
BMP7 and SHH regulate< i> Pax2 in mouse retinal astrocytes by relieving TLX repression
30
BMP7 and SHH Regulate Pax2 in Mouse Retinal Astrocytes by Relieving TLX Repression Rachna Sehgal 1,4 , Nader Sheibani 2 , Simon J Rhodes 3,4 , and Teri L Belecky Adams 1,4 1 Department of Biology, Indiana University-Purdue University Indianapolis, 723 W Michigan St, Indianapolis, IN-46202 2 Department of Ophthalmology and Visual Sciences, University of Wisconsin, School of Medicine and Public Health, 600 Highland Avenue, K6/458 CSC, Madison, WI-53792-4673 3 Department of Cellular and Integrative Physiology, Indiana University School of Medicine, MS 362, 635 N. Barnhill Drive, Indianapolis, IN-46202 4 Center for Regenerative Biology and Medicine, Indiana University- Purdue University Indianapolis, 723 W Michigan St, Indianapolis, IN-46202 Abstract Pax2 is essential for development of the neural tube, urogenital system, optic vesicle, optic cup and optic tract. In the eye, Pax2 deficiency is associated with coloboma, a loss of astrocytes in the optic nerve and retina, and abnormal axonal pathfinding of the ganglion cells axons at the optic chiasm. Thus, appropriate expression of Pax2 is essential for astrocytes determination and differentiation. Although BMP7 and SHH have been shown to regulate Pax2 expression, the molecular mechanism by which this regulation occurs is not well understood. In this study, we determined that BMP7 and SHH activate Pax2 expression in mouse retinal astrocyte precursors in vitro. SHH appeared to play a dual role in Pax2 regulation; 1) SHH may regulate BMP7 expression, and 2) the SHH pathway cooperates with the BMP pathway to regulate Pax2 expression. BMP and SHH pathway members can interact separately or together with TLX, a repressor protein in the tailless transcription factor family. Here we show that the interaction of both pathways with TLX relieves the repression of Pax2 expression in mouse retinal astrocytes. Together these data reveal a new mechanism for the cooperative actions of signaling pathways in astrocyte determination and differentiation and suggest interactions of regulatory pathways that are applicable to other developmental programs. Keywords SHH; BMP7; TLX; PAX2; astrocytes; retina Correspondence should be sent to following address: Teri L Belecky Adams, Department of Biology, SL306, Center for Regenerative Biology and Medicine, Indiana University-Purdue University Indianapolis, 723 W Michigan St., Indianapolis, IN-46202, Tel. (317)278-5715, Fax (317) 274-2846, [email protected]. Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. NIH Public Access Author Manuscript Dev Biol. Author manuscript; available in PMC 2010 August 15. Published in final edited form as: Dev Biol. 2009 August 15; 332(2): 429–443. doi:10.1016/j.ydbio.2009.05.579. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
Transcript of BMP7 and SHH regulate< i> Pax2 in mouse retinal astrocytes by relieving TLX repression

BMP7 and SHH Regulate Pax2 in Mouse Retinal Astrocytes byRelieving TLX Repression
Rachna Sehgal1,4, Nader Sheibani2, Simon J Rhodes3,4, and Teri L Belecky Adams1,41 Department of Biology, Indiana University-Purdue University Indianapolis, 723 W Michigan St,Indianapolis, IN-462022 Department of Ophthalmology and Visual Sciences, University of Wisconsin, School of Medicineand Public Health, 600 Highland Avenue, K6/458 CSC, Madison, WI-53792-46733 Department of Cellular and Integrative Physiology, Indiana University School of Medicine, MS362, 635 N. Barnhill Drive, Indianapolis, IN-462024 Center for Regenerative Biology and Medicine, Indiana University- Purdue UniversityIndianapolis, 723 W Michigan St, Indianapolis, IN-46202
AbstractPax2 is essential for development of the neural tube, urogenital system, optic vesicle, optic cupand optic tract. In the eye, Pax2 deficiency is associated with coloboma, a loss of astrocytes in theoptic nerve and retina, and abnormal axonal pathfinding of the ganglion cells axons at the opticchiasm. Thus, appropriate expression of Pax2 is essential for astrocytes determination anddifferentiation. Although BMP7 and SHH have been shown to regulate Pax2 expression, themolecular mechanism by which this regulation occurs is not well understood. In this study, wedetermined that BMP7 and SHH activate Pax2 expression in mouse retinal astrocyte precursors invitro. SHH appeared to play a dual role in Pax2 regulation; 1) SHH may regulate BMP7expression, and 2) the SHH pathway cooperates with the BMP pathway to regulate Pax2expression. BMP and SHH pathway members can interact separately or together with TLX, arepressor protein in the tailless transcription factor family. Here we show that the interaction ofboth pathways with TLX relieves the repression of Pax2 expression in mouse retinal astrocytes.Together these data reveal a new mechanism for the cooperative actions of signaling pathways inastrocyte determination and differentiation and suggest interactions of regulatory pathways thatare applicable to other developmental programs.
KeywordsSHH; BMP7; TLX; PAX2; astrocytes; retina
Correspondence should be sent to following address: Teri L Belecky Adams, Department of Biology, SL306, Center for RegenerativeBiology and Medicine, Indiana University-Purdue University Indianapolis, 723 W Michigan St., Indianapolis, IN-46202, Tel.(317)278-5715, Fax (317) 274-2846, [email protected]'s Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to ourcustomers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review ofthe resulting proof before it is published in its final citable form. Please note that during the production process errors may bediscovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
NIH Public AccessAuthor ManuscriptDev Biol. Author manuscript; available in PMC 2010 August 15.
Published in final edited form as:Dev Biol. 2009 August 15; 332(2): 429–443. doi:10.1016/j.ydbio.2009.05.579.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

IntroductionDuring induction of the eye, the interaction of microenvironmental cues, signalingpathways, and transcription factors are critical to the proper patterning of the optic cup, lens,and optic stalk. There are a large variety of inductive factors, such as retinoic acid, fibroblastgrowth factors (FGFs), sonic hedgehog (SHH), WNTs, activins, and bone morphogeneticproteins (BMPs) that are essential for various aspects of patterning. In addition, there are alarge number of transcription factors whose expression patterns rely in part on the presenceof these inductive factors (Fuhrmann et al., 2000; Vogel-Hopker et al., 2000; Zhao et al.,2001). Although the regulation of transcription factor expression by these factors is partiallycharacterized, our understanding of the mechanisms by which these factors cooperate needsinvestigation.
One protein that has been identified as critical to optic cup development is PAX2, a memberof the Paired Box family of transcription factors. PAX2 plays a pivotal role in eye, ear,spinal cord and kidney development (Dressler et al., 1990; Eccles et al., 1992; Nornes et al.,1990). Loss of Pax2 expression has been linked to various developmental and functionalabnormalities such as coloboma (lack of choroid fissure formation), a marked reduction inoptic nerve and retinal astrocytes, misprojections of the optic nerve, inner earmalformations, and kidney hypoplasia (Dressler and Douglass, 1992). Interestingly, Pax2overexpression in the optic cup can phenocopy the loss of Pax2 expression in some respects,namely the formation of colobomas, albeit the formation of the colobomas in gain- and loss-of-function studies appears to be different (Sehgal et al., 2008). In the vertebrate eye, PAX2is co-expressed with other transcription factors like PAX6 and CHX10, throughout the opticvesicle stage. As development proceeds, inductive factors restrict the expression patterns ofthe various transcription factors to lens, retina, retinal pigmented epithelium (RPE) or opticstalk. At the optic cup stage, PAX2 is expressed in ventral optic cup and optic stalk andeventually becomes restricted entirely to the optic stalk, the cells of which eventually giverise to the astrocytes of the optic nerve (Chu et al., 2001). A subpopulation of the optic nerveastrocytes then migrate into the optic cup to generate retinal astrocytes (Chan-Ling andStone, 1991; Zhang and Stone, 1997)
The BMPs are a large family of secreted proteins that are known to be critical for theformation of a large array of tissues in the developing organism (Chen et al., 2004). BMPscan signal through SMAD-dependent and SMAD-independent pathways (Derynck andZhang, 2003; Herpin and Cunningham, 2007). In the canonical signaling pathway, BMPsignaling is mediated by the receptor SMADs (R-SMADs), 1, 5, and 8. Upon activation,these SMADs form a complex with SMAD4 and the complex is translocated to the nucleusto regulate transcription (Dudley and Robertson, 1997). SMADs can regulate geneexpression by binding directly to DNA sequences, or by interacting with co-activators andco-repressors (Lee et al., 2000; Miyazono, 1999; Miyazono, 2000a; Miyazono, 2000b;Miyazono et al., 2005; Zhang and Derynck, 2000).
Many of the BMPs are expressed in tissues surrounding the eyefield, optic vesicles, andoptic cups, and within portions of the eye itself in the developing and mature organ(Belecky-Adams and Adler, 2001; Dale and Jones, 1999; Hocking and McFarlane, 2007;Koshiba-Takeuchi et al., 2000; Wilson et al., 2007; Wordinger et al., 2002). Organogenesisof the eye appears to be reliant on BMPs, in particular BMP4 and 7 (Adler and Belecky-Adams, 2002; Dudley et al., 1995; Koshiba-Takeuchi et al., 2000; Sakuta et al., 2001;Trousse et al., 2001). BMP7 is hypothesized to be critical for many steps in the progressivedevelopment of the eye, including the patterning of the eyefield, optic stalk, and optic nervehead, and differentiation of the retinal pigmented epithelium, lens placode, anterior segment,and retinal ganglion cells (Adler and Belecky-Adams, 2002; Dale et al., 1997; Dale and
Sehgal et al. Page 2
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Jones, 1999; Dudley and Robertson, 1997; Furuta et al., 1997; Hung et al., 2002; Jena et al.,1997; Morcillo et al., 2006; Muller et al., 2007; Wawersik et al., 1999; Wordinger et al.,2002; Zhao et al., 2002). At the eye field stage, BMP7, is expressed along with SHH in theprechordal mesoderm underlying the diencephalon and controls the identity of the rostraldiencephalon (Dale et al., 1997). BMP7 has been shown by several investigators to regulatethe expression of Pax2, although it is unclear whether this regulation is direct or indirect(Adler and Belecky-Adams, 2002; Dale et al., 1997; Morcillo et al., 2006).
SHH is a member of the hedgehog family that, much like the BMPs, appears to be involvedin the development of many tissues, including midline structures, limbs, gastrointestinaltract, and central nervous system (CNS; (Chari and McDonnell, 2007). In vertebrates, thehedgehog proteins signal through the GLI1, GLI2 and GLI3 transcription factors (Hooperand Scott, 2005). How the GLI proteins are regulated in vertebrates is still not wellunderstood. In both Drosophila and vertebrates, the SHH binds to the patched homologues(PTC) and relieves repression on another transmembrane protein, smoothened (SMO). InDrosophila, active SMO controls the proteolytic processing of the GLI homologue cubitusinterruptus (Ci). Ci is bifunctional; in its full length form the protein acts as a transcriptionalactivator, but following cleavage, the protein will act as a transcriptional repressor (Parkinand Ingham, 2008). SMO activity regulates the Ci processing through a complex thatincludes kinesin protein Cos-2, a serine-threonine kinase fused, and a novel protein knownas suppressor of fused (Huangfu and Anderson, 2006). The signaling of the SMO proteinand the conservation of pathway members is not clear in vertebrates, and only one of theGLI proteins, GLI3, has been shown to be proteolytically cleaved to give rise to thetranscriptional repressor form of the protein (Huangfu and Anderson, 2006). Hence, GLI3can function as a functional repressor and activator, while GLI1 and GLI2 are thought to actonly as activators (Bai et al., 2004; Motoyama et al., 2003).
