
Stauroneis lacusvulcani sp. nov. (Bacillariophyceae), a new diatom from volcanic lakes in...
10
Accepted by David M. Williams: 31 Oct. 2013; published: 28 Nov. 2013 47 PHYTOTAXA ISSN 1179-3155 (print edition) ISSN 1179-3163 (online edition) Copyright © 2013 Magnolia Press Phytotaxa 148 (1): 47–56 (2013) www.mapress.com/ phytotaxa/ Article http://dx.doi.org/10.11646/phytotaxa.148.1.3 Stauroneis lacusvulcani sp. nov. (Bacillariophyceae), a new diatom from volcanic lakes in northeastern China PATRICK RIOUAL 1* , QIANG GAO 1 , YUMEI PENG 2 & GUOQIANG CHU 1 1 Key Laboratory of Cenozoic Geology and Environment, Institute of Geology and Geophysics, Chinese Academy of Sciences, #19 Beitucheng Xilu, P.O. box 9825, Beijing 100029, China * E-mail: [email protected] (Corresponding author). 2 State Key Laboratory of Loess and Quaternary Geology, Institute of Earth Environment, Chinese Academy of Sciences, Xi’an 710075, China *Corresponding author: E-mail: [email protected] Abstract Stauroneis lacusvulcani sp. nov. is described from two small volcanic lakes in northeastern China. The morphology of S. lacusvulcani is illustrated with light and scanning electron micrographs and discussed in comparison with several species of the Stauroneis gracilior group. S. lacusvulcani can be distinguished by the size of the valves and its long-protracted and strongly capitate apices. Diatom analysis of the sedimentary record from Lake Xiaolongwan (Jilin Province) showed that this new species was most abundant during the early Holocene (~8950 to 10,640 yrs BP). Key words: China, morphometric analysis, new species, palaeolimnology, Stauroneis Introduction In recent years, our understanding of the taxonomy and species diversity in the genus Stauroneis Ehrenberg (1843: 45) has considerably improved thanks to the work of Lange-Bertalot et al. (2003), Van de Vijver et al. (2004) and Bahls (2010). In their overview for China, Li & Qi (2010) listed 43 species and varieties of Stauroneis. In the past few years, sediment cores were retrieved from numerous volcanic lakes in northeastern China. During this research, a new species of Stauroneis was discovered and is described herein using light (LM) and scanning electron microscopy (SEM). Material and methods Samples collection and preparation In the context of an ongoing project that aims at investigating the past and present diversity and distribution of diatoms in Northeastern China (Fig. 1), surface-sediment and sediment core samples have been investigated from a suite of lakes. The samples were retrieved from the deepest point of the lakes using either an Uwitec® gravity corer or a modified piston corer (Chu et al. 2009). In parallel with the collection of sediment samples, surface water samples (~0.3 m depth) were also collected and analysed for a wide range of chemical and physical variables. The methods used to measure these various environmental variables are given in Rioual et al. (2013). The species described herein was found in 2 localities: first in the surface-sediment of Lake Xiaolongwan (October 2005) and then in the surface sediment of Sifangshan Tianchi (April 2012). We also investigated the distribution of this new species in a sediment core retrieved in 2006 from Lake Xialongwan. The age model TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Transcript of Stauroneis lacusvulcani sp. nov. (Bacillariophyceae), a new diatom from volcanic lakes in...

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
PHYTOTAXA
ISSN 1179-3155 (print edition)
ISSN 1179-3163 (online edition)Copyright © 2013 Magnolia Press
Phytotaxa 148 (1): 47–56 (2013)
www.mapress.com/phytotaxa/Article
http://dx.doi.org/10.11646/phytotaxa.148.1.3
Stauroneis lacusvulcani sp. nov. (Bacillariophyceae), a new diatom from volcanic
lakes in northeastern China
PATRICK RIOUAL1*, QIANG GAO1, YUMEI PENG2 & GUOQIANG CHU1
1Key Laboratory of Cenozoic Geology and Environment, Institute of Geology and Geophysics, Chinese Academy of Sciences, #19
Beitucheng Xilu, P.O. box 9825, Beijing 100029, China * E-mail: [email protected] (Corresponding author).2State Key Laboratory of Loess and Quaternary Geology, Institute of Earth Environment, Chinese Academy of Sciences, Xi’an 710075,
China
*Corresponding author: E-mail: [email protected]
Abstract
Stauroneis lacusvulcani sp. nov. is described from two small volcanic lakes in northeastern China. The morphology of S.
