
Bone marrow stromal cell transplantation for treatment of sub-acute spinal cord injury in the rat
16
Research Report Bone marrow stromal cell transplantation for treatment of sub-acute spinal cord injury in the rat Chizuka Ide a,c, ⁎ , Yoshiyasu Nakai c , Norihiko Nakano c , Tae-Beom Seo c , Yoshihiro Yamada a,b , Katsuaki Endo b , Toru Noda b , Fukuki Saito f , Yoshihisa Suzuki d , Masanori Fukushima e , Toshio Nakatani f a Department of Occupational Therapy, Faculty of Nursing and Rehabilitation, Aino University, Higashi-ohda, 4-5-4, Ibaragi City, Osaka, Japan b Department of Physical Therapy, Faculty of Nursing and Rehabilitation, Aino University, Higashi-ohda, 4-5-4, Ibaragi City, Osaka, Japan c Institute of Regeneration and Rehabilitation, Aino University, Higashi-ohda, 4-5-4, Ibaragi City, Osaka, Japan d Department of Plastic and Reconstructive Surgery, Tazuke Medical Research Institute, Kitano Hospital, Osaka, Japan e Translational Research Center, Kyoto University Graduate School of Medicine, Kyoto, Japan f Emergency and Clinical Care Center, Kansai Medical University, Osaka, Japan ARTICLE INFO ABSTRACT Article history: Accepted 15 March 2010 Available online 19 March 2010 Bone marrow stromal cells (BMSCs) have been studied as effective transplants for the treatment of spinal cord injury (SCI). Our previous study showed that BMSCs infused into the cerebrospinal fluid (CSF) exhibited distinct effects on the recovery of acute SCI. The present study examined the effects of BMSCs in sub-acute SCI (2 weeks post-injury) by transplanting them directly into the lesion. The spinal cord was crush-injured at the Th8-9 level in rats, and 2 weeks later, cultured BMSCs (5 × 10 5 ) derived from GFP-transgenic rats of the same strain were transplanted into the lesion. Tissue repair and nerve regeneration were examined by immunohistochemistry and electron microscopy. GFP-labeled BMSCs survived as cell assemblies in the spinal cord for 1-2 weeks after transplantation. The dorsal side of BMSC assemblies in the spinal cord usually showed an expanded GFAP-negative, astrocyte- devoid area, in which extracellular matrices including collagen fibrils were deposited. Numerous regenerating axons associated with Schwann cells grew out through such astrocyte-devoid extracellular matrices. Ascending (CGRP-containing) and descending (5HT- and TH-containing) axons were included in these regenerating axons. Regenerated axons were myelinated by Schwann cells beyond 2 weeks post-transplantation. Cavity formation was reduced in the cell transplantation group. Locomotory behavior assessed by the BBB scale improved to 9.8 points in the cell transplantation group, while it was to 5.5-5.7 in the control. BMSC transplantation into lesions of advanced SCI has markedly beneficial effects on tissue repair and axonal outgrowth, leading to improved locomotion in rats. © 2010 Elsevier B.V. All rights reserved. Keywords: Bone marrow stromal cell Sub-acute spinal cord injury Delayed cell transplantation Axonal regeneration Behavior Trophic factor BRAIN RESEARCH 1332 (2010) 32 – 47 ⁎ Corresponding author. Institute of Regeneration and Rehabilitation, Department of Occupational Therapy, Faculty of Nursing and Rehabilitation, Aino University, 4-5-11 Higashi-ohta, Ibaraki City, Osaka, 567-0012, Japan. Fax: + 81 72 627 7882. E-mail address: [email protected] (C. Ide). 0006-8993/$ – see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.brainres.2010.03.043 available at www.sciencedirect.com www.elsevier.com/locate/brainres
Transcript of Bone marrow stromal cell transplantation for treatment of sub-acute spinal cord injury in the rat

B R A I N R E S E A R C H 1 3 3 2 ( 2 0 1 0 ) 3 2 – 4 7
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /b ra i n res
Research Report
Bone marrow stromal cell transplantation for treatment ofsub-acute spinal cord injury in the rat
Chizuka Idea,c,⁎, Yoshiyasu Nakaic, Norihiko Nakanoc, Tae-Beom Seoc,Yoshihiro Yamadaa,b, Katsuaki Endob, Toru Nodab, Fukuki Saito f, Yoshihisa Suzukid,Masanori Fukushimae, Toshio Nakatanif
aDepartment of Occupational Therapy, Faculty of Nursing and Rehabilitation, Aino University, Higashi-ohda, 4-5-4, Ibaragi City, Osaka, JapanbDepartment of Physical Therapy, Faculty of Nursing and Rehabilitation, Aino University, Higashi-ohda, 4-5-4, Ibaragi City, Osaka, JapancInstitute of Regeneration and Rehabilitation, Aino University, Higashi-ohda, 4-5-4, Ibaragi City, Osaka, JapandDepartment of Plastic and Reconstructive Surgery, Tazuke Medical Research Institute, Kitano Hospital, Osaka, JapaneTranslational Research Center, Kyoto University Graduate School of Medicine, Kyoto, JapanfEmergency and Clinical Care Center, Kansai Medical University, Osaka, Japan
A R T I C L E I N F O
⁎ Corresponding author. Institute of RegenerRehabilitation, Aino University, 4-5-11 Higas
E-mail address: [email protected] (C. I
0006-8993/$ – see front matter © 2010 Elsevidoi:10.1016/j.brainres.2010.03.043
A B S T R A C T
Article history:Accepted 15 March 2010Available online 19 March 2010
Bone marrow stromal cells (BMSCs) have been studied as effective transplants for thetreatment of spinal cord injury (SCI). Our previous study showed that BMSCs infused into thecerebrospinal fluid (CSF) exhibited distinct effects on the recovery of acute SCI. The presentstudy examined the effects of BMSCs in sub-acute SCI (2 weeks post-injury) by transplantingthem directly into the lesion. The spinal cord was crush-injured at the Th8-9 level in rats,and 2 weeks later, cultured BMSCs (5×105) derived from GFP-transgenic rats of the samestrain were transplanted into the lesion. Tissue repair and nerve regeneration wereexamined by immunohistochemistry and electronmicroscopy. GFP-labeled BMSCs survivedas cell assemblies in the spinal cord for 1-2 weeks after transplantation. The dorsal side ofBMSC assemblies in the spinal cord usually showed an expanded GFAP-negative, astrocyte-devoid area, in which extracellular matrices including collagen fibrils were deposited.Numerous regenerating axons associated with Schwann cells grew out through suchastrocyte-devoid extracellular matrices. Ascending (CGRP-containing) and descending(5HT- and TH-containing) axons were included in these regenerating axons. Regeneratedaxons were myelinated by Schwann cells beyond 2 weeks post-transplantation. Cavityformation was reduced in the cell transplantation group. Locomotory behavior assessed bythe BBB scale improved to 9.8 points in the cell transplantation group, while it was to 5.5-5.7in the control. BMSC transplantation into lesions of advanced SCI has markedly beneficialeffects on tissue repair and axonal outgrowth, leading to improved locomotion in rats.
© 2010 Elsevier B.V. All rights reserved.
Keywords:Bone marrow stromal cellSub-acute spinal cord injuryDelayed cell transplantationAxonal regenerationBehaviorTrophic factor
ation and Rehabilitation, Department of Occupational Therapy, Faculty of Nursing andhi-ohta, Ibaraki City, Osaka, 567-0012, Japan. Fax: +81 72 627 7882.de).
er B.V. All rights reserved.
