
Effects of chlorate on the sulfation process of Trypanosoma cruzi glycoconjugates. Implication of...
13
Acta Tropica 137 (2014) 161–173 Contents lists available at ScienceDirect Acta Tropica jo ur nal home p age: www.elsevier.com/locate/actatropica Effects of chlorate on the sulfation process of Trypanosoma cruzi glycoconjugates. Implication of parasite sulfates in cellular invasion Maximiliano R. Ferrero a , Luciana L. Soprano a , Diana M. Acosta a , Gabriela A. García a , Mónica I. Esteva a , Alicia S. Couto b , Vilma G. Duschak a,∗ a Area de Bioquímica de Proteínas y Glicobiología de Parásitos, Departamento de Investigación, Instituto Nacional de Parasitologia, “Dr Mario Fatala Chaben”, ANLIS-Malbrán, Ministerio de Salud de la Nación, Paseo Colòn 568, CABA (1063), Buenos Aires, Argentina b CIHIDECAR, Departamento de Química Orgánica, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Buenos Aires, Argentina a r t i c l e i n f o Article history: Received 2 March 2014 Received in revised form 17 May 2014 Accepted 20 May 2014 Available online 28 May 2014 Keywords: Trypanosoma cruzi Sulfation Invasion process Cruzipain Sulfoglycosphingolipids a b s t r a c t Sulfation, a post-translational modification which plays a key role in various biological processes, is inhibited by competition with chlorate. In Trypanosoma cruzi, the agent of Chagas’ disease, sulfated structures have been described as part of glycolipids and we have reported sulfated high-mannose type oligosaccharides in the C-T domain of the cruzipain (Cz) glycoprotein. However, sulfation pathways have not been described yet in this parasite. Herein, we studied the effect of chlorate treatment on T. cruzi with the aim to gain some knowledge about sulfation metabolism and the role of sulfated molecules in this parasite. In chlorate-treated epimastigotes, immunoblotting with anti-sulfates enriched Cz IgGs (AS- enriched IgGs) showed Cz undersulfation. Accordingly, a Cz mobility shift toward higher isoelectric points was observed in 2D-PAGE probed with anti-Cz antibodies. Ultrastructural membrane abnormalities and a significant decrease of dark lipid reservosomes were shown by electron microscopy and a significant decrease in sulfatide levels was confirmed by TLC/UV-MALDI-TOF-MS analysis. Altogether, these results suggest T. cruzi sulfation occurs via PAPS. Sulfated epitopes in trypomastigote and amastigote forms were evidenced using AS-enriched IgGs by immunoblotting. Their presence on trypomastigotes surface was demonstrated by flow cytometry and IF with Cz/dCz specific antibodies. Interestingly, the percentage of infected cardiac HL-1 cells decreased 40% when using chlorate-treated trypomastigotes, suggesting sul- fates are involved in the invasion process. The same effect was observed when cells were pre-incubated with dCz, dC-T or an anti-high mannose receptor (HMR) antibody, suggesting Cz sulfates and HMR are also involved in the infection process by T. cruzi. © 2014 Elsevier B.V. All rights reserved. 1. Introduction Chagas disease, caused by the parasitic protozoan Trypanosoma cruzi represents a major health problem in Latin America. The World Health Organization estimates that 8–15 million persons currently have T. cruzi infection in 18 endemic countries in Cen- tral and South America and approximately 50 million people are at risk (WHO, 2010). The disease has also emerged as a public health problem elsewhere in the world due to infected people migrating to other regions of the world (Pérez-Molina et al., 2012). The most relevant clinical manifestation is chronic chagasic cardiomyopathy, causing the death by heart failure (Moolani et al., 2012). The biolog- ical cycle of T. cruzi involves vertebrate and invertebrate hosts. In ∗ Corresponding author. Tel.: +54 11 4331 4016; fax: +54 1143317142. E-mail addresses: [email protected], [email protected] (V.G. Duschak). mammals, the replicative form of the parasite behaves as an obli- gate intracellular pathogen that displays multiple mechanisms to manipulate a variety of host cell processes. T. cruzi attaches and invades a large variety of mammalian cells, including cardiomyocytes, by receptor-mediated interac- tions. Although the nature of the ligand/receptor components in both T. cruzi and target cells has not been completely elucidated, several lines of evidence indicate that interactions with oligosac- charides are involved (Meirelles et al., 1999; Barbosa and Meirelles, 1992; Bonay and Fresno, 1995; Araújo-Jorge and De Souza, 1998; Calvet et al., 2012). It has been demonstrated that cellular high mannose receptor (HMR) is involved in T. cruzi invasion and sur- vival (Soeiro et al., 1999; Garrido et al., 2011) and it has been proposed that its interaction with cruzipain (Cz), one of the major cysteine proteinase (CP) of T. cruzi, increases receptor recycling (Garrido et al., 2011). HMR is a multi-domain protein composed of a cysteine-rich domain with lectin activity that binds to sulfated http://dx.doi.org/10.1016/j.actatropica.2014.05.014 0001-706X/© 2014 Elsevier B.V. All rights reserved.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Effects of chlorate on the sulfation process of Trypanosoma cruzi glycoconjugates. Implication of...

Eg
MMa
Cb
a
ARRAA
KTSICS
1
cWctrptrci
h0
Acta Tropica 137 (2014) 161–173
Contents lists available at ScienceDirect
Acta Tropica
jo ur nal home p age: www.elsev ier .com/ locate /ac ta t ropica
ffects of chlorate on the sulfation process of Trypanosoma cruzilycoconjugates. Implication of parasite sulfates in cellular invasion
aximiliano R. Ferreroa, Luciana L. Sopranoa, Diana M. Acostaa, Gabriela A. Garcíaa,ónica I. Estevaa, Alicia S. Coutob, Vilma G. Duschaka,∗
Area de Bioquímica de Proteínas y Glicobiología de Parásitos, Departamento de Investigación, Instituto Nacional de Parasitologia, “Dr Mario Fatalahaben”, ANLIS-Malbrán, Ministerio de Salud de la Nación, Paseo Colòn 568, CABA (1063), Buenos Aires, ArgentinaCIHIDECAR, Departamento de Química Orgánica, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Buenos Aires, Argentina
r t i c l e i n f o
rticle history:eceived 2 March 2014eceived in revised form 17 May 2014ccepted 20 May 2014vailable online 28 May 2014
eywords:rypanosoma cruziulfationnvasion processruzipainulfoglycosphingolipids
a b s t r a c t
Sulfation, a post-translational modification which plays a key role in various biological processes, isinhibited by competition with chlorate. In Trypanosoma cruzi, the agent of Chagas’ disease, sulfatedstructures have been described as part of glycolipids and we have reported sulfated high-mannose typeoligosaccharides in the C-T domain of the cruzipain (Cz) glycoprotein. However, sulfation pathways havenot been described yet in this parasite. Herein, we studied the effect of chlorate treatment on T. cruziwith the aim to gain some knowledge about sulfation metabolism and the role of sulfated molecules inthis parasite. In chlorate-treated epimastigotes, immunoblotting with anti-sulfates enriched Cz IgGs (AS-enriched IgGs) showed Cz undersulfation. Accordingly, a Cz mobility shift toward higher isoelectric pointswas observed in 2D-PAGE probed with anti-Cz antibodies. Ultrastructural membrane abnormalities anda significant decrease of dark lipid reservosomes were shown by electron microscopy and a significantdecrease in sulfatide levels was confirmed by TLC/UV-MALDI-TOF-MS analysis. Altogether, these resultssuggest T. cruzi sulfation occurs via PAPS. Sulfated epitopes in trypomastigote and amastigote forms wereevidenced using AS-enriched IgGs by immunoblotting. Their presence on trypomastigotes surface was
demonstrated by flow cytometry and IF with Cz/dCz specific antibodies. Interestingly, the percentage ofinfected cardiac HL-1 cells decreased 40% when using chlorate-treated trypomastigotes, suggesting sul-fates are involved in the invasion process. The same effect was observed when cells were pre-incubatedwith dCz, dC-T or an anti-high mannose receptor (HMR) antibody, suggesting Cz sulfates and HMR arealso involved in the infection process by T. cruzi.© 2014 Elsevier B.V. All rights reserved.
. Introduction
Chagas disease, caused by the parasitic protozoan Trypanosomaruzi represents a major health problem in Latin America. The
orld Health Organization estimates that 8–15 million personsurrently have T. cruzi infection in 18 endemic countries in Cen-ral and South America and approximately 50 million people are atisk (WHO, 2010). The disease has also emerged as a public healthroblem elsewhere in the world due to infected people migratingo other regions of the world (Pérez-Molina et al., 2012). The most
elevant clinical manifestation is chronic chagasic cardiomyopathy,ausing the death by heart failure (Moolani et al., 2012). The biolog-cal cycle of T. cruzi involves vertebrate and invertebrate hosts. In∗ Corresponding author. Tel.: +54 11 4331 4016; fax: +54 1143317142.E-mail addresses: [email protected], [email protected] (V.G. Duschak).
ttp://dx.doi.org/10.1016/j.actatropica.2014.05.014001-706X/© 2014 Elsevier B.V. All rights reserved.
mammals, the replicative form of the parasite behaves as an obli-gate intracellular pathogen that displays multiple mechanisms tomanipulate a variety of host cell processes.
T. cruzi attaches and invades a large variety of mammaliancells, including cardiomyocytes, by receptor-mediated interac-tions. Although the nature of the ligand/receptor components inboth T. cruzi and target cells has not been completely elucidated,several lines of evidence indicate that interactions with oligosac-charides are involved (Meirelles et al., 1999; Barbosa and Meirelles,1992; Bonay and Fresno, 1995; Araújo-Jorge and De Souza, 1998;Calvet et al., 2012). It has been demonstrated that cellular highmannose receptor (HMR) is involved in T. cruzi invasion and sur-vival (Soeiro et al., 1999; Garrido et al., 2011) and it has been
proposed that its interaction with cruzipain (Cz), one of the majorcysteine proteinase (CP) of T. cruzi, increases receptor recycling(Garrido et al., 2011). HMR is a multi-domain protein composed ofa cysteine-rich domain with lectin activity that binds to sulfated
1 Tropi
scd
h(sbdTmYcTlnMtafs(
ooparOdaesg2taha(
rgomTappssdp
itktsostiemt
62 M.R. Ferrero et al. / Acta
ugars, a fibronectin type-II domain that mediates binding toollagen and eight C-type-lectin-like or carbohydrate-recognitionomains (CRD) (Martinez-Pomares et al., 2006).
