
Biological influences on modern sulfates: Textures and composition of gypsum deposits from Guerrero...
16
Biological influences on modern sulfates: Textures and composition of gypsum deposits from Guerrero Negro, Baja California Sur, Mexico Marilyn B. Vogel a, ⁎, David J. Des Marais b , Mary N. Parenteau c , Linda L. Jahnke b , Kendra A. Turk a,1 , Michael D.Y. Kubo a a SETI Institute, 415 N. Whisman Rd., Mountain View, CA 94043, United States b NASA Ames Research Center, Astrobiology Branch N239-4, Moffett Field, CA, 94035, United States c Oak Ridge Associated Universities NPP, NASA/ARC Astrobiology Branch, Moffett Field, CA, 94035, United States abstract article info Article history: Received 11 May 2009 Received in revised form 24 November 2009 Accepted 24 November 2009 Keywords: Gypsum Sulfates Biological influence Sabkha Biofilm–mineral interactions Carbonate replacement Gypsum (CaSO 4 ·2H 2 O) deposits from a range of sedimentary environments at Guerrero Negro, Baja California Sur, Mexico were investigated for microscale texture and composition in order to differentiate features formed under substantial microbial influence from those for which microbial effects were relatively minor or absent. Gypsum deposits were classified according to their sedimentary environment, textures, crystal habit, brine composition and other geochemical factors. The environments studied included subaqueous sediments in anchialine pools and in solar salterns, as well as subsurface sediments of mudflats and saltpans. Gypsum that developed in the apparent absence of biofilms included crystals precipitated in the water column and subsedimentary discs that precipitated from phreatic brines. Subsedimentary gypsum developed in sabkha environments exhibited a sinuous microtexture and poikilitically enclosed detrital particles. Water column precipitates had euhedral prismatic habits and extensive penetrative twinning. Gypsum deposits influenced by biofilms included bottom nucleated crusts and gypsolites developing in anchialine pools and saltern ponds. Gypsum precipitating within benthic biofilms, and in biofilms within subaerial sediment surfaces provided compelling evidence of biological influences on crystal textures and habits. This evidence included irregular, high relief surface textures, accessory minerals (S°, Ca-carbonate, Sr/ Ca-sulfate and Mg-hydroxide) and distinctive crystal habits such as equant forms and crystals having distorted prism faces. © 2009 Elsevier B.V. All rights reserved. 1. Introduction Gypsum (CaSO 4 ·2H 2 O) is an important phase in biogeochemistry and sedimentology as a mineral sink for sulfur and as an endolithic niche for phototrophic and chemotrophic microorganisms. The petrology of classical ancient gypsum deposits such as the Miocene and Permian successions has been systematically studied (i.e. Hardie et al., 1985; Spencer and Lowenstein, 1988; Michalzik, 1996) but reports describing textural and compositional characteristics of primary sulfates forming in modern environments are relatively scarce. The discovery of gypsum on Mars by the Mars Express Orbiter (Bibring et al., 2006) and the Mars Reconnaissance Orbiter (Murchie et al., 2007) underscores the need to identify factors that influence gypsum morphology and mineral associations. Instruments on the Mar Science Laboratory mission, currently scheduled for launch in 2011, will be able to image surface sedimentary features at a scale of 12 μm per pixel, highlighting the need for better understanding of evaporite textures on microscopic scales. This report focuses on gypsum forming in subaerial, subsedimen- tary, and subaqueous environments of a modern coastal sabkha and saltern near Guerrero Negro, Baja California Sur, Mexico. This locality includes a typical sabkha with mud and gypsum (see Schreiber and Tabakh, 2000; Warren, 2006), natural anchialine pools and a marine solar saltern managed by Exportadora de Sal, S.A. (ESSA). The range of depositional environments provided an opportunity to study textural and compositional characteristics of gypsum that formed in subaque- ous, subaerial and subsedimentary environments. Although fine scale textures and morphologies such as those described here are typically lost or modified on Earth over geologic timescales, ancient Martian evaporites have retained remarkably well-preserved features that formed during deposition and early diagenesis (McLennan et al., 2005). A clearer understanding of primary textures in evaporites also can assist in interpretation of replacement phases and stable isotope patterns (e.g. Shen et al., 2001; Runnegar et al. 2001; Gandin et al., 2005). Early diagenesis of primary gypsum includes dissolution/repreci- pitation cycles that often lead to carbonate or polyhalite replacement Sedimentary Geology 223 (2010) 265–280 ⁎ Corresponding author. E-mail address: [email protected] (M.B. Vogel). 1 Current Address: Ocean Sciences, University of California, Santa Cruz, Santa Cruz, CA 95064, United States. 0037-0738/$ – see front matter © 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.sedgeo.2009.11.013 Contents lists available at ScienceDirect Sedimentary Geology journal homepage: www.elsevier.com/locate/sedgeo
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Biological influences on modern sulfates: Textures and composition of gypsum deposits from Guerrero...

Sedimentary Geology 223 (2010) 265–280
Contents lists available at ScienceDirect
Sedimentary Geology
j ourna l homepage: www.e lsev ie r.com/ locate /sedgeo
Biological influences on modern sulfates: Textures and composition of gypsumdeposits from Guerrero Negro, Baja California Sur, Mexico
Marilyn B. Vogel a,⁎, David J. Des Marais b, Mary N. Parenteau c, Linda L. Jahnke b,Kendra A. Turk a,1, Michael D.Y. Kubo a
a SETI Institute, 415 N. Whisman Rd., Mountain View, CA 94043, United Statesb NASA Ames Research Center, Astrobiology Branch N239-4, Moffett Field, CA, 94035, United Statesc Oak Ridge Associated Universities NPP, NASA/ARC Astrobiology Branch, Moffett Field, CA, 94035, United States
⁎ Corresponding author.E-mail address: [email protected] (M.B. Vog
1 Current Address: Ocean Sciences, University of CaliCA 95064, United States.
0037-0738/$ – see front matter © 2009 Elsevier B.V. Aldoi:10.1016/j.sedgeo.2009.11.013
a b s t r a c t
a r t i c l e i n f oArticle history:Received 11 May 2009Received in revised form 24 November 2009Accepted 24 November 2009
Keywords:GypsumSulfatesBiological influenceSabkhaBiofilm–mineral interactionsCarbonate replacement
Gypsum (CaSO4·2H2O) deposits from a range of sedimentary environments at Guerrero Negro, BajaCalifornia Sur, Mexico were investigated for microscale texture and composition in order to differentiatefeatures formed under substantial microbial influence from those for which microbial effects were relativelyminor or absent. Gypsum deposits were classified according to their sedimentary environment, textures,crystal habit, brine composition and other geochemical factors. The environments studied includedsubaqueous sediments in anchialine pools and in solar salterns, as well as subsurface sediments of mudflatsand saltpans. Gypsum that developed in the apparent absence of biofilms included crystals precipitated inthe water column and subsedimentary discs that precipitated from phreatic brines. Subsedimentary gypsumdeveloped in sabkha environments exhibited a sinuous microtexture and poikilitically enclosed detritalparticles. Water column precipitates had euhedral prismatic habits and extensive penetrative twinning.Gypsum deposits influenced by biofilms included bottom nucleated crusts and gypsolites developing inanchialine pools and saltern ponds. Gypsum precipitating within benthic biofilms, and in biofilms withinsubaerial sediment surfaces provided compelling evidence of biological influences on crystal textures andhabits. This evidence included irregular, high relief surface textures, accessory minerals (S°, Ca-carbonate, Sr/Ca-sulfate and Mg-hydroxide) and distinctive crystal habits such as equant forms and crystals havingdistorted prism faces.
el).fornia, Santa Cruz, Santa Cruz,
l rights reserved.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
Gypsum (CaSO4·2H2O) is an important phase in biogeochemistryand sedimentology as a mineral sink for sulfur and as an endolithicniche for phototrophic and chemotrophic microorganisms. Thepetrology of classical ancient gypsum deposits such as the Mioceneand Permian successions has been systematically studied (i.e. Hardieet al., 1985; Spencer and Lowenstein, 1988; Michalzik, 1996) butreports describing textural and compositional characteristics ofprimary sulfates forming in modern environments are relativelyscarce. The discovery of gypsum on Mars by the Mars Express Orbiter(Bibring et al., 2006) and the Mars Reconnaissance Orbiter (Murchieet al., 2007) underscores the need to identify factors that influencegypsum morphology and mineral associations. Instruments on theMar Science Laboratory mission, currently scheduled for launch in2011, will be able to image surface sedimentary features at a scale of
12 μm per pixel, highlighting the need for better understanding ofevaporite textures on microscopic scales.
This report focuses on gypsum forming in subaerial, subsedimen-tary, and subaqueous environments of a modern coastal sabkha andsaltern near Guerrero Negro, Baja California Sur, Mexico. This localityincludes a typical sabkha with mud and gypsum (see Schreiber andTabakh, 2000; Warren, 2006), natural anchialine pools and a marinesolar saltern managed by Exportadora de Sal, S.A. (ESSA). The range ofdepositional environments provided an opportunity to study texturaland compositional characteristics of gypsum that formed in subaque-ous, subaerial and subsedimentary environments. Although fine scaletextures and morphologies such as those described here are typicallylost or modified on Earth over geologic timescales, ancient Martianevaporites have retained remarkably well-preserved features thatformed during deposition and early diagenesis (McLennan et al.,2005). A clearer understanding of primary textures in evaporites alsocan assist in interpretation of replacement phases and stable isotopepatterns (e.g. Shen et al., 2001; Runnegar et al. 2001; Gandin et al.,2005).