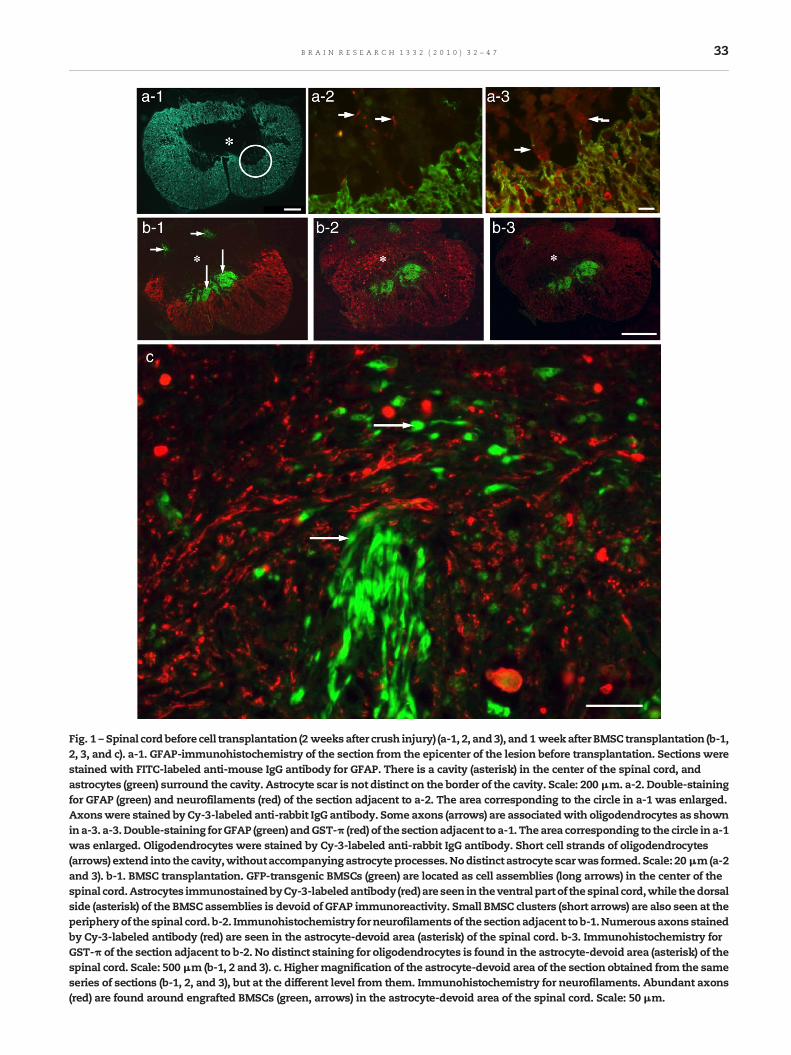
Fig. 1 – Spinal cordbefore cell transplantation (2 weeksafter crush injury) (a-1, 2, and3), and1 weekafter BMSC transplantation (b-1,2, 3, and c). a-1. GFAP-immunohistochemistry of the section from the epicenter of the lesion before transplantation. Sections werestained with FITC-labeled anti-mouse IgG antibody for GFAP. There is a cavity (asterisk) in the center of the spinal cord, andastrocytes (green) surround the cavity. Astrocyte scar is not distinct on the border of the cavity. Scale: 200 μm. a-2. Double-stainingfor GFAP (green) and neurofilaments (red) of the section adjacent to a-2. The area corresponding to the circle in a-1 was enlarged.Axonswere stained by Cy-3-labeled anti-rabbit IgG antibody. Some axons (arrows) are associatedwith oligodendrocytes as shownina-3. a-3.Double-staining forGFAP (green) andGST-π (red) of the sectionadjacent to a-1. Thearea corresponding to the circle in a-1was enlarged. Oligodendrocytes were stained by Cy-3-labeled anti-rabbit IgG antibody. Short cell strands of oligodendrocytes(arrows) extend into the cavity,withoutaccompanyingastrocyte processes.Nodistinct astrocyte scarwas formed. Scale: 20μm(a-2and 3). b-1. BMSC transplantation. GFP-transgenic BMSCs (green) are located as cell assemblies (long arrows) in the center of thespinal cord.Astrocytes immunostainedbyCy-3-labeledantibody (red)are seen in theventral part of thespinal cord,while thedorsalside (asterisk) of the BMSC assemblies is devoid of GFAP immunoreactivity. Small BMSC clusters (short arrows) are also seen at theperipheryof the spinal cord. b-2. Immunohistochemistry forneurofilaments of the sectionadjacent to b-1.Numerousaxons stainedby Cy-3-labeled antibody (red) are seen in the astrocyte-devoid area (asterisk) of the spinal cord. b-3. Immunohistochemistry forGST-π of the section adjacent to b-2. No distinct staining for oligodendrocytes is found in the astrocyte-devoid area (asterisk) of thespinal cord. Scale: 500 μm (b-1, 2 and 3). c. Highermagnification of the astrocyte-devoid area of the section obtained from the sameseries of sections (b-1, 2, and 3), but at the different level from them. Immunohistochemistry for neurofilaments. Abundant axons(red) are found around engrafted BMSCs (green, arrows) in the astrocyte-devoid area of the spinal cord. Scale: 50 μm.
33B R A I N R E S E A R C H 1 3 3 2 ( 2 0 1 0 ) 3 2 – 4 7

34 B R A I N R E S E A R C H 1 3 3 2 ( 2 0 1 0 ) 3 2 – 4 7
1. Introduction
Various kinds of cells have so far been used in transplantationstudies of spinal cord injury (SCI) (Enzmann et al., 2006), whichinclude Schwann cells (Xu et al., 1999; Golden et al.,2007; Someya et al., 2008), embryonic stem cells (McDonaldet al., 1999), macrophages (Rapalino et al., 1998; Knoller et al.,2005), neural stem cells (Wu et al., 2002; Hofstetter et al., 2005),olfactory ensheathing cells (Li et al., 1997; Ramon-Cueto et al.,2000; Plant et al., 2003), choroid plexus (Ide et al., 2001;Matsumoto et al., 2003), epidermal neural crest stem cells(Sieber-Blum et al., 2006), and bone marrow stromal cells(BMSCs) (Wu et al., 2002; Hofstetter et al., 2002; Ohta et al.,2004; Himes et al., 2006; Lu et al., 2007).
Our previous study showed that BMSCs exhibited distincteffects on the recovery of injured spinal cords when they wereinfused into the cerebrospinal fluid (CSF) of rats with acutecrush injury of the spinal cord (Ohta et al., 2004; Yoshiharaet al., 2007). Locomotion in the rat was improved, and cavityformation in the spinal cord was reduced. Based on theseresults, followed by validation of the safety employing rhesusmonkeys, we applied BMSCs clinically to a patient with acutespinal cord injury under a strict protocol in March 2006 (Saitoet al., 2008).
Many cell transplantation studies of SCI, including ourprevious studies, have primarily focused on acute SCI (Coyneet al., 2006; Hofstetter et al., 2002; Lu et al., 2005; Swanger et al.,2005). However, considering that all patients with SCI atpresent are in sub-acute or chronic stages, the treatment ofSCI through delayed cell transplantation is an importantstrategy for translational studies on SCI (Neuhuber et al., 2005;Himes et al., 2006; Lu et al., 2007; Zurita et al., 2008; NandoeTewarie et al., 2009). Considering that the affected tissuesmight have been lost by degeneration, resulting in cavityformation in sub-acute and chronic SCI, BMSCs were directlyengrafted into the lesion of SCI in the present study.
The present study demonstrated that BMSC transplanta-tion produced extensive outgrowth of regenerating axons
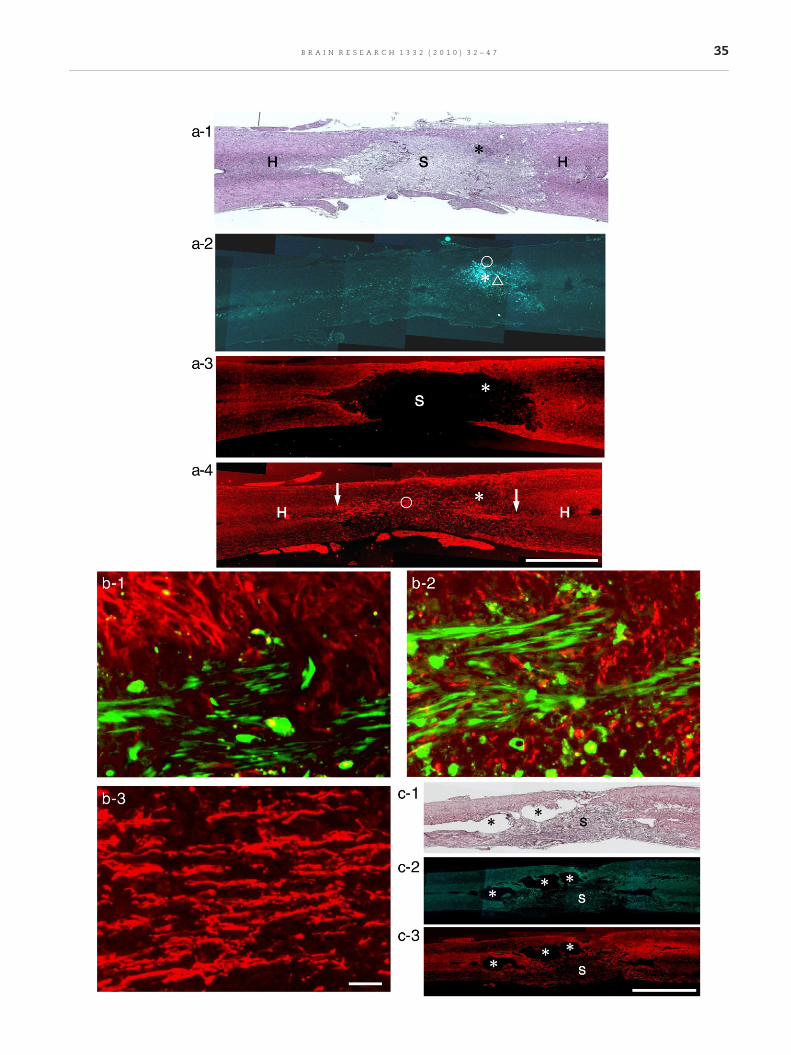
Fig. 2 – One week after transplantation. Horizontal sections. Left tregenerating axons extend longitudinally through the astrocyte-(S) apparently different from spinal cord parenchyma (H). No distiBMSCs shown in a-2. a-2. The section adjacent to a-1. Engraftedborder of the lesion shown in a-1. a-3. GFAP immunohistochemistarea (S) extends rostro-caudally. Engrafted BMSCs (site indicateda-4. Immunohistochemistry for neurofilaments in the section adextend in bundles along the whole length of the astrocyte-devoihost nerve networks in the transitional zones on the rostral as wengrafted BMSCs shown in a-2. H: spinal cord parenchyma. Scalmarked with a circle in a-2, showing the relationship between eimmunohistochemistry). Most BMSCs extend longitudinally, andMagnification of the part marked with a triangle in a-2, showing(red, neurofilament immunohistochemistry). Regenerating axonMagnification of regenerating axons marked with a circle in a-4.area. Scale: 50 μm (b-1, 2, and 3). c-1. Control (vehicle injection).and a tissue area (S) different from the host spinal cord tissue inAstrocytes are found around cavities (asterisks). c-3. The sectionremain around cavities (asterisks); however, few axons are foun
associated with Schwann cells through the extracellularmatrices consisting of collagen fibrils in the sub-acute spinalcord injury. The cavity formation was reduced, and distinctlocomotory improvement occurred.
2. Results
2.1. Prior to BMSC transplantation
Two weeks after crushing, the affected spinal cord tissueswere lost, resulting in cavity formation inmost cases (Fig. 1a).Astrocytes surrounded the cavity, but with no apparentastrocyte scar (Fig. 1a-1). Some axons (Fig. 1a-2) andoligodendrocytes (Fig. 1a-3) were located on the inner cavitywall.