lacusvulcani is illustrated with light and scanning electron micrographs and discussed in comparison with several species
of the Stauroneis gracilior group. S. lacusvulcani can be distinguished by the size of the valves and its long-protracted
and strongly capitate apices. Diatom analysis of the sedimentary record from Lake Xiaolongwan (Jilin Province) showed
that this new species was most abundant during the early Holocene (~8950 to 10,640 yrs BP).
Key words: China, morphometric analysis, new species, palaeolimnology, Stauroneis
Introduction
In recent years, our understanding of the taxonomy and species diversity in the genus Stauroneis Ehrenberg
(1843: 45) has considerably improved thanks to the work of Lange-Bertalot et al. (2003), Van de Vijver et al.
(2004) and Bahls (2010). In their overview for China, Li & Qi (2010) listed 43 species and varieties of
Stauroneis. In the past few years, sediment cores were retrieved from numerous volcanic lakes in northeastern
China. During this research, a new species of Stauroneis was discovered and is described herein using light
(LM) and scanning electron microscopy (SEM).
Material and methods
Samples collection and preparation
In the context of an ongoing project that aims at investigating the past and present diversity and
distribution of diatoms in Northeastern China (Fig. 1), surface-sediment and sediment core samples have been
investigated from a suite of lakes. The samples were retrieved from the deepest point of the lakes using either
an Uwitec® gravity corer or a modified piston corer (Chu et al. 2009). In parallel with the collection of
sediment samples, surface water samples (~0.3 m depth) were also collected and analysed for a wide range of
chemical and physical variables. The methods used to measure these various environmental variables are
given in Rioual et al. (2013).
The species described herein was found in 2 localities: first in the surface-sediment of Lake Xiaolongwan
(October 2005) and then in the surface sediment of Sifangshan Tianchi (April 2012). We also investigated the
distribution of this new species in a sediment core retrieved in 2006 from Lake Xialongwan. The age model
Accepted by David M. Williams: 31 Oct. 2013; published: 28 Nov. 2013 47

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
established for this core combines the count of annual laminations (= varves) and 14C dating (Chu et al. 2009).
For diatom analysis, at least 500 diatom valves were enumerated to establish abundances values expressed as
relative percentages. Diatom concentration was estimated using the microsphere method of Battarbee &
Kneen (1982) and expressed as number of valves per gram dry matter (valves g dm-1). Collection data and
details on the localities of the samples investigated are presented in Table 1.
TABLE 1: Physical and chemical parameters recorded at the localities of the samples studied.
The sediment samples were prepared using the water-bath method (Renberg 1990). Sub-samples of the
cleaned, homogenized suspension were diluted with distilled water and left to settle onto glass coverslips and
dry overnight in ambient conditions. Coverslips were mounted on glass slides with Naphrax® (a commonly
used mountant with a refraction index of 1.73). To establish abundance values expressed as relative
percentages at least 500 diatom valves were enumerated.
Counting was performed with a Leica DM LB2 light microscope using bright-field and phase contrast oil
immersion optics at 1000× magnification.
Morphometric analysis
For LM analysis, measurements of valve length, width and striae density were made on digitized images
calibrated against a slide micrometer using the public domain ImageJ software (http://imagej.nih.gov/ij/). In
total, 85 valves were investigated under the LM.
For SEM analyses, aliquots of cleaned samples were air-dried onto glass coverslips of 12 mm diameter.
These were mounted onto aluminum stubs using Ted Pella® double-coated carbon conductive adhesive tape
and coated with gold using a Polaron SC7640 sputter coater for viewing on a LEO 1530 VP at the State Key
Laboratory of Paleobiology and Stratigraphy (Chinese Academy of Sciences, Nanjing). For areolae density,
the length of 5 areolae was measured on 5 striae located immediately near the stauros. These values were
converted to a number of areolae per 10 μm (Cortese & Gersonde 2007) and averaged. Only 7 valves could be
investigated under the SEM.
