
Cell-type dependent response of melanoma cells to aloe emodin
10
(This is a sample cover image for this issue. The actual cover is not yet available at this time.) This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Cell-type dependent response of melanoma cells to aloe emodin

(This is a sample cover image for this issue. The actual cover is not yet available at this time.)
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Cell-type dependent response of melanoma cells to aloe emodin
J. Radovic a, D. Maksimovic-Ivanic a, G. Timotijevic b, S. Popadic c, Z. Ramic c, V. Trajkovic c, D. Miljkovic a,S. Stosic-Grujicic a, S. Mijatovic a,⇑a Department of Immunology, Institute for Biological Research ‘‘Sinisa Stankovic’’, University of Belgrade, Bulevar Despota Stefana 142, 11060 Belgrade, Serbiab Institute of Molecular Genetics and Genetic Engineering, Belgrade University, Vojvode Stepe 444a, 11000 Belgrade, Serbiac Institute of Microbiology and Immunology, School of Medicine, University of Belgrade, Dr. Subotica 1, 11000 Belgrade, Serbia
a r t i c l e i n f o
Article history:Received 3 May 2012Accepted 26 May 2012Available online 7 June 2012
Keywords:Aloe emodinMelanomaERK1/2AktApoptosisDifferentiation
a b s t r a c t
Intrinsic characteristics of melanoma cells such as expression of inducible nitric oxide synthase (iNOS),redox status, and activity of signaling pathways involved in proliferation, differentiation and cell deathdefine the response of the cells to the diverse treatments. In this context we compared the effectivenessof herbal antaquinone aloe emodin (AE) against mouse B16 melanoma and human A375, different in ini-tial activity of ERK1/2, constitutive iNOS expression and basal level of reactive oxygen species (ROS). Bothcell lines are sensitive to AE treatment. However, while the agent induces differentiation of B16 cellstoward melanocytes, in A375 cells promoted massive apoptosis. Differentiation of B16 cells, character-ized by enhanced melanin production and tyrosinase activity, was mediated by H2O2 production synchro-nized with rapid p53 accumulation and enhanced expression of cyclins D1 and D3. Caspase mediatedapoptosis triggered in A375 cells was accompanied with Bcl-2 but not iNOS down-regulation. In addition,opposite regulation of Akt-ERK1/2 axis in AE treated B16 and A375 cells correlated with different out-come of the treatment. However, AE in a dose-dependent manner rescued both B16 and A375 cells fromdoxorubicin- or paclitaxel-induced killing. These data indicate that caution is warranted when AE isadministrated to the patients with conventional chemotherapy.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Although naturally occurring anthracene derivatives present inthe extracts of the roots, bark or leaves of numerous plants wereprimary known as laxatives and are intensively used for that pur-pose until today, in last few decades anticancer properties of thesecompounds are in the focus of scientific interest (Mueller et al.,1998; Fullbeck et al., 2005; Huang et al., 2005; Kim et al., 2005;Mijatovic et al., 2005a; Su et al., 2005; Esmat et al., 2006; Leeet al., 2006; Lin et al., 2006). Indeed, numerous conventional cyto-static drugs are anthracyclin-type of molecules (Caponigro et al.,2005; Wu and Hasinoff, 2005). Most important natural hydrox-ianthraquinones with confirmed biological potential, aloe emodin(AE) and emodin, possess antibacterial, antifungal, antiviral, diure-tic, immunosuppressive, anti-inflammatory, vasorelaxant and,finally, anticancer activities (Huang et al., 1991; Barnard et al.,1992; Huang et al., 1992; Yagi et al., 1997; Kumar et al., 1998;
Mueller et al., 1998; Kim et al., 2001; Basu et al., 2005; Fullbecket al., 2005; Huang et al., 2005; Kim et al., 2005; Mijatovic et al.,2005a; Su et al., 2005; Esmat et al., 2006; Lee et al., 2006; Linet al., 2006; Tabolacci et al., 2010). AE, present in highest concen-tration in the cortex of Aloe vera leaves, induces cell death inseveral tumor cell lines, as well as the regression of neuroectoder-mal tumors in mice with severe combined immunodeficiencywithout appreciable signs of acute and chronic toxicity (Pecereet al., 2000; Lee et al., 2001; Kuo et al., 2002; Wasserman et al.,2002; Yeh et al., 2003; Mijatovic et al., 2004; Lin et al., 2006; Leeet al., 2006). Also there is no confirmed genotoxicity of the com-pound in vivo in healthy animals after the high acute exposure toAE when the drug concentrations in the blood were almost in thesame range as highly genotoxic concentration in vitro (Heidemannet al., 1996). The multiple mechanisms are involved in tumor cellgrowth inhibitory effect of AE. The outcome of the drug treatmenthas been described as induction of apoptosis (Acevedo-Duncanet al., 2004; Yeh et al., 2003; Lee et al., 2005; Lin et al., 2006), auto-phagic cell death Mijatovic et al., 2005a), or the cytostatic activitymanifested by cell cycle arrest at the different stages of celldivision (Chen et al., 2004; Lin et al., 2006).
Recently, Tabolacci et al. (2010) demonstrated the inhibitoryeffects of AE on B16-F10 melanoma cell proliferation, invasionand differentiation, accompanied by a remarkable increase of the
0278-6915/$ - see front matter � 2012 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.fct.2012.05.047
Abbreviations: AE, aloe emodin; ROS, reactive oxygen species; CFSE, carboxy-fluorescein diacetate succinimidyl ester; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; iNOS, inducible nitric oxide synthase; DOXO, doxo-rubicin; Pct, paclitaxel; F.I.C., fraction inhibitory concentration.⇑ Corresponding author. Tel.: +381 11 2078 390; fax: +381 11 2761 433.