2.2. One week after cell transplantation
Implanted GFP-transgenic BMSCs were identified as greenfluorescent cell assemblies of various sizes in the spinal cord1 week after transplantation (Figs. 1b, 2a-2). GFP-labeled BMSCassemblies were partly in contact with astrocytes (Figs. 1b-1,2b-1), however, in most cases they were surrounded by largespaces displaying no astrocytic immunoreactivity as observedin coronal (Fig. 1b-1) and horizontal (Fig. 2a-3) sections of thespinal cord. Astrocyte-devoid areas were usually located onthe dorsal side of GFP-labeled BMSC assemblies. Althoughapparently vacant by immunohistochemistry, these astro-cyte-devoid areas in fact contained densely packed tissues(Fig. 2a-1). Immunohistochemistry demonstrated numerousaxons around engrafted BMSCs in astrocyte-devoid areas(Figs. 1b-2, c). Almost no oligodendrocytes were found in theastrocyte-devoid area (Fig. 1b-3). In coronal sections, mostaxons appeared cut transversely (Figs. 1b-2, c), while inhorizontal sections they were cut longitudinally (Figs. 2a-4,b-3). It was remarkable that numerous axons extendedlongitudinally (rostro-caudal direction) along the length of
o right: rostro-caudal direction. Thesemicrographs show thatdevoid area. a-1. HE staining. The lesion is filled with tissuesnct cavity is formed. An asterisk indicates the site of engraftedBMSCs (green) are located as cell assemblies (asterisk) at thery (red) in the section adjacent to a-2. A large astrocyte-devoidby asterisk) are located at the border of astrocyte-devoid area.jacent to a-3. It is remarkable that numerous axons (red)d area shown in a-3. These axons appear in continuity withell as caudal side (arrows). Asterisk indicates the site ofe: 2 mm (a-1, 2, 3 and 4). b-1. Magnification of the partngrafted BMSCs (green) and astrocytes (red, GFAPare only partly in association with astrocytes. b-2.
the relationship between engrafted BMSCs (green) and axonss are in intimate association with engrafted BMSCs. b-3.Axons extend longitudinally through the astrocyte-devoidHorizontal section. HE staining. There are cavities (asterisks)the lesion. c-2. The section adjacent to c-1. GFAP staining.adjacent to c-2. Neurofilament staining. Some axons (red)d in the lesion (S). Scale: 2 mm (c-1, 2 and 3).
35B R A I N R E S E A R C H 1 3 3 2 ( 2 0 1 0 ) 3 2 – 4 7

36 B R A I N R E S E A R C H 1 3 3 2 ( 2 0 1 0 ) 3 2 – 4 7
the astrocyte-devoid area (Fig. 2a-4). Axons were in closeassociation with engrafted BMSCs (Fig. 2b-2). Axons within theastrocyte-devoid area were reasonably regarded as regenerat-ing, and appeared in continuity with the neural networks ofthe host spinal cord (Fig. 2a-4). BMSCs appeared to be arrangedalmost longitudinally in cell assemblies (Figs. 2b-1, b-2), but
Fig. 3 – Immunoelectron microscopy of engrafted BMSCs, and histtransplantation. a. Epon section from the same series of spinal corimmunostained with anti-GFP antibody, and visualized by HRP-DAidentified as dark-stained cell assemblies in the center (long arrow)clear thatdense tissue (asterisk)occupies theastrocyte-devoidareab. Electronmicrograph taken from Epon section as shown in “a”. Echaracteristic of containing numerous mitochondria, within engravessel (V) is seennearBMSCs.Somecells (asterisks) arenot immunin this micrograph. Scale: 5 μm. c. Enlargement of engrafted BMSCmatrices (C) are deposited around BMSCs (M). Scale: 2 μm. d. ThisMany blood vessels (some are labeled with arrows) of various diam
exhibited no tendency to migrate into the host spinal cordtissue. In the vehicle-injected control, cavities were formedand only a few axons were found in the lesion (Fig. 2c).
Immuno-electron microscopy showed that DAB-stainedBMSCs were clearly identified as dark, elongated cells in theextracellular matrices containing collagen fibrils (Fig. 3b, c).
ology of blood vessels in the lesion. One week afterd sections as shown in Fig. 1b. GFP-labeled BMSCs wereB staining for electron microscopy. Engrafted BMSCs areaswell as in the periphery (short arrows) of the spinal cord. It isdorsal to theBMSCassembliesas seen inFig. 1b.Scale: 200 μm.ngrafted BMSCs (B) are stained dark. There is a growth cone (G)fted BMSC assemblies. What appears to be a developing bloodostained forGFP, the cell lineageofwhichcannotbedetermineds and extracellular matrices. Collagen-containing extracellularsection was obtained from the same specimen as in Fig. 2a-1.eters are found in the lesion. HE staining. Scale: 20μm.

37B R A I N R E S E A R C H 1 3 3 2 ( 2 0 1 0 ) 3 2 – 4 7
Some growing axons were found in association with BMSCs(Fig. 3b). Blood vessels were occasionally found in theseextracellular matrices (Fig. 3b), suggesting that not onlynerve regeneration but also blood vessel formation might bepromoted in these areas. Hematoxylin-eosin (HE)-stainedsections showed thatmany blood vessels of various diameterswere found in the lesion around BMSCs (Fig. 3d).
For conventional electron microscopy, engrafted BMSCswere identified as cell assemblies different from host spinal
Fig. 4 – Conventional electronmicroscopy. Oneweek after transplas engrafted BMSCs is located in the white matter of the spinal cmyelin sheaths is marked by a dotted line. The dorsal part (doubmatter, corresponding to the astrocyte-devoid area seen in Fig. 1dorsal to the cell assembly in “a”. There are numerous unmyelinprocesses (arrows) or a cell body (S), suggesting that these axon-unmyelinated immature axons correspond to the abundant axonand 2a-4 and c-2. Scale: 2 μm. c-1. Magnification of an unmyelinacell processes (S) with basal laminae (arrows) on their surface. ThSchwann cell processes. Collagen fibrils (Co) are seen around thean developing Schwann cell (S) covered by basal lamina (arrows
cord tissue in Epon section (Fig. 4a). Corresponding to theabundant axons in astrocyte-devoid areas shown by immu-nohistochemistry in Figs. 1 and 2, numerous unmyelinatedimmature axons were identified near the presumed BMSCassembly by electron microscopy (Fig. 4b). These axonswere surrounded, singly or in bundles, by cell processes(Figs. 4b, c-1) or cell bodies (Figs. 4b, c-2) covered by basallaminae on the surface. Extracellular matrices containingcollagen fibrils were deposited in the space associated with
antation. a. Epon section. A cell assembly (asterisk) presumedord. The border of host white matter containing degeneratedle asterisks) of the cell assembly is lacking in the host whiteb. Scale: 50 μm. b. Electron micrograph taken from the areaated immature axons (some are labeled “A”) covered by cellassociated cells might be developing Schwann cells. Theses demonstrated by immunohistochemistry in Figs. 1b-2 and c,ted immature axon bundle. Two axons (A) are surrounded byis feature indicates growing axons surrounded by developingnerve bundle. Scale: 500 nm. c-2. Thismicrograph shows that) surrounds unmyelinated immature axons (A). Scale: 1 μm.

38 B R A I N R E S E A R C H 1 3 3 2 ( 2 0 1 0 ) 3 2 – 4 7
basal laminae. These features were similar to those seen inperipheral nerve regeneration. Cell processes and cell bodiessurrounding axons were regarded as developing Schwanncells. Immunohistochemistry showed that Schwann cellswere closely associated with axons (see Supplement Fig. 1a).
2.3. Two and 8 weeks after cell transplantation
Immunohistochemical findings were almost the same 2-8 weeks after cell transplantation. The astrocyte-devoid areaas noted in the previous stage was also found in horizontal(Fig. 5a-2) and coronal (Fig. 5c-1) sections of spinal cord inthese stages. BMSCs were no longer identified beyond 2 weeksafter transplantation. There is no tendency suggesting thatastrocyte-devoid areas were reduced in size by invasion ofastrocytes or their cell processes. It is remarkable thatnumerous axons were found in the astrocyte-devoid areaalso in these stages. Regenerating axons filled the whole spaceof the astrocyte-devoid area (Figs. 5a-3, c-2). In coronalsections, axons were cut transversely (Figs. 5c-2, d), and inhorizontal sections cut longitudinally (Figs. 5a-3, b), indicatingthat axons extended in a rostro-caudal direction through theastrocyte-devoid area. As in the previous stages, theseregenerating axons appeared to be in continuity with neuralnetworks of the host spinal cord (Fig. 5a-3). These regeneratingaxons were grouped into small bundles consisting of a fewindividual axons (Fig. 5d). In the control, various cavities werefound, and only a small number of axons were noted in thelesion of the spinal cord (Fig. 5e).