Cz is a glycoprotein found in all stages of the parasite whichas been purified from epimastigotes as a complex mix of isoformsCazzulo et al., 2001). Although the bulk of the enzyme is lysosomal,everal observations indicate the presence of Cz-like membrane-ound isoforms located at the cell surface in the three mainevelopmental stages of the parasite (Souto-Padron et al., 1990;omas et al., 1997; Parussini et al., 1998) and the release to theedium of Cz-like CPs by metacyclic trypomastigotes (Yokoyama-
asunaka et al., 1994; Duschak et al., 2006). The mature enzymeonsists of the catalytic domain and a C-terminal (C-T) extension.he C-T domain contains high mannose- and complex-type-N-inked glycans (Parodi et al., 1995) and showed to be antigenic inatural and experimental infections (Scharfstein et al., 1983, 1985;artinez et al., 1991; Cazzulo et al., 2001). It is worth mentioning
hat Cz has been extensively studied not only as proteinase, antigennd glycoprotein (Duschak and Couto, 2009) but also as candidateor vaccine development (Cazorla et al., 2009) and it was also con-idered a very promising target for the chemotherapy of the diseaseDuschak and Couto, 2010; Duschak, 2011).
We have reported the presence of sulfated high mannose typeligosaccharides in the N-glycosylation site of the C-T domainf Cz (Barboza et al., 2005), constituting the first report on theresence of a sulfated glycoprotein in trypanosomatids. We havelso demonstrated that these sulfate groups elicit specific immuneesponses and are involved in heart damage (Acosta et al., 2008).ur results in conjunction with those reported in Dictyosteliumiscoydeum are, as far as we know, the only ones describing thetypical antigenic properties of this type of structures (Freezet al., 1984; Acosta et al., 2008). Furthermore, in T. cruzi, sulfatedtructures have been described as part of glycolipids such as sulfo-lycosphingolipids (or sulfatides) (Uhrig et al., 1992; Acosta et al.,012). UV-MALDI-TOF-MS analysis has shown that these struc-ures are mainly composed of sphingosine as the long chain basecylated with estearic acid (Acosta et al., 2012). Inhibition ELISAas demonstrated cross-reactivity between the C-T domain of Cznd sulfatides which was attributed to common sulfated epitopesAcosta et al., 2012).
Sulfation, widely observed from bacteria to humans, plays a keyole in various biological processes such as cell communication,rowth, and development (Kovensky, 2009) and sulfated N-linkedligosaccharides have been mainly implicated in several specificolecular recognition processes (Kawasaki et al., 2000; Honke and
aniguchi, 2002). Sulfation reaction occurs via the transfer of anctivated sulfate group from the donor, 3′-phosphoadenosine 5′-hosphosulfate (PAPS) (Klaassen and Boles, 1997) to a specificosition on a variety of acceptor sugar residues on numerousubstrates (Fukuda et al., 2001; Esko and Lindahl, 2001). ATP-ulfurylase, a key enzyme in the synthesis of the high energy sulfateonor PAPS is known to be inhibited by chlorate treatment (Sup-lementary data, Fig. 1) (Baeuerle and Huttner, 1986).
As we mentioned above, sulfated glycoconjugates were foundn T. cruzi, however, none of the enzymes involved in the sulfa-ion pathway has been described so far. With the aim to gain somenowledge about this process and the role of sulfated molecules inhis parasite, here we studied the effect of chlorate treatment onulfated epitopes, including those present in Cz and sulfatides, andn the infection of cardiac cells. Epimastigotes treated with chloratehowed Cz undersulfation, ultrastructural membrane abnormali-ies and a decrease of dark lipid reservosomes and sulfatide levels,
ndicating sulfation may occur via PAPS. On the other hand, sulfatedpitopes were demonstrated in both forms of T. cruzi present in theammalian host, trypomastigote and amastigote, by immunoblot-ing and interestingly, chlorate-treated trypomastigotes showed
ca 137 (2014) 161–173
less capacity to infect cardiac cells, suggesting sulfates are involvedin the invasion process.
2. Methods
2.1. Parasites
Tulahuen (stock Tul2), Brazil and CL strains of T. cruzi were used.Epimastigotes were grown in axenic Brain Heart Tryptose (BHT)medium (Cazzulo et al., 1985). Trypomastigote and amastigoteforms of the parasite were maintained in Vero cell monolayers. Try-pomastigotes were obtained from the culture medium of infectedVero cells free of cellular debris by leaving them to swim off the cen-trifuged pellet for 1 h at 37 ◦C and then collected by centrifugationfor 10 min at 2300 rpm in a Sorvall SS34 rotor. Amastigotes wereobtained from infected Vero cells treated with RPMI 5% FCS and0.06% SDS for 30 s. The resulting suspension was centrifuged twice,first at 800 rpm to discard cellular debris and then at 3200 rpm tocollect the amastigotes. Both steps were performed for 10 min inan Eppendorf 5804R centrifuge (Andrews and Colli, 1982).
2.2. Purification of Cz and C-T and desulfation treatment
Cz was purified to homogeneity from epimastigotes of Tulahuenstrain by affinity chromatography on Concanavalin A-Sepharosefollowed by Mono Q chromatography step (Acosta et al., 2008).The C-T domain was obtained by self-proteolysis of highly purifiedCz active preparations and purified by gel filtration in a Biogel P-30 column as previously described (Barboza et al., 2003). Chemicaldesulfation of Cz and C-T was achieved with the addition of pyridine(0.015 ml) over the lyophilized sample, which was then dissolvedin dimethylsulfoxide: methanol (9:1 (v/v), 0.2 ml), adjusted to pH4 with diluted HCl, heated at 100 ◦C for 2 h and freeze dried (Freezeet al., 1983). Purification was followed by 10% SDS-PAGE stainedwith silver nitrate. Protein content was measured by Bradford’smethod.
2.3. Obtainment of sera and purification of IgGs
Rabbits (New Zealand white lineage) were immunized withpurified Cz and dCz extracted from polyacrylamide gels or puri-fied CT and dC-T (three doses of 50 �g of protein/dose/rabbit byintradermal injection). Experimental procedures were conductedin the INP, Dr Mario Fatala Chaben, Ministerio de Salud, Argentina,in accordance with the ethical legislations and regulatory enti-ties established in Argentina and International Guides for careand use of laboratory animals. Precipitation of antibodies fromrabbit sera was done with ammonium sulfate and, afterwards,preimmune, anti-Cz, anti-dCz, anti-C-T and anti-dC-T IgGs werepurified by passing over a ProteinA-Sepharose column and elutingthe adsorbed IgG with 0.1 M-citrate buffer at pH 3.5 according toEy et al. (1978).
2.4. Preparation of the AS-enriched IgG fraction and evaluationby ELISA
Adsorption protocol with dCz was performed as previouslydescribed (Acosta et al., 2011). Briefly, 3.34 �g of dCz in 10 �l of PBSwere adsorbed onto a nitrocellulose membrane and next blockedwith PBS-3% skimmed milk. After blocking, anti-Cz IgGs (3.34 �gin 100 �l of PBS-1% skimmed milk) were incubated with the mem-brane for 1 h at 37 ◦C. Anti-Cz IgGs were incubated sequentially with
two nitrocellulose-containing dCz.An ELISA was performed as follows in order to evaluate ifadsorption with dCz led to enrichment in anti-sulfates IgGs: flat-bottomed plates were coated overnight at 4 ◦C with 0.3 �g per well

Tropi
omwepdpwFdda
2
taaschcd
2
t(jmwsw1rDrtdF
2
(iBBbstctw
2
a(npt
M.R. Ferrero et al. / Acta
f Cz or dCz. Both proteins were previously cracked in 0.15 M 2-ercaptoetanol. After blocking with PBS-3% skimmed milk, platesere incubated with a 1/4000 final dilution of anti-Cz IgGs or AS-
nriched IgGs for 1 h at RT. After washing with PBS-0.01% Tween,lates were incubated with mouse anti-rabbit IgG HRP (Jackson)iluted 1/2000 for 40 min at RT. Color was developed with o-henylenediamine dihydrochloride (OPD) and optical density (OD)as read at 490 nm using an ELISA microplate reader (Dynatech).
or competition ELISA, 50 �l of both IgGs, anti-Cz and AS-enriched,iluted 1/1000 in PBS-1% skimmed milk were pre-incubated withifferent amounts of GlnNAc6S in a final volume of 200 �l for 2 ht 37 ◦C in the presence of 0.1 M 2-mercaptoethanol.
.5. Parasite treatment with chlorate
Epimastigotes were grown in 15 ml tubes with an initial concen-ration of 2 × 106 parasites/ml (final volume: 4 ml) with differentmounts of sodium chlorate (Sigma–Aldrich) (0, 10, 25, 50 mM)nd collected after 24, 48 and 72 h. Trypomastigotes of Tulahuéntrain were grown in Vero cells with different concentrations ofhlorate (0, 10, 25) for 96 h. After washing, trypomastigotes werearvested, counted in a Neubauer chamber and used to infect HL1ells as described below. In all cases, culture medium was renewedaily to maintain chlorate concentration in each sample.
.6. Western blot analysis
Epimastigotes (2 × 105/lane), with or without chloratereatment, amastigotes (1 × 106/lane) and trypomastigotes2 × 106/lane) lysates and purified Cz (1 �g) as control were sub-ected to 10% SDS-PAGE and electrotransferred to nitrocellulose
embranes for 2 h at 200 mA. After blotting, nitrocellulose sheetsere coated with tris-buffered saline solution containing 3%
kimmed milk (TBS-M). The transferred proteins were incubatedith AS-enriched IgGs or anti-Cz IgGs (diluted 1/500 in TBS-M) for
h at RT and afterwards, incubated with peroxidase mouse anti-abbit IgG HRP (Jackson) (diluted 1/2000 in TBS-M) for 1 h at RT.etection was performed with the Enhanced chemiluminescence
eagent (ECL) (Amersham Biosciences, Cambridge, UK). Quanti-ation of luminic intensity was performed with a FujiLAS1000ensitometer equipped with IMAGE GAUGE 3.122 software (Fujiilm, Tokyo, Japan).
