
Freezing tolerance and cold acclimation in guava (Psidium ...
Upload
washingtonCategory
view
0download
0
Cryobiology 68 (2014) 294–302
Contents lists available at ScienceDirect
Cryobiology
journal homepage: www.elsevier .com/ locate/ycryo
Biotransport and intracellular ice formation phenomena in freezinghuman embryonic kidney cells (HEK293T) q
http://dx.doi.org/10.1016/j.cryobiol.2014.02.0110011-2240/� 2014 Elsevier Inc. All rights reserved.
q Statement of funding: This work was supported by the National Natural ScienceFoundation of China (Nos. 51076149, 51276179), and the Fundamental ResearchFunds for the Central Universities (WK 2100230009).⇑ Corresponding authors.
E-mail addresses: [email protected] (G. Zhao), [email protected] (X. Zhou).
Yunpeng Xu a, Gang Zhao a,⇑, Xiaoming Zhou b,⇑, Weiping Ding a, Zhiquan Shu c, Dayong Gao c
a Department of Electronic Science and Technology, University of Science and Technology of China, Hefei 230027, Chinab School of Mechanical, Electronic and Industrial Engineering, University of Electronic Science and Technology of China, Chengdu 611731, Chinac Department of Mechanical Engineering, University of Washington, Seattle, WA 98195, USA
a r t i c l e i n f o a b s t r a c t
Article history:Received 7 November 2013Accepted 20 February 2014Available online 28 February 2014
Keywords:HEK293TCryomicroscopyCryopreservationWater transportIntracellular ice formation (IIF)Optimal cooling rate
The objective of this study is to determine the cryobiological characteristics of human embryonic kidney(HEK293T) cells. The cell membrane hydraulic conductivity (Lpg) and the activation energy of watertransport (ELp) were determined in the absence/presence of cryoprotectant agent (CPA), while thenucleation rate kinetic and thermodynamic parameters (XSCN
o and jSCNo ) were determined in the absence
of CPA. Since dehydration and intracellular ice formation (IIF) are two factors that may cause damage tocells during the freezing process, systematical freezing experiments were carried out at different coolingrates (5, 10, 15, 20, 30, and 60 �C/min) under the commercial available cryomicroscopy (FDCS 196, Link-ham, Waterfield, UK) to further explore the cryoinjury mechanism for HEK293T cells. By simultaneouslyfitting the water transport equation to the experimentally measured volumetric shrinkage data at 5, 10,and 15 �C/min, the ‘‘combined best fit’’ membrane permeability parameters for HEK293T cells in bothphosphate buffer saline (PBS) and CPA media (0.75M Me2SO in PBS) are determined. They are Lpg = 2.85 �10�14 m/s/Pa (0.17 lm/min/atm), ELp = 142.91 kJ/mol (34.13 kcal/mol) (R2 = 0.990), and Lpg[cpa] = 2.73 ±0.44 � 10�14 m/s/Pa (0.16 ± 0.03 lm/min/atm), ELp[cpa] = 152.52 ± 27.69 kJ/mol (36.42 ± 6.61 kcal/mol)(R2 = 0.993), respectively. An optimal cooling rate Bopt (the highest cooling rate without IIF) wasdetermined to be 14.24 �C/min in the absence of CPA. Additionally, the ice nucleation parameters (XSCN
o
and jSCNo ) were averaged to be 1.31 ± 0.11 � 108 m�2 s�1 and 7.67 ± 2.55 � 109 K5 for the cooling rates
20, 30, and 60 �C/min.� 2014 Elsevier Inc. All rights reserved.
Introduction
Cryopreservation refers to the techniques of long-term preser-vation of cells or tissues by cooling them to a certain low temper-ature (e.g. �196 �C) to reduce the metabolic rates to an extremelylow level. At low temperatures, any life activities includingbiochemical reactions tend to cease. However, damage to the cellsor tissues usually happens because of excessive dehydration andthe formation of a significant amount of intracellular ice duringthe freezing process [32]. Either cellular water loss in too slowfreezing or intracellular ice formation in too fast freezing has beenproven to be deleterious factors, which will decrease the post-thawsurvival rate [28,33]. Therefore, the optimal cooling rate does exist
between the ‘too low’ and ‘too high’ freezing rates. Whether a cool-ing rate is ‘too low’ or ‘too high’ could be evaluated using a functionof cell membrane permeability to water and the probability ofintracellular ice formation. Difference in membrane permeabilityto water and probability of IIF leads to different optimal coolingrates. In general, the optimal cooling rate is defined to be theone, which minimizes both the solute injury and the IIF injury,i.e. cool as fast as possible without forming damaging intracellularice [35]. On the other hand, IIF has always been considered as themore lethal factor due to its destruction to the subcellularstructure and the cell membrane [8,9]. Thus, study on themechanism of intracellular ice formation and its growth turns tobe of great importance as to applying cryogenic technology to cellpreservation [5,16,30].
Human embryonic kidney cells (HEK293T), a specific cell lineoriginally derived from human embryonic kidney cells grown intissue culture, have been widely used in cell biology researchinvolving transfecting in a gene and then analyzing the expressedprotein or the propagation of adenoviral vectors [1,15,24,37]. They

Y. Xu et al. / Cryobiology 68 (2014) 294–302 295
are also used by the biotechnology industry to produce therapeuticproteins and viruses for gene therapy [18,27]. Cryopreservation isan important issue in such application. So far, cryopreservationof the HEK293T cells is still challenging in minimizing cryoinjuryto improve the post-thaw survival rate. Fundamental cryobiologi-cal characteristics, including the permeability of the cell mem-brane to water and the probability of IIF of the HEK293T cells,are necessary for optimization of the cryopreservation protocols.
In this work, a comprehensive study was carried out to betterunderstand the biotransport and IIF phenomena during freezingof HEK293T cells. To avoid overlarge supercooling, ice seedingwas applied to induce extracellular ice formation at a specificsubzero temperature. To clearly observe the cellular response tofreezing, the water transport and intracellular ice formationphenomena were studied under the cryomicroscopy [3,9,45,46].The experimental data was fitted to the widely used models ofwater transport and ice nucleation to calculate the cryobiologicalcharacteristics of HEK293T cells. Then, the influence of CPA onthe water permeability parameters was evaluated. Eventually,the optimal cooling rate was predicted theoretically based on thewater permeability parameters.
Materials and methods
Cell collection and culture
The HEK293T cells were obtained from Prof. Guoqiang Bi’s Labat USTC as a gift [40]. The cells were cultured in Dulbecco’smodified Eagle’s medium (Hyclone, Thermo Fisher Scientific Inc.,UK) with the 10% FBS (Hyclone, Thermo Fisher Scientific Inc.,UK), 100 U/L penicillin G and 100 U/L streptomycin (Hyclone,Thermo Fisher Scientific Inc., UK) at 37 �C and 5% CO2 incubator(MCO-5AC, SANYO, Japan). The cells were collected at approxi-mately 80% confluence. The adhered cells were treated with0.25% Trypsin (Biosharp, China) after washing with 1 � PBS. Thenthe cells were centrifuged at 80�g for 5 min and re-suspended to1 � 105/ml with 1 � PBS.
Cryomicroscopy study
For cryomicroscopy imaging study of HEK293T cells, a speciallymade copper wire mounted on the edge of a cryostage’s surfacewas used for ice-seeding in order to keep the moisture from enter-ing the cryostage. The traditional ice-seeding method, using asyringe needle precooled in liquid nitrogen, would lead to a dimvision during freezing and cause a negative influence on subse-quent observation. Prior to the cooling step, a circular cover glass(16 mm in diameter) was placed on the top of the cryostage to holdthe cell suspension, then a small drop (�5lL) of 1 � PBS withHEK293T cells was pipetted onto the center of the cover glass withanother cover glass covered on it slightly. The Linkham cryostagewas fixed to a microscope (BX 53, Olympus, Japan). Meanwhile, aQICAM (Qimaging, Survey, Canada) was used for real-timemonitoring of both water dehydration and intracellular ice forma-tion during freezing (resolution, 960 � 720; frame rate, 5fps).
