
6-Bromoindirubin-3'-Oxime Inhibits JAK/STAT3 Signaling and Induces Apoptosis of Human Melanoma Cells
Upload
institutoadolfolutzCategory
view
0download
0
Available online at www.sciencedirect.com
www.elsevier.com/locate/ygcen
General and Comparative Endocrinology 156 (2008) 218–223
Biological effects of insulin on murine melanoma cells and fisherythrophoroma cells: A comparative study
Adriana Luchs a,*, Doris Hissako Sumida b,1, Maria Aparecida Visconti b,Ana Maria de Lauro Castrucci a,b,c
a Pos-Graduation Program, Department of Physiology and Biophysics, Biomedical Science Institute, University of Sao Paulo, Sao Paulo, SP, Brazilb Department of Physiology, Biosciences Institute, University of Sao Paulo, Sao Paulo, SP, Brazil
c Department of Biology, University of Virginia, Charlottesville, VA, USA
Received 25 August 2007; revised 19 January 2008; accepted 5 February 2008Available online 12 February 2008
Abstract
Insulin is the hormone that plays an essential role in metabolism and mitosis of normal and tumor cells, exerting its pleiotropic effectsthrough binding to specific membrane receptors and promoting the phosphorylation of tyrosine residues of the receptor itself and ofother components of the signaling pathway. The aim of this study was to investigate the effects of insulin on melanogenesis and cellgrowth in three different cell lines: the goldfish GEM-81 erythrophoroma cells (undifferentiated and differentiated with 1.5% dimethyl-sulfoxide-DMSO), and the murine B16F10 and Cloudman S91 melanoma cells. Undifferentiated GEM-81 and B16F10 cells responded toinsulin with a small increase of cell proliferation, whereas S91 cells responded with a decrease of growth. In the two mammalian cell lines,and in DMSO-differentiated GEM-81 cells, the hormone strongly inhibited melanogenesis, by decreasing tyrosinase activity. In undif-ferentiated GEM-81 cells, insulin had no effect on tyrosinase activity. An increase in the tyrosine phosphorylation status of pp185 (insulinreceptor substrate 1 and 2—IRS-1/2) phosphorylation degree was observed in S91 mouse melanoma and in differentiated GEM-81 ery-throphoroma cells, suggesting that this specific protein was maintained during transformation process and participates in insulin signal-ing. Our results imply an ancient and diverse history of the insulin signaling system in vertebrate pigment cells.� 2008 Elsevier Inc. All rights reserved.
Keywords: Pigment cells; Insulin; Melanogenesis; Cell proliferation; Signaling pathway
1. Introduction
An increasing number of hormones has been found toaffect the growth and metabolism of cell lines in vitro (Row-bottom et al., 1981). Among the polypeptide hormones,insulin is one of the most complex because it plays diverseroles including stimulation of glucose transport, proteinand glycogen synthesis, inhibition of lipolysis, regulation
0016-6480/$ - see front matter � 2008 Elsevier Inc. All rights reserved.
doi:10.1016/j.ygcen.2008.02.001
* Corresponding author. Present address: Laboratory of Enteric Virus,Virology Service, Medical Biology Division, Adolfo Lutz Institute, SaoPaulo, SP, Brazil. Fax: +55 11 30883041.
E-mail address: [email protected] (A. Luchs).1 Present address: Department of Basics Sciences, College of Odontol-
ogy, University of Paulista State Julio de Mesquita Filho, Aracatuba, SP,Brazil.
of gene transcription and translation, and cell growthand proliferation (Kim and Novak, 2007).
Insulin appeared early in evolution, as demonstrated bythe presence of insulin-like peptides in various inverte-brates, including nematodes, mollusks, and insects (Claeyset al., 2002; Wu and Brown, 2006). The insulin receptor ispresent in virtually all vertebrate tissues, although its den-sity ranges from 40 receptors in erythrocytes to 200,000 inadipocytes and hepatocytes (White and Kahn, 1994). Insu-lin rapidly stimulates tyrosine phosphorylation of intracel-lular proteins with 165 and 190 kDa. These proteins arecollectively called pp185 and include the insulin receptorsubstrates IRS (Sun et al., 1991,1995; Miralpeix et al.,1992). Among the six family members, IRS 1, 2, and 4 existin humans and mice, IRS 1 and 2 been widely distributed in

A. Luchs et al. / General and Comparative Endocrinology 156 (2008) 218–223 219
most tissues (Cai et al., 2003). Tyrosine-phosphorylatedIRS is able to bind effector molecules thus amplifying insu-lin receptor signaling (White and Kahn, 1994; Cheathamand Kahn, 1995; White, 2002).