During development, SHH is released from the prechordal mesoderm to induce theformation of the floorplate in the midline neural plate (Dale et al., 1997). The floorplateitself then expresses SHH which is proposed to regulate the formation of ventral neural tubecell fates. The eyefield, which initially forms in the region overlying the prechordalmesoderm, is cut into two pieces as the floorplate of the neural plate is induced by SHH(Ekker et al., 1995; Macdonald et al., 1995; Nagase et al., 2005; Ohkubo et al., 2002). At amolecular level, SHH induces Pax2 expression in this region, which in turn reciprocallyinhibits Pax6. The region that continues to express Pax2 in the ventral diencephalonbecomes the optic stalk. Later in development, SHH expression is restricted to retinalganglion cells and SHH derived from ganglion cell axons is thought to drive theproliferation of optic nerve astrocytes (Dakubo et al., 2008; Dakubo et al., 2003; Wallaceand Raff, 1999). SHH has also been shown by several investigators to regulate theexpression of Pax2 in the eye (Macdonald et al., 1995; Schimmenti et al., 2003; Wang et al.,2005). Bovolenta and colleagues found that Pax2 is regulated by sequential activity of SHHand BMP7 (Morcillo et al., 2006). While it is generally accepted that SHH regulates Pax2expression, little is known about the mechanism of Pax2 regulation by these signalingmolecules and whether the regulation is direct or indirect (Macdonald et al., 1995; Morcilloet al., 2006; Nakamura, 2001).
Pax2 is also regulated in the vertebrate eye by TLX, a member of the tailless class of orphannuclear receptors, which has been shown to repress expression of Pax2 (Yu et al., 2000; Yuet al., 1994). In early stages of development, Tlx is expressed mainly in the dorsal retinawith low levels of expression in the ventral retina, whereas Pax2 expression is limitedprimarily to the ventral retina and optic stalk (Nornes et al., 1990). In the chick optic vesicle,expression of Tlx and Pax2 is dynamic and transiently overlaps, particularly in thepresumptive optic stalk. Later in development, Tlx is expressed in retinal progenitor cells
Sehgal et al. Page 3
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

and Müller glial cells in the retina. TLX represses Pax2 by binding to a conservedrecognition sequence (AAGTCA) ~ 80base pairs upstream of the TATA box (Miyawaki etal., 2004; Yu et al., 2000).
The studies reported here were directed at understanding the molecular mechanism by whichSHH and BMP7 regulate Pax2 expression in mouse retinal astrocytes. We show that 1)BMP7 expression appears to be regulated by SHH, and 2) SHH and BMP7 interact withTLX on the Pax2 promoter and relieve repression by TLX in retinal astrocytes.
ResultsPhospho-SMAD1 and GLI2 and 3 are present in the developing optic vesicle, optic stalk,and nerve
Previous reports from our laboratory and others have proposed that SHH and BMP7 arecritical to the development of the optic stalk astrocytes and have shown that mRNAencoding intracellular signaling components of the BMP and SHH pathways are present inthe developing optic stalk (Belecky-Adams and Adler, 2000; Dakubo et al., 2003; Furimskyand Wallace, 2006; Morcillo et al., 2006). Immunohistochemistry was performed tocompare the localization of PAX2 with GLI2, GLI3, and pSMAD1 proteins at embryonicday 9.5 (E9.5) optic vesicle, E10.5 optic stalk, and E16 optic nerve. As has been describedin previous reports (Nornes et al., 1990), PAX2 was not present in the optic stalk at E9.5(Fig 1A–C;. However, GLI2 (Fig 1D–F), GLI3 (Fig 1G–I) and pSMAD1 (Fig 1J–L) were allpresent in the optic vesicle at E9.5. GLI2 and phospho-SMAD1 (pSMAD1) were bothlocalized primarily in cells at the ventricular edge of the optic vesicle, where cells wereundergoing mitosis (compare Fig 1F and L), while GLI3 immunolabel was presentthroughout the optic vesicle (Fig 1I). Further, GLI3 label was also present in thepresumptive lens ectoderm and in a subpopulation of mesenchyme surrounding the opticvesicle (Fig 1I). At E10.5, PAX2 was localized primarily to the ventral optic cup and opticstalk (Fig 2A–D). GLI2 protein was localized to the optic stalk, optic cup and lens vesicle(Fig 2E–H). GLI3 was widely expressed throughout the optic stalk, retinal pigmentedepithelium (RPE), optic cup, and lens vesicle (Fig 2I–L). Phospho-SMAD1 was alsolocalized to the optic stalk, RPE, optic cup and lens placode (Fig 2M–P). As was noted atE9.5, GLI2 and pSMAD1 immunolabel appeared to be more intense at the ventricular edgeof the dorsal and ventral optic stalk and optic cup, where dividing cells are located(arrowheads, Fig 2H and P). In the E16 eye, PAX2 is restricted to the optic nerve head andoptic nerve (Fig 3A–C). GLI2 (Fig 3D–F), GLI3 (Fig 3G–I), and pSMAD1 (Fig 3J–L) wereall localized to subpopulations of cells in the optic nerve head (ONH) and optic nerve (ON).
Characteristics of retinal astrocyte precursors in vitroTo begin to unravel the molecular mechanism by which the BMP7 and SHH pathwaysmight regulate the differentiation of retinal astrocytes, we turned to an in vitro model systemwhich would allow us to uniformly treat cells with various combinations of the two growthfactors and probe molecular interactions between the two pathways. We chose to useprimary retinal astrocyte precursors isolated from dissociated retina obtained from 4-week-old transgenic mice that ubiquitously express a temperature sensitive SV-40 large T antigen(immorto-mice; (Jat et al., 1991)). Collagenase-treated cells were cultured following thedepletion of endothelial cells using antibody-coated magnetic beads (Scheef et al., 2005).Cells were plated and grown at the permissive temperature of 33°C and surviving cellsallowed to reach confluence over a 2–3 week period. Confluent cells were subsequentlycharacterized using immunocytochemistry and fluorescence-activated cell sorting (FACs)analyses. Figure 4A and B show the morphology of the cells under phase contrastmicroscopy. Cells were positive for PAX2 (Fig 4C), GFAP (Fig 4D), and NG2 (Fig 4E).
Sehgal et al. Page 4
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Consistent with previous analyses of the cells, FACs analyses indicated that nearly 100% ofthe cells expressed GFAP and NG2 (Fig 4F, G;(Scheef et al., 2005)). Theimmunocytochemistry and FACs analyses coupled with the previous analysis of the cellsindicated that the cells showed characteristics consistent with multipotent retinal astrocyteprecursors (Scheef et al., 2005; Stallcup and Beasley, 1987; Yokoyama et al., 2006).
SHH and BMP7 activate PAX2 and phospho PAX2To determine if addition of BMP7 and/or SHH to retinal astrocytes affected morphology,cells were treated with vehicle, BMP7, SHH, or BMP7+ SHH for 2 days and immunolabeledfor PAX2 or GFAP. While there were no detectable changes in appearance or number (datanot shown) of the cells, there were evident increases in the amount of PAX2immunofluorescence of treated cells in comparison to the control vehicle-treated cells (Fig5A). To verify the increase in levels of PAX2, lysates of treated cells were subjected towestern analysis. Cells were treated with vehicle, BMP7, SHH, or both BMP7 and SHH for4 hours, cell lysates separated by SDS-PAGE, and immunoblot for PAX2 performed. Asshown in Fig 5; B, there was a notable increase in both isoforms of PAX2 in lysates fromtreated cells in comparison to the vehicle control (Fig 5B). These assays showed the 46 and48 kDa forms seen in previous studies (Dressler and Douglass, 1992) also were present inthese cells (Fig 5B). Densitometric analysis revealed a 3-fold increase in PAX2 protein inlysates treated with BMP7 or SHH and a 4-fold increase in lysates co-treated with BMP7and SHH compared to control cultures treated with vehicle alone (Fig 5B′).
PAX2 activity is regulated by phosphorylation of a carboxyl-terminal trans- activationdomain that is rich in serine and threonine residues (Cai et al., 2002). To determine if therewas a concomitant increase in phospho-PAX2 following treatment of retinal astrocytes withBMP7, SHH, or both, western blotting of lysates from treated cells was performed using aphospho-PAX2 specific antibody. Immunoblots showed 1.6-fold and 2-fold increases inphospho-PAX2 levels in lysates treated with BMP7 or SHH, respectively, in comparison tovehicle treated controls, while lysates co-treated with BMP7 and SHH showed anapproximately 3-fold increase over controls (Fig 5C, C′).
Cells treated with SHH have increased levels of phospho SMAD1Because the observed increases in both PAX2 and phospho-PAX2 in the cells treated withBMP7 and SHH were sub-additive, the possibility of cross talk between the two signalingpathways was investigated. Retinal astrocytes were treated with control vehicle, BMP7,SHH, or both BMP7 and SHH and lysates of cells were assayed for the downstreamsignaling component of the canonical BMP pathway, pSMAD1. Vehicle-treated cellsshowed a basal level of pSMAD1 (Fig 6A). As expected, BMP7-treated cells showed anincrease in the levels of pSMAD1. Interestingly, there was also an increase in pSMAD1following SHH treatment, and co-treatment yielded a sub-additive increase in pSMAD1similar to what had been seen in previous experiments in which BMP7 and SHH were addedto the cultures.
A densitometric analysis of the western blot using β-tubulin as a loading control showed a 2-fold increase in pSMAD1 levels in BMP7- treated, a 2.7-fold increase in SHH-treated and a3-fold increase in co-treated cells over vehicle (Fig 6A, A′). As a negative control the sameblot was subjected to an antibody specific for phospho-SMAD2, the downstream signalingcomponent of activin and other TGFβ growth factors. Quantitative analysis showed nosignificant changes in phospho-SMAD2 following treatment of cells in comparison to thevehicle controls (Fig 6B, B′).
Sehgal et al. Page 5
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Follistatin abolishes the increase in phospho SMAD1 protein in SHH-treated cellsThe previous result suggested that the SHH pathway was regulating pSMAD1 levels, eitherthrough pathway cross talk or by regulating expression of one or more members of BMPpathway. If SHH regulates BMP7 expression, we hypothesized that treatment of cells with aBMP7 inhibitor should abolish the effects of both BMP7 and SHH on pSMAD1 levels. Toblock BMP signaling, we treated cells with each growth factor separately or in combinationwith follistatin and a BMP7 function blocking antibody. Cell lysates were analyzed forlevels of pSMAD1. Consistent with our prior observations, treatment of the cells withBMP7, SHH or both factors yielded an increase in pSMAD1 that was greater in the lysatesfrom control-treated cells (Fig 6D). As would be expected if SHH signaling were regulatingBMP expression, co-treatment with follistatin and BMP7 function blocking antibodyblocked pSMAD1 signaling following treatment with each factor alone or in combination. Adensitometric analysis of the blots using α-tubulin as a loading control showed a 2-foldincrease in lysates treated with BMP7, a 2.5-fold increase in SHH-treated lysates and a 3.3-fold increase in lysates co-treated with BMP7 and SHH. Lysates treated with growth factorsand combination of follistatin and BMP7 function blocking serum showed >10-fold decreasein levels of pSMAD1 (Fig 6D, D′).
Cyclopamine treatment decreases BMP7 protein levels in vitroTo further test if BMP7 expression is regulated by SHH, cells were treated with vehicle orcyclopamine (an inhibitor of SHH signaling), and analyzed for levels of BMP7 by westernblotting. BMP7 protein levels in cyclopamine treated cells were decreased as compared tothe vehicle treated cells. Densitometric analysis showed a 2-fold decrease in BMP7 proteinlevels in cyclopamine treated cells compared to vehicle controls (Fig 6E, E′).
BMP7 and SHH decrease the binding of TLX protein on Pax2 promoterTailless (TLX) is an orphan nuclear receptor that acts as a suppressor of Pax2 expressionand is essential for eye development (Yu et al., 2000). We hypothesized that TLX repressionwould have to be removed to allow Pax2 expression and induction of astrocyte fate in theoptic stalk. To begin to examine whether the intracellular signaling pathways activated byBMP7 and SHH decreased TLX availability to interact with the proximal Pax2 promotersequences, we performed EMSA reactions using nuclear extracts of treated astrocytes andbiotin labeled oligonucleotides containing the TLX binding site in the Pax2 promoter. If theBMP and/or SHH pathways interfered with TLX repression, then one might expect to see adecrease in TLX binding to its consensus sequences in the presence of nuclear extracts fromtreated cells. As was hypothesized, TLX binding activity for the Pax2 promoter sequencewas decreased in cells treated with both BMP7 and SHH (Fig 7A, C; n=4). Parallel EMSAreactions using in vitro translated TLX protein (Fig 7B, Lane 2) and nuclear extracts fromastrocytes (Fig 7B, Lane 3) served as controls for TLX protein/DNA complex migration(Fig 7B). Further, treatment of EMSA reactions containing nuclear extracts from astrocytesincubated with the oligo representing the TLX binding site followed by incubation with anantibody specific for TLX showed a supershift of the detected band confirming the presenceof TLX in the complex (Fig 7B, lane 4).