E-mail address: [email protected] (S. Mijatovic).
Food and Chemical Toxicology 50 (2012) 3181–3189
Contents lists available at SciVerse ScienceDirect
Food and Chemical Toxicology
journal homepage: www.elsevier .com/locate / foodchemtox

Author's personal copy
activity of transglutaminase 2. Melanoma represent heterogeneousgroup of aggressive and high metastatic tumors (Hoang andEichenfield, 2000; Russo et al., 2009). One of the most seriousbarriers for their therapy is intrinsic resistance to induction ofapoptosis by natural mediators of immune response and chemo-therapeutic drugs (Hersey and Zhang, 2001). In the present studywe reported that the cell specific response is crucial for determina-tion of AE action. In fact, intracellular features of melanoma cellslike expression of iNOS, redox status, and activity of signaling path-ways crucial for proliferation, differentiation and cell death coulddefine the response of the cells to the treatment and even convertthe same treatment from cytostatic to cytotoxic (Demary et al.,2001; Tang and Grimm, 2004; Grimm et al., 2008). In this contextwe evaluated the molecular base of melanoma cell reaction to AEexposure and tried to explain why the same treatment in murineB16 cells resulted in cell differentiation while in human A375 linepromoted massive apoptosis. We confirmed that basal level ofreactive oxygen species (ROS) as well as opposite regulation ofAkt-ERK1/2 axis in AE treated B16 and A375 cells is associated withthe outcome of the treatment.
2. Materials and methods
2.1. Reagents, cells and animals
All chemicals used in the experiments were purchased from Sigma (St. Louis,USA), unless specified otherwise. Mayer’s hematoxylin was from Bio Optica (Milan-o, Italy). Carboxyfluorescein diacetate succinimidyl ester (CFSE) and 20 ,70-dichloro-dihydrofluorescein diacetate (H2DCFDA) were from Molecular Probes (Eugene,USA). AE was stored at �20 �C at concentration of 200 lM in DMSO, and it was di-luted in culture medium immediately before use. Murine melanoma B16 cell linewas a kind gift from Dr. Sinisa Radulovic (Institute for Oncology and Radiology ofSerbia, Belgrade, Serbia) while A375 cells were kind gift from Prof Ferdinando Nic-oletti (Department of Biomedical Sciences, University of Catania, Italy). Cells weregrown in HEPES-buffered RPMI 1640 medium supplemented with 5% or 10% FCS,2 mM L-glutamine, antibiotics and 0.01% sodium pyruvate (culture medium) at37 �C in humidified atmosphere with 5% CO2. Cells were used for experiments afterconventional trypsinization procedure. Control cell cultures contained the amountof DMSO corresponding to its content in the solution with the highest concentrationof AE used in the particular experiment.
Inbred C57BL/6 mice were obtained from our own facility at the Institute forBiological Research ‘‘Sinisa Stankovic’’, University of Belgrade (Belgrade, Serbia)and kept under standard laboratory conditions (non specific pathogen free) withfree access to food and water. The handling of animals and the study protocol werein accordance to the requirements of the European Union and approved by theInstitutional Animal Care and Use Committee at the Institute for Biological Research‘‘Sinisa Stankovic’’, University of Belgrade.
2.2. MTT-test and CFSE staining
The cells were seeded in flat-bottom 96-well plates (104cells/well) overnight,and then treated with different concentration of AE as indicated. After 24 h incuba-tion, 3–4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) testwas performed as described previously (Mijatovic et al., 2005a). The results ofMTT test, obtained from triplicate cultures were presented as % of control values ob-tained in untreated cell cultures. Rate of cell proliferation was determined usingflow cytometric analysis of cells labeled with CFSE. Briefly, cells were detachedand loaded with 1.5 lM CFSE for the 15 min at 37 �C, washed two times and thenseeded in 6-well plates. Cells were treated for the 24 and 48 h, trypsinized andwashed two times. Finally, cells were resuspended in PBS and analyzed by flowcytometry. Green fluorescence emission from cells illuminated with excitation lightof 488 nm was measured with a FACSCalibur (BD, Heidelberg, Germany) and ana-lyzed using Cell Quest Pro software (BD).
2.3. Determination of apoptosis and cell cycle analysis
Cells (2.5 � 105/well) were treated with 40 lM of AE in 6-well plate for 24 h,then trypsinized and stained with AnnV-FITC/EtBr according to the manufac-turer’s instruction (Biotium, Hayward, CA). Altermativly, cell cycle analysis wasperformed by propidium iodide (PI) staining as previously described (Mijatovicet al., 2005a; Maksimovic-Ivanic et al., 2008). Cells were analysed with FACSCal-ibur flow cytometer (BD, Heidelberg, Germany) using Cell Quest Pro software(BD).
2.4. Staining of cells with Mayer’s hematoxylin
Cells were grown (3 � 104/well) in a chamber slide for 24 h in the presence orabsence of AE or 0.5 lM of H2O2. Subsequently, they were fixed in 4% paraformal-dehyde for 20 min at RT, washed twice in PBS and stained with Mayer’s hematoxilinfor 30 s. Finally, cells were washed with tap water, mounted with glycergel and ana-lyzed under light microscope.
