
Bis(4-hydroxy-2H-chromen-2-one): Synthesis and effects on leukemic cell lines proliferation and...
8
Bis(4-hydroxy-2H-chromen-2-one): Synthesis and effects on leukemic cell lines proliferation and NF-jB regulation Oualid Talhi a,1 , Michael Schnekenburger b,1 , Jana Panning b,c , Diana G. C. Pinto a , José A. Fernandes d , Filipe A. Almeida Paz d , Claus Jacob c , Marc Diederich e,⇑ , Artur M. S. Silva a,⇑ a QOPNA, Department of Chemistry, University of Aveiro, 3810-193 Aveiro, Portugal b Laboratoire de Biologie Moléculaire et Cellulaire du Cancer, Hôpital Kirchberg, L-2540 Luxembourg, Luxembourg c School of Pharmacy, Building B 2.1., Room 1.13, University of Saarland, Campus, 66123 Saarbrucken, Germany d CICECO, Department of Chemistry, University of Aveiro, 3810-193 Aveiro, Portugal e College of Pharmacy, Seoul National University, 599 Gwanak-ro, Gwanak-gu, Seoul 151-742, Republic of Korea article info Article history: Received 11 February 2014 Revised 27 March 2014 Accepted 30 March 2014 Available online 6 April 2014 Keywords: Biscoumarins 2D-NMR X-ray crystallography Leukaemia cell lines NF-jB inhibition abstract Synthesis of the bis-4-hydroxycoumarin-type compound, 3,3 0 -[3-(2-hydroxyphenyl)-3-oxopropane-1,1- diyl]bis(4-hydroxy-2H-chromen-2-one), was performed by two alternative pathways, either involving a basic organocatalyzed 1,4-conjugate addition tandem reaction of 4-hydroxycoumarin on chromone-3- carboxylic acid, or a double condensation of 4-hydroxycoumarin on x-formyl-2 0 -hydroxyacetophenone. The anti-proliferative effects of the bis-4-hydroxycoumarin-type compound on human K-562 (chronic myeloid leukaemia) and JURKAT (acute T-cell leukaemia) cell lines using trypan blue staining, as well as its involvement in nuclear factor-kappa B (NF-jB) regulation analyzed by luciferase reporter gene assay, gene expression analysis and western blots were analysed. This compound inhibited TNFa-induced NF-jB activation in K-562 (IC 50 17.5 lM) and JURKAT (IC 50 19.0 lM) cell lines, after 8 h of incubation. Interestingly, it exerted mainly cytostatic effects at low doses on both cell lines tested, whereas it decreased JURKAT cell viability starting at 50 lM from 24 h of treatment. Importantly, it did not affect the viability of peripheral blood mononuclear cells (PBMCs) from healthy donors, even at concentrations above 100 lM. Ó 2014 Elsevier Ltd. All rights reserved. 1. Introduction Cancer is characterized by uncontrolled growth and spreading of abnormal cells. Smoking, alcohol intake, obesity and physical inac- tivity are seen as main cancer risk factors. Also genetic predisposi- tion, inflammatory environment, activation of oncogenes and deactivation of tumour suppressor genes contribute to development of this disease. 1–3 A central molecular mechanism involved in inflammatory and innate immune response is the transcription fac- tor nuclear factor kappa-B cell (NF-jB) comprising five members including, relA (p65), relB, cRel, NF-jB1 (p50) and NF-jB2 (p52). Canonical NF-jB pathway is activated by Toll-like-, tumour necrosis factor (TNF-a) or T-cell receptors. RelA-p50 dimers are associated with inhibitor of jB (IjB) retaining them in an inactive form in the cytoplasm. After stimulation of cell surface receptors with pro- inflammatory cytokines, IjB kinase (IKK) is activated and phosphor- ylates IjB. As a consequence IjB is ubiquitinated and subsequently degraded by the proteasome. The released NF-jB transcription fac- tor then translocates to the nucleus and transactivates target genes involved in cancer initiation, promotion and progression. 4–6 A large majority (79.8%) of anticancer drugs developed between 1981 and 2010 derive from natural origins. 7 The flavonoid family is a prominent group of natural compounds showing anti-prolifera- tive effects; the flavone quercetin, for example, is able to inhibit the growth of several tumour cell lines, 8 while chalcones were described as NF-jB pathway inhibitors, inducing cell death via apoptosis. 9,10 Other interesting bioorganic compounds including polysulfides from garlic, 11 curcumin from the rhizome of Curcuma longa 12–15 and the sesquiterpene heteronemin from the marine organism Hyrtios erecta, 16 possess anti-tumour, anti-inflammatory and anti-tumorigenic abilities based on the anti-proliferative, pro- apoptotic character and/or cytotoxicity leading to cell death by apoptosis or autophagy depending on the cellular context. Coumarins constitute an important category of natural products as more than thousand coumarin derivatives have been described http://dx.doi.org/10.1016/j.bmc.2014.03.046 0968-0896/Ó 2014 Elsevier Ltd. All rights reserved. ⇑ Corresponding authors. Tel.: +82 2 880 8919 (M.D.); tel.: +351 234 370714; fax: +351 234 370084 (A.M.S.S.). E-mail addresses: [email protected] (M. Diederich), [email protected] (A.M.S. Silva). 1 Equal contribution. Bioorganic & Medicinal Chemistry 22 (2014) 3008–3015 Contents lists available at ScienceDirect Bioorganic & Medicinal Chemistry journal homepage: www.elsevier.com/locate/bmc
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Bis(4-hydroxy-2H-chromen-2-one): Synthesis and effects on leukemic cell lines proliferation and...

Bioorganic & Medicinal Chemistry 22 (2014) 3008–3015
Contents lists available at ScienceDirect
Bioorganic & Medicinal Chemistry
journal homepage: www.elsevier .com/locate /bmc
Bis(4-hydroxy-2H-chromen-2-one): Synthesis and effectson leukemic cell lines proliferation and NF-jB regulation
http://dx.doi.org/10.1016/j.bmc.2014.03.0460968-0896/� 2014 Elsevier Ltd. All rights reserved.
⇑ Corresponding authors. Tel.: +82 2 880 8919 (M.D.); tel.: +351 234 370714; fax:+351 234 370084 (A.M.S.S.).
E-mail addresses: [email protected] (M. Diederich), [email protected](A.M.S. Silva).