Normal melanocytes depend on a combination of insu-lin-like growth factor (IGF-I) or insulin, 12-O-tetradeca-noyl phorbol-13-acetate (TPA), a-melanocyte stimulatinghormone (a-MSH), and basic fibroblast growth factor(bFGF) for in vitro proliferation. Cells from primary mel-anomas require only IGF-I or insulin for continuousgrowth, and metastatic melanoma cells can proliferate inbasic medium without addition of any growth factors orproteins (Graeven and Herlyn, 1992; Oka et al., 2004).
Human and mice melanoma cell lines are common cul-ture models used to investigate insulin effects on prolifera-tion and related pathways (Kahn et al., 1980; Androlewiczet al., 1989; Rodeck et al., 1991; Slominski et al., 1992;Benathan and Labidi, 1997), but little is known about itsaction in pigment cell lines of ectothermic vertebrates.GEM-81, an erythrophoroma cell line from a cutaneousred pigment cell-derived tumor of the teleost Carassius
auratus (Matsumoto et al., 1980), is a useful cultured fishpigment cell line. Treatment with 1–2% dimethylsulfoxide(DMSO) induces terminal differentiation of these cells,which exhibit increased melanization (Matsumoto et al.,1982, 1984).
The aim of this study was to investigate the effects ofinsulin on melanogenesis and cell growth of DMSO-differ-entiated and undifferentiated fish cells and compare theresponses with those of two mouse melanoma cells lines,B16F10 and Cloudman S91.
2. Materials and methods
2.1. Cell lines
The murine B16F10 melanoma cell line and the GEM-81 goldfish ery-throphoroma cell line were kind gifts of Dr. Vincent Hearing (NationalInstitute of Health, Bethesda, MD, USA) and Dr. Jiro Matsumoto (KeioUniversity, Yokohama, Japan), respectively. Murine Cloudman S91 mel-anoma cell line was purchased from the American Type Culture of CellCollection repository (ATCC, Rockville, MD, USA). All cell lines weregrown as monolayer cultures in Ham’s F-10 medium (Cultilab, Campinas,SP, Brazil), supplemented with 5% (S91) or 10% (B16F10 and GEM-81)fetal bovine serum (Gibco, Carlsbad, California USA), in the absence ofantibiotics, at 37 �C in 5% CO2 (S91 and B16F10) or 25 �C (GEM-81)humidified atmosphere. The culture medium was replaced every 3–4 days.
2.2. Cell treatments
S91 and B16F10 were seeded at 2 � 105 cells and GEM-81 at 5 � 105
cells per 25 cm2 flasks. After 24 h in culture, insulin (Sigma, St. Louis,MO, USA) was added at final concentrations of 0.1, 1, 10, 100, and1000 ng/mL for 24, 48, 72, and 96 h treatment. The concentration of fetalbovine serum was reduced to 2% (S91) and 4% (B16F10 and GEM-81),during the experiments. Same treatments were performed with GEM-81cells previously differentiated with 1.5% of DMSO (Merck S.A., White-house Station, NJ, USA or Labsynth Ltd., SP, Brazil) for 4 days, afterwhich period the hormone was added to the medium. Three independentassays were performed in triplicate, and the medium was replacedevery 48 h.
2.3. Cell proliferation
At the end of each experiment, cells were harvested with Tyrode/EDTA (ethylenediaminetetracetic acid, Sigma, St. Louis, MO, USA) solu-tion, resuspended in sodium phosphate-buffered saline (PBS) and countedwith a hemocytometer in a phase contrast microscope. The cell numberswere expressed as percent of control.
2.4. Tyrosinase activity
Twenty-four hours before ending the experiment, the medium wasreplaced with fresh medium containing 0.7 lCi/mL of [3H]-tyrosine (spe-cific activity 57 Ci/mmol, GE Healthcare, Piscataway, NJ, USA), to deter-mine tyrosinase activity (Pomerantz, 1969, modified according toMengeaud and Ortonne, 1994). Briefly, 10% of charcoal (Sigma, St. Louis,MO, USA) dissolved in 10% trichloroacetic acid (Sigma, St. Louis, MO,USA) was added to the medium. The mixture was vortexed vigorously,incubated at 9 �C for 20 min, and centrifuged at 3000g for 10 min. Thesupernatant (500 ll) was added to 5 ml of scintillation fluid and cpm val-ues were obtained in a beta counter (Tri-Carb, 2100 TR, Packard, Ram-sey, MN, USA), and converted to percentage of control.
