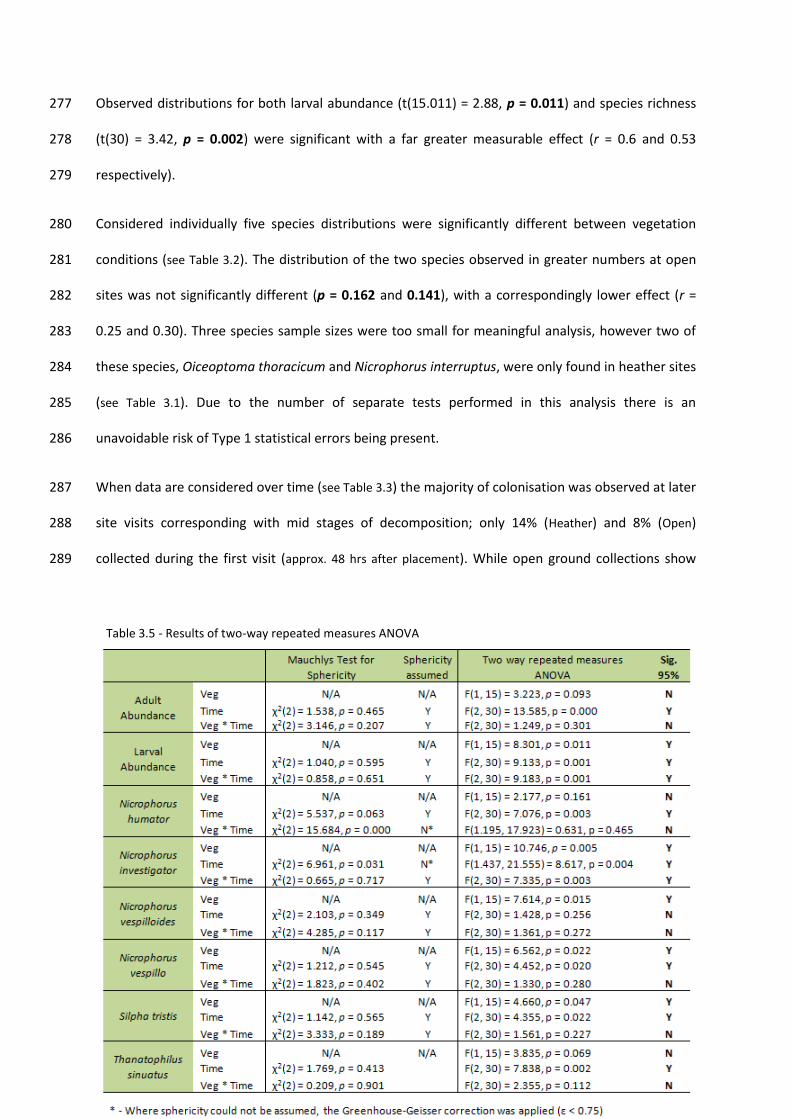
(see Fig 2.1). This reduced third week was a result of problems sourcing carrion bait. This gave a total 253
of 96 collection events across the study period. 254

13
3. Results 255
In total 940 Silphidae were found from ten species and six genera, representing 46% of British 256
Silphidae species (21 total - Booth, 2012). Sixteen species of Silphidae found in Britain are associated 257
with carrion (Dekeirsschieter et al., 2011b) of which nine (56%) were found; in addition one species not 258
specialising in carrion was also found - Silpha atrata. Of this total, 61% (577) were found in the 259
mature heather sites compared to 39% (363) in open sites. Individual species presence ranged from 260
34-100% (Heather) and 0-66% (Open) at species level. Only two species (Nicrophorus humator and 261
Thanatophilus sinuatus) were more abundant in open areas. Abundance of individual species ranged 262
from 2 to over 300 (see Table 3.1). Larval stages of Silphidae species were also counted, although 263
neither identification to species level or aging was attempted. In total 209 larvae were found, with 264
all but 2 (< 1%) found in heather areas. 265
The results of analysis using the independent samples t - test to determine the significance of these 266
data, collectively and individually, are shown in Table 3.2. Levene’s test for equality of variances was 267
employed to determine if equal variance could be assumed for each data set. Kolmogorov-Smirnov’s 268
test for normality showed that only the species abundance data (total number of species found at 269
each site) could be considered to be normally distributed (D(32) = 0.147, p = 0.76), with other data 270

14
sets exhibiting pronounced positive skew and kurtosis. To avoid problems caused by violations of the 271
assumption of normality bootstrapping was used on all other data (using IBM SPSS Statistics v.22) to 272
ensure obtained results could be considered robust (Field, 2009). The size of the effect vegetation 273
type had on all variables was then calculated as a Pearson’s correlation co-efficient (r). 274
Despite a large disparity, the observed difference in abundance between vegetation types was not 275
significant (t(30) = 1.43, p = 0.162) and had only a small effect upon observed differences (r = 0.25). 276
Table 3.2 - Results of Independent samples t - test comparing vegetation types means
Table 3.3 - Total abundance over time
Table 3.4 - Species abundance over time
15
Observed distributions for both larval abundance (t(15.011) = 2.88, p = 0.011) and species richness 277
(t(30) = 3.42, p = 0.002) were significant with a far greater measurable effect (r = 0.6 and 0.53 278
respectively). 279
Considered individually five species distributions were significantly different between vegetation 280
conditions (see Table 3.2). The distribution of the two species observed in greater numbers at open 281
sites was not significantly different (p = 0.162 and 0.141), with a correspondingly lower effect (r = 282
0.25 and 0.30). Three species sample sizes were too small for meaningful analysis, however two of 283
these species, Oiceoptoma thoracicum and Nicrophorus interruptus, were only found in heather sites 284
(see Table 3.1). Due to the number of separate tests performed in this analysis there is an 285
unavoidable risk of Type 1 statistical errors being present. 286
When data are considered over time (see Table 3.3) the majority of colonisation was observed at later 287
site visits corresponding with mid stages of decomposition; only 14% (Heather) and 8% (Open) 288
collected during the first visit (approx. 48 hrs after placement). While open ground collections show 289
Table 3.5 - Results of two-way repeated measures ANOVA