To confirm that there was indeed less TLX bound to the TLX consensus site of the Pax2promoter, a chromatin immunoprecipitation (ChIP) using TLX antibody was performedusing lysates from retinal astrocytes treated with vehicle, BMP7, SHH, or BMP7 + SHH.Quantitative PCR (qPCR) was done using primers specific for a 60 bp region of the Pax2promoter encompassing the TLX-binding region of the Pax2 promoter (Figure 7D).Comparable to the EMSA analysis, treatment of cells with BMP7, SHH, or BMP7 + SHHdecreased the amount of Pax2 amplified, indicating that less TLX was immunoprecipitatedas a complex with the Pax2 promoter in treated cells (Fig 7D).
Sehgal et al. Page 6
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

TLX, GLI2 and pSMAD1 interact with each otherThere are several ways which the SHH and BMP pathways might decrease TLX binding toits consensus site. For instance, the SMADs and/or GLIs might 1) bind to TLX to keep itfrom binding to DNA or release it from the consensus site, or 2) bind to DNA and stericallyinterfere with TLX binding to its consensus sequences. To analyze whether decreasedbinding of TLX following BMP7 and SHH signaling involves interaction of theirdownstream components pSMAD1 and GLI2 respectively, co-immunoprecipitation (Co-Ip)was performed. Retinal astrocytes were treated with vehicle and total proteins were isolated.Extracted nuclear proteins from treated cells were precipitated with control β-tubulin, GLI2or pSMAD1 antibodies and then blotted with antibody specific to TLX (Fig 7E; a),pSMAD1 (Fig 7E; b) or GLI2 (Fig 7E; c). As expected, lysates incubated with control β-tubulin antibody do not co-precipitate TLX, GLI2, or pSMAD1 (Fig 7E; first lane). All ofthe proteins were detected following immunoprecipitation using the pSMAD1 or GLI2antibodies, consistent with the hypothesis that these proteins interact with each other (n=3).To determine if TLX, GLI2 and/or pSMAD1 could also form complexes in treated cells, Co-Ips were performed using nuclear extracts from cells treated with BMP7, SHH, or BMP7 +SHH (n=3). As was seen in vehicle-treated cells, TLX, GLI2, and pSMAD1 wereimmunoprecipitated as a complex when cells were incubated with pSMAD1 or GLI2antibody (data not shown).
An alternate hypothesis would be that BMP7 and SHH treatment of astrocytesdownregulates the expression of TLX. To rule out BMP7 or SHH regulation of TLXexpression, western blotting was performed on BMP7, SHH, and co-treated cells. Nosignificant differences in amounts of TLX protein were apparent in the treated cellscompared to controls (Fig 7F, F′).
To test whether SMAD1 or GLI2 could bind to sequences within the proximal promoter ofPax2 and thereby sterically hinder TLX binding, we performed an avidin-biotin complexDNA-binding (ABCD) assay. This assay is used to evaluate the interactions of endogenousproteins (SMAD1 and GLI2) with the biotinylated double-stranded DNA oligonucleotidescontaining the binding element of interest. Total cellular proteins from retinal astrocyteswere isolated and incubated with biotin-labeled oligonucleotides representing the TLXbinding site in the Pax2 promoter. Proteins bound to the TLX binding site oligonucleotideswere purified by streptavidin agarose chromatography, eluted from the beads, separated onSDS-PAGE gels, transferred and blotted with antibody specific for TLX, pSMAD1, orGLI2. Consistent with our other observations, TLX protein was observed to bind to the TLXbinding site (Fig 7G). Interestingly, neither pSMAD1 or GLI2 could be detected, althoughcontrol experiments showed that pSMAD1 and GLI2 were detectable in assays using theconsensus SMAD or GLI sites respectively (data not shown).
SHH and BMP7 regulate Pax2 promoter activity in the presence of TLXTo determine if SHH and BMP7 can regulate the activity of the Pax2 promoter withinretinal astrocytes, luciferase assays were performed. Astrocytes were transiently co-transfected with an expression vector for TLX (CMV-TLX) and a luciferase reporter genecontaining 900 bp of the proximal Pax2 upstream region (Pax2-Luc). Cells were incubatedwith vehicle or growth factors for 24 hours, lysed, and total proteins were analyzed forluciferase activity. As expected, cells co-transfected with CMV-TLX and Pax2-Luc vectorsshowed decreased levels of luciferase activity in comparison to those transfected with Pax2-Luc plus control expression vector (Fig 8A). Cells transfected with Pax2-Luc andsubsequently treated with either BMP7 and/or SHH showed at least a 2-fold increase inluciferase activity. Consistent with our hypothesis, cells co-transfected with CMV-TLX and
Sehgal et al. Page 7
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Pax2-Luc and treated with either BMP7, SHH or BMP7 and SHH were able to relieve Pax2repression to varying degrees (Fig. 8A).
P-SMAD1 and GLI2 decrease TLX-binding at the Pax2 proximal promoterTo verify that pSMAD1 and GLI2 are found at the Pax2 promoter in situ, cells were treatedwith vehicle or BMP7 + SHH and a ChIP analysis performed on the lysates using antibodiesspecific for pSMAD1, GLI2, or TLX. The results of the ChIP were then analyzed by qPCRusing primers that amplified the proximal Pax2 promoter (Fig 8B). As was seen in theprevious ChIP analysis, TLX appeared to be abundant at the Pax2 promoter in vehicle-treated cells. However, in lysates from treated cells there was a decrease in Pax2amplification of approximately 2-fold. Furthermore, treated cells showed a dramaticincrease in the levels of amplified Pax2 when pSMAD1 and GLI2 were used for the ChIPanalysis. Together these results support the hypothesis that untreated retinal astrocyteprecursors have high levels of TLX bound to the Pax2 promoter, while cells treated withBMP7 and/or SHH have relieved TLX repression of Pax2 expression by removal of TLXand a concomitant increase in pSMAD1 and GLI2 bound to the proximal Pax2 promoter.
DiscussionThe present study focuses on the mechanism of regulation of Pax2 by SHH and BMP7.Indirect immunohistochemistry showed that critical members of the BMP and SHHpathways were present in the optic vesicle just prior to Pax2 expression and were present inthe PAX2-expressing optic stalk and optic nerve. Primary cultures of retinal astrocytes wereused to probe the interactions of the two pathways in retinal astrocytes. Cells were treatedwith BMP7 and SHH and the results analyzed by western blotting, electrophoretic mobilityshift assay, ABCD assay, co-immunoprecipitation, luciferase assays, and ChIP-qPCR. Thefollowing conclusions could be drawn from these experiments; 1) BMP7 and SHH increasePAX2 and phospho- PAX2 levels in astrocytes, 2) SHH regulates BMP7 expression inastrocytes, 3) SHH and BMP7 relieve Pax2 repression by decreasing the binding of TLX,and 4) the modulation of TLX repression of Pax2 expression is likely to include binding ofboth pSMAD1 and GLI2 to TLX, rather than regulation of TLX mRNA or protein levels orsteric interference of TLX binding to the Pax2 promoter.
Cooperation of SHH and BMP pathwaysAlthough the SHH and BMP pathways are typically described as having antagonistic effectsin patterning of the central nervous system, our results, as well as the results of other labs,indicate that this may be a simplistic model (Arkell and Beddington, 1997; Liem et al.,1995). An excellent example of this is the interactions of BMPs and SHH in the neural tube.BMPs have been proposed as one of the “dorsalizing” factors, along with wnts and activins,whereas SHH has been proposed as the “ventralizing” factor of the neural tube (Ulloa andBriscoe, 2007). What is rarely acknowledged is the fact that some of the BMPs (as well ascertain wnts) are also expressed in the prechordal mesoderm underlying the forebrain andmidline portion of the neural tube (Dale et al., 1997; Furuta et al., 1997; Placzek andBriscoe, 2005). Dale et al. (1997) have shown that both BMP7 and SHH are co-expressed inthe prechordal mesoderm and are required to induce the formation of ventral midline cells ofthe rostral diencephalon. BMP7 reportedly acts on the midline neural cells to modify theirresponse to SHH, inducing the expression of markers found only in rostral diencephalon(Dale et al., 1997). Interestingly, studies by Morcillo et al (2006) have also suggested thatthe activity of BMP7 and SHH are necessary for the proper patterning of the optic nervehead and optic fissure and that the activity of BMP7 prior to SHH signaling is necessary forPax2 expressing cells in the optic nerve head.
Sehgal et al. Page 8
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

These findings are not restricted to the optic nerve and Pax2. Investigations in other tissuesalso suggest that SHH and BMPs can act cooperatively (Suzuki et al., 2004; Yuasa et al.,2002). The cooperation of SHH and BMPs has been characterized at several different levels.First, SHH and BMPs have been shown by various investigators to regulate each other’sexpression (Kawai and Sugiura, 2001; Zhang et al., 2000), although the level of BMPspresent appeared to determine whether the SHH expression is upregulated or inhibited(Zhang et al., 2000). That BMPs may regulate the expression of SHH is consistent with theidea that BMP7 and SHH sequentially regulate patterning and differentiation in thediencephalon (Dale et al., 1997; Morcillo et al., 2006). A second mechanism by which SHHand BMPs may act coordinately may be crosstalk of the pathways with one another. Forinstance, there is evidence that the BMP pathways interact with the notch, Toll, and MAPKinase pathways (reviewed in (Herpin and Cunningham, 2007). Finally, the last way inwhich the BMP and SHH pathways might act coordinately is through co-regulation ofgenes. The SMADs have been shown to regulate gene expression either by binding to DNAdirectly with co-activators or co-repressors, or by binding to other transcription factors toregulate their binding to DNA (Miyazono et al., 2005). There are at least 3 consensus DNArecognition sequences for the BMP-activated SMADs (Kim et al., 1997; Kusanagi et al.,2000; Shim et al., 2002). The SMADs have also been shown to interact with many differenttranscription factors, although only a handful of factors have been implicated in binding tothe BMP-related SMADs, including Runx, Menin, YY1, Msx1, Vent2, MyoD, Hoxc8, SIP1,FoxG1, p300, CBP, cSki, SnoN, and GCN5 (Miyazono et al., 2005). This is one of the firststudies to show that BMP and SHH pathways may interact with each other to regulate geneexpression.
TLX and Pax2 regulationTLX is an orphan nuclear receptor that plays an important role in regulating cell numbersand astrocyte development in the developing retina (Miyawaki et al., 2004). TLX is alsoimplicated in forebrain development and emotional behavior (Roy et al., 2002). TLXregulates Pax2 by binding the promoter at a recognition sequence which has been conservedin mouse and humans (Yu et al., 2000). In addition, TLX is expressed in proangiogenicastrocytes and its expression is controlled by oxygen concentration. TLX acts asproangiogenic switch which becomes active in low concentrations of oxygen. After retinalangiogenesis, TLX is downregulated and its angiogenic activity is switched off whenastrocytes come in contact with the blood vessels (Uemura et al., 2006). BMPs also areexpressed in endothelial cells as well as playing key roles to regulate the developmentalprogram of hematopoietic cells in humans (Bhatia et al., 1999; Fan et al., 2002). In thisstudy, BMP7 and SHH were shown to relieve the repression of Pax2 by TLX by interactingwith TLX. Astrocyte migration into the retina precedes the formation of blood vessels(Fruttiger, 2007; Stone and Dreher, 1987). PAX2 (+) perinatal astrocytes closely associatewith blood vessels (Chu et al., 2001). In addition, after completion of blood vesselformation, astrocytes lose their proangiogenic properties but continue maintain vascularhomeostasis by promoting the formation and maintenance of blood-retina barrier (Abbott,2002; Gardner et al., 1997; Janzer and Raff, 1987). We hypothesize that after hypoxia-induced blood vessel formation occurs, TLX repression of Pax2 is relieved by BMPs fromthe blood and then Pax2 continues its expression in astrocytes.