Immunohistochemistry for Schwann cells showed thatregenerating axons were closely associated with Schwanncells, indicating that each nerve fiber was composed of axonsand Schwann cells (Supplement Fig. 1a) in the lesion. Thesimilar close association of regenerating axons and Schwanncells was found at 1-week, and 4- and 8-weeks post-
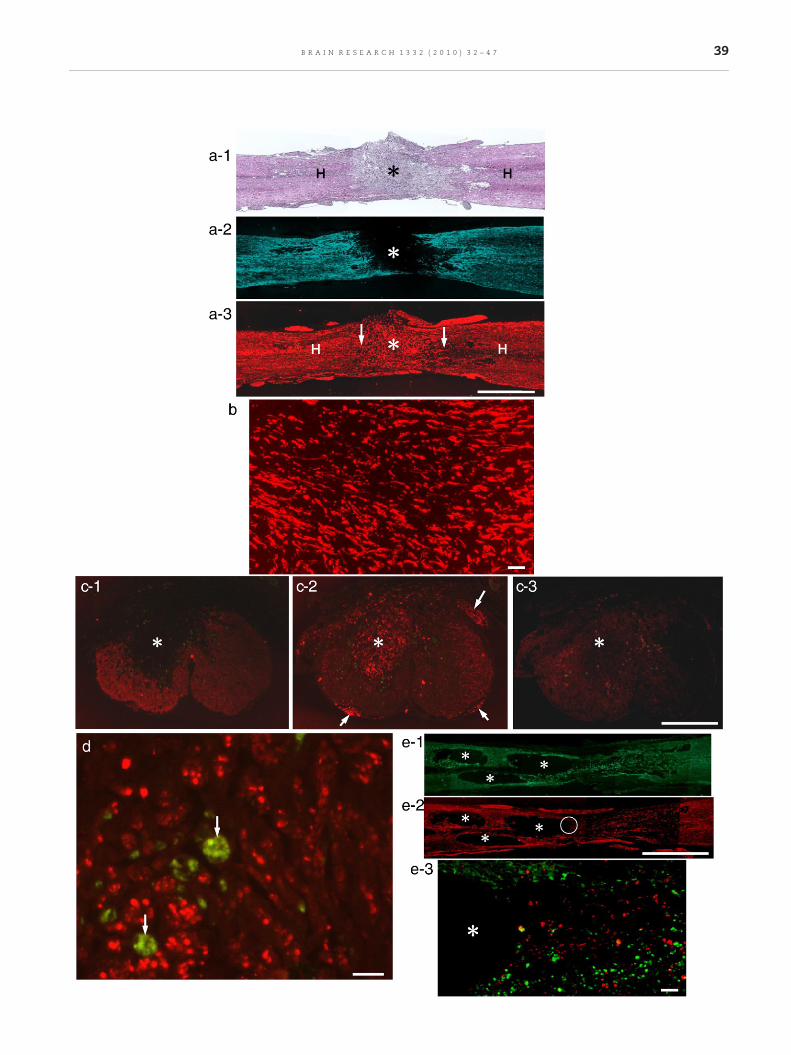
Fig. 5 – Two to 8 weeks after BMSC transplantation or vehicle injimmunostaining. a-1. Two weeks after transplantation. Horizon(asterisk) different from the host spinal cord tissue (H). a-2. Immu(the red color of astrocyteswas converted to blue for clear referencof the tissue different from the host spinal cord parenchyma shoa-3.Immunohistochemistry for neurofilaments (red) in the sectionabundant regenerating axons extended longitudinally, filling theaxons are in continuity with neural networks of the host spinal2 mm (a-1, 2 and 3). b. Magnification of axon bundles at the asteScale: 50 μm. c-1. Eight weeks after transplantation. GFAP immudorsal part (asterisk) of the spinal cord is devoid of GFAP immunpost-transplantation. c-2. Immunohistochemistry for neurofilamremarkable that numerous axons fill the astrocyte-devoid area (aFig. 1b-2. The dorsal (long arrow) and ventral roots (short arrowssection adjacent to c-2. The spinal cord lesion is only faintly staiastrocyte-devoid area (asterisk). Scale: 500 μm (c-1, 2 and 3). d. Maxons are cut transversely, indicating that they extend longitudiinto bundles of various sizes. Macrophages (arrows) showing no20 μm. e-1. Control. Eight weeks after vehicle injection. GFAP imthe lesion. e-2. The section adjacent to e-1. Neurofilament immuaround cavities (asterisks) in the lesion. Scale: 2 mm (c-1 and 2).corresponding to the site of a circle in e-2. Double-staining for Gamong astrocytes (green). Part of the cavity (asterisk) is seen on
transplantation. These findings were compatible with thoseobtained by electron microscopy at 1-week (Fig. 4), and 2-(Fig. 6a) and 8-weeks (Fig. 6b) post-transplantation.
Immunohistochemistry for specific types of axons con-taining calcitonin gene-related peptide (CGRP) (SupplementFig. 1b), serotonin (5HT) (Supplement Fig. 1c), and tyrosinehydroxylase (TH) (Supplement Fig. 1d) showed that thesetypes of axons were found among regenerating axons in thelesion. Many CGRP-immuno-positive axons (primary senso-ry fibers) grew throughout the lesion. Some 5HT-immuno-positive (raphespinal fibers) and TH-immuno-positive (coer-ulospinal fibers) axons extended from the rostral border intothe lesion. None or only a few 5HT- or TH-immuno-positiveaxons were found in the caudal side. These findingsindicated that ascending (primary sensory) and descending(raphespinal and coerulospinal) axons extended into thelesion.
Electron microscopy demonstrated the presence of axonbundles containing several unmyelinated axons in the extra-cellular matrices (Fig. 6a). These axons corresponded to thosenoted by immunohistochemistry in Fig. 5b. Axons appearedfurther developed in these stages from those 1-week post-transplantation (Fig. 4b): axons became thicker and separatedinto individual axons, and some were thinly myelinated bySchwann cells 2 weeks after transplantation (Fig. 6a). Eightweeks after transplantation, most axons were thickly myelin-ated by Schwann cells (Fig. 6b), and were grouped in bundles asnoted by immunohistochemistry in Fig. 5d. These featuresweresimilar to those observed in peripheral nerve regeneration.
2.4. Cavity formation
Cavity formation was assessed using HE sections. Cavityformation was markedly reduced in the BMSC-transplantedspinal cord; only very small cavities were identified in the
ection. Coronal and horizontal sections. HE staining andtal section. HE staining. The lesion is filled with dense tissuenohistochemistry for astrocytes in the section adjacent to a-1e). The astrocyte-devoid area (asterisk) corresponds to the sitewn in a-1. Engrafted BMSCs had disappeared at this stage.adjacent to a-2. Although engrafted BMSCs had disappeared,whole space of the astrocyte-devoid area (asterisk). These
cord (H) on the rostral as well as caudal side (arrows). Scale:risk in a-3. Numerous axons extend almost longitudinally.nohistochemistry. Coronal section. As seen in Fig. 1b, theoreactivity. Engrafted BMSCs disappeared from 2 weeksents in the section adjacent to c-1. Comparing with c-1, it issterisk) of the spinal cord. This feature is the same as that in) are identified. c-3. Immunohistochemistry for GST-π in thened. No distinct staining for oligodendrocytes is seen in theagnification of axons in a part (asterisk) of c-2. Numerous
nally through the astrocyte-devoid area. Axons are separatedn-specific fluorescence are scattered among axons. Scalemunohistochemistry. Large cavities (asterisks) are formed innohistochemistry. Few axons are found in remaining arease-3. Magnification of part of the section adjacent to e-2,FAP and neurofilaments. Only a few axons (red) are foundthe left. Scale: 50 μm.
39B R A I N R E S E A R C H 1 3 3 2 ( 2 0 1 0 ) 3 2 – 4 7
40 B R A I N R E S E A R C H 1 3 3 2 ( 2 0 1 0 ) 3 2 – 4 7
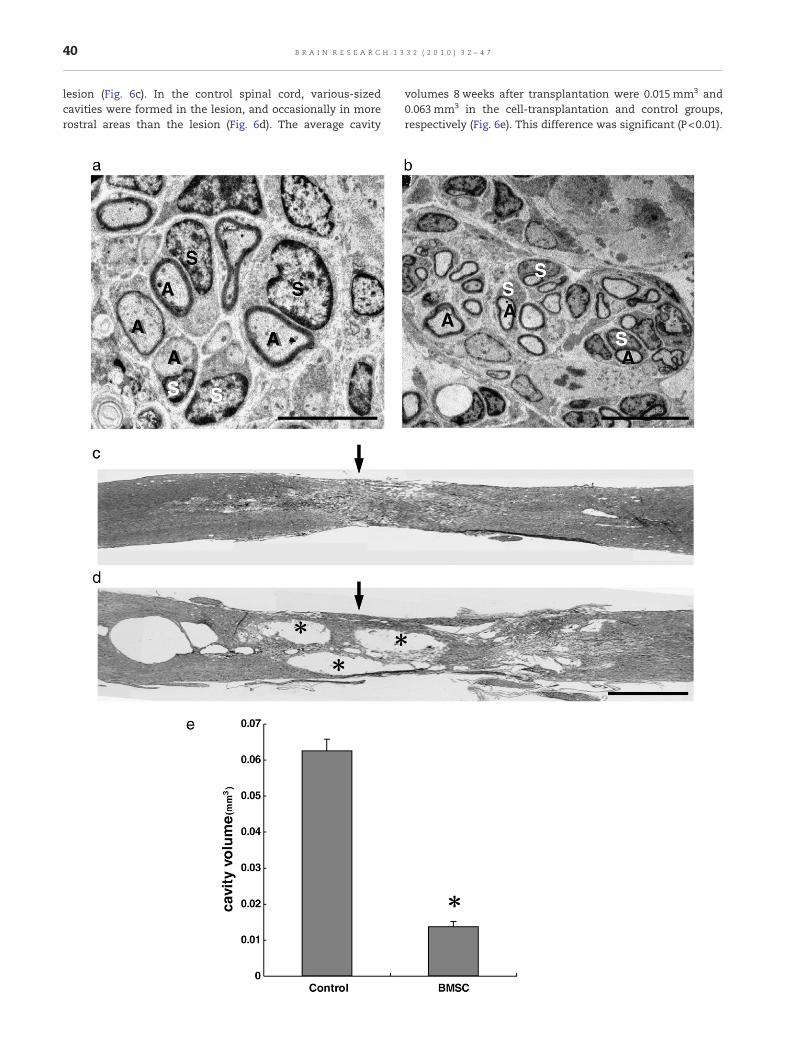
lesion (Fig. 6c). In the control spinal cord, various-sizedcavities were formed in the lesion, and occasionally in morerostral areas than the lesion (Fig. 6d). The average cavity
volumes 8 weeks after transplantation were 0.015 mm3 and0.063 mm3 in the cell-transplantation and control groups,respectively (Fig. 6e). This difference was significant (P<0.01).