16
consistent colonisation rates at the later collection events (Visit 2 & 3: 46% each) heather areas are 290
less consistent, showing a peak at the second observation (Visit 2: 53%; Visit 3: 33%). By assessing this 291
at species level much greater variability is observed, consistent with different preferences exhibited 292
by different species (Dekeirsschieter et al., 2011b). These data were analysed using two-way repeated 293
measure ANOVA (results in Table 3.5) to test the effect of vegetation conditions and time (related to 294
the stage of decomposition), and interaction between variables. This test assumes that the data 295
exhibits sphericity which is tested using Mauchlys test; requiring a correction if conditions of 296
sphericity are not met (Field, 2009). 297
This test was used to analyse total abundance (adults & larva) and six of the ten observed species. 298
Species with a small sample size (total abundance of < 20 from 96 collection events) were excluded. 299
As a test of variation between vegetation types, this analysis mirrored the earlier t-test results, 300
supporting the robustness of these tests. When comparing how observed abundance changed over 301
time, the results showed that both adult and larval abundance differed significantly (F(2, 30) = 13.58, 302

17
p = 0.000 & F(2, 30) = 9.13, p = 0.001 respectively). Of the six species tested five showed significant 303
variation over time (Fig 3.1). Nicrophorus vespilloides (Visit 1: 32%, Visit 2: 45%, Visit 3: 23%) abundance 304
did not vary significantly (F(2, 30) = 1.43, p = 0.256). 305
Only larval abundance and Nicrophorus investigator showed a significance variation in the 306
interaction between the two variables (F(2, 30) = 9.18, p = 0.001 and F(2, 30) = 7.33, p = 0.003 307
respectively). Fig 3.2 shows the difference in species colonisations between vegetation conditions. 308
Fig 3.2 - Species colonisation rates; comparison between habitats

18
Environmental variables were recorded for each site at each visit to allow comparison. These 309
variables were then compared using independent samples t-tests (see Table 3.6). Of the three 310
variables recorded, only Soil Temperature showed a significant difference between vegetation types 311
(F(1, 94) = -1.917, p = 0.000), with open habitats being significantly warmer than heather areas (21.0 312
OC (± 0.49) open; 19.1 OC (± 0.56) heather sites). It is recognised however, that other measurable 313
environmental and micro-climatic variables will affect invertebrate occupation of these areas, and 314
that these offer only a narrow perspective of differences between these areas. 315
Appendix I contains distribution maps of eight of the observed species; these show abundance of 316
species collected at each sampling site. (Necrodes littoralis and Nicrophorus interruptus distributions were 317
not mapped due to the small samples sizes of 2 and 3 respectively). 318
Tested Variables Levenes test result
Equal Var.
Assumed? Mean diff
Confidence Interval
(Bca 95%) Result
Significant
@ 95%?
Effect size
(r) *
Air Temperature F(1, 94) = 3.85, p = 0.053 Y 0.7604 -0.533, 2.085 t (94) = 1.10, p = 0.274 N 0.11
Soi l Temperature F(1, 94) = 0.11, p = 0.742 Y -1.917 -2.918, -0.930 t (94) = -3.86, p = 0.000 Y 0.37
Humidity F(1, 94) = 1.48, p = 0.226 Y -0.7584 -6.092, 4.187 t (94) = -0.304, p = 0.762 N 0.03
Table 3.6 - Results of independent samples t - test comparing measured environmental variables between vegetation types

19
4. Discussion 319
Silphidae on heathland 320
The specific topic of this study has, so far as the author is aware, no precedent. This prevents critical 321
comparison with other directly comparable studies; however both heathland invertebrates and 322
Silphidae have been studied in sufficient detail to provide some context for the discussion of these 323
results. As such many points raised by this study may go unanswered, instead being a suggestion for 324
further study. 325
The results of the primary research aim - to record species presence and abundance - show that a 326
large proportion of necrophagous British Silphidae (56%) will utilise carrion in heathland areas. 327
Previous records exist for seven of the ten observed species in the 10km grid square in which the 328
study area falls (UK Grid Square TM34). Of particular note is the observation of Nicrophorus 329
interruptus, a ‘Nationally Notable B’ species not previously recorded in this grid square although 330
records exist in adjacent squares, of which three were found. Neither Thanatophilus sinuatus (334 331
found) nor Silpha tristis (91 found) had previously been recorded in this grid square, although 332
records did exist for one or more adjacent grid squares (NBN, Undated). It should be noted that one 333
necrophagous Silphidae species found in Britain, Thanatophilus dispar, is inactive at the time of year 334
the study was conducted (Active: May - July) (Dekeirsschieter et al., 2011b) and therefore may be 335
missed in this study, even if it were present on site. However, it is a rare species (Dekeirsschieter et al., 336
2011b) with no records within 100 miles of the study site (NBN, Undated) (for an assessment of 337
species overall abundance see Table 4.1). 338
Dekeirsschieter et al. (2011b) in reviewing Silphidae in Western Europe suggested habitat 339
preferences for species found in that geographic range (see Table 4.1) (also Martin-Vega & Baz, 2012). 340
The habitats described in this table are not further defined and are therefore difficult to use in great 341
detail. It can reasonably be assumed that heathland habitat, or elements thereof constitute an open 342
habitat; ‘field’ habitats conditions may be met by the areas of acid grassland on site; forest 343