During the HEK293T cells’ freezing experiment, both celldehydration and intracellular ice formation were investigated withice-seeding. Cells were firstly cooled from room temperature(�23 �C) to �1 �C (�3 �C for 0.75M Me2SO) with a cooling rate of10 �C/min. The temperature was maintained at �1 �C (�3 �C for0.75M Me2SO) for 100 s in order to induce ice-seeding with thecopper wire. Then the sample was rewarmed to �0.5 �C (�2.5 �Cfor 0.75M Me2SO) temporarily with a heating rate of 10 �C/minand held for 180 s to achieve equilibrium. This procedure couldreduce the influence of ice-seeding on initial cell volume and
provide a better visualization during subsequent freezing. After-wards, the cells were further cooled to a deep subzero temperature(e.g. �50 �C) with different cooling rates of 5, 10, 15, 20, 30, and60 �C/min. Finally, the cells were rewarmed to the room tempera-ture at the heating rate of 50 �C/min. The whole processes of freez-ing and thawing were observed under a 50� objective lens withlong working distance.
Modeling of water transportation during freezing
In a state of thermodynamic equilibrium, the chemical potentialof water in the extracellular solution is equal to that in the intracel-lular cytoplasm. When extracellular ice forms due to ice-seeding,extracellular solution tends to be concentrated and intracellularwater begins to permeate through the membrane to restoreequilibrium. The water transport phenomenon can be describedby the following equations originally proposed by Mazur and latermodified by Levin et al. and Fahy [12,26,29,31]:
dVc
dT¼ � LpAcRT
Bmw� DHf
Rð 1TR� 1
TÞ � ln
Vc � Vb � Vs
Vc � Vb � Vs þ /snsvw
� �ð1Þ
Lp ¼ Lpg exp � ELp
R1T� 1
TR
� �� �: ð2Þ
The above water transport model was further modified byKarlsson et al. to incorporate the influence of CPA on the cellvolume during freezing as follows [25]:
dVc
dT¼�LpAcRT
Bvw
� DHf
R1TR�1
T
� �� ln
Vc�Vb�Vs�ncpavcpa
Vc�Vb�Vs�ncpavcpaþð/snsþncpaÞvw
� �ð3Þ
Lp ¼ Lpg ½cpa� exp �ELp½cpa�R
1T� 1
TR
� �� �ð4Þ
where Vc is the cell volume (lm3); T is the absolute temperature(K); Ac is the cell surface area (lm2); R is the universal gas constant(8.314 J mol�1 K�1); B is the cooling rate (�C/min); DHf is the latentheat of water fusion (6016.52 J mol�1); TR is the reference tempera-ture; Vb is the osmotically inactive volume of the cell (lm3); Vs isthe volume of salt (lm3); /s is the dissociation constant of NaCl(2 for NaCl); ns is the number of moles of NaCl (mol); ncpa is thenumber of moles of CPA in solution; vcpa is the molar volume ofCPA (in this study, 69 � 1012 lm3/mol for 0.75M Me2SO) [17]; vw
is the molar volume of water (18.02 � 1012 lm3/mol); Lpg is themembrane hydraulic conductivity at the reference temperature of272.65 K and Lpg[cpa] at 270.65 K (m/s/Pa or lm/min/atm); ELp orELp[cpa] is the membrane permeability activation energy (kJ/molor kcal/mol).
Prediction of the optimal cooling rate Bopt
Once the cryobiological parameters are determined, the optimalcooling rate Bopt, defined as the highest cooling rate that traps 5% ofthe initial intracellular water at the end temperature of �15 �C, canbe calculated by incorporating these parameters into the GenericOptimal Cooling Rate Equation (GOCRE) as follows [39]:
Bopt ¼ 1009:5 expð�0:0546ElpÞ � Lpg �SA
WV
� �ð5Þ
where SA is the available cell membrane surface area (lm2); SAWV is
the ratio of the available surface for water transport to the initialvolume of intracellular water (WV = Vo � Vb, Vo is the isotonic orinitial cell volume and Vb is the osmotically inactive cell volume).

296 Y. Xu et al. / Cryobiology 68 (2014) 294–302
Modeling of probability of intracellular ice formation during freezing
The probability of IIF (PIF) parameters can be determined usingthe SCN models as follows [42,45]:
PSCNIIF ¼ 1� exp �
Z t
0AcISCNdt
� �
¼ 1� exp �Z Tfo
TAcISCNð1=BÞdT
� �ð6Þ
ISCN ¼ XSCNo
NSCN
NSCNo
g0
gT
Tfo
� �0:5
exp�jSCN
o ðTf =TfoÞ4
ðT � Tf Þ2T3
" #ð7Þ
where ‘o’ refers to isotonic condition; t is the time; Tfo is the equilib-rium freezing temperature under the isotonic condition (272.65 K);ISCN refers to the nucleation rate of surface-catalyzed; XSCN
o is thepre-exponential factor (m�2 s�1); jSCN
o is the exponential factor(K5); NSCN is the number of water molecules in contract with the
substrate; and NSCN
NSCNo
can be simplified as A/Ao, where A is the surface
area of the plasma membrane; Tf is the equilibrium freezing tem-perature, and it can be calculated using the following equation [26]:
Tf ¼1
Tfo� R
DHf
� �� ln
nw
nw þ ns
� �� ��1
: ð8Þ
g is the viscosity of the cytoplasm, and it can be estimated withthe undermentioned equations [26,45]:
g ¼ gw exp2:5us
1� 0:61us
� �ð9Þ
where us is the volume fraction of salts, and gw is the viscosity ofpure water described using the following free volume model[21,45]:
gw ¼ gw;o expv�w
ðK11=kÞðK21 � Tgw þ TÞ
� �ð10Þ
where v�w is the specific volume at 0 K (0.91 ml g�1); k is the overlapfactor because the same free volume is available to more than onemolecule; K11/k (1.945 ml g�1 K�1) and K21 (�19.73 K) are two freevolume parameters for solvent (water); Tgw is the glass transitiontemperature of water (136 K). The values for these parameters inthis study were adopted from Ref. [45].
Cryobiological parameter estimation
The model parameters for water transport (Lpg, ELp or Lpg[cpa],ELp[cpa]) and IIF (XSCN
o , jSCNo ) were determined by fitting the exper-
imental data to the above models. The HEK293T cell’s osmoticallyinactive volume, Vb was not derived by the conventional Boyle-Van’t Hoff method as described in Ref. [3]. Instead, it was obtainedby a combined-fitting method together with water permeabilityparameters, Lpg and ELp, as has been accomplished by Zhao [48].This method was also adopted for the recent study of our groupon the permeability coefficients of SF21 cells [44]. The mean cell-uar volumes with the relevant standard errors during freezingwere calculated prior to fitting. The fitting was accomplished basedon the nonlinear least-squares strategy. The best fit waterpermeability parameters were determined by adjusting the initialvalue of the model parameters to approach a maximum R2. Then,the nucleation rate kinetic and thermodynamic parameters (XSCN
o
and jSCNo ) were obtained by fitting the PIF data to the SCN model,
coupled with the model of cell dehydration during freezing[3,19,20,38,45].
Results
Water transport
Fig. 1 showed HEK293T cells’ morphology at �0.5 �C after ice-seeding (A) and the transient volume change of a representativecell during freezing at 5 �C intervals (B). Clearly, the cell shrankalong with the outflow of cell water during freezing. The cell wasassumed spherical to quantitatively determine its volume fromthe projected area. Those non-spherical, over-large or over-smallcells were excluded.