2.5. Staining of cells with H2DCFDA and DAB
Cells were stained with H2DCFDA according to the manufacturer’s instructionfor 30 min and then treated with AE (40 lM). After 5, 15, 30 and 45 min cells werewashed and presence of reactive oxygen species was detected by flow cytometry.
Cells were exposed to AE (40 lM) in parallel with 3,30-Diaminobenzidine (DAB,1 mg/ml) and appearance of black precipitates was evaluated by light microscopy.
2.6. Tyrosinase activity assay and melanin determination
Tyrosinase activity was determined by measuring the rate of oxidation of L-dopa. Briefly, sub-confluent cultures in 6-well plate were lysed in 100 ll phosphatebuffer pH 6.8–1% Triton X-100 and than centrifuged at 10000 rpm for 5 min. 40 llof each extract was mixed with 100 ll of L-dopa substrate solution (2 mg/ml). Enzy-matic reaction was carried out at 37 �C and the absorbance at 570 nm was readevery 10 min for at least 1 h. The final activity was corrected by the total amountof protein estimated by Bradford assay. For melanin determination cells were incu-bated in 6-well plate for 24 h, trypsinized, counted and than lysed in 100 ll of 1 MNaOH. 400 ll of distilled water was added and samples were incubated at 60 �C for1 h. Thereafter, absorbance of dissolved dye was measured at 492 nm.
2.7. In vivo administration of B16 cells
For in vivo application, B16 cells were cultivated for 24 h in the presence of40 lM AE or adequate amount of DMSO, then trypsinized, counted and finallyresuspended in PBS. Primary tumors were induced by subcutaneous (s.c.) injectionof 5 � 105 melanoma cells in the dorsal right lumbosacral region of C57BL/6 mice.Tumor growth was observed every day and tumor size was measured in threedimensions. Tumor volume was calculated by the following formula: [0.52 ab2](Maksimovic-Ivanic et al., 2008), where a is the longest and b is the shortestdiameter.
2.8. Caspase detection
Cells (2.5 � 105/well) were treated with 40 lM AE for 24 h, then trypsinized andstained with Apostat (R&D Systems, Minneapolis, MN) in phenol red free RPMI 1640(5% FCS) for 15 min at 37 �C. At the end of cultivation cells were washed and ana-lysed with FACSCalibur flow cytometer (BD, Heildelberg, Germany) using Cell QuestPro software (BD).
2.9. Immunoblot analysis of AKT, p-ERK, p53, cyclin D1 and D3, Bcl-2, iNOS and IjB
Cells (1 � 106) were seeded in flasks (25 cm3), and treated with 40 lM AE forthe indicated time intervals. At the end of incubation, cells were harvested in pro-tein lysis buffer containing 62.5 mM Tris–HCl (pH 6.8 at 25� C), 2% w/v SDS, 10%glycerol, 50 mM DTT, and 0.01% w/v bromophenol blue, and subjected to electro-phoresis on 12% SDS–polyacrylamide gels or 10% SDS–polyacrylamide gels fordetection of iNOS. Electro-transfer to polyvinylidene difluoride membranes at5 mA/cm2 was done with a semi-dry blotting system (Fastblot B43, Biorad, Goettin-gen, Germany). The membranes were blocked with 5% w/v nonfat dry milk in PBSwith 0.1% Tween-20, and blots were probed with specific antibodies to pIkB, p53,cyclin D1, cyclin D3 and b-actin (all from Santa Cruz Biotechnology, Santa Cruz,CA), iNOS, (Sigma, St. Louis, MO), AKT, p-AKT, ERK, p-ERK, Bcl-2 (all from Cell Sig-naling Technology), followed by incubation with secondary antibody (ECL donkeyanti-rabbit HRP linked, GE Healthcare, Buckinghamshire, UK). Bands were visual-ized using chemiluminescence detection system (ECL, GE Healthcare).
2.10. Isobologram analysis
To determine the type of interaction between AE and conventionally used che-motherapeutic isobologram analysis was performed. Isobolograms were made fromtreatments with diverse concentrations of AE (40, 20, 10 lM) combined with differ-ent concentrations of doxorubicin (DOXO) 0.125–1 lM or 0.015–0.1 lM for A375and B16, respectively, and with paclitaxel (Pct) 3.7–25 lM and 0.38–3 lM forA375 and B16, respectively. Combinations attaining 30–50% of cytotoxicity wereexpressed as concentration of single agent alone making this amount of toxicity.Analysis was prepared on the basis of dose–response curves of cell viability treatedwith AE alone, Pct or DOXO alone or their combination for 24 h. Fraction inhibitoryconcentration (F.I.C.): concentration of each agent in combination/concentration ofeach agent alone <1 is considered synergistic while F.I.C. > 1 is consideredantagonistic.
3182 J. Radovic et al. / Food and Chemical Toxicology 50 (2012) 3181–3189

Author's personal copy
2.11. Statistical analysis
The significance of the differences between various treatments was analyzed byANOVA followed by Student–Newman-Keuls test or Mann–Whitney U-test forin vivo experiments. A p value less than 0.05 was considered significant.