20
conditions may be met by the small areas of semi-mature and mature trees on site, however these 344
are small areas. These assumptions may not fully explain the presence of these species. Some 345
possible explanations are discussed here. 346
Webb (1989) found that the vegetation communities and structure in habitats surrounding 347
heathland areas affects the diversity of invertebrates found there. In Webb’s study the diversity of 348
heathland invertebrates increased (a sign of habitat degradation through loss of specialist 349
communities (Webb & Hopkins, 1984a)) as a result of different habitats bordering heathland areas. 350
This effect was particularly noticeable in areas where heathlands were fragmented and interspersed 351
by modified or intensively managed habitats. The study site is a relatively small area of the 352
heathland in the region: even the full SSSI area of 461 hectares represents only a portion of the total 353
remnant heathland in the ‘Sandlings’ area (SSSI Citation, Sutton & Hollesley Heaths), although much of 354
what does remain is in poor condition. Formerly a much larger tract of heathland, significant 355
portions have been converted to commercial forestry, arable agriculture or commercial 356
development, or allowed to succeed to woodland habitats. Habitats adjacent to the study site 357
include deciduous woodland, coniferous plantation, arable agriculture, a road and unsurfaced tracks 358
and associated verges and other areas of heathland, with different management strategies. With 359
this range of bordering habitats, species not normally found in heathland habitats may use the study 360
site as additional, albeit marginal habitat, especially when there is a specific resource, in this case 361
carrion, available (Webb & Hopkins, 1984a). 362
Lin et al. (2007) found in a study of commercial forestry planted on former heathland in East Anglia 363
that a range of open forest habitats providing refuges suitable for overlapping Carabidae 364
(Coleoptera) communities associated with both closed canopy forest and heathland. If the reverse 365
could also be applied, that heathland provides some suitable habitat for species normally associated 366
with open forest areas, then the presence of those Silphidae deemed to be forest species by 367

21
Dekeirsschieter et al. (2011b) may be using small patches of suitable habitat or marginal habitats on 368
heathland, particularly those areas bordered by or interspersed with forest. 369
Milne & Milne (1976) using mark-release-recapture studies showed that some species of Silphidae 370
can detect and disperse to find carrion resources from as far as 4km. This is achieved through 371
detection of volatile organic compounds (VOC’s), including organosulphates, given off during the 372
decay process (Kalinova et al., 2009; Podskalsa et al., 2009; Forbes, 2014). These are detected by sensitive 373
olfactory glands in the beetle’s antennae (Milne & Milne, 1976). If beetles were drawn from this order 374
of distance to the bait on the study site then species occupying a wide range of habitats may be 375
present. This study has no way of determining distance travelled by individuals to exploit this 376
resource, and therefore it cannot be concluded how inter-habitat dispersal has influenced the 377
observed communities, but it should be borne in mind as a possible influencing factor. 378
There is opportunity for substantial additional study into this area of Silphidae ecology. Further study 379
into habitat occupation and preference could clarify understanding into what habitat features are of 380
greatest import. From the view point of forensic science, which is the motive for a large portion of 381
the study into Silphidae ecology, additional understanding into habitat occupation may be beneficial. 382
Aside from the calculation of PMImin, carrion entomofauna carrion can be beneficial in determining 383
whether a body has been relocated after death (Matuszewski et al., 2013). The presence on a carcase 384
of either larva or adults of species not found in a certain habitat may indicate that a body has been 385
relocated, but an in depth and consistent understanding of species habitat preferences is needed for 386
this to be reliable (Huffman & Wallace, 2012). 387
Variation between vegetation conditions 388
The secondary aim of research was to determine if differences in observed species use of habitat 389
features were significant, specifically areas of mature heather and open areas (see Box 2.1). The 390
results (see Table 3.2) show that significant differences were observed in most of the tested data. All 391
species exhibiting significant variation showed a preference (implied through higher abundance) for 392

22
heather areas over bare ground. Further, of the three species not tested due to insufficient sample 393
size, two were only found in heather areas, suggesting a preference, although untestable with these 394
data. No data shows this more clearly than the collection of Silphidae larva - of 209 collected all but 395
2 were from mature heather sites. 396
Pearson’s correlation co-efficient values for all data which exhibited significance was only less than r 397
= 0.5 (threshold for a large effect - Field, 2009) for one data set, that of Thanatophilus sinuatus (r = 398
0.48) (range: r = 0.52 - 0.60; see Table 3.2). This indicates that the difference in vegetation had a large 399
statistical effect on distribution. 400
Few studies have been published in regard to Silphidae habitat preferences at these small scales, 401
although some are mentioned in studies covering a range of arthropod species, therefore any 402
attempted discussion of causal factors for these habitat preferences by necessity must draw upon 403
non-species specific studies as a back ground. Significant differences in invertebrate communities 404
found in different vegetation structures are widely recorded (e.g. Gardner, 1991; De Bruyn et al., 2001; 405
Blake et al., 2003; Dennis, 2003; Cuesta et al., 2006; Schirmel & Buchholz, 2011; Martin-Vega & Baz, 2012). 406
There are a wide range of factors, unrecorded mostly in the restrictions of this study, which may 407
explain this disparity. Micro-climatic conditions (including but not limited to humidity, temperature 408
and shelter) are widely understood to dictate occupation and use of different habitats by 409
invertebrates even at very small scales (Thomas et al., 1999; Gillingham, 2010; Schirmel et al., 2011), but 410
there are other factors such as vegetation structure and species; presence of prey species; soil types, 411
content and moisture; aspect; elevation and bordering habitats which also play a role (De Jong & 412
Chadwick, 1999; Kirby, 2001). Only one of the three environmental variables measured (Soil 413
Temperature) showed a significant difference between vegetation conditions. 414