2.5. Western blotting
Twenty-four hours after seeding (1 � 106 cells/75 cm2 flask), S91 cellsand GEM-81 cells were treated for 5, 10, 15, 30, and 60 min with 1000 ng/mL insulin, in the absence of fetal calf serum. The cells were lysed with1 mL RIPA buffer (1.2% aprotinin, 4 lM antipain, 5 mM EDTA, 5 lMleupeptin, 1.1 mM Na3VO4, 150 mM NaCl, 1.4 mM PMSF, 0.1% SDS,10 mM Tris–HCl, 1% Triton X-100, pH 7.4; Sigma Chem. Co., St. Louis,MO, USA) and incubated at 100 �C for 10 min. The samples were thencooled on ice, and centrifuged at 16000g for 40 min at 4� C. The proteinconcentration was determined by DC protein assay (Bio-Rad, Hercules,CA, USA) and kept in Laemmli buffer (0.1% bromophenol blue, 50% glyc-erol, 10% SDS, 1 M sodium phosphate pH 7, 15% DTT, Sigma Chem.Co., St. Louis, MO, USA). Equal amounts of protein (185 lg) from eachsample were separated by SDS–PAGE (100 V), and then transferred ontoa nitrocellulose membrane. The membranes were initially incubated withblocking solution (150 mM NaCl, 10 mM Tris, 0.02% Tween 20, SigmaChem. Co., St. Louis, MO, USA, added to 5% free fat milk) for 2 h at25 �C. After the washing step, the membranes were incubated in a solutioncontaining monoclonal anti-phosphotyrosine antibody (1:200, SantaCruz,Ca, USA) for 4 h at 25 �C. Membranes were washed and incubated for 1 hwith the secondary anti-mouse antibody conjugated to horseradish perox-idase (1:10,000, GE Healthcare, Piscataway, NJ, USA). After a final wash-ing step, the membranes were revealed using a chemioluminescence kit,according to the manufacturer instructions (GE Healthcare, Piscataway,NJ, USA), and exposed to X-ray film (Hyperfilm ECL, GE Healthcare,Piscataway, NJ, USA) for 5 min at 25 �C. The band intensities were quan-tified by optic densitometry (NIH IMAGE software, Version 1.62, byWayne Rasband, National Institutes of Health, Bethesda) of the devel-oped autoradiographs.
2.6. Statistical analysis
Each experiment was carried out at least three times (in triplicate),yielding similar results in each occasion. Before the percentage transfor-mation, data were analyzed by one-way ANOVA followed by Student–Newman–Keuls (SNK) test and the differences were considered significantfor p < 0.05.
3. Results
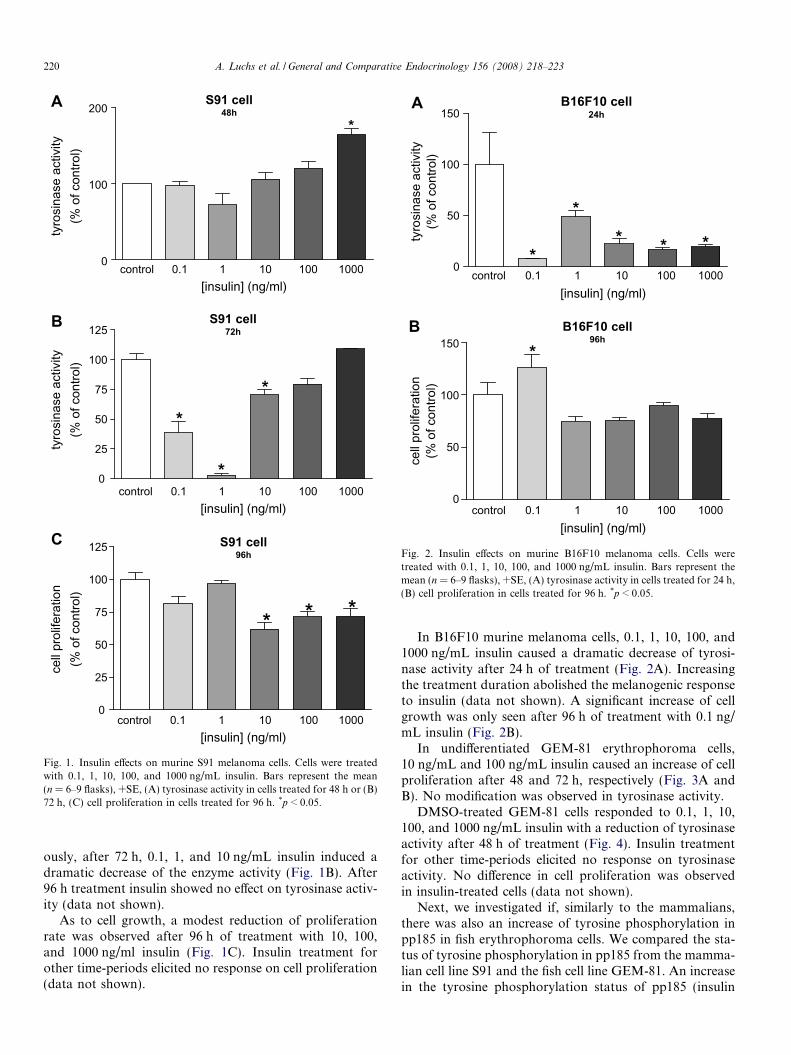
In S91 murine melanoma cells, 1000 ng/mL insulincaused a small increase of tyrosinase activity after 48 h.Lower concentrations exhibited no effect (Fig. 1A). Curi-
0
100
200*
tyro
sina
se a
ctiv
ity(%
of c
ontro
l)ty
rosi
nase
act
ivity
(% o
f con
trol)
S91 cell48h
A
0
25
50
75
100
125
*
*
*
S91 cell72h
B
control 0.1 1 10 100 10000
25
50
75
100
125
* * *
[insulin] (ng/ml)
control 0.1 1 10 100 1000[insulin] (ng/ml)
control 0.1 1 10 100 1000[insulin] (ng/ml)
cell
prol
ifera
tion
(% o
f con
trol)
S91 cell96h
C
Fig. 1. Insulin effects on murine S91 melanoma cells. Cells were treatedwith 0.1, 1, 10, 100, and 1000 ng/mL insulin. Bars represent the mean(n = 6–9 flasks), +SE, (A) tyrosinase activity in cells treated for 48 h or (B)72 h, (C) cell proliferation in cells treated for 96 h. *p < 0.05.
0
50
100