Lake
Parameter Sifangshan Xiaolongwan
Sample collector G.Chu P.Rioual
No water samples and dates 1; Apr–2012 8; from Jul–05 to Oct–12
Elevation (masl) 869 657
Latitude (N) 49º 22'33'' 42º 17'59''
Longitude (E) 123º 27'51' 126º 21'35''
Lake area (ha) 0.1 7.7
Max depth (m) 0.5 15
pH 5.99 6.54–6.93
Elec. Conductivity (µS/cm) 146 52–76
Alkalinity (µeq/L) 1177 364–498
TP (µg/L) 330 4–26
TN (µg/L) 5186 784–1818
DOC (mg/L) 35.7 2.4–12.3
Cl (mg/L) 5.1 1.0–2.5
NO3 (mg/L) 1.6 0.0–4.2
SO4 (mg/L) 1.9 3.1–5.3
Ca (mg/L) 22.1 5.7–9.4
K (mg/L) 5.6 1.3–2.9
Mg (mg/L) 5.4 1.5–2.2
Na (mg/L) 2.4 0.4–1.4
Dissolved Si (mg/L) 38.3 0.1–1.3
% Stauroneis lacusvulcani in surface sediment 0.7 0.2
RIOUAL ET AL.48 • Phytotaxa 148 (1) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURE 1: Geographical location of the study sites. Lake Xiaolongwan is marked by an X, Sifangshan Tianchi by a S. Shaded areas
represent mountainous regions with an elevation >1000 m.
Statistical analyses
To check if the populations of Stauroneis found at the two localities show significant morphometric
differences, single characters analyses (boxplots) and bivariate analyses (scatter plots) were performed. In
particular, the relationship between valve length and width is traditionally used to distinguish diatom taxa
(Mann et al. 2004). For single characters analyses, non-overlapping notches on boxplots were used to infer
statistically significant dissimilarity at the 95% confidence interval (Chambers et al. 1983, Paull et al. 2008).
Boxplots and scatterplots were drawn using R (http://www.r-project.org/).
Abbreviations
Light microscopy (= LM); scanning electron microscopy (= SEM). Author abbreviations follow the
International Plant Name Index (IPNI; http://www.ipni.org/); herbarium acronyms follow Index
Herbariorum (http://sciweb.nybg.org/science2/IndexHerbariorum.asp) with IGGDC = Herbarium of
Bacilliarophyceae, Institute of Geology and Geophysics, Chinese Academy of Sciences, Beijing, People's
Republic of China
New species description
Stauroneis lacusvulcani Rioual sp. nov. (Figs 2–21)
Type:—CHINA: Inner Mongolia, Great Xing’An Mountains, Sifangshan Tianchi, 869 m, 49º 22'33''N , 123º 27'51''E,
April-2008, surface sediment collected by Dr Guoqiang Chu, IGG-CAS, Beijing. (holotype slide IGGDC-DB-SIFA-
1204!, holotype specimen illustrated in Fig 7, located using England Finder K45/1); BM 101 674 (isotype slide).
Phytotaxa 148 (1) © 2013 Magnolia Press • 49STAURONEIS LACUSVULCANI SP. NOV.

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 2–15. Light micro-photographs of Stauroneis lacusvulcani. Scale bar = 10 µm. Figs 3,4,7,8. Specimen from Sifangshan
Tianchi, surface-sediment. Figs 2, 5, 6. Specimens from Xialongwan surface-sediment. Figs 9–15. Specimens from Xiaolongwan core
sample.
RIOUAL ET AL.50 • Phytotaxa 148 (1) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
LM (Figs 2–15): Valves lanceolate with long protracted capitate apices (Figs 2–15); length 62–83 μm, width
9.2–12.0 μ m. Raphe moderately lateral, becoming filiform at distal ends, with strongly unilaterally deflected
(hooked) distal ends. Axial area narrow, linear. Shape of central area varies from narrow rectangular (Figs 2–
3, 5–6, 10–13) to asymmetrical stauros expanding towards valve margins (Figs 1, 4, 7–9, 14–15). Shortened
striae may be present at valve margins (Figs 2, 5–8, 9–12) or not (Figs 3–4, 13–15). Striae parallel along
border of stauros becoming radiate to more steeply radiate near valve apices, 21–28 in 10 µm.