3. Results
3.1. AE reduces the growth of B16 and A375 cells
To evaluate the anti-melanoma capacity of AE in vitro, B16 andA375 cells were incubated in the presence of various AE concentra-tions, and tumor cell viability was determined after 24 h by MTTtest. Mitochondrial respiration in both cell lines exposed to thedrug was dose-dependently reduced in comparison to untreatedcontrol (Fig. 1(a)). Cell cycle analysis performed after 18 h of treat-ment with AE showed more pronounced cell cycle arrest in G2/Mphase in B16 cells in comparison to A375 cells (Fig. 1(b)). In addi-tion, Ann/PI staining revealed the absence of apoptotic B16 cellseven after 48 h of incubation with AE (Fig. 1(c)). In the same time-frame, treatment of A375 cells with AE resulted in exaggeratedaccumulation of early and also late apoptotic cells. Light micro-scopic analysis of B16 cells in the presence of AE revealed that ab-sence of apoptosis correlated with cellular morphological changes(Fig. 1(d)). Compared to untreated cells with polygonal shape, cellsincubated with AE displayed lopsided shape with increasedvolume of cytoplasm and membrane outgrowths resembling den-drites. On the other hand, numerous rounded apoptotic cells wereobserved upon the treatment of A375 cells with AE (Fig. 1(d)).These results suggest that besides obvious anti-cancer potentialof AE against both cell lines tested, the route of AE action is quitedifferent between them.
3.2. AE promotes differentiation of B16 cells
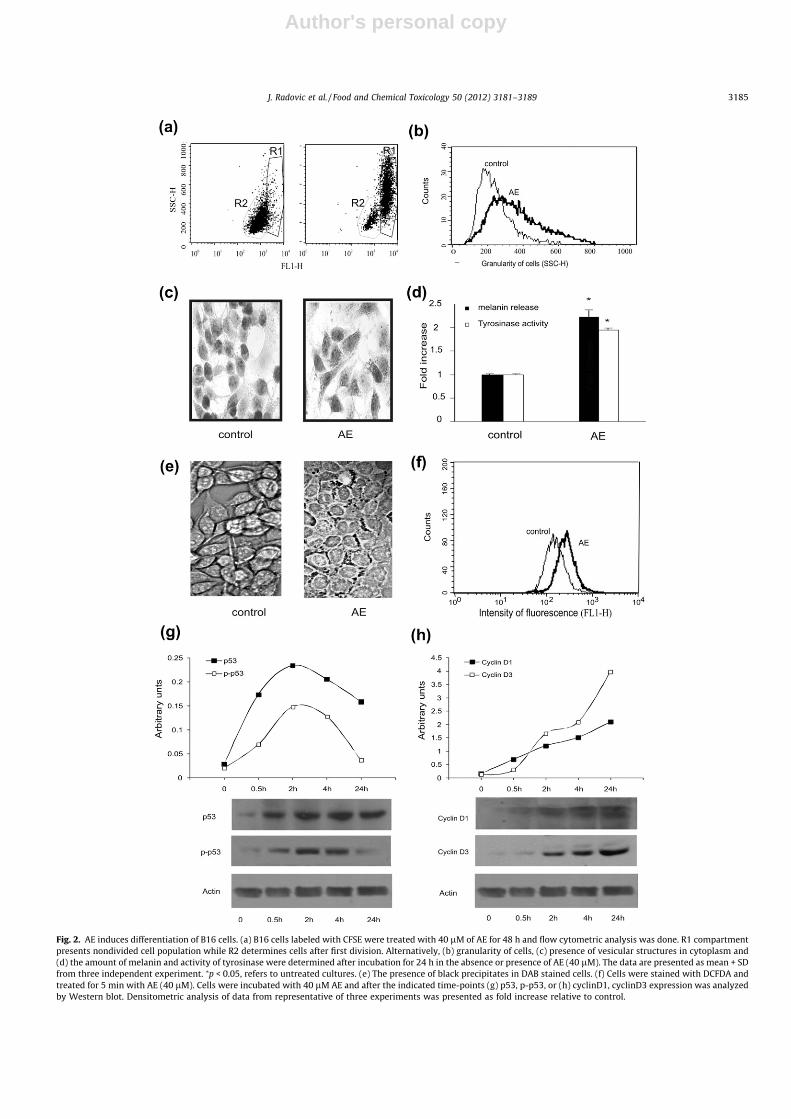
The morphological transformation of cells, as well as G2M blockwithout apoptosis induction, implied that B16 cells underwent toprocess opposite to malignant transformation. To further examinethe anti-proliferative activity of AE we performed cytofluorimetricanalysis of CFSE-stained cells. Compared to control untreated cells,the percentage of the divided cells after 48 h of incubation in thepresence of AE was significantly lower (95% in untreated cells vs.15% in AE treated cells) (Fig. 2(a)). Concordantly with this andthe previously described data (Fig. 1(d)), the appearance of in-creased granularity in cytoplasm of melanoma cells was detectedby flow cytometric analysis (Fig. 2(b)). In contrast to small vesiclesevident in control untreated cells (Fig. 2(c)), large vesicles spread-ing from perinuclear area to growing dendrites of AE-treated cellswere readily detectable upon staining with Mayer’s haematoxylin(Fig. 2(c)). To further examine whether the morphological transfor-mation of B16 melanoma cells was accompanied by the inductionof melanogenesis, we assessed both melanin synthesis and tyrosi-nase activity. Compared to control cells, the amount of melanin re-leased from the lysed cells was significantly elevated in culturesexposed to the drug for 24 h (Fig. 2(d)). In addition, AE treatmentinduced twofold increase in tyrosinase activity in B16 melanomacells (Fig. 2(d)). These data confirmed previously described differ-entiation of B16 cells toward melanocytes upon the treatment withAE (Tabolacci et al., 2010). Having in mind the described role ofH2O2 in differentiation of different types of cells, as well as the factthat this molecule is one of the products of melanin metabolism,we next investigated its production in cells exposed to AE. Theaddition of DAB led to appearance of black precipitates, furtherconfirming previous findings (Fig. 2(e)). Treatment of cells with0.5 lM H2O2 resulted in the appearance of granular structures inthe cytoplasm of cells mimicking the effect of AE (not shown).
