
Bortezomib induces apoptosis by interacting with JAK/STAT pathway in K562 leukemic cells
10
RESEARCH ARTICLE Bortezomib induces apoptosis by interacting with JAK/STAT pathway in K562 leukemic cells Nur Selvi & Burçin Tezcanli Kaymaz & Cumhur Gündüz & Çağdaş Aktan & Hatice Demet Kiper & Fahri Şahin & Melda Cömert & Ali Fatih Selvi & Buket Kosova & Güray Saydam Received: 11 April 2014 /Accepted: 1 May 2014 # International Society of Oncology and BioMarkers (ISOBM) 2014 Abstract In the current study, we aimed to identify the cyto- toxic and apoptotic effects of bortezomib (BOR) on human K562 chronic myelogenous leukemia cells and to evaluate the potential roles of Janus kinase/signal transducers and activa- tors of transcription (JAK/STAT) pathway members STAT3, STAT5, and JAK2 on BOR-induced cell death of leukemic cells. Cell viability was assessed via trypan blue dye exclusion test, and cytotoxicity of the BOR-treated cells was conducted by 2,3-bis(2-methoxy-4-nitro-5-sulphophenyl)-2H-tetrazoli- um-5-carboxanilide inner salt (XTT) assay. The relative mes- senger RNA (mRNA) expression levels of STAT3, STAT5A, STAT5B, and JAK2 were analyzed by quantitative reverse transcriptase-polymerase chain reaction (qRT-PCR). On the other hand, their protein expression levels were detected by western blot method. The obtained results indicated that BOR treatment reduced cell viability and induced leukemic cell apoptosis in a dose- and time-dependent manner as compared to untreated control cells. While mRNA expression levels of STAT5A, STAT5B, and STAT3 were significantly reduced following BOR treatment when compared to untreated con- trols, it had no effect upon JAK2 mRNA expression. As for protein levels, STAT expressions were downregulated after BOR treatment especially at 72nd and 96th hours. Our results pointed out that BOR treatment had a significant potential of being an anticancer agent for chronic myelogenous leukemia therapy, and this effect could be due to the expressional downregulations of JAK/STAT pathway members. Keywords BOR . CML . JAK/STAT pathway . Apoptosis Introduction Chronic myeloid leukemia (CML) is a chronic blood disorder characterized by a reciprocal translocation between chromo- somes 9 and 22, leading to the creation of a chimeric gene encoding BCR-ABL fusion protein with a constitutive tyro- sine kinase activity [1]. The resultant Bcr-Abl oncoprotein, a constitutively active tyrosine kinase, activates numerous sig- nal transduction pathways, leading to uncontrolled cell prolif- eration and reduced rates of apoptosis [2]. CML history has been significantly improved by the usage of imatinib—a tyrosine kinase inhibitor (TKI)—as first line therapy that was newly diagnosed as chronic-phase CML [3]. Imatinib has revolutionized the treatment of CML by transforming the process from an invariably fatal disease to a chronic, but still in a manageable, condition [4]. However, despite its advances, it was not curable among all patients due to intolerance to side effects, fails in therapy response, or gain of resistance. For these patients, other TKIs nilotinib and dasatinib are used as second-generation therapy that are ap- proved for the treatment of CML, after imatinib failure or intolerance. Besides these, additional genetic defects have also been observed in more than 80 % of patients in the blast crisis (BC) phase of CML, which is defined as the most aggressive phase of the disease [5]. To overcome these limitations, new strategies and alternative and effective therapy agents for controlling drug resistance exhibit an increasing interest of area among researchers. Hematopoietic cell proliferation and differentiation is reg- ulated by cytokines that are also known as interferons, N. Selvi (*) : B. T. Kaymaz : C. Gündüz : Ç. Aktan : B. Kosova Department of Medical Biology, Medical Faculty, Ege University, Bornova 35100, Izmir, Turkey e-mail: [email protected] H. D. Kiper : F. Şahin : M. Cömert : G. Saydam Department of Haemotology, Medical Faculty, Ege University, Bornova 35100, Izmir, Turkey A. F. Selvi Selimiye State Hospital, 22030 Edirne, Turkey Tumor Biol. DOI 10.1007/s13277-014-2048-0
Transcript of Bortezomib induces apoptosis by interacting with JAK/STAT pathway in K562 leukemic cells

RESEARCH ARTICLE
Bortezomib induces apoptosis by interacting with JAK/STATpathway in K562 leukemic cells
Nur Selvi & Burçin Tezcanli Kaymaz & Cumhur Gündüz & Çağdaş Aktan &
Hatice Demet Kiper & Fahri Şahin & Melda Cömert & Ali Fatih Selvi & Buket Kosova &
Güray Saydam
Received: 11 April 2014 /Accepted: 1 May 2014# International Society of Oncology and BioMarkers (ISOBM) 2014
Abstract In the current study, we aimed to identify the cyto-toxic and apoptotic effects of bortezomib (BOR) on humanK562 chronic myelogenous leukemia cells and to evaluate thepotential roles of Janus kinase/signal transducers and activa-tors of transcription (JAK/STAT) pathway members STAT3,STAT5, and JAK2 on BOR-induced cell death of leukemiccells. Cell viability was assessed via trypan blue dye exclusiontest, and cytotoxicity of the BOR-treated cells was conductedby 2,3-bis(2-methoxy-4-nitro-5-sulphophenyl)-2H-tetrazoli-um-5-carboxanilide inner salt (XTT) assay. The relative mes-senger RNA (mRNA) expression levels of STAT3, STAT5A,STAT5B, and JAK2 were analyzed by quantitative reversetranscriptase-polymerase chain reaction (qRT-PCR). On theother hand, their protein expression levels were detected bywestern blot method. The obtained results indicated that BORtreatment reduced cell viability and induced leukemic cellapoptosis in a dose- and time-dependent manner as comparedto untreated control cells. While mRNA expression levels ofSTAT5A, STAT5B, and STAT3 were significantly reducedfollowing BOR treatment when compared to untreated con-trols, it had no effect upon JAK2 mRNA expression. As forprotein levels, STAT expressions were downregulated afterBOR treatment especially at 72nd and 96th hours. Our resultspointed out that BOR treatment had a significant potential ofbeing an anticancer agent for chronic myelogenous leukemia
therapy, and this effect could be due to the expressionaldownregulations of JAK/STAT pathway members.