150
*
** * *
control 0.1 1 10 100 1000[insulin] (ng/ml)
control 0.1 1 10 100 1000[insulin] (ng/ml)
tyro
sina
se a
ctiv
ity(%
of c
ontro
l)
B16F10 cell24h
A
0
50
100
150 *
cell
prol
ifera
tion
(% o
f con
trol)
B16F10 cell96h
B
Fig. 2. Insulin effects on murine B16F10 melanoma cells. Cells weretreated with 0.1, 1, 10, 100, and 1000 ng/mL insulin. Bars represent themean (n = 6–9 flasks), +SE, (A) tyrosinase activity in cells treated for 24 h(B) cell proliferation in cells treated for 96 h. *p < 0.05.
220 A. Luchs et al. / General and Comparative Endocrinology 156 (2008) 218–223
ously, after 72 h, 0.1, 1, and 10 ng/mL insulin induced adramatic decrease of the enzyme activity (Fig. 1B). After96 h treatment insulin showed no effect on tyrosinase activ-ity (data not shown).
As to cell growth, a modest reduction of proliferationrate was observed after 96 h of treatment with 10, 100,and 1000 ng/ml insulin (Fig. 1C). Insulin treatment forother time-periods elicited no response on cell proliferation(data not shown).
,
In B16F10 murine melanoma cells, 0.1, 1, 10, 100, and1000 ng/mL insulin caused a dramatic decrease of tyrosi-nase activity after 24 h of treatment (Fig. 2A). Increasingthe treatment duration abolished the melanogenic responseto insulin (data not shown). A significant increase of cellgrowth was only seen after 96 h of treatment with 0.1 ng/mL insulin (Fig. 2B).
In undifferentiated GEM-81 erythrophoroma cells,10 ng/mL and 100 ng/mL insulin caused an increase of cellproliferation after 48 and 72 h, respectively (Fig. 3A andB). No modification was observed in tyrosinase activity.
DMSO-treated GEM-81 cells responded to 0.1, 1, 10,100, and 1000 ng/mL insulin with a reduction of tyrosinaseactivity after 48 h of treatment (Fig. 4). Insulin treatmentfor other time-periods elicited no response on tyrosinaseactivity. No difference in cell proliferation was observedin insulin-treated cells (data not shown).
Next, we investigated if, similarly to the mammalians,there was also an increase of tyrosine phosphorylation inpp185 in fish erythrophoroma cells. We compared the sta-tus of tyrosine phosphorylation in pp185 from the mamma-lian cell line S91 and the fish cell line GEM-81. An increasein the tyrosine phosphorylation status of pp185 (insulin

GEM-81 cell48h
A
control 0.1 1 10 100 10000
100
200
*
[insulin] (ng/ml)
control 0.1 1 10 100 1000
[insulin] (ng/ml)
cell
prol
ifera
tion
(% o
f con
trol)
cell
prol
ifera
tion
(% o
f con
trol)
0
50
100
150 *GEM-81 cell
72hB
Fig. 3. Insulin effects on undifferentiated GEM-81 erythrophoroma cells.Bars represent the mean (n = 6–9 flasks), +SE, cell proliferation in cellstreated for (A) 48 h or (B) 72 h, with 0.1, 1, 10, 100, and 1000 ng/mLinsulin. *p < 0.05.
control 0.1 1 10 100 10000
25
50
75
100
125
** *
*
*
[insulin] (ng/ml)
tyro
sina
se a
ctiv
ity
(% o
f con
trol
)
GEM-81 cell48h
Fig. 4. Insulin effects on differentiated GEM-81 erythrophoroma cells.Bars represent the mean (n = 6–9 flasks), +SE, tyrosinase activity. Cellswere treated for 48 h with 0.1, 1, 10, 100, and 1000 ng/mL insulin.*p < 0.05.