SEM (Figs 16–21): Areolae round to transapically oblong, irregularly spaced, 18–28 in 10 µm. Weakly
developed hyaline area present, thus striae next to stauros appear discontinuous (Figs 17, 20). Raphe bordered
by wide sternum on each side. Proximal raphe ends straight and slightly expanded (Fig. 17), apical external
raphe endings hooked with hooks opening toward secondary side (Fig. 18). Internally, raphe-sternum bears
longitudinal ribs and well-developed helictoglossae (Fig. 21).
FIGURES 16–21. SEM of Stauroneis lacusvulcani from Sifangshan. 16. Exterior view of whole valve. 17. Close-up of valve centre.
18. Close-up of distal end. 19. Interior view of whole valve. 20. Interior view of centre showing ridge bearing raphe fissure. 21.
Interior view of distal end showing helictoglossa.
Phytotaxa 148 (1) © 2013 Magnolia Press • 51STAURONEIS LACUSVULCANI SP. NOV.

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Etymology:—Based on its occurrence in volcanic lakes.
Distribution and ecology:—Stauroneis lacusvulcani was found in the surface sediment of two volcanic
lakes of NE China, Sifangshan Tianchi and Lake Xiaolongwan. Lake Xiaolongwan located in the Long Gang
volcanic field in Jilin Province, is a small maar lake (i.e. formed by a phreatomagmatic eruption) with a
maximum water depth of 15 m (Chu et al. 2009). Sifangshan Tianchi, located in the Great Xing’An Mountains
(Inner Mongolia) is a much smaller water body as only a small pond, less than 1 m deep, persists at the center of
the crater. In both sites, the water is slightly acidic with low electrical conductivity and relatively high dissolved
organic carbon concentration (Table 1). For Sifangshan, however, the values for total phosphorus (TP), total
nitrogen (TN) and dissolved silica were measured on a water sample that was contaminated with sediment.
In both surface-sediment samples, S. lacusvulcani is rare and represents less than 1% of the assemblages.
In the sediment core from Lake Xiaolongwan, S. lacusvulcani is most abundant (14.4%) in the sample taken at
400 cm core depth from the early part of the Holocene.
Discussion
Stauroneis lacusvulcani can be usefully compared with the group of species that include Stauroneis gracilior
Reichardt (1995: 34) and Stauroneis neohyalina Lange-Bertalot & Krammer (in Lange-Bertalot & Metzeltin
1996: 104) (Table 2), the most similar being Stauroneis gracilior “Zackenberg” (a manuscript name for a taxon
described, but not named, by Van de Vijver et al. 2004: 37) from East Greenland (Zackenberg Area) and
Stauroneis pikuni Bahls (2010: 115) described from Montana (USA). Bahls (2011) suggested that these two taxa
are synonyms. S. lacusvulcani differs from both by its larger dimension (valve length is 62–83 instead of 42–62
µm for S. gracilior ‘Zackenberg’, 53–64 µm for S. pikuni), its lanceolate shape (instead of rhombic-lanceolate),
the long-protracted and strongly capitate apices, the presence of a weakly developed hyaline zone and, in most
specimens, the presence of shortened striae in the stauros. S. gracilior sensu stricto, originally described from
French Guyana, is smaller and has a more conspicuous hyaline area around the stauros. S. neohyalina is much
smaller, has more linear valves and has a higher striae density. Stauroneis sofia Van De Vijver & Le Cohu (in
Van de Vijver, Gremmen, Beyens, & Le Cohu 2004: 104), described from the sub-Antarctic region, has valves
of similar dimension to S. lacusvulcani but its apices are less protracted and is characterized by a large hyaline
area. These differences justify the description of S.lacusvulcani as a new species.