The BMPs have recently been implicated in the development and formation of glial scarsfollowing injury and in disease states throughout the central nervous system (Hall andMiller, 2004). The BMP-related SMADs interact with members of the ciliary neurotrophicfactor(CNTF)/leukemia inhibitory factor (Stayner et al.) pathways, known as the signaltransducers and transcription factors (STATs), via the FRAP-STAT pathway (Rajan andMcKay, 1998; Rajan et al., 2003). The STATs and SMADs form a complex with the p300/
Sehgal et al. Page 9
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

CBP proteins to increase transcription of the type III intermediate filament protein GFAP(Yanagisawa et al., 2001). Together these two pathways potentiate astrocyte induction(Nakashima et al., 1999). BMP7 has been shown to be upregulated following spinal cordinjury and to play a neuroprotective role in the central nervous system (Cox et al., 2004;Harvey et al., 2004; Setoguchi et al., 2001; Tsai et al., 2007). However, we hypothesize, thatwhile BMPs may play a role initially in neuroprotection, they may also initiate reactivegliosis and/or maintain glial scars that prevent regeneration of central nervous systemconnections.
Future studies will be focused on determining the role of BMP7 and SHH in both astrocytedevelopment as well as the in glial scar formation following injury and in disease states suchas glaucoma, diabetic retinopathy, and age-related macular degeneration.
Materials and MethodsMaterials used
Dulbecco’s Modified Eagle’s Medium (DMEM), cell dissociation solution (Sigma; St.Louis, MO), FBS, Penicillin/Streptomycin, Trypsin-EDTA, Phenol:Choroform, DTT,ProLong antifade mounting media with DAPI (Invitrogen; Carlsbad, CA), INFγ, mouse N-terminus SHH peptide, mouse BMP7 (R&D Systems; Minneapolis, MN), Hepes, L-Glutamine, NEAA (Cellgro; Herndon, VA), EC growth supplement, protease inhibitorcocktail, RNase free water, Heparin (Sigma; St. Louis, MO), PVDF membranes (Millipore;Bedford, MA), NaCl, Tris, NP-40, Sodium deoxycholate, EDTA, IGPEAL, Glycerol, SDS,L-Glutamine, Sodium Pyruvate, CellBind ™ –treated dishes and flasks (Fisher Scientific;Hanover Park, IL), Roche buffer M, FuGene HD Transfection Reagent (Roche;Indianapolis, IN), Nuclear extraction kit, EMSA kit (Panomics; Fremont, CA), ECLChemiluminescent substrate, Protein G plus beads (Pierce), Chemiluminescence films(Amersham Bio- sciences, UK), DC assay kit (Bio-Rad; Hercules, CA), Streptavidin agaroseresin (Thermo Scientific; Rockford, IL), Cyclopamine (Calbiochem), SHH, BMP7,Follistatin (R&D systems, Minneapolis, MN), Lucifearse Assay System, TNT QuickCoupled Transcription/Translation system (Promega, Madison, WI), pCMV-TLX neo6expression vector (Origene; Rockville, MD), mouse BMP7 and BMP7 function blockingserum (kind gift from Pamela J. Lein, Oregon Health and Science University, Portland,Oregon). Antibodies: Rabbit anti- PAX2 (Covance; 1μg/ml), rabbit anti-phospho PAX2(Zymed; 2μg/ml), rabbit anti acetyl-Histone H3 (Millipore 17-615, 4μg/ml ChIP), rabbitanti-SMAD1 (Zymed; 1μg/ml), rabbit anti- phospho SMAD1 (Millipore; 1μg/ml forwesterns; 10ug/ml for immunofluorescence), rabbit anti-phospho SMAD1 (Santa Cruzsc-12353; 4μg/ml for ChIP), rabbit anti-SMAD2 (Zymed; 2μg/ml), rabbit anti-TLX (Abcamab30942; 0.4μg/ml for westerns, 4μg/ml for ChIP), mouse anti-TLX (abcam 10ug/ml forimmunofluorescence), rabbit anti-GLI2 (Abcam; 1μg/ml for westerns, 10mg/ml forimmunofluorescence), rabbit anti-GLI2 (Santa Cruz sc-20290; 4μg/ml ChIP); rabbit anti-GLI3 (Abcam; 10ug/ml); rabbit anti-GLI1 (Cell Signaling; 1:100), mouse anti-β-tubulin(Sigma; 1μg/ml), mouse anti-α tubulin (Sigma; 0.5μg/ml), human anti-BMP7 (R&Dsystems; 2μg/ml for westerns; 10ug/ml for immunofluorescence), rabbit anti-GFAP (Dako;1:1000 for immunofluorescence; 2μg/ml for FACs); mouse anti-NG2 chondroitin sulfateproteoglycans (Fisher Scientific; 2μg/ml); ECL anti-rabbit HRP (1:5000; GE Healthcare,UK), anti-rabbit HRP for CO-IP (1:20000; e Bioscience, San Diego, CA), donkey anti-goatIgG conjugated to alexa-fluor 594 (Molecular Probes Invitrogen, 1:1000); goat anti-mouseIgG conjugated to alexa-fluor 594 (Molecular Probes Invitrogen, 1:1000), and goat anti-rabbit IgG conjugated to alexa-fluor 594 (Molecular Probes Invitrogen, 1:1000).
Sehgal et al. Page 10
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Experimental animalsAll animal experimentation described in this study was conducted in accordance withaccepted standards of humane animal care, the NIH guidelines for the care and use ofexperimental animals, and protocols approved by the institutional animal care and usecommittees for Indiana University School of Medicine and University of Wisconsin. Wild –type C3H mice and immorto-mice were housed under pathogen-free conditions in controlledlight cycle and temperatures, and provided tap water and commercial mouse chow adlibidum. Mating pairs were monitored daily for vaginal plugs.
Cell Culture and Gene Regulation AssaysRetinal astrocytes were isolated from “immorto” mice back-crossed to C57BL/6J and grownas previously described (Scheef et al., 2005) and were grown on CellBind™-treated dishesand flasks. For luciferase assays, a luciferase reporter gene containing 900 bp upstream ofthe start site of the mouse Pax2 promoter (including the TLX binding site) was constructedin the pGL4.14 vector (Promega). Astrocytes were cultured in 6×35mm dishes. 5×105
astrocytes were transiently co-transfected using Fugene HD transfection reagent with 300 ngof reporter plasmid and 200 ng of expression vector. Parallel control samples were co-transfected with equal amounts of empty vectors (n=6 for each condition). After 24 hours oftransfection, cells were treated with BMP7, SHH and, BMP7 & SHH. 24 hours later, cellswere lysed and luciferase activity was measured using Beckman Coulter LD400 platereader/luminometer (Beckman Coulter, Fullerton, CA). Total protein concentration wasdetermined using Bio-Rad DC assay and luciferase activities were normalized to proteinconcentration. Each experiment was repeated at least three times (n= 4+).
SDS-PAGE and Western AnalysisAstrocytes were seeded in 10 mm culture dishes at a concentration of 5×105 cells/ml. Theywere treated with BMP7 (200 ng/ml), SHH (200 ng/ml) either alone or in combination.After 12–16 hours of treatment, cells were lysed by adding lysis buffer (50mM Tris pH 7.4,1% NP-40, 0.25% sodium deoxycholate, 150mM sodium chloride, 1mM EDTA,supplemented with 1 μg/ml protease inhibitors) for 30 min on ice. Cell lysates werecentrifuged at 14000 rpm for 15 min at 4°C and supernatant was collected. Total proteinconcentration was determined using DC-Assay (Bio-Rad). Proteins were transferred toPVDF membranes overnight at 4°C and immunoblotted as described in (Nofziger et al.,2005). Densitometry of the blots was performed using ImageJ 1.34 s software. western datavalues for treated samples were compared with vehicle samples using one-tailed student’s t-test. Differences were considered significant when p ≤ 0.06. For total PAX2 proteindensitometric analysis, the PAX2 dimers were totaled together because previous reportshave shown no difference in the ability of the two alternatively spliced products to bind toDNA and activate expression (Phelps and Dressler, 1996)
Nuclear Extract PreparationNuclear extracts were prepared from wild type retinal astrocytes using a nuclear extractionkit according to the manufacturer’s instructions (Panomics, Fremont, CA). Cells were grownto 90% confluence and washed with PBS. Buffer A working reagent containing Buffer A(supplied with the kit), DTT, 10% IGEPAL and protease inhibitor cocktail was added to theculture flasks, kept on ice bucket and incubated for 10 minutes on rocking platform. Cellswere removed using a sterile scraper and transferred to 1.5ml microfuge tube andcentrifuged at 14,000 rpm for 3 min at 4°C. To the pellet, Buffer B working solutioncontaining Buffer B (supplied with the kit), DTT and protease inhibitor cocktail was added.Tubes were vortexed and incubated horizontally on an ice bucket which was then transferred
Sehgal et al. Page 11
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

to a rocking platform for 2 hours. Samples were centrifuged and supernatants weretransferred to a fresh microfuge tube. Nuclear extracts were stored at −80°C until analyzed.
Electrophoretic Mobility Shift Assay (EMSA) and In Vitro TranslationMobility shift assays were performed using EMSA kits (Panomics) according to themanufacturer’s instructions with the following additions or changes. Probes used for EMSAanalysis were as follows: Anti-sense probe (5′-3′):AGCTTTGTCTGACAAGTCATCCATCTAGCT; Sense probe (5′-3′):AGCTAGATGGATGACTTGTCAGACAAAGCT. 4 μg nuclear extract was incubated with5x binding buffer, Poly d (I–C) for 5 min at room temperature before adding 1 μl of biotinlabeled Pax2 probe (prepared by annealing sense and biotin labeled antisense oligos byboiling 100ng/μl of each oligo with 10x Roche buffer M for 5 minutes and kept at roomtemperature overnight). In vitro translations were performed as in Pfaeffle et al (2007).
Avidin Biotin Conjugated DNA Binding assay (ABCD)The ABCD assay was performed as in Glass et al. (1987). Briefly, sense and biotin labeledantisense oligos were first annealed by boiling 100 pmole/μl of each oligo with 10x Rochebuffer M for 5 minutes and kept at room temperature overnight. 10μl of the total sample wasused for each reaction. 500μg total proteins from astrocytes were mixed with 100μl ofprotease inhibitor cocktail, 10μl of probe and DNA pull down buffer pH 7.5 (containing25mM HEPES, 15mM NaCl, 0.5mM DTT, 0.5% IGEPAL, 0.1mM EDTA and 10%glycerol) was added to make total volume to 1ml in 1.5ml microfuge tubes. The solutionwas incubated on rocker shaker at 4°C overnight. 40 μl of 50% streptavidin-agarose resinwas added and was incubated for 6hours at 4°C on rocker shaker. Tubes were centrifuged at12000 rpm for 2 minutes and the pellet was then washed five times with DNA pull downbuffer. 1x SDS-PAGE loading buffer was added to the pellet and boiled for 5 minutes. Thesamples were stored at −20°C until further analyzed by western blotting.
Co-immunoprecipitationCo-immunoprecipitation was performed using the universal magnetic Co-IP kit from ActiveMotif with minor modifications to the manufacturer’s instructions. Briefly, nuclei wereisolated form cells using enzymatic digestion. Three hundred micrograms of nuclear proteinextract was incubated with 5μg of antibody specific for b-tubulin, pSMAD1, or GLI2overnight at 4°C on a rotator. Following incubation with antibody, lysates were incubatedwith magnetic protein G beads for 3–4 hours at 4°C on a rotator. After washing beads 6times in ice cold wash buffer, beads were suspended in 2X reducing loading buffer (130mmTris pH6.8, 4% SDS, 0.02% bromophenol blue, 20% glycerol, 100mmDTT), vortexed tomix and stored at −80°C until western blotting was performed. Prior to loading, beads wereboiled for 10 minutes, spun, and the elute was subjected to SDS-PAGE. Following transfer,blots were immunoblotted as in western analysis section.
ImmunofluorescencePrior to immunohistochemistry, antigen retrieval was performed by placing sections in10mM sodium citrate pH 6.0 at 65°C and heating in a microwave on high for 20 minutes.Sections were then allowed to come to room temperature prior to proceeding with theimmunofluorescence protocol. Immunofluorescence on sections was performed as describedin Wilson et al.(Wilson et al., 2007), with the exception that slides were mounted usingProLong Gold with DAPI. Immunocytochemistry was performed as described in Belecky-Adams et al (Belecky-Adams et al., 1996). Antibodies were used at the following dilutions:Pax2,1:200; TLX, 10μg/ml; GLI1, 1:100, Gli2, 10μg/ml, Gli3, 10μg/ml, SHH, 10μg/ml,BMP7, 10μg/ml, pSMAD1, 10μg/ml.