Fig. 7 – Locomotory behavior. This graph shows BBB scores inrats of cell transplantation and control groups 10 weeks afterinjury (8 weeks after cell transplantation). There aresignificant differences (P<0.05) between cell transplantationand control groups 2, 4, and 8 weeks after BMSCtransplantation (asterisks).
41B R A I N R E S E A R C H 1 3 3 2 ( 2 0 1 0 ) 3 2 – 4 7
2.5. Locomotory behavior
Dropping a weight from a height of 7.5 cm yielded low BBBscores 2 weeks after crush injury, and no distinct improve-ment occurred thereafter in the control group.
BBB scores improved from 5.3±0.5 before transplantation to6.5±0.7, 7.7±0.7, 9.6±0.7, and 9.8±0.8 at 1, 2, 4, and 8 weeks aftertransplantation, respectively. On the other hand, rats in thevehicle-injection group showed BBB scores of 5.3±0.5, 5.3±1.0,5.6±1.2, 5.7±1.4, and 5.7±1.4, and those of the non-treatmentgroup showed 5.3±0.5, 5.3±0.5, 5.4±0.5, 5.5±0.04, and 5.5±0.6,respectively, at the above time points (Fig. 7). There weresignificant differences (P<0.05) between the cell transplantationand control groups 2, 4, and 8 weeks after transplantation.
3. Discussion
The present study demonstrated that BMSCs transplantedinto an SCI lesion 2 weeks post-injury produced tissuematrices primarily composed of collagen fibrils, into whichastrocytes were not allowed to migrate, resulting in theformation of astrocyte-devoid areas in the spinal cord. It is
Fig. 6 – Conventional electron microscopy of axons extending thcavity formation at 8 weeks after BMSC transplantation (c, d and eseparated from each other, and covered by Schwann cells (S). Somimmunohistochemistry in Figs. 5a-3 and b. Scale: 5 μm. b. EightIndividual axons (A) are thickly myelinated by Schwann cells (S)immunohistochemistry in Figs. 5c-2 and d. Scale: 10 μm. c. Horizlesion (arrow) is filledwith tissue as seen in Figs. 2a-1 and 5a-1. HEformed in the lesion (arrow). This HE section is from a series of Fig2 mm (c and d). e. This graph shows a comparison of the volumetransplantation groups. The difference is significant (asterisk, P<
remarkable that numerous axons grew out through suchastrocyte-devoid tissue matrices, as demonstrated by immu-nohistochemistry and electron microscopy. Schwann cellsmight be associated with growing axons and formed myelinsheaths, as in the case of peripheral nerve regeneration. Theseelectron microscopic findings are compatible with the resultsof other immunohistochemical studies that Schwann cellsand laminin are associated with growing axons in the BMSC-transplanted spinal cord (Lu et al., 2007). Although BMSCsdisappeared from the host tissue 1-2 weeks after transplanta-tion, cavity formation was markedly reduced, and locomotorybehavior improved to 9.8 versus 5.5-5.7 of the control 8 weeksafter transplantation.
3.1. Fate of BMSCs
In the present study, BMSCs disappeared between 1 and2 weeks after transplantation in SD rats. Our previous studyshowed that BMSCs infused into CSF survived for 2-3 weeksafter infusion in SD rats (Ohta et al., 2004). This study alsoshowed that administration of the standard dose immuno-suppressant FK506 did not change the survival time oftransplanted BMSCs. Our other studies employing the sameexperimental paradigm, however, showed that neural stemcells survived and proliferated for more than 3 weeks aftertransplantation in SD rats (Wu et al., 2002; Bai et al., 2003).It was reported that the disappearance of BMSCs 1-2 weeksafter transplantation might be mainly due to an inflam-matory reaction in the normal brain of SD rats (Coyne etal., 2006). On the other hand, BMSCs can survive longer butwith a gradual decrease in number in the spinal cord ofinbred Fisher 344 (Lu et al., 2005; Swanger et al., 2005) or SDrats treated with high-dose immunosuppressant (Swangeret al., 2005). It should be emphasized that BMSCs, evenwhen their survival time was limited, enhanced tissuematrix formation and axonal outgrowth in sub-acute SCI. Itis conceivable that the longer their survival, the strongertheir effects. Considering that BMSCs gradually decrease innumber with time in syngeneic (Lu et al., 2007), andimmunosuppressed allogeneic grafts (Swanger et al., 2005;Himes et al., 2006; Ohtaki et al., 2008), it appears thatBMSCs might not survive long enough to be integrated intothe spinal cord tissue.
The transplantation of BMSCs has beneficial effects eventhough BMSCs disappear from the spinal cord after transplan-tation. It might be conceivable that ensuing death of BMSCsfollowing transplantation might elicit some cellular reactionsthat lead to the tissue repair including axonal outgrowth inSCI. In this respect, living BMSCs might not be needed for
rough the lesion at 2 and 8 weeks (a and b), and reduction in). a. Twoweeks after transplantation. Individual axons (A) aree aremyelinated. These axons correspond to those shown byweeks after transplantation. There are many nerve fibers.. These axons correspond to those shown byontal section of the spinal cord of BMSC transplantation. Thestaining. d. Control. Vehicle injection. Cavities (asterisks) are
. 5e, and adjacent to the section in Fig. 5e-3. HE staining. Scale:of the spinal cord cavity between control and BMSC0.01).

42 B R A I N R E S E A R C H 1 3 3 2 ( 2 0 1 0 ) 3 2 – 4 7
transplantation. Nonviable BMSCs could be transplanted inSCI to test this hypothesis in the future.
Several studies report that BMSCs differentiate into neuralcells, including astrocytes and neurons, by labeling cells with5-bromo-2-deoxyuridine (BrdU) or bis benzamide (BBZ) (Choppet al., 2000; Ankeny et al., 2004; Zurita, et al., 2008); however,BrdU and BBZ tend to stain host cells by diffusion, leading tomisunderstanding of the results (Coyne et al., 2006). AlthoughBMSCs are known to exhibit the capacity to differentiate intomyocytes, Schwann cells, and neurons in vitro (Dezawa et al.,2005), it has been recognized that transplanted BMSCs do notdifferentiate into neural cells in the host brain (Coyne et al.,2006) or spinal cord (Castero et al., 2002; Li and Chopp, 2009;Sheth et al., 2008).
3.2. The effects of BMSCs
It was demonstrated that implanted BMSCs provided extra-cellular matrices including collagen fibrils, through whichnumerous regenerating axons associated with Schwann cellsgrew out longitudinally. This might probably contribute to thedistinct improvement of locomotory behavior as well as thereduction of cavity formation.
Although they survived for only a limited time, BMSCsexerted beneficial effects on tissue repair and locomotoryimprovement in the present sub-acute as well as previousacute SCI study (Ohta et al., 2004). It is reported that delayedBMSC transplantationhasnobeneficial effects on tissue sparingin sub-acute or chronic SCI (Nandoe Tewarie et al., 2009).However, the present study indicates that sub-acute BMSCtransplantation has more broad effects including tissue repair,axonal regenerationand locomotory improvement. BMSCshavebeen suggested to release diffusible neuroprotective factors(Chopp et al., 2000; Ankeny et al., 2004; Himes et al., 2006;Swanger et al., 2005; Shintani et al., 2007). In fact, it has beenshown that BMSCs release BDNF, VEGF, and cytokines such asIL-6 and stem cell factor (SCF) (Neuhuber et al., 2005), and IGF-1(Ohtaki et al., 2008). Neuroprotective properties of BMSCs arealso shown in the coculture (Wu et al., 2003; Crigler et al., 2006;Isele et al., 2007; Wright et al., 2007), and organotypic sliceculture (Kamei et al., 2007; Shichinohe et al., 2008). Exogenouslyapplied trophic factors, including BDNF, IGF-1, NT-3, and GDNF(Lu et al., 2007; Sharma, 2007), and HGF (Kitamura et al., 2007)have been shown to be effective for SCI. BMSCs might beregarded as a small molecular factory of neuroprotectivefunction (Li and Chopp, 2009).
Electron microscopy in the present study has demon-strated that BMSCs induce tissue matrices composedprimarily of collagen fibrils, which probably act as aneffective scaffold for axonal outgrowth. It can be said thatBMSCs have dual functions: the release of trophic factorsand induction of tissue matrix formation for neuroprotec-tion and tissue repair scaffolds, respectively. Immunohis-tochemistry has shown that Schwann cells are closelyassociated with axons: regenerating nerve fibers composedof axons and Schwann cells grow in the spinal cord lesion.Electron microscopy has demonstrated that such nerveregeneration occurs in collagen fibril-containing extracellu-lar matrices. This feature is the same as seen in peripheralnerve regeneration (Ide et al., 1983). Schwann cell involve-
ment in CNS nerve regeneration has been suggested also byimmunohistochemistry in many studies (Lu et al., 2007;Ankeny et al., 2004; Himes et al., 2006; Hofstetter et al.,2002). The immunohistochemical data shown by Lu et al.(2007) might correspond to cellular components in therepaired tissue demonstrated by the present study: NG2might be related to proteoglycans of extracellular matrices,L1 might be expressed at the contact of axons withSchwann cells, and laminin might be derived from thebasal laminae on the surface of Schwann cells associatedwith axons.
Sources of Schwann cells were not specified in the presentstudy.