The normalized cell volumes (the transient cell volume dividedby the volume at the ice-seeding temperature, i.e., Vc/Vc,ice-seeding)versus temperature at three different cooling rates was depictedin Fig. 2. The initial volume referred to the one at �0.5 �C, i.e. theequilibrium temperature. The temperature for cell dehydrationapproximately ranged from the �0.5 to �25.0 �C. The equilibriumwas retained later as the cooling rate rose from 5 to 15 �C/min. Itwas also noticed that there was no significant cell volume changebelow �20 �C for all the cooling rates. At slow cooling rate of 5 �C/min, the cells dehydrated excessively. In contrast, at cooling rate of15 �C/min, the cell dehydration process tended to be more gradual.This phenomenon may be attributed to the strong temperaturedependence of the membrane hydraulic conductivity, that is, Lpg
has dropped sharply as the temperature decreased. When the cellswere cooled at 5 �C/min, only cell dehydration phenomenon wasobserved (no IIF), indicating that in this circumstance, the cell in-jury was mainly caused by excessive dehydration. In contrast, a lit-tle intracellular ice formation was observed at 10 �C/min in thecooling process. However, at 15 �C/min, both cell dehydrationand IIF were observed, indicating that the cells underwent twotypes of injuries. Similar phenomenon was also observed in thefreezing of peromyscus oocytes [45]. After thawing, most of thecells lost their membrane integrity and their regular shapes, indi-cating some kinds of direct injuries.
The HEK293T cells’ volumetric responses to different coolingrates were investigated in the present study. Fig. 2A–C depictedthe average normalized cell volume versus temperature at the cool-ing rates of 5, 10, and 15 �C/min. The open symbols of circle, uppertriangle, and lower triangle represented the measured cell volu-metric responses at 5 (22 cells), 10 (17 cells), and 15 (13cells) �C/min respectively, while the solid lines represented thepredicted cell volumes at the corresponding cooling rates. Sincethe fitting process was accomplished separately for each coolingrate, it is called ‘‘separate-fitting’’, while there is also a ‘‘com-bined-fitting’’ where all parameters are estimated simultaneouslyusing all the experimental data in spite of the cooling rates. Com-paratively, a composite curve obtained by the combined-fittingwas given in Fig. 2D. The corresponding water permeability param-eters were Lpg = 2.85 � 10�14 m/s/Pa (0.17 lm/min/atm) at the ref-erence temperature 272.65 K, and ELp = 142.91 kJ/mol (34.13 kcal/mol). These parameters would be used in the subsequent fittingfor intracellular ice formation.
The fitting results along with the goodness of fit (R2) at differentcooling rates were shown in Table 1. An extraordinary good corre-lations were acquired between all the experimental data and thetheoretical model, where a goodness of fit ranged from 0.990 to0.998. The average values of the separate-fitting wereLpg = 3.02 ± 0.28 � 10�14 m/s/Pa (0.18 ± 0.02 lm/min/atm) at thereference temperature of 272.65 K, and ELp = 145.33 ± 6.57 kJ/mol(34.71 ± 1.57 kcal/mol). The combined-fitting and the seperate-fitting methods demonstrated very similar results, for both theR-squared and the parameter values. The derived parameters inthe present work were compared with some other types of humancells in Table 2. The Vb/Vo value of HEK293T cells, 0.29 determined
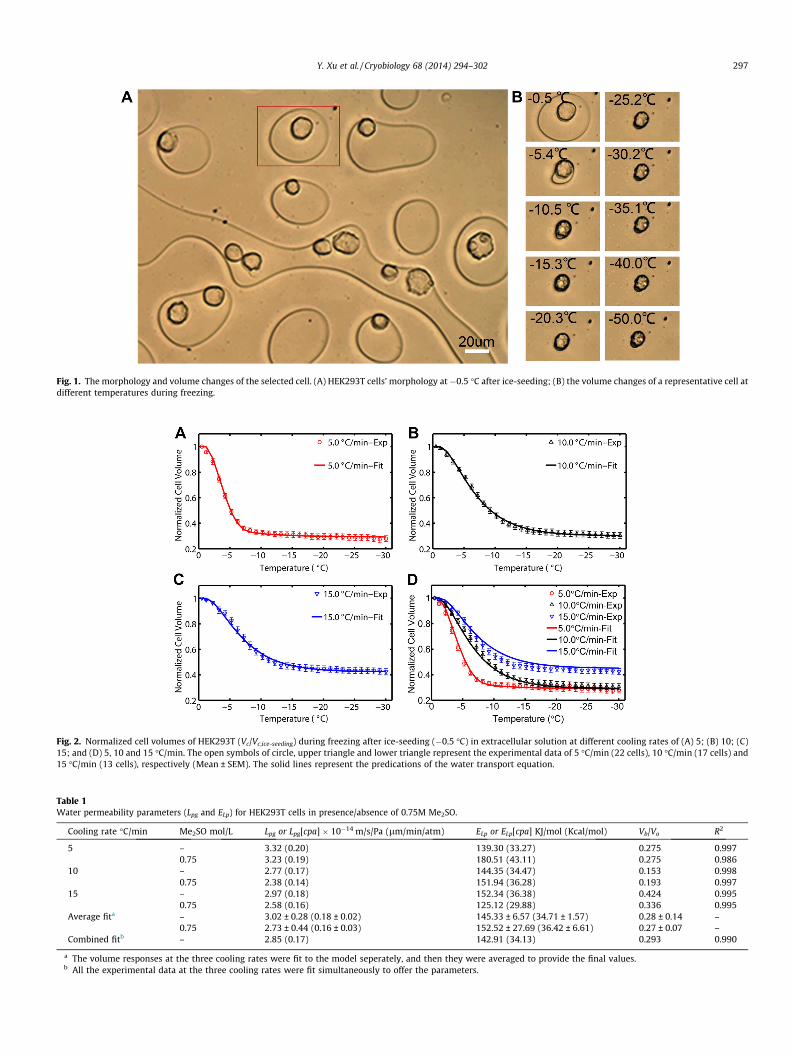
Fig. 1. The morphology and volume changes of the selected cell. (A) HEK293T cells’ morphology at�0.5 �C after ice-seeding; (B) the volume changes of a representative cell atdifferent temperatures during freezing.
Fig. 2. Normalized cell volumes of HEK293T (Vc/Vc,ice-seeding) during freezing after ice-seeding (�0.5 �C) in extracellular solution at different cooling rates of (A) 5; (B) 10; (C)15; and (D) 5, 10 and 15 �C/min. The open symbols of circle, upper triangle and lower triangle represent the experimental data of 5 �C/min (22 cells), 10 �C/min (17 cells) and15 �C/min (13 cells), respectively (Mean ± SEM). The solid lines represent the predications of the water transport equation.
Table 1Water permeability parameters (Lpg and ELp) for HEK293T cells in presence/absence of 0.75M Me2SO.
Cooling rate �C/min Me2SO mol/L Lpg or Lpg[cpa] � 10�14 m/s/Pa (lm/min/atm) ELp or ELp[cpa] KJ/mol (Kcal/mol) Vb/Vo R2
5 – 3.32 (0.20) 139.30 (33.27) 0.275 0.9970.75 3.23 (0.19) 180.51 (43.11) 0.275 0.986
10 – 2.77 (0.17) 144.35 (34.47) 0.153 0.9980.75 2.38 (0.14) 151.94 (36.28) 0.193 0.997
15 – 2.97 (0.18) 152.34 (36.38) 0.424 0.9950.75 2.58 (0.16) 125.12 (29.88) 0.336 0.995
Average fita – 3.02 ± 0.28 (0.18 ± 0.02) 145.33 ± 6.57 (34.71 ± 1.57) 0.28 ± 0.14 –0.75 2.73 ± 0.44 (0.16 ± 0.03) 152.52 ± 27.69 (36.42 ± 6.61) 0.27 ± 0.07 –
Combined fitb – 2.85 (0.17) 142.91 (34.13) 0.293 0.990
a The volume responses at the three cooling rates were fit to the model seperately, and then they were averaged to provide the final values.b All the experimental data at the three cooling rates were fit simultaneously to offer the parameters.