Early diagenesis of primary gypsum includes dissolution/repreci-pitation cycles that often lead to carbonate or polyhalite replacement
266 M.B. Vogel et al. / Sedimentary Geology 223 (2010) 265–280
(e.g. Pierre, 1983; Anadón et al., 1992). Primary gypsum in modernenvironments is also extensively affected by microbial biogeochem-ical factors. Microbial communities leave clear evidence of theiractivity in the microscale textures and mineralogy of sub-aerial andsubaqueous gypsum deposits. Criteria for assessing the biogenicity oftextures in gypsum however have not been systematically defined.Effective classification of gypsum facies requires assessment ofwhether specific morphologies are unique to certain processes and/or environments. Detailed description of sedimentary environmentsand microbial community diversity is provided elsewhere (Vogel
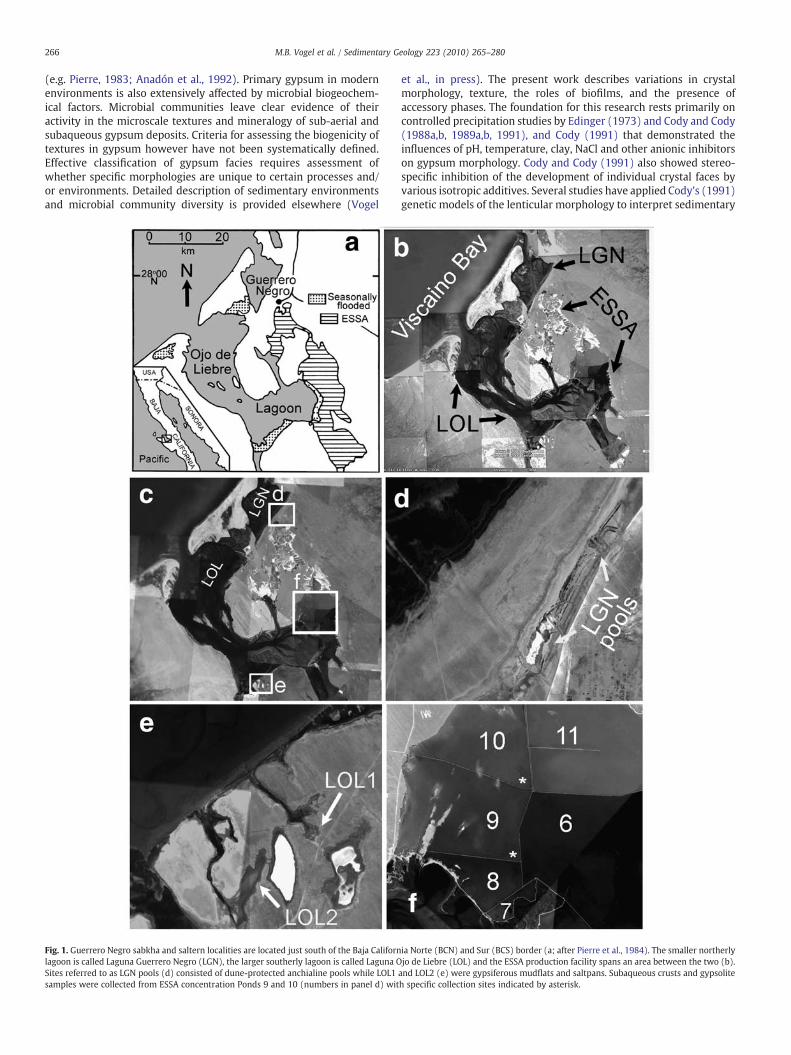
Fig. 1. Guerrero Negro sabkha and saltern localities are located just south of the Baja Californlagoon is called Laguna Guerrero Negro (LGN), the larger southerly lagoon is called Laguna OSites referred to as LGN pools (d) consisted of dune-protected anchialine pools while LOL1samples were collected from ESSA concentration Ponds 9 and 10 (numbers in panel d) wit
et al., in press). The present work describes variations in crystalmorphology, texture, the roles of biofilms, and the presence ofaccessory phases. The foundation for this research rests primarily oncontrolled precipitation studies by Edinger (1973) and Cody and Cody(1988a,b, 1989a,b, 1991), and Cody (1991) that demonstrated theinfluences of pH, temperature, clay, NaCl and other anionic inhibitorson gypsum morphology. Cody and Cody (1991) also showed stereo-specific inhibition of the development of individual crystal faces byvarious isotropic additives. Several studies have applied Cody's (1991)genetic models of the lenticular morphology to interpret sedimentary
ia Norte (BCN) and Sur (BCS) border (a; after Pierre et al., 1984). The smaller northerlyjo de Liebre (LOL) and the ESSA production facility spans an area between the two (b).and LOL2 (e) were gypsiferous mudflats and saltpans. Subaqueous crusts and gypsoliteh specific collection sites indicated by asterisk.

267M.B. Vogel et al. / Sedimentary Geology 223 (2010) 265–280
environments (e.g. Aref, 1998; Buck and Van Hoesen, 2002), howeverno studies have addressed the effects of microbial biofilms on gypsummorphology.
2. Depositional environment and sedimentology
The field study site included two subtidal embayments adjacent tothe Bahia de Sebastian Vizcaino located just south of the borderbetween Baja California Norte and Baja California Sur (lat. 28°00′N–27°30′N; Fig. 1a). Laguna Guerrero Negro (LGN) and the larger LagunaOjo de Liebre (LOL) are protected by the barriers of Isla Arena and thenortheast cusp of Punta Eugenia, respectively (Fig. 1b). The geographyand sedimentology of the area were described in detail by Phleger(1962), prior to establishment of the salt production facility. Primarygypsum deposits occur in the natural littoral, supralittoral and sub-aerial environments (the sabkha) and in the subaqueous environ-ments of anchialine pools and channels, and the artificial concentra-tion ponds of the Exportadora de Sal, S.A. (ESSA) salt productionfacility (saltern). Sabkha gypsum deposits were studied at localitiesalong the margins of LGN where several natural depressions arejuxtaposed against sandy berms (Fig. 1d), and at sites along sandy highground accessed by tidal channels along the southern margin of LOL(Fig. 1e). Saltern deposits were studied in the marginal splash zonesand subaqueous environments of ESSA concentration Ponds 9 and 10(Fig. 1f; salinity increases with increasing pond #). These ponds hostthe saltern's most voluminous gypsum deposits and are locateddirectly upstream from ponds where halite precipitation commences.
2.1. Sabkha—LGN and LOL
The LGN site includes anchialine pools and channels that attaindepths of up to 0.5 m. The pools host both abundant benthic microbialbiofilmswith internally crystallized gypsum granules andmoremassivecrystalline gypsum deposits with a biofilm component. These poolsranged in salinity from 130‰ (measured by on-site refractometer) tohalite saturation (N300‰) but only gypsum depositing environmentswere sampled for this study (Table 1). Pools with thick benthic biofilmshad salinities between ∼130‰ and 150‰. Density stratified pools in themore southerly areas of LGNhad salinities gradients from136‰ to 220‰and temperature gradients from 29 °C to 40 °C. Black surface biofilmscoated the gypsumdeposits in the lower regions of these stratified pools(black mantle gypsum). Water column precipitate crystals werecollected from LGN pools with salinities of 174 to 176‰.
Table 1Locality, pH, salinity and major cation and anion concentration data for saltern and sabkha snatural anchialine pools and saltern ponds. LOL samples were collected from gypsiferous mand lower most layers of a density- and temperature-stratified anchialine pool.
Locality Description pH Salinitya
(ppt)Na(ppm)
Mg(ppm)
K(
Pond 9 Subaqueous saltern—Sept. 2007 7.3 172 51,300 6650 2Pond 9 Subaqueous saltern—April 2006 7.9 171 52,700 6840 2Pond 10 Subaqueous saltern—Sept. 2007 7.7 212 68,200 8670 2Pond 10 Subaqueous saltern—Feb. 2006 7.4 300/o.s. 99,900 11,800 3LGN1-1 Subaqueous benthic mats; biofilm-
supported gypsum7.5 162 52,500 6690 2
LGN1-2 Subaqueous—upper layer of densitystratified w/black gypsum
7.8 139 41,000 4890 1
LGN1-3 Subaqueous—lower layer densitystratified w/black gypsum
7.4 220 73,500 9160 2
LOL1-1 Biofilm/sediment surface—brine sheet 7.4 300/o.s. 89,500 20,000 5LOL2-3 Biofilm/sediment surface—brine sheet 7.1 300/o.s. 78,700 27,200 6LOL3-1 Biofilm/sediment surface,
Pump Station #17.3 241 79,700 11,000 2
LOL2-1 Sediment subsurface 6.9 252 82,200 13,800 3LOL2-2 Sediment subsurface 7.3 300/o.s. 86,600 16,300 4
a Salinity (ppt) measurements were collected with a refractometer with detection limitb Seawater concentration factors are based on average seawater concentration values for
The LOL site consists of evaporiticmudflats at the terminus of two tidalchannels (LOL1 and LOL2; Fig. 1e). Gypsum and halite form a thinprecipitate surfaceabovediscretepink, greenandpurple subsurface layers(1–2 cm) consisting of mud sized clastic sediment, gypsum precipitategrains and microbial communities. The gypsum surface and sediment/biofilm layer are underlain by black sulfidic mud and unconsolidated tanto orange/brown carbonate sand. The water table was observed in thevicinity of the carbonate sand at depths of 15 to 20 cm. Subsedimentarygypsum develops in both the vadose and phreatic horizons beneath thegypsumandbiofilm/sediment surface. Vadose gypsumoccurred in browncalcareous mud horizons (4–5 cm depth) as white, mm scale flakes orlaths. Phreatic gypsum occurs as 1 to 5 cm lenticular crystals and aggre-gates colored brown by abundant detrital inclusions.
2.2. Saltern—Exportadora de Sal, S.A. (ESSA) Ponds 9 and 10
Gypsum is deposited as gypsolites, cements and massive botryoidalcrusts in Pond 9 (150–170‰ salinity) and Pond 10 (N210‰ salinity) ofthe ESSA salterns. The turbidity, agitation, and water depths are similarfor both ponds, but greater in magnitude than that of the protectedanchialine pools of the LGN site. Although the saltern ponds attaindepths of up to 1.5 m in places, samples described herewere collected inshallower areas (b1 mdeep). Gypsumcrusts develop atop sulfidicmuds,gypsiferous sand, or shelly gravels and range in thickness from 5 to10 cm in Pond 9 up to several meters in Pond 10. These crusts have anouter mantle of coarse, growth-aligned gypsum twins inhabited bylayered orange, purple and green endolithicmicrobial communities, andan inner,mottled granular corewithdark red, brownandblackmicrobialbiofilms. Gypsum spherules, referred to as gypsooids, occurred either asunconsolidated coarse sands or as gypsolites that are stabilized andconsolidated by mm scale layers of gypsum cement. Cement-stabilizedgypsooids located in the near subsurface of the precipitate–waterinterface hosted discretely layered orange, green and purple biofilms.
3. Methodology
3.1. Gypsum crystallographic descriptions
Due to the variety of ways inwhich gypsum can be indexed, salientpoints of the descriptive framework for gypsum crystallography arereviewed here. To facilitate comparisons, the Miller indices of Dana(1892) and Cody and Cody (1988a) are used, with F-morphologic cellcentering and unit cell elongation along the c-axis. Crystal faces are
amples. Saltern pond and LGN samples represent subaqueous environments, includingudflats and saltpans. LGN1-2 and LGN1-3 represent samples taken from the uppermost
ppm)Ca(ppm)
Sr(ppm)
Br(ppm)
SO4
(ppm)Cl(ppm)
Na xs.w.b
Ca xs.w.b
Mg xs.w.b
K xs.w.b
110 1500 34.60 53.12 15,752 100,729 4.7 3.6 5.0 5.2180 1180 35.20 56.76 16,041 184,428 4.8 2.8 5.2 5.3700 932 38.00 56.81 17,925 131,991 6.2 2.2 6.6 6.6500 277 9.90 69.12 21,329 103,102 9.1 0.7 8.9 8.6020 1160 35.20 55.08 15,477 98,353 4.8 2.8 5.1 5.0
500 1260 22.80 51.78 13,139 80,031 3.7 3.0 3.7 3.7
540 740 19.60 61.06 18,430 138,344 6.7 1.8 6.9 6.2
350 299 12.60 74.69 37,634 182,211 8.1 0.7 15.2 13.1910 207 11.50 88.72 49,277 189,416 7.2 0.5 20.6 16.9860 840 34.90 63.12 16,251 156,596 7.2 2.0 8.3 7.0
400 471 9.80 64.54 27,152 159,617 7.5 1.1 10.5 8.3220 363 14.50 71.65 30,455 173,823 7.9 0.9 12.4 10.3
of b300 ppt (o.s. = off scale).each cation from Fontes and Matray (1993).