The present study has shown that regenerating axonsappear in continuity with neural networks on the rostral aswell as caudal side of the host spinal cord. The outgrowth of5HT-, TH- and CGRP-immunostained axons into the lesionsupports this hypothesis: descending fibers includingraphespinal and coerulospinal axons, and ascending fibersgrow out from the rostral and caudal borders, respectively,into the lesion (Lu et al., 2005). The connection of regeneratingaxons including these ascending and descending ones withhost neural networks is unclear in the present study. However,the apparent continuity of regenerating axons with hostneural components at the rostral and caudal sides as shownin Figs. 2a-4 and 5a-3, suggests that some regenerating axonshave extended further into the other side. It is probable thatsuch regenerating axons might have at least partly contribut-ed to locomotory improvement in the present study. Theneural connection of regenerating axons with host networkswill be an important research subject in the future. It seemsthat astrocytes do not contribute directly to axonal outgrowththrough the lesion in the present study. Oligodendrocyteshave no relation with regenerating axons in astrocyte-devoidareas.
The finding that blood vessels were frequently foundaround BMSCs indicates that BMSCs might also facilitateblood vessel formation in SCI (Yoshihara et al., 2007). Bloodvessel formation is critical for nutritional supply to thelesion.
Improvements in locomotion were distinct in the BMSCtransplantation group.
The volume of cell solution or PBS injected into the spinalcord is relatively large compared with that of the cavitiesformed in the lesion. The possibility that a large volume ofsolution might cause some damage to the remaining hostintact tissues should be considered. As far as we examined,there was no transient drawback or deteriorating change inlocomotory behavior that could result from the tissue damagefollowing the injection of cell solution or PBS. It can be said thatthe injection of 30 μl of solution has no serious adverseinfluence on the spinal cord.
3.3. Acute vs. sub-acute or chronic SCI
The present study examined delayed cell transplantation forSCI in rats. There are no definite criteria to categorize sub-acute or chronic SCI in the rat. Considering that thedegeneration of crush-injured spinal cord tissues advancedup to cavity formation 2 weeks after crush injury, and that

43B R A I N R E S E A R C H 1 3 3 2 ( 2 0 1 0 ) 3 2 – 4 7
almost no significant changes in the BBB score were notedthereafter, the spinal cord 2 weeks after injury might beregarded as a fairly advanced stage of SCI. It would beexperimentally realistic to consider that this condition isalmost stable from histological as well as functional points ofview. Therefore, the SCI 2 weeks post-crush might beregarded as sub-acute or early chronic condition of SCI.Houle and Tessler (2003) stated that 2-week post-injury issub-chronic, while 4-week post-injury is chronic, and thatacute and sub-chronic tissues might respond to the trophicfactor, but chronic ones might not. It is conceivable that glialcells and neurons might be critical conditions for survival inacute SCI, whereas they might have already degenerated andbeen lost in sub-acute and chronic SCI. Unlike the previousacute SCI study, in which BMSCs were infused through CSF,BMSCs were implanted directly into the lesion in the presentstudy as in other delayed transplantation studies (Chopp etal., 2000; Neuhuber et al., 2005; Himes et al., 2006; Lu et al.,2007; Zurita et al., 2008). Direct injection of BMSCs into thelesion might be an effective method of transplantation insub-acute and chronic SCI; however, considering that infu-sion of BMSCs through CSF by lumbar puncture is the mostpreferable method clinically, the effect of infusing BMSCsthrough CSF in an sub-acute and chronic SCI should beexamined.
3.4. Clinical application of BMSCs
Researchers involved in “regeneration studies” should striveto clinically apply their achievements. BMSCs are character-istic in that they are autologous and readily obtainable fromthe patient's bone marrow. The fact that BMSCs are safe andhave a marked ability to promote the repair of SCI tissue,including axonal regeneration, means that they are promisingcells for translational therapies. Based on our studies andconfirmation of safety in monkeys, a patient with acute SCIwas treated for the first time with BMSC transplantation bylumbar puncture in 2006 (Saito et al., 2008). Clinical application
Table 1 – This table shows numbers of rats used in the present
of BMSC transplantation was discussed (Nandoe Tewarieet al., 2006), and treatments for patients with chronic SCIwere also reported using different techniques of cell applica-tion (Syková et al., 2006; Yoon et al., 2007; Wright et al., 2008;Pal et al., 2009).
4. Experimental procedure
4.1. Surgery and crush injury
A hundred and eighteen Sprague-Dawley (SD) rats (4-6 weeksold, female) were used in the present study (Table 1), whichwas performed with approval from the Animal Committee ofAino University, and all animal experiments were performedin accordance with the Guidelines for Animal Experiments ofAino University that were laid down in compliance with theJapanese Regulations for Animal Welfare. All efforts weremade to minimize the number of animals and their sufferingthroughout the experiments. Rats were anesthetized byintraperitoneal injection (0.3 ml) of a mixture of Nembutal(pentobarbiturate sodium) and atropine (1:1). Laminectomywas performed at the Th8-9 vertebrate, and the spinal cordwith the dura mater was exposed. The spinal cord covered bydura mater was crush-injured by spontaneously dropping a10 g metal rod from a height of 7.5 cm using an NYU impactor.Crush injury has usually been made by dropping a weightfrom 2.5-5.0 cm (Chopp et al., 2000; Himes et al., 2006);however, our experiences showed that thesemild ormoderateinjuries tended to produce high spontaneous locomotoryrecovery in control rats. Therefore, in the present study aweight was dropped from 7.5 cm, in which no distinctimprovement was noted beyond 2 weeks post-injury in thecontrol. The spinal cord lesion was covered with a Goatexsheet, and the muscle and skin were sutured layer by layer.Goatex sheets enabled access to the injury site of the spinalcord in the second surgery for cell transplantation. Aftersurgery, 5-10 ml physiological saline was injected subcutane-
study.

44 B R A I N R E S E A R C H 1 3 3 2 ( 2 0 1 0 ) 3 2 – 4 7
ously to prevent dehydration of the rat. An antibiotic (gentacin3 mg/kg; Schering-Plough, Osaka Japan) was administeredsubcutaneously for 3 days after surgery.
4.2. Cell culture
Bonemarrow cells were obtained from GFP-transgenic SD rats(4 weeks old) by perfusion through femurs and tibias withculture medium, and cultured in Dulbecco's modified Eagle'smedium (DMEM) with 10% fetal calf serum (FCS). Within 5-7 days of culture, BMSCs proliferated to a density of ca. 107 inone dish. BMSCs were long, rectangular cells adhering to thebase of the culture flasks, exhibiting “green” fluorescence anda positive reaction for CD90 and CD29, and a negative reactionfor CD34 and CD11b on immunohistochemistry. Cells werelifted by incubation in 0.05% trypsin solution for 3 min at 37 °C.After three washings, cells were used for transplantation.
4.3. Cell transplantation
Cell transplantationwas performed 2 weeks after crush injury.For cell transplantation, 48 rats that had been crush-injured2 weeks previously were again anesthetized, and the spinalcord lesion was exposed by removing the Goatex sheet.Cultured cells were suspended at a density of 5×105 in 30 μlphosphate-buffered saline (PBS), and injected using an insulinsyringe with a 29 gauge needle into the spinal cord at threepoints: epicenter, and rostral and caudal borders of the lesion(10 μl each). The insulin syringe was attached to the stereo-tactic device (model SR-6N, Narishige). The needle wasinserted vertically to a depth of 1.5 mm (ca. 1 mmat the needlepore) from the spinal cord surface. The injection time was3 min. The needle was retained for 3 min in the same positionafter injection (cell transplantation group). For the control, PBSwithout BMSCs was injected to 42 rats in the same manner asabove (vehicle injection group), or neither cells nor PBS wasinjected after the crush injury to 22 rats (non-treatment group).
4.4. Locomotory assessment
The locomotory capacity of the hind limbs was assessedemploying theBBBscale inanopenarea (ca. 1.0 m2) byobservingrats for 5 min (Basso et al., 1995). Ten rats from each groupwereassessed at each time point (2 weeks post-injury, and 1, 2, 4, and8 weeks after cell transplantation or vehicle injection). Locomo-tion was recorded by digital video camera at each time point asabove, and analyzed by 2 researcherswhowere not aware of theexperiment.
4.5. Preparations for histology and immunohistochemistry
Rats were deeply anesthetized by an intraperitoneal injec-tion of Nembutal (100 mg/kg), and perfused intracardiallywith 50 ml PBS, followed by 200 ml of a fixative containing2% paraformaldehyde in 0.1 M phosphate buffer, pH7.4.Specimens were processed in a standard procedure forembedding in OCT compound, and cut coronally or horizo-natally into 7-μm-thick frozen sections with a cryostat(CM1510S; Leica). Frozen sections were mounted on siran-coated glass slides.
For histology, horizontal sections were stained withhematoxylin and eosin (HE), and used for observing bloodvessels, and measuring the volume of cavities in the spinalcord. For immunohistochemistry, after washing three timeswith PBS, and blocking, with a 0.1% bovine serum albuminsolution containing 0.1% Tween 20 in PBS for 30 min, sectionswere incubated overnight with a solution containing primaryantibodies as follows: anti-glial fibrillary acidic protein (GFAP)monoclonal antibody (1:300; Sigma) for astrocytes, anti-neurofilament 200 kD rabbit polyclonal antibody (1:100;Chemicon) for axons, anti-GST-πrabbit polyclonal antibody(1:300; MBL) for oligodendrocytes, anti-Schwann cell mousemonoclonal antibody (1:1000; Cosmo Bio) for Schwann cells,anti-5HT rabbit polyclonal antibody (1:2000; Sigma) forraphespinal fibers, anti-TH mouse monoclonal antibody(1:150; Chemicon) for coerulospinal fibers, and anti-CGRPrabbit polyclonal antibody (1:2000; Sigma) for primary sensoryfibers. After washing, the sections were incubated overnightwith secondary antibodies as follows: FITC- or Cy-3-labeledanti-mouse IgG antibody (1:1000; Amersham Biosciencea) forastrocytes, and coerulospinal fibers, or Cy-3-labeled anti-rabbit IgG antibody (1:1000; Amersham Biosciences) foroligodendrocytes, axons, raphespinal fibers, and primarysensory fibers. Sections were mounted and examined by afluorescence microscope (Axio Imager MI; Carl Zeiss).