Moreover, staining of cells by DCFDA showed a remarkable in-crease in H2O2 production, detected in cells even 5 min upon addi-tion of the drug (Fig. 2(f)). Moreover, Western blot analysisrevealed that process of differentiation was accompanied withtransient up-regulation of total p53 which correlated with theinhibited proteolytic degradation of this molecule, as judged bythe synchronized time-kinetics of the p-p53 form phosphorylatedon Ser 20 (Fig. 2(g)). In parallel, AE treatment significantly elevatedthe expression of cyclins D1 and D3 (Fig. 2(h)). Taken together, allthese data clearly indicated that AE reversed melanoma B16 cellsinto non-malignanat phenotype. In order to estimate whether theloss of malignant potential by the treatment with AE is permanent,B16 cells were incubated for 24 h with drug, and then injected intosyngeneic C57BL/6 mice. Animals were sacrificed on day 30, tu-mors isolated and their size was measured. Mice injected withB16 cells pretreated with AE (AE-B16) showed significant delayin tumor appearance (14 vs. 24 day post-tumor challenge in con-trol and AE-B16, respectively) and dramatic decrease in tumor vol-ume compared to the animals that received vehicle-treated B16cells (1.5 + 0.71 vs. 0.4 ± 0.36 in control and AE-B16, respectively).In summary, it is evident that in vitro pretreatment of B16 cellswith AE dramatically reduced their malignant potential in vivo.
3.3. AE induces caspase dependent apoptosis of A375 cells
To clarify the mechanism underlying the apoptosis of A375 cellstriggered by AE, total caspase activity was estimated. Treatmentwith AE resulted in the intensified caspase activity in tested cells(Fig. 3(a)). Having in mind that in melanoma cells anti-apoptoticsignal is dominant and responsible for their resistance to therapy,Bcl-2 protein expression was analyzed. Apoptosis triggered by AEwas followed with the early decrease of cytoprotective Bcl-2(Fig. 3(b)). In the light of the fact that A375 cells constitutively ex-press iNOS which is essential for they growth (Grimm et al., 2008),and that AE is able to inhibit iNOS (Mijatovic et al., 2004) we nextinvestigated iNOS protein expression, as well as protein expressionof one of its most important regulators- NFjB. The expression ofiNOS was not affected during the 12 h of treatment (Fig. 3(c)).Moreover, the expression of p-IjB, as the parameter of NF-jBactivity was also not changed during this period of cultivation withAE (Fig. 3(c)), confirming that AE prompted apoptosis throughdown-regulation of cytoprotective molecule Bcl-2 rather then iNOSexpression.
3.4. AE oppositely regulates Akt and ERK1/2 activity in B16 and A375cells
To explore the molecular basis of the anti-melanoma effect ofAE, B16 and A375 cells were treated with AE and Akt and ERK1/2activation were estimated by Western blot at different time points.As seen in Fig. 4, the relative expression of p-ERK in B16 cells wasamplified in time-dependent manner while the same treatment inA375 cells caused dramatic inhibition of ERK even after 30 min ofexposure. In parallel, the expression of Akt was in negative corre-lation with ERK in both cell lines. Whereas in B16 cells the rateof Akt phosphorylation on Ser 473 was significantly diminished,in A375 cells it was markedly elevated. Taken together, these datashow that different mechanisms of AE antitumor action in A375and B16 cells are associated with opposite regulation of thesetwo signaling pathways.
3.5. AE antagonizes conventional chemotherapy
The inhibited proliferation in B16 cells as well as decreased le-vel of anti-apoptotic signals in A375 cells raised a possibility thattreatment with AE could have chemo-sensitizing potential. For that
J. Radovic et al. / Food and Chemical Toxicology 50 (2012) 3181–3189 3183

Author's personal copy
Fig. 1. AE reduces the growth of B16 and A375 melanoma cells. (a) B16 and A375cells (104/well) were treated with different concentrations of AE. After 24 h of cultivation cellviability was determined by MTT. The data are presented as mean ± SD from representative of three independent experiments performed in triplicate. ⁄p < 0.05, refers tountreated cultures. After the 18 and 48 h of treatment with 40 lM AE cell cycle analysis (b) and AnnV/PI (c) double staining were done, respectively. (d) Morphology of cellswas analyzed by light microscopy after incubation for 48 h in the absence or presence of AE (40 lM).
3184 J. Radovic et al. / Food and Chemical Toxicology 50 (2012) 3181–3189
Author's personal copy
Fig. 2. AE induces differentiation of B16 cells. (a) B16 cells labeled with CFSE were treated with 40 lM of AE for 48 h and flow cytometric analysis was done. R1 compartmentpresents nondivided cell population while R2 determines cells after first division. Alternatively, (b) granularity of cells, (c) presence of vesicular structures in cytoplasm and(d) the amount of melanin and activity of tyrosinase were determined after incubation for 24 h in the absence or presence of AE (40 lM). The data are presented as mean + SDfrom three independent experiment. ⁄p < 0.05, refers to untreated cultures. (e) The presence of black precipitates in DAB stained cells. (f) Cells were stained with DCFDA andtreated for 5 min with AE (40 lM). Cells were incubated with 40 lM AE and after the indicated time-points (g) p53, p-p53, or (h) cyclinD1, cyclinD3 expression was analyzedby Western blot. Densitometric analysis of data from representative of three experiments was presented as fold increase relative to control.