1 Equal contribution.
Oualid Talhi a,1, Michael Schnekenburger b,1, Jana Panning b,c, Diana G. C. Pinto a, José A. Fernandes d,Filipe A. Almeida Paz d, Claus Jacob c, Marc Diederich e,⇑, Artur M. S. Silva a,⇑a QOPNA, Department of Chemistry, University of Aveiro, 3810-193 Aveiro, Portugalb Laboratoire de Biologie Moléculaire et Cellulaire du Cancer, Hôpital Kirchberg, L-2540 Luxembourg, Luxembourgc School of Pharmacy, Building B 2.1., Room 1.13, University of Saarland, Campus, 66123 Saarbrucken, Germanyd CICECO, Department of Chemistry, University of Aveiro, 3810-193 Aveiro, Portugale College of Pharmacy, Seoul National University, 599 Gwanak-ro, Gwanak-gu, Seoul 151-742, Republic of Korea
a r t i c l e i n f o
Article history:Received 11 February 2014Revised 27 March 2014Accepted 30 March 2014Available online 6 April 2014
Keywords:Biscoumarins2D-NMRX-ray crystallographyLeukaemia cell linesNF-jB inhibition
a b s t r a c t
Synthesis of the bis-4-hydroxycoumarin-type compound, 3,30-[3-(2-hydroxyphenyl)-3-oxopropane-1,1-diyl]bis(4-hydroxy-2H-chromen-2-one), was performed by two alternative pathways, either involving abasic organocatalyzed 1,4-conjugate addition tandem reaction of 4-hydroxycoumarin on chromone-3-carboxylic acid, or a double condensation of 4-hydroxycoumarin on x-formyl-20-hydroxyacetophenone.The anti-proliferative effects of the bis-4-hydroxycoumarin-type compound on human K-562 (chronicmyeloid leukaemia) and JURKAT (acute T-cell leukaemia) cell lines using trypan blue staining, as wellas its involvement in nuclear factor-kappa B (NF-jB) regulation analyzed by luciferase reporter gene assay,gene expression analysis and western blots were analysed. This compound inhibited TNFa-inducedNF-jB activation in K-562 (IC50 17.5 lM) and JURKAT (IC50 19.0 lM) cell lines, after 8 h of incubation.Interestingly, it exerted mainly cytostatic effects at low doses on both cell lines tested, whereas itdecreased JURKAT cell viability starting at 50 lM from 24 h of treatment. Importantly, it did not affectthe viability of peripheral blood mononuclear cells (PBMCs) from healthy donors, even at concentrationsabove 100 lM.
� 2014 Elsevier Ltd. All rights reserved.
1. Introduction
Cancer is characterized by uncontrolled growth and spreading ofabnormal cells. Smoking, alcohol intake, obesity and physical inac-tivity are seen as main cancer risk factors. Also genetic predisposi-tion, inflammatory environment, activation of oncogenes anddeactivation of tumour suppressor genes contribute to developmentof this disease.1–3 A central molecular mechanism involved ininflammatory and innate immune response is the transcription fac-tor nuclear factor kappa-B cell (NF-jB) comprising five membersincluding, relA (p65), relB, cRel, NF-jB1 (p50) and NF-jB2 (p52).Canonical NF-jB pathway is activated by Toll-like-, tumour necrosisfactor (TNF-a) or T-cell receptors. RelA-p50 dimers are associatedwith inhibitor of jB (IjB) retaining them in an inactive form in thecytoplasm. After stimulation of cell surface receptors with pro-
inflammatory cytokines, IjB kinase (IKK) is activated and phosphor-ylates IjB. As a consequence IjB is ubiquitinated and subsequentlydegraded by the proteasome. The released NF-jB transcription fac-tor then translocates to the nucleus and transactivates target genesinvolved in cancer initiation, promotion and progression.4–6
A large majority (79.8%) of anticancer drugs developed between1981 and 2010 derive from natural origins.7 The flavonoid family isa prominent group of natural compounds showing anti-prolifera-tive effects; the flavone quercetin, for example, is able to inhibitthe growth of several tumour cell lines,8 while chalcones weredescribed as NF-jB pathway inhibitors, inducing cell death viaapoptosis.9,10 Other interesting bioorganic compounds includingpolysulfides from garlic,11 curcumin from the rhizome of Curcumalonga12–15 and the sesquiterpene heteronemin from the marineorganism Hyrtios erecta,16 possess anti-tumour, anti-inflammatoryand anti-tumorigenic abilities based on the anti-proliferative, pro-apoptotic character and/or cytotoxicity leading to cell death byapoptosis or autophagy depending on the cellular context.
Coumarins constitute an important category of natural productsas more than thousand coumarin derivatives have been described

4-PPy O
O
HOOC
- CO2
O
HO O
O
O-HOOC
O
O
O
O
O-
O
O
O
O
O
O
O
OH
4PPy
O
OH
O
O- OHO
O
O OO
O
21
Intermediate
O
H
OHOH3
AB
- H2O
OHO
O
OH
O OHO
O
OHO
O
O
OH OO
OH
4
Scheme 1. Synthesis of biscoumarin 4.
O
OO
HO
CH2
O
OH
O
H
OH
H
2
3'2'
4'
1
HMBC
3
O
OO
HO
CH2
O
OH
O
H
OH
H
2
3'2'
4'
1
3
NOESY
6'''
Figure 1. Main HMBC correlations and NOE effects observed in the 2D NMR spectraof the biscoumarin 4.
O. Talhi et al. / Bioorg. Med. Chem. 22 (2014) 3008–3015 3009
and isolated from more than 800 plant species.17 Certain deriva-tives exhibit useful pharmaceutical activities, as they were creditedwith potent antioxidant, anti-inflammatory, anticancer and mol-luscicidal activities.18 They induce apoptosis by cytochrome crelease and caspase activation.19 Recent reports described theselective cytotoxicity to cancer cells of some structurally simplecoumarins, such as 7,8-dihydroxy-4-methylcoumarin, 7,8-diacet-oxy-4-methylcoumarin20 and 7-hydroxycoumarin,21 which inhibitproliferation of lung cancer cells without affecting the normalgrowth of healthy peripheral blood mononuclear cells (PBMCs).In the quest for novel natural-mimicking compounds, we describehere the anticancer and anti-inflammatory activities of a new syn-thetic bis-4-hydroxycoumarin-type compound, which has beenelaborated by updated synthetic routes.
Figure 2. Schematic representation of the molecular unit present in the crystalstructure of the biscoumarin compound 4. Non-hydrogen atoms are represented asthermal ellipsoids drawn at the 50% probability level and hydrogen atoms as smallspheres with arbitrary radii. The atomic labelling is provided for all non-hydrogenatoms. Dashed green and gold lines represent, respectively, strong O–H� � �O andweak C–H� � �O hydrogen bonds. Hydrogen bonding geometry: O2–H2� � �O1,dO� � �O = 2.575(5) Å, <(OHO) = 152�; O5–H5� � �O6, dO� � �O = 2.614(5) Å, <(OHO) = 158�;O8–H8� � �O3, dO� � �O = 2.665(5) Å, <(OHO) = 158�; C2–H2B� � �O5, dO� � �O = 3.145(5) Å,<(CHO) = 125�. The inset (top view of the molecule) depicts the tilted nature of the3-(2-hydroxyphenyl)propion-1-yl unit with respect to the sp3 average plane of thecentral C1–C2 moiety.