268 M.B. Vogel et al. / Sedimentary Geology 223 (2010) 265–280
referred to as generalized {hkl} forms or by letters as shown in Fig. 2.Most of the crystals described here range from euhedral prismaticmorphologies having prominent {010} pinacoids or b faces, tolenticular morphologies with enlarged l{111} and e{103} faces, andb{010} and m{110} faces that are diminished or absent (Fig. 2a). Themonoclinic 2/m symmetry class consists exclusively of pinacoid andprism forms, and has no symmetrically unmatched faces. Literaturesources however sometimes refer to individual e pinacoids as pedions.As outlined in Chang et al. (1995), gypsum can be described by asmany as six different unit cells using body (I-cell) or face (F- and A-cell) centered lattices. The high degree of symmetry of the gypsumcrystal alsomeans that assignment of the first- and third-order prisms(l andm for gypsum) depends on assignment of the elongated edge ofthe unit cell to either the a- or c-axis. Crystallographic elongation isknown to favor the [001] zone axis, which is usually referred to as thec-axis (e.g. Bosbach and Ramensee, 1994, and here) and sometimesreferred to as the a-axis (e.g. Hünger and Henning, 1988). Etch pitsand nucleation islands may also assume this [001] elongation, andboth theoretical and experimental research have shown that itprovides a visual clue for orienting prismatic crystals (Bosbach andHochella, 1996; Fan and Teng, 2007). Etch pits were sometimesobserved in the samples described here but could be difficult tointerpret (Fig. 2b).
Many of the crystals described here have faceted to semi-faceted,equant or distorted morphologies with unusual aspect ratios among
Fig. 2. Crystallographic orientation and nomenclature for gypsum samples after Dana anelongation along the c-axis (panel a). Crystals with prominent b{010} pinacoids were descrLensoidal/lenticular crystals were flattened along [001] and have l prism and e pinacoid faceparallel to [001] and the shelved textures ofm prisms are parallel to [100] respectively (c). Torder prisms (panels b–d).
prisms and pinacoids (Fig. 2a). Distinguishing between prisms wasdifficult for lenticular crystals that showed rounding and shorteningalong [001], and for other morphologies that did not show clear signsof [001] elongation. Gypsum exhibits perfect cleavage parallel to the{010} face due to the arrangement of water molecules between Ca2+-bound SO4
2 tetrahedron layers (Cole and Lancucki, 1974; Pedersen andSemmingsen, 1982). Fracture parallel to {100} can yield a conchoidalsurface, and fracture on the {011} prism can yield a splintery or fibroussurface. Prismatic gypsum crystals in this study exhibited distinctlydifferent microtextures on the {110} and {111} prisms. The {110} ormprisms have ridged or shelf-like textures with ridge axes orientedparallel to [100], whereas {111} or l prisms have a striated or sinuoustexture with irregular elongated void spaces parallel to [001], similarto the fibrous texture predicted for fracture of this surface (Fig. 2c–d).Similar shelved and striated surfaces are apparent in micrographs ofgoniometrically referenced gypsum crystals shown in Cody and Cody(1991). The striated and shelved textures, apparent in crystalsdescribed here, are thus used in orienting the l{111} and m{110}prisms, respectively for crystals with distorted, equant or lenticularforms.
3.2. Analytical techniques
Samples and on-site hydrochemical data were collected in Apriland September of 2007. Brine cation data was collected using ion
d Cody and Cody (1989a,b), are based on F-morphologic cell indexing and unit cellibed as prismatic. Equant crystals had enlarged m and l prisms relative to b pinacoids.s that are enlarged with indistinct edges. The sinuous/fibrous texture of the l prisms arehe sinuous/fibrous and shelved textures were used to visually orient the first and third-
269M.B. Vogel et al. / Sedimentary Geology 223 (2010) 265–280
chromatography (Ca, Mg, K, Na), coulometry (Fe) and ICP-MS (Sr) atMonterey Bay Analytical Laboratories in Monterey, CA. Brine anionconcentrations (Cl−, Br− and SO4
−) were measured at Ames ResearchCenter using a Dionex ICS2000 ion chromatograph system with KOHcarrier reagents. Samples were gently rinsed in de-ionized water toremove brine and residual salts for petrographic imaging with highvacuum scanning electron microscopy (SEM). SEM imaging wasperformed on a JOEL 5600 SEM with an energy dispersive X-rayspectroscopic detector (EDS) at the Stanford/USGS Micro-AnalyticalCenter. X-ray diffraction (XRD) was also used to identify accessoryphases in several samples. XRD samples were analyzed using Terra(inXitu, Inc. Mountain View, CA), a field-portable version of theCheMin X-ray diffraction/X-ray fluorescence (XRD/XRF) instrumentchosen to fly on the 2011 Mars Science Laboratory Mission. Sampleswith grain sizes of b150 μm and weighing ∼50 mg were run on Terraat 25 keV and ∼10 W for 30 min to 1 h (Taylor et al., 2008). The
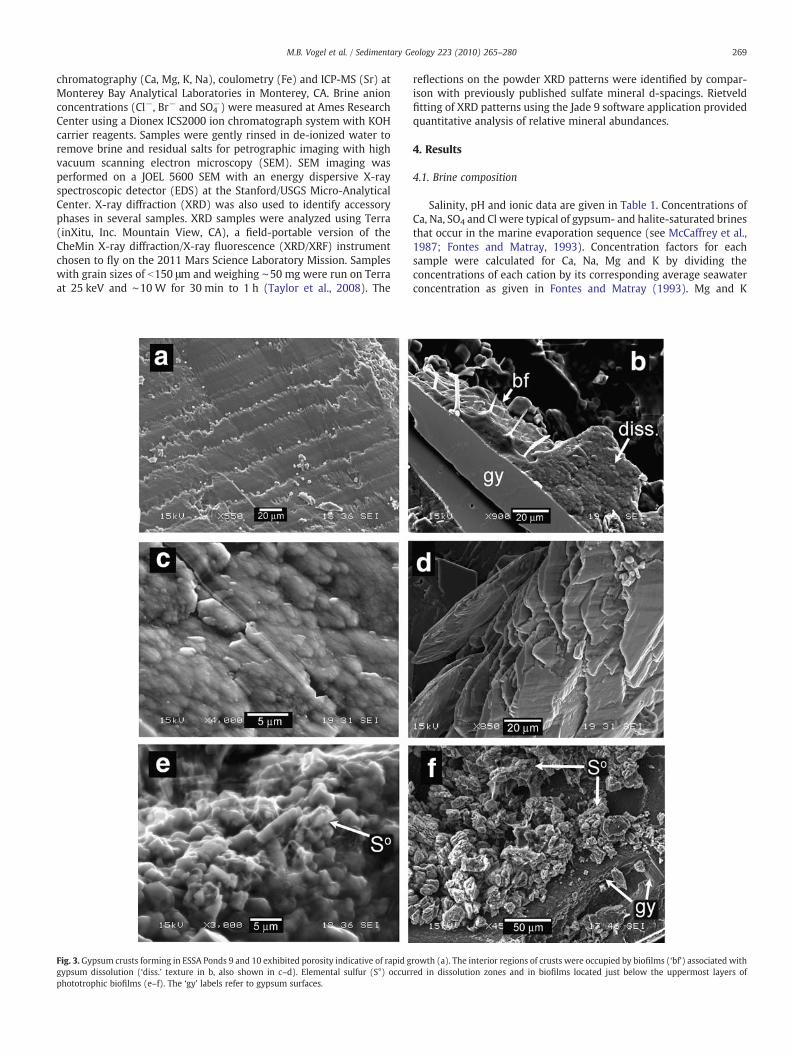
Fig. 3. Gypsum crusts forming in ESSA Ponds 9 and 10 exhibited porosity indicative of rapid ggypsum dissolution (‘diss.’ texture in b, also shown in c–d). Elemental sulfur (S°) occurphototrophic biofilms (e–f). The ‘gy’ labels refer to gypsum surfaces.
reflections on the powder XRD patterns were identified by compar-ison with previously published sulfate mineral d-spacings. Rietveldfitting of XRD patterns using the Jade 9 software application providedquantitative analysis of relative mineral abundances.
4. Results
4.1. Brine composition
Salinity, pH and ionic data are given in Table 1. Concentrations ofCa, Na, SO4 and Cl were typical of gypsum- and halite-saturated brinesthat occur in the marine evaporation sequence (see McCaffrey et al.,1987; Fontes and Matray, 1993). Concentration factors for eachsample were calculated for Ca, Na, Mg and K by dividing theconcentrations of each cation by its corresponding average seawaterconcentration as given in Fontes and Matray (1993). Mg and K
rowth (a). The interior regions of crusts were occupied by biofilms (‘bf’) associated withred in dissolution zones and in biofilms located just below the uppermost layers of

270 M.B. Vogel et al. / Sedimentary Geology 223 (2010) 265–280
concentration factors ranged from ∼3 to 20 and serve as proxies forrelative salinities among brine samples. The Ca and Na concentrationfactors are shown for comparison with differences in concentrationfactors reflecting the extent of gypsum and halite precipitation.Equivalent factors indicate little or no precipitation and disparatefactors indicate active or significant precipitation of Ca- and Na-bearing phases. The lowest Ca concentration factors measuredcorrespond to subsedimentary brine sheets from the LOL (LagunaOjo de Liebre) sabkha. These brines also had the highest salinities(measured by refractometer) and low pH values, relative to brinesfrom other environments. Subaqueous brines from LGN (LagunaGuerrero Negro) exhibited slightly lower concentration factors thanPond 9 and 10 brines. Concentrations of Sr were below typical valuesfor gypsum-precipitating brines. Brine samples from the LOL sabkhalocality had Sr concentrations that were only slightly higher thannormal (i.e. unconcentrated) seawater values. Low Sr concentrationvalues may be due to brine depletion of Sr caused by precipitation ofSr-bearing phases within LOL sabkha biofilm/sediment surfaces (seebelow). Br was also lower than expected for gypsum domain brines,occurring in concentrations equivalent to, or below those of normalseawater. Only one sample from a subsurface sample, LOL2-2, haddetectible Fe concentrations (90 ppm; not shown in Table 1).
4.2. Texture and morphology descriptions
4.2.1. Gypsum crusts: subaqueous; sediment–water interface;ESSA Ponds 9 and 10
Botryoidal crusts and pavements (crusts) in subaqueous environ-ments exhibited coarse twinned crystalline textures. Crusts andgypsolites described below form in areas of the saltern that hostsmaximal gypsum precipitation. Gypsum crusts consist of bottomnucleated, growth-aligned cm-scale twins with chevron growth
Fig. 4. Gypsooids were mostly 0.5 to 1 mm in diameter and had both gypsum and non-gypcrystals (a). Some gypsooids had carbonate cores (‘cc’ in b). Surface crystals of associated wilenticular crystals had coarser habits where biofilms were sparse (d). Arrows indicate faint
bands. Euhedral crystals in the upper portions of the crust exhibitvery little texture in micrographs except for growth-aligned porosity.This porosity appeared as growth bands of elongate μm scale voids(Fig. 3a). At depths of 1 cm or greater, pigmented biofilms occupyinterstitial spaces and defect sites. Biofilms occur as discrete orange,green purple and brown layers within the crust (see Vogel et al.,2009). The inner regions of the crust have dissolution zones whereeuhedral crystals verged into uncemented granular residues associ-ated with thicker biofilms. Dissolution zone gypsum exhibit labiate(meaning resembling lips) or irregular etched surfaces with subtle 2-to 5-μm sized globular features often drapedwith biofilms (Fig. 3b–d).Elemental sulfur occurs within gypsum crusts from Pond 10 atsalinities of ∼210–300 ppt. Crusts in Pond 10 form as thicker depositswith biofilms occurring at greater depths within the surfaces. Sulfur issometimes present embedded in yellow to light-olive biofilms, eitheron grains at the base of the uppermost orange-colored zones, orintermingled with the inner pink-colored gypsum dissolution zone.Sulfur phases were typically 1 to 5 μm granules that were either tooscarce to be detected by XRD or had no defined crystal structure(Fig. 3e). Larger (∼10 mm) tabular aggregates were also observedfrom lower layers of an alternate crust (Fig. 3f).