For immunohistochemical examination, 5 rats each fromthe cell transplantation and vehicle injection groups weresacrificed at each time point (1, 2, 4, and 8 weeks after celltransplantation or vehicle injection). For the non-treatmentcontrol, 2 rats at 2 weeks post-injury (pre-transplantation),and one each at 3, 4, 6 and 10 weeks post-injury (1, 2, 4, and8 weeks, respectively, in Table 1) were used.
4.6. Immuno-electron microscopy
Three rats each at 1 and 2 weeks post-transplantation wereused for immuno-electronmicroscopy. Cryostat sections wereincubated in 0.5% H2O2 for 20 min at room temperature. Afterblocking and permeation with 0.01% saponin in PBS, thesections were then incubated with rabbit anti-GFP antibodyovernight at 4 °C. After washing, sections were incubated withan Fab’ fraction of goat anti-rabbit IgG conjugated withhorseradish peroxidase (1:1,000) overnight at 4 °C, and post-fixed with 1% glutaraldehyde for 10 min at room temperature.Subsequently, the sections were reacted with a solutioncontaining 0.04% 3,3-diaminobenzidine (DAB) and 0.005%H2O2 for 5 min to visualize the HRP, osmicated with 1%osmium tetroxide in 0.1 M PB for 30 min at room temperature,dehydrated by ethanol, and embedded in Epon 812 on glassslides. The embedded sections were removed from the glassslides by heating, and mounted on metal stubs. Semi-thinEpon sections were prepared for light microscopy. Thinsections were cut and, without metal staining, observed withan electron microscope (H-7650; Hitachi) at 80 kV.
4.7. Conventional electron microscopy
Three rats each from cell transplantation and vehicleinjection groups at each time point (1, 2, 4 and 8 weeksafter cell transplantation or vehicle injection) were used. For

45B R A I N R E S E A R C H 1 3 3 2 ( 2 0 1 0 ) 3 2 – 4 7
the non-treatment control, 2 rats at 2 weeks post-injury (pre-transplantation), and one rat each at 3, 4, 6 and 10 weekspost-injury (1, 2, 4, and 8 weeks, respectively, in Table 1) wereused. Rats were transcardially perfused with 50 ml PBS,followed by a fixative containing 1% glutaraldehyde and 1%paraformaldehyde in 0.1 M phosphate buffer, pH 7.4. Thespinal cord including the injured site was transversely cutinto several slices. These tissue slices were then post-fixedin 1% osmium tetroxide in 0.1 M PB, dehydrated, andembedded in Epon 812. Thin sections were stained withuranyl acetate and lead citrate, and observed by electronmicroscopy. Semi-thin Epon sections were stained withtoluidine blue for observation by light microcopy.
4.8. Cavity volume measurement
Whole spinal cord segments were cut horizontally into 7-μmthick frozen sections on cryostat, and stained with HE. Theareas of cavities were measured in every 5 sections (35-μminterval) using the Image Filing System (Flovel; Tokyo). Thecavity volume was calculated by multiplying the average areaof cavities by the total depth of the sections examined.
4.9. Statistics
The BBB scores were analyzed by repeated measures ANOVA,and post-hoc analysis was performed by Tukey's test to assessthe significance of differences at each time point (P<0.05).
For cavity formation, the mean values were analyzed usingStudent's t-test for difference between cell transplantationand control groups.
Acknowledgments
The authors would like to sincerely thank Dr. Akio Koyama,Representative Director of the Aino Gakuin, for establishingthe Institute of Regeneration and Rehabilitation.
This study was supported in part by grants from theJapanese Ministry of Education, Culture, Sports, Science andTechnology (No. 17300110 and 20300122) to C.I.
Appendix A. Supplementary data
Supplementary data associated with this article can be found,in the online version, at doi:10.1016/j.brainres.2010.03.043.
R E F E R E N C E S
Ankeny, D.P., McTigue, D.M., Takeman, L.B., 2004. Bone marrowtransplants provide tissue protection and directional guidancefor axons after contusive spinal cord injury in rats. Exp. Neurol.190, 17–31.
Bai, H., Suzuki, Y., Noda, T., Wu, S., Kataoka, K., Kitada, M., Ohta,M., Chou, H., Ide, C., 2003. Dissemination and proliferationof neural stem cells on the spinal cord by injection into thefourth ventricle of the rat. J. Neurosci. Methods 124,181–187.
Basso, D.M., Beattie, M.S., Bresnahan, J.C., 1995. A sensitive andreliable locomotor rating scale for open field testing in rats.J. Neurotrauma 12, 1–21.
Castero, R., Jackson, K.A., Goodell, M.A., Robertson, C.S., Liu, H.,Shine, H.D., 2002. Failure of bone marrow cells totransdifferentiate into neural cells in vivo. Science 297, 1299.
Chopp, M., Zhang, X.H., Li, Y., Wang, L., Chen, J., Lu, D., Rosenblum,M., 2000. Spinal cord injury in rat: treatment with bonemarrowstromal cell transplantation. Neuroreport 11, 3001–3005.
Coyne, T.M., Marcus, A.J., Woodbury, D., Black, I.B., 2006. Marrowstromal cells transplanted to the adult brain are rejected by aninflammatory response and transfer donor labels to hostneurons and glia. Stem Cells 24, 2483–2492.
Crigler, L., Robey, R.C., Asawachaicharn, A., Gaupp, D., Phinney,D.G.,2006. Human mesenchymal stem cell subpopulations express avariety ofneuro-regulatorymoleculesandpromoteneuronal cellsurvival and neuritogenesis. Exp. Neurol. 198, 54–64.
Dezawa, M., Hoshino, M., Nabeshima, Y., Ide, C., 2005. Marrowstromal cells: implications in health and disease in the nervoussystem. Curr. Mol. Med. 5, 723–732.
Enzmann, G.U., Lenton, R.L., Talbott, J.F., Cao, Q., Whittemore, S.R.,2006. Functional considerations of stem cell transplantationtherapy for spinal cord repair. J. Neurotrauma 23, 479–485.
Golden,K.L., Pearse,D.D., Blits, B., Garg,M.S., Oudega,M.,Wood,P.M.,Bunge, M.B., 2007. Transduced Schwann cells promote axonsgrowthandmyelinationafter spinal cord injury. Exp.Neurol. 207,203–217.
Himes, B.T., Neuhuber, B., Coleman, C., Kushner, R., Swanger, S.A.,Kopen, G.C., Wagner, J., Shumsky, J.S., Fischer, I., 2006.Recovery of function following grafting of human bonemarrow-derived stromal cells into the inured spinal cord.Neurorehabil. Neural Repair 20, 278–296.
Hofstetter, C.P., Schwarz, E.J., Hess, D., Widenfalk, J., El Manira, A.,Prockop, D.J., Olspn, L., 2002. Marrow stromal cells form guidingstrands in the injured spinal cord and promote recovery. Proc.Natl. Acad. Sci. U. S. A. 99, 2199–2204.
Hofstetter, C.P., Holmsrom, N.A., Lilja, J.A., Schweinhardt, P., Hao,J., Spenger, C., Wiesenfeld-Hallin, Z., Kurpad, S.N., Frisen, J.,Olson, L., 2005. Allodynia limits usefulness of intraspinalneural stem cell grafts; directed differentiation improvesoutcome. Nat. Neurosci. 8, 346–355.
Houle, J.D., Tessler, A., 2003. Repair of chronic spinal cord injury.Exp. Neurol. 182, 247–260.
Ide, C., Tohyama, K., Yokota, R., Nitatori, T., Onodera, S., 1983.Schwann cell basal lamina and nerve regeneration. Brain Res.288, 61–75.
Ide, C., Kitada, M., Chakrabortty, S., Taketomi, M., Matsumoto, N.,Kikukawa, S., Mizoguchi, A., Kawaguchi, S., Endoh, K., Suzuki,Y., 2001. Grafting of choroid plexus ependymal cells promotesthe growth of regenerating axons in the dorsal funiculus of ratspinal cord: a preliminary report. Exp. Neurol. 167, 242–251.
Isele, N.B., Lee, H.S., Landshanner, S., Straube, A., Padovan, C.S.,Plesnia, N., Culmsee, C., 2007. Bone marrow stromal cellsmediate protection through stimulation of PI3-K/Akt andMAPK signaling in neurons. Neurochem. Int. 2007 (50), 243–250.
Kamei, N., Tanaka, N., Oishi, Y., Ishikawa, M., Hamasaki, T.,Nishida, K., Nakanishi, K., Sakai, N., Ochi, M., 2007. Bonemarrow stromal cells promoting corticospinal axon growththrough the release of humoral factors in organotypiccocultures in neonatal rats. J. Neurosurg. Spine 6, 412–419.
Kitamura, K., Iwanami, A., Nakamura, M., Yamane, J., Watanabe,K., Suzuki, Y., Miyazawa, D., Shibata, S., Funakoshi, H.,Miyatake, S., Coffin, R.S., Nakamura, T., Toyama, Y., Okano, H.,2007. Hepatocyte growth factor promotes endogenous repairand functional recovery after spinal cord injury. J. Neurosci.Res. 85, 2332–2342.
Knoller, N., Auerbach, G., Fulga, V., Zelig, G., Attias, J., Bakimer, R.,Marder, J.B., Yoles, E., Belkin, M., Schwartz, M., Hadani, M., 2005.Clinical experience using incubated autologous macrophages

46 B R A I N R E S E A R C H 1 3 3 2 ( 2 0 1 0 ) 3 2 – 4 7
as a treatment for complete spinal cord injury: phase I studyresults. J. Neurosurg. Spine 3, 173–181.
Li, Y., Chopp, M., 2009. Marrow stromal cell transplantationin stroke and traumatic brain injury. Neurosci. Lett. 456,120–123.