2. Results and discussion
2.1. Chemistry
The well-known biological activities of coumarins oriented ustowards a biscoumarin scaffold as a potential biologically activedyad presenting a quite symmetrical geometry and containingthe relevant bioactive benzopyran-2-one ring. The targetbiscoumarin 4 was first synthesized by reacting two equivalentsof 4-hydroxycoumarin 1 on chromone-3-carboxylic acid 2 usinga catalytic amount of 4-pyrrolidinopyridine (4PPy) in refluxingchloroform. Mechanistically, an initial 1,4-conjugate addition/decarboxylation tandem process has led to the conjugatedintermediate which suffers a subsequent classical Michael additionon the existing a,b-unsaturated ketone system giving rise to thebiscoumarin 4 (72%) (Scheme 1, pathway A).
The action of 4-hydroxycoumarin 1 on aliphatic and aromaticaldehydes has been reported in the literature giving similar bis-coumarin structures.11 In consequence, we decided to study thereaction of 4-hydroxycoumarin 1 on the cheap x-formyl-2-hydroxyacetophenone 3, utilizing our optimized conditions(catalytic amount of 4PPy in refluxing chloroform). Following thisprotocol, we prepared biscoumarin 4 with a higher yield (83%),which is readily isolated in a pure state after solvent removal undervacuum and direct re-crystallisation from ethanol (Scheme 1,pathway B). This alternative and economic reaction follows asuccession of organo-base promoted double condensations of two4-hydroxycoumarin units 1 through their activated 3-C methylene

3010 O. Talhi et al. / Bioorg. Med. Chem. 22 (2014) 3008–3015
group on the aldehyde function in x-formyl-2-hydroxyacetophe-none 3.
Structural elucidation of 4 was first made on the basis of exten-sive 1D and 2D NMR techniques. From 1H NMR spectrum (CDCl3)we concluded that a pair of 4-hydroxycoumarin nuclei and a 3-(20-hydroxyphenyl)-3-oxopropyl moiety is involved in the structure ofthe obtained compound. Resonances of the aromatic protons of both4-hydroxycoumarin nuclei appear as broad and unresolved signals,while both exchangeable protons of hydroxyl groups (40-OH/400-OH)occur at different chemical shift values (11.45 and 12.10 ppm).These data seem to indicate that both 4-hydroxycoumarin ringsmay not be absolutely positioned in a symmetrical bilateral mannerrelative to the sp3 medium plane of the tetrahedral carbon C-1(26.7 ppm in the 13C NMR spectrum). Moreover, the H-2 signalappears as a pseudo triplet, which also confirms some asymmetry
Prol
ifera
tion
(cel
ls/m
L)
µ
A
B
0
25
50
75
10024 h 48 h 72 h
0
25
50
75
100
10 25 50 100
Compound 4 ( µM)
0.0E+00
4.0E+05
8.0E+05
1.2E+06
1.6E+0624h 48h 72h
0.0E+00
5.0E+05
1.0E+06
1.5E+06
0 10 25 50 100 0%
25%
50%
75%
100%
0%
25%
50%
75%
100%
0%
25%
50%
75%
100%
0
0%
25%
50%
75%
100%
Cel
ls
C
Viab
ility
(% o
f con
trol
)
0
25
50
75
100
10 50 100
24h 48h D
Compound 4 ( µM)
Figure 3. Compound 4 decreases cancer cell proliferation by blocking cell cycle progrconcentrations of compound 4. (A) Cell viability and (B) proliferation were assessed aftePBMCs from healthy donors were incubated with compound 4 and then cell viabilityindependent experiments. ⁄ and ⁄⁄ indicate p <0.05 and p <0.01 versus untreated cells, r
of both coumarin moieties (Fig. S1 of ESI). When the 1H NMR spec-trum was acquired at 50 �C (CDCl3), all signals were tentativelyresolved and H-2 clearly appears as the expected doublet, whilethe signals of both hydroxyl protons (40-OH/400-OH) appear veryclose 11.61 and 11.95 ppm (Fig. S2 of ESI). This result indicates thatan increase in the temperature greatly affects the mobility of themolecule moieties, making the structure more symmetric.
Alongside with HSQC, the HMBC experiment of biscoumarin 4allows the assignments of several non-protonated and quaternarycarbons resonances. H-1 showed correlations with C-30/300 and thecarbonyl carbons in the structure 4, including C-3 (202.6 ppm) ofthe 3-(20-hydroxyphenyl)propion-1-yl fragment, C-20/200 and C-40/4 (164.1 and 165.3) of the 4-hydroxycoumarin units (Fig. 1, HMBC).In addition, H-2 shows weak connectivity with C-3 and C-30/300
enabling the assignment of this latter to the signals at 105.3 and
8h
10 25 50 100
48h
24h
72h
* ** **
* *
** **
** ** **
0%
25%
50%
75%
100%
0%
25%
50%
75%
100%
0%
25%
50%
75%
100%
0%
25%
50%
75%
100%
0 10 25 50 100
**
* ** ** **
** ** **
** ** **
JURKAT K-562 G2/M S G1
Compound 4 ( µM) Compound 4 ( µM)
ession in G0/G1 phase. K-562 and JURKAT cells were treated with the indicatedr 24, 48 and 72 h. (C) Cell cycle distribution was analyzed between 8 and 72 h. (D)was evaluated after 24 and 48 h of treatment. Data are the mean ± SD of three
espectively.
O. Talhi et al. / Bioorg. Med. Chem. 22 (2014) 3008–3015 3011
106.0 ppm, once again confirming the absence of symmetry atroom temperature (Figs. S3–S6 of ESI).
In the NOESY experiment of biscoumarin 4 (Fig. 1), the main NOEeffects are observed between H-6000 and H-2 and also between both40/400-OH protons and H-1 and H-2. However, 2000-OH proton pre-sents only NOE cross effect with one of the two 40/400-OH protonswhich is a clear indication that the coumarin units are not disposedin a symmetric manner around the central tertiary carbon atom atroom temperature (Fig. S7 of ESI). This assumption was unequivo-cally confirmed using single-crystal X-ray diffraction studies, whichshowed that at �123 �C compound 4 crystallizes in the centrosym-metric P21/n space group with the asymmetric unit being com-posed of a whole molecular unit as depicted in Figure 2. The two4-hydroxycoumarin moieties attached to the tertiary carbon atomare mutually rotated. In this way the hydroxyl groups are engagedin strong and highly directional intra-molecular O–H� � �O hydrogenbonds with the carbonyl moieties of the neighbouring 4-hydroxy-coumarin unit, ultimately boosting the structural robustness of thismolecule. The same type of interaction is also observed within the3-(20-hydroxyphenyl)propion-1-yl moiety, but the presence of themethylene C-2 moiety prevents the existence of strong supramo-lecular interactions with the substituent 4-hydroxycoumarins.