Y. Xu et al. / Cryobiology 68 (2014) 294–302 297
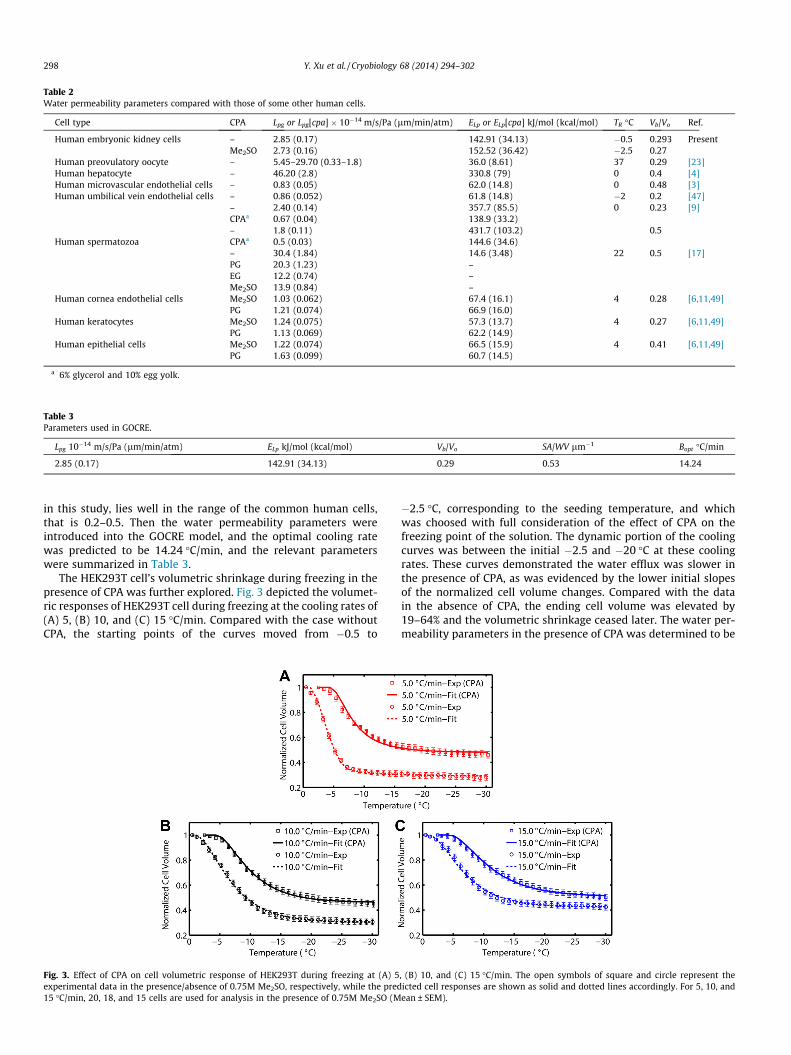
Table 2Water permeability parameters compared with those of some other human cells.
Cell type CPA Lpg or Lpg[cpa] � 10�14 m/s/Pa (lm/min/atm) ELp or ELp[cpa] kJ/mol (kcal/mol) TR �C Vb/Vo Ref.
Human embryonic kidney cells – 2.85 (0.17) 142.91 (34.13) �0.5 0.293 PresentMe2SO 2.73 (0.16) 152.52 (36.42) �2.5 0.27
Human preovulatory oocyte – 5.45–29.70 (0.33–1.8) 36.0 (8.61) 37 0.29 [23]Human hepatocyte – 46.20 (2.8) 330.8 (79) 0 0.4 [4]Human microvascular endothelial cells – 0.83 (0.05) 62.0 (14.8) 0 0.48 [3]Human umbilical vein endothelial cells – 0.86 (0.052) 61.8 (14.8) �2 0.2 [47]
– 2.40 (0.14) 357.7 (85.5) 0 0.23 [9]CPAa 0.67 (0.04) 138.9 (33.2)– 1.8 (0.11) 431.7 (103.2) 0.5
Human spermatozoa CPAa 0.5 (0.03) 144.6 (34.6)– 30.4 (1.84) 14.6 (3.48) 22 0.5 [17]PG 20.3 (1.23) –EG 12.2 (0.74) –Me2SO 13.9 (0.84) –
Human cornea endothelial cells Me2SO 1.03 (0.062) 67.4 (16.1) 4 0.28 [6,11,49]PG 1.21 (0.074) 66.9 (16.0)
Human keratocytes Me2SO 1.24 (0.075) 57.3 (13.7) 4 0.27 [6,11,49]PG 1.13 (0.069) 62.2 (14.9)
Human epithelial cells Me2SO 1.22 (0.074) 66.5 (15.9) 4 0.41 [6,11,49]PG 1.63 (0.099) 60.7 (14.5)
a 6% glycerol and 10% egg yolk.
Table 3Parameters used in GOCRE.
Lpg 10�14 m/s/Pa (lm/min/atm) ELp kJ/mol (kcal/mol) Vb/Vo SA/WV lm�1 Bopt �C/min
2.85 (0.17) 142.91 (34.13) 0.29 0.53 14.24
298 Y. Xu et al. / Cryobiology 68 (2014) 294–302
in this study, lies well in the range of the common human cells,that is 0.2–0.5. Then the water permeability parameters wereintroduced into the GOCRE model, and the optimal cooling ratewas predicted to be 14.24 �C/min, and the relevant parameterswere summarized in Table 3.
The HEK293T cell’s volumetric shrinkage during freezing in thepresence of CPA was further explored. Fig. 3 depicted the volumet-ric responses of HEK293T cell during freezing at the cooling rates of(A) 5, (B) 10, and (C) 15 �C/min. Compared with the case withoutCPA, the starting points of the curves moved from �0.5 to
Fig. 3. Effect of CPA on cell volumetric response of HEK293T during freezing at (A) 5experimental data in the presence/absence of 0.75M Me2SO, respectively, while the pred15 �C/min, 20, 18, and 15 cells are used for analysis in the presence of 0.75M Me2SO (M
�2.5 �C, corresponding to the seeding temperature, and whichwas choosed with full consideration of the effect of CPA on thefreezing point of the solution. The dynamic portion of the coolingcurves was between the initial �2.5 and �20 �C at these coolingrates. These curves demonstrated the water efflux was slower inthe presence of CPA, as was evidenced by the lower initial slopesof the normalized cell volume changes. Compared with the datain the absence of CPA, the ending cell volume was elevated by19–64% and the volumetric shrinkage ceased later. The water per-meability parameters in the presence of CPA was determined to be
, (B) 10, and (C) 15 �C/min. The open symbols of square and circle represent theicted cell responses are shown as solid and dotted lines accordingly. For 5, 10, andean ± SEM).