Keywords BOR . CML . JAK/STAT pathway . Apoptosis
Introduction
Chronic myeloid leukemia (CML) is a chronic blood disordercharacterized by a reciprocal translocation between chromo-somes 9 and 22, leading to the creation of a chimeric geneencoding BCR-ABL fusion protein with a constitutive tyro-sine kinase activity [1]. The resultant Bcr-Abl oncoprotein, aconstitutively active tyrosine kinase, activates numerous sig-nal transduction pathways, leading to uncontrolled cell prolif-eration and reduced rates of apoptosis [2].
CML history has been significantly improved by the usageof imatinib—a tyrosine kinase inhibitor (TKI)—as first linetherapy that was newly diagnosed as chronic-phase CML [3].Imatinib has revolutionized the treatment of CML bytransforming the process from an invariably fatal disease toa chronic, but still in a manageable, condition [4]. However,despite its advances, it was not curable among all patients dueto intolerance to side effects, fails in therapy response, or gainof resistance. For these patients, other TKIs nilotinib anddasatinib are used as second-generation therapy that are ap-proved for the treatment of CML, after imatinib failure orintolerance. Besides these, additional genetic defects have alsobeen observed in more than 80 % of patients in the blast crisis(BC) phase of CML, which is defined as the most aggressivephase of the disease [5]. To overcome these limitations, newstrategies and alternative and effective therapy agents forcontrolling drug resistance exhibit an increasing interest ofarea among researchers.
Hematopoietic cell proliferation and differentiation is reg-ulated by cytokines that are also known as interferons,
N. Selvi (*) : B. T. Kaymaz : C. Gündüz : Ç. Aktan :B. KosovaDepartment of Medical Biology, Medical Faculty, Ege University,Bornova 35100, Izmir, Turkeye-mail: [email protected]
H. D. Kiper : F. Şahin :M. Cömert :G. SaydamDepartment of Haemotology, Medical Faculty, Ege University,Bornova 35100, Izmir, Turkey
A. F. SelviSelimiye State Hospital, 22030 Edirne, Turkey
Tumor Biol.DOI 10.1007/s13277-014-2048-0

interleukins, and colony-stimulating factors. One of the mostimportant signaling cascades that regulate cell proliferationand apoptosis giving rise to the development of leukemia isthe Janus kinase/signal transducers and activators of transcrip-tion (JAK/STAT) signaling pathway [6]. Following dimeriza-tion and phosphorylation, STATs migrate into nucleus wherethey are activated and thus affecting cellular processes such astranscriptional regulations of several growth factors andoncoproteins [7]. In the pathway, constitutive JAK activationleads to persistent activation of STATs, and in consequence ofthis, several cancers exhibit permanent STAT activity [1].Among STATs, STAT3 activation has more effect upon lym-phomas and myelomas, and it has also been implicated inmalignant transformation, and its suppression directs tumorcells to apoptosis [8, 9]. STAT5A and STAT5B have also beenshown to be necessary for the development of malignancy,and they exhibit regulatory roles in the development of leu-kemia [10]. On the other hand, IL-6 induces transcription ofthe IL-6 gene via JAK2 and STAT3. This induction gives riseto increased autocrine production of IL-6 that is observed indifferent cancer cell lines. Thus, regulating JAK2 and/orSTAT3 could reduce IL-6 production, thereby impairing cellgrowth and enhancing their susceptibility to other treatments[11].
Bortezomib is a first class of proteasome inhibitor, current-ly approved in the USA and Europe for the treatment ofpatients diagnosed with multiple myeloma (MM) and relapsedor refractory mantle cell lymphoma [12]. However, it is un-known whether BOR could be an alternative drug in leukemiaor induce leukemic cell differentiation as being an apoptosisindicator. In this study, we aimed to investigate the apoptoticeffect of BOR treatment on leukemic cells and to clarify thepotential role of JAK/STAT pathway members upon BOR-induced apoptosis.
Material and methods
Cells
Human chronic myelogenous leukemia cell line K562 waspurchased from ECACC (European Collection of Cell Cul-tures). Cells were cultured in RPMI-1640 medium containing10 % (v/v) heat-inactivated fetal calf serum, 100 units ofpenicillin-streptomycin/ml, 1 %L-glutamine at 37 °C in hu-midified air containing 5 % CO2. Cells that were in logarith-mic growth phase were used in the experiments.