0 5’ 10’ L 30’ 60’
pp185
Fig. 5. Typical autoradiogram of insulin-stimulated tyrosine phosphory-lation status of pp185 from murine S91 melanoma cells. These cells weretreated for 0, 5, 10, 30, and 60 min (lanes 1, 2, 3, 4, and 5, respectively)with 1000 ng/mL insulin (L = protein ladder).
0 5’ 10’ L 30’ 60’
pp185
Fig. 6. Typical autoradiogram of insulin-stimulated tyrosine phosphory-lation status of pp185 from differentiated GEM-81 erythrophoroma cells.These cells were treated for 0, 5, 10, 30, and 60 min (lanes 1, 2, 3, 4, and 5,respectively) with 1000 ng/mL insulin (L = protein ladder).
A. Luchs et al. / General and Comparative Endocrinology 156 (2008) 218–223 221
receptor substrate 1 and 2—IRS-1/2) phosphorylationdegree was observed in S91 mouse melanoma (Fig. 5)
and differentiated GEM-81 erythrophoroma cells (Fig. 6)after 5, 10, 30 and 60 min (lanes 2, 3, 5, and 6) of treatment.
4. Discussion
Long term color change may occur in vertebrates by pig-ment cell proliferation and/or pigment synthesis (a processknown as melanogenesis in the case of melanocytes), whichcan be modulated by several hormones (Nery and Cast-rucci, 1997). Since its isolation in 1921, insulin mechanismof action has been intensively studied reflecting the biolog-ical importance of the hormone for human biology andmedicine (al-Habori, 1993). The molecular mechanisms ofinsulin action are far from being completely understood,moreover because its signaling pathway shares many com-ponents with several growth factors, including IGF-I(Rowbottom et al., 1981; Argiles and Lopez-Soriano,2001).
Although insulin pleiotropic effects have been widelydescribed, it is not known whether this hormone plays arole in vertebrate melanogenesis. Tyrosinase, a highly con-served (Slominski et al., 2004) rate-limiting enzyme formelanogenesis, catalyzes the conversion of L-tyrosine todopaquinone, which is required for the synthesis of eithereumelanin or phaeomelanin (Tsatmali et al., 2002). Weobserved a small increase of melanin production in murineS91 melanoma cells, after 48 h treatment with the highestconcentration of insulin. This effect on melanogenesis wasreversed after 72 h treatment, and a marked inhibition oftyrosinase activity was elicited by 0.1, 1, and 10 ng/mLinsulin. In B16F10 cells, insulin at all concentrations dras-tically decreased tyrosinase activity after 24 h treatment, inaccordance to what had already been reported for mouse(Fuller et al., 1990) and human melanoma cells (Benathanand Labidi, 1997).
In addition, we also demonstrated that insulin can affectpigmentation in mammals through the modulation of cell

222 A. Luchs et al. / General and Comparative Endocrinology 156 (2008) 218–223
proliferation. Murine S91 Cloudman melanoma cellsexhibited a significant decrease in proliferation after 96 htreatment with 10, 100, and 1000 ng/mL insulin. Otherincubation periods with the hormone did not affect cellgrowth. In fact, the effects of insulin on S91 cells seem tobe controversial, since the hormone has been reported byvarious laboratories to either stimulate or inhibit murineS91 melanoma growth (Kahn et al., 1980; Haring et al.,1984; Bregman et al., 1985; Fleischmann and Pawelek,1985). This may be explained by the fact that S91 line isheterogeneous; when the variant clones were individuallyisolated from the wild-type Cloudman melanoma by Paw-elek and colleagues (Kahn et al., 1980; Fleischmann andPawelek, 1985; Slominski et al., 1992), the cloned cell lineswere demonstrated to be either resistant to insulin ordependent upon insulin for growth.
In opposition to S91 cells, murine B16F10 melanoma cellsexhibited only a small increase in proliferation after 96 h incontact with the lowest insulin concentration. In fact, Row-bottom et al. (1981) showed that insulin enhanced the growthof various tumor cell lines, including melanoma lines. Kahnet al. (1980) tested the insulin effect on five melanoma celllines, including Cloudman and B16F10 melanoma, and theinhibition seemed to be unique to the S91 Cloudman line.Wortmannin, an inhibitor of PI 3-kinase, has been reportedto potently inhibit the protective effect of insulin on melano-cytes, without affecting the ability of TPA, bFGF, or IBMXto rescue the cells from apoptosis (Oka et al., 2004). Thisresult may indicate that insulin stimulates cell proliferationthrough PI 3-kinase activation.