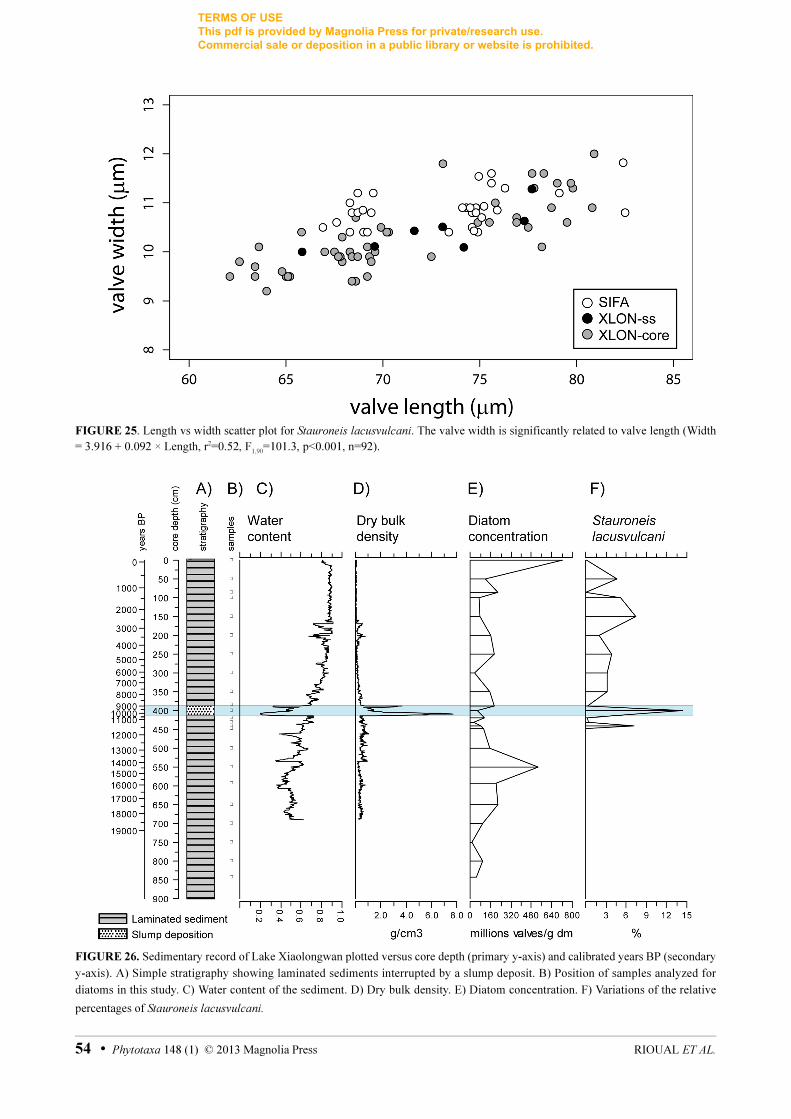
The boxplots for valve length (Fig. 22) and width (Fig. 23) indicate that the population from Sifangshan
has larger valve dimension than that from Xiaolongwan. However, there was no significant difference in striae
density (Fig. 24). Moreover, the ‘length vs width’ scatter plot shows that the two populations overlap and that
they do not occupy different size trajectories (Fig. 25). There is therefore no reason to suspect that these two
populations belong to different taxa. The difference in the distribution of lengths and widths between the two
populations may be due to a difference in the number of generations included in the samples (Tropper 1975).
The down-core sample from Xiaolongwan represents a much longer time span than the surface sediment
collected in Sifangshan and therefore may include a population than underwent more divisions and is hence
skewed towards the smaller sizes.
Most species of Stauroneis species are motile, epipelic diatoms (Round et al. 1990, Bahls 2011) and this is
probably also true for S. lacusvulcani. Currently in Lake Xiaolongwan, the development of the epipelon is
limited by a dense belt of Phragmites in the littoral zone and by rapid light attenuation under the deeper water.
Consequently, the assemblage found in the uppermost section of Lake Xiaolongwan sedimentary record is
dominated by planktonic (e.g. Discostella, Fragilaria) and epiphytic (e.g. Achnanthidium, Brachysira,
Encyonopsis, Tabellaria flocculosa) diatoms (Panizzo et al. 2013). The reduced extent of the lake area
available for the development of the epipelic community may explain why S. lacusvulcani is so rare in the
surface sediment sample we analyzed.