Cell proliferation assay
XTT cell proliferation assay was performed using differentconcentrations of BOR in order to obtain the IC50 value of itfor K562 cells with duration of 72 h. IC50 is the dose of an
ingredient that inhibits 50 % of the cells, and the value iscalculated via the detected cell proliferation curve. The assaywas triplicated.
Bortezomib treatment
K562 cells were treated with a concentration of 17.7 nMBORwhich stands for the calculated IC50 value. The experimentalsetup was designed for 24–96 h time course treatment of BORwith leukemia cells also with a nontreated control group. TheBOR was applied into the cells with reverse transfectionprotocol in order to supply the same number of cells at theend of 96 h. BOR treatment was revealed in order to define theexpressional changes of STATs and JAK2 at both messengerRNA (mRNA) and protein levels, and also the apoptotic caseof the cells. All experiments were triplicated and the averageof the results was taken.
Total RNA extraction and cDNA synthesis
After treatment with BOR for 0–96 h, total RNAs wereextracted from the cells according to the manufacturer’s pro-tocol (High Pure RNA Extraction Kit, Roche Diagnostic). Theamount and quality of the isolated RNAs were measured byNanoDrop spectrophotometer, and 100 ng RNAwas reverse-transcribed into complementary DNA (cDNA) via “FirstStrand cDNA Synthesis Kit, Roche Diagnostic” followingthe instructions.
Quantitative assessment of STAT mRNAs by real-timeqRT-PCR
The primer pairs and probes used for the amplification ofSTAT3 (NM_003150), STAT5A (NM_003152), and STAT5B(NM_012447) are previously described in our study [13].Target cDNA sequences were amplified by quantitative re-verse transcriptase-PCR (qRT-PCR) using a fluorescence-based real-time detection method with LightCycler v. 2.0instrument. The thermal cycling profiles were as follows:denaturation step for 10 min at 95 °C; amplification step for10 s at 95 °C, 11 s at 56 °C, 11 s at 72 °C with 45 cycles; and acooling step for 30 s at 40 °C. “LightCycler h-G6PDHHouse-keeping Gene Set” (Roche Diagnostic) kit was used as ahousekeeping gene according to the kit manual in order todefine the relative expression levels of STATs.
In each PCR reaction, 10-μl reaction mixture was preparedfor STAT3, STAT5A, or STAT5B target genes by adding3 mM MgCl2, 0.5 pmol/μl of each primer, 0.2 pmol/μl eachfluorogenic probes (Tib Molbiol), and 1 μl FastStart DNAMaster Hybridization probes enzyme with 2.5 μl of cDNA.As for 10-μl reference gene master mix, the reaction involved3 mM MgCl2, 1 μl of ready-to-use h-G6PDH detection mix,and 1 μl of FastStart DNA Master Hybridization probes
Tumor Biol.

enzyme with 2.5 μl of cDNA. The relative expression levelsof STATs were determined by the proportion of the targetvalue to reference value after using reference gene standardcurve established from different copy numbers of G6PDH.
Quantitative assessment of JAK2 mRNA level by qRT-PCR
Another qRT-PCR assay was carried out in order to define theexpression level of JAK2 with LightCycler 480 instrument andits software. Primers and universal probe library (UPL) probestargeting β-actin were used as housekeeping gene internal con-trols. PCR assay was performed by using TaqMan Master Kit(Roche Diagnostics) according to the instructions of the manu-facturer. The used UPL probes targeting either JAK2 or β-actinwere obtained from Roche Diagnostics’ “Universal Probe Li-brary,” and they were labeled with 6-carboxyfluorescein (FAM)at their 5′ end with a dark quencher dye near the 3′- end. Thethermal cycling profile was as follows: denaturation step for10 min at 95 °C, amplification step for 10 s at 95 °C, 30 s at56 °C, 1 s at 72 °C of 45 cycles, and a cooling step for 30 s at40 °C. Relative ratio (RR) of gene expressions was calculatedusing the following formula: target value/reference value (Ctvalues). Fold change of a target gene was calculated by dividingthe RR of control group to RR of BOR-treated group.
Determining STAT protein expression levels by western blotanalyses
For evaluating expressional changes of STAT proteins, initial-ly, protein extraction was done from BOR-treated cells, andafter determining protein amounts, western blot assay wascarried out.
Preparation of cytosolic fraction
K562 cells (2×106) were washed two times with ice-coldphosphate-buffered saline (PBS), and then cells were centri-fuged at 2,500 rpm for 5 min. After centrifugation step, pelletswere mixed with lysis reagent (ProteoJET Mammalian CellLysis Reagent, Fermantas), and protease inhibitors 0.1 mMleupeptin and 0.02 mM N-alpha-p-tosyl-L-lysinechloromethyl ketone (TLCK) were added. The homogenateswere immediately spinned at 1,000g for 10 min, and thesupernatant was spun at 100.000g for 1 h. The resultingsupernatant was used as the cytosolic fraction in order tomeasure protein amounts.
Assessment of protein amount
Total protein concentrations from the extracts were measuredby Bradford method via Bradford reagent (Fermentas). Theassay was standardized with bovine serum albumin (BSA)proteins ranging between 0.25 and 2 mg/ml concentrations,
and the protein amounts were detected with NanoDrop spec-trometer using the standard curve obtained from BSAstandards.