23
Silphidae habitat use 415
In light of these results a review of what is understood of Silphidae ecology, and to some extent, the 416
habits of other necrophagous species would be beneficial. Within the group Silphidae two 417
subfamilies, Nicrophorinae and Silphinae, exhibit different strategies for breeding and resource 418
selection (Dekeirsschieter et al., 2011b). In addition both habitat and food resource selection has been 419
shown to differ even within species depending on the level of reproductive maturity of individuals 420
(Lowe & Lauff, 2012). 421
Nicrophorinae: Also known as ‘Burying beetles’, this group are unusual in their breeding strategy. 422
Small vertebrate carcases are buried or covered with soil or organic litter, to a greater or lesser 423
depth dependant on species, to protect the resource from competition. This is done co-operatively 424
with a breeding pair often working together to achieve this. When interred the carcase is 425
manipulated, both physically and chemically, to provide optimum conditions for the development 426
and feeding of larvae (Milne & Milne 1976; Cooter & Barclay, 2006; Dekeirsschieter et al., 2011b). Soil 427
conditions suitable for burial are therefore a key determinant of Nicrophorinae presence (Scott, 428
1998). 429
Due to the described behaviour sexually mature Nicrophorinae show a marked preference for 430
smaller carcases which can be buried more easily (Matuszewski et al., 2008; Lowe & Luaff, 2012); both 431
Dekeirsschieter et al. (2011b) and Wilhelm et al. (2001) suggest that carcases smaller than 100g are 432
most favoured, although burial of carcases up to 300g have been observed. Scott (1998) does record 433
anecdotal evidence of larger carcases, including Rabbits as used in this study, being used by several 434
pairs of Nicrophorinae for breeding in the absence of more suitable resources. Typically however, 435
Nicrophorinae found on larger carcases (> 300g) tend to be sexually immature, or mature but 436
subordinate non-breeding individuals. Their presence is likely for their own food requirements, 437
either by feeding directly on the carrion or through predation of other species, especially Diptera 438
larva. Immature individuals occupying larger carcases are therefore less likely to conform to habitat 439

24
preferences because breeding efficiency is not a consideration in resource selection (Putman, 1983; 440
Dekeirsschieter et al., 2011b; Lowe & Luaff, 2012). 441
Without recording the sex or determining the sexual maturity of individuals in this study it cannot be 442
accurately determined whether these patterns were demonstrated, but it may allow the reasonable 443
assumption that a large number of Nicrophorinae species found in this study were sexually 444
immature. Supporting this assumption; carcases used were all well in excess of 300g (approx. 750 - 445
1500g), and no visual evidence of attempts to bury the carcase was observed. The highest number of 446
Nicrophorinae individuals found on a single carcase in a single visit was 61, from all 5 species: a 447
group of species which aim to limit competition, both intra and inter-specific, when seeking 448
reproductive habitats and resources, are unlikely to be found in these numbers when attempting to 449
breed. 450
Wilhelm et al. (2001) state that smaller Nicrophorinae favour damp soil conditions were burial is 451
more easily achieved and are therefore found more frequently in closed conditions such as 452
woodland; in contrast larger species tend to favour open habitats. In this study, the largest 453
Nicrophorinae species found, Nicrophorus humator, was also the only species not to show a 454
significant preference for heather areas (for species size ranges see Table 4.1). This potentially 455
supports the above statements. Heather dominated areas are a more closed habitat and with 456
greater moisture holding potential due to higher proportions of organic matter than present in bare 457
soil or very short grass sward areas (Gupta & Larson, 1979). However, this should be considered 458
tentatively: soil moisture was not recorded so this assumption cannot be considered conclusive. 459
Also, with the assumption that most individuals observed were sexually immature, preferentially 460
selection of habitat conditions may not have been a causal factor in the significance of these 461
distributional differences. 462
Wilhelm et al. (2001) also indicated that soil temperature affected larval development, with warmer 463
temperatures coinciding with faster larval development rates than in cooler conditions. They 464

25
concluded that this may lead to preferential habitat selection for warm soil conditions. The observed 465
results of this study, in so far as they can be applied, contradict these findings: Nicrophorinae 466
species, with the exception of N. humator were all significantly more abundant in mature heather 467
environments, which showed a significantly lower soil temperature. 468
To more accurately assess the above points, further study could be undertaken, where the sexual 469
maturity of individuals was identified to allow mature and immature individuals to be considered as 470
separate populations. 471
One feature of the soil conditions which is not made mention of in any relevant literature is the 472
presence of root networks and the effect of these on the ability of Nicrophorinae to effectively bury 473
carrion. The areas of mature heather exhibited often dense root networks. These were not 474
measured or recorded during the study, but did present a challenge in placing pitfall traps. Data 475
from this study cannot determine any conclusion to this informal hypothesis, but it again identifies 476
unanswered questions in this field of study. 477
Silphinae: In contrast to Nicrophorinae species, Silphinae do not bury carrion. This dictates a 478
significantly different, almost opposite, association with the resource. Larger carcases (> 300g) are 479
favoured by sexually mature individuals (Wilhelm et al., 2001; Martin-Vega & Baz, 2010). Silphinae eggs 480
are laid in the soil near to carcases allowing larva when hatched to colonise and utilise the food 481
resource, with no parental care (Dekeirsschieter et al., 2011b). Immature species meanwhile will utilise 482
a wider range of carcase sizes, including much smaller carcases than would be used as a site for 483
oviposition (Dekeirsschieter et al., 2011b).This implies that sexually mature individuals of the two 484
groups will not be in direct competition with each other for breeding resources, but that immature 485
individuals of the different subfamilies may compete for resources. Niche separation may be 486
achieved through a preference for different stages of decomposition (Dekeirsschieter et al., 2011b). 487
Without the need to bury carrion soil conditions are largely removed as significant feature, although 488