4.2.2. Gypsolite: subaqueous; sediment–water interface; ESSA Ponds 9and 10
Gypsolites composed of gypsooids (gypsum spheres with internalnuclei that are 0.5–2 mm in diameter) develop in marginal subaque-ous environments at the southeastern corner of Pond 9. This area ofPond 9 experiences significant wave agitation from easterly winds.The surfaces of individual gypsooids exhibited sub-millimeter scale,petal-like texture consisting of randomly oriented, interlockingprismatic/lenticular and lenticular crystals (Fig. 4a). (In this report aprismatic/lenticular crystal is flattened but retains straight crystal
sum cores. Surfaces were composed of aggregates of prismatic/lenticular to lenticularth biofilms were flattened with ribbed or ridged edges (‘x’ and ‘*’ in panel c). Prismatic/edges that intersect the acute point of the b{010} pinacoid.

271M.B. Vogel et al. / Sedimentary Geology 223 (2010) 265–280
edges, whereas a lenticular crystal is flattened with round edges.) Theinteriors of gypsooid consisted of randomly oriented sub-micron scalespar and coarser millimeter scale spar in the core regions of the grain.Some gypsooids had allochthonous cores of carbonate (Fig. 4b) orother detrital particles. Gypsooid layers stabilizedby syngenetic gypsumcementshost biofilms of stratified phototrophicmicrobial communities,some of which precipitate carbonate granules (see below). Biofilmresidueswereobserveddrapedon crystal faces and showednoevidenceof microbial boring activity.
The surfaces of gypsooids exhibit individual crystals havingprismatic/lenticular to lenticular forms. Lenticular forms tend to beassociated with thicker and more pervasive biofilms whereas theprismatic/lenticular forms have sparser biofilms. Lenticular crystalsare seldom N5 μm thick and had anhedral habits with round edges.The flattened surfaces of lenticular crystals are either smooth, orribbed and irregular. The irregular edges have sawtooth ridges or rib-likemorphologies (Fig. 4c, ‘x’ and ‘*’ respectively). Prismatic/lenticularcrystals are thicker (N5 μm) than lenticular crystals and exhibit a b
Fig. 5. Calcium carbonate (‘cc’) from gypsum crusts (a–c) and gypsolite (d–f) exhibited acbiofilms (‘bf’) that apparently replaced the gypsum matrix (‘gy’; b). Rounded carbonate gcarbonate grains shown in (f).
{010} pinacoid adjoined by prism faces with shelved texture (Fig. 4d).The flattened faces have irregular textures and faint edges thatintersect the acute point of the b{010} pinacoid (arrows, Fig. 4d). Theprism faces are interpreted to be the {110} faces, and the flattenedsurfaces are interpreted to be l{111} and e{103} forms.
4.2.3. Calcium carbonateCalcium carbonate occurs in trace amounts in the dissolution zone
of crystalline deposits of the saltern and anchialine pools of the sabkhawhere gypsum precipitation predominates over the development ofbenthic biofilms. Neither type of Ca-carbonate occurred in greatenough abundance for crystal structure identificationwith XRD. UsingEDS, acicular forms were identified within biofilms in dissolved re-gions of gypsum crusts and as residues on irregular corroded gypsumgrains (Fig. 5a). Crystals occur in radiating clusters, embedded in ordrapedwith biofilms, lodged inmicron scale gypsum cavities (Fig. 5b).Most carbonate needles are 1 to 2 μm inwidth and 5 to 15 μm in lengthwith irregular splintered or steeply pyramidal terminations (Fig. 5c).
icular and rounded morphologies. Acicular morphologies occurred as clusters withinrains had mammillated textures and were penetratively twinned (e). EDS spectra of

272 M.B. Vogel et al. / Sedimentary Geology 223 (2010) 265–280
Elongated grains are usually multifaceted and sometimes penetra-tively twinned. Acicular carbonate was associated with biofilms thatappear to corrode the host gypsum surface, indicating gypsumreplacement by carbonate.
Spheroidal calcium carbonate granules are also found in biofilmsdraped on surfaces of prismatic/lenticular and lenticular crystals ofgypsooids (Fig 5d). These crystals occur as ∼1 to 5 μm ovate-shapedgrains with fine sub-μm scale mammillated textures (Fig. 5e).Aggregates exhibit botryoidal textures and occasional penetrativetwinning (‘⁎’ in Fig. 5e). The spheroidal carbonate grains occur mostlyon surfaces of gypsum crystals but are sometimes observed in cavitiesof surface crystals (EDS spectra shown in Fig. 5). On the whole, thesecrystals appear to develop without significant corrosion of thegypsum host matrix. Ovate Ca-carbonate grains were more abundantamong prismatic/lenticular gypsum petals.
4.2.4. Black mantle gypsum: subaqueous; sediment–water interface; LGNpools
In shallow anchialine pools of LGNwith density- and temperature-stratified brines, gypsum forms as crusts with dense, darkly coloredpigmented microbial communities. Gypsum forms near the bottomsof pools in which temperatures range from 28.7 °C at the surface(136 ppt salinity) to 40.1 °C at depth (220 ppt). Concentration factorsindicate that this salinity gradient spans the gypsum precipitationdomain with the upper region near the beginning stage of gypsumsaturation and the lower region nearing halite saturation. Crusts arecoated with black to dark green biofilms that exhibit ‘frothy’ orhoneycomb textures in SEM images (Fig. 6a). These carbonaceousbiofilms are highly recalcitrant and show significant Mg, Si and Scomponents according to EDS analyses. XRD analyses howeverindicate that this material has nomineral component. The honeycombstructures may have formed during the preparation procedure due toosmotic collapse of the organic sample matrix. Douglas et al. (2008)reported a similar type of black epilithic biofilm from gypsumdeposits
Fig. 6. Black mantled gypsum had a distinctive biofilm mantle with frothy texture (a). Undeare observed to be entombed by gypsum (‘⁎’ in panel c). Biofilms also develop in cavities (
in Death Valley, CA. In a hydrated state, the Death Valley biofilmdisplayed a cauliform morphology reproduced in environmental SEMimages by Douglas et al. (2008). For Guerrero Negro samples, cm-scale gypsum crystals forming underneath the black biofilm mantleare twinned and opaque. In SEM images these grains appear asstacked tabular pseudo-hexagonal crystals with defect sites occupiedby biofilms (Fig. 6b–c). The extremely thin tabular crystals aredominated by enlarged b faces and distorted prism forms. Edges oftabular leaves of black mantle gypsum were poorly aligned givingaggregate crystal forms a ‘dog-eared’ appearance. Some grains showencrusted spherical and filamentous cells (Fig. 6d) indicating thatprecipitation can outpace the microbial motility and growth in thisenvironment.
4.2.5. Biofilm-supported gypsum: subaqueous; sediment–waterinterface; LGN pools
Gypsum forming within columnar and benthic mat cyanobacterialbiofilms includes a variety of euhedral to anhedral morphologiesranging in scale from 1 to 0.1 mm, as well as larger lensoidal aggre-gates. Concentration factors indicate that brines from this environ-ment are at the beginning of gypsum precipitation. Euhedral grainshave prominent b{010} pinacoid faces and equant to distorted, semi-elongated morphologies (Fig. 7a). The l{111} faces of some crystalsdisplay striated porosity but many faces have smooth or labiatetexture, indicating biofilm occupancy. Some grains exhibit discretezonation of porosity (Fig. 7a inset, b) indicating episodic crystalliza-tion. Subhedral and anhedral grains are the most prevalent morphol-ogy and exhibit subangular habits, with l, b, m and possible e faces ofsimilar dimensions (Fig 7b). Among distorted grains, the surface areaof the opposing [010] and [0 −1 0] faces of both l and m prisms oftendiffers significantly (Fig. 7c–d), sometimes by as much as a factor ofthree. Equant and distorted forms usually have discernible edges andpoints but also exhibit cavities or irregular surfaces affected bybiofilms (Fig. 7d–e). Anhedral grains have highly distorted or
rlying gypsum was highly tabular in form (b) with distorted prisms. Cells and filamentsd).

Fig. 7. Subaqueousbiofilm-supportedgrains inpanels a–dhavedistinctive aspect ratios, signs of episodic crystallization and irregular textures. The inset inpanel amarked by an ‘⁎’, and thegrain at left in panel b show porosity zonation which indicates episodic crystallization. An equant grain with labiate surface (‘lab.’) is shown in panel b. Distorted grains (c and d) in somecases were asymmetric (c) and exhibited varying degrees of preferential growth among forms (‘b’ labels indicate {010} pinacoid). Enlargedm and l faces are also shown in panel e (withletters e, l,m and b indicating crystal faces). Irregular surfaces and cavities often were draped or filled by biofilms (‘bf’ in panel f).
273M.B. Vogel et al. / Sedimentary Geology 223 (2010) 265–280
flattened and rounded forms (lenticular shape). The anhedrallenticular forms usually have irregular cleaved or rounded crystalfaces and cavities occupied by biofilms (Fig. 7f). Biofilms are observeddraping over areas with irregular textures and occupying surfacesbetween anhedral grains. Larger aggregates (b200 μm) are anhedraland have irregular labiate surfaces and biofilm drapes.
4.2.6. Water column gypsum: subaqueous; water column; LGN poolsGypsum precipitates in the water column as euhedral crystals or
aggregates (∼1 mm diameter) with pervasive penetrative twinningand distinctive void space textures (Fig. 8a–b). Euhedral crystals havewell-developed faceted morphology ranging from prismatic (prom-inent {010} faces) to needle-like (elongated on [001]) forms. Elongatemembers of aggregates sometimes show contact ‘swallowtail’ twin-ning on the [100] plane. Void textures (sub-micron scale pores) areelongate and aligned parallel to [001] on b faces. Third-order m{110}prisms are larger and more developed than l{111} forms, whichusually occurred only as subtle prisms on many crystals. Few e{103}
faces are observed. Water column precipitate crystals do not exhibitirregular labiate textures, flattened surfaces, inclusions, or other ir-regular textures. Sedimentological literature suggests that thesecrystals can fall to the sediment–water interface and accumulateinto laminar beds, in which oriented crystals similar to thosedescribed here may be recognized (see Scholle, 1979).