To our knowledge, insulin action on erythrophoromacells has never been reported in the literature. Insulininduced what makes this paper unique. Insulin inducedsmall growth stimulation in undifferentiated GEM-81 cellsafter 48 and 72 h, at 10 and 100 ng/mL, respectively. Nomelanogenic effect was observed. Interestingly, differenti-ated GEM-81 cells exhibited the opposite response, nomodulation of cell growth was observed but a dramaticreduction of tyrosinase activity occurred after 48 h treat-ment, at all concentrations of insulin. DMSO-treatmentseems to have increased the sensitivity of the melanogenicpathway to insulin. DMSO is known to induce, per se, asignificant decrease of GEM-81 cell proliferation (Matsum-oto et al., 1982, 1984; Filadelfi et al., 2004). Endothelinsand sarafotoxins (Filadelfi et al., 2004) and melanin-con-centrating hormone (MCH) (Isoldi et al., 2004) increasethe proliferation of both GEM-81 cells types through theactivation of phospholipase C and PKCb. Most probably,however, insulin is stimulating proliferation through theactivation of the mitogen-activated protein kinases(MAPK) cascade (Kim and Novak, 2007).
Apparently, the DMSO treatment of GEM-81 cells doesnot induce melanogenesis, since melanin levels were similarand low in both undifferentiated and DMSO-differentiatedGEM-81 cells (Filadelfi et al., 2004). In DMSO-differenti-ated GEM-81 cells, however, insulin induced a decreasein melanogenesis, as observed in the two murine cell lines.
Then, insulin could be considered as a lightening agent inGEM-81 as well as in mammalian cells. The inhibition oftyrosinase activity may be due to the activation of phos-phodiesterase 3 by Akt (Kim and Novak, 2007), leadingto a decrease of cAMP, well known second messenger ofmelanogenesis (Abdel-Malek et al., 1995).
In addition, although the hormone induced significantmitogenic effects on undifferentiated GEM-81 andB16F10 cells, the increase in proliferation was small, incontrast with what has been reported for several tumor celllines (Goodwin et al., 2002; Rowbottom et al., 1981;Argiles and Lopez-Soriano, 2001). In teleost GEM-81 cells,and murine B16F10 and S91 melanoma cells, the strongesteffects of insulin were seen on tyrosinase activity, and noton cell growth.
In S91 cells, we observed a remarkable increase in tyro-sine phosphorylation of pp185 after insulin treatment. Thepp185 protein corresponds to the IRS-1/2 migration pat-tern on SDS–PAGE (Sun et al., 1991; Miralpeix et al.,1992). The IRS-proteins belong to a class of docking pro-teins which mediate insulin, growth factor and a growingnumber of cytokine actions, especially activated by tyro-sine kinase receptors (White, 1998). The protein phosphor-ylation pattern in melanoma cells suggests that insulinintracellular pathways are constitutively activated in thesecells and the IRS family leads to activation of Ras (mito-gen-activated protein kinase), MAPKs, and phosphoinosi-tol-3-kinase (PI3 K), events involved in the mitogeniccascade (All-Ericsson et al., 2002). The pp185 presenceand its enhanced tyrosine phosphorylation suggests thatthis protein was conserved during the tumor developmentand it plays a role in insulin signaling pathway of mamma-lian transformed pigment cells.
For the first time pp185 was identified in differentiatederythrophoroma GEM-81 cells, and insulin elicits a high sta-tus of tyrosine phosphorylation, similar to what wasobserved in mammalian cells. This finding permits to con-clude that pp185 function was maintained during the trans-formation process and corroborates the ancientevolutionary history of the IRS-protein family in thevertebrates.
In conclusion, our results suggest a long and diverse his-tory of insulin/insulin receptor/insulin signaling system invertebrate pigment cells. As perspective for further study,we suggest the investigation of other components of theinsulin signaling cascade in GEM-81 cells to compare towhat is known in the mammalian cell and to confirm ourfinding that the signaling of insulin in fish pigment cellwas conserved along evolution. The GEM-81 tumor modelis only beginning to be exploited and provides the uniqueopportunity for novel insights into the cellular and molec-ular biology of pigment cell signal transduction.