Sehgal et al. Page 12
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

FACScan analysisFACS analysis was performed as described in Scheef et al., 2005. Briefly, cells were rinsedwith phosphate buffered saline (PBS) with 0.4% EDTA. Cells were incubated with 2ml ofdissociation solution and rinsed from the plates with 5ml of DMEM containing 10% fetalbovine serum. Cells were spun, washed once one with tris-buffered saline (TBS),resuspended in 0.5ml of TBS with 1% goat serum, incubated for 20 minutes at 4°C,incubated with 0.5ml of rabbit anti GFAP (2μg/ml) or rabbit anti-NG2 (2μg/ml) for 30minutes at 4°C. FACS of labeled cells was compared to samples incubated with control IgGin place of primary antibodies.
Chromatin Immunoprecipitation and Quantitative PCRCells were rinsed with cold PBS, removed from dish with cell scraper, and crosslinkingcarried out in a final concentration of 1% formaldehyde for 15 minutes on ice. Cells weresubsequently incubated with glycine to stop the crosslinking and were washed twice byspinning at 4500RPM for 7 minutes and rinsing the pellet in PBS containing 1X proteaseinhibitor mix (Roche) and 0.1mM PMSF at 4°C. Following rinses, pellets were resuspendedin ChIP dilution buffer (0.01% SDS, 1.1% Triton X-100, 1.2mM EDTA, 16.7 mM Tris HClpH8.1, 167mM NaCl with 1X protease inhibitor mix and 0.1mM PMSF) and stored at−80°C. Upon thaw, cells were spun at 6000 RPM for 5 minutes and pellets resuspended in200mL of SDS lysis buffer (1% SDS, 10mM EDTA, 50mM Tris-HCl, pH8.1 with 1Xprotease inhibitor mix and 0.1mM PMSF) and incubated on ice for 10 minutes. The mixturewas sonicated for 3–4 sets of 10-second pulses using a Cole Palmer, High IntensityUltrasonic Processor/Sonicator, 50-watt model (with 2mm tip) set to 30% of maximumpower. Debris was removed by spinning at 13,000 RPM for 10 minutes at 4°C, supernatanttransferred to a fresh tube and 1.8ml of ChIP dilution buffer added. Two hundred microlitersof sample was saved as an input control. For immunoprecipitation, antibody was added at afinal concentration of 2μg/ml and rotated overnight at 4°C. After overnight incubation,samples were incubated with 60ml of protein A agarose, rotated for 1 hour at 4°C, andcentrifuged at 1000RPM for 1 min at 4°C. Beads were washed at room temperature for 5minutes on a rocker once in 1.0ml of low salt wash buffer (0.1% SDS, 1% Triton X-100,2mM EDTA, 20mM Tris HCl, pH8.1, 150mM NaCl), four times in 1.0ml high salt washbuffer ( 0.1% SDS, 1% Triton X-100, 2mM EDTA, 20mM Tris HCl, pH8.1, 500mM NaCl),once in LiCl wash buffer (0.25M LiCl, 1% NP-40, 1% deoxycholate, 1mM EDTA, 10mMTris HCl, pH8.1) and twice in TE. Samples were incubated in 250ul of freshly preparedelution buffer (1% SDS,0.1M NaHCO3), incubated at room temp. for 15 minutes, beadsspun out and supernatant transferred to a fresh tube. Elution was repeated with beads oncemore and eluates were combined. The sample and input control were incubated with 20ul of5.0M NaCl for 4 hours at 65°C, then incubated in 20μg of proteinase K. Samples weretreated with phenol chloroform and ethanol precipitated. Pellets were resuspended in 50μl ofTE and stored at −20°C until qPCR performed. qPCR was performed using Taqman PCRReagent kit and Applied Biosystems -7500 Real time PCR system according to themanufacturer’s instructions. Primer Express 3.0 by Applied Biosystems was used to designPrimers and Probes. Oligonucleotide Probe had 5′ FAM and 3′TAMRA reporter andquencher dyes respectively. GAPDH (used as endogenous control) and Pax2 primers usedwere: GAPDH Forward Primer; 5′-GAAGGTGAAGGTCGGAGTC-3′, GAPDH ReversePrimer; 5′-GAAGATGGTGATGGGATTTC-3′, GAPDH Probe FAM-CAAGCTTCCCGTTCTCAGCC-TAMRA, PAX2 Forward Primer;5′CGGCGCTGGCGAATC3′, PAX2 Reverse Primer;5′CGGGAGATGGATGACTTGTCA3′ Pax2 Probe; FAM-AGAGTGGTGGAATCTA-TAMRA. Each reaction was performed in triplicates and each experiment was performed atleast 3 times. The data was expressed as Log10 fold increase (mean±SD) above control.
Sehgal et al. Page 13
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

AcknowledgmentsThe authors would like to thank Pam Lein (Oregon Health and Science University, Portland, OR) for mouse BMP7and function-blocking BMP7 antibody and Greg Dressler (University of Michigan, Ann Arbor, MI) for sharing 9.1Kbp of 5′ untranslated mouse Pax2 sequences. The authors also gratefully acknowledge Rachel Mullen for helpwith immunostaining. TBA is supported by the March of Dimes, American Cancer Society, and American HealthAssistance Foundation, while SJR is supported by R01 HD42024 from the NIH/NICHD. NS is supported byEY016695 and Retina Research Foundation.
ReferencesAbbott NJ. Astrocyte-endothelial interactions and blood-brain barrier permeability. J Anat
2002;200:629–38. [PubMed: 12162730]Adler R, Belecky-Adams TL. The role of bone morphogenetic proteins in the differentiation of the
ventral optic cup. Development 2002;129:3161–71. [PubMed: 12070091]Arkell R, Beddington RS. BMP-7 influences pattern and growth of the developing hindbrain of mouse
embryos. Development 1997;124:1–12. [PubMed: 9006062]Bai CB, Stephen D, Joyner AL. All mouse ventral spinal cord patterning by hedgehog is Gli dependent
and involves an activator function of Gli3. Dev Cell 2004;6:103–15. [PubMed: 14723851]Belecky-Adams T, Adler R. Developmental expression patterns of bone morphogenetic proteins,
receptors, and binding proteins in the chick retina. J Comp Neurol 2001;430:562–72. [PubMed:11169487]
Belecky-Adams T, Cook B, Adler R. Correlations between terminal mitosis and differentiated fate ofretinal precursor cells in vivo and in vitro: analysis with the “window-labeling” technique.Developmental Biology 1996;178:304–15. [PubMed: 8812131]
Belecky-Adams TL, Adler R. Developmental expression patterns of bone morphogenetic proteins,receptors and binding proteins in the chick retina. J Comp Neurol. 2000 in press.
Bhatia M, Bonnet D, Wu D, Murdoch B, Wrana J, Gallacher L, Dick JE. Bone morphogenetic proteinsregulate the developmental program of human hematopoietic stem cells. J Exp Med1999;189:1139–48. [PubMed: 10190905]
Cai Y, Lechner MS, Nihalani D, Prindle MJ, Holzman LB, Dressler GR. Phosphorylation of Pax2 bythe c-Jun N-terminal kinase and enhanced Pax2-dependent transcription activation. J Biol Chem2002;277:1217–22. [PubMed: 11700324]
Chan-Ling T, Stone J. Factors determining the migration of astrocytes into the developing retina:migration does not depend on intact axons or patent vessels. J Comp Neurol 1991;303:375–86.[PubMed: 2007655]
Chari NS, McDonnell TJ. The sonic hedgehog signaling network in development and neoplasia. AdvAnat Pathol 2007;14:344–52. [PubMed: 17717435]
Chen D, Zhao M, Harris SE, Mi Z. Signal transduction and biological functions of bonemorphogenetic proteins. Front Biosci 2004;9:349–58. [PubMed: 14766372]
Chu Y, Hughes S, Chan-Ling T. Differentiation and migration of astrocyte precursor cells andastrocytes in human fetal retina: relevance to optic nerve coloboma. FASEB J 2001;15:2013–5.[PubMed: 11511521]
Cox S, Harvey BK, Sanchez JF, Wang JY, Wang Y. Mediation of BMP7 neuroprotection by MAPKand PKC IN rat primary cortical cultures. Brain Res 2004;1010:55–61. [PubMed: 15126117]
Dakubo GD, Beug ST, Mazerolle CJ, Thurig S, Wang Y, Wallace VA. Control of glial precursor celldevelopment in the mouse optic nerve by sonic hedgehog from retinal ganglion cells. Brain Res2008;1228:27–42. [PubMed: 18625210]
Dakubo GD, Wang YP, Mazerolle C, Campsall K, McMahon AP, Wallace VA. Retinal ganglion cell-derived sonic hedgehog signaling is required for optic disc and stalk neuroepithelial celldevelopment. Development 2003;130:2967–80. [PubMed: 12756179]
Dale JK, Vesque C, Lints TJ, Sampath TK, Furley A, Dodd J, Placzek M. Cooperation of BMP7 andSHH in the induction of forebrain ventral midline cells by prechordal mesoderm. Cell1997;90:257–69. [PubMed: 9244300]
Sehgal et al. Page 14
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Dale L, Jones CM. BMP signalling in early Xenopus development. Bioessays 1999;21:751–60.[PubMed: 10462415]
Derynck R, Zhang YE. Smad-dependent and Smad-independent pathways in TGF-beta familysignalling. Nature 2003;425:577–84. [PubMed: 14534577]
Dressler GR, Deutsch U, Chowdhury K, Nornes HO, Gruss P. Pax2, a new murine paired-box-containing gene and its expression in the developing excretory system. Development1990;109:787–95. [PubMed: 1977574]
Dressler GR, Douglass EC. Pax-2 is a DNA-binding protein expressed in embryonic kidney andWilms tumor. Proc Natl Acad Sci U S A 1992;89:1179–83. [PubMed: 1311084]
Dudley AT, Lyons KM, Robertson EJ. A requirement for bone morphogenetic protein-7 duringdevelopment of the mammalian kidney and eye. Genes Dev 1995;9:2795–807. [PubMed:7590254]
Dudley AT, Robertson EJ. Overlapping expression domains of bone morphogenetic protein familymembers potentially account for limited tissue defects in BMP7 deficient embryos. Dev Dyn1997;208:349–62. [PubMed: 9056639]
Eccles MR, Wallis LJ, Fidler AE, Spurr NK, Goodfellow PJ, Reeve AE. Expression of the PAX2 genein human fetal kidney and Wilms’ tumor. Cell Growth Differ 1992;3:279–89. [PubMed: 1378753]
Ekker SC, Ungar AR, Greenstein P, von Kessler DP, Porter JA, Moon RT, Beachy PA. Patterningactivities of vertebrate hedgehog proteins in the developing eye and brain. Curr Biol 1995;5:944–55. [PubMed: 7583153]
Fan X, Valdimarsdottir G, Larsson J, Brun A, Magnusson M, Jacobsen SE, ten Dijke P, Karlsson S.Transient disruption of autocrine TGF-beta signaling leads to enhanced survival and proliferationpotential in single primitive human hemopoietic progenitor cells. J Immunol 2002;168:755–62.[PubMed: 11777969]
Fruttiger M. Development of the retinal vasculature. Angiogenesis 2007;10:77–88. [PubMed:17322966]
Fuhrmann S, Levine EM, Reh TA. Extraocular mesenchyme patterns the optic vesicle during early eyedevelopment in the embryonic chick. Development - Supplement 2000;127:4599–609.