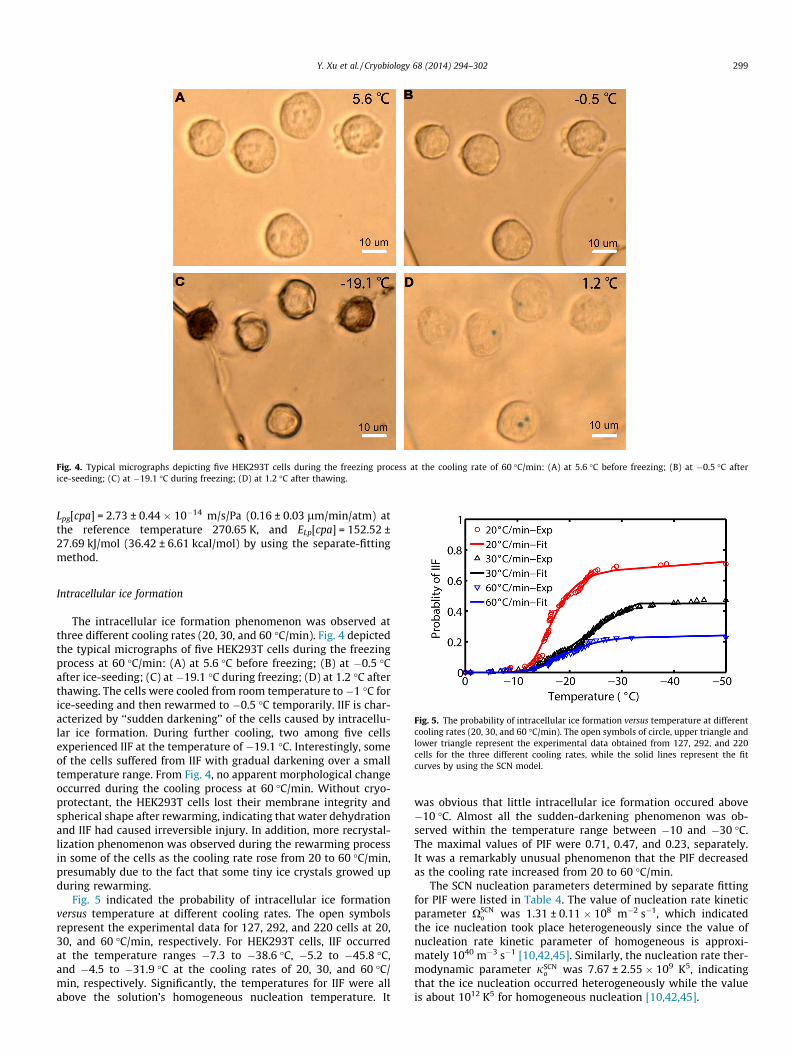
Fig. 4. Typical micrographs depicting five HEK293T cells during the freezing process at the cooling rate of 60 �C/min: (A) at 5.6 �C before freezing; (B) at �0.5 �C afterice-seeding; (C) at �19.1 �C during freezing; (D) at 1.2 �C after thawing.
Y. Xu et al. / Cryobiology 68 (2014) 294–302 299
Lpg[cpa] = 2.73 ± 0.44 � 10�14 m/s/Pa (0.16 ± 0.03 lm/min/atm) atthe reference temperature 270.65 K, and ELp[cpa] = 152.52 ±27.69 kJ/mol (36.42 ± 6.61 kcal/mol) by using the separate-fittingmethod.
Fig. 5. The probability of intracellular ice formation versus temperature at differentcooling rates (20, 30, and 60 �C/min). The open symbols of circle, upper triangle andlower triangle represent the experimental data obtained from 127, 292, and 220cells for the three different cooling rates, while the solid lines represent the fitcurves by using the SCN model.
Intracellular ice formation
The intracellular ice formation phenomenon was observed atthree different cooling rates (20, 30, and 60 �C/min). Fig. 4 depictedthe typical micrographs of five HEK293T cells during the freezingprocess at 60 �C/min: (A) at 5.6 �C before freezing; (B) at �0.5 �Cafter ice-seeding; (C) at �19.1 �C during freezing; (D) at 1.2 �C afterthawing. The cells were cooled from room temperature to �1 �C forice-seeding and then rewarmed to �0.5 �C temporarily. IIF is char-acterized by ‘‘sudden darkening’’ of the cells caused by intracellu-lar ice formation. During further cooling, two among five cellsexperienced IIF at the temperature of �19.1 �C. Interestingly, someof the cells suffered from IIF with gradual darkening over a smalltemperature range. From Fig. 4, no apparent morphological changeoccurred during the cooling process at 60 �C/min. Without cryo-protectant, the HEK293T cells lost their membrane integrity andspherical shape after rewarming, indicating that water dehydrationand IIF had caused irreversible injury. In addition, more recrystal-lization phenomenon was observed during the rewarming processin some of the cells as the cooling rate rose from 20 to 60 �C/min,presumably due to the fact that some tiny ice crystals growed upduring rewarming.
Fig. 5 indicated the probability of intracellular ice formationversus temperature at different cooling rates. The open symbolsrepresent the experimental data for 127, 292, and 220 cells at 20,30, and 60 �C/min, respectively. For HEK293T cells, IIF occurredat the temperature ranges �7.3 to �38.6 �C, �5.2 to �45.8 �C,and �4.5 to �31.9 �C at the cooling rates of 20, 30, and 60 �C/min, respectively. Significantly, the temperatures for IIF were allabove the solution’s homogeneous nucleation temperature. It
was obvious that little intracellular ice formation occured above�10 �C. Almost all the sudden-darkening phenomenon was ob-served within the temperature range between �10 and �30 �C.The maximal values of PIF were 0.71, 0.47, and 0.23, separately.It was a remarkably unusual phenomenon that the PIF decreasedas the cooling rate increased from 20 to 60 �C/min.
The SCN nucleation parameters determined by separate fittingfor PIF were listed in Table 4. The value of nucleation rate kineticparameter XSCN
o was 1.31 ± 0.11 � 108 m�2 s�1, which indicatedthe ice nucleation took place heterogeneously since the value ofnucleation rate kinetic parameter of homogeneous is approxi-mately 1040 m�3 s�1 [10,42,45]. Similarly, the nucleation rate ther-modynamic parameter jSCN
o was 7.67 ± 2.55 � 109 K5, indicatingthat the ice nucleation occurred heterogeneously while the valueis about 1012 K5 for homogeneous nucleation [10,42,45].
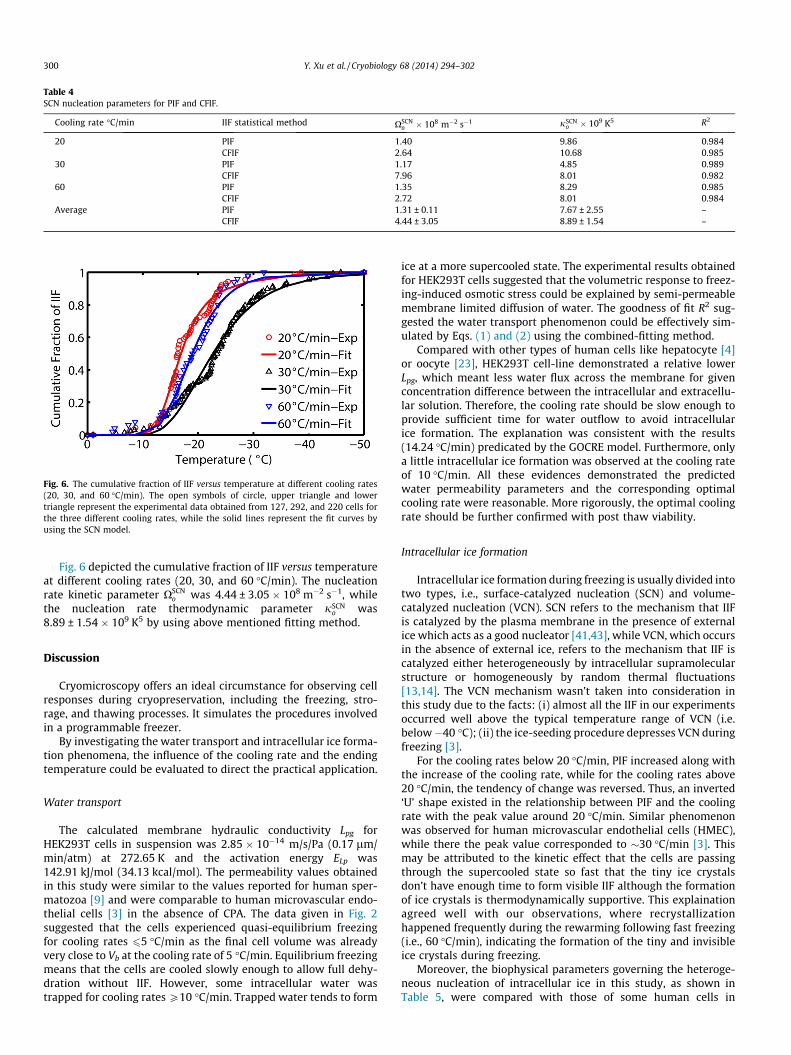
Table 4SCN nucleation parameters for PIF and CFIF.