26
soil temperature may again affect early development as eggs are deposited into the soil - this is not 489
discussed in published literature. 490
Based on illustrations and descriptions in Cooter & Barclay (2006) and Dekeirsschieter et al. (2011b) 491
it is believed that all larvae found were Silphinae species although formal identification was not 492
attempted. Nicrophorus species are morphologically distinct and their typical breeding habit would 493
make it unusual to find these larvae feeding out in the open. 494
One interesting note is that in this study larvae were observed on carcases as early as 2 days after 495
carcase placement. Dekeirsschieter et al. (2011b) state that eggs hatch in 4 - 5 days. Even 496
discounting the time period needed for adults to colonise a carcase site and oviposit, this does not 497
allow sufficient time for egg development and hatching (far higher larval abundance was observed at 498
later collections: Visit 1: 38; Visit 2: 105; Visit 3: 66). This shows that larvae, as well as adults, must be 499
dispersing to the resource from the surrounding area: without flight it is certain that the dispersal 500
capability of these larvae is less than adults of the species, but no studies can be found suggesting 501
what dispersal distances are possible for larva. Instar number was not determined, nor were larvae 502
measured, but the range of sizes (approx. from <5mm to >25mm) suggested a range of instars from 503
across the developmental spectrum were present throughout the study. 504
This identifies another area of research of potential interest to forensic investigation: Silphinae are 505
of greater use than Nicrophorinae in forensic investigations because of their preference for larger 506
carcases (Dekeirsschieter et al., 2011b). However, if their larvae are used in this context then 507
understanding the dispersal abilities of these larvae could be a significant factor in accurately 508
determining PMImin. Lewis & Benbow (2011) have shown that a lack of understanding of dispersal 509
habits and capabilities in Diptera larva may have led to misinterpretation of forensic evidence 510
previously. It is believed that this could be an additional area where lack of published research may 511
lead to misinterpretation. 512

27
Matuszewski et al. (2013) conducted a study of necrophagous invertebrates across a gradient of 513
habitats from open areas to forest. Three Silphinae species also found in this study were observed: 514
Necrodes littoralis and Thanatophilus sinuatus were found across the range of habitats as in this 515
study. The final species, Oiceoptoma thoracicum, was considered to be a forest specialist, and 516
avoided breeding in open areas. In the current study O. thoracicum was observed only in heather 517
habitats, although in very low abundance, supporting to a degree the study in that they favoured 518
more closed vegetation conditions. However, at a landscape scale, heathland areas generally would 519
be classified as open. The above study used the findings to state that O. thoracicum found on a 520
carcase in open areas would be a sign of carcase relocation, but the findings of this study contradict 521
that at a wider habitat scale. This demonstrates the need for clarity in defining habitats in studies 522
such as these. 523
Entomofaunal Succession 524
The final research aim was to observe the colonisation patterns of the observed species or 525
entomofaunal succession. Fig 3.1 shows that differences were observed between species 526
colonisation rates; Fig 3.2 allows the colonisation rates to be compared between habitats showing 527
some differences although only Nicrophorus investigator demonstrated a significant interaction 528
between habitat and time; Table 3.5 shows which data exhibited significant differences over time. 529
Unfortunately the meaningful analysis of this data is limited due to the variance displayed in 530
decomposition rate. Entomofaunal succession is dictated not by time since death but stage of 531
decomposition which is dictated by biotic, abiotic and seasonal variables (Wilhelm et al., 2001; 532
Matuszewski et al., 2010). The scale of this variation was poorly understood to the author prior to data 533
collection and so no provision was made for recording the stage of decomposition. This has 534
compromised the usefulness of these data. 535
The two sub-families again exhibit behavioural differences and contrasting preferences between 536
stages of decomposition in which they colonise a carcase. Adults of both sub-families will occupy a 537

28
carcase to feed themselves; either directly from the carrion or by predating the other invertebrates 538
found there or to use carrion as a resource for reproduction (Chapman & Sankey, 1955; Putman, 1983). 539
Immature individuals seeking to feed themselves are less restricted to prime conditions so will 540
occupy carrion at a wider range of stages of decomposition than sexually mature individuals whose 541
focus is reproduction (Matuszewski et al., 2008). 542
Sexually mature Nicrophorinae species, as a result of their reproductive habit will colonise small 543
carcases early in the decomposition stage to prevent competition when the carcase is interred 544
(Dekeirsschieter et al., 2011b). All Nicrophorinae species (except N. interruptus with a total sample size 545
of 3) were found at the first visit just two days after placement and then throughout the study. The 546
carcases used in this study were larger than is preferentially chosen and so the colonisation habits of 547
these species cannot be accurately tested fully in this setting, although the results do seem to 548
support this behaviour. 549
In contrast, Silphinae species are more associated with mid stages of decomposition: bloated and 550
decay (Putman, 1983; Dekeirsschieter et al., 2011b). In this study Silphinae were found across the time 551
spectrum, although, because this cannot be reliably linked with a stage of decomposition, it cannot 552
be definitively stated whether or not this matches a specific stage of decomposition. 553
Different decomposition rates are the result of a variety of factors including the colonisation by 554
different invertebrates, especially Diptera (Putman, 1983; Matuszewski et al., 2008). The presence of 555
Diptera larvae and in some cases the larvae of other necrophagous species including Silphinae 556
species has a significant impact on the rate at which decomposition progresses (De Bruyn et al., 2001; 557
Matuszewski et al., 2010). Matuszewski et al. (2010) showed that decomposition progresses fastest in 558
more humid habitats, largely as a result of the increased Diptera colonisation. Therefore the 559
colonisation of carrion by Silphidae is also affected by the wider necrophagous invertebrate 560
community. 561