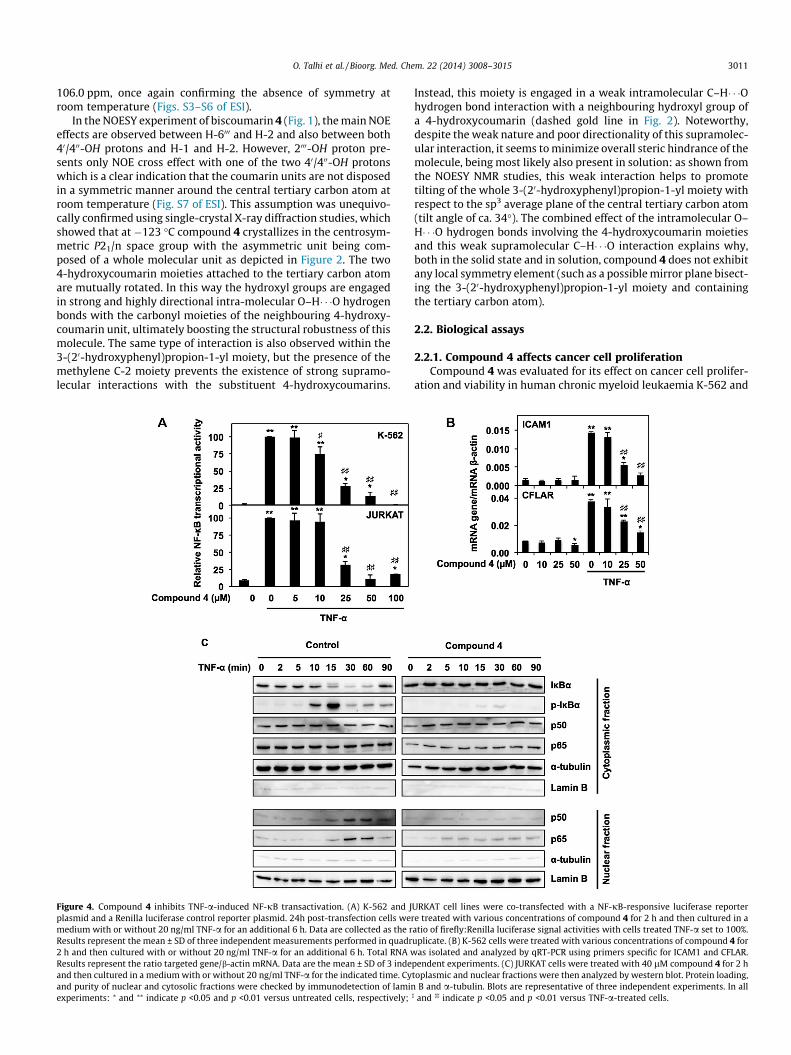
Figure 4. Compound 4 inhibits TNF-a-induced NF-jB transactivation. (A) K-562 and Jplasmid and a Renilla luciferase control reporter plasmid. 24h post-transfection cells wermedium with or without 20 ng/ml TNF-a for an additional 6 h. Data are collected as the rResults represent the mean ± SD of three independent measurements performed in quadr2 h and then cultured with or without 20 ng/ml TNF-a for an additional 6 h. Total RNA wResults represent the ratio targeted gene/b-actin mRNA. Data are the mean ± SD of 3 indeand then cultured in a medium with or without 20 ng/ml TNF-a for the indicated time. Cyand purity of nuclear and cytosolic fractions were checked by immunodetection of laminexperiments: ⁄ and ⁄⁄ indicate p <0.05 and p <0.01 versus untreated cells, respectively;
Instead, this moiety is engaged in a weak intramolecular C–H� � �Ohydrogen bond interaction with a neighbouring hydroxyl group ofa 4-hydroxycoumarin (dashed gold line in Fig. 2). Noteworthy,despite the weak nature and poor directionality of this supramolec-ular interaction, it seems to minimize overall steric hindrance of themolecule, being most likely also present in solution: as shown fromthe NOESY NMR studies, this weak interaction helps to promotetilting of the whole 3-(20-hydroxyphenyl)propion-1-yl moiety withrespect to the sp3 average plane of the central tertiary carbon atom(tilt angle of ca. 34�). The combined effect of the intramolecular O–H� � �O hydrogen bonds involving the 4-hydroxycoumarin moietiesand this weak supramolecular C–H� � �O interaction explains why,both in the solid state and in solution, compound 4 does not exhibitany local symmetry element (such as a possible mirror plane bisect-ing the 3-(20-hydroxyphenyl)propion-1-yl moiety and containingthe tertiary carbon atom).
2.2. Biological assays
2.2.1. Compound 4 affects cancer cell proliferationCompound 4 was evaluated for its effect on cancer cell prolifer-
ation and viability in human chronic myeloid leukaemia K-562 and
URKAT cell lines were co-transfected with a NF-jB-responsive luciferase reportere treated with various concentrations of compound 4 for 2 h and then cultured in aatio of firefly:Renilla luciferase signal activities with cells treated TNF-a set to 100%.uplicate. (B) K-562 cells were treated with various concentrations of compound 4 for
as isolated and analyzed by qRT-PCR using primers specific for ICAM1 and CFLAR.pendent experiments. (C) JURKAT cells were treated with 40 lM compound 4 for 2 htoplasmic and nuclear fractions were then analyzed by western blot. Protein loading,
B and a-tubulin. Blots are representative of three independent experiments. In all] and ]] indicate p <0.05 and p <0.01 versus TNF-a-treated cells.

OOHOO
OHOOH
O
OOHOO
OH
+
4 5 1
Scheme 2. Chemical substructures of the biscoumarin 4.
0.0E+00 5.0E+05 1.0E+06 1.5E+06 2.0E+06
0 24 48 72
A
0.0E+00
5.0E+05
1.0E+06
1.5E+06
2.0E+06 0 µM 10 µM 100 µM
Compound 1
Compound 5
Prol
ifera
tion
(cel
ls/m
L)
Hours
B
100
120
140
B tiv
ity
3012 O. Talhi et al. / Bioorg. Med. Chem. 22 (2014) 3008–3015
acute T-cell leukaemia JURKAT cell lines (Fig. 3A and B). Resultsshowed that treatments with compound 4 affected cell prolifera-tion in both cell lines with JURKAT cells being more sensitive thanK-652 cells. Indeed, we observed a moderate effect at 10 lM after48 h of treatment only in JURKAT cells and then a more pro-nounced effect in both cell lines starting from 25 lM and 24 h oftreatment. The drastic decrease of proliferation observed from50 lM was accompanied by a relatively restrained reduction of cellviability but solely in JURKAT cells. Decreased tumour cell prolifer-ation was associated to an inhibition of cell cycle progression inG0/G1 phase in both cell lines starting at 8 h from 50 lM and at24 h from 25 lM in JURAKT and K-562 cells, respectively(Fig. 3C). Nevertheless, compound 4 did not alter the viability ofPBMCs isolated from healthy donors (Fig. 3D).