.7. Two-dimensional (2D) SDS-PAGE
Epimastigotes (2 × 105), with or without chlorate treatment, Cz1 �g) or Cz + dCz (1 �g each) were resuspended in isoelectric focus-ng (IEF) sample buffer and were separated by IEF a 4–1 ratio ofio-Lyte 5/7 and Bio-Lyte 3/10 (Bio-Rad). The tubes containing theio-Lytes were pre-electrophoresed at 200 V for 10 min followedy 300 V for 15 min and 400 V for 15 min more. After loading theample the electrophoresis was performed at 500 V for 10 min andhen 1000 V for 3.5 h. The second dimension separation was thenonducted in 10% SDS-PAGE reducing gels. Proteins were electro-ransferred to nitrocellulose membranes and western blot assayas performed as described in the above section.
.8. Isolation and purification of glycosphingolipids
Pellets of epimastigotes were extracted and fractionated bynionic exchange chromatography on a DEAE-Sephadex A-25
acetate form) column, which was differentially eluted to recovereutral GSLs and zwitterionic lipids as well as anionic lipids asreviously described (Acosta et al., 2011). The purified acidic frac-ion was analyzed by TLC in n-propanol/NH3/water (75:5:5, v/v/v).ca 137 (2014) 161–173 163
Spots were developed with I2. Spots corresponding to the sul-foglycolipids were scrapped from the plate and extracted withchloroform/methanol/water (10:10:1, v/v/v) to be subjected toUV-MALDI-TOF-MS analysis (Acosta et al., 2012) The analysis ofsulfatides undersulfation was evaluated by TLC after extraction andpurification of AGSLs from parasites treated with 10, 25 and 50 mMchlorate for 24 h.
2.9. Electron microscopy analysis
Parasite cells were fixed and processed for electron microscopyanalysis. Thin sections were sliced in a Porter-Blum MT2 Sorvallultramicrotome and the sections picked up in 300-mesh single holdgrids. Staining was done first with uranyl acetate and then withReynolds lead citrate stain. The samples were examined under aSiemens Zeiss C10 (Siemens, Germany) electron microscope. Pho-tographs were taken with a Kodak electron imaging film (EastmanKodak, Rochester, NY, USA). The number of dark or electronlucentreservosomes was counted per cell (200 cells were analyzed).
2.10. Surface labeling of parasites
For flow cytometry assays, live trypomastigotes of Tulahuenstrain stock Tul2 were washed with FACS buffer (0.1% FCS in PBS)and then incubated in a final volume of 100 �l with 10 �g of IgGpurified from rabbit pre-immune serum, anti-Cz IgGs or anti-dCzIgGs for 60 min at RT. Another washing step with FACS buffer wasdone before the incubation with a 1/100 dilution of a FITC-labeledgoat anti-rabbit-IgG antibody. Following 40 min of incubation, sam-ples were washed twice with FACS buffer and fixed with 2% PFA.A negative control sample incubated with the secondary antibodyalone was included. Samples were acquired on a FACSCalibur (Bec-ton Dickinson), and data were analyzed with FlowJo software.
Epimastigote and trypomastigote forms of Tulahén strain ofT. cruzi were stained by IF. Parasites were incubated with anti-Cz, anti-C-T, anti-dCz and anti-dC-T IgGs diluted 1/500 for 30 minat 37 ◦C. After washing, parasites were incubated with FITC-conjugated anti-rabbit IgG (Dako A/S, Denmark) diluted 1/40 for1 h at 37 ◦C. Slides were mounted in buffered glycerin and visual-ized by fluorescence microscopy (Camargo, 1966), and processedas described in the section: Amastigote staining for IF.
2.11. Infection assays of HL-1 cells
HL-1 cells (Claycomb et al., 1998) were cultivated on T-25 flaskscoated with 0.02% gelatin (DIFCO) and 0.005 mg/ml fibronectin(Sigma), in Claycomb medium (SAFC biosciences) supplementedwith 10% fetal bovine serum (FBS; Sigma), 2 mM L-glutamine(Sigma), plus antibiotics and maintained at 37 ◦C in a 5% CO2 atmo-sphere. For HL-1 infection assays, 2 × 104 HL-1 cells were platedinto 24-well culture dishes containing glass cover-slips (12 mmdiameter) coated with gelatin and fibronectin 24 h prior to theinfection. Control or chlorate-treated trypomastigotes (2 × 105 perwell) were added to the cells and after 4 h, parasites were removedand cells were washed twice with fresh medium. In some exper-iments, prior to and during the infection step, HL-1 cells werepre-incubated for 2 h at 37 ◦C with either 20 �g/ml of E-64 inactiv-ated Cz, dCz, C-T, dC-T or a rabbit monoclonal anti-HMR antibody(5 �g/ml) (CD-206-H 300 Santa Cruz Biotechnology Inc, CA, USA).Exclusively for this assay, in order to conserve the 3 dimensionalstructure of Cz, enzymatic desulfation of purified and active Cz was
performed with sulfatase VII from A. entrailis (Sigma–Aldrich) aspreviously described (Freeze et al., 1983). Infection was allowedto proceed for 3 days. Glass cover-slips were removed and cellswere fixed and then stained by May Grünwald Giemsa technique.
1 Tropi
Ai
fiims
2
c45fiiTaae1rTpEal
2
ES(loGmTaBdwpd
3
3
swpaaCcaIa2oT
64 M.R. Ferrero et al. / Acta
lternatively, amastigotes were stained by IF (see below). All exper-ments were performed in triplicates.
The percentage of infected cells was assessed in 80 microscopicelds per sample. Infectivity was calculated as 100 × number of
nfected cell in samples with treatment (chlorate-treated trypo-astigotes or pre-treated cells)/number of infected cells in control
amples (non-treated trypomastigotes or cells).
.12. Amastigote staining for IF
Three days after infection, HL-1 infected cells grown onto glassover-slips were fixed with 2% paraformaldehyde in PBS ON at◦C and then permeabilized with 0.1% Triton-X-100 in PBS for
min. After blocking covers in 1% BSA in PBS for 90 min at RT,xed cells were incubated ON at 4 ◦C with a serum from a chron-
cally infected mouse with Tulahuen strain of T. cruzi (1/1000).he covers were washed three times for 15 min each with PBSnd then incubated with goat anti-mouse IgG Alexa Fluor 594ntibody (1/1000) (Invitrogen) for 1 h at RT. After washes, cov-rs were stained with ethidium bromide (0.66 �g/ml in PBS) for0 min and were mounted over slides with 5 �l of Fluoromounteagent (Diagnostic Biosystems Inc, CA, USA) between both glasses.he slides were conserved at −20 ◦C in the dark. The assays wereerformed three times by triplicate. Cells were observed in a Nikonclipse E200 fluorescence microscope. Serial images (200×) werecquired with a Nikon Coldpix S10 camera covering all the mono-ayer, processed by blind deconvolution using the Image J software.
.13. Statistical analysis
Results are presented as the mean plus standard deviation (SD).ach experiment was performed independently three to four times.tatistical analysis was generally performed using PRISM softwareGraphPad Software, San Diego, CA, USA). One-way ANOVA fol-owed by Tukey’s test was used to compare: 1 – the RI of Cz bandbtained by western blot, 2 – the densitometric quantitation ofSLs spots in TLC, 3 – the number of reservosomes in electronicroscopy pictures at different chlorate concentrations and 4 –
. cruzi infectivity in HL-1 cells, except in those experiments usingnti-HMR antibodies in which T-test was used. (Two-Way ANOVA,onferroni post test was used for ELISA in preadsorption proce-ure with dCz). Fluorescence intensities of parasites confrontedith IgGs against Cz and its derivatives were analyzed by non-arametric Kruskal–Wallis test followed by multiple comparisonata of Dunns.
. Results
.1. Enrichment in antibodies specific for sulfated epitopes
With the aim to have a tool to study the effect of chlorate onulfated epitopes in T. cruzi, purified anti-Cz IgGs were adsorbedith desulfated Cz (dCz). An ELISA using Cz and dCz as antigens waserformed in order to evaluate whether the IgG fraction obtainedfter adsorption was enriched in anti-sulfated epitopes. Although,s expected, Cz was better recognized than dCz for both IgGs, anti-z and anti-Cz adsorbed with dCz, the Cz/dCz recognition ratio (R)hanged from 1.31 to 3.51 when dCz-adsorbed IgGs were used. Inddition, Cz recognition by dCz-adsorbed IgGs, but not by anti-CzgGs, decreased significantly when growing amounts of GlcNAc6S,
synthetic sugar which mimics the sulfated Cz epitope (Couto et al.,012), were added (Fig. 1C) and therefore, a decrease in R value wasnly observed when dCz-adsorbed IgGs were used (Fig. 1A and B).hese results indicate that the anti-Cz IgG fraction adsorbed with
ca 137 (2014) 161–173
dCz is enriched in anti-sulfates (AS) antibodies, so it was calledAS-enriched IgG.