29
A diverse range of factors affect the behaviour of Silphidae in any habitat they occupy. This study has 562
attempted to test and expand current knowledge on a few of these factors in a habitat in which they 563
as a group are poorly studied. The gap in ecological understanding that was identified was far too 564
large for a single study to attempt to address; hence the aim of this study was largely to use a 565
research opportunity to identify additional research which could be undertaken to fill this gap. Some 566
further opportunities for study have been identified in the course of comparing the results of this 567
study with previously published work. 568

30
Acknowledgements 569
I would like to thank and acknowledge the assistance of: Dr P. Mitchell at all stages of project 570
planning, execution and write up; B. Calversbert on behalf of Suffolk Wildlife Trust, for allowing me 571
to undertake this research project at Sutton Heath Nature Reserve, Suffolk; P. Shires from Stoke-on-572
Trent City Council, for allowing me to undertake pilot studies at Park Hall Country Park; finally my 573
family, especially my wife, for numerous acts of support and assistance throughout this project and 574
all other study I have undertaken. 575

31
References 576
Aerts, R., Huiszoon, A., Van Oostrum, J. H. A., Van der Vijver, C. A. D. M., & Wilhelms, J. H. (1995) The 577
potential for heathland restoration on formerly arable land at a site in Drenthe, The Netherlands, Journal of 578
Applied Ecology, 32: 827-835. 579
Anon. (2013) Promoting Habitat Mosaics for Invertebrates: Lowland Heath, Buglife, Peterborough. 580
Blake, S., McCracken, D. I., Eyre, M. D., Garside A. & Foster, G. N. (2003) The relationship between the 581
classification of Scottish ground beetle assemblages (Coleoptera, Carabidae) and the National Vegetation 582
Classification of British plant communities, Ecography, 26 (5): 602-616. 583
Booth, A. G. (2012) Silphidae, in: Duff, A.G. (Ed.) Checklist of Beetles of the British Isles (2nd
Ed), Pemberley 584
Books, Iver. 585
Buglife (Undated) Lowland Heath [Online], Available from: www.buglife.org.uk/advice-and-586
publications/advice-on-managing-bap-habitats/lowlandheathland (Accessed on: 29/11/2013). 587
Byrd, J. H. & Castner, J. L. (Eds.) (2010) Forensic Entomology: The Utility of Arthropods in Legal Investigations 588
(2nd
Ed.), CRC Press, Boca Raton, FL, USA. 589
Castro, C. P., Garcia, M. D., da Silva, P. M., Silva I. F. & Serrano, A. (2013) Coleoptera of forensic interest: A 590
study of seasonal community composition and succession in Lisbon, Portugal, Forensic Science International, 591
232: 73–83. 592
Chapman, R. F. & Sankey, J. H. P (1955) The Larger Invertebrate Fauna of three Rabbit Carcases, Journal of 593
Animal Ecology, 24: 395-402. 594
Chick, A. (2008) Some novel suggestions for the collection and study of Diptera from carrion. Bulletin of the 595
Dipterists Forum, 65: 24-26. 596
Cooter, J. & Barclay, M. V. L. (Eds.) (2006) A Coleopterists Handbook (4th Edition), The Amateur Entomologists 597
Society, Orpington. 598
Cuesta, D., Taboada, A., Calvo, L. & Salgado, J. M. (2006) A preliminary investigation of ground beetles 599
(Coleoptera: Carabidae) assemblages and vegetation community structure in Calluna vulgaris heathlands in 600
NW Spain, Entomologica Fennica, 17: 241-252. 601
De Bruyn, L., Thys, S., Scheirs, J. & Verhagen, R. (2001) Effects of vegetation and soil on species diversity of 602
soil dwelling Diptera in a heathland ecosystem, Journal of Insect Conservation, 5: 87-97. 603
De Jong, G. D. & Chadwick, J. W. (1999) Decomposition and Arthropod Succession on Exposed Rabbit Carrion 604
During Summer at High Altitudes in Colorado, USA, Journal of Medical Entomology, 36 (6): 833-845. 605