4.2.7. Biofilm/sediment surface gypsum: subaerial/subsedimentary saltpans; LOL
Gypsum from biofilm/sediment surfaces exhibits diverse euhedralto anhedral morphologies with smooth, rounded and irregularsurfaces. The euhedral to subhedral morphologies include prismaticgrains, and those with equant aspect ratios (equal b, l, and m forms)and distorted forms (Fig. 8c). Distorted and elongated grains arecommon among subhedral morphologies. The l, m and b faces exhibitstriations, shelved textures, and rectangular etch pits, respectively.Some grains contain detrital particles and some displayed penetrativetwinning (Fig. 8d). Enclaves or cavities on surfaces host biofilms with

Fig. 8. Water column precipitate grains and aggregates were prismatic and highly twinned (a and b). Porosity textures for these grains were aligned with crystallographic axes.Biofilm/sediment surface gypsum has a range of prismatic equant and lensoidal habits (c and d). Asterisks in panel c indicate grains with distorted prisms. Euhedral and subhedralexamples may have surficial growths of biofilm-hosted Sr/Ca-sulfate and Mg-hydroxide (panels d and e showing Sr/Ca phase embedded gypsum). Panel f shows EDS analyses ofthese phases. The EDS analysis shown in panel f may incorporate trace amounts of gypsum or biofilm material (small peaks in the Mg-hydroxide spectra).
274 M.B. Vogel et al. / Sedimentary Geology 223 (2010) 265–280
nests of fine (N1 μm thick) acicular, tabular and spheroidal Sr/Ca-sulfate and magnesium hydroxides detected with EDS (Fig. 8e–f). TheSr/Ca-sulfate phase is similar in morphology to celestite butconstituted b1 wt.% of the matrix and thus could not be confirmedby XRD. An unknown phase rich in Mg and O occurs in closeassociated with Sr/Ca-sulfate and biofilms, and exhibits a range ofacicular, thin tabular and blocky morphologies. Anhedral gypsummorphologies are lenticular with highly irregular, cleaved andtextured surfaces (Fig. 9a–b). These grains exhibit similar dimensionsalong [001] and [100] axes and labiate textures with high relief.
A disordered high-Mg calcite (MgxCayCO3) is detected in thebiofilm/sediment surface gypsum via EDS and XRD. The Mg-calcitephase occurs in the sediment surface and immediate subsurface assub-micron scale irregular particles or larger peloidal forms embed-ded in biofilms that coated flattened and distorted gypsum grains
(Fig. 9c–d). The Mg-calcite constituted around 25 wt.% of one samplefrom an uppermost orange/pink biofilm layer that also contained 64%gypsum and 10% halite. Another sample taken from a similar horizonof the biofilm/sediment surface consisted only of gypsum (79%) andhalite (21%) indicating that the Mg-calcite phase is not uniformlydistributed throughout the gypsum surface.
4.2.8. Subsedimentary gypsum: subaerial/subsedimentary salt pans; LOLSubsedimentary discs and cements occur beneath surficial mi-
crobial biofilms and in subsurface sediments of the sabkha, but theyare not found in the subsediment of subaqueous gypsum-precipitat-ing environments. Subsedimentary gypsum formed as micro- to cm-scale biconvex discs (similar to lenticular gypsum, see Cody; 1979;Warren, 2006) or disc aggregates (Fig. 9e). Detrital material is uni-formly distributed within these discs and constitutes around 10% of

Fig. 9. Biofilm/sediment surface gypsum has distinctive lensoidal habits (a–b). Lensoidal grains have irregular high relief surface textures and co-occur with Mg-calcite shown inpanel c. Panel d shows gypsum discs set among Mg-calcite biofilm complex. Subsedimentary gypsum that is not associated with biofilms occurs as flattened and rounded discs withstriations and detrital inclusions (e), or as fibrous cements developing among clastic particles and aggregates (f).
275M.B. Vogel et al. / Sedimentary Geology 223 (2010) 265–280
the matrix volume. Elongate void spaces or striations are orientedparallel to the [001] axis of the discs. Subsedimentary gypsum alsoforms as sinuous cements and frameworks in clastic muds and sands(Fig. 9f). Sinews develops into striated, sheet-like and smoothframeworks enclosing sands, pebbles and shells. Concentration factorsfrom subsedimentary environments indicate that brines were pastmaximum gypsum precipitation and nearing halite saturation.
5. Discussion
Organization of gypsum microtextures and accessory phasesrequires exploration of the links between crystal/grain features andbiological and environmental factors. Of the morphologies describedabove, only the water column precipitate grains and subsedimentarylenticular gypsum are not associated with biofilms. Biological
influence on their formation is thus considered minor or absent.Subsedimentary gypsum is distinguished by both its flat and roundmorphology, and its high proportions of detrital inclusions. Thepresence of sedimentary particles and organic compounds, as well assubsedimentary saturation and pH conditions are known causal factorin development of this morphology (Cody and Cody, 1988a,b; Cody,1991). Water column precipitate grains are euhedral, prismatic toelongated, and highly penetratively twinned. The morphologies ofsubsedimentary gypsum and water column precipitates are distinctfrom morphologies of gypsum associated with biofilms. This lattercategory includes gypsum occupied by endoevaporitic biofilms, andgypsum particles that crystallize while enveloped in a biofilm matrix.Gypsum types that contain endolithic or epilithic biofilms includebottom nucleated crusts, gypsolites and black mantle gypsum.Gypsum types that developwithin biolfims include biofilm-supported

276 M.B. Vogel et al. / Sedimentary Geology 223 (2010) 265–280
gypsum and biofilm/sediment surface gypsum. Together thesebiofilm-associated gypsum types register the effects of the biofilmsin the form of irregular textures, accessory minerals, and distinctiveaspect ratios among crystal faces.
5.1. Irregular textures
Irregular and rounded, labiate surfaces with μm scale globularfeatures are characteristic textures of biofilm-supported gypsum,biofilm/sediment surface gypsum, and the dissolution zones ofgypsum crusts. These textures are absent in water column precipitatesamples and subsedimentary gypsum types that formed in theabsence of well-developed biofilms. Gypsum in areas occupied bybiofilms exhibits rounded edges, labiate surfaces, cavities and highmorphological relief textures. Dissolution zones in the interior ofgypsum crusts shows rounded and globular textures. Pits, borings,and networks of etchings are not observed, indicating that euendo-lithic processes do not create these irregular textures. Dissolutionsurfaces in gypsum have been recognized by previous workers in bothmodern and ancient deposits (Vai and Lucchi, 1977; Douglas, 2004).Vogel et al. (2009) discuss potential mechanisms by which the biofilmEPSmay influence precipitation and dissolution of gypsum but furtherstudy is necessary to resolve the biofilm's specific role in theformation of irregular surface textures. The unique association ofirregular textures with biofilms in samples from several differentdepositional settings however demonstrates a potential utility ofthese features in supporting biofacies interpretations.
5.2. Biofilms and accessory phases—sulfur
Elemental sulfur (S°)was detectedwith EDS as 1- to 5-μm inclusionsand globules in dissolution zone biofilms of gypsum crusts, but was notdetectedwith XRD. Elemental sulfur is present inmany ancient gypsumdeposits and can be formed by a range of syn-depositional and post-depositional processes (e.g. Schreiber and Kinsman; 1975; Machel,2001, and references therein). The S° described here could be a non-crystalline phase such as polysulfide or cyclic S°, both of which weredetected in abundance in solvent-extracted organic moieties fromgypsum crust biofilms (Vogel et al., 2006). Non-crystalline phasesconstitutemost of the S° inmarine sediments and are often observed insediments with high rates of sulfate reduction (see Zopfi et al., 2004).These phases can arise from abiotic sulfide oxidation by Fe(III) and Mn(IV) oxides, oxidation of FeS, and as intermediate species in bacterialsulfide oxidation. Bacterial sulfur storage globules from variousphotosynthetic and non-photosynthetic species have been character-ized as non-crystalline α-S8 and sulfur chains encapsulated byfunctionalized sulfur moieties (Pickering et al., 2001; Prange et al.,2002). The size distribution of sulfur particles from our samples isconsistent with that of microbial inclusions as reported in Stolz (1990).
Oren et al. (2009) encountered purple single celled phototrophsand colorless bacteria containing sulfur globules in gypsum crustsfrom Eilat, Israel. In a study of saltpan evaporites from Death Valley,CA, Douglas (2004) reported crystalline S° (rosickyite; γ-sulfur)associated with phototrophic biofilms and corroded grain surfaces.Non-crystalline S° in samples from Guerrero Negro also occurred nearthe base of the uppermost phototrophic layers near the dissolutionzones within the gypsum crusts, but was volumetrically less abundantthan the rosickyite, which was described as a major mineral phase inthe Death Valley samples (Douglas, 2004). Douglas and Yang (2002)attributed rosickyite formation in the interior of the Death Valleygypsum crusts to a high pH (N7.9) and activity of sulfide-oxidizingbacteria located at an O2 and H2S interface. Studies of similar gypsumdeposits have measured microbial sulfide oxidation occurring inlower layers of the gypsum crust and found that dynamics betweensulfide and oxygen species were spatially complex and temporallyvariable (Sørensen et al., 2004; Canfield et al., 2004; Sørensen et al.,
2005; Oren et al., 2009). In spite of the complexity of chemicalgradients within the crust, elemental sulfur species can be interpretedas intermediates of redox cycling of sulfur by microbial sulfatereduction and the various sulfide oxidation processes. Sulfur species,minerals, isotopic patterns, and textures provide evidence of thiscycling (Visscher et al., 2000; Machel, 2001; Tekin, 2006).
5.3. Ca-carbonate
Acicular and spheroidal Ca-carbonate were observed via SEM insamples of subaqueous gypsum in the dissolution zone of crusts andwithin biofilms located on gypsooid surfaces. Mg-calcite wasdefinitively identified using XRD in samples of biofilm/sedimentsurface gypsum. The acicular carbonate always occurs in associationwith biofilms and corrosion features, in an apparent replaciverelationship with the gypsum host matrix. The spheroidal carbonateoccurred with gypsooid-hosted biofilms that were mostly epilithicand did not appear to replace gypsum phases. The contrastingoccurrences of carbonate in subaqueous environments may be relatedto the greater porosity of the gypsooid framework relative to that ofthe crusts fabrics, and due to the effects of porosity on redox gradientsand carbonate alkalinity. Rates of organic matter burial and oxidation(and thus carbonate alkalinity) could also be greater in depositionalenvironments where wind action and agitation lead to gypsooidformation. Schultze-Lam and Beveridge (1994) and Thompson andFerris (1990) showed epicellular biomineralization of gypsum, calcite,celestite and magnesite by Synechoccocus sp. from alkaline lakeenvironments but few other studies have addressedmicrobial ecologyof primary carbonate precipitation in gypsiferrous environments.