3.2. Undersulfation of Cz by chlorate treatment
The inhibitory effect of chlorate on Cz sulfation was evalu-ated in T. cruzi epimastigote forms. With this aim, epimastigoteswere treated with increasing concentrations of chlorate (10, 25 and50 mM) and sulfated epitopes on Cz were analyzed by western blotprobed with the AS-enriched IgG. It is worth mentioning that chlo-rate had effect neither on parasite growth nor on protein synthesis,as shown by SDS-PAGE profile (Supplementary data, Fig. 2; Fig. 3;Fig. 4). A gradual decrease in recognition of Cz sulfated epitopeswas observed at growing amounts of chlorate after 48 h of treat-ment (Fig. 2IA). TcAKR, a cytosolic non-sulfated protein (Garavagliaet al., 2010), was run in parallel and used as a loading control. Czundersulfation was significant after treatment with 25 and 50 mMchlorate, showing a sulfation inhibition of 50 and 60% relative to thecontrol (0 mM chlorate) with p values of 0.05 and 0.01 respectively(Fig. 2IB). In addition, epimastigotes treated with 25 mM chloratewere subjected to two-dimensional gel electrophoresis and ana-lyzed by western blot with anti-Cz IgGs. Cz (Fig. 2IIA), dCz (Fig. 2IIB)and parasite lysates without chlorate treatment (Fig. 2IIC) wereused as controls. Clearly, treatment with 25 mM chlorate produceda shift of Cz toward higher isoelectric points (Fig. 2IID) than thecontrol sample (Fig. 2IIC), in agreement with Cz undersulfation
3.3. Effects of chlorate on sulfation of glycosphingolipids (GSLs)
Epimastigotes were treated with increasing concentrations ofchlorate (10, 25 and 50 mM), acidic GSLs (AGSLs) were extracted,purified and further analyzed by thin layer chromatography (TLC).A gradual decrease of the spot B which migrates similar to thesulfatide standard (SM4s) was observed (Fig. 3A and B). Analy-sis by UV-MALDI-TOF mass spectrometry confirmed that spot Bcorresponds to sulfatidic structures similar to those recently char-acterized (Acosta et al., 2012) (Supplementary data, Fig. 5). Thescanned spot B values evidenced a significant decrease in thebiosynthetic levels of these AGSLs at growing chlorate concentra-tions (Fig. 3A and B). On the contrary, a significant increase in spotA, corresponding to neutral GSLs structures (NGSLs), was obtainedafter treatment with 25 and 50 mM chlorate (Fig. 3A and C). NGSLsstructures were also confirmed by UV-MALDI-TOF resulting coinci-dent with those previously described (de Lederkremer et al., 1985)(data not shown).
3.4. Morphological effects of chlorate treatment in epimastigotes
Morphological structure of epimastigotes treated with chlo-rate was analyzed by electron microscopy. While no alterationswere detected in the ultrastructural analysis of epimastigotestreated with 25–30 mM chlorate (data not shown), some abnor-malities were observed at 40 mM and several after 50 mM chloratetreatment when comparing with untreated parasites (Fig. 4Aa).Chlorate-treated parasites evidenced swollen mitochondria, alter-ations in reservosomes, membranes and Golgi complex and showedatypical containing-membrane structures such as compartmen-talized mielinic- and electrodense material-containing figures(Fig. 4Ab–f). Membrane alterations may be attributed to theincrease in neutral lipids as consequence of undersulfation of GSLsdue to chlorate treatment. Reservosomes are round organelles(average diameter of 400–600 nm) delimited by a membrane,
commonly localized at the posterior region of epimastigotes, andcomposed mainly of either an electrodense protein matrix (Rd) orelectronlucent lipidic inclusions (RI) (De Souza et al., 1978; Soareset al., 1987; Cunha-e-Silva et al., 2006). When these particles were
M.R. Ferrero et al. / Acta Tropica 137 (2014) 161–173 165
A: Anti-Cz IgGs
0.0 2.5 5.0 10.0 20.00.0
0.5
1.0
1.5
2.0
Cz dCz
R: 1,31 1,42 1,40 1,40 1,30
GlcNAc6S (��M)
OD
49
0 n
m
C: Cz recog nition inh ibition
0 5 10 15 200
10
20
30
40
Anti-Cz IgGs
AS-enriched-IgGs**
*
*
GlcNAc6S (��M)
Inh
ibit
ion
(%
)
B: AS-enriched-IgGs
0.0 2.5 5.0 10.0 20.00.0
0.1
0.2
0.3
Cz dCz
R: 3,51 3,38 3,18 2,55 2,09
GlcNAc6S (��M)
DO
49
0 n
m
Fig. 1. Enrichment of IgG antibodies specific for sulfates by adsorption of anti-Cz IgGs with dCz. ELISA results showing the recognition of Cz (�) and dCz (�) as antigens bya c6S. Ro entagd hibit
ctc2ebbhicisc
3d
dbofia
nti-Cz IgGs (A) or AS-enriched IgGs (B) competing with growing amounts of GlnNAbtained with dCz as antigens for each GlnNAc6S concentration. (C) Inhibition percata obtained from A and B graphics. The absence of GlnNAc6S corresponds to 0% in
ounted in control and chlorate-treated parasites, it was observedhat in parasites treated with chlorate the number of Rd particles,orresponding to newly forming reservosomes (Sant’Anna et al.,009), decreased (Fig. 4B). As new reservosomes are acidic, verylectrodense and poor in lipidic inclusions, the decrease in the num-er of Rd particles observed in treated parasites could be explainedy the diminution in the AGSLs induced by chlorate. On the otherand, the number of RI particles increased with chlorate treatment
n a dose dependent way (Fig. 4C). RI particles in treated parasitesorresponded to reservosomes filled with lipidic electronlucentnclusions or refringent particles of varied aspects, some round,ome looking like planar bilayers, others with lipid inclusions ofrystalloid aspect; suggesting accumulation of neutral lipids.
.5. Immune recognition of sulfated epitopes in the threeevelopmental stages of T. cruzi
In order to evaluate the presence of sulfated epitopes in differentevelopmental forms of T. cruzi other than epimastigotes, westernlot assays of epimastigote, amastigote and trypomastigote lysates
f three T. cruzi strains (Tulahuén, CL Brener and Brazil) were con-ronted with anti-Cz IgGs and AS-enriched IgGs (Fig. 5I). Differencesn the immune recognition of Cz and sulfated epitopes patternsmong different stages and strains of the parasite were evidencedis the recognition ratio calculated as the OD value obtained with Cz/the OD valuee of Cz recognition by anti-Cz IgGs (�) and AS-enriched-IgGs (�) achieved with theion. *p < 0.05.
by both pools of IgGs. The presence of immunoreactive bands withthe AS-enriched IgGs in all T. cruzi strains and in all of the parasiteforms indicates that sulfated epitopes are ubiquitous. In these con-ditions the sulfated epitopes appeared to be present in different Czisoforms. Protein isoforms with similar apparent molecular weightto control Cz were revealed with anti-Cz IgGs. However, when AS-enriched IgGs were probed on trypomastigote lysates, the main Czband was not observed, indicating the lack of sulfated epitopes inthis Cz isoform. In addition, the presence of sulfated Cz isoforms oflower and higher apparent molecular weight than the major bandwere observed” (Duschak et al., 2006). In addition to Cz recognition,other molecules were observed which may correspond to either Cz-like isoforms or cross-reactive sulfated epitopes (Fig. 5I). In orderto study the presence of sulfated epitopes on parasite surface, IFassays were performed in epimastigotes and trypomastigotes withanti-Cz, anti-dCz, anti-C-T, and anti-dC-T IgGs (Fig. 5II). Parasitesconfronted with anti-dCz and anti-dC-T IgGs recognized parasitesurface to a lesser extent than anti-Cz and anti-C-T ones, suggest-ing the presence of Cz with a sulfated C-T domain on the surfaceof both forms of T. cruzi (Fig. 5II, A and B). The presence of sul-
fated epitopes on trypomastigote surface was confirmed by flowcytometry using anti-Cz and anti-dCz IgGs. As expected, anti-Czshowed higher signals than anti-dCz IgGs (Fig. 5III, C and D). It isworth mentioning that even though we used anti-Cz and anti-dCz
166 M.R. Ferrero et al. / Acta Tropica 137 (2014) 161–173
Fig. 2. Cz undersulfation in epimastigotes treated with chlorate. I: Cz recognition by AS-enriched IgG in chlorate-treated epimastigotes. A: western blot analysis of epimastigotesamples probed with AS-enriched IgG. Cz and dCz lanes contain 1 �g of purified proteins. Equivalence in protein loading was controlled by immune-detection of TcAKR.B: Measurement of relative Cz levels at different concentrations of chlorate. Ratio: RI Cz band/RI TcAKR band. RI: relative intensity. The RI obtained without chlorate wastaken as 100%. * indicates a p value < 0.05 and ** indicates a p value < 0.01 both relative to 0 mM chlorate. II: Cz mobility in two-dimensional gel electrophoresis after chloratetreatment. A: Cz (1 �g); B: Cz, (1 �g) plus dCz (1 �g); C: 2 × 105 parasite without chlorate treatment and D: 2 × 105 parasite treated with 25 mM chlorate for 48 h. On the top,t e secow
Isbfobd
3
iiVtmswbc
he first dimension left to right is isoelectric focusing from pH range 3 to 10, and thith anti-Cz IgGs.
gGs and we conclude about the presence of sulfated Cz on para-ite surface, we cannot rule out that these results may either reflectinding to Cz isoforms that were secreted and adsorbed to the sur-ace of parasites or recognition of cross-reactive sulfated epitopesn unrelated surface antigens. In this regard, the cross-reactivityetween sulfates from surface Cz and sulfatides has been alreadyemonstrated (Acosta et al., 2012).