Furimsky M, Wallace VA. Complementary Gli activity mediates early patterning of the mouse visualsystem. Dev Dyn 2006;235:594–605. [PubMed: 16342201]
Furuta Y, Piston DW, Hogan BL. Bone morphogenetic proteins (BMPs) as regulators of dorsalforebrain development. Development 1997;124:2203–12. [PubMed: 9187146]
Gardner TW, Lieth E, Khin SA, Barber AJ, Bonsall DJ, Lesher T, Rice K, Brennan WA Jr. Astrocytesincrease barrier properties and ZO-1 expression in retinal vascular endothelial cells. InvestOphthalmol Vis Sci 1997;38:2423–7. [PubMed: 9344364]
Hall AK, Miller RH. Emerging roles for bone morphogenetic proteins in central nervous system glialbiology. J Neurosci Res 2004;76:1–8. [PubMed: 15048925]
Harvey BK, Mark A, Chou J, Chen GJ, Hoffer BJ, Wang Y. Neurotrophic effects of bonemorphogenetic protein-7 in a rat model of Parkinson’s disease. Brain Res 2004;1022:88–95.[PubMed: 15353217]
Herpin A, Cunningham C. Cross-talk between the bone morphogenetic protein pathway and othermajor signaling pathways results in tightly regulated cell-specific outcomes. FEBS J2007;274:2977–85. [PubMed: 17521337]
Hocking JC, McFarlane S. Expression of Bmp ligands and receptors in the developing Xenopus retina.Int J Dev Biol 2007;51:161–5. [PubMed: 17294367]
Hooper JE, Scott MP. Communicating with Hedgehogs. Nat Rev Mol Cell Biol 2005;6:306–17.[PubMed: 15803137]
Huangfu D, Anderson KV. Signaling from Smo to Ci/Gli: conservation and divergence of Hedgehogpathways from Drosophila to vertebrates. Development 2006;133:3–14. [PubMed: 16339192]
Hung FC, Zhao S, Chen Q, Overbeek PA. Retinal ablation and altered lens differentiation induced byocular overexpression of BMP7. Vision Res 2002;42:427–38. [PubMed: 11853758]
Janzer RC, Raff MC. Astrocytes induce blood-brain barrier properties in endothelial cells. Nature1987;325:253–7. [PubMed: 3543687]
Sehgal et al. Page 15
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Jat PS, Noble MD, Ataliotis P, Tanaka Y, Yannoutsos N, Larsen L, Kioussis D. Direct derivation ofconditionally immortal cell lines from an H-2Kb-tsA58 transgenic mouse. Proc Natl Acad Sci U SA 1991;88:5096–100. [PubMed: 1711218]
Jena N, Martin-Seisdedos C, McCue P, Croce CM. BMP7 null mutation in mice: developmentaldefects in skeleton, kidney, and eye. Exp Cell Res 1997;230:28–37. [PubMed: 9013703]
Kawai S, Sugiura T. Characterization of human bone morphogenetic protein (BMP)-4 and -7 genepromoters: activation of BMP promoters by Gli, a sonic hedgehog mediator. Bone 2001;29:54–61.[PubMed: 11472891]
Kim J, Johnson K, Chen HJ, Carroll S, Laughon A. Drosophila Mad binds to DNA and directlymediates activation of vestigial by Decapentaplegic. Nature 1997;388:304–8. [PubMed: 9230443]
Koshiba-Takeuchi K, Takeuchi JK, Matsumoto K, Momose T, Uno K, Hoepker V, Ogura K,Takahashi N, Nakamura H, Yasuda K, Ogura T. Tbx5 and the retinotectum projection. Science2000;287:134–7. [PubMed: 10615048]
Kusanagi K, Inoue H, Ishidou Y, Mishima HK, Kawabata M, Miyazono K. Characterization of a bonemorphogenetic protein-responsive Smad-binding element. Mol Biol Cell 2000;11:555–65.[PubMed: 10679014]
Lee KS, Kim HJ, Li QL, Chi XZ, Ueta C, Komori T, Wozney JM, Kim EG, Choi JY, Ryoo HM, BaeSC. Runx2 is a common target of transforming growth factor beta1 and bone morphogeneticprotein 2, and cooperation between Runx2 and Smad5 induces osteoblast-specific gene expressionin the pluripotent mesenchymal precursor cell line C2C12. Mol Cell Biol 2000;20:8783–92.[PubMed: 11073979]
Liem KF Jr, Tremml G, Roelink H, Jessell TM. Dorsal differentiation of neural plate cells induced byBMP-mediated signals from epidermal ectoderm. Cell 1995;82:969–79. [PubMed: 7553857]
Macdonald R, Barth KA, Xu Q, Holder N, Mikkola I, Wilson SW. Midline signalling is required forPax gene regulation and patterning of the eyes. Development 1995;121:3267–78. [PubMed:7588061]
Miyawaki T, Uemura A, Dezawa M, Yu RT, Ide C, Nishikawa S, Honda Y, Tanabe Y, Tanabe T. Tlx,an orphan nuclear receptor, regulates cell numbers and astrocyte development in the developingretina. J Neurosci 2004;24:8124–34. [PubMed: 15371513]
Miyazono K. Signal transduction by bone morphogenetic protein receptors: functional roles of Smadproteins. Bone 1999;25:91–3. [PubMed: 10423029]
Miyazono K. Positive and negative regulation of TGF-beta signaling. J Cell Sci 2000a;113(Pt 7):1101–9. [PubMed: 10704361]
Miyazono K. TGF-beta signaling by Smad proteins. Cytokine Growth Factor Rev 2000b;11:15–22.[PubMed: 10708949]
Miyazono K, Maeda S, Imamura T. BMP receptor signaling: transcriptional targets, regulation ofsignals, and signaling cross-talk. Cytokine Growth Factor Rev 2005;16:251–63. [PubMed:15871923]
Morcillo J, Martinez-Morales JR, Trousse F, Fermin Y, Sowden JC, Bovolenta P. Proper patterning ofthe optic fissure requires the sequential activity of BMP7 and SHH. Development 2006;133:3179–90. [PubMed: 16854970]
Motoyama J, Milenkovic L, Iwama M, Shikata Y, Scott MP, Hui CC. Differential requirement for Gli2and Gli3 in ventral neural cell fate specification. Dev Biol 2003;259:150–61. [PubMed: 12812795]
Muller F, Rohrer H, Vogel-Hopker A. Bone morphogenetic proteins specify the retinal pigmentepithelium in the chick embryo. Development 2007;134:3483–93. [PubMed: 17728349]
Nagase T, Nagase M, Yoshimura K, Fujita T, Koshima I. Angiogenesis within the developing mouseneural tube is dependent on sonic hedgehog signaling: possible roles of motor neurons. GenesCells 2005;10:595–604. [PubMed: 15938717]
Nakamura H. Regionalization of the optic tectum: combinations of gene expression that define thetectum. Trends Neurosci 2001;24:32–9. [PubMed: 11163885]
Nakashima K, Yanagisawa M, Arakawa H, Kimura N, Hisatsune T, Kawabata M, Miyazono K, TagaT. Synergistic signaling in fetal brain by STAT3-Smad1 complex bridged by p300. Science1999;284:479–82. [PubMed: 10205054]
Sehgal et al. Page 16
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Nofziger C, Chen L, Shane MA, Smith CD, Brown KK, Blazer-Yost BL. PPARgamma agonists do notdirectly enhance basal or insulin-stimulated Na(+) transport via the epithelial Na(+) channel.Pflugers Arch 2005;451:445–53. [PubMed: 16170524]
Nornes HO, Dressler GR, Knapik EW, Deutsch U, Gruss P. Spatially and temporally restrictedexpression of Pax2 during murine neurogenesis. Development 1990;109:797–809. [PubMed:1977575]
Ohkubo Y, Chiang C, Rubenstein JL. Coordinate regulation and synergistic actions of BMP4, SHHand FGF8 in the rostral prosencephalon regulate morphogenesis of the telencephalic and opticvesicles. Neuroscience 2002;111:1–17. [PubMed: 11955708]
Parkin CA, Ingham PW. The adventures of Sonic Hedgehog in development and repair. I. Hedgehogsignaling in gastrointestinal development and disease. Am J Physiol Gastrointest Liver Physiol2008;294:G363–7. [PubMed: 18063705]
Phelps DE, Dressler GR. Identification of novel Pax-2 binding sites by chromatin precipitation. J BiolChem 1996;271:7978–85. [PubMed: 8626478]
Placzek M, Briscoe J. The floor plate: multiple cells, multiple signals. Nat Rev Neurosci 2005;6:230–40. [PubMed: 15738958]
Rajan P, McKay RD. Multiple routes to astrocytic differentiation in the CNS. J Neurosci1998;18:3620–9. [PubMed: 9570793]
Rajan P, Panchision DM, Newell LF, McKay RD. BMPs signal alternately through a SMAD or FRAP-STAT pathway to regulate fate choice in CNS stem cells. J Cell Biol 2003;161:911–21. [PubMed:12796477]
Roy K, Thiels E, Monaghan AP. Loss of the tailless gene affects forebrain development and emotionalbehavior. Physiol Behav 2002;77:595–600. [PubMed: 12527005]
Sakuta H, Suzuki R, Takahashi H, Kato A, Shintani T, Iemura S, Yamamoto TS, Ueno N, Noda M.Ventroptin: a BMP-4 antagonist expressed in a double-gradient pattern in the retina. Science2001;293:111–5. [PubMed: 11441185]
Scheef E, Wang S, Sorenson CM, Sheibani N. Isolation and characterization of murine retinalastrocytes. Mol Vis 2005;11:613–24. [PubMed: 16148882]
Schimmenti LA, de la Cruz J, Lewis RA, Karkera JD, Manligas GS, Roessler E, Muenke M. Novelmutation in sonic hedgehog in non-syndromic colobomatous microphthalmia. Am J Med Genet A2003;116A:215–21. [PubMed: 12503095]
Sehgal R, Karcavich R, Carlson S, Belecky-Adams TL. Ectopic Pax2 expression in chick ventral opticcup phenocopies loss of Pax2 expression. Dev Biol 2008;319:23–33. [PubMed: 18485342]
Setoguchi T, Yone K, Matsuoka E, Takenouchi H, Nakashima K, Sakou T, Komiya S, Izumo S.Traumatic injury-induced BMP7 expression in the adult rat spinal cord. Brain Res 2001;921:219–25. [PubMed: 11720729]
Shim S, Bae N, Han JK. Bone morphogenetic protein-4-induced activation of Xretpos is mediated bySmads and Olf-1/EBF associated zinc finger (OAZ). Nucleic Acids Res 2002;30:3107–17.[PubMed: 12136093]
Stallcup WB, Beasley L. Bipotential glial precursor cells of the optic nerve express the NG2proteoglycan. J Neurosci 1987;7:2737–44. [PubMed: 3305800]
Stayner CK, Cunliffe HE, Ward TA, Eccles MR. Cloning and characterization of the human PAX2promoter. J Biol Chem 1998;273:25472–9. [PubMed: 9738017]
Stone J, Dreher Z. Relationship between astrocytes, ganglion cells and vasculature of the retina. JComp Neurol 1987;255:35–49. [PubMed: 3819008]
Suzuki T, Takeuchi J, Koshiba-Takeuchi K, Ogura T. Tbx Genes Specify Posterior Digit Identitythrough Shh and BMP Signaling. Dev Cell 2004;6:43–53. [PubMed: 14723846]
Trousse F, Esteve P, Bovolenta P. Bmp4 mediates apoptotic cell death in the developing chick eye. JNeurosci 2001;21:1292–301. [PubMed: 11160400]