Western blot assay
Protein extracts were heated at 100 °C for 5 min with 4×concentrated Laemmli solution. Equal amount of protein sam-ples (20 μgrprotein/well) were loaded on to polyacrylamidegel to run electrophoresis (SDS-PAGE, %10). Then, the gelwas transferred on PVDF membrane (Biorad) using a wettransfer system (Bio-Rad). The blotting stage was done inprepared blocking buffer 5 % (w/v) milk powder in Tris-buffered saline (TBS) for 60 min at room temperature. Afterwashing steps, the membrane was exposed to varying primaryantibodies (STAT3, STAT5A, STAT5B, or β-actin) with1:2,000 dilutions for overnight at 4 °C. After washing stepswith TBS/T, secondary antibody IgG was applied with1:2,500 dilution for 90 min at room temperature. The proteinlevels of STATs in each line were detected by using theenhanced colorimetric detection kit (Immun-Blot alkalinephosphatase Assay kit, Bio-Rad), according to manufacturer’sinstructions. The western blot results were evaluated with gelimaging and analyses system (Chemi Smart 2000).
Apoptosis analyses
PE annexin V assay
Apoptotic case of the BOR-induced cells was evaluated by PEannexin V staining that allows detection of apoptosis anddifferentiates apoptosis from necrosis (PE Annnexin V, BDBiosciences). For this purpose, 2.5×105 cells were taken andthe kit procedure was applied. PE annexin V staining canidentify apoptosis at an earlier stage than assays based onnuclear changes such as DNA fragmentation. The number ofalive, apoptotic, and necrotic cells were evaluated at BDAccuri C6 Flow Cytometry (Becton–Dickinson, USA).
ApoDirect in situ DNA fragmentation assay
To evaluate apoptosis in BOR-treated cells, a single-stepstaining assay for labeling DNA breaks with FITCdUTPwas done using ApoDirect in situ DNA fragmentation kit(Biovision). For this experiment, K562 cells were seeded in6-well plates. Media was removed after 24-h incubation andreplaced with RPMI-1640 medium containing BOR at a con-centration of 17.7 nM. Control and BOR-treated cells wereincubated for 24, 48, 72, and 96 h. After centrifugation, thecell pellet was resuspended in 500 μl of PBS. Paraformalde-hyde (1 %) in PBS was then added to the cell suspension andincubated for 15 min on ice. After two times washing withPBS, the cell pellet again was resuspended in 70 % ethanol
Tumor Biol.

and stored at −20 °C. The cell samples were then pelleted,washed, and resuspended in 50 μl of staining solution con-taining reaction buffer, terminal deoxynucleotidyl transferaseenzyme, FITC-labeled deoxyuridine triphosphate nucleotides,and deionized water. After 60-min incubation at 37 °C, cellswere again washed, pelleted, and resuspended in 250 μl ofpropidium iodine/RNase solution and incubated for 30 mi-nutes in RT. The protocol was provided by the manufacturerand followed accordingly including the use of positive andnegative controls. Apoptosis was evaluated using BD AccuriC6 Flow Cytometry (Becton–Dickinson, USA) utilizing a488-nm argon laser light source for excitation and a 530-nmband-pass filter for FITC fluorescence (FL-1H) and a fluores-cence detector equipped with a 585/42 band-pass filter forFL2A. A total of 20,000 events were acquired for analysisusing Cell Quest Software.
Statistical analyses
The mRNA expression levels and apoptosis analyses wereevaluated with Student’s t test referring to significance ofp<0.05.
Results
Assessment of cellular proliferation by XTT
XTT cell proliferation assay was carried out by using in-creased concentrations of BOR, also with untreated cells intriplicates in a time-dependent manner. This assay was result-ed in gain of cell proliferation curves that enabled the calcu-lation of IC50 value, determined as 17.7 nM at the 72nd hour(Fig. 1).
BOR reduces STAT mRNA expression levels in K562 cells
Following determination of IC50 value, this BOR dose wasapplied to cells, and their relative mRNA expression rates forSTATs and JAK2were analyzed via qRT-PCR. Following 24–96 h of BOR treatment and their untreated counterparts,significant decreases were detected at both mRNA and proteinexpression levels of STATs especially for the last 2 days.
STAT5B mRNA expression was significantly dwindlingby 92 and 98 % in a time-dependent manner for the 72nd and96th hours (4.25-fold, p<0.0001; 4.38-fold, p=0.022), re-spectively, when compared to untreated control group(Fig. 2). As for STAT5A, its expression was prominentlyreduced by 85 and 90 % (3.85-fold, p=0.003; 4.02-fold, p=0.041) especially at the 48th and 72nd hours, respectively, inBOR-treated K562 cells compared to control group (Fig. 3).