.6. Effect of sulfates in T. cruzi invasion of HL-1 cells
In order to study the participation of parasite sulfates in thenvasion process, chorate-treated trypomastigotes were used tonfect the cardiac cell line HL-1. Trypomastigotes were grown inERO cells in the presence of chlorate during 96 h and afterwards,
hey were used to infect HL-1 cells. While 10 mM chlorate treat-ent of trypomastigotes did not show effect on HL-1 infection, a
ignificant decrease in the number of infected cells was observedith 25 mM chlorate (Fig. 6), suggesting that parasite sulfates may
e relevant for infection. No significant differences in the per-entage of infected cells were observed with 25 mM and 30 mM
nd dimension (top to bottom) is 10% SDS-PAGE gels. Western blot was confronted
chlorate treatment (data not shown). Chlorate concentrationshigher than 40 mM were avoided to discard possible pleiotropiceffects since they induced some membrane alterations and ultra-structural abnormalities (Section 3.4). With the aim to betterunderstand the mechanisms used by T. cruzi sulfates in the inva-sion process, HL-1 cells were incubated with Cz, dCz, C-T or dC-Tprior to and during the infection and the percentage of infectedcells was analyzed. Treatment of HL-1 cells with Cz or C-T did notaffect the infectivity in comparison with untreated control cells(Fig. 7A1–A3, B and C). However, a significant decrease in infectionwas observed after incubation of cells with desulfated molecules,either dCz (Fig. 7A4 and B) or dC-T (Fig. 7A5 and C). The fact that sim-ilar results were obtained with either Cz or C-T and dCz or dC-T is inagreement with the presence of sulfates in the C-T domain of Cz as ithas been previously described (Barboza et al., 2005). As it has beendescribed that Cz can interact with cellular receptors, such as HMR
(Soeiro et al., 1999: Garrido et al., 2011) and, based on our results,we may hypothesize that sulfated molecules from T. cruzi, eithersoluble or on parasite surface, are necessary for parasite infec-tion since they may trigger cellular signals necessary for parasite
M.R. Ferrero et al. / Acta Tropica 137 (2014) 161–173 167
Fig. 3. Analysis of AGSLs in chlorate treated epimastigotes. (A) Epimastigote forms were treated with different concentrations of chlorate (0, 10, 25 and 50 mM) for 24 h.The AGSLs fractions obtained from 5 × 109 parasites, with and without chlorate treatment were analyzed by TLC. Ganglioside (GM1) and sulfatide (SM4s) standards useda uantitc M (**
iinufeiiHmtdlmHtr
s controls are shown with arrows. The bars graphs represent the densitometric qorresponding to spot B. Spot A: 0 mM vs 25 mM 50 mM (**p < 0.01); 25 mM vs 50 m
nternalization (Santos Barrias et al., 2013). On the other hand, tak-ng into account that HMR has ability to bind sulfated and alsoon-sulfated oligosaccharidic chains (Liu et al., 2001), we may spec-late that desulfated forms of Cz might either block the binding siteor Cz, probably due to steric hindrance, or HMR recycling (Garridot al., 2011), leading to cells less susceptible to be infected. Accord-ng to this hypothesis, a significant decrease in the percentage ofnfection (Fig. 7A6 and D) was observed after pre-incubation ofL1 cells with anti-HMR antibodies reactive to its non-sulfatedannose binding domains, suggesting HMR is involved in the infec-
ion process by T. cruzi. A participative role of sulfates from C-Tomain of Cz, as well as, non-sulfated Cz isoforms in T. cruzi cel-
ular invasion through this receptor is strongly feasible. It is worth
entioning that the presence of the mannose receptor over theL1 cell line was confirmed by immunofluorescence staining overhe cells with a rabbit specific monoclonal antibody against thiseceptor (Supplementary data, Fig. 6).
ation of the spot signals. (B) Bar graph corresponding to spot A and (C) Bar graph*p < 0.001). Spot B: 0 mM vs 10 mM, 25 mM and 50 mM (***p < 0.001).
4. Discussion
Sulfation is a post-translational modification which is criticalfor many biological events such as ligand-receptor recognition,receptor-mediated signaling, cellular adhesion and structuralmaintenance (Kovensky, 2009; Fukuda et al., 2001; Rapraegeret al., 1991). In T. cruzi, sulfated structures have been identi-fied as part of glycolipids (Freeze et al., 1984) and glycoproteins(Barboza et al., 2005), however nothing is known about themetabolic pathways involved in their synthesis. In previous works,we have determined the chemical composition of sulfatides (Acostaet al., 2012) and reported for the first time the presence of asulfated glycoprotein when we described the N-glycans present
in the C-T domain of the Cz (Barboza et al., 2005). Moreover,we demonstrated that sulfates are main targets of immuno-logical responses against Cz and are capable to induce tissuedamage. Immunization of BALB/c mice with C-T in the absence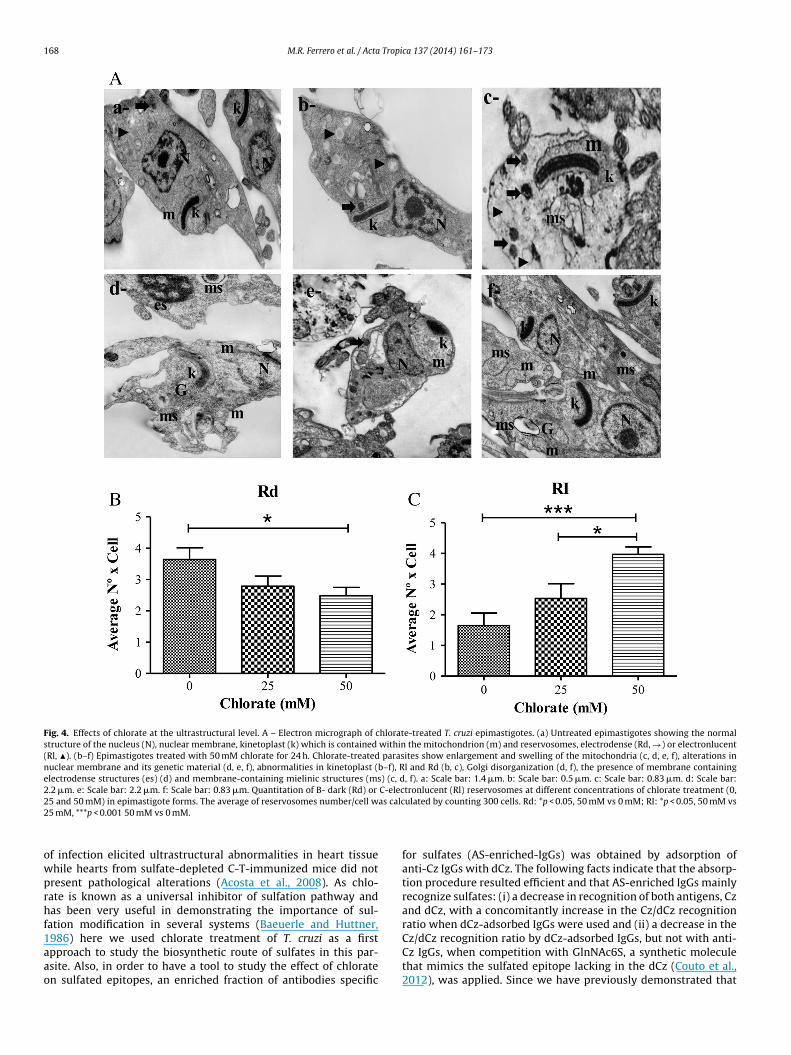
168 M.R. Ferrero et al. / Acta Tropica 137 (2014) 161–173
Fig. 4. Effects of chlorate at the ultrastructural level. A – Electron micrograph of chlorate-treated T. cruzi epimastigotes. (a) Untreated epimastigotes showing the normalstructure of the nucleus (N), nuclear membrane, kinetoplast (k) which is contained within the mitochondrion (m) and reservosomes, electrodense (Rd, →) or electronlucent(Rl, �). (b–f) Epimastigotes treated with 50 mM chlorate for 24 h. Chlorate-treated parasites show enlargement and swelling of the mitochondria (c, d, e, f), alterations innuclear membrane and its genetic material (d, e, f), abnormalities in kinetoplast (b–f), Rl and Rd (b, c), Golgi disorganization (d, f), the presence of membrane containingelectrodense structures (es) (d) and membrane-containing mielinic structures (ms) (c, d, f). a: Scale bar: 1.4 �m. b: Scale bar: 0.5 �m. c: Scale bar: 0.83 �m. d: Scale bar:2.2 �m. e: Scale bar: 2.2 �m. f: Scale bar: 0.83 �m. Quantitation of B- dark (Rd) or C-electronlucent (Rl) reservosomes at different concentrations of chlorate treatment (0,2 as calc2
owprhf1aao
5 and 50 mM) in epimastigote forms. The average of reservosomes number/cell w5 mM, ***p < 0.001 50 mM vs 0 mM.
f infection elicited ultrastructural abnormalities in heart tissuehile hearts from sulfate-depleted C-T-immunized mice did notresent pathological alterations (Acosta et al., 2008). As chlo-ate is known as a universal inhibitor of sulfation pathway andas been very useful in demonstrating the importance of sul-
ation modification in several systems (Baeuerle and Huttner,
986) here we used chlorate treatment of T. cruzi as a firstpproach to study the biosynthetic route of sulfates in this par-site. Also, in order to have a tool to study the effect of chloraten sulfated epitopes, an enriched fraction of antibodies specificulated by counting 300 cells. Rd: *p < 0.05, 50 mM vs 0 mM; RI: *p < 0.05, 50 mM vs
for sulfates (AS-enriched-IgGs) was obtained by adsorption ofanti-Cz IgGs with dCz. The following facts indicate that the absorp-tion procedure resulted efficient and that AS-enriched IgGs mainlyrecognize sulfates: (i) a decrease in recognition of both antigens, Czand dCz, with a concomitantly increase in the Cz/dCz recognitionratio when dCz-adsorbed IgGs were used and (ii) a decrease in the
Cz/dCz recognition ratio by dCz-adsorbed IgGs, but not with anti-Cz IgGs, when competition with GlnNAc6S, a synthetic moleculethat mimics the sulfated epitope lacking in the dCz (Couto et al.,2012), was applied. Since we have previously demonstrated that
M.R. Ferrero et al. / Acta Tropica 137 (2014) 161–173 169
Fig. 5. The presence of sulfate moieties in Cz-like isoforms in the three developmental forms from different strains of Trypanosoma cruzi. Immunerecognition of surfacesulfated epitopes. I: Western blot of the three developmental stages of T. cruzi: E, epimastigotes, A, amastigotes and T, trypomastigotes, with anti-Cz IgGs (a) and AS-enrichedIgGs (b) in three T. cruzi strains: Tulahuen (Tul), CL Brener (CL) and Brazil. Purified Cz (1 �g) was used as control. Purified Cz (1 �g) was used as control. II: IF assay fromfixed epimastigotes (A) and trypomastigotes (B) confronted with IgGs purified from rabbit pre-immune serum (a1, b1), anti-Cz IgGs (a2, b2), anti-dCz IgGs (a3, b3), anti-C-TIgGs (a4, b4) and anti-dC-T IgGs (a5, b5). Fluorescence was quantified (in arbitrary units) with ImajeJ 1.46. Magnification: 400×. *p < 0.05, **p < 0.01 and ***p < 0.001. III: Flowc ary anI s (D) f
ctep
ytometry analysis of Tulahuen strain trypomastigotes incubated with the secondgGs purified from rabbit pre-immune serum (B), anti-Cz IgGs (C), and anti-dCz IgG
ross-reaction exists between sulfates groups from sulfatides andhe C-T domain of Cz (Acosta et al., 2012), we may assume that AS-nriched IgGs recognize sulfated epitopes in general, not only thoseresent on Cz.
tibody alone (FITC-labeled goat anti-rabbit-IgG antibody) as negative control (A),ollowed by the corresponding secondary antibody.