Cooling rate �C/min IIF statistical method XSCNo � 108 m�2 s�1 jSCN
o � 109 K5 R2
20 PIF 1.40 9.86 0.984CFIF 2.64 10.68 0.985
30 PIF 1.17 4.85 0.989CFIF 7.96 8.01 0.982
60 PIF 1.35 8.29 0.985CFIF 2.72 8.01 0.984
Average PIF 1.31 ± 0.11 7.67 ± 2.55 –CFIF 4.44 ± 3.05 8.89 ± 1.54 –
Fig. 6. The cumulative fraction of IIF versus temperature at different cooling rates(20, 30, and 60 �C/min). The open symbols of circle, upper triangle and lowertriangle represent the experimental data obtained from 127, 292, and 220 cells forthe three different cooling rates, while the solid lines represent the fit curves byusing the SCN model.
300 Y. Xu et al. / Cryobiology 68 (2014) 294–302
Fig. 6 depicted the cumulative fraction of IIF versus temperatureat different cooling rates (20, 30, and 60 �C/min). The nucleationrate kinetic parameter XSCN
o was 4.44 ± 3.05 � 108 m�2 s�1, whilethe nucleation rate thermodynamic parameter jSCN
o was8.89 ± 1.54 � 109 K5 by using above mentioned fitting method.
Discussion
Cryomicroscopy offers an ideal circumstance for observing cellresponses during cryopreservation, including the freezing, stro-rage, and thawing processes. It simulates the procedures involvedin a programmable freezer.
By investigating the water transport and intracellular ice forma-tion phenomena, the influence of the cooling rate and the endingtemperature could be evaluated to direct the practical application.
Water transport
The calculated membrane hydraulic conductivity Lpg forHEK293T cells in suspension was 2.85 � 10�14 m/s/Pa (0.17 lm/min/atm) at 272.65 K and the activation energy ELp was142.91 kJ/mol (34.13 kcal/mol). The permeability values obtainedin this study were similar to the values reported for human sper-matozoa [9] and were comparable to human microvascular endo-thelial cells [3] in the absence of CPA. The data given in Fig. 2suggested that the cells experienced quasi-equilibrium freezingfor cooling rates 65 �C/min as the final cell volume was alreadyvery close to Vb at the cooling rate of 5 �C/min. Equilibrium freezingmeans that the cells are cooled slowly enough to allow full dehy-dration without IIF. However, some intracellular water wastrapped for cooling rates P10 �C/min. Trapped water tends to form
ice at a more supercooled state. The experimental results obtainedfor HEK293T cells suggested that the volumetric response to freez-ing-induced osmotic stress could be explained by semi-permeablemembrane limited diffusion of water. The goodness of fit R2 sug-gested the water transport phenomenon could be effectively sim-ulated by Eqs. (1) and (2) using the combined-fitting method.
Compared with other types of human cells like hepatocyte [4]or oocyte [23], HEK293T cell-line demonstrated a relative lowerLpg, which meant less water flux across the membrane for givenconcentration difference between the intracellular and extracellu-lar solution. Therefore, the cooling rate should be slow enough toprovide sufficient time for water outflow to avoid intracellularice formation. The explanation was consistent with the results(14.24 �C/min) predicated by the GOCRE model. Furthermore, onlya little intracellular ice formation was observed at the cooling rateof 10 �C/min. All these evidences demonstrated the predictedwater permeability parameters and the corresponding optimalcooling rate were reasonable. More rigorously, the optimal coolingrate should be further confirmed with post thaw viability.
Intracellular ice formation
Intracellular ice formation during freezing is usually divided intotwo types, i.e., surface-catalyzed nucleation (SCN) and volume-catalyzed nucleation (VCN). SCN refers to the mechanism that IIFis catalyzed by the plasma membrane in the presence of externalice which acts as a good nucleator [41,43], while VCN, which occursin the absence of external ice, refers to the mechanism that IIF iscatalyzed either heterogeneously by intracellular supramolecularstructure or homogeneously by random thermal fluctuations[13,14]. The VCN mechanism wasn’t taken into consideration inthis study due to the facts: (i) almost all the IIF in our experimentsoccurred well above the typical temperature range of VCN (i.e.below�40 �C); (ii) the ice-seeding procedure depresses VCN duringfreezing [3].
For the cooling rates below 20 �C/min, PIF increased along withthe increase of the cooling rate, while for the cooling rates above20 �C/min, the tendency of change was reversed. Thus, an inverted‘U’ shape existed in the relationship between PIF and the coolingrate with the peak value around 20 �C/min. Similar phenomenonwas observed for human microvascular endothelial cells (HMEC),while there the peak value corresponded to �30 �C/min [3]. Thismay be attributed to the kinetic effect that the cells are passingthrough the supercooled state so fast that the tiny ice crystalsdon’t have enough time to form visible IIF although the formationof ice crystals is thermodynamically supportive. This explainationagreed well with our observations, where recrystallizationhappened frequently during the rewarming following fast freezing(i.e., 60 �C/min), indicating the formation of the tiny and invisibleice crystals during freezing.
Moreover, the biophysical parameters governing the heteroge-neous nucleation of intracellular ice in this study, as shown inTable 5, were compared with those of some human cells in

Table 5SCN nucleation parameters compared with those of some other human cells.
Cell type CPA XSCNo � 108 m�2 s�1 jSCN
o � 109 K5 B �C/min Ref.
Human embryonic kidney cells – 1.31 7.67 20, 30, 60 PresentHuman hepatocyte – 7.6 7.5 – [7]
Glucose 10.8 1.04 100 [4]Human microvascular endothelial cells – 6.87a 8.36 25, 50, 100, 130 [3]Human dermal-fibroblast – 0.92 1.2 130 [2]
a Obtained with combine-fitting for the cooling rates 25, 50, 100, and 130 �C/min.
Y. Xu et al. / Cryobiology 68 (2014) 294–302 301
literatures. The values of HEK293T cells were very close to those ofthe HMECs [3], and they were also comparable to those of humanhepatocytes [4] and human dermal-fibroblast [2]. It implied thatthe parameters for SCN with ice seeding measured in this study werereasonable.
Probability of IIF and cumulative fraction of IIF
The concepts of probability of IIF (PIF) and the cumulative frac-tion of IIF (CFIF) are somewhat confused, and are often lumped to-gether or even mixed up. Either PIF or CFIF is often used to describethe mechanism of nucleation, and to fit the ice nucleation kineticand thermodynamic parameters.
To further comprehend the mechanism of intracellular ice for-mation, CFIF defined as follows [46] was discussed and comparedwith PIF for all the cooling rates.
PIF¼ Number of cells undergone IIF at a certain temperatureNumber of cells in the observed field ðsuffered IIF plus no IIFÞ
CFIF ¼ Number of cells suffered IIF at a certain temperatureTotal number of cells suffered IIF
All the cells in the observed field are taken into account in thedefinition of PIF, while only the cells suffered IIF are referred toin the definition of CFIF.
Comparatively, the CFIF data was also fitted using the same for-mulas, and the corresponding results were listed in Table 4. Evi-dently, the parameters for CFIF, XSCN
o and jSCNo , were somewhat
bigger than those for PIF, especially for the pre-exponential factorXSCN
o . According to Figs. 5 and 6, and Table 4, the effect of increasedXSCN
o was to shift the kinetics of IIF to higher temperatures, while atthe same time the range of temperatures over which IIF occuredwas narrower. Another interesting phenomenon was that the CFIFcurve for 60 �C/min was higher than that for 30 �C/min in the tem-perature range �10 to �35 �C, while for the PIF curve, as was re-verse. This result indicated that the intracellular ice formationwere not uniformly scattered in the temperature range duringfreezing. For 60 �C/min, IIF concentrated in relative higher temper-ature intervals. It should be pointed out that the ice nuclei formedat lower temperatures are sometimes too tiny to be detected underthe light microscropy. In conclusion, the comparative results indi-cated that the CFIF was a more quantitative and detailed descrip-tion of the temperature distribution of the incident of ‘‘suddendarkening’’ IIF, while PIF depicted the phenomenon of IIF from aglobal perspective and revealed the fundamental effect of freezingon a group of cells. Therefore, it seems that PIF is more suitable forstudying the features of intracellular nucleation from the perspec-tive of species, while CFIF is more appropriate for research on spe-cies internal characteristics.