Li, Y., Field, P.M., Raisman, G., 1997. Repair of adult ratcorticospinal tract by transplants of olfactory ensheathingcells. Science 277, 2000–2002.
Lu, P., Jones, L.L., Tuszynski, M.H., 2005. BDNF-expressing marrowstromal cells support extensive axonal growth at sites of spinalcord injury. Exp. Neurol. 191, 344–360.
Lu, P., Jones, L.L., Tuszynski, M.H., 2007. Axon regenerationthrough scars and into sites of chronic spinal cord injury. Exp.Neurol. 203, 8–21.
Matsumoto, N., Kitayama, H., Kitada, M., Kimura, K., Noda, M., Ide,C., 2003. Isolation of a set of genes expressed in the choroidplexus of the mouse using suppression subtractivehybridization. Neuroscience 117, 405–415.
McDonald, J.W., Liu, X.Z., Qu, Y., Liu, S., Mickey, S.K., Turetsky, D.,Gottlieb, D.I., Choi, D.W., 1999. Transplanted embryonic stemcells survive, differentiate and promote recovery in injured ratspinal cord. Nat. Med. 5, 1410–1412.
Nandoe Tewarie, R.D., Hurtado, A., Levi, A.D., Grotenhuis, J.A.,Oudega, M., 2006. Bone marrow stromal cells for repair of thespinal cord; towards clinical application. Cell Transplant. 15,560–577.
Nandoe Tewarie, R.D.S., Hurtado, A., Ritfeld, G.J., Rahien, S.,Werdell, D.F., Burroso, M.M.S., Grotenhuis, J.A., Oudega, M.,2009. Bone marrow stromal cells elicit tissue sparing afteracute but not delayed transplantation into the contused adultrat thoracic spinal cord. J. Neurotrauma 26, 1–10.
Neuhuber, B., Himes, B.T., Shumsky, J.S., Gallo, G., Fischer, I., 2005.Axonal growth and recovery of function supported by humanbone marrow stromal cells in the injured spinal cord exhibitdonor variation. Brain Res. 1035, 73–85.
Ohta, M., Suzuki, Y., Noda, T., Ejiri, Y., Dezawa, M., Katoka, K.,Chou, H., Ishikawa, N., Matsumoto, N., Iwashita, Y., Mizuta, E.,Kuno, S., Ide, C., 2004. Bone marrow stromal cells infused intothe cerebrospinal fluid promote functional recovery of theinjured rat spinal cord with reduced cavity formation. Exp.Neurol. 187, 266–278.
Ohtaki, H., Ylostalo, J.H., Foraker, J.E., Robinson, A.P., Reger, R.L.,Shioda, S., Prockop, D.J., 2008. Stem/progenitor cells from bonemarrow decrease neuronal death in global ischemia bymodulation of inflammatory/immune responses. Proc. Natl.Acad. Sci. U. S. A. 105, 14638–14643.
Pal, R., Venkataramana, N.K., Jan, M., Bansal, A., Balaraju, S., Joan,M., Chandra, R., Dixit, A., Ranthan, A., Murgod, U., Totey, S.,2009. Ex-vivo-expanded autologous bone marrow–derivedmesenchymal stromal cells in human spinal cord injury/paraplegia: a pilot clinical study. Cytotherapy 11, 897–911.
Plant, G.W., Christensen, C.L., Oudega, M., Bunge, M.S., 2003.Delayed transplantation of olfactory ensheathing gliapromotes sparing/regeneration of supraspinal axons in thecontused rat spinal cord. J. Neurotrauma 20, 1–16.
Ramon-Cueto, A., Cordeo, M.I., Santos-Benito, F.F., Avila, J., 2000.Functional recovery of paraplegic rats and motor axonregeneration in their spinal cords by olfactory ensheathingglia. Neuron 25, 425–435.
Rapalino, O., Lazarov-Spiegler, O., Agranov, E., Velan, G.J., Yoles, E.,Fraidakis, M., Solomon, A., Gepstein, R., Katz, A., Belkin, M.,Hadani, M., Schwartz, M., 1998. Implantation of stimulatedhomologous macrophages results in partial recovery ofparaplegic rats. Nat. Med. 4, 814–821.
Saito, F., Nakatani, T., Iwase, M., Maeda, Y., Murao, Y., Suzuki, Y.,Onodera, R., Fukushima, M., Ide, C., 2008. Spinal cord injurytreatmentwith intrathecal autologous bonemarrowstromal celltransplamtation: the first clinical trial case report. J. Trauma 64,53–59.
Sharma, H.S., 2007. A select combination of neurotrophinsenhances neuroprotection and functional recovery followingspinal cord injury. Ann. N. Y. Acad. Sci. 1122, 95–111.
Sheth, R.N., Manzano, G., Li, X., Levi, A.D., 2008. Transplantationof human bone marrow-derived stromal cells into thecontused spinal cord of nude rats. Neurosurg. Spine 8,153–162.
Shichinohe, H., Kuroda, S., Tsuji, S., Yamaguchi, S., Yano, S., Lee, J.B.,Kobayashi, H., Kikuchi, S., Hida, K., Iwasaki, Y., 2008. Bonemarrow stromal cells promote neurite extension in organotypicspinal lord slice: significance for cell transplantation therapy.Neurorehabil. Neural Repair 22, 447–457.
Shintani, A., Nakao, N., Kakishita, K., Itakura, T., 2007. Protectionof dopamine neurons by bone marrow stromal cells. Brain Res.1186, 48–55.
Sieber-Blum, M., Schnell, L., Grim, M., Hu, Y.E., Schneider, R.,Schwab, M.E., 2006. Characterization of epidermal neural creststem cell (EPI-NCSC) grafts in the lesioned spinal cord. Mol.Cell. Neurosci. 32, 67–81.
Someya, Y., Koda, M., Dezawa, M., Kadota, T., Hashimoto, M.,Kamada, T., Nishio, Y., Kadota, R., Mannoji, C., Miyashita, T.,Okawa, A., Yoshinaga, K., Yamazaki, M., 2008. Reductionof cystic cavity, promotion of axonal regeneration andsparing, and functional recovery with transplanted bonemarrow stromal cell-derived Schwann cells after contusioninjury to the adult rat spinal cord. J. Neurosurg. Spine 9,600–610.
Swanger, S.A., Neuhuber, B., Himes, T., Bakshi, A., Fischer, I., 2005.Analysis of allogeneic and syngeneic bone marrow stromalcell graft survival in the spinal cord. Cell Transplant. 14,775–786.
Syková, E., Homola, A.,Mazanec, R., Lachmann, H., Konrádová, S.L.,Kobylka, P., Pádr, R., Meuwirth, J., Komrska, V., Vávra, V., Stulík,J., Bojar, M., 2006. Autologous bone marrowtransplantation in patients with subacute and chronic spinalcord injury. Cell Transplant. 15, 675–687.
Wright, K.T., Masri, W.E., Osman, A., Roberts, S., Chamberlain, G.,Ashton, B.A., Johnson, W.E.B., 2007. Bone marrow stromal cellsstimulate neurite outgrowth over neural proteoglycans (CSPG),myelin associated glycoprotein andNogo-A. Biochem. Biophys.Res. Commun. 354, 559–566.
Wright, K.T., Masri, W.E., Osman, A., Roberts, S., Trivedi, J., Ashton,B.A., Johnson, W.E., 2008. The cell culture expansion of bonemarrow stromal cells from human with spinal cord injury:implication for future cell transplantation therapy. Spinal Cord46, 811–817.
Wu, S.F., Suzuki, Y., Noda, T., Bai, H.L., Kitada, M., Kataoka, K.,Nishimura, Y., Ide, C., 2002. Immunohistochemical andelectron microscopic study of invasion and differentiationin spinal cord lesion of neural stem cells graftedthrough cerebrospinal fluid in rat. J. Neurosci. Res. 69,940–945.
Wu, S.F., Suzuki, Y., Noda, T., Bai, H., Kitada, M., Kataoka, K., Chou,H., Ide, C., 2003. Bone marrow stromal cells enhancedifferentiation of co-cultured neurosphere cells and promoteregeneration of injured spinal cord. J. Neurosci. Res. 72,343–351.
Xu, X.M., Zhang, S.X., Li, H., Aebischer, P., Bunge, M.B., 1999.Regrowth of axons into the distal spinal cord through aSchwann-cell-seeded mini-channel implanted intohemisected adult rat spinal cord. Eur. J. Neurosci. 11,1723–1740.
Yoon, S.H., Shim, Y.S., Park, Y.H., Chung, J.K., Nam, J.H., Kim, M.O.,Park, H.C., Park, S.R., Min, B.H., Kim, E.Y., Choi, B.H., Park, H., Ha,Y., 2007. Complete spinal cord injury treatment usingautologous bone marrow cell transplantation and bonemarrow stimulation with granulocyte macrophage-colonystimulating factor: phase I/II clinical trial. Stem Cells 25,2066–2073.

47B R A I N R E S E A R C H 1 3 3 2 ( 2 0 1 0 ) 3 2 – 4 7
Yoshihara, T., Ohta, M., Itokazu, Y., Matsumoto, N., Dezawa, M.,Suzuki, Y., Taguchi, A., Watanabe, Y., Adachi, Y., Ikehara, S.,Sugimoto, H., Ide, C., 2007. Neuroprotective effect of bonemarrow-derived mononuclear cells promoting functionalrecovery from spinal cord injury. J. Neurotrauma 24, 1026–1036.
Zurita, M., Vaquero, J., Bonilla, C., Santos, M., De Haro, J.,Ova, S., Aguayo, C., 2008. Functional recovery ofchronic paraplegic pigs after autologous transplantationof bone marrow stromal cells. Transplantation 86,845–853.




