32
Dekeirsschieter J., Verheggen F. J., Haubruge E. & Brostaux Y. (2011a) Carrion beetles visiting pig carcasses 606
during early spring in urban, forest and agricultural biotypes of Western Europe, Journal of Insect Science, 11: 607
73. 608
Dekeirsschieter, J., Verheggen, F. J., Lognay, G. & Haubruge, E. (2011b) Large carrion beetles (Coleoptera, 609
Silphidae) in Western Europe: a Review, Biotechnology, Agronomy, Environment and Society, 15 (3): 435 - 447. 610
Dennis, P. (2003) Sensitivity of upland arthropod diversity to livestock grazing, vegetation structure and 611
landform, Food, Agriculture & Environment, 1 (2): 301-307. 612
DeVault, T. L., Brisbin, I. L. & Rhodes, O. E. (2004) Factors influencing the acquisition of rodent carrion by 613
vertebrate scavengers and decomposers, Canadian Journal of Zoology, 82: 502 - 509. 614
DeVault, T. L., Olson, Z. H., Beasley, J. C. & Rhodes, O. E. (2011) Mesopredators dominate competition for 615
carrion in an agricultural landscape, Basic and Applied Ecology, 12: 268–274. 616
Field, A. (2009) Discovering Statistics using SPSS (3rd Ed.), Sage Publications Ltd., London. 617
Forbes, S.L. & Perrault, K. A. (2014) Decomposition Odour Profiling in the Air and Soil Surrounding Vertebrate 618
Carrion. PLoS ONE 9(4): e95107. 619
Gardner, S. M. (1991) Ground Beetle (Coleoptera: Carabidae) communities on upland heath and their 620
association with heathland flora, Journal of Biogeography, 18: 281-289. 621
Gillingham, P. (2010) The relative importance of microclimate and land use to biodiversity, PhD Thesis, 622
University of York, York 623
Grassberger, M. & Frank C. (2004) Initial Study of Arthropod Succession on Pig Carrion in a Central European 624
Urban Habitat, Journal of Medical Entomology, 41 (3): 511-523. 625
Gupta, S. C. & Larsen, W. E. (1979) Estimating Soil Water Retention Characteristics From Particle Size 626
Distribution, Organic Matter Percent, and Bulk Density, Water Resources Research, 15 (6): 1633-1635. 627
Hackston, M. (2009) Key to the British Species of genus Nicrophorus [Online], Available at: www.record-628
lrc.co.uk/Downloads/KeytotheBritishspeciesofgenusNicrophorus[07052011].pdf, (Accessed: June 2014). 629
Hackston, M. (2014) Key to genus of the British Silphidae [Online], Available at: 630
https://sites.google.com/site/mikesinsectkeys/Home/keys-to-coleoptera/silphidae, (Accessed: June 2014). 631
Hawley, G., Anderson, P., Gash, M., Smith, P., Higham, N., Alonso, I., Ede, J. & Holloway, J. (2008) Impact of 632
heathland restoration and re-creation techniques on soil characteristics and the historical environment: Natural 633
England Research report RIN010, Natural England, Peterborough. 634

33
Huffman, J. E. & Wallace J. R. (Eds.) (2012) Wildlife Forensics: Methods and Applications, Wiley-Blackwell, 635
Chichester. 636
Hurlbert, S. H. (1984) Psuedoreplication and the design of ecological field experiments, Ecological 637
Monographs, 54 (2): 187-211. 638
JNCC (Undated) UK Lowland Heathland Habitats [Online], Available from: http://jncc.defra.gov.uk/page-1432 639
(Accessed on: 3/12/2013). 640
Kalinová, B., Podskalská, H., Růžička, J. & Hoskovec, M. (2009) Irresistible bouquet of death—how are burying 641
beetles (Coleoptera: Silphidae: Nicrophorus) attracted by carcasses, Naturwissenschaften, 96 (8): 889-899. 642
Kirby, P. (2001) Habitat Management for Invertebrates: A Practical Handbook. RSPB, Sandy. 643
Kirby, P. (2004) Habitat Fragmentation. Species at Risk. Invertebrate Group Identification: 89. English Nature, 644
Peterborough. 645
Kulshretha, P. & Satpathy, D. K. (2001) Use of Beetles in forensic entomology, Forensic Science International, 646
120 (1-2): 15-17. 647
Lewis, A. J. & Benbow, M. E. (2011) When Entomological Evidence Crawls Away: Phormia regina en masse 648
Larval Dispersal, Journal of Medical Entomology, 48 (6): 1112-1119. 649
Lin, Y.-C., James, R. & Dolman, P. M. (2007) Conservation of heathland ground beetles (Coleoptera: 650
Carabidae): the value of lowland coniferous plantations, Biodiversity and Conservation, 16: 1337-1358. 651
Lowe, A. J. & Lauff, R. E. (2012) Arboreal Burials in Nicrophorus spp. (Coleoptera: Silphidae), Psyche, 2012: 1-6 652
(Article ID 578650). 653
Martin-Vega, D. & Baz, A. (2012) Spatiotemporal Distribution of Necrophagous Beetles (Coleoptera: 654
Dermestidae, Silphidae) Assemblages in Natural Habitats of Central Spain, Annuls of the Entomological Society 655
of America, 105 (1): 44-53. 656
Matuszewski, S., Szafałowicz, M. & Jarmusz, M. (2013) Insects colonising carcasses in open and forest habitats 657
of Central Europe: Search for indicators of corpse relocation, Forensic Science International, 231: 234–239. 658
Matuszewski, S., Bajerlein, D., Konwerski, S. & Szpila, K. (2008) An initial study of insect succession and 659
carrion decomposition in various forest habitats of Central Europe, Forensic Science International, 180 (2-3): 660
61-69. 661
Matuszewski, S., Bajerlein, D., Konwerski, S. & Szpila, K. (2010) Insect succession and carrion decomposition 662
in selected forests of Central Europe. Part 1: Pattern and rate of decomposition, Forensic Science International, 663
194: 85–93. 664