J. Radovic et al. / Food and Chemical Toxicology 50 (2012) 3181–3189 3185

Author's personal copy
purpose, both cell lines were exposed to conventional therapeuticsdoxorubicine and paclitaxel in the presence of AE (Fig. 5). Therange of doses was determined for each cell line. Surprisingly, inboth cell lines co-treatment with AE and chemotherapeutic drugssignificantly antagonized the activity of the latter, thus indicatingthat AE acted in a cyto-protective manner in the presence of othertoxic stimuli.
4. Discussion
In the present study we have provided evidence for the strongcytostatic and cytotoxic activity of herbal pigment AE against
B16 and A375 melanoma cells, respectively. The cytostatic effectin B16 cells could be attributed to differentiation-inducing prop-erty of the drug through ROS production, while the observed cyto-toxic activity in A375 cells is due to a decrease in thecytoprotective anti-apoptotic molecules which promoted switchto apoptotic phenotype.
We and others have shown that AE down-regulated growth ofhepatoma, fibrosarcoma, neuroblastoma, astrocytoma, lung carci-noma, leukemia, bladder carcinoma, Merkel cell lines and mela-noma B16/F10 cells (Pecere et al., 2000; Lee et al., 2001; Leeet al., 2006; Kuo et al., 2002; Wasserman et al., 2002; Yeh et al.,2003; Mijatovic et al., 2004, 2005a; Lin et al., 2006; Tabolacciet al., 2010). The tumoricidal activity of naturally occurring com-pounds and numerous chemotherapeutics is usually ascribed tothe induction of apoptotic or autophagic cell death (Mijatovicet al., 2005a). The anti-neuroblastoma effect of AE was explainedby apoptotic cell death promoted by specific energy-dependentpathway of the drug incorporation (Pecere et al., 2000). Our studiesof tumoricidal action of AE on astrocytoma C6 cells suggested theimportance of programmed cell death type II, known as autophagiccell death (Mijatovic et al., 2005a). In concordance with previouslymentioned data, AE treatment induced massive apoptosis of A375cells. Apoptotic process coincided with the augmented activity ofcaspases and diminished expression of the cyto-protective mole-cule Bcl-2. The effect of AE on Bcl-2 expression has previously beenshown (Chiu et al., 2009). In addition to these reports, AE is de-scribed as a differentiation inducing agent. Our results confirmedrecently published data by Tabolacci et al. (2010) that AE triggereddifferentiation process in B16/F10 cells accompanied with reduc-tion of their metastatic potential. Diverse responses of B16 andA375 cells to various stimuli are well documented (Demary et al.,2001). We therefore attempted to evaluate the mechanisms under-lying the different responses of the two melanoma cell lines to AEtreatment. It has previously been reported that A375 cells constitu-tively produce NO which is their endogenous growth factor(Grimm et al., 2008). Thus, its elimination would be negatively re-flected on melanoma expansion. Constitutive expression of iNOSand the fact that they have high intracellular ROS levels makeA375 cells more resistant to oxidative stress and determine the re-sponse to agents which mode of action is related to induction ofROS and RNS (Gruber et al., 1992; Demary et al., 2001). On theother hand, B16 cells are known as low ROS producing cells. Hence,the outcome of retinoic acid (RA) treatment in those cells is differ-ent (Gruber et al., 1992; Demary et al., 2001). In fact, A375 cellswere resistant to RA treatment, while B16 cells entered the differ-entiation process toward melanocytes, ascribed to H2O2 produc-tion, increased expression of aPKC, and stimulated activity ofERK and transcription factor AP1. Similarly, treatment with AE ini-tiated the phenotypical transformation accompanied with elevatedtyrosinanse activity and consequently up-regulated melanine pro-duction. As judged by the diminished potential of AE-treated B16cells to induce tumor in C57BL/6 mice, it seems that conversionof malignant toward primary phenotype was permanent. The lostof proliferative potential was accompanied with significant intra-cellular changes, such as up-regulation of cyclin D type and tumorsuppressor p53. Cyclin D1 has the affinity for the proliferating cellnuclear antigen and thus inhibits synthesis of DNA, inducing cellcycle arrest similar to one observed upon the AE treatment(Ullmannová et al., 2003; Alao, 2007). Cyclin D3 is described as akey regulator in oligodendrocyte differentiation, where togetherwith p53 regulates the transcription of genes involved in this pro-cess (Tokumoto et al., 1999; Billon et al., 2004). We found that AEinduced fast accumulation of total p53, as well as phosphorylatedform of this protein on Ser 20 in B16 cells. Having in mind thatphosphorylation on this position protected p53 molecule frommdm- mediated degradation it seems that enhanced p53
Fig. 3. AE induces caspase dependent apoptosis in A375 cells. (a) Cells wereincubated in the presence of 40 lM AE and pancaspase activity was assessed. (b, c)Cells were incubated with 40 lM AE for indicated time and Bcl-2, iNOS and pIjBexpression was analyzed by Western blot. Densitometric analysis of data fromrepresentative of three experiments was presented as fold increase relative tocontrol.