The influence of CPA
The model parameters obtained in the absence of CPA in thisstudy was indispensable for theoretical simulations according tothe diffusion-limited IIF models developed by Karlsson et al. [26]
and later improved by Zhao et al. [50]. The model makes it possibleto predict the critical cooling rate versus the initial cryoprotectantconcentration to design an optimal cryopreservation protocol for agiven biological cell. Therefore, the parameters acquired in thiswork can be applied to predict the probability of IIF and the growthof intracellular crystals in HEK293T cells under various CPA con-centration and cooling rates.
However, many former studies reported that the presence ofCPA would decrease the membrane hydraulic conductivity (Lpg),although the exact mechanism is as yet unknown. In the presentwork, the cryomicroscopy was further used to evaluate the influ-ence of CPA on the water permeability parameters. Similar effectof CPA on Lpg was observed in our experimental data, where anaverage of 9.6% drop of the water membrane permeability oc-curred, i.e., from 3.02 ± 0.28 � 10�14 m/s/Pa (0.18 ± 0.02 lm/min/atm) in the absence of CPA to 2.73 ± 0.44 � 10�14 m/s/Pa(0.16 ± 0.03 lm/min/atm) in the presence of 0.75M Me2SO. Thisconclusion was further confirmed by the date of the previous liter-atures. Table 2 depicted the comparisons of some human cells’water permeability parameters in the absence/presence of CPA.Devireddy et al. demonstrated that an increase of the solutes’ con-centration (CPA: 6% glycerol and 10% egg yolk) in the extracellularmedium would decrease Lpg of human sperm cells by 70% [9]. Sim-ilar effects were observed that the values of Lpg obtained in thepresence of different CPA [PG, EG, and Me2SO] were 30–60% lowerthan those without CPAs [17]. Hempling and White reported thatrat megakaryocytopoietic cells equilibrated in 0.6M Me2SO in-creased their osmotically inactive volume, while reduced thehydraulic conductivity by 50% [22]. As to the activation energy,ELp, no such consistant conclusion was drawn to date. Our experi-mental data demonstrated the similar consequence for ELp, wherethe introduction of CPA increased it for the cooling rates 5 and10 �C/min, while decreased it for 15 �C/min, when separate-fittingwas applied. Mazur demonstrated that ELp remained unchangeddue to the increase of the extracellular concentration [34], whileGilmore et al. measured an increase in the value of ELp for humansperm cells [17]. Mccaa et al. pointed out that human monocytespre-equilibrated with Me2SO had a lower Lpg and a higher activa-tion energy than the ones without additive [36].
Due to the fact that the effective functional relationship be-tween CPA type, CPA concentration and water permeability param-eters is as yet unknown, and the fact that few studies have beenfocused on the impact of CPA on intracellular ice formation, furthersystematical investigations are necessary.
Summary and conclusions
In this work, comprehensive and systematical experimentswere carried out to explore the mechanism of water transportand probability of intracellular ice formation during freezing ofHEK293T cells at different cooling rates. For better visualizationduring freezing, a new method of ice-seeding with copper wirewas put forward for the first time to avoid the moisture fromentering the cryostage. The water permeability parameters (Lpg,ELp) and the probability of intracellular ice formation parameters(XSCN
o , jSCNo ) were determined by fitting the experimental data to

302 Y. Xu et al. / Cryobiology 68 (2014) 294–302
the water transport and SCN equations. Rather than the conven-tional method for determination of Vb by using the Boyle-Van’tHoff equation, a combined fit approach was used to calculate Vb,which turned out to own an outstanding goodness of fit. Further-more, an optimal cooling rate (the highest cooling rate withoutIIF) of 14.24 �C/min was predicted using the GOCRE method. Fur-ther experiments with 0.75M Me2SO were implemented to evalu-ate the effect of CPA on water permeability parameters. Similaritiesand differences of PIF and CFIF were discussed in detail accordingto the experimentally obtained curves in this study. All these re-sults provide useful and indispensable information for optimiza-tion of the cryopreservation of the human embryonic kidney cells.
Acknowledgments
This work was supported by the National Natural Science Foun-dation of China (Nos. 51076149, 51276179), and the FundamentalResearch Funds for the Central Universities (WK2100230009). Theauthors would like to acknowledge Mr. Jianye Wang for the guid-ance on cell culture and Miss Jingru Yi for the assistance of dataprocessing.
References
[1] L. Amar, M. Desclaux, N. Faucon-Biguet, J. Mallet, R. Vogel, Control of smallinhibitory RNA levels and RNA interference by doxycycline induced activationof a minimal RNA polymerase III promoter, Nucleic Acids Res. 34 (2006).
[2] S.K. Balasubramanian, J.C. Bischof, A. Hubel, Water transport and IIFparameters for a connective tissue equivalent, Cryobiology 52 (2006) 62–73.
[3] M.S. Berrada, J.C. Bischof, Evaluation of freezing effects on humanmicrovascular-endothelial cells (HMEC), Cryoletters 22 (2001) 353–366.
[4] J.C. Bischof, C.M. Ryan, R.G. Tompkins, M.L. Yarmush, M. Toner, Ice formation inisolated human hepatocytes and human liver tissue, ASAIO J. 43 (1997) 271–278.
[5] V. Brzhezovskiy, T. Tabolinovskaya, V. Lyubaev, I. Pustynskiy, A. Aidarbekova,T. Aketova, S. Subramanian, N. Malyshev, Cryosurgery in the treatment of oralcancer, Oral Oncol. 1 (2005) 190.
[6] S. Chuenkhum, Z.F. Cui, The parameter conversion from the Kedem–Katchalskymodel into the two-parameter model, Cryoletters 27 (2006) 185–199.
[7] T.B. Darr, A. Hubel, Freezing characteristics of isolated pig and humanhepatocytes, Cell Transplant. 6 (1997) 173–183.
[8] R.V. Devireddy, D.J. Swanlund, K.P. Roberts, J.C. Bischof, Subzero waterpermeability parameters of mouse spermatozoa in the presence ofextracellular ice and cryoprotective agents, Biol. Reprod. 61 (1999) 764–775.
[9] R.V. Devireddy, D.J. Swanlund, K.P. Roberts, J.L. Pryor, J.C. Bischof, The effect ofextracellular ice and cryoprotective agents on the water permeabilityparameters of human sperm plasma membrane during freezing, Hum.Reprod. 15 (2000) 1125–1135.
[10] M.F. Dowgert, P.L. Steponkus, Effect of cold-acclimation on intracellular iceformation in isolated protoplasts, Plant Physiol. 72 (1983) 978–988.
[11] S.L. Ebertz, L.E. McGann, Cryoinjury in endothelial cell monolayers,Cryobiology 49 (2004) 37–44.
[12] G.M. Fahy, Simplified calculation of cell water-content during freezing andthawing in nonideal solutions of cryoprotective agents and its possibleapplication to the study of solution effects injury, Cryobiology 18 (1981)473–482.
[13] F. Franks, S.F. Mathias, P. Galfre, S.D. Webster, D. Brown, Ice nucleation andfreezing in undercooled cells, Cryobiology 20 (1983) 298–309.
[14] F. Franks, T. Wakabayashi, S.F. Mathias, Nucleation kinetics of ice inundercooled yeast-cells – long-term stability against freezing, J. Gen.Microbiol. 133 (1987) 2807–2815.