RIOUAL ET AL.52 • Phytotaxa 148 (1) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 22–24. Box plots showing quantile distributions of valves characters for populations of Stauroneis lacusvulcani observed
in the 2 localities: Sifangshan (n=35) and Xialongwan (n=7 for surface sediment and n=50 for core sample). 22. Valve length. 23.
Valve width. 24. Striae density (= number of striae in 10 µm). The 25–75 percent quartiles (excluding outliers) are drawn using a box.
The median is shown with a horizontal line inside the box. The whiskers represent the upper and lower “inner fence”, i.e. are drawn
from the edge of the box up to the largest/lowest data point less than 1.5 times the box height. Outliers, i.e. values outside the inner
fences, are shown as circles if they lie further from the edge of the box than 3 times the box height.
The diatom analysis of the sedimentary record from Lake Xiaolongwan indicates that S. lacusvulcani has
been present in that lake since at least the Late glacial with four peaks in relative abundances at ~11,450
(7.2%), between ~10,650 and ~8950 (14.4%), at ~2350 (7.4%) and ~600 (4.6%) calibrated years BP (Fig. 26).
The palaeolimnological investigation of this sedimentary record is still on-going and at this stage of our study
we cannot propose a definitive interpretation of these peaks of Stauroneis. However, it is interesting to note
that high values of S. lacusvulcani coincide with low values for diatom concentration. The peaks in
S.lacusvulcani may therefore correspond with intervals when the productivity of planktonic and epiphytic
diatoms communities decreased and when conditions became more favorable to the epipelon. The highest peak
of S. lacusvulcani occurred during an interval when the deposition of laminated sediment was interrupted by a
slump deposit (between the core depths 388 and 414 cm) clearly identified by sharp changes in water content
and dry bulk density (Fig. 26). This disturbance in sedimentation was most likely related to the volcanic
eruption of regional origin that occurred at ~10,460 cal. yrs BP (Stebich et al. 2009).
Phytotaxa 148 (1) © 2013 Magnolia Press • 53STAURONEIS LACUSVULCANI SP. NOV.
TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURE 25. Length vs width scatter plot for Stauroneis lacusvulcani. The valve width is significantly related to valve length (Width
= 3.916 + 0.092 × Length, r2=0.52, F1,90
=101.3, p<0.001, n=92).
FIGURE 26. Sedimentary record of Lake Xiaolongwan plotted versus core depth (primary y-axis) and calibrated years BP (secondary
y-axis). A) Simple stratigraphy showing laminated sediments interrupted by a slump deposit. B) Position of samples analyzed for
diatoms in this study. C) Water content of the sediment. D) Dry bulk density. E) Diatom concentration. F) Variations of the relative
percentages of Stauroneis lacusvulcani.
RIOUAL ET AL.54 • Phytotaxa 148 (1) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
TA
BL
E 2
: C
ompa
rison
of m
orph
olog
ical
feat
ures
for s
elec
ted
spec
ies o
f Sta
uron
eis
S.
lacu
s-vu
lcan
i S.
grac
ilior
–‘Z
acke
nber
g’
S.pi
kuni
S.
neoh
yalin
a S.
baco
nian
a S.
grac
ilior
S.
subg
raci
lior
S.so
fia
Ref
eren
ce
This
stud
y V
an d
e V
ijver
et a
l. (2
004)
B
ahls
(201
1)
Bah
ls (2
011)
B
ahls
(201
1)
Van
de
Vijv
er e
t al
. (20
04)
Van
de
Vijv
er e
t al.
(200
4)
Van
de
Vijv
er e
t al
. (20
04)
Val
ve le
ngth
62–8
3 46–6
2 53–6
4 32–4
9 73–1
39
47–5
3 30–3
8 47–8
2 V
alve
wid
th
9.2–
12.0
8.
5–10
.5
9–12
6.
0–9.
0 9.
0–18
.0
9 6.
5–8.