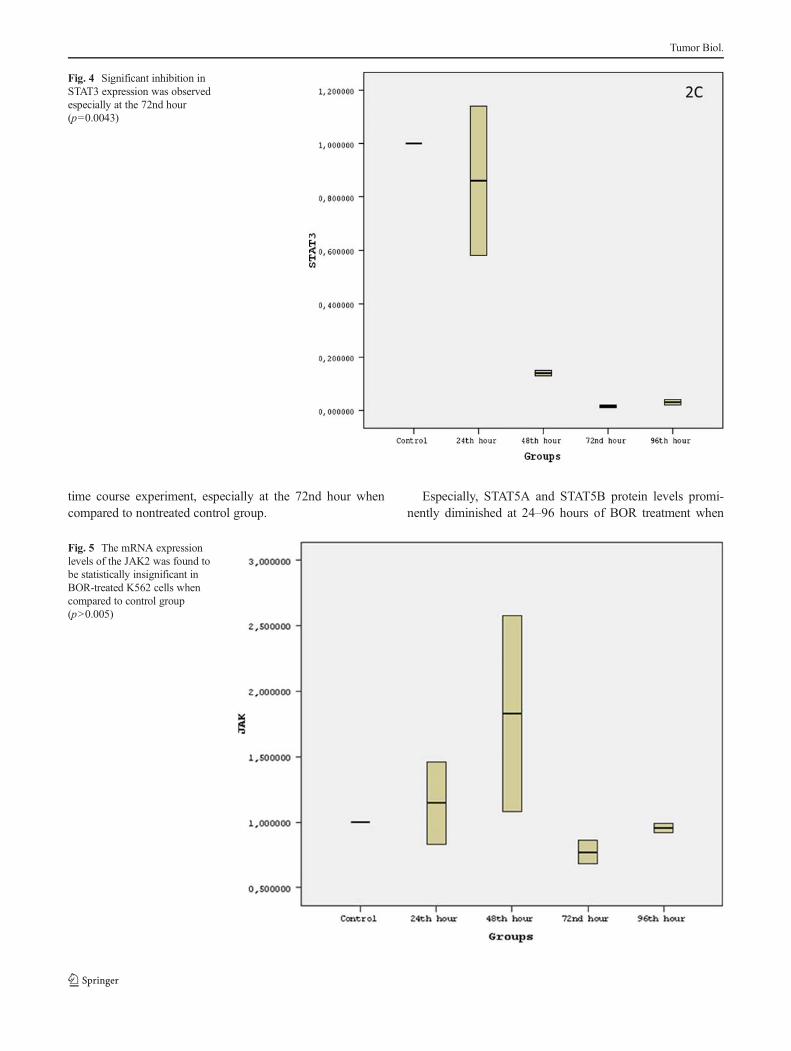
On the other hand, while STAT3 expression was signifi-cantly decreased by 66 % (2.86-fold; p=0.0043) at the 72ndhour (Fig. 4), the changes in the expression levels of JAK2was found to be insignificant in BOR-treated K562 cells whencompared to control group in any day of time course experi-ment (Fig. 5) giving rise to a conclusion that JAK2 did notplay major roles on biological effect of BOR upon leukemiccells. As a result, our mRNA expression results indicated that,initially, STAT3 and especially STAT5A and STAT5B couldhave played crucial roles in BOR mechanism upon CMLcells.
Bortezomib induced apoptosis in leukemic cells
Apoptotic cells were examined with flow cytometer by using“BDAnnexin V” and “ApoDirect in situ DNA fragmentation”kits, and changes in apoptosis rates were identified. As for ourBD annexin Vapoptosis assay results, the number of apoptoticcells was found to be increasing following BOR treatment anddetected as 57.2, 60.7, and 52.5 % of apoptotic cells for 24–72-h duration, but a dramatic increase was found at the 96thhour of the treatment by 70.3 % of apoptosis. As forApoDirect in situ DNA fragmentation assay results, we deter-mined induction of apoptosis in a time-dependent manner.While 24.3 and 30.3 % of apoptosis rates were seen at 24thand 48th hours, 57.5 and 58.8 % apoptosis induction wasdetected especially at the 72nd and 96th hours. Eventually,flow cytometry results showed that the percentage of apopto-tic cells were increased gradually following BOR treatment byboth of the assays. All results are shown in Fig. 6.
Western blot analysis results
The leukemic cells that were treated with BOR resulted indecreased expression of STAT proteins obtained at 24th–96thhours as in the same line with mRNA expression level results.STAT3 protein illustrated a decrease in its expression in our
Fig. 1 Percentage changes in cytotoxicity of bortezomib treatment. Theconcentration of BOR that inhibited cell growth by 50 % (IC50) wascalculated from cytotoxicity plots. The XTT assay was performed usingtriplicate samples in at least two independent experiments. The IC50 valuewas calculated as 17.7 nM at the 72nd hour
Tumor Biol.