Tsai MJ, Weng CF, Shyue SK, Liou DY, Chen CH, Chiu CW, Yang TH, Pan HA, Liao RI, Kuo HS,Huang MC, Huang WC, Hoffer BJ, Cheng H. Dual effect of adenovirus-mediated transfer ofBMP7 in mixed neuron-glial cultures: neuroprotection and cellular differentiation. J Neurosci Res2007;85:2950–9. [PubMed: 17628501]
Sehgal et al. Page 17
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Uemura A, Kusuhara S, Wiegand SJ, Yu RT, Nishikawa S. Tlx acts as a proangiogenic switch byregulating extracellular assembly of fibronectin matrices in retinal astrocytes. J Clin Invest2006;116:369–77. [PubMed: 16424942]
Ulloa F, Briscoe J. Morphogens and the control of cell proliferation and patterning in the spinal cord.Cell Cycle 2007;6:2640–9. [PubMed: 17912034]
Vogel-Hopker A, Momose T, Rohrer H, Yasuda K, Ishihara L, Rapaport DH. Multiple functions offibroblast growth factor-8 (FGF-8) in chick eye development. Mech Dev 2000;94:25–36.[PubMed: 10842056]
Wallace VA, Raff MC. A role for Sonic hedgehog in axon-to-astrocyte signalling in the rodent opticnerve. Development 1999;126:2901–9. [PubMed: 10357934]
Wang Y, Dakubo GD, Thurig S, Mazerolle CJ, Wallace VA. Retinal ganglion cell-derived sonichedgehog locally controls proliferation and the timing of RGC development in the embryonicmouse retina. Development 2005;132:5103–13. [PubMed: 16236765]
Wawersik S, Purcell P, Rauchman M, Dudley AT, Robertson EJ, Maas R. BMP7 acts in murine lensplacode development. Dev Biol 1999;207:176–88. [PubMed: 10049573]
Wilson JM, Sato K, Chernoff EA, Belecky-Adams TL. Expression patterns of chick Musashi-1 in thedeveloping nervous system. Gene Expr Patterns 2007;7:817–25. [PubMed: 17544341]
Wordinger RJ, Agarwal R, Talati M, Fuller J, Lambert W, Clark AF. Expression of bonemorphogenetic proteins (BMP), BMP receptors, and BMP associated proteins in human trabecularmeshwork and optic nerve head cells and tissues. Mol Vis 2002;8:241–50. [PubMed: 12131877]
Yanagisawa M, Nakashima K, Takizawa T, Ochiai W, Arakawa H, Taga T. Signaling crosstalkunderlying synergistic induction of astrocyte differentiation by BMPs and IL-6 family ofcytokines. FEBS Lett 2001;489:139–43. [PubMed: 11165238]
Yokoyama A, Sakamoto A, Kameda K, Imai Y, Tanaka J. NG2 proteoglycan-expressing microglia asmultipotent neural progenitors in normal and pathologic brains. Glia 2006;53:754–68. [PubMed:16534776]
Yu RT, Chiang MY, Tanabe T, Kobayashi M, Yasuda K, Evans RM, Umesono K. The orphan nuclearreceptor Tlx regulates Pax2 and is essential for vision. Proc Natl Acad Sci U S A 2000;97:2621–5.[PubMed: 10706625]
Yu RT, McKeown M, Evans RM, Umesono K. Relationship between Drosophila gap gene tailless anda vertebrate nuclear receptor Tlx. Nature 1994;370:375–9. [PubMed: 8047143]
Yuasa T, Kataoka H, Kinto N, Iwamoto M, Enomoto-Iwamoto M, Iemura S, Ueno N, Shibata Y,Kurosawa H, Yamaguchi A. Sonic hedgehog is involved in osteoblast differentiation bycooperating with BMP-2. J Cell Physiol 2002;193:225–32. [PubMed: 12385000]
Zhang Y, Derynck R. Transcriptional regulation of the transforming growth factor-beta -induciblemouse germ line Ig alpha constant region gene by functional cooperation of Smad, CREB, andAML family members. J Biol Chem 2000;275:16979–85. [PubMed: 10748032]
Zhang Y, Stone J. Role of astrocytes in the control of developing retinal vessels. Invest OphthalmolVis Sci 1997;38:1653–66. [PubMed: 9286254]
Zhang Y, Zhang Z, Zhao X, Yu X, Hu Y, Geronimo B, Fromm SH, Chen YP. A new function ofBMP4: dual role for BMP4 in regulation of Sonic hedgehog expression in the mouse tooth germ.Development 2000;127:1431–43. [PubMed: 10704389]
Zhao S, Chen Q, Hung FC, Overbeek PA. BMP signaling is required for development of the ciliarybody. Development 2002;129:4435–42. [PubMed: 12223402]
Zhao S, Hung FC, Colvin JS, White A, Dai W, Lovicu FJ, Ornitz DM, Overbeek PA. Patterning theoptic neuroepithelium by FGF signaling and Ras activation. Development 2001;128:5051–60.[PubMed: 11748141]
Sehgal et al. Page 18
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1. GLI2, GLI3, and pSMAD1 are expressed in E9.5 optic vesicle prior to PAX2Ten micron cryosections through E9.5 optic vesicle were co-labeled with DAPI andantibodies against PAX2 (A–C), GLI2 (D–F), GLI3 (G–I), or pSMAD1 (pSMAD1; J–L).Panels A–L show Nomarski and corresponding fluorescent images for DAPI and antibodylabel. (A–C) PAX2 was not detectable in the optic vesicle at E9.5. (D–F) GLI2-expressingcells were found primarily at the ventricular edge of the optic vesicle (arrowheads, F). (G–I)GLI3 was detected throughout the optic vesicle (arrowheads, I) and in the presumptive lensplacode and a subpopulation of mesenchymal cells. (J–L) Like GLI2, pSMAD1 wasdetected primarily in cells lining the ventricular edge of the developing optic vesicle(arrows, L). Magnification Bar in A = 50μm; applies to A–L.
Sehgal et al. Page 19
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 2. Expression of PAX2, GLI2, GLI3, and pSMAD1 in E10.5 optic cupTen micron cryosections through E10.5 optic cup were co-labeled with DAPI and antibodiesagainst PAX2 (A–D), GLI2 (E–H), GLI3 (I–L), or pSMAD1 (M–P).Panels (A–T) show Nomarski and corresponding fluorescent images for DAPI and antibodylabel. (A–D) As shown by arrowheads in D, PAX2 was localized to the ventral optic cupand optic stalk at E10.5. (E–H) Similar to localization at E9.5, stronger GLI2 signals werefound at the ventricular edge of the optic stalk (arrowheads, H), and weaker signals weredetected throughout the rest of the optic stalk, optic cup, and lens. (I–L) GLI3 was foundthroughout the optic stalk, retinal pigmented epithelium, and the lens placode. (M–P).Phospho-SMAD1 localization was similar to that noted for GLI2, with higher signal
Sehgal et al. Page 20
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

detected in cells at the ventricular edge (arrowheads, p) and weaker signals noted throughoutthe rest of the optic stalk, optic cup, and lens. (Q–T) Negative controls in which primaryantibody was omitted show no background label in the optic stalk, optic cup, and lens.Magnification Bar in A = 50μm and applies to photomicrographs in A–C, E–G, I–K, M–O,and Q–S. Bar in D= 50μm and applies to photomicrographs in D, H, L, P, and T.Abbreviations: DOS; dorsal optic stalk, VOS; ventral optic stalk, RPE; retinal pigmentedepithelium, R; retina, LP; lens placode, L, lens.
Sehgal et al. Page 21
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 3. Expression of PAX2, GLI2, GLI3, and pSMAD1 in E16 optic nerveTen micron cryosections through E16 optic nerve head and nerve were co-labeled withDAPI and antibodies against PAX2 (A–C), GLI2 (D–F), GLI3 (G–I), or pSMAD1 (J–L).Panels (A–O) show Nomarski and corresponding fluorescent images for DAPI and antibodylabel. (A–C) PAX2 was detected in nuclei of the optic nerve head (ONH) and optic nerve(ON) at E16 (arrowheads, B & C). GLI2 (arrowheads E & F), GLI3 (arrows, H & I) andpSMAD1 (arrowheads, K & L) were also detected in the nuclei of cells in the ONH and ON.Negative controls in which the primary antibody was omitted are shown in M–O.Magnification Bar in A = 50μm and applies to photomicrographs in A–O.
Sehgal et al. Page 22
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Abbreviations: RPE; retinal pigmented epithelium, ON; optic nerve, ONH; optic nerve head,R; retina.
Sehgal et al. Page 23
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
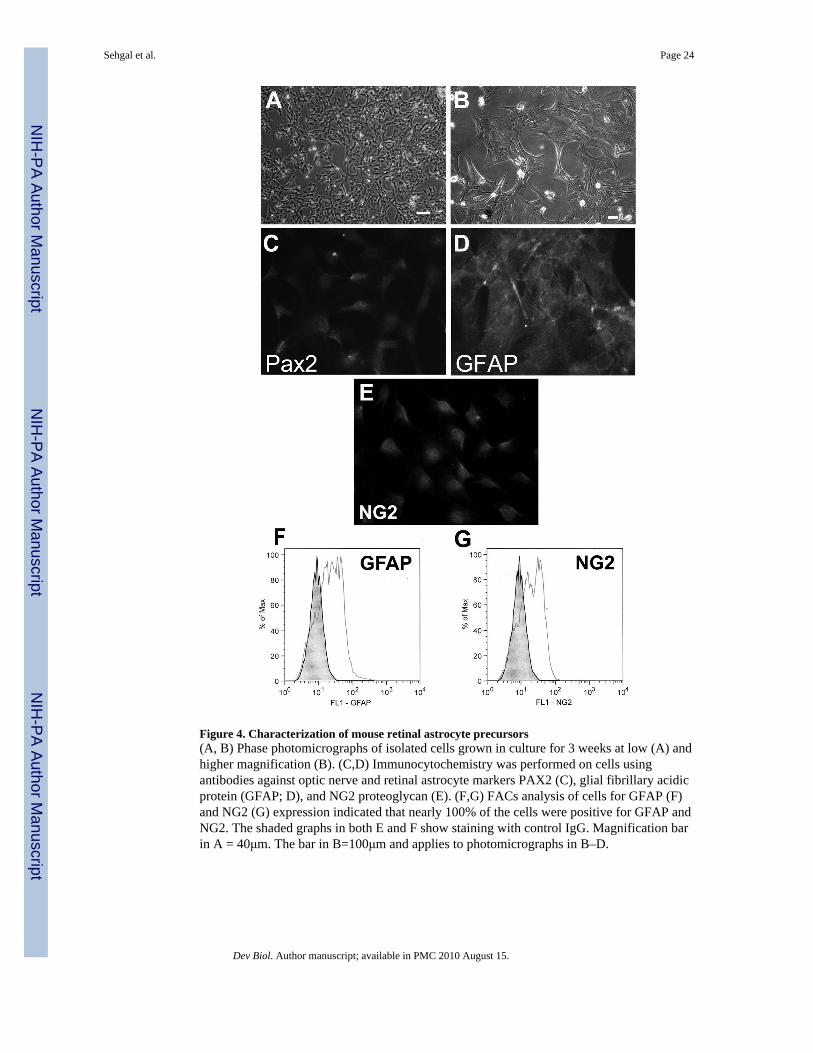
Figure 4. Characterization of mouse retinal astrocyte precursors(A, B) Phase photomicrographs of isolated cells grown in culture for 3 weeks at low (A) andhigher magnification (B). (C,D) Immunocytochemistry was performed on cells usingantibodies against optic nerve and retinal astrocyte markers PAX2 (C), glial fibrillary acidicprotein (GFAP; D), and NG2 proteoglycan (E). (F,G) FACs analysis of cells for GFAP (F)and NG2 (G) expression indicated that nearly 100% of the cells were positive for GFAP andNG2. The shaded graphs in both E and F show staining with control IgG. Magnification barin A = 40μm. The bar in B=100μm and applies to photomicrographs in B–D.
Sehgal et al. Page 24
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 5. SHH and BMP7 promote increased levels of PAX2 and phospho PAX2 in retinalastrocytes(A) Expression of PAX2 (a–d) and GFAP (e–h) in retinal astrocytes treated with controlvehicle (a,e), BMP7 (b,f), SHH (c,g) and BMP7 +SHH (d,h). Digital photomicrographs weretaken at the same exposure and are unprocessed. Note the increase in PAX2immunoreactivity of treated cells (b, c, d) in comparison to vehicle-treated cells (a). (B)PAX2 levels were measured by western blotting after treatment of retinal astrocytes withBMP7, SHH, or BMP7 and SHH (n=3). Note the increase in the amount of PAX2 in isolatesof treated cells in comparison to control. (B′) Densitometric analysis of experiments shownin B normalized to the β-tubulin loading control. Values shown above histogram barsindicate the level of statistical significance for each treatment in comparison to controls. (C)Phospho-PAX2 levels were assayed by western blotting in astrocytes treated with BMP7,SHH, or BMP7 and SHH (n=3). (C′) Densitometric analysis of immunoblot shown in Cnormalized to β-tubulin. Values shown above histogram bars indicate the level of statisticalsignificance for each treatment in comparison to controls. Magnification bar in a = 50μmand applies to photomicrographs in a–h.