3186 J. Radovic et al. / Food and Chemical Toxicology 50 (2012) 3181–3189

Author's personal copy
expression is rather the consequence of decreased degradationthen de novo synthesis (Chehab et al., 1999). In addition, fast accu-mulation of p53 correlated with arrest of cells in G2/M phase of thecell cycle. As previously mentioned, ROS production upon AE treat-ment could be considered as initial trigger for the differentiation. Itwas documented that H2O2 induced differentiation of many celltypes (Steinbeck et al., 1998; Kang et al., 2003; Xiao et al., 2009).Staining with DCFDA dye confirmed that AE provoked rapid pro-duction of peroxy radicals, while exposure of cells to DAB led toformation of dark precipitates indicating the activity of peroxydaseenzyme. Corresponding to this, treatment of cells with H2O2 in-duced morphological changes similar to AE (not shown). Strong
activation of ERK1/2 in AE-exposed B16 cells could also be the con-sequence of H2O2 generation. Increased activation of ERK is con-nected to RA-induced differentiation in the same melanoma cellline (Gruber et al., 1992; Demary et al., 2001). In parallel, the sametreatment did not induce ROS production in A375 cells and accord-ingly, relevant intracellular events were different from those ob-served in B16 cells. Oppositely to B16 cells, dramaticallyinhibited ERK in A375 cells upon AE exposure could be the causeof the detected apoptosis. Surprisingly, ERK inhibition did not re-flect to iNOS expression, excluding the importance of iNOS activityfor tumoricidal effect of AE. The ability of AE to abrogate ERKexpression was described previously and the drug was discussed
Fig. 4. AE differentially regulates p-Akt and ERK1/2 expression. Cells were incubated with 40 lM AE for indicated time and p-Akt and ERK1/2 expression was analyzed byWestern blot. Densitometric analysis of data from representative of three experiments was presented as fold increase relative to control.
J. Radovic et al. / Food and Chemical Toxicology 50 (2012) 3181–3189 3187

Author's personal copy
as eventual ERK inhibitor (Mijatovic et al., 2005a,b). The presentstudy clearly shows that this phenomenon is not explicitly a fea-ture of AE, but could be influenced by cell specificity. The highERK expression in A375 cells could be related to elevated expres-sion of PTK receptors. Similarly, in C6 astrocytoma and L929 fibro-sarcoma cells ERK inhibition was shown as a basic mechanism oftumoricidal activity of AE (Mijatovic et al., 2005a,b). In melanomacells tested here, ERK and Akt were reciprocally regulated. We havepreviously reported this relation in A375 cell exposed to HIV pro-tease inhibitor, saquinavir, and its NO modified form (Mijatovicet al., 2011). In addition, the enhanced activity of Akt could bethe consequence of mTORC2 release from S6K suppression(Delgoffe et al., 2011; Fenton and Gout, 2011), indicating possibil-ity that downstream rather than upstream part of PI3-Akt pathwayis affected by AE. This speculation is supported by cell cycle arrest,decreased expression of Bcl-2 and late degradation of actin(unpublished data) in A375 cells, which are all under S6K control(Fenton and Gout, 2011). However, the key step in cascade ofevents triggered by AE leading to different responses of A375 andB16 is still unknown.
Although AE targeted many signaling pathways involved in cellresponse to therapy it was of interest to investigate the effect of AEon the activity of chemotherapeutic drugs. Combined treatment ofAE and paclitaxel or doxorubicin resulted in weakening of thedrugs’ effects in both lines tested independently from differentintracellular response to AE. The absence of the expected antraqui-none mediated sensitization to chemotherapy already described innumerous cell lines such as Merkel cell carcinoma, esophageal car-cinoma, HER-2/neu-overexpressing breast cancer cells etc., re-vealed the complexity of relevant signaling pathways andpossible interactions between them in response to both agents(Zhang et al., 1999; Fenig et al., 2004; Yang et al., 2004). The signal-ing network triggered by the therapy is basically defined by the cell
specificity rather than predictable way of drug action, especially incombined treatments. Following our previous data about AE poten-tial to neutralize cisplatin toxicity in rat astrocytoma and mousefibrosarcoma (Mijatovic et al., 2005b), as well as the potential toabrogate apoptosis induced by TNF in the same cell line, (Harhajiet al., 2007), it could be concluded that naturally occurring com-pounds such as AE have intrinsic potential of to be cyto-protectivein the circumstances when cells are exposed to other toxic stimuli.
Having in mind our and numerous literature data, it is clear thatanti-tumor potential of this plant compound is undoubted but itmust be evaluated in the context of the tumor cell specificity.Application of AE could be undesirable and even dangerous iftumor is already exposed to chemotherapeutic drugs.
Conflict of Interest
The authors are not aware of any conflicts of interest.
Acknowledgement
This work was supported by the funds of The Ministry of Edu-cation and Science of the Republic of Serbia (GrantNo. 173013).
References
Acevedo-Duncan, M., Russell, C., Patel, S., Patel, R., 2004. Aloe-emodin modulatesPKC isozymes, inhibits proliferation, and induces apoptosis in U-373MG gliomacells. Int. Immunopharmacol. 4, 1775–1784.
Alao, J.P., 2007. The regulation of cyclin D1 degradation: roles in cancerdevelopment and the potential for therapeutic invention. Mol. Cancer 6, 24.
Barnard, D.L., Huffman, J.H., Morris, J.L., Wood, S.G., Hughes, B.G., Sidwell, R.W.,1992. Evaluation of the antiviral activity of anthraquinones, anthrones andanthraquinone derivatives against human cytomegalovirus. Antiviral Res. 17,63–77.
Fig. 5. AE attenuates the effectiveness of chemotherapy. B16 (upper panel) and A375 cells (lower panel) (104/well) were treated with of 10, 20 and 40 lM of AE in parallelwith DOXO or Pct. After 24 h of cultivation cell viability was determined by MTT and isobologram curves from representative of three independent experiments werepresented. (F.C.I. >1 is considered antagonistic).