Chlorate treatment of epimastigotes forms of T. cruzi showedseveral effects which can be attributed to alterations in themetabolism of sulfation. Cz undersulfation in treated parasites wasshown not only by western blot probed with AS-enriched IgGs but

170 M.R. Ferrero et al. / Acta Tropi
Chlorate-treated trypomastigotes
0 10 25
0
50
100
150
*
Chlorate (mM)
infe
ctiv
ity
(%
)
Fig. 6. Effects of chlorate on trypomastigote infection of cardiac cell line HL-1. 24 hHL-1 cells monolayers were infected with chlorate-treated trypomastigotes. At 3days post-infection, infected HL-1 cells were fixed and stained with May-GrungwaldGiemsa. The number of infected cells was counted and infectivity was calculatedas 100 × infected cell number with chlorate-treated trypomastigotes/infected cellnumber with non-treated trypomastigotes (0 mM chlorate). HL-1 cells infected withnv
aImfrab(toatpremiotbcmoprtits1erftwwtttidsdw
cruzi (Soeiro et al., 1999), contain two portions with distinct car-
on-treated trypomastigotes yielded 30% of infected cells. *p < 0.05, 25 mM chlorates 0 mM chlorate.
lso by bidimensional gel electrophoresis tested with anti-Cz IgGs.t is important to mention that the C-T domain from Cz was for-
erly detected by isoelectrofocusing showing pI values rangingrom 4.5 to 5.0 (Stoka et al., 1995; Cazzulo et al., 1995), the sameange we obtained here for Cz from non-treated epimastigotes. Inddition, a similar 2D pattern was obtained by others in radiola-eled precipitates probed with an anti-Cz monoclonal antibodyScharfstein et al., 1986). The shift of Cz toward a higher isoelec-ric point in chlorate-treated epimastigotes is in line with the lackf an acidic component of this protein. On the other hand, thenalysis of AGSLs biosynthesized by epimastigotes under chloratereatment evidenced a marked reduction of sulfatides (spot B) inarallel to a manifest increase of NGSLs. When the effect of chlo-ate on morphological structure of epimastigotes was analyzed bylectron microscopy, we observed drastic membrane alterations,itochondrial swelling, abnormal concentric membrane structures
nside some organelles, formation of myelin figures, and Golgi dis-rganization. While these ultrastructural changes could be relatedo the altered proportion of sulfatides and NGSLs and/or mem-rane alterations could be attributed to the increase in NGSLs asonsequence of undersulfation of GSLs induced by chlorate treat-ent, it is important to note that these alterations have also been
bserved in other studies using different drugs affecting lipidicathways (Menna-Barreto et al., 2009). Moreover, quantization ofeservosomes in chlorate-treated epimastigotes showed a reduc-ion in the number of electrodense reservosomes and an increasen the number of the electronlucent reservosomes. It is knownhat the matrix of reservosomes contains electronlucent inclu-ions of lipidic nature (Soares et al., 1987; Soares and De Souza,988) and that newly formed ones are acidic organelles (Sant’Annat al., 2009). Therefore, the reduction in number of electrodenseeservosomes could be explained as a result of the decrease in sul-atides induced by chlorate while the increase in the number ofhose electronlucent may reflect the accumulation of NGSLs. It isorth mentioning that membrane alterations were not observedhen parasites were treated with 25 mM chlorate, indicating that
he chlorate dosis needed for ultrastructure alterations are higherhat those required for producing effects on glycoconjugates sulfa-ion at molecular level. Altogether, the effect of chlorate treatmentn epimastigote forms strongly suggests the presence of a PAPS-ependent sulfation mechanism in T. cruzi. The existence of a
equence with homology to a PAPS reductase in the T. cruzi genomeatabase reinforces this hypothesis. However, no other sequencesith homology to enzymes involved in sulfation metabolism wereca 137 (2014) 161–173
found in this database, even alignments with conserved sulfotrans-ferases motifs showed no matching results, thus suggesting thatsignificant differences in sequences between T. cruzi and conven-tional sulfation enzymes may exist.
Western blot analysis using the AS-enriched IgGs showed thatsulfated epitopes are present in the three parasite stages of differentparasite strains demonstrating that these moieties are ubiquitous.Taking into account that these AS-enriched IgGs were obtainedfrom anti-Cz IgGs, we consider that they might mainly recog-nize Cz isoforms. However, for the reasons mentioned above, wecannot rule out that other sulfated proteins and/or sulfatides arerecognized by these antibodies. Furthermore, differences in theelectrophoretic pattern among all stages of T. cruzi suggest somestage-dependent regulation in the sulfation mechanism. On theother side, we have also demonstrated that sulfated epitopes areon the surface of epimastigotes and trypomastigotes. This is inaccordance with a previous report showing by immunoelectronmicroscopy the presence of cross-reactive material with Cz onthe surface of all the developmental stages of the parasite (Souto-Padron et al., 1990). We have to take into account that the presenceof sulfate moieties in parasite surface may either reflect binding toCz isoforms linked to the membrane or secreted and adsorbed to theparasite surface (Parussini et al., 1998; Yokoyama-Yasunaka et al.,1994; Duschak et al., 2006). The surface location of Cz isoforms is inagreement with the antigenic characteristics of the sulfated groupspresent in the C-T domain we have previously described (Barbozaet al., 2005; Acosta et al., 2008; Couto et al., 2012). Although sul-fated proteoglycans are involved in the cell recognition processin several human pathogens (Pancake et al., 1992; Ortega-Barriaand Boothroyd, 1999; Carruthers et al., 2002; Fleckenstein et al.,2002), including T. cruzi (Herrera et al., 1994; Oliveira et al., 2008;Bambino-Medeiros et al., 2011), no data are available regardinginvolvement of Cz as glycoprotein in the invasion process.
In this report, we show (i) the participation of parasite sul-fates in invasion of trypomastigotes to a cardiomyocyte cell line,since chlorate treatment of trypomastigotes lead to a decreasein infectivity, and (ii) the involvement of Cz sulfates in thisprocess, as the decrease was reproduced after pre-incubationof cells with dCz and dC-T but not with Cz and C-T. This resultmay indirectly suggest a participation of Cz-sulfated epitopes inpromoting the internalization of trypomastigotes by HL-1 cells,probably by allowing the receptor to trigger an intracellularsignaling (Yoshida and Cortez, 2008; Vázquez-Mendoza et al.,2013). In this sense, mannose receptor-pathogen interactions havebeen implicated in the initiation of signaling pathways leadingto the production of lysosomal enzymes, IL-12, among others(Lefkowitz et al., 1997; Shibata et al., 1997). On the other hand,we confirm that cellular HMR is involved in parasite infection(Soeiro et al., 1999; Vázquez-Mendoza et al., 2013). The blockageof the HL-1 HMR with monoclonal antibodies specific for twoout of eight carbohydrate-recognition domains (CRD) whichbind to non-sulfated molecules also resulted in a decrease of theinfection percentage. It was documented that Cz interacts withcellular HMR, activates intracellular signals, increases intracellularamastigotes growth and induces HMR recycling (Stempin et al.,2004, 2008; Garrido et al., 2011). In addition, all these effects canbe reproduced by mannose coupled to BSA (Garrido et al., 2011).Moreover, it was observed that pre-treatment of HMR with mon-oclonal antibodies reduces parasite intracellular growth (Garridoet al., 2011). Finally, taking into account (i) that sulfate-containingCz is a high-mannose-type glycoprotein, (ii) that HMR localized atthe surface of cardiac cells are involved in binding and uptake of T.
bohydrate recognition properties with the ability to bind sulfatedand non-sulfated oligosaccharidic chains (Liu et al., 2001) and areexpressed by antigen-presenting cells in non-lymphoid peripheral

M.R. Ferrero et al. / Acta Tropica 137 (2014) 161–173 171
Fig. 7. Effects of pre-treatments with Cz, Cz-derivatives and anti-HMR antibodies on the infection of HL-1 cells. (A) IF pictures of HL-1 infected cells which were treated 2 hprior to and during the infection with: C-T (2), Cz (3), dC-T (4), dCz (5) and anti-HMR (6). HL-1 cells without treatment were used as control (1). After 3 days post-infection,c ed by5 ty wasn
othHCwd
r
ells were fixed and stained by IF using a chronically-infected mouse serum follow0 �m. (B)–(D) Infectivity graphs obtained with the different treatments. Infectiviumber samples without treatment. *p < 0.05; **p < 0.01.
rgans counting cardiac muscle in mice (Linehan, 2005), and (iii)hat HL1 is a cardiac cell line (Claycomb et al., 1998), althoughere we do not demonstrate the interaction between Cz and theMR from HL1 cells, based on our results, we can speculate thatz may participate in the infection process through interaction
ith the HMR and, in particular, through the sulfates from the C-Tomain.In addition to the fact that sulfates mount a humoral specificesponse in Chagas disease patients (Acosta et al., 2008), that
a goat antimouse Alexa Fluor 594 antibody. Magnification: 200×. Scale bar size: calculated as 100 × infected cell number in samples with treatment/infected cell
sulfate moieties are common epitopes between Cz and sulfatidesand specific antibodies for sulfates inversely correlate with sever-ity of human Chagas disease (Acosta et al., 2012), our findingsconstitute a new step contributing to elucidate all the intrinsicaspects of T. cruzi-host cell interplay displayed during the estab-
lishment of the Chagas disease. Finally, these results suggest thatsurface sulfate groups of Cz due to their overall presence mightbe also involved in immunopathogenicity, immunomodulationmechanisms and in the progression of the disease.