The acicular carbonate occurs where biofilm-associated texturesindicate gypsum dissolution inside the crust. Microbially mediateddiagenetic replacement of gypsum by carbonate has been documen-ted in soils, and in marine and lacustrine sediments (see Machel,2001). Peckman et al. (1999) described neoformed diageneticaragonite with organic aggregates and S° in Permian Zechstein car-bonates. Gypsum replacement by dolomite in Miocene sedimentaryunits is described in Pierre and Rouchy (1988), Sanz-Montero et al.(2006) and Ayllón-Quevedo et al. (2007). These reports cited grainmorphology, enrichment of biologically significant elements, andstable isotope values as evidence of early diagenetic or syn-depositional bacterial sulfate reduction using gypsum as a sulfatesource. The enrichment of elements such as Mg, P, S, Si, Al, and Fe inthe biofilms examined here supports the link between elementalenrichment and the biogenicity of carbonates in these reports. Featuresthat have been cited as evidence of former endolithic microbialcommunities include irregular spheroid, rod-shaped and filamentousmorphologies assumed by carbonate accessory phases, aswell as boringtextures, corrosion and pitting of the gypsum host crystals (Peckmanet al., 1999; Sanz-Montero et al., 2006; Barbieri et al., 2006; Ayllón-Quevedo et al., 2007; Panieri et al., 2008).
5.4. Mg-calcite
A disordered Mg-calcite (MgxCayCO3) were the only authigenicmineral found in significant abundance (∼25 wt.%)within gypsum froma biofilm/sediment surface of the LOL sabkha. The Mg-calcite occurs asfine-grained precipitates enveloped within biofilms. As with otheraccessory phases, Mg-calcite precipitates in near surface phototrophicbiofilms and therefore is interpreted to be a biologically induced phase.Magnesian carbonates have been found in association with gypsum inboth modern environments and ancient sediments. Mg-calcite similarto thephase reportedherewasdetectedwithin gypsifiedmicrobialmatsfrom an anchialine pool in the Bahamas (Douglas, 2002), and frommicrobial mats in Miocene lacustrine evaporites of the Madrid Basin(Sanz-Montero et al., 2006; described as a poorly ordered, Ca-rich, non-stoichiometric dolomite). Magnesite and dolomite have been

277M.B. Vogel et al. / Sedimentary Geology 223 (2010) 265–280
documented in associationwith gypsum in strata fromLate Tertiary andQuaternary basins connected to the paleo-Mediterranean (Pérez et al.2002; Sanz-Montero et al., 2006; Barbieri et al., 2006;Ayllón-Quevedoetal., 2007). With regard to modern environments, Thompson and Ferris(1990) demonstrated epicellular precipitation of gypsum and magne-sitebySynechococcus sp. under alkaline freshwater conditions. Dolomiteand Mg-calcite precipitation has also been reported from modernhypersaline lagoons in Brazil where it has been linked with sulfideoxidation based on pore water composition and isotopic signatures(Vasconcelos and McKenzie, 1997; Moreira et al., 2004). These lagoonshowever were below gypsum saturation and high-Mg calcite may haveformed under contrasting pH and salinity conditions relative to thosenecessary for dolomite precipitation (Arvidson and MacKenzie; 1999;Moreira et al., 2004). Precipitation ofMg-calcite in the biofilm/sedimentsurfaces at Guerrero Negro was likely driven by both microbialprocesses that generate carbonate alkalinity in biofilms (see Corzo etal., 2005; Dupraz et al., 2008), and by the high salinity of sabkha brinesheets. Folk (1974) showed that Mg content of carbonates increaseswith increasingMg2+ and Na+ concentrations in the parent brines, andcategorized Mg-calcites as typical of hypersaline and sabkha environ-ments. Brines sheets from the LOL sabkhas had unusually high-Mgcontents and lower Ca concentrations than brines from subaqueousenvironments (see Table 1).
5.5. Sr/Ca-sulfate and Mg-hydroxides
Sr/Ca-sulfate and Mg-hydroxides occur in biofilm/sedimentsurface gypsum as tabular, acicular and spheroidal forms. Theseminerals are interpreted to be biofilm-associated replacement phasesbased on their occupancy of cavities within gypsum host crystals. Thebrine associatedwith these sabkha samples (biofilm/sediment surfaceand subsedimentary gypsum) had the highest concentrations of mostmajor ions with the exception of Sr. Low Sr concentrations may havebeen due to Sr uptake by the Sr/Ca-sulfate phase. Celestite has beenshown to occur as an early diagenetic phase associated with gypsum(Evans and Shearman, 1964) and has been identified in gypsumreplacement fabrics with organic matter and dolomite (Rodríguez-Aranda et al., 2005; Sanz-Montero et al., 2006). Taberner et al. (2002)used δ34S and δ18O isotopes to argue that celestite in Eocene marinecarbonates was formed from residual sulfate after bacterial sulfatereduction. Douglas (2004) found strontianite and celestite precipitat-ed on mineralized cells in interior zones gypsum crusts from DeathValley salt pans. Schultze-Lam and Beveridge (1994) demonstratedsimilar cyanobacterial celestite biomineralization in culture experi-ments. Magnesium hydroxides such as those associated with biofilmSr/Ca-sulfates typically do not precipitate from marine brines. Brucite(Mg(OH)2) has been reported in trace amounts in association withprimary low temperature evaporitic sulfates and carbonates (Sedel-nikov et al., 1968; Rodgers and Courtney, 1988). Brucite has beenshown to form at low temperatures, very high alkalinities, and lowpCO2 (Gulbrandsen and Cremer, 1970; Luce, 1971). Biologicallyinduced brucite formation has been reported from coral microenvir-onments inhabited by biofilms (Nothdurft et al., 2005) and from acultured diatom species (Tesson et al., 2008). These reports linkedbrucite formation with photosynthetically induced alkalization andlowering of pCO2, and with adsorption of Mg2+ by biofilms. Similarprocesses may have occurred within biofilm/sediment gypsummicroenvironments with photosynthetic biofilms facilitating Mg-hydroxide precipitation.
5.6. Gypsum crystal forms: dimensions and aspect ratios
Both subaqueous and biofilm/sediment surface samples forming inthe presence of biofilms exhibit distinctive aspect ratios compared tothose forming where biofilms were absent or sparse (i.e. watercolumn grains and subsedimentary gypsum discs). Gypsooid surface
crystals exhibit flattening along [001] and distinctive [hk0] edges.Black mantle gypsum crystals also exhibit enigmatic distorted tabularhabits. Both of these gypsum types however develop under unusualconditions of high salinity, density stratification, wave action, etc.These conditions may eclipse the effects of biofilms occupancy oncrystal habit modification. Discussion of aspect ratios is thereforelimited to biofilm-supported and sediment/biofilm gypsum types.Both of these types of gypsum crystals are enveloped in biofilmsduring growth, develop in sedimentary environments with lowturbulence or water column agitation, and exhibit compellingevidence of interfacial growth preferences. The biofilms are alsohighly enriched in elemental impurities (Mg, Si, S, P, Al, Fe, etc.) asevidenced by the EDS analysis of biofilm residues.
The preponderant equant and distorted to lenticular habits ofbiofilm grains suggest biofilm occlusion may favor growth of l and mprisms along with e pinacoids over that of b pinacoids. Smallerrelative b{010} pinacoids may formed due to high concentrations ofimpurities, organic and enzymatic compounds, or unknownmicrobialinteractions during precipitation. Many biofilm occluded grains havem faces similar in size to l prisms butm prism faces never significantlyexceed l prism faces in size. Equant and distorted grains comprise asignificant subpopulation of biofilm gypsum crystals, indicating thatbiofilms interact with specific faces or symmetry elements of crystals.Lenticular crystals associated with biofilms retain evidence of l prismforms or exhibit highly rounded surfaces (l prisms not discernible)with irregular, high relief textures (e.g. Fig. 8d). The l faces are absentfrom the surfaces of biofilm-free subsedimentary gypsum discs.
Studies of diverse environmental compounds have shown thatinhibitor induced habit modification ultimately favors the l prismleading to lenticular rounding (Cody and Cody, 1989a; Cody; 1991).Equant gypsum crystals (i.e. those having proportionate m and lprisms) have not been specifically described in previous reports.Distorted crystals are described in Cody and Cody (1991) as productsof controlled crystallization in the presence of isotropic additives.These include stereospecific enzymes, biologically relevant organo-phosphates and carboxylic acids. The distorted biofilm-hosted crystalmorphologies described here could similarly have resulted from thechirality of the predominant inhibitors present in the biofilm.
Due to the importance of gypsum in industrial applications, aconsiderable body of research has sought to elucidate parameters thataffect gypsum shape, size and other crystallization factors (seediscussion in Cody, 1991). Studies addressing the physiochemistryof gypsum precipitation have found that growth in the crystallo-graphic c direction is favored under saturation conditions in theabsence of inhibitors due to differences in the specific surface tensionof molecular units that comprise each face (Hünger and Henning,1988; [001] indexed therein as the ‘a’ direction). Other studies havedemonstrated how selective adsorption (a variant of surface energy)of cations and anions leads to differing growth rates on each face class.Specifically, l{111} faces comprised of exposed Ca2+ ions offerpreferential sites for adsorption of foreign anions, m{110} facesconsisting of both Ca2+ and SO4
2− ions offer sites for both positive andnegative adsorbants but are preferentially bound by foreign cations(Edinger, 1973). Specific surface terminations (configurations of ionson a given surface) and corresponding surface energies of gypsumcrystal forms are computed in Hill and Plank (2004). These workersreported differential binding effects each surface by organic acidscommonly used as growth retardants in industrial applications. Hilland Plank (2004) concluded that under alkaline conditions, ioniccomposition of terminations and their corresponding surface energiesof each crystal face are determining factors in the growth andmorphology of gypsum. By these mechanisms, crystal forms that arebound by foreign ions grow more slowly while other forms growrapidly, with the bound or inhibited faces becoming enlarged relativeto other faces. Barcelona and Atwood (1978) demonstrated selectiveinhibition of the l{111} face by free fatty acids and attributed their

278 M.B. Vogel et al. / Sedimentary Geology 223 (2010) 265–280
results to the higher relative density of Ca2+ ions on the l{111} face. Asimilar study by Dogan et al. (2004) attributed this effect to the similarspacing of Ca2+ ions the l{111} face (3.7 Å), to the spacing of functionalgroups on carboxylic acids (3.5 Å). Organic acids such as carboxylic,phosphoric, phosphonic and phenolic hydroxyl compounds that arereadily deprotonated (anionic), are biologically relevant and havebeen demonstrated to be highly effective crystallization inhibitors(Cody, 1991). These prior observations support the inference thatbiofilm with predominantly anionic functional groups can effectivelybind the l{111} surfaces, leading to rapid growth and diminished sizeofm and b forms. Equant crystals observed to have largerm{110} facesindicate that biofilmsmay also have selectively inhibited growth of them prism (Fig. 10). Enlarged relativem faces raise the specter of cationicfunctional groups in addition to anionic functional groups as potentialinhibitors in the biofilm environment. Differential growth of differentfaces could also be due to lower relative surface energy of the m facesunder alkaline conditions, corresponding to theoretical findings of Hilland Plank (2004).