2 8–
10.5
V
alve
out
line
Lanc
eola
te
Rho
mbi
c-la
nceo
late
La
nceo
late
to
rhom
bic-
lanc
eola
te
Line
ar-
lanc
eola
te
Line
ar-
lanc
eola
te
Nar
row
ly
lanc
eola
te
Lanc
eola
te
Nar
row
ly
lanc
eola
te to
w
eakl
y rh
ombi
cal-
lanc
eola
te
Api
ces
Long
pro
tract
ed-
capi
tate
Pr
otra
cted
cap
itate
to
rost
rate
-cap
itate
N
arro
wed
ca
pita
te
Prot
ract
ed a
nd
narr
owly
rost
rate
to
subc
apita
te
Prot
ract
ed
rost
rate
Pr
otra
cted
cap
itate
Pr
otra
cted
rost
rate
to
rost
rate
-cap
itate
Pr
otra
cted
ro
stra
te-c
apita
te
Cen
tral a
rea
Rec
tang
ular
to
wea
kly
expa
nded
V
ery
wea
kly
expa
nded
N
arro
w,
Rec
tang
ular
R
ecta
ngul
ar
Nar
row
, re
ctan
gula
r R
ecta
ngul
ar
Rec
tang
ular
D
istin
ctly
ex
pand
ed
Hya
line
zone
ar
ound
fasc
ia
Wea
kly
deve
lope
d A
bsen
t A
bsen
t A
bsen
t A
bsen
t Pr
esen
t A
bsen
t Pr
esen
t
Cen
tral r
aphe
en
ding
s St
raig
ht, s
light
ly
expa
nded
St
raig
ht, i
ndis
tinct
St
raig
ht, n
ot
expa
nded
or
defle
cted
Stra
ight
, not
in
flate
d St
raig
ht, n
ot
expa
nded
St
raig
ht, s
light
ly
expa
nded
Sl
ight
ly b
ut
dist
inct
ly d
efle
cted
St
raig
ht,
indi
stin
ct
Rap
he ri
dge
Pres
ent
Pres
ent
Pr
esen
t A
bsen
t Pr
esen
t #s
triae
/10
µm
21–2
7 28–3
0 24–2
8 32–4
0 16
in th
e ce
ntre
, 20
at t
he e
nds
25–2
6 28–3
1 25–2
6
#pun
cta/
10
µm
18–2
8 25–2
7 20–2
4
14–1
8 15–2
5 26–3
1 25–3
0
Stria
tion
patte
rn
Alm
ost p
aral
lel i
n th
e m
iddl
e, m
ore
radi
ate
near
the
apic
es
Mod
erat
ely
radi
ate
in th
e m
iddl
e, m
ore
radi
ate
near
the
apic
es
Rad
iate
th
roug
hout
R
adia
te
thro
ugho
ut
Para
llel i
n th
e m
iddl
e to
radi
ate
near
the
apic
es
Mod
erat
ely
radi
ate
in th
e m
iddl
e, m
ore
radi
ate
near
the
apic
es
Mod
erat
ely
radi
ate
in th
e m
iddl
e, m
ore
radi
ate
near
the
apic
es
Mod
erat
ely
radi
ate
in th
e m
iddl
e, m
ore
radi
ate
near
the
apic
es
Phytotaxa 148 (1) © 2013 Magnolia Press • 55STAURONEIS LACUSVULCANI SP. NOV.

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
So far, S. lacusvulcani has been found in only two lakes. However, its distribution is not restricted to a
small geographical zone as the two lakes are located more than 800 km apart and the type of lake in which it
was found is rather common in northeastern China.
Acknowledgements
Dr Yan Fang (State Key Laboratory of Paleobiology and Stratigraphy, Nanjing) is thanked for assistance with
the SEM. This research was supported by the "Strategic Priority Research Program" of the Chinese Academy
of Sciences, Climate Change: Carbon Budget and Relevant Issues (grant # XDA05120201-2) and the National
Science Foundation of China (grant # 40502018).
References
Bahls, L. (2010) Stauroneis in the Northern Rockies: 50 species of Stauroneis sensu stricto from western Montana, northern Idaho,
northeastern Washington and southwestern Alberta, including 16 species described as new. Northwest Diatoms, 4. 172 pp.
Bahls, L. (2011) Stauroneis pikuni. In: Diatoms of the United States. In: http://westerndiatoms.colorado.edu/taxa/species/
stauroneis_pikuni/ (accessed: 2 August 2013)
Battarbee, R.W. & Kneen, M.J. (1982) The use of electronically counted microspheres in absolute diatom analysis. Limnology &
Oceanography 27: 184–188.