1 Tropi
A
CsaCOHttt
A
i2
R
A
A
A
A
A
B
B
B
B
B
B
C
C
C
C
C
C
C
C
72 M.R. Ferrero et al. / Acta
cknowledgements
V.G.D., A.S.C. and G.A.G are Research Career Investigators fromONICET, Argentina. D.M.A., M.R.F. and LLS thank CONICET fellow-hips granted. We thank grants from CONICET (PIP-2008-1548)nd ANPCyT (PICT-2006-00145). We also thank to Dr. Williamlaycomb from Louisiana State University Medical Center, Newrleans, LA, USA for kindly providing to researchers from INP theL-1 cardiac cells, to Cristina Maidana, member of the Produc-
ion Department, INP Dr M. Fatala Chaben, ANLIS-Malbrán ando Margarita Lopez from LANAIS-MIE, for the electron microscopyechnical assistance.
ppendix A. Supplementary data
Supplementary data associated with this article can be found,n the online version, at http://dx.doi.org/10.1016/j.actatropica.014.05.014.
eferences
costa, D.M., Arnaiz, M.R., Esteva, M.I., Barboza, M., Stivale, D., Orlando, U.D., Torres,S., Laucella, S.A., Couto, A.S., Duschak, V.G., 2008. Sulfates are main targets ofimmune responses to cruzipain and are involved in heart damage in BALB/cimmunized mice. Int. Immunol. 20 (4), 461–470.
costa, D.M., Soprano, L.L., Ferrero, M., Landoni, M., Esteva, M.I., Couto, A.S., Duschak,V.G., 2011. A striking common O-linked N-acetylglucosaminyl moiety betweencruzipain and myosin. Parasite Immunol. 33 (7), 363–370.
costa, D.M., Soprano, L.L., Ferrero, M.R., Esteva, M.I., Riarte, A., Couto, A.S., Duschak,V.G., 2012. Structural and immunological characterization of sulfatides: rel-evance of sulfate moieties in Trypanosoma cruzi glycoconjugates. ParasiteImmunol. 34 (11), 499–510.
ndrews, N.W., Colli, W., 1982. Adhesion and interiorization of Trypanosoma cruziin mammalian cells. J. Protozool. 29 (2), 264–269.
raújo-Jorge, T.C., De Souza, W., 1998. Interaction of Trypanosoma cruzi withmacrophages. Involvement of surface galactose and N-acetyl-d-galactosamineresidues on the recognition process. Acta Trop. 45, 127–136.
arbosa, H.S., Meirelles, M.N., 1992. Ultrastructural detection in vitro of WGA-, RCAI-, and Con A-binding sites involved in the invasion of heart muscle cells byTrypanosoma cruzi. Parasitol. Res. 78 (5), 404–409.
arboza, M., Duschak, V.G., Cazzulo, J.J., de Lederkremer, R.M., Couto, A.S., 2003.Presence of sialic acid in N-linked oligosaccharide chains and O-linked N-acetylglucosamine in cruzipain, the major cysteine proteinase of Trypanosomacruzi. Mol. Biochem. Parasitol. 126, 293–296.
arboza, M., Duschak, V.G., Fukuyama, Y., Nonami, H., Erra-Balsells, R., Cazzulo, J.J.,Couto, A.S., 2005. Structural analysis of the N glycans present in cruzipain, themajor cysteine proteinase of Trypanosoma cruzi. Identification of sulfated highmannose type oligosaccharides. FEBS J. 272, 3803–3815.
aeuerle, P.A., Huttner, W.B., 1986. Chlorate. A potent inhibitor of protein sulfationin intact cells. Biochem. Biophys. Res. Commun. 141 (2), 870–877.
ambino-Medeiros, R., Oliveira, F.O., Calvet, C.M., Vicente, D., Toma, L., Krieger, M.A.,Meirelles, M.N., Pereira, M.C., 2011. Involvement of host cell heparan sulfateproteoglycan in Trypanosoma cruzi amastigote attachment and invasion. Para-sitology 138 (5), 593–601.
onay, P., Fresno, M., 1995. Characterization of carbohydrate binding proteins inTrypanosoma cruzi. J. Biol. Chem. 270 (19), 11062–11070.
alvet, C.M., Melo, T.G., Garzoni, L.R., Oliveira Jr., F.O., Neto, D.T., Meirelles, L., Pereira,M.C., 2012. Current understanding of the Trypanosoma cruzi-cardiomyocyteinteraction. Front. Immunol. 3 (327), 1–8.
amargo, M.E., 1966. Fluorescent antibody test for serodiagnosis of American Try-panosomiasis. Technical modification employing preserved culture forms ofTrypanosoma cruzi in a slide test. Rev. Inst. Med. Trop. Sao Paulo 8 (5), 227–234.
arruthers, V.B., Hakansson, S., Giddings, O.K., Sibley, L.D., 2002. Toxoplasma gondiiuses sulfated proteoglycans for substrate and host cell attachment. Infect.Immun. 68 (7), 4005–4011.
azorla, S.I., Frank, F.M., Malchiodi, E.L., 2009. Vaccination approaches against Try-panosoma cruzi infection. Expert Rev. Vacc. 8, 921–935.
azzulo, J.J., Franke de Cazzulo, B.M., Engel, J.C., Cannata, J.J., 1985. End productsand enzyme levels of aerobic glucose fermentation in trypanosomatids. Mol.Biochem. Parasitol. 16, 329–343.
azzulo, J.J., Labriola, C., Parussini, F., Duschak, V.G., Martinez, J., Franke de Cazzulo,B.M., 1995. Cysteine proteinases in Trypanosoma cruzi and other Trypanosomatidparasites. Acta Chim. Sloven. 42, 409–418.
azzulo, J.J., Stoka, V., Turk, V., 2001. Cruzipain, the major cysteine proteinase from
the protozoan parasite Trypanosoma cruzi. Curr. Pharm. Des. 7, 1143–1156.laycomb, W.C., Lanson Jr., N.A., Stallworth, B.S., Egeland, D.B., Delcarpio, J.B., Bahin-ski, A., Izzo Jr., N.J., 1998. HL-1 cells: a cardiac muscle cell line that contracts andretains phenotypic characteristics of the adult cardiomyocyte. Proc. Natl. Acad.Sci. U.S.A. 95 (6), 2979–2984.
ca 137 (2014) 161–173
Couto, A.S., Soprano, L.L., Landoni, M., Pourcelot, M., Acosta, D.M., Bultel, L., Parente,J., Ferrero, M.R., Barbier, M., Dussouy, C., Esteva, M.I., Kovensky, J., Duschak, V.G.,2012. An anionic synthetic sugar containing 6-SO3-NAcGlc mimics the sulfatedcruzipain epitope that plays a central role in immune recognition. FEBS J. 279(19), 3665–3679.
Cunha-e-Silva, N., Sant’Anna, C., Pereira, M.G., Porto-Carreiro, I., Jeovanio, A.L., deSouza, W., 2006. Reservosomes: multipurpose organelles? Parasitol. Res. 99 (4),325–327.
de Lederkremer, R.M., Zingales, B., Confalonieri, A.N., Couto, A.S., Martin, N.F., Colli,W., 1985. In vivo incorporation of palmitic acid and galactose in glycolipids ofTrypanosoma cruzi epimastigotes. Biochem. Int. 10, 79–88.
De Souza, W., Carvalho, T.U., Benchimol, M., 1978. Trypanosoma cruzi: ultrastruc-tural, cytochemical and freeze-fracture studies of protein uptake. Exp. Parasitol.45, 101–115.
Duschak, V.G., Barboza, M., García, G.A., Lammel, E.M., Couto, A.S., Isola, E.L., 2006.Novel cysteine proteinase in Trypanosoma cruzi metacyclogenesis. Parasitology132, 345–355.
Duschak, V.G., Couto, A.S., 2009. Cruzipain, the major cysteine protease of Try-panosoma cruzi: a sulfated glycoprotein antigen as relevant candidate for vaccinedevelopment and drug target. A review. Curr. Med. Chem. 16 (24), 3174–3202.
Duschak, V.G., Couto, A.S., 2010. Targets and patented drugs for chemotherapy ofChagas disease. In: Atta-ur-Rahman, F.R.S., Choudhary, M.I. (Eds.), Anti-InfectiveDrug Discovery, vol. 1. Frontiers, Bentham Science Publishers, pp. 323–408(Chapter 18).
Duschak, V.G., 2011. A decade of targets and patented drugs for chemotherapy ofChagas disease. Recent Pat. Antiinfect Drug Discov. 6 (3), 216–259.
Esko, J.D., Lindahl, U., 2001. Molecular diversity of heparan sulfate. J. Clin. Invest. 108(2), 169–173.
Ey, P.L., Prowse, S.J., Jenkin, C.R., 1978. Isolation of pure IgG1, IgG2a and IgG2bimmunoglobulins from mouse serum using protein A-sepharose. Immunochem-istry 15, 429–436.
Fleckenstein, J.M., Holland, J.T., Hasty, D.L., 2002. Interaction of an outer membraneprotein of enterotoxigenic Escherichia coli with cell surface heparan sulfate pro-teoglycans. Infect. Immun. 70 (3), 1530–1537.
Freeze, H.H., Yeh, R., Miller, A.L., Kornfeld, S., 1983. Structural analysis of theasparagine-linked oligosaccharides from three lysosomal enzymes of Dic-tyostelium discoideum. Evidence for an unusual acid-stable phosphodiester. J.Biol. Chem. 258 (24), 14874–14879.
Freeze, H.H., Mierendorf, R.C., Wunderlich, R., Dimond, R.L., 1984. Sulfated oligosac-charides block antibodies to many Dictyostelium discoideum acid hydrolases. J.Biol. Chem. 259 (16), 10641–10643.
Fukuda, M., Hiraoka, N., Akama, T.O., Fukuda, M.N., 2001. Carbohydrate-modifyingsulfotransferases: structure, function, and pathophysiology. J. Biol. Chem. 276,47747–47750.
Garavaglia, P.A., Cannata, J.J., Ruiz, A.M., Maugeri, D., Duran, R.,Galleano, M., García, G.A., 2010. Identification, cloning and charac-terization of an aldo-keto reductase from Trypanosoma cruzi withquinone oxido-reductase activity. Mol. Biochem. Parasitol. 173 (2),132–141.
Garrido, V.V., Dulgerian, L.R., Stempin, C.C., Cerbàn, F.M., 2011. The increase inmannose receptor recycling favors arginase induction and Trypanosoma cruzisurvival in macrophages. Int. J. Biol. Sci. 7 (9), 1257–1272.