6. Conclusions
Petrographic and geochemical analyses of samples from sabkhaand saltern environments at Guerrero Negro, Baja California Sur,Mexico reveal textural characteristics in gypsum that can be linked toenvironmental and biological factors. Gypsum from subaqueousenvironments includes bottom nucleated crusts, gypsolites, watercolumn precipitates and individual gypsum grains precipitated withinbenthic microbial biofilms. Gypsum forming at the LOL sabkha saltpans includes gypsum developing in sediment surface biofilms and
Fig. 10. Summary illustration of possiblemechanisms for differential development of l{111} andsurface have surface terminations dominated by exposed Ca2+ ions, whereas planes of them{reader to Hill and Plank (2004) and animations of gypsum crystal lattice onwww.mindat.org fogrowth of the other crystal faces (c). Equant crystal morphologies could be caused by lower regrowth rate of the l{111} prism (b). A higher relative growth rate (t) of any surface will lead t
gypsum from the subsediment. Water column precipitates andsubsedimentary gypsum discs develop in the absence of biofilms.Water column precipitates are characterized by euhedral prismatic toelongate habits, homogeneous, porous surface textures and pervasivepenetrative twinning. Subsedimentary gypsum discs are highlyrounded, flattened along [001], and have sinuous axial surfacetextures with detrital inclusions. Gypsum precipitated in the presenceof biofilms includes bottom nucleated crusts, gypsolites, black mantlegypsum, biofilm-supported gypsum, and sediment/biofilm gypsum.Each of these gypsum types develops in subaqueous environmentsexcept sediment/biofilm gypsumwhich occurs on mudflat surfaces orjust beneath salt pan crusts.
Irregular textures in high relief, accessory minerals and distinctivecrystal face aspect ratios are interpreted as evidence of the influenceof biofilms during gypsum formation. Irregular textures includelabiate and globular features associated with both dissolution andcrystallization processes that are likely affected by biofilm micro-environments. The accessory phases forming in association withgypsum-hosted biofims include non-crystalline S°, Ca-carbonate, Mg-calcite Sr/Ca-sulfate and Mg-hydroxides. With the exception of Mg-hydroxides, these phases have been recognized as products orintermediates of microbially mediated sulfate reduction in bothmodern environments and ancient sedimentary deposits. The Ca-carbonate detected in gypsum samples studied here is volumetricallyscarce but demonstrates that carbonate can arise from earlydiagenetic replacement driven by biofilms. Biofilm-supported andsediment/biofilm gypsum differs from morphological types devel-oped in the absence of pervasive biofilms in terms of their crystalaspect ratios and irregular surface textures. Populations of biofilm
m{011} prisms. In atomic packing of the gypsumcrystal, planes parallel to the l{111} prism011} prisms have interspersed rows of Ca2+ and SO4− ions exposed (c). The ‘⁎’ refers ther further details. Anions (besides sulfate) can thus bind the l{111} surfaces leading to rapidlative surface energy of them{011} prism and binding therein that leads to higher relativeo enlargement of the other surfaces, which grow at a slower rate.

279M.B. Vogel et al. / Sedimentary Geology 223 (2010) 265–280
gypsum crystals include prismatic to lensoidal morphologies withunique equant and distorted intermediates. The equant to distortedmorphologies have been attributed to form and face specific in-hibition by biologically relevant functional groups (Cody and Cody1991; Cody, 1991). The biofilm thus likely plays an active role in brinesaturation states, and crystallization phases of these crystals, signs ofwhich are been preserved in grain morphology.
Acknowledgements
This work was funded by grants from the NASA Exobiology andEvolutionary Biology program and the NASA Astrobiology Institute(NAI) to DJD and the Ames Team of the NAI. MBV acknowledgessupport from Oak Ridge Associated Universities NASA PostdoctoralProgram. We also thank the staff of the Stanford/U.S.G.S SUMACfacility for microscopy support. F. Mazdab of the U.S. GeologicalSurvey provided helpful feedback on crystallographic descriptions. D.Blake and D. Cardace are thanked for use of and assistance with theTerra XRD/XRF instrument. The manuscript greatly benefitted fromreviews by C. Pierre and B.C. Schrieber.
References
Anadón, P., Rosell, L., Talbot, M.R., 1992. Carbonate replacement of lacustrine gypsumdeposits in two Neogene continental basins, eastern Spain. Sediment. Geol. 78,201–216.
Aref, M.A.M., 1998. Holocene stromatolites and microbial laminites associated withlenticular gypsum in a marine-dominated environment, Ras El Shatan area, Gulf ofAqaba, Egypt. Sedimentology 45, 245–262.
Arvidson, R.S., MacKenzie, F.T., 1999. The dolomite problem: control of precipitationkinetics by temperature and saturation state. Am. J. Sci. 299, 257–288.
Ayllón-Quevedo, F., Souza-Egipsy, V., Sanz-Montero, M.E., Rodriguez-Aranda, J.P., 2007.Fluid inclusion analysis of twinned selenite gypsum beds from the Miocene of theMadrid basin (Spain). Implication on dolomite bioformation. Sediment. Geol. 201,211–230.
Barbieri, R., Stivaletta, N., Mariangeli, L., Ori, G.G., 2006. Microbial signatures in sabkhaevaporite deposits of Chott el Gharsa (Tunisia) and their astrobiological implica-tions. Planet. Space Sci. 54, 726–736.
Barcelona, M.J., Atwood, D.K., 1978. Gypsum–organic interaction in the natural seawater:effect of organics on precipitation kinetics and crystal morphology. Mar. Chem. 6,99–115.
Bibring, J.P., Langevin, Y., Mustard, J.F., Poulet, F., Arvidson, R., Gendrin, A., Gondet, B.,Mangold, N., Pinet, P., Forget, F., 2006. Global mineralogical and aqueous Marshistory derived from OMEGA/Mars express data. Science 312, 400–404.
Bosbach, D., Hochella, M.F., 1996. Gypsum growth in the presence of growth inhibitors:a scanning force microscopy study. Chem. Geol. 132, 227–236.
Bosbach, D., Ramensee, W., 1994. In situ investigation of growth and dissolution of the(010) surface of gypsum by Scanning ForceMicroscopy. Geochim. Cosmochim. Acta58, 843–849.
Buck, B.J., Van Hoesen, J.G., 2002. Snowball morphology and SEM analysis of pedogenicgypsum, southern New Mexico, U.S.A. J. Arid Environ. 51, 469–487.
Canfield, D.E., Sørensen, K.B., Oren, A., 2004. Biogeochemistry of a gypsum-encrustedmicrobial ecosystem. Geobiology 2, 133–150.
Chang, L.L.Y., Howie, R.A., Zussman, J., 1995. Non-silicates: Sulphates, Carbonates,Phosphates, Halides (Rock-forming Minerals). Longman Group, Harlow, UK.
Cody, R.D., 1979. Lenticular gypsum: occurrence in nature and experimental determina-tions of soluble green plant material on its formation. J. Sediment. Petrol. 49,1015–1028.
Cody, R.D., 1991. Organo-crystalline interactions in evaporite systems: the effects ofcrystallization inhibition. J. Sediment. Petrol. 61, 704–718.
Cody, R.D., Cody, A.M., 1988a. Gypsum nucleation and crystal morphology in analogueterrestrial environments. J. Sediment. Petrol. 58, 247–255.
Cody, R.D., Cody, A.M., 1988b. Tannic acid induced penetration twinning in gel-growncalcium sulfate dihydrate. Cryst. Res. Technol. 23, 1437–1441.
Cody, R.D., Cody, A.M., 1989a. Evidence for micro-biological induction of (101)Montmartre twinning of Gypsum (CaSO4·2H2O). J. Cryst. Growth 98, 721–730.
Cody, R.D., Cody, A.M., 1989b. SEM and polarization analyses updating early lightmicroscope studies related to 101 twin formation in gypsum. J. Cryst. Growth 98,731–738.
Cody, R.D., Cody, A.M., 1991. Chiral habit modifications of gypsum from epitaxial-likeadsorption of stereospecific growth inhibitors. J. Cryst. Growth 113, 508–519.
Cole, W.F., Lancucki, C.J., 1974. A refinement of the crystal structure of gypsumCaSO4·2H2O. Acta Cryst. B30, 921–929.
Corzo, A., Luzon, A., Mayayo, M.J., van Bergijk, S.A., Mata, P., Garcia de Lomas, J., 2005.Carbonate mineralogy along a biogeochemical gradient in recent lacustrinesediments of Gallocanta Lake (Spain). Geomicrobio. J. 22, 283–298.
Dana, E.S., 1892. The System of Mineralogy, Sixth Edition with Appendices I, II, and III,Completing the Work to 1915. John Wiley & Sons, New York.
Dogan, Ö., Akyol, E., Öner, M., 2004. Polyelectrolytes inhibition effect on crystallizationof gypsum. Cryst. Res. Technol. 39, 1108–1114.
Douglas, S., 2002. ESEM-EDS and XRD study of micromineralogical layering in amicrobial mat from a hypersaline pond on Lee Stocking Island, Bahamas: formationof celestite in microbial exopolymers. Geological Society of America AnnualMeeting, Abstract with Programs. 216-4.
Douglas, S., 2004. Microbial biosignatures in evaporite deposits: evidence from DeathValley, California. Planet. Space Sci. 52, 223–227.
Douglas, S., Yang, H., 2002. Mineral biosignatures in evaporites: presence of rosickyitein an endoevaporitic microbial community from Death Valley, California. Geology30, 1075–1078.
Douglas, S., Abbey, W., Mielke, R., Conrad, P., Kanik, I., 2008. Textural and mineralogicalbiosignatures in an unusual microbialite from Death Valley, California. Icarus 193,620–636.
Dupraz, C., Reid, P.R., Braissant, O., Decho,A.W., Norman, R.S., Visscher, P.T., 2008. Processesof carbonate precipitation in modern microbial mats. Earth-Sci. Rev. 11–17 avail.
Edinger, S.E., 1973. The growth of gypsum. J. Cryst. Growth 18, 217–224.Evans, G., Shearman, D.J., 1964. Recent celestine from the sediments of the Trucial Coast
of the Persian Gulf. Nature 202, 385–386.Fan, C., Teng, H.H., 2007. Surface behavior of gypsum during dissolution. Chem. Geol.
245, 242–253.Folk, R.L., 1974. The natural history of crystalline calcium carbonate: effect of
magnesium content and salinity. J. Sediment. Petrol. 44, 40–53.Fontes, J.C., Matray, J.M., 1993. Geochemistry and origin of formation brines from the
Paris Basin, France. Chem. Geol. 109, 177–200.Gandin, A., Wright, D.T., Melezhik, V., 2005. Vanished evaporites and carbonate
formation in the Neoarchaean Kogelbeen and Gamaohaan formations of theCampbellrand Subgroup, South Africa. J. Afr. Earth Sci. 41, 1–23.
Gulbrandsen, R.A., Cremer, M., 1970. Coprecipitation of carbonate and phosphate fromseawater. U.S.G.S. Prof. Paper 700-C, pp. 125–126.
Hardie, L.A., Lowenstein, T.K., Spencer, R.J., 1985. The problem of distinguishing betweenprimary and secondary feature in evaporates. In: Schreiber, B.C. (Ed.), 6th Symposiumon Salt. Salt Institute, Alexandria, Virginia, pp. 1–59.
Hill, J.R., Plank, J., 2004.Retardationof settingofplasterofParisbyorganicacids:understandingthe mechanism through molecular modeling. J. Comput. Chem. 25, 1438–1448.
Hünger, K.J., Henning, O., 1988. On the crystallization of gypsum from supersaturatedsolutions. Cryst. Res. Technol. 23, 1135–1143.
Luce, R.W., 1971. Brucite identified as crystallizing from a natural alkaline spring gel.Clays Clay Miner. 19, 335–336.