Acknowledgments
This work was partially supported by FAPESP (Grants96/6490-0, 97/06896-1, and 99/00131-9) and CNPq (Grant

A. Luchs et al. / General and Comparative Endocrinology 156 (2008) 218–223 223
550676/2002-3), Brazil. A.L. and D.H.S. were FAPESP fel-lows (00/08785-7 and 01/10469-9, respectively). We areespecially thankful to Ana Cristina Scarparo, Ana MariaCaliman Filadelfi and Luciana Nogueira de Sousa And-rade, who in different ways, contributed to this work. Weare grateful to the valuable technical assistance of TelmaPazzini and Marcio Patto Lima.
References
Abdel-Malek, Z., Swope, V.B., Suzuki, I., Akcali, C., Harriger, M.D.,Boyce, S.T., Urabe, K., Hearing, V.J., 1995. Mitogenic and melano-genic stimulation of normal human melanocytes by melanotropicpeptides. Proc. Natl. Acad. Sci. USA 92, 1789–1793.
al-Habori, M., 1993. Mechanism of insulin action, role of ions and thecytoskeleton. Int. J. Biochem. 25, 1087–1099.
All-Ericsson, C., Girnita, L., Seregard, S., Bartolazzi, A., Jager, M.J.,Larsson, O., 2002. Insulin-like growth factor-1 receptor in uvealmelanoma: a predictor for metastatic disease and a potential thera-peutic target. Invest. Ophthalmol. Vis. Sci. 43, 1–8.
Androlewicz, M.J., Brandenburg, D.F., Straus, D.S., 1989. Insulinreceptor internalization defect in an insulin-resistant mouse melanomacell line. Biochemistry 28, 9750–9757.
Argiles, J.M., Lopez-Soriano, F.J., 2001. Insulin and cancer (Review). Int.J. Oncol. 18, 683–687.
Benathan, M., Labidi, F., 1997. Insulin inhibits tyrosinase activity and 5-S-cysteinyldopa formations in human melanoma cells. Acta Derm.Venereol. 77, 281–284.
Bregman, M.D., Abdel Malek, Z.A., Meyskens Jr., F.L., 1985. Anchor-age-independent growth of murine melanoma in serum-less media isdependent on insulin or melanocyte-stimulating hormone. Exp. CellRes. 157, 419–428.
Cai, D., Dhe-Paganon, S., Melendez, P.A., Lee, J., Shoelson, S.E., 2003.Two new substrates in insulin signaling, IRS5/DOK4 and IRS6/DOK5. J. Biol. Chem. 278, 25323–25330.
Cheatham, B., Kahn, C.R., 1995. Insulin action and the insulin signalingnetwork. Endocr. Rev. 16, 117–142.
Claeys, I., Simonet, G., Poels, J., Van Loy, T., Vercammen, L., De Loof,A., Vanden Broeck, J., 2002. Insulin-related peptides and theirconserved signal transduction pathway. Peptides 23, 807–816.
Filadelfi, A.M., Ramanzini, G.C., Visconti, M.A., Castrucci, A.M., 2004.The endothelin/sarafotoxin-induced increase of the proliferation ofundifferentiated and DMSO-differentiated GEM-81 goldfish erythro-phoroma cells is mediated by ETB receptors. Pigment Cell Res. 17,480–487.
Fleischmann, R.D., Pawelek, J.M., 1985. Evidence that a 90-kDaphosphoprotein, an associated kinase, and a specific phosphatase areinvolved in the regulation of Cloudman melanoma cell proliferation byinsulin. Proc. Natl. Acad. Sci. USA 82, 1007–1011.
Fuller, B.B., Niekrasz, I., Hoganson, G.E., 1990. Down-regulation oftyrosinase mRNA levels in melanoma cells by tumor promoters and byinsulin. Mol. Cell Endocrinol. 72, 81–87.
Goodwin, P.J., Ennis, M., Pritchard, K.I., Trudeau, M.E., Koo, J.,Madranas, Y., Hartwick, W., Hoffman, B., Hood, N., 2002. Fastinginsulin and outcome in early-stage breast cancer: results of aprospective cohort study. J. Clin. Oncol. 20, 42–51.
Graeven, U., Herlyn, M., 1992. In vitro growth patterns of normal humanmelanocytes and melanocytes from different stages of melanomaprogression. J. Immunother. 12, 199–202.
Haring, H.U., White, M.F., Kahn, C.R., Kasuga, M., Lauris, V.,Fleischmann, R., Murray, M., Paewlek, J., 1984. Abnormality ofinsulin binding and receptor phosphorylation in an insulin-resistantmelanoma cell line. J. Cell Biol. 99, 900–908.
Isoldi, M.C., de Pina Benabou, M.H., Schumacher, R.I., Gois, C.C.,Scarparo, A.C., Rebouc�as, N.A., Visconti, M.A., 2004. Mechanismsof action of melanin-concentrating hormone in the teleost fish
erythrophoroma cell line (GEM-81). Gen. Comp. Endocrinol. 136,270–275.