Sehgal et al. Page 25
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
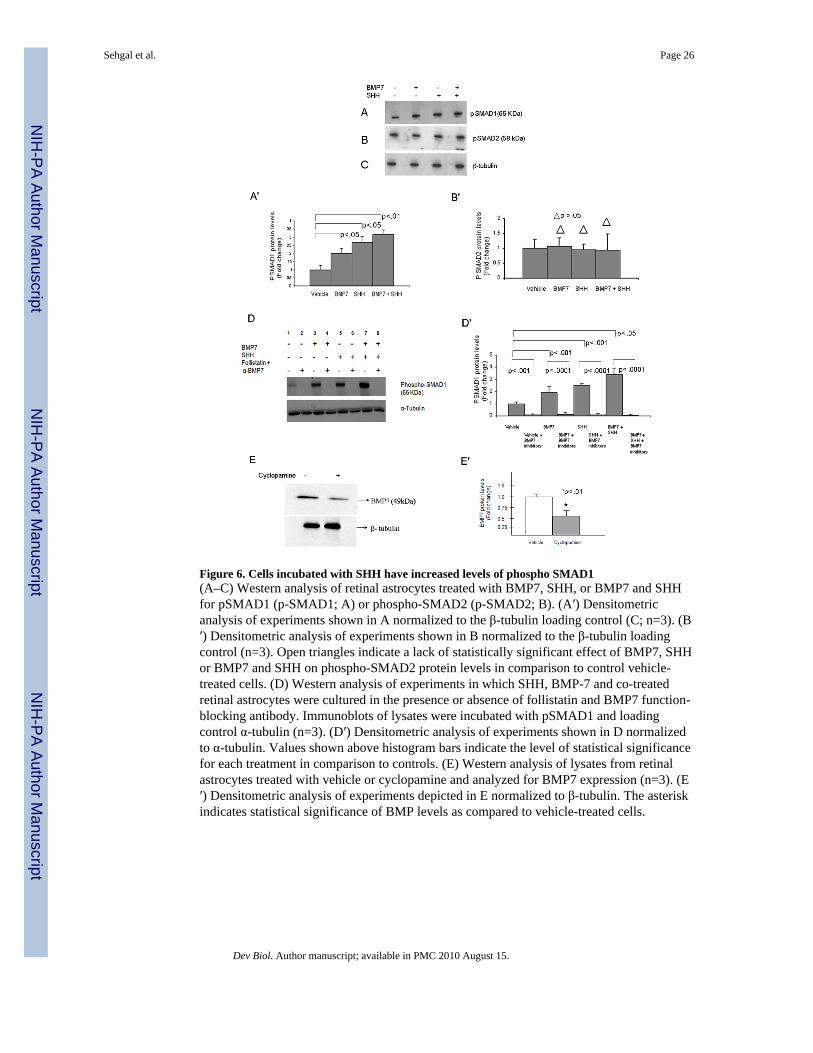
Figure 6. Cells incubated with SHH have increased levels of phospho SMAD1(A–C) Western analysis of retinal astrocytes treated with BMP7, SHH, or BMP7 and SHHfor pSMAD1 (p-SMAD1; A) or phospho-SMAD2 (p-SMAD2; B). (A′) Densitometricanalysis of experiments shown in A normalized to the β-tubulin loading control (C; n=3). (B′) Densitometric analysis of experiments shown in B normalized to the β-tubulin loadingcontrol (n=3). Open triangles indicate a lack of statistically significant effect of BMP7, SHHor BMP7 and SHH on phospho-SMAD2 protein levels in comparison to control vehicle-treated cells. (D) Western analysis of experiments in which SHH, BMP-7 and co-treatedretinal astrocytes were cultured in the presence or absence of follistatin and BMP7 function-blocking antibody. Immunoblots of lysates were incubated with pSMAD1 and loadingcontrol α-tubulin (n=3). (D′) Densitometric analysis of experiments shown in D normalizedto α-tubulin. Values shown above histogram bars indicate the level of statistical significancefor each treatment in comparison to controls. (E) Western analysis of lysates from retinalastrocytes treated with vehicle or cyclopamine and analyzed for BMP7 expression (n=3). (E′) Densitometric analysis of experiments depicted in E normalized to β-tubulin. The asteriskindicates statistical significance of BMP levels as compared to vehicle-treated cells.
Sehgal et al. Page 26
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 7. BMP7 and SHH decrease the binding of TLX protein on Pax2 promoter(A) Electrophoretic mobility shift assay (EMSA) performed using nuclear extracts fromretinal astrocytes treated with vehicle, BMP7, SHH, or BMP + SHH (n=4). Biotinylatedoligonucleotide representing the TLX binding site (AAGTCA) that lies ~80bp upstream ofPax2 start site was used to probe nuclear extracts. Note the decrease in TLX bound in cellsco-treated with BMP7 and SHH. (B) EMSA performed in which in vitro translated TLXprotein was incubated with labeled oligo representing the TLX-binding site in the Pax2promoter (lane 2), nuclear extracts from untreated astrocytes were incubated withbiotinylated oligo (lane 3), or nuclear extracts from untreated retinal astrocytes wasincubated with labeled oligo in the presence of TLX antibody (Lane 4). Lane 4 shows
Sehgal et al. Page 27
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

supershift in TLX band in presence of TLX antibody. No shift or supershift was detected insamples incubated without labeled oligo or TLX antibody (lane1). (C) Quantitation ofEMSA represented in A. Values above the histogram bars indicate the statistical significanceof each treatment in comparison to control vehicle-treated cells. (D) Quantitativepolymerase chain reaction (qPCR) of a chromatin immunoprecipitation (ChIP) assay usingextracts from retinal astrocytes treated with vehicle, BMP7, SHH, or BMP + SHH (n=6).TLX antibody was used to precipitate DNA/protein complexes and primers to the TLXbinding site within the Pax2 promoter used to amplify with Taqman qPCR. As a positivecontrol, antibody against acetyl-Histone H3 was used to precipitate protein/DNA complexesand primers for Glyceraldehyde-3-phosphate (GAPDH) used for amplification (first bar).The values for the acetyl-histone H3 were automatically set to a value of 1. GAPDH wasused as an internal control for all experiments and values shown in D are normalized toGAPDH. Values for the rabbit IgG negative control were subtracted from experimentalvalues prior to plotting. Compare the decrease in the amount of Pax2 amplified in D as aresult of growth factor treatment with the decrease in the amount of TLX bound in EMSAs(C). E) Co-immunoprecipitation (Co-Ip) was performed by incubating nuclear lysates withantibody against control β-tubulin (Lane1), pSMAD1 (Lane 2) or GLI2 (Lane 3) andanalyzed by western blotting for TLX (a), pSMAD1 (b), or GLI2 (c). (c). Lane 2 and 3shows binding of TLX protein with pSMAD1 and GLI2 respectively. (b) Lane 2 acts as apositive control to show that pSMAD1 was immunoprecipitated by pSMAD1 antibody.Lane 3 shows presence of pSMAD1 signal in Co-Ip with GLI2. (c) Lane 2 confirms theinteraction of GLI2 with pSMAD1. Lane3 acts as a positive control. (F) TLX protein levelsin astrocytes treated overnight with BMP7, SHH and, BMP7 and SHH. Co-Ip experimentswere all performed a total of 3 times. (F′) Densitometric analysis of experiments depicted inF normalized to β-tubulin. Statistical significance is indicated. (G) Avidin–biotin complexDNA binding assay (ABCD assay) was used to determine if pSMAD1 or GLI2 werebinding at or near the TLX binding site. Biotinylated oligonucleotide representing the TLXbinding site in the proximal Pax2 promoter was incubated with nuclear lysates fromuntreated retinal astrocytes. No signal was present in samples incubated without labeledprobe (lane1). TLX was the only protein detected when the TLX binding site was present(lane2).
Sehgal et al. Page 28
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
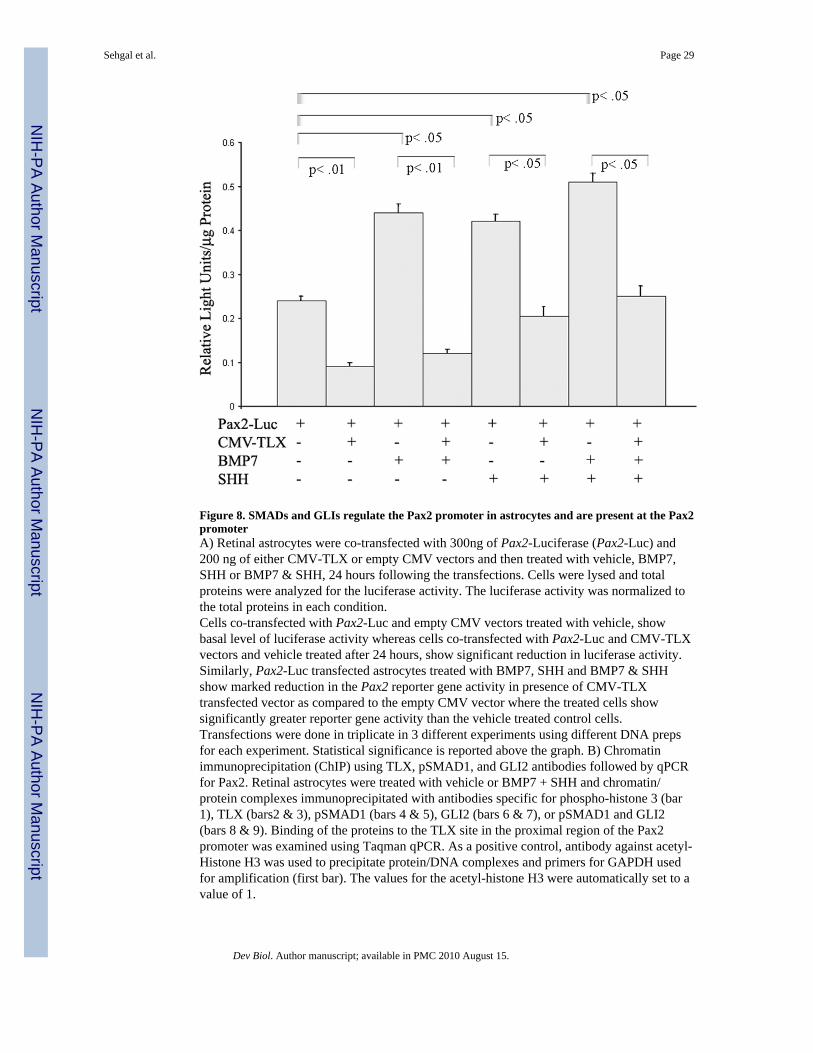
Figure 8. SMADs and GLIs regulate the Pax2 promoter in astrocytes and are present at the Pax2promoterA) Retinal astrocytes were co-transfected with 300ng of Pax2-Luciferase (Pax2-Luc) and200 ng of either CMV-TLX or empty CMV vectors and then treated with vehicle, BMP7,SHH or BMP7 & SHH, 24 hours following the transfections. Cells were lysed and totalproteins were analyzed for the luciferase activity. The luciferase activity was normalized tothe total proteins in each condition.Cells co-transfected with Pax2-Luc and empty CMV vectors treated with vehicle, showbasal level of luciferase activity whereas cells co-transfected with Pax2-Luc and CMV-TLXvectors and vehicle treated after 24 hours, show significant reduction in luciferase activity.Similarly, Pax2-Luc transfected astrocytes treated with BMP7, SHH and BMP7 & SHHshow marked reduction in the Pax2 reporter gene activity in presence of CMV-TLXtransfected vector as compared to the empty CMV vector where the treated cells showsignificantly greater reporter gene activity than the vehicle treated control cells.Transfections were done in triplicate in 3 different experiments using different DNA prepsfor each experiment. Statistical significance is reported above the graph. B) Chromatinimmunoprecipitation (ChIP) using TLX, pSMAD1, and GLI2 antibodies followed by qPCRfor Pax2. Retinal astrocytes were treated with vehicle or BMP7 + SHH and chromatin/protein complexes immunoprecipitated with antibodies specific for phospho-histone 3 (bar1), TLX (bars2 & 3), pSMAD1 (bars 4 & 5), GLI2 (bars 6 & 7), or pSMAD1 and GLI2(bars 8 & 9). Binding of the proteins to the TLX site in the proximal region of the Pax2promoter was examined using Taqman qPCR. As a positive control, antibody against acetyl-Histone H3 was used to precipitate protein/DNA complexes and primers for GAPDH usedfor amplification (first bar). The values for the acetyl-histone H3 were automatically set to avalue of 1.
Sehgal et al. Page 29
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

GAPDH was used as an internal control for all experiments and values shown in B arenormalized to GAPDH. Values for the rabbit IgG negative control were subtracted fromexperimental values prior to plotting.
Sehgal et al. Page 30
Dev Biol. Author manuscript; available in PMC 2010 August 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript




