2.2.2. Compound 4 inhibits TNFa-induced NF-jB activationIt is well established that NF-jB signaling pathway is implicated in
the regulation of inflammation, proliferation and apoptosis. In addi-tion, its activation is linked to cancer development and progression.Therefore, we evaluated interference of compound 4 with TNF-a-induced NF-jB transactivation. We observed that compound 4 abro-gated TNF-a-induced NF-jB activation in a dose-dependent mannerwith IC50 values of 17.5 and 19.0 lM in K-562 and JURKAT cells,respectively (Fig. 4A). These values are within the range of IC50 valuespreviously reported by our laboratory for various compounds againstTNF-a-induced NF-jB activation in K-562 cells using the same exper-imental conditions (Table 1). In order to further confirm the inhibitoryeffect of compound 4 on TNF-a-induced NF-jB signaling, we per-formed gene expression analyses of NF-jB target genes in K-562 cells(Fig. 4B). Results demonstrated that compound 4 dose-dependentlyimpaired expression of ICAM (intercellular adhesion molecule) 1and caspase 8 and FADD-like apoptosis regulator (CFLAR; c-Flip)genes induced by TNF-a. Since NF-jB activation and its subsequenttranslocation form cytoplasm to nucleus requires degradation of itscytoplasmic inhibitor IjB, we determined the effect of compound 4on IjBa integrity as well as on translocation of p50 and p65 NF-jBsubunits (Fig. 4C).Western blot analyses revealed that compound 4prevented TNF-a-induced IjBa degradation and the subsequenttranslocation of NF-jB subunits into the nucleus. In the past fewyears, several natural and synthetic coumarin derived compoundswere reported to act as NF-jB inhibitors. For instance, decursinolangelate, a coumarin compound isolated from Angelica gigas, blocksthe transmigration and inflammatory activation of cancer cellsthrough inhibition of phosphoinositide 3-kinase (PI3K), extracellularsignal-regulated kinases (ERK) and NF-jB activation.22 Decursin andnodakenin, two other coumarins isolated from Angelicae giga, were
Table 1Inhibitors of TNF-a-induced NF-jB activation in K-562 cells
Compound IC50 (lM) Refs.
4-Hydroxychalcone 24 (Orlikova, 2012)16
4-Methoxychalcone 29 (Orlikova, 2012)16
Altersolanol A 0.8 (Teiten, 2013)27
Butein 38 (Orlikova, 2012)16
Calomelanone 11 (Orlikova, 2012)16
Curcumin 8 (Duvoix, 2003)13
Embellicine A 3 (Ebrahim, 2013)28
Embellicine B 0.4 (Ebrahim, 2013)28
Flavokawain C 8 (Orlikova, 2012)16
Goniothalamin 7 (Orlikova, 2013)29
Heteronamin 5 (Schumacher, 2010)16
Homobutein 38 (Orlikova, 2012)16
Isoliquiritigenin 32 (Orlikova, 2012)16
Phloretin 41 (Orlikova, 2012)16
Sanguinarine 2 (Duvoix, 2004)28
Tetraacetylaltersolanol 4.6 (Teiten, 2013)27
UNBS1450 0.03 (Juncker, 2011)30
b-Lapachone 4 (Duvoix, 2004)28
reported to block the UVB-induced activation of NF-jB and suppressthe transcriptional activity and translocation of NF-jB by inhibitingIjBa degradation and IKK phosphorylation, respectively.23,24 Fur-thermore, it has been shown that increased IjB phosphorylationand NF-jB nuclear translocation induced by ischemia/reperfusioninjury were significantly decreased following pre-treatment with ost-hole, a natural derivative of coumarin found in several plants.25 Basedon these findings, we believe that our novel dicoumarinic compoundcould further provide increased inhibition potential. However, clearlythe initial(s) target(s) of coumarin-mediated NF-jB inhibitionremains to be elucidated.
We selected here two leukaemia cell lines where activation ofNF-jB is under the control of differential molecular mechanisms.
0
20
40
60
80
100 10 100 10
TNF-α (20ng/ml)
0 0 Compound (µM)
Compound 1 Compound 5
Rel
ativ
e N
F-κ
tran
scrip
tiona
l ac
Figure 5. Effect of compound 1 and 5 on cell proliferation and NF-jB transcrip-tional activity. (A) K-562 cells were treated with the indicated concentrations ofcompound and cell proliferation was evaluated after 24, 48 and 72 h. Data are themean ± SD of three independent experiments. (B) K-562 cells were co-transfectedwith a NF-jB-responsive luciferase reporter plasmid and the Renilla luciferasecontrol reporter plasmid. 24 h post-transfection cells were treated with variousconcentration of compound 1 and 5 for 2 h and then cultured in a medium with orwithout 20 ng/ml TNF-a for an additional 6 h. Data are collected as the ratio offirefly:Renilla luciferase signal activities with cell treated TNF-a set to 100%. Resultsrepresent the mean ± SD of three independent measurements performed inquadruplicate. ⁄⁄ indicates p <0.01 versus untreated cells.

O. Talhi et al. / Bioorg. Med. Chem. 22 (2014) 3008–3015 3013
Whereas JURKAT cells activate NF-jB via a canonical pathwayinvolving degradation of the IjB inhibitor of the NF-jB transcrip-tion factor allowing translocation into the nucleus and transactiva-tion of target genes, K-562 cells rely on oncokinase Bcr-Abl.According to Baldwin and co-workers expression of Bcr-Abl leadsto activation of NF-jB-dependent transcription by causing nucleartranslocation of NF-jB as well as by increasing the transactivationfunction of the RelA/p65 subunit of NF-jB. Importantly, this activa-tion is dependent on the tyrosine kinase activity of Bcr-Abl’.26 Inour hands and in other labs, degradation of IjB inhibitor is notobservable in K-562 cell line and accordingly was not investigatedhere. We essentially investigate effects of anticancer compoundson leukaemia and lymphoma cell models and as chronic myeloge-nous leukaemia patients show increased resistance against clini-cally used Gleevec (Imatinib) Bcr-Abl inhibitor, additional novelinhibitors are urgently required.
2.2.3. Substructures 1 and 5 present in compound 4 fail toproduce biological effects
In order to clearly demonstrate that the newly synthetizedhybrid compound 4 possesses an inhibitory potential superior tothe substructures present in its structure 4, namely 4-hydroxy-coumarin 1 and 20-hydroxypropiophenone 5 (Scheme 2), we theninvestigated the effect of such compounds on cell viability andTNF-a-activation of NF-jB transcriptional activity in K-562 cells(Fig. 5A and B). Results revealed that both substructures failed toproduce the effects induced by compound 4.
3. Conclusions
Our results underline two alternative synthetic methods of the bis-4-hydroxycoumarin-type compound 3,30-[3-(2-hydroxyphenyl)-3-oxopropane-1,1-diyl]bis(4-hydroxy-2H-chromen-2-one. We demon-strated that the new compound inhibits proliferation of K-562 andJUKAT leukaemia cell lines associated to an accumulation of cells inG0/G1 phase of cell cycle without affecting the viability of healthyPBMCs. In addition, this compound blocks the activation of the pro-inflammatory NF-jB pathway. We have also rationalized the conceptof the active molecule built from inactive constructive fragments, assubstructures of the biscoumarin-type compound, 20-hydroxyphenyl-propione and 4-hydroxycoumarin, do not show any activity.