[15] S. Fredj, K.J. Sampson, H.J. Liu, R.S. Kass, Molecular basis of ranolazine block ofLQT-3 mutant sodium channels: evidence for site of action, Br. J. Pharmacol.148 (2006) 16–24.
[16] A.A. Gage, Cryosurgery in the treatment of cancer, Surg. Gynecol. Obstet. 174(1992) 73–92.
[17] J.A. Gilmore, L.E. Mcgann, J. Liu, D.Y. Gao, A.T. Peter, F.W. Kleinhans, J.K. Critser,Effect of cryoprotectant solutes on water permeability of human spermatozoa,Biol. Reprod. 53 (1995) 985–995.
[18] H.M. Guy, L. McCloskey, G.J. Lye, K.A. Mitrophanous, T.K. Mukhopadhyay,Characterization of lentiviral vector production using microwell suspensioncultures of HEK293T-derived producer cells, Hum. Gene Ther. Methods 24(2013) 125–139.
[19] X. He, S. Mcgee, J.E. Coad, F. Schmidlin, P.A. Iaizzo, D.J. Swanlund, S. Kluge, E.Rudie, J.C. Bischof, Investigation of the thermal and tissue injury behaviour inmicrowave thermal therapy using a porcine kidney model, Int. J. Hyperthermia20 (2004) 567–593.
[20] X.M. He, J.C. Bischof, The kinetics of thermal injury in human renal carcinomacells, Ann. Biomed. Eng. 33 (2005) 502–510.
[21] X.M. He, A. Fowler, M. Toner, Water activity and mobility in solutions ofglycerol and small molecular weight sugars: implication for cryo- andlyopreservation, J. Appl. Phys. 100 (2006).
[22] H.G. Hempling, S. White, Permeability of cultured megakaryocytopoietic cellsof the rat to dimethyl sulfoxide, Cryobiology 21 (1984) 133–143.
[23] J. Hunter, A. Bernard, B. Fuller, J. Mcgrath, R.W. Shaw, Plasma-membrane waterpermeabilities of human oocytes – the temperature-dependence of water-movement in individual cells, J. Cell. Physiol. 150 (1992) 175–179.
[24] T. Kanno, H. Yamamoto, T. Yaguchi, R. Hi, T. Mukasa, H. Fujikawa, T. Nagata, S.Yamamoto, A. Tanaka, T. Nishizaki, The linoleic acid derivative DCP-LAselectively activates PKC-epsilon, possibly binding to the phosphatidylserinebinding site, J. Lipid Res. 47 (2006) 1146–1156.
[25] J.O.M. Karlsson, E.G. Cravalho, I.H.M.B. Rinkes, R.G. Tompkins, M.L. Yarmush,M. Toner, Nucleation and growth of ice crystals inside cultured-hepatocytesduring freezing in the presence of dimethyl-sulfoxide, Biophys. J. 65 (1993)2524–2536.
[26] J.O.M. Karlsson, E.G. Cravalho, M. Toner, A model of diffusion-limited icegrowth inside biological cells during freezing, J. Appl. Phys. 75 (1994) 4442–4445.
[27] A. Krakowiak, R. Pecherzewska, R. Kaczmarek, A. Tomaszewska, B. Nawrot, W.J.Stec, Evaluation of influence of Ap4A analogues on Fhit-positive HEK293Tcells; cytotoxicity and ability to induce apoptosis, Bioorg. Med. Chem. 19(2011) 5053–5060.
[28] S.P. Leibo, P. Mazur, E.H.Y. Chu, 2 factors responsible for freezing injury inhamster tissue culture cells, Cryobiology 6 (1970) 580.
[29] R.L. Levin, E.G. Cravalho, C.E. Huggins, Membrane model describing effect oftemperature on water conductivity of erythrocyte-membranes at subzerotemperatures, Cryobiology 13 (1976) 415–429.
[30] M.O. Maiwand, J.M. Evans, J.E. Beeson, The application of cryosurgery in thetreatment of lung cancer, Cryobiology 48 (2004) 55–61.
[31] P. Mazur, Kinetics of water loss from cells at subzero temperatures andlikelihood of intracellular freezing, J. Gen. Physiol. 47 (1963) 347.
[32] P. Mazur, Cryobiology – freezing of biological systems, Science 168 (1970) 939.[33] P. Mazur, Role of intracellular freezing in death of cells cooled at supraoptimal
rates, Cryobiology 14 (1977) 251–272.[34] P. Mazur, Equilibrium, quasi-equilibrium, and nonequilibrium freezing of
mammalian embryos, Cell Biophys. 17 (1990) 53–92.[35] P. Mazur, S.P. Leibo, E.H.Y. Chu, 2-Factor hypothesis of freezing injury –
evidence from Chinese-hamster tissue-culture cells, Exp. Cell Res. 71 (1972)345.
[36] C. Mccaa, K.R. Diller, S.J. Aggarwal, T. Takahashi, Cryomicroscopicdetermination of the membrane osmotic properties of human monocytes atsubfreezing temperatures, Cryobiology 28 (1991) 391–399.
[37] H. Mustafa, B. Strasser, S. Rauth, R.A. Irving, K.L. Wark, Identification of afunctional nuclear export signal in the green fluorescent protein asFP499,Biochem. Biophys. Res. Commun. 342 (2006) 1178–1182.
[38] D.J. Smith, M. Schulte, J.C. Bischof, The effect of dimethylsulfoxide on the watertransport response of rat hepatocytes during freezing, J. Biomech. Eng. 120(1998) 549–558.
[39] S. Thirumala, R.V. Devireddy, A simplified procedure to determine the optimalrate of freezing biological systems, J. Biomech. Eng. 127 (2005) 295–300.
[40] P. Thomas, T.G. Smart, HEK293 cell line: a vehicle for the expression ofrecombinant proteins, J. Pharmacol. Toxicol. Methods 51 (2005) 187–200.
[41] M. Toner, E.G. Cravalho, D.R. Armant, Water transport and estimatedtransmembrane potential during freezing of mouse oocytes, J. Membr. Biol.115 (1990) 261–272.
[42] M. Toner, E.G. Cravalho, M. Karel, Thermodynamics and kinetics ofintracellular ice formation during freezing of biological cells, J. Appl. Phys.67 (1990) 1582–1593.
[43] M. Toner, E.G. Cravalho, M. Karel, D.R. Armant, Cryomicroscopic analysis ofintracellular ice formation during freezing of mouse oocytes withoutcryoadditives, Cryobiology 28 (1991) 55–71.
[44] J.Y. Wang, K.X. Zhu, G. Zhao, J. Ren, C. Yue, D.Y. Gao, Dual dependence ofcryobiogical properties of Sf21 cell membrane on the temperature and theconcentration of the cryoprotectant, PLoS One 8 (2013).
[45] G. Yang, M. Veres, G. Szalai, A.L. Zhang, L.X. Xu, X.M. He, Biotransportphenomena in freezing mammalian oocytes, Ann. Biomed. Eng. 39 (2011)580–591.
[46] G. Yang, A.L. Zhang, L.X. Xu, Intracellular ice formation and growth in MCF-7cancer cells, Cryobiology 63 (2011) 38–45.
[47] A. Zhang, L.X. Xu, G.A. Sandison, S. Cheng, Morphological study of endothelialcells during freezing, Phys. Med. Biol. 51 (2006) 6047–6060.
[48] G. Zhao, A universal self-adaptive time-varying function for extracellularconcentration during osmotic shift for curve-fitting permeability coefficientsof cell membrane, Cryoletters 31 (2010) 279–290.
[49] G. Zhao, K. Kurata, H. Takamatsu, Measurement of membrane hydraulicconductivity of bovine carotid artery endothelial cells using a perfusionmicroscope, Cryoletters 33 (2012) 232–240.
[50] G. Zhao, D.W. Luo, D.Y. Gao, Universal model for intracellular ice formation andits growth, AIChE J. 52 (2006) 2596–2606.
Copyright © 2022 FDOKUMEN




