Machel, H.G., 2001. Bacterial and thermochemical sulfate reduction in diageneticsettings — old and new insights. Sediment. Geol. 140, 143–175.
McCaffrey, M.A., Lazar, B., Holland, H.D., 1987. The evaporation path of seawater and thecoprecipitation of Br− and K+ with halite. J. Sediment. Petrol. 57, 928–937.
McLennan, S.M., Bell, J.F., Calvin, W.M., Christensen, P.R., Clark, B.C., de Souza, P.A.,Farmer, J., Farrand,W.H., Fike, D.A., Gellert, R., Ghosh, A., Glotch, T.D., Grotzinger, J.P.,Hahn, B., Herkenhoff, K.E., Hurowitz, J.A., Johnson, J.R., Johnson, S.S., Jolliff, B.,Klingelhofer, G., Knoll, A.H., Learner, Z., Malin, M.C., McSween Jr., H.Y., Pocock, J.,Ruff, S.W., Soderblom, L.A., Squyres, S.W., Tosca, N.J.,Watters,W.A.,Wyatt,M.B., Yen,A., 2005. Provenance and diagenesis of the evaporite-bearing Burns formation,Meridiani Planum, Mars. Earth Planet. Sci. Lett. 240, 95–121.
Michalzik, D., 1996. Lithofacies, diagenetic spectra and sedimentary cycles of Messinian(Late Miocene) evaporites in SE Spain. Sediment. Geol. 106, 203–222.
Moreira, N.F., Walter, L.M., Vasconcelos, C., McKenzie, J.A., McCall, P.J., 2004. Role ofsulfide oxidation in dolomitization: sediment and pore-water geochemistry of amodern hypersaline lagoon system. Geology 32, 701–704.
Murchie, S., and the CRISM Science and EngineeringTeams, 2007. First results from theCompact Reconnaissance Imaging Spectrometer for Mars (CRISM). LPSC XXXV111,League City, TX, LPI Contr. No. 1338, p. 1478.
Nothdurft, L.D., Webb, G.E., Buster, N.A., Holmers, C.W., Sorauf, J.E., Kloprogge, J.T., 2005.Brucite microbialites in living coral skeletons: indicators of extreme microenviron-ments in shallow marine settings. Geology 33, 169–172.
Oren, A., Sorensen, K.B., Canfield, D.E., Teske, A.P., Ionescu, D., Lipski, A., Altendorf, K.,2009. Microbial communities and processes within a hypersaline gypsum crust in asaltern evaporation pond (Eilat, Israel). Hydrobiologia 626, 15–26.
Panieri, G., Lugli, S., Manzi, V., Palinska, K.A., Roveri, M., 2008. Microbial communities inMessinian evaporite deposits of the Vena del Gesso (northern Apennines, Italy).Stratigraphy 5, 347–356.
Peckman, J., Paul, J., Thiel, V., 1999. Bacterially mediated formation of diageneticaragonite and native sulfur in Zechstein carbonates (Upper Permian, CentralGermany). Sediment. Geol. 126, 205–222.
Pedersen, B.F., Semmingsen, D., 1982. Neutron diffraction refinement of the structure ofgypsum, CaSO4×2H2O. Acta Cryst. B38, 1074–1077.
Pérez, A., Luzón, A., Roc, A.C., Soria, A.R., Mayayo, M.J., Sánchez, J.A., 2002. Sedimentaryfacies distribution and genesis of a recent carbonate-rich saline lake: GallocantaLake, Iberian Chain, NE Spain. Sediment. Geol. 148, 185–202.
Phleger, F.B., 1962. Sedimentology of Guerrero Negro Lagoon, Baja California, Mexico.Contribution No. 48, Marine Foraminifera Laboratory. Contribution from theScripps Institute of Oceanography, San Diego, CA, pp. 205–237.
Pickering, I.J., George, G.N., Yu, E.Y., Brune, D.C., Tuschak, C., Overmann, J., Beatty, J.T.,Prince, R.C., 2001. Analysis of sulfur biochemistry of sulfur bacteria using X-rayabsorption spectroscopy. Biochemistry 40, 8138–8145.
Pierre, C., 1983. Polyhalite replacement after gypsum at Ojo de Liebre Lagoon (BajaCalifornia, Mexico): an early diagenesis by mixing of marine brines and continentalwaters. 6th Int. Symp. on Salt, vol. 1, pp. 257–265.
Pierre, C., Rouchy, J.M., 1988. Carbonate replacements after sulfate evaporites in theMiddle Miocene of Egypt. J. Sediment. Petrol. 58, 446–456.

280 M.B. Vogel et al. / Sedimentary Geology 223 (2010) 265–280
Pierre, C., Ortlieb, L., Person, A., 1984. Supratidal evaporitic dolomite at Ojo de LiebreLagoon: mineralogical and isotopic arguments for primary crystallization. J.Sediment. Petrol. 54, 1049–1061.
Prange, A., Chauvistre, R., Modrow, H., Hormes, J., Truper, H.G., Dahl, C., 2002. Quantitativespeciation of sulfur in bacterial sulfur globules: X-ray absorption spectroscopy revealsat least three different species of sulfur. Microbiology 148, 267–276.
Rodgers, K.A., Courtney, S.F., 1988. Mineral records from Funafuti, Tuvalu: gypsum,brucite, ettringite. Miner. Mag. 52, 411–414.
Rodríguez-Aranda, J.P., Sanz-Montero, M.E., Ayllón-Quevedo, F., Souza-Egypsi, V., 2005.Formación de celestina dentro de un contexto microbiano en ambiente lacustresalino. Mioceno inferior de la Cuenca de Madrid. Macla 3, 171–172.
Runnegar, B., Dollase,W.A., Ketcham, R.A., Colbert, M., Carlson,W.D., 2001. Early Archaeansulfates from Western Australia first formed as hydrothermal barites not gypsumevaporates. Geol. Soc. of Amer., Annual Meeting and Exposition Abstracts A404.
Sanz-Montero, M.E., Rodríguez-Aranda, J.P., Calvo, J.P., 2006. Mediation of endoeva-poritic microbial communities in early replacement of gypsum by dolomite: a casestudy from Miocene Lake deposits of the Madrid Basin, Spain. J. Sediment. Res. 76,1257–1266.
Scholle, P., 1979. Constituents, textures, cements, and porosities of sandstones andassociated rocks. AAPG Mem. 28.
Schreiber, B.C., Kinsman, D.J., 1975. New observations on the Pleistocene evaporites ofMontallegro, Sicily and a modern analog. J. Sediment. Petrol. 45, 469–479.
Schreiber, B.C.El, Tabakh, M., 2000. Deposition and early alteration of evaporites.Sedimentology 47, 215–238.
Schultze-Lam, S., Beveridge, T.J., 1994. Nucleation of celestite and strontianite on acyanobacterial S-Layer. Appl. Environ. Microbiol. 60, 447–453.
Sedelnikov, G.S., Pustylnikov, A.M., Bailieva, G.G., 1968. Mineralogical-petrographicstudies of sediments in the Kara-Bogaz-Gol mixing zone. Chem. Abstr. 70 60,695 k.
Shen, Y., Buick, R., Canfield, D.E., 2001. Isotopic evidence for microbial sulphatereduction in the Early Archaean era. Nature 410, 77–81.
Sørensen, K.B., Canfield, D.E., Oren, A., 2004. Salinity responses of benthic microbialcommunities in a solar saltern (Eilat, Israel). Appl. Environ. Microbiol. 70, 1608–1616.
Sørensen, K.B., Canfield, D.E., Teske, A.P., Oren, A., 2005. Community composition of ahypersaline endoevaporiticmicrobialmat. Appl. Environ.Microbiol. 71, 7352–7365.
Spencer, R.J., Lowenstein, T.K., 1988. Evaporites. In: McIlreath, I.A., Morrow, D.W. (Eds.),Diagenesis: Geosciences Canada Report Serial, vol. 4, pp. 141–163.
Stolz, J.F., 1990. Distribution of phototrophic microbes in the stratified microbialcommunity at Laguna Figueroa, Baja California, Mexico. BioSys. 23, 345–357.
Taberner, C., Marshall, J.D., Hendry, J.P., Pierre, C., Thirlwall, M.F., 2002. Celestiteformation, bacterial sulfate reduction and carbonate cementation of Eocene reefsand basinal sediments (Igualada, NE Spain). Sedimentology 49, 171–190.
Taylor, G.J., Blake, D., Gillis-Davis, J., Chipera, S.J., Bish,D.,Hammer, J., Lucey, P., Vaniman,D.T.,Sarrizin, P., 2008. X-ray diffraction in thefield and lab on themoon. Lunar and PlanetaryInstitute, Joint Ann. Meet. of LEAG-ICEUM-SRR, Abstr. # 4095.
Tekin, E., 2006. Elemental sulfur of bacterial origin in the lacustrine evaporites of thePolatli-Sivrihisar Neogene Basin (Central Anatolia, Turkey). Carb. Evap. 21, 33–39.
Tesson, Benoit, Gaillard, C., Martin-Jézéquel, V., 2008. Brucite formation mediated bythe diatom Phaeodactylum tricornutum. Mar. Chem. 109, 60–76.
Thompson, J.B., Ferris, F.G., 1990. Cyanobacterial precipitation of gypsum, calcite, andmagnesite from natural alkaline lake water. Geology 18, 995–998.
Vai, G.B., Lucchi, F.R., 1977. Algal crusts, autochthonous and clastic gypsum in acannibalistic evaporite basin: a case history from the Messinian of NorthernApennines. Sedimentology 24, 211–244.
Vasconcelos, C., McKenzie, J.A., 1997. Microbial mediation of modern dolomiteprecipitation and diagenesis under anoxic conditions (Lagoa Vermelha, Rio deJaneiro, Brazil). J. Sediment. Res. 67, 378–390.
Visscher, P.T., Reid, R.P., Bebout, B.M., 2000. Microscale observations of sulfate reduction:correlation of microbial activity with lithified micritic laminae in modern marinestromatolites. Geology 28, 919–922.
Vogel, M.B., Des Marais, D.J., Jahnke, L.L., Turk, K.A., Kubo, M.D., 2006. Organicgeochemistry of endoevaporitic environments: microbial diversity and lipidbiomarkers from gypsum deposits at the E.S.S.A. Salt Works, Guerrero Negro,Baja, Mexico. EOS Trans. Am. Geophys. Union 87 (52) Fall Meet. Suppl., B11D-07.
Vogel, M.B., Des Marais, D.J., Turk, K.A., Parenteau, M.N., Jahnke, L.L., Kubo, M.D.Y., 2009.The role of biofilms in actively forming gypsum deposits at Guerrero Negro, Mexico.Astrobio. 9, 875–893.
Warren, J.K., 2006. Evaporites: Sediments, Resources and Hydrocarbons. Springer-Verlag, Berlin.
Zopfi, J., Ferdelman, T.G., Fossing, H., 2004. Distribution and fate of sulfur intermediates—sulfur tetrathionate, thiosulfate, and elemental sulfur—in marine systems. In: Amend,J.P., Edwards, K.J., Lyons, T.W. (Eds.), Sulfur Biogeochemistry— Past and Present: Geol.Soc. of Amer. Special Paper, vol. 379, pp. 97–116.




