Herrera, E.M., Ming, M., Ortega-Barria, E., Pereira, M.E., 1994. Mediation of Try-panosoma cruzi invasion by heparan sulfate receptors on host cells and penetrincounter-receptors on the trypanosomes. Mol. Biochem. Parasitol. 65 (1),73–83.
Honke, K., Taniguchi, N., 2002. Sulfotransferases and sulfated oligosaccharides. Med.Res. Rev. 22, 637–654.
Kawasaki, N., Ohta, M., Hyuga, S., Hyuga, M., Hayakawa, T., 2000. Application of liquidchromatography mass spectrometry and liquid chromatography with tandemmass spectrometry to the analysis of the site specific carbohydrate heterogene-ity in erythropoietin. Anal. Biochem. 285, 82–91.
Klaassen, C.D., Boles, J.W., 1997. Sulfation and sulfotransferases 5: the importanceof 3′-phosphoadenosine 5′-phosphosulfate (PAPS) in the regulation of sulfation.FASEB J. 11 (6), 404–418.
Kovensky, J., 2009. Sulfated oligosaccharides: new targets for drug development?Curr. Med. Chem. 16, 2338–2344.
Lefkowitz, D.L., Lincoln, J.A., Lefkowitz, S.S., Bollen, A., Moguilevsky, N., 1997.Enhancement of macrophage-mediated bactericidal activity by macrophage-mannose receptor-ligand interaction. Immunol. Cell Biol. 75, 136–141.
Linehan, S.A., 2005. The mannose receptor is expressed by subsets of APC in non-lymphoid organs. BMC Immunol. 6, 4–14.
Liu, Y., Misulovin, Z., Bjorkman, P.J., 2001. The molecular mechanism of sulfatedcarbohydrate recognition by the cysteine-rich domain of mannose receptor. J.Mol. Biol. 305, 481–490.
Martinez, J., Campetella, O., Frasch, A.C.C., Cazzulo, J.J., 1991. The major cysteineproteinase (cruzipain) from Trypanosoma cruzi is antigenic in human infections.Infect Immun. 59 (11), 4275–4277.
Martinez-Pomares, L., Wienke, D., Stillion, R., McKenzie, E.J., Arnold, J.N., Harris, J.,McGreal, E., Sim, R.B., Isacke, C.M., Gordon, S., 2006. Carbohydrate-independentrecognition of collagens by the macrophage mannose receptor. Eur. J. Immunol.
36, 1074–1082.Meirelles, M.N., Pereira, M.C., Singer, R.H., Soeiro, M.N., Garzoni, L.R., Silva, D.T., Bar-bosa, H.S., Araujo-Jorge, T.C., Masuda, M.O., Capella, M.A., Lopes, A.G., Vermelho,A.B., 1999. Trypanosoma cruzi-cardiomyocytes: new contributions regarding abetter understanding of this interaction. Mem. Inst. Oswaldo Cruz 94, 149–152.

Tropi
M
M
O
O
P
P
P
P
R
S
S
S
S
S
M.R. Ferrero et al. / Acta
enna-Barreto, R.F., Salomão, K., Dantas, A.P., Santa-Rita, R.M., Soares, M.J., Barbosa,H.S., de Castro, S.L., 2009. Different cell death pathways induced by drugs inTrypanosoma cruzi: an ultrastructural study. Micron 40, 157–168.
oolani, Y., Bukhman, G., Hotez, P.J., 2012. Neglected tropical diseases as hiddencauses of cardio-vascular disease. PLoS Negl. Trop. Dis. 6 (6), e1499.
liveira Jr., F.O., Alves, C.R., Calvet, C.M., Toma, L., Bouc as, R.I., Nader, H.B., CastroCôrtes, L.M., Krieger, M.A., Meirelles, M.N., Souza Pereira, M.C., 2008. Try-panosoma cruzi heparin-binding proteins and the nature of the host cell heparansulfate-binding domain. Microb. Pathog. 44 (4), 329–338.
rtega-Barria, E., Boothroyd, J.C., 1999. A Toxoplasma lectin-like activity specific forsulfated polysaccharides is involved in host cell infection. J. Biol. Chem. 274,1267–1276.
ancake, S.J., Holt, G.D., Mellouk, S., Hoffman, S.L., 1992. Malaria sporozoites andcircumsporozoite proteins bind specifically to sulfated glycoconjugates. J. CellBiol. 117, 1351–1357.
arodi, A.J.A., Labriola, C., Cazzulo, J.J., 1995. The presence of complex-type oligosac-charides at the C-terminal domain glycosylation site of some molecules ofcruzipain. Mol. Biochem. Parasitol. 69, 247–255.
arussini, F., Duschak, V.G., Cazzulo, J.J., 1998. Membrane-bound cysteine proteinaseisoforms in different developmental stages of Trypanosoma cruzi. Cell. Mol. Biol.44, 513–519.
érez-Molina., J.A., Norman, F., López-Vélez, R., 2012. Chagas disease in non-endemiccountries: epidemiology, clinical presentation and treatment. Curr. Infect. Dis.Rep. 14, 263–274.
apraeger, A.C., Krufka, A., Olwin, B.B., 1991. Requirement of heparan sulfate forßFGF-mediated fibroblast growth and myoblast differentiation. Science 252,1705–1708.
ant’Anna, C., Nakayasu, E.S., Pereira, M.G., Lourenc o, D., de Souza, W., Almeida, I.C.,Cunha-E-Silva, N.L., 2009. Subcellular proteomics of Trypanosoma cruzi reservo-somes. Proteomics 9 (7), 1782–1794.
antos Barrias, E., Ulisses de Carvalho, T.M., De Souza, W., 2013. Trypanosoma cruzi:entry into mammalian host cells and parasitophorous vacuole formation. Front.Immunol. 4 (186), 1–10.
charfstein, J., Rodriguez, M.M., Alves, C.A., De Souza, W., Previato, J.O., Mendonca-Previato, L., 1983. Trypanosoma cruzi: description of a highly purified surfaceantigen defined by human antibodies. J. Immunol. 131 (2), 972–976.
charfstein, J., Luquetti, A., Murta, A.C., Senna, M., Rezende, J.M., Rassi, A., Mendonc a-Previato, L., 1985. Chagas’ disease: serodiagnosis with purified Gp25 antigen.Am. J. Trop. Med. Hyg. 34 (6), 1153–1160.
charfstein, J., Schechter, M., Senna, M., Peralta, J.M., Mendonc a-Previato, L., Miles,M.A., 1986. Trypanosoma cruzi: characterization and isolation of 57/51000 m.w.
ca 137 (2014) 161–173 173
surface glycoprotein (GP 57/51) expressed by epimastigotes and bloodstreamtrypomastigotes. J. Immunol. 137 (4), 1336–1344.
Shibata, Y., Metzger, W.J., Myrvik, Q.N., 1997. Chitin particle-induced cell-mediatedimmunity is inhibited by soluble mannan: mannose receptor-mediated phago-cytosis initiates IL-12 production. J. Immunol. 159, 2462–2467.
Soares, M.J., De Souza, M.F., De Souza, W., 1987. Ultrastructural visualization of lipidsin trypanosomatids. J. Protozool. 34, 199–203.
Soares, M.J., De Souza, W., 1988. Cytoplasmic organelles of trypanosomatids. A cyto-chemical and stereological study. J. Submicrosc. Cytol. Pathol. 20, 349–363.
Soeiro, M.N., Paiva, M.M., Barbosa, H.S., Meirelles, M.N., Araújo-Jorge, T.C., 1999.A cardiomyocytes mannose receptor system is involved in Trypanosoma cruziinvasion and is down-modulated after infection. Cell Struct. Funct. 24, 139–149.
Souto-Padron, T., Campetella, O.E., Cazzulo, J.J., De Souza, W., 1990. Cys-teine proteinase in Trypanosoma cruzi: immunocytochemical localization andinvolvement in parasite–host cell interaction. J. Cell Sci. 96, 485–490.
Stempin, C.C., Tanos, T.B., Coso, O.A., Cerbán, F.M., 2004. Arginase induction pro-motes Trypanosoma cruzi intracellular replication in cruzipain-treated J774 cellsthrough the activation of multiple signaling pathways. Eur. J. Immunol. 34 (1),200–209.
Stempin, C.C., Garrido, V.V., Dulgerian, L.R., Cerbán, F.M., 2008. Cruzipain andSP600125 induce p38 activation, alter NO/arginase balance and favor the sur-vival of Trypanosoma cruzi in macrophages. Acta Trop. 106 (2), 119–127.
Stoka, V., Nycander, M., Lenarcic, B., Labriola, C., Cazzulo, J.J., Björk, I., Turk, V., 1995.Inhibition of cruzipain, the major cysteine proteinase of the protozoan parasite,Trypanosoma cruzi, by proteinase inhibitors of the cystatin superfamily. FEBSLett. 370, 101–104.
Tomas, A.M., Miles, M.M., Kelly, J.M., 1997. Overexpression of cruzipain, the majorcysteine proteinase of Trypanosoma cruzi, is associated with enhanced metacy-clogenesis. Eur. J. Biochem. 244, 596–603.
Uhrig, M.L., Couto, A.S., de Lederkremer, R.M., Zingales, B., Colli, W., 1992. Metaboliclabelling and partial characterisation of a sulfoglycolipid in Trypanosoma cruzitrypomastigotes. Carbohydr. Res. 231, 329–334.
Vázquez-Mendoza, A., Carrero, J.C., Rodriguez-Sosa, M., 2013. Parasitic infections: arole for C-type lectins receptors. BioMed Res. Int. 2013, 1–12.
WHO, 2010. Available at: http://www.who.int/mediacentre/factsheets/fs340/en/index.html
Yokoyama-Yasunaka, J.K.U., Pral, E.M.F., Oliveira Jr., O.C., Alfieri, S.C., Stolf, A.M.S.,1994. Trypanosoma cruzi: identification of proteinases in shed components oftrypomastigote forms. Acta Trop. 57, 307–315.
Yoshida, N., Cortez, M., 2008. Trypanosoma cruzi: parasite and host cell signalingduring the invasion process. Subcell. Biochem. 47, 82–91.