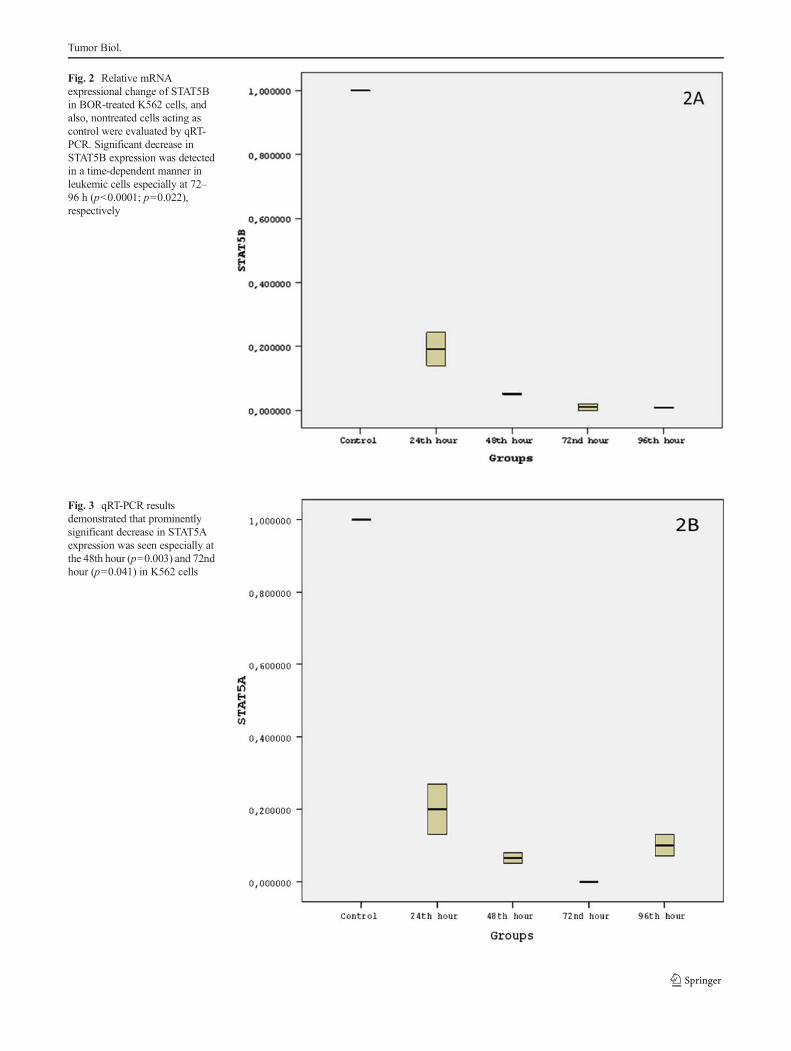
Fig. 2 Relative mRNAexpressional change of STAT5Bin BOR-treated K562 cells, andalso, nontreated cells acting ascontrol were evaluated by qRT-PCR. Significant decrease inSTAT5B expression was detectedin a time-dependent manner inleukemic cells especially at 72–96 h (p<0.0001; p=0.022),respectively
Fig. 3 qRT-PCR resultsdemonstrated that prominentlysignificant decrease in STAT5Aexpression was seen especially atthe 48th hour (p=0.003) and 72ndhour (p=0.041) in K562 cells
Tumor Biol.
time course experiment, especially at the 72nd hour whencompared to nontreated control group.
Especially, STAT5A and STAT5B protein levels promi-nently diminished at 24–96 hours of BOR treatment when
Fig. 4 Significant inhibition inSTAT3 expression was observedespecially at the 72nd hour(p=0.0043)
Fig. 5 The mRNA expressionlevels of the JAK2 was found tobe statistically insignificant inBOR-treated K562 cells whencompared to control group(p>0.005)
Tumor Biol.

confronted with untreated control group. These inhibitions atprotein levels of STATs could be the result of triggered apo-ptosis. But, no expressional change or inhibition was detectedon JAK2 protein level (Fig. 7).
Discussion
Normal hematopoiesis has signaling pathways in which cyto-kines and their receptors play essential roles. Anomalies,which can be arisen by these ways, result in malignant trans-formation, decreasing apoptosis, and uncontrolled prolifera-tion. For these reasons, signal transduction pathways havebecome convenient targets in advanced leukemia therapy.The roles of JAK/STAT, Raf/MEK/Erk, and PI3K/Akt ininduction of leukemic progression are cell cycle regulatoryand signal transduction pathways which point out the impor-tance of apoptosis in hematopoiesis [14–17]. Although manychemotherapeutic agents have been used for the treatment ofleukemia, there are still some limitations due to gain of resis-tance or intolerance, although targeted and alternative therapystrategies are important phenomenons nowadays [18–21].
In this study, we initially aimed to determine the potentialcytotoxic and apoptotic effects of bortezomib on K562 cellline and to investigate whether this probable effect was uponJAK/STAT pathway components STAT3, STAT5A, STAT5B,and JAK2. We have revealed that BOR treatment causedcytotoxicity upon leukemic cells and triggered their apoptosisin a time-dependent manner. The apoptotic effect was also
described in another study by Zheng et al. [22] in whichBortezomib potently inhibited growth and increased the apo-ptosis rate in the described cell lines: a multifactor-resistantcell line K562/A02 and its sensitive parental cell.
Although there is no reported study that included the im-pact of BOR on leukemia upon JAK/STAT pathway compo-nents in response to treatment until now, Zhou et al. exploredthe effect of bortezomib on the drug sensitivity of imatinib-resistant chronic myeloid leukemia cell line K562/G0, andthey concluded that BOR enhanced imatinib sensitivity ofimatinib in resistant K562/G01 cells. In their study, the authorsspeculated that this mechanism might be related to cell cyclephase arrested at G2/M and downregulation of COX-2 andMDR1 expression [23].