4. Experimental section
4.1. General
Melting points are measured on Büchi B-540 equipment and areuncorrected. NMR spectra were recorded on Bruker Avance 300spectrometer (300.13 MHz for 1H and 75.47 MHz for 13C), withCDCl3 as solvent and TMS as the internal standard. Chemical shifts(d) are reported in ppm and coupling constants (J) in Hz which allare calculated using MESTRENOVA 8 (Free Trail License) analyticalchemistry software suite for NMR, LC, GC and MS. Unequivocal 13Cassignments were made with the aid of 2D HSQC and HMBC exper-iments (delays for one bond and long-range JC/H couplings wereoptimized for 145 and 7 Hz, respectively). Compounds 1, 2, 3, 5and 4-pyrrolidinopyridine (4PPy) have been purchased formSigma–Aldrich.
4.2. Single-crystal X-ray diffraction studies
Crystals of compound 4 were harvested from the crystallizationvial and were immersed in highly viscous FOMBLIN Yperfluoropolyether vacuum oil (LVAC 140/13) purchased fromSigma–Aldrich. A suitable single-crystal was selected and mounted
on a Hampton Research CryoLoop31 with the help of a Stemi 2000stereomicroscope equipped with Carl Zeiss lenses. Data were col-lected at 150(2) K on a Bruker X8 Kappa APEX II charge-coupleddevice (CCD) area-detector diffractometer (MoKa graphite-mono-chromated radiation, k = 0.71073 Å) controlled by the APEX2 soft-ware package,32 and equipped with an Oxford Cryosystems Series700 cryostream monitored remotely using the software interfaceCryopad.33 Images were processed using the software packageSAINT+,34 and data were corrected for absorption by the multi-scansemi-empirical method implemented in SADABS.35 The crystalstructure was solved using the direct methods algorithm imple-mented in SHELXS-97,36,37 allowing the immediate location ofmost of non-hydrogen atoms composing the molecular unit ofcompound 4. All remaining non-hydrogen atoms were locatedfrom difference Fourier maps calculated from successive full-matrix least squares refinement cycles on F2 using SHELXL-97.37,38 Non-hydrogen atoms were successfully refined usinganisotropic displacement parameters.
Hydrogen atoms bound to carbon were placed at their idealizedpositions using appropriate HFIX instructions in SHELXL: 43 for thearomatic, 23 for the –CH2– groups and 13 for the tertiary carbonatom. The hydrogen atoms associated with the –OH groups weredirectly located from difference Fourier maps and were includedin the structural model with the O–H distances restrained to0.95(1) Å so to ensure a chemically reasonable geometry for thesemoieties. All these hydrogen atoms were included in subsequentrefinement cycles in riding-motion approximation with isotropicthermal displacements parameters (Uiso) fixed at 1.2 (those boundto carbon) or 1.5 (for the –OH groups) times Ueq of the atom towhich they are attached.
The last difference Fourier map synthesis showed the highestpeak (1.274 e�3) and deepest hole (�0.363 e�3) located at1.10 Šand 0.58 Šfrom O5 and O7, respectively. Structural draw-ings have been created using the software package Crystal ImpactDiamond.39
Crystal data for 3,30-[3-(2-hydroxyphenyl)-3-oxopropane-1,1-diyl]bis(4-hydroxy-2H-chromen-2-one 4: C27H18O8, MW = 470.41g mol�1, colourless block (0.16 � 0.08 � 0.05 mm3), monoclinic,space group P21/n, a = 11.1193(13) Å, b = 14.1801(16) Å, c =14.4024(14) Å, b = 112.559(7), V = 2097.1(4) Å3, Z = 4, Dc = 1.490g cm�3, l(Mo-Ka) = 0.111 mm�1, 19163 reflections were collectedin the range 2.10 6 h 6 25.35, index ranges �12 6 h 6 13,�11 6 k 6 17, �17 6 l 6 17, of which 3847 were independent(Rint = 0.0753), completeness to h = 25.35� = 100.0%, final R1 ([I >2r(I)]) = 0.0879, wR2 ([I>2r(I)]) = 0.2354, R1 (all data) = 0.1214,wR2 (all data) = 0.2613. CCDC 920034.
4.3. Synthesis of 3,30-[3-(2-hydroxyphenyl)-3-oxopropane-1,1-diyl]bis(4-hydroxy-2H-chromen-2-one) 4
4-Hydroxycoumarin 1 (2.00 g, 12.34 mmol) is added to chro-mone-3-carboxylic acid 2 (1.174 g, 6.17 mmol, 0.5 equiv, pathwayA) or x-formyl-20-hydroxyacetophenone 3 (1.013 g, 6.17 mmol,0.5 equiv, pathway B) in chloroform (20 mL). A catalytic amountof 4PPy (0.09 g, 0.62 mmol, 0.05 equiv) is dropped into the solu-tion, which is allowed to reflux under stirring overnight. AfterTLC monitoring, the solvent is evaporated and the resulting resin-ous solid is directly recrystallized from ethanol to afford the bis-coumarin 4: C27H18O8 (yellowish white solid, MW = 470.43 g/mol, 2.410 g, yield 83%, mp = 223–225 �C).
1H NMR (300.13 MHz, CDCl3): d = 4.21 (pseudo t, J = 6.9 Hz, 2H,H-2), 5.44 (t, J = 6.7 Hz, 1H, H-1), 6.86–6.97 (m, 2H, H-3000, H-5000),7.368 (br s, 2H, H-60/600), 7.369 (dd, J = 7.8 and 0.9 Hz, 2H, H-80/800), 7.46 (ddd, J = 7.8, 7.2 and 1.4 Hz, 1H, H-4000), 7.60 (t, J = 7.8 Hz,2H, H-70/700), 7.87 (dd, J = 8.1 and 1.4 Hz, 1H, H-6000), 8.01 (br s,

3014 O. Talhi et al. / Bioorg. Med. Chem. 22 (2014) 3008–3015
2H, H-50/500), 11.45 and 12.10 (2 br s, 2H, 40/400-OH), 11.92 (s, 1H,2000-OH) ppm.
1H NMR (300.13 MHz, 50 �C, CDCl3): d = 4.18 (d, J = 6.7 Hz, 2H,H-2), 5.44 (t, J = 6.7 Hz, 1H, H-1), 6.87–6.94 (m, 2H, H-3000, H-5000),7.32–7.37 (m, 4H, H-60/600, H-80/800), 7.44 (ddd, J = 7.7, 7.2 and1.6 Hz, 1H, H-4000), 7.60 (dt, J = 7.6 and 1.6 Hz, 2H, H-70/700), 7.85(dd, J = 8.1 and 1.6 Hz, 1H, H-6000), 8.01 (br d, 2H, J = 7.9 Hz, H-50/500), 11.61 and 11.95 (2 br s, 2H, 40/400-OH), 11.85 (s, 1H, 2000-OH)ppm.