Chambers, J.M., Cleveland, W.S., Kleiner, B. & Tukey, P.A. (1983) Graphical methods for data analysis. Wadsworth & Brooks/Cole
Publishing Company, Pacific Grove, USA, 395 pp.
Chu, G., Sun, Q., Gu, Z., Rioual, P., Liu, Q., Wang, K., Han, J. & Liu, J. (2009) Dust records from varved lacustrine sediments of two
neighboring lakes in northeastern China over the last 1400 years. Quaternary International 194: 108–118.
http://dx.doi.org/10.1016/j.quaint.2008.08.005
Cortese, G. & Gersonde, R. (2007) Morphometric variability in the diatom Fragilariopsis kerguelensis: implications for Southern
Ocean paleoceanography. Earth and Planetary Science Letters 257: 526–544.
http://dx.doi.org/10.1016/j.epsl.2007.03.021
Lange-Bertalot, H., Cavacini, P., Tagliaventi, N. & Alfinito, S. (2003) Diatoms of Sardinia. Rare and 76 new species in rock pools and
other ephemeral waters. Iconographia Diatomologica 12, A.R.G. Gantner Verlag K.G, Ruggell, 438 pp.
Li, J. & Qi, Y. (2010) Flora algarum sinicarum aquae dulcis. Timus XIV, Bacillariophyta, Naviculaceae (I). Science Press, Beijing,
177 pp.
Mann, D.G., McDonald,S.M., Bayer,M.M., Droop,S.J.M., Chepurnov,V., Loke,R.E., Ciobanu,A. & du Buf,H. (2004) The Sellaphora
pupula species complex (Bacillariophyceae): morphometric analysis, ultrastucture and mating data provide evidence for five new
species. Phycologia 43: 459–482.
Panizzo, V.N., Mackay, A.W., Rose, N.L., Rioual, P. & Leng, M.J. (2013). Recent palaeolimnological change recorded in Lake
Xiaolongwan, northeast China: climatic versus anthropogenic forcing. Quaternary International 290–291: 322–334.
http://dx.doi.org/10.1016/j.quaint.2012.07.033
Paull, T.M., Hamilton, P.B., Gajewski, K. & LeBlanc, M. (2008) Numerical analysis of small Arctic diatoms (Bacillariophyceae)
representing the Staurosira and Staurosirella species complexes. Phycologia 47: 213–224.
http://dx.doi.org/10.2216/07-17.1
Renberg, I. (1990) A procedure for preparing large sets of diatom slides from sediment cores. Journal of Paleolimnology 4: 87–90.
http://dx.doi.org/10.1007/BF00208301
Rioual, P., Lu, Y., Yang, H., Scuderi, L.A., Chu, G., Holmes, J.A., Zhu, B. & Yang, X. (2013) Diatom-environment relationships and a
transfer function for conductivity in lakes of the Badain Jaran Desert, Inner Mongolia, China. Journal of Paleolimnology 50:
207–229.
http://dx.doi.org/10.1007/s10933-013-9715-9
Round, F.E., Crawford, R.M. & Mann, D.G. (1990) The diatoms. Biology and morphology of the genera. Cambridge University Press,
Cambridge, UK, 747 pp.
Stebich, M., Mingram, J. Han, J. & Liu, J. (2009) Late Pleistocene spread of (cool-)temperate forests in Northeast China and climate
changes synchronous with the North Atlantic region. Global and Planetary Change 65: 56–70.
http://dx.doi.org/10.1016/j.gloplacha.2008.10.010
Tropper, C.B. (1975) Morphological variation of Achnanthes hauckiana (Bacillariophyceae) in the field. Journal of Phycology 11:
297–302.
Van de Vijver, B., Beyens, L. & Lange-Bertalot, H. (2004) The genus Stauroneis in the Arctic and (Sub-) Antarctic regions.
Bibliotheca Diatomologica 51. J.Cramer, Berlin. 317 pp.
RIOUAL ET AL.56 • Phytotaxa 148 (1) © 2013 Magnolia Press




