Kahn, R., Murray, M., Pawelek, J., 1980. Inhibition of proliferationof Cloudman S91 melanoma cells by insulin and characterizationof some insulin-resistant variants. J. Cell Physiol. 103, 109–119.
Kim, S.K., Novak, R.F., 2007. The role of intracellular signaling ininsulin-mediated regulation of drug metabolizing enzyme gene andprotein expression. Pharmacol. Ther. 113, 88–120.
Matsumoto, J., Ishikawa, T., Masahito, P., Takayama, S., 1980. Perma-nent cell lines from erythrophoromas in goldfish (Carassius auratus). J.Natl. Cancer Inst. 64, 879–890.
Matsumoto, J., Ishikawa, T., Masahito, P., Takayama, S., Taylor, J.D.,Tchen, T.T., 1984. Clonal heterogeneity in physiological properties ofmelanized cells induced from goldfish erythrophoroma cell lines.Differentiation 27, 36–45.
Matsumoto, J., Lynch, T.J., Grabowski, S.M., Taylor, J.D., Tchen,T.T., 1982. Induction of melanized cells from a goldfish erythro-phoroma: isolation of pigment translocation variants. Science 217,1149–1151.
Mengeaud, V., Ortonne, J.P., 1994. Regulation of melanogenesis inducedby 5-methoxypsoralen without ultraviolet light in murine melanomacells. Pigment Cell Res. 7, 245–254.
Miralpeix, M., Sun, X.J., Backer, J.M., Myers Jr., M.G., Araki, E., White,M.F., 1992. Insulin stimulates tyrosine phosphorylation of multiplehigh molecular weight substrates in Fao hepatoma cells. Biochemistry31, 9031–9039.
Nery, L.E., Castrucci, A.M., 1997. Pigment cell signalling for physio-logical color change. Comp. Biochem. Physiol. A Physiol. 118,1135–1144.
Oka, M., Kageyama, A., Fukunaga, M., Bito, T., Nagai, H., Nishigori,C., 2004. Phosphatidylinositol 3-kinase/Akt-dependent and -indepen-dent protection against apoptosis in normal human melanocytes. J.Invest. Dermatol. 123, 930–936.
Pomerantz, S.H., 1969. L-tyrosine-3,5-3H assay for tyrosinase develop-ment in skin of newborn hamsters. Science 164, 838–839.
Rodeck, U., Melber, K., Kath, R., Menssen, H.D., Varello, M., Atkinson,B., Herlyn, M., 1991. Constitutive expression of multiple growth factorgenes by melanoma cells but not normal melanocytes. J. Invest.Dermatol. 97, 20–26.
Rowbottom, L.A., Whitehead, R.H., Roberts, G.P., Hughes, L.E., 1981.A study of the growth requirements of human tumour cells in tissueculture. Aust. J. Exp. Biol. Med. Sci. 59, 91–100.
Slominski, A., McNeely, T., Pawelek, J., 1992. Defect of insulin receptorin insulin-resistant variants of Cloudman S91 mouse melanoma cells.Melanoma Res. 2, 115–122.
Slominski, A., Tobin, D.J., Shibahara, S., Wortsman, J., 2004. Melaninpigmentation in mammalian skin and its hormonal regulation. Physiol.Rev. 84, 1155–1228.
Sun, X.J., Rothenberg, P., Kahn, C.R., Backer, J.M., Araki, E., Wilden,P.A., Cahill, D.A., Goldstein, B.J., White, M.F., 1991. Structure of theinsulin receptor substrate IRS-1 defines a unique signal transductionprotein. Nature 352, 73–77.
Sun, X.J., Wang, L.M., Zhang, Y., Yenush, L., Myers Jr., M.G.,Glasheen, E., Lane, W.S., Pierce, J.H., White, M.F., 1995. Roleof IRS-2 in insulin and cytokine signaling. Nature 377,173–177.
Tsatmali, M., Ancans, J., Thody, A.J., 2002. Melanocyte function and itscontrol by melanocortin peptides. J. Histochem. Cytochem. 50, 125–133.
White, M.F., Kahn, C.R., 1994. The insulin signaling system. J. Biol.Chem. 269, 1–4.
White, M.F., 1998. The insulin-signaling system: a network of dockingproteins that mediate insulin and cytokine action. Recent Prog. Horm.Res. 53, 119–138.
White, M.F., 2002. IRS proteins and the common path to diabetes. Am. J.Physiol. Endocrinol. Metab. 283, E413–E422.
Wu, Q., Brown, M.R., 2006. Signaling and function of insulin-likepeptides in insects. Ann. Rev. Entomol. 51, 1–24.
Copyright © 2022 FDOKUMEN




