13C NMR (75.47 MHz, CDCl3): d = 26.7 (C-1), 37.8 (C-2), 105.3and 106.0 (C-30,300), 116.6 (C-80,800), 118.5 (C-3000), 119.0 (C-1000),119.1 (C-5000), 124.0 (C-100,1000), 124.4 (C-50,500), 125.0 (C-60,600),129.6 (C-6000), 132.8 (C-70,700), 136.7 (C-4000), 152.2 (C-90/900), 162.2(C-2000), 164.1 and 165.3 (C-20,200, C-40,400), 202.6 (C-3) ppm.
HRMS (ESI+): m/z calcd for [C27H18O8+Na]+: 493.0899; found:493.0904. Anal. Calcd: C, 68.94; H, 3.86. Found: C, 68.96; H, 3.89.
4.4. Cell culture, treatments and viability assay
The human K-562 (chronic myeloid leukaemia) and JURKAT(acute T-cell leukaemia) cell lines were obtained from the Deut-sche Sammlung von. Mikroorganismen und Zellkulturen. PBMCswere obtained as previously reported.40 All cell types werecultured in RPMI 1640 (Lonza) supplemented with 10% heat-inac-tivated foetal calf serum (Lonza) and 1% antibiotic-antimycotic(Lonza) at 37 �C in humid atmosphere and 5% CO2.
Compounds 1, 4 and 5 were dissolved at 50 mM in DMSO(Sigma–Aldrich), aliquoted and stored at �20 �C upon treatment.TNFa (Sigma–Aldrich) was dissolved in 5% (w/v) BSA in 1� PBSto a concentration of 10 lg/mL. Cells in exponential phase weretreated at a concentration of 200,000 cells/mL. Control cells weretreated with the same volume of DMSO as the one required forcompounds. Cell proliferation and viability were measured as pre-viously reported.40
4.5. Analysis of cell cycle distribution
Cell cycle analyses were performed as previously described.41
4.6. Transient transfection and luciferase reporter gene assay
Cells were transiently transfected with 5 lg of a luciferasereporter gene construct containing five repeats of a consensusNF-jB binding site (Stratagene, Genomics Agilent) and 5 lg of aRenilla luciferase control reporter plasmid (ph-RG-TK, Promega,Leiden, Netherlands) and reporter gene assays were carried outas previously defined.42
4.7. Protein extraction and Western blot
Nuclear and cytosolic fractions were extracted as previouslyreported.43 Western blot experiments were carried out as previ-ously described with minor modifications.44 Proteins of interestwere detected with ECL Prime Western Blotting Detection Systemreagent (GE Healthcare) using an ImageQuant LAS 4000 mini (GEHealthcare). Antibodies were purchased from Santa Cruz Biotech-nology: anti-IjBa (sc-371), anti-lamin B (sc-6216), anti-p50(sc-7178), anti-p65 (sc-8008); form Cell Signaling: anti-pIjBa(#9246); and from Calbiochem: anti-a-tubulin (CP06).
4.8. Gene expression analyses
Total RNA extraction, cDNA synthesis and qRT-PCR were per-formed as previously described.39 Primer sequences are availableon request.
4.9. Statistical analysis
Differences between mean values were tested for statisticalsignificance by the two-tailed Student’s t-test. p-Values below0.05 were considered as statistically significant.
Acknowledgments
We would like to thank Fundação para a Ciência e a Tecnologia(FCT, Portugal), the European Union, QREN, FEDER, COMPETE, forfunding the Organic Chemistry Research Unit (QOPNA) (ProjectPEst-C/QUI/UI0062/2013), Laboratório Associado Centro de Inves-tigação em Materiais Cerâmicos e Compósitos—CICECO (ProjectPEst-C/CTM/LA0011/2013) and the Portuguese National NMRNetwork (RNRMN). European Community’s Seventh FrameworkProgramme (FP7/2007-20139 under Grant agreement n� 215009)is also acknowledged for financial support. We further wish tothank FCT for the post-doctoral Grant SFRH/BD/63736/2009 (toJ.A.F.) and for specific funding toward the purchase of the single-crystal X-ray diffractometer. MiS is supported by a ‘WaxweilerGrant for cancer prevention research’ from the Action Lions‘Vaincre le Cancer’. The work performed at Laboratoire de BiologieMoléculaire et Cellulaire du Cancer was supported by TélévieLuxembourg, the «Recherche Cancer et Sang» foundation, and the«Recherches Scientifiques Luxembourg» and «Een Häerz firKriibskrank Kanner» associations. M.D. is supported by the NRFby the MEST of Korea for Tumor Microenvironment GCRC2012-0001184 Grant, by the Seoul National University (SNU)Research Grant, by the Research Settlement Fund for the new fac-ulty of SNU and by the Research Institute of PharmaceuticalSciences.
Supplementary data
Supplementary data (1H, 13C, HSQC, HMBC and NOESY NMRspectra) associated with this article can be found, in the online ver-sion, at http://dx.doi.org/10.1016/j.bmc.2014.03.046.
References and notes
1. Balkwill, F.; Mantovani, A. Lancet 2001, 357, 539.2. Byers, T. Annu. Rev. Public Health 2010, 31, 121.3. Schnekenburger, M.; Diederich, M. Curr. Colorectal Cancer Rep. 2012, 8, 66.4. Baud, V.; Karin, M. Nat. Rev. Drug. Disc. 2009, 8, 33.5. Gilmore, T. D. Oncogene 2006, 25, 6680.6. Li, Q.; Verma, I. M. Nat. Rev. Immunol. 2002, 2, 725.7. Newman, D. J.; Cragg, G. M. J. Nat. Prod. 2012, 75, 311.8. Ren, W.; Qiao, Z.; Wang, H.; Zhu, L.; Zhang, L. Med. Res. Rev. 2003, 23, 519.9. Orlikova, B.; Schnekenburger, M.; Zloh, M.; Golais, F.; Diederich, M.; Tasdemir,
D. Oncol. Rep. 2012, 28, 797.10. Orlikova, B.; Tasdemir, D.; Golais, F.; Dicato, M.; Diederich, M. Genes Nutr. 2011,
6, 125.11. Busch, C.; Jacob, C.; Anwar, A.; Burkholz, T.; AichaBa, L.; Cerella, C.; Diederich,
M.; Brandt, W.; Wessjohann, L.; Montenarh, M. Int. J. Oncol. 2010, 36, 743.12. Duvoix, A.; Blasius, R.; Delhalle, S.; Schnekenburger, M.; Morceau, F.; Henry, E.;
Dicato, M.; Diederich, M. Cancer Lett. 2005, 223, 181.13. Duvoix, A.; Morceau, F.; Delhalle, S.; Schmitz, M.; Schnekenburger, M.; Galteau,
M. M.; Dicato, M.; Diederich, M. Biochem. Pharmacol. 2003, 66, 1475.14. Duvoix, A.; Morceau, F.; Schnekenburger, M.; Delhalle, S.; Galteau, M. M.;
Dicato, M.; Diederich, M. Ann. N.Y. Acad. Sci. 2003, 1010, 389.15. Teiten, M. H.; Eifes, S.; Dicato, M.; Diederich, M. Toxins (Basel) 2010, 2, 128.16. Schumacher, M.; Cerella, C.; Eifes, S.; Chateauvieux, S.; Morceau, F.; Jaspars, M.;
Dicato, M.; Diederich, M. Biochem. Pharmacol. 2010, 79, 610.17. Murray, R. D. H.; Mendez, J.; Brown, S. A. The Natural Coumarins: Occurrence,
Chemistry and Biochemistry; Wiley: New York, 1982.18. O’Kennedy, R.; Douglas, R. Coumarins: Biology, Applications and Mode of Action;
Wiley: New York, 1997.19. Kostova, I. Expert Opin. Drug Discov. 2007, 2, 1605.20. Goel, A.; Prasad, A. K.; Parmar, V. S.; Ghosh, B.; Saini, N. Chem. Biol. Interact.