34
Midgley, J. M., Richards, C. S. & Villet, M. H. (2010) The Utility of Coleoptera in Forensic Investigations, pp. 57-665
68 in: Amendt, J., Goff, M. L., Campobasso, C. P. & Grassberger, M. (Ed’s) (2010) Current Concepts in Forensic 666
Entomology, Springer. 667
Milne, L. J. & Milne, M. (1976) The Social Behaviour of Burying Beetles, Scientific American, 235 (2): 84 - 89. 668
Moreno-Opo, R. & Margalida, A. (2013) Carcasses provide resources not exclusively to scavengers: patterns of 669
carrion exploitation by passerine birds. Ecosphere, 4 (8): 105. 670
NBN (National Biodiversity Network) (Undated) NBN Gateway Interactive Species Mapping [Online], available 671
at: https://data.nbn.org.uk/imt/, (Accessed on: June 2014). 672
Podskalská, H., Růžička, J., Hoskovec, M. and Šálek, M. (2009) Use of infochemicals to attract carrion beetles 673
into pitfall traps. Entomologia Experimentalis et Applicata, 132: 59–64. 674
Price, E. (2003) Lowland Grassland and Heathland Habitats, Routledge, London 675
Putman, R. J. (1983) Carrion and Dung: the decomposition of animal wastes, Studies in Biology No 156, Edward 676
Arnold, London. 677
Rackham, O. (2003) The Illustrated History of the Countryside, Weidenfield & Nicolson, London. 678
Ray, R.–R., Seibold, H. & Heurich, M. (2014) Invertebrates outcompete vertebrate facultative scavengers in 679
simulated lynx kills in the Bavarian Forest National Park, Germany. Animal Biodiversity and Conservation, 37 680
(1): 77–88. 681
Schirmel, J. & Buchholz, S. (2011) Response of carabid beetles (Coleoptera: Carabidae) and spiders (Araneae) 682
to coastal heathland succession, Biodiversity and Conservation, 20: 1469-1482. 683
Schirmel, J., Mantilla-Contreras, J., Blindow, I. & Fartmann, T. (2011) Impacts of succession and grass 684
encroachment on heathland Orthoptera, Journal of Insect Conservation, 15: 633-642. 685
Schirmel, J. (2010) Short-term effects of modern heathland management measures on carabid beetles 686
(Coleoptera: Carabidae), Applied Ecology and Environmental Research, 8 (3): 165-175. 687
Scott, M. P. (1998) The Ecology and Behaviour of Burying Beetles, Annual Review of Entomology, 43: 595-618. 688
Sharanowski, B. J., Walker, E. G. & Anderson, G. S. (2008) Insect succession and decomposition patterns on 689
shaded and sunlit carrion in Saskatchewan in three different seasons, Forensic Science International, 179: 219-690
240. 691
Sutherland, W. & Hill, D. (Eds.) (1995) Managing Habitats for Conservation, Cambridge University Press, 692
Cambridge. 693

35
Symes, N. & Day, J. (2003) A practical guide to the restoration and management of Lowland Heathland, RSPB, 694
Sandy. 695
Tantawi, T. I., El-Kady, E. M.; Greenberg, B. & El-Ghaffar, H. A. (1996) Arthropod Succession on Exposed 696
Rabbit Carrion in Alexandria, Egypt, Journal of Medical Entomology, 33 (4): 566-580. 697
Thomas, J. A., Rose, R. J., Clarke, R. T., Thomas, C. D. & Webb, N. R. (1999) Intraspecific variation in habitat 698
availability among ectothermic animals near their climatic limits and their centres of range. Functional Ecology, 699
13: 55–64. 700
Usher, M. B. (1992) Management and diversity of arthropods in Calluna heathland, Biodiversity and 701
Conservation, 1: 63-79. 702
Van den Berg, L. J. L., Vergeer, P. & Roelofs, J. G. M. (2003) Heathland restoration in the Netherlands: Effects 703
of turf cutting depth on germination of Arnica montana, Applied Vegetation Science, 6: 117-124 704
Watson, E. J. & Carlton, C. E. (2003) Spring succession of necrophilous insects on wildlife carcasses in 705
Louisiana, Journal of Medical Entomology, 40 (3): 338 - 347. 706
Watson, E. J. & Carlton, C. E. (2005) Insect succession and decomposition of wildlife carcasses during fall and 707
winter in Louisiana, Journal of Medical Entomology, 42 (2): 193 - 203. 708
Webb, J. R., Drewitt, A. L. & Measures G. H. (2010) Managing for species: integrating the needs of England’s 709
priority species into habitat management. NERR024. Natural England, Sheffield. 710
Webb, N. R. & Hopkins P. J. (1984a) Invertebrate diversity on fragmented Calluna heathland, Journal of 711
Applied Ecology, 21: 921-933. 712
Webb, N. R. & Hopkins, P. J. (1984b) The composition of the Beetle and Spider faunas on fragmented 713
heathlands, Journal of Applied Ecology, 21: 935-946. 714
Webb, N. R. (1989) Studies on the invertebrate fauna of fragmented heathland in Dorset, UK, and the 715
implications for conservation, Biological Conservation, 47 (2): 145-156. 716
Webb, N. R., Clarke, R. T. & Nicholas, J. T. (1984) Invertebrate diversity on fragmented Calluna-heathland: 717
effects of surrounding vegetation, Journal of Biogeography, 11: 41-46. 718
Wilhelm, S. I., Larson, D. J. & Storey, A. E. (2001) Habitat Preference Of Two Burying Beetles (Coleoptera: 719
Silphidae: Nicrophorus) Living Among Seabirds, North-eastern Naturalist, 8 (4): 435-442. 720
Wilson, E. E. & Wolkovich, E. M. (2011) Scavenging: how carnivores and carrion structure communities, Trends 721
in Ecology and Evolution, 26 (3): 129-135. 722
Wright, R. (2009a) Beginners guide Silphidae 1: Nicrophorus, Beetle News, 1 (1): 6. 723

36
Wright, R. (2009b) Beginners guide: Silphidae 2, Beetle News, 1 (3): 5-9. 724
725

37
Appendix I - Species Distribution and Abundance Maps 726
Appendix I: Species Distribution & Abundance Maps

38
727




