In the second part of our study, we examined STATs andJAK2 expressions following BOR treatment. It is known thatSTAToverexpression is frequently observed in human cancers[24–26], and among STATs, while STAT3 protein acts as anoncogene, other STAT proteins may function in oncogenictransformation [8]. In this study, STAT3 was found to be theleast downregulated gene among STATs, giving rise to aconclusion that STAT3 was more likely to be related withlymphomas rather than CML. But, we have shown that BORtreatment reduced STAT5B mRNA relative expression levelssignificantly in K562 cells in a time-dependent manner, espe-cially at the 72nd and 96th hours with inhibition rates of 92and 98 %, respectively. Also, a prominent downregulation ofSTAT5A mRNA expression level was observed with a sup-pression rate of 90 % for K562 cells at the 72nd hour of BOR
Fig. 6 Apoptosis assay in K562 cells. Untreated control and bortezomib-treated cells for a duration of 24–96 h. Apoptotic cells were assessed byqualitative flow cytometry. a The BD annexin Vassay: While left plateau
shows viable cells, the right plateau illustrates apoptotic, respectively. bApoDirect In Situ DNA fragmentation assay: While upper cell groupexhibits apoptotic cells, the lower group shows viable cells, respectively
Tumor Biol.

treatment. Finally, we can accomplish that STAT5A andSTAT5B are very crucial and play major roles in the effectmechanism of BOR upon leukemia, indicating the magnitudeof JAK/STAT pathway in targeted therapy of leukemia. As forJAK2, JAKs likely to have additional roles in promoting drugresistance in several cancer cell types [11], and they serve asuseful targets in the clinic [7]. On the other hand, constitutive-ly active STAT proteins could be effective in malign processby stimulating antiapoptotic ways. In our study, we did notobserve any expressional downregulation of JAK2 followingBOR treatment.
In the last part of the study, we evaluated STAT proteinlevels by western blot analysis. The used antibodiesagainst STAT5A and STAT5B showed expressional in-hibition in a time-dependent manner parallel to mRNAexpression results. As for STAT3 protein levels, weagain observed similar results with STAT3 mRNA ex-pression levels. This observed decreases in STAT pro-tein expression could be related with high-apoptosis rates. Atthe end of this assay, we concluded that STATs might havefundamental roles in the inhibition mechanism of BOR uponof leukemic cells.
Fig. 7 K562 cells were culturedfor 96 h in the presence of17.7 nM BOR, also withnontreated control groups. Cellextracts were analyzed by westernblot analysis everyday in order toassess the content ofJAK2/STATs;β-actin was used asa loading control. The targetSTAT5A and STAT5B proteinexpressions were downregulatedin K562 cells, and STAT3expression was downregulated atthe 72nd hour. M marker, Ccontrol group, 24–96 time courseof study period
Tumor Biol.

There can be two explanations for decreased mRNA andprotein expression levels related with BOR effect mech-anism upon leukemic cells. First, we think that BORmight be effective by inhibiting antiapoptotic proteins.In a study, anticancer mechanism of BOR was describedas, which included inhibition of Nf-KB and itsantiapoptotic target genes, suppression of severalantiapoptotic proteins ( e.g., BcL-XL, BCL-2, andSTAT-3) [27]. Another approach is that the suppressionof STATs following BOR treatment might be due to theactivation of suppressors of cytokine signaling (SOCS)proteins that function as STAT inhibitors. Especially,SOCS1 and SOCS3 are inhibitors of JAK-STAT path-way [28]. In this recent study, they have reported thatSOCS-1 was highly tyrosine phosphorylated in one offive Bcr-Abl-positive CML samples. Disrupting the ty-rosine phosphorylation of SOCS-1 and SOCS-3 promotedthe apoptosis of K562 cells and blocked the tumor formationin nude mice. Taken together, these results revealed a require-ment for tyrosine phosphorylation of SOCS-1 and SOCS-3 inBcr-Abl-induced tumorigenesis in the presence of these SOCSproteins [28].
When all observed qRT-PCR and western blot resultswere evaluated together, we concluded that STAT5A,STAT5B, and STAT3 appear to be playing more effi-cient roles in the pathogenesis and development ofCML. As for BD annexin V assay and ApoDirect in situDNA fragmentation assays, our results indicated that, espe-cially, suppression of STAT5A, STAT5B, and STAT3 expres-sions leads to induction of apoptosis and referred as candidatetarget genes for CML. Also, there is another study that sug-gested STAT molecules as novel therapeutic targets for onco-genic transformation [29].
In summary, in this study, we have shown that BOR hadcytotoxic and apoptotic effects on K562 chronic myelogenousleukemia cells, and STAT5A and STAT5B might have criticalroles in BOR-induced cell death. If these in vitro applicationscould be applied into clinics, we think that BOR might con-tribute to CML treatment in the future.
References
1. Nasr R, Bazarbachi A. Chronic myeloid leukemia. "Archetype" ofthe impact of targeted therapies. Pathol Biol. 2012;60(4):230–45.
2. Ramirez P, Dipersio JP. Therapy options in imatinib failures. TheOncologist Leukemias. 2008;13:424–34.
3. Bixby DL. Managing inadequate responses to frontline treatment ofchronic myeloid leukemia: a case-based review. Cancer Treat Rev.2013;39(3):241–51.
4. Kantarjian HM, Shah NP, Cortes JE, Baccarani M, Agarwal MB,Undurraga MS, et al. Dasatinib or imatinib in newly diag-nosed chronic-phase chronic myeloid leukemia: 2-year follow-up
from a randomized phase 3 trial (DASISION). Blood. 2012;119(5):1123–9.
5. Steinberg M, Steinberg M. BCOP. New drug dasatinib: a tyrosinekinase inihibitor for the treatment of chronic myelegenous leukemiaand philadelphia chrosome-positive acute lymphoblastic leukemia.Clinical Thepeutics. 2007;29(11):2289–308.