2009, 179, 363.21. Lopez-Gonzalez, J. S.; Prado-Garcia, H.; Aguilar-Cazares, D.; Molina-Guarneros,
J. A.; Morales-Fuentes, J.; Mandoki, J. J. Lung Cancer 2004, 43, 275.22. Kim, W. J.; Lee, M. Y.; Kim, J. H.; Suk, K.; Lee, W. H. Cancer Lett. 2010, 296, 35.

O. Talhi et al. / Bioorg. Med. Chem. 22 (2014) 3008–3015 3015
23. Hwang, B. M.; Noh, E. M.; Kim, J. S.; Kim, J. M.; Hwang, J. K.; Kim, H. K.; Kang, J.S.; Kim, D. S.; Chae, H. J.; You, Y. O.; Kwon, K. B.; Lee, Y. R. Int. J. Mol. Med. 2013,31, 477.
24. Rim, H. K.; Cho, W.; Sung, S. H.; Lee, K. T. J. Pharmacol. Exp. Ther. 2012, 342, 654.25. Dong, W.; Zhang, Z.; Liu, Z.; Liu, H.; Wang, X.; Bi, S.; Wang, X.; Ma, T.; Zhang, W.
Int. J. Mol. Med. 2013, 31, 1367.26. Reuther, J. Y.; Reuther, G. W.; Cortez, D.; Pendergast, A. M.; Baldwin, A. S., Jr.
Genes Dev. 1998, 12, 968.27. Duvoix, A.; Delhalle, S.; Blasius, R.; Schnekenburger, M.; Morceau, F.; Fougere,
M.; Henry, E.; Galteau, M. M.; Dicato, M.; Diederich, M. Biochem. Pharmacol.2004, 68, 1101.
28. Ebrahim, W.; Aly, A. H.; Wray, V.; Mandi, A.; Teiten, M. H.; Gaascht, F.; Orlikova,B.; Kassack, M. U.; Lin, W.; Diederich, M.; Kurtan, T.; Debbab, A.; Proksch, P. J.Med. Chem. 2013, 56, 2991.
29. Orlikova, B.; Schumacher, M.; Juncker, T.; Yan, C. C.; Inayat-Hussain, S. H.;Hajjouli, S.; Cerella, C.; Dicato, M.; Diederich, M. Food Chem. Toxicol. 2013, 59,572.
30. Juncker, T.; Cerella, C.; Teiten, M. H.; Morceau, F.; Schumacher, M.; Ghelfi, J.;Gaascht, F.; Schnekenburger, M.; Henry, E.; Dicato, M.; Diederich, M. Biochem.Pharmacol. 2011, 81, 13.
31. Kottke, T.; Stalke, D. J. Appl. Crystallogr. 1993, 26, 615.32. APEX2, Data Collection Software Version 2.1-RC13, 2006, Bruker AXS, Delft,
The Netherlands.
33. Cryopad, Remote monitoring and control, Version 1.451, 2006, OxfordCryosystems, Oxford, United Kingdom.
34. SAINT+, Data Integration Engine v. 7.23a�, 1997–2005, Bruker AXS, Madison,Wisconsin, USA.
35. Sheldrick, G. M. SADABS v.2.01, Bruker/Siemens Area Detector AbsorptionCorrection Program, 1998, Bruker AXS, Madison, Wisconsin, USA.
36. Sheldrick, G. Acta Crystallogr. Sect. A 2008, 64, 112.37. Sheldrick, G. M. SHELXS-97, Program for Crystal Structure Solution, University
of Göttingen, 2008.38. Sheldrick, G. M. SHELXL-97, Program for Crystal Structure Refinement, 1997,
University of Göttingen.39. Brandenburg, K. DIAMOND, Version 3.2f. Crystal Impact GbR, 1997–2010,
Bonn, Germany.40. Schnekenburger, M.; Grandjenette, C.; Ghelfi, J.; Karius, T.; Foliguet, B.; Dicato,
M.; Diederich, M. Biochem. Pharmacol. 2011, 81, 364.41. Seidel, C.; Schnekenburger, M.; Dicato, M.; Diederich, M. Cancer Lett. 2014, 343,
134.42. Morceau, F.; Schnekenburger, M.; Blasius, R.; Buck, I.; Dicato, M.; Diederich, M.
Cancer Lett. 2006, 240, 203.43. Schnekenburger, M.; Morceau, F.; Duvoix, A.; Delhalle, S.; Trentesaux, C.;
Dicato, M.; Diederich, M. Biochem. Pharmacol. 2004, 68, 1269.44. Karius, T.; Schnekenburger, M.; Ghelfi, J.; Walter, J.; Dicato, M.; Diederich, M.
Biochem. Pharmacol. 2011, 81, 1329.












![Antitubercular effect of 8-[(4-Chloro phenyl) sulfonyl]-7-Hydroxy-4-Methyl-2H-chromen-2-One in guinea pigs](https://static.fdokumen.com/doc/165x107/63336ec5b6829c19b80c6a0b/antitubercular-effect-of-8-4-chloro-phenyl-sulfonyl-7-hydroxy-4-methyl-2h-chromen-2-one.jpg)


![Ethyl 2-(6-amino-5-cyano-3,4-dimethyl-2H,4H-pyrano[2,3-c]pyrazol-4-yl)acetate](https://static.fdokumen.com/doc/165x107/630bead9dffd3305850820dd/ethyl-2-6-amino-5-cyano-34-dimethyl-2h4h-pyrano23-cpyrazol-4-ylacetate.jpg)