6. Smoak KA, Cidlowski JA. Mechanisms of glucocorticoid receptorsignaling during inflammation. Mech Ageing Dev. 2004;125:697–706.
7. Bromberg JF. Activation of STAT proteins and growth control.Biogeosciences. 2001;23:161–9.
8. Bromberg J. Stat proteins and oncogenesis. J Clin Invest. 2002;109:1139–42.
9. Catlett-Falcone R, Landowski TH, Oshiro MM, et al. Constitutiveactivation of Stat3 signaling confers resistance to apoptosis in humanU266 myeloma cells. Immunity. 1999;10:105–15.
10. Levy DE, Gilliland DG. Divergent roles of Stat1 and Stat5 in malig-nancy as revealed by gene disruptions in mice. Oncogene. 2000;19:2505–10.
11. Costa-Pereira AP, Bonito NA, Seckl MJ. Dysregulation of januskinases and signal transducers and activators of transcription incancer. Am J Cancer Res. 2011;1(6):806–16.
12. Richardson PG, Hideshima T, Anderson KC. Bortezomib (PS-341).A novel, first-in-class proteasome inhibitor for the treatment ofmultiple myeloma and other cancer. Cancer Control. 2003;10(5):361–9.
13. Kaymaz BT, Selvi N, Saydam G, Şahin F, Kosova B.Methylprednisolone induces apoptosis by interacting with theJAK/STAT pathway in HL–60 and K–562 leukemic cells.Hematology. 2012;17(2):93–9.
14. Lee Jr JT, McCubrey JA. The Raf/MEK/ERK signal transductioncascade as a target for chemotherapeutic intervention in leukemia.Leukemia. 2002;16:486–507.
15. Steelman LS, Abrams SL, Whelan J, Bertrand FE, Ludwig DE,Bäsecke J, et al. Contributions of the Raf/MEK/ERK, PI3K/PTEN/Akt/mTOR and Jak/STAT pathways to leukemia. Leukemia.2008;22(4):686–707.
16. McCubrey JA, Steelman LS, Abrams SL, Bertrand FE, Ludwig DE,Bäsecke J, et al. Targeting survival cascades induced by activation ofRas/Raf/MEK/ERK, PI3K/PTEN/Akt/mTOR and Jak/STAT path-ways for effective leukemia therapy. Leukemia. 2008;22(4):708–22.Review.
17. Steelman LS, Pohnert SC, Shelton JG, Franklin RA, Bertrand FE,McCubrey JA. JAK/STAT, Raf/MEK/ERK, PI3K/Akt and BCR-ABL in cell cycle progression and leukemogenesis. Leukemia.2004;18(2):189–218. Review.
18. Sacha T, Hochaus A, Hanfstein B, Muller MC, Rudzki Z, Czopek J,et al. Abl-kinase domain point mutation as a cause of imatinib crisis.Leuk Res. 2003;27:1163–6.
19. Hochaus A. Cytogenetic and molecular mechanisms of resistance toimatinib. Semin Hematol. 2003;40:69–79.
20. Tipping AJ, Melo JV. Imatinib mesylate in combination with otherchemotherapeutic drugs: in vitro studies. Semin Hematol. 2003;40:83–91.
21. Roche-Lestiennc C, Preudhomme C. Mutations in the Abl kinasedomain pre-exist the conset of imatinib treatment. Semin Hematol.2003;40:80–2.
22. Zheng B, Zhou R, Gong Y, Yang X, Shan Q. Proteasomeinhibitor bortezomib overcomes P-gp-mediated multidrug re-sistance in resistant leukemic cell lines. Int Jnl Lab Hem. 2012;34:237–47.
23. Zhou Y, Ma LM, Li XY, Zhang HP, Wang T, Niu YY, et al. Effect ofbortezomib on the drug sensitivity of imatinib resistant K562/G01cells. Zhonghua Xue Ye Xue Za Zhi. 2011;32(6):292–5.
24. Danial NN, Pernis A, Rothman PB. Jak–STAT signaling induced bythe v-abl oncogene. Science. 1995;269:1875–7.
Tumor Biol.

25. Danial NN, Rothman P. JAK–STAT signaling activated by Abloncogenes. Oncogene. 2000;19:2523–31.
26. Migone TS, Lin JX, Cereseto A, Mulloy JC, O’Shea JJ, Franchini G,et al. Constitutively activated Jak–STAT pathway in T cells trans-formed with HTLV-I. Science. 1995;269:79–81.
27. Muj Taba T, Dou QP. Advences in the understanding of mechanismand therapeutic use of Bortezomib. DiscovMed. 2011;12(67):471–80.
28. Qui X, Guo G, Chen K, Kashiwada M, Druker BJ, Rothman PB,et al. A requirement for SOCS-1 and SOCS-3 phosphorylation inBcr–ABL–induced tumorigenesis. Neoplasia. 2012;14(6):547–58.
29. Steelman LS, Pohnert SC, Shelton JG, Franklin RA, Bertrand FE,McCubrey JA. JAK/STAT, Raf/MEK/ERK, PI3K/Akt and BCR-ABL in cell cycle progression and leukemogenesis. Leukemia.2004;18:189–218.
Tumor Biol.




