3188 J. Radovic et al. / Food and Chemical Toxicology 50 (2012) 3181–3189

Author's personal copy
Basu, S., Ghosh, A., Hazra, B., 2005. Evaluation of the antibacterial activity ofVentilago madraspatana Gaertn. Rubia cordifolia Linn. and Lantana camara Linn.:isolation of emodin and physcion as active antibacterial agents. Phytother. Res.19, 888–894.
Billon, N., Terrinoni, A., Jolicoeur, C., McCarthy, A., Richardson, W.D., Melino, G., Raff,M., 2004. Roles for p53 and p73 during oligodendrocyte development.Development 131, 1211–1220.
Caponigro, F., Basile, M., de Rosa, V., Normanno, N., 2005. New drugs in cancertherapy, National Tumor Institute, Naples, 17–18 June 2004. Anticancer Drugs16, 211–221.
Chehab, N.H., Malikzay, A., Stavridi, E.S., Halazonetis, T.D., 1999. Phosphorylation ofSer-20 mediates stabilization of human p53 in response to DNA damage. Proc.Natl. Acad. Sci. USA 96, 13777–13782.
Chen, H.C., Hsieh, W.T., Chang, W.C., Chung, J.G., 2004. Aloe-emodin induced in vitroG2/M arrest of cell cycle in human promyelocytic leukemia HL-60 cells. FoodChem. Toxicol. 42, 1251–1257.
Chiu, T.H., Lai, W.W., Hsia, T.C., Yang, J.S., Lai, T.Y., Wu, P.P., Ma, C.Y., Yeh, C.C., Ho,C.C., Lu, H.F., Wood, W.G., Chung, J.G., 2009. Aloe-emodin induces cell deaththrough S-phase arrest and caspase-dependent pathways in human tonguesquamous cancer SCC-4 cells. Anticancer Res. 29, 4503–4511.
Delgoffe, G.M., Pollizzi, K.N., Waickman, Heikamp, E., Meyers, D.J., Horton, M.R.,Xiao, B., Worley, P.F., Powell, J.D., 2011. The kinase mTOR regulates thedifferentiation of helper T cells through the selective activation of signaling bymTORC1 and mTORC2. Nat. Immunol. 12, 295-303.
Demary, K., Wong, L., Liou, J.S., Faller, D.V., Spanjaard, R.A., 2001. Redox control ofretinoic acid receptor activity: a novel mechanism for retinoic acid resistance inmelanoma cells. Endocrinology 142, 2600–2605.
Esmat, A.Y., Tomasetto, C., Rio, M.C., 2006. Cytotoxicity of a natural anthraquinone(Aloin) against human breast cancer cell lines with and without ErbB-2:topoisomerase IIalpha coamplification. Cancer Biol. Ther. 5, 97–103.
Fenig, E., Nordenberg, J., Beery, E., Sulkes, J., Wasserman, L., 2004. Combined effect ofaloe-emodin and chemotherapeutic agents on the proliferation of an adherentvariant cell line of Merkel cell carcinoma. Oncol. Rep. 11, 213–217.
Fenton, T.R., Gout, I.T., 2011. Functions and regulation of the 70 kDa ribosomal S6kinases. Int. J. Biochem. Cell. Biol. 43, 47–59.
Fullbeck, M., Huang, X., Dumdey, R., Frommel, C., Dubiel, W., Preissner, R., 2005.Novel curcumin- and emodin-related compounds identified by in silico 2D/3Dconformer screening induce apoptosis in tumor cells. BMC Cancer 5, 97.
Grimm, E.A., Ellerhorst, J., Tang, C.H., Ekmekcioglu, S., 2008. Constitutiveintracellular production of iNOS and NO in human melanoma: possible role inregulation of growth and resistance to apoptosis. Nitric Oxide 19, 133–137.
Gruber, J.R., Ohno, S., Niles, R.M., 1992. Increased expression of protein kinase Calpha plays a key role in retinoic acid-induced melanoma differentiation. J. Biol.Chem. 267, 13356–13360.
Harhaji, L., Mijatovic, S., Maksimovic-Ivanic, D., Popadic, D., Isakovic, A., Todorovic-Markovic, B., Trajkovic, V., 2007. Aloe emodin inhibits the cytotoxic action oftumor necrosis factor. Eur. J. Pharmacol. 568, 248–259.
Heidemann, A., Volkner, W., Mengs, U., 1996. Genotoxicity of aloe emodin in vitroand in vivo. Mutat. Res. 367, 123–133.
Hersey, P., Zhang, X.D., 2001. How melanoma cells evade trail-induced apoptosis.Nat. Rev. Cancer 1, 142–150.
Hoang, M.T., Eichenfield, L.F., 2000. The rising incidence of melanoma in childrenand adolescents. Dermatol. Nurs. 12, 188–189.
Huang, H.C., Chang, J.H., Tung, S.F., Wu, R.T., Foegh, M.L., Chu, S.H., 1992.Immunosuppressive effect of emodin, a free radical generator. Eur. J.Pharmacol. 211, 359–364.
Huang, H.C., Lee, C.R., Chao, P.D., Chen, C.C., Chu, S.H., 1991. Vasorelaxant effect ofemodin, an anthraquinone from a Chinese herb. Eur. J. Pharmacol. 205, 289–294.
Huang, Q., Shen, H.M., Ong, C.N., 2005. Emodin inhibits tumor cell migrationthrough suppression of the phosphatidylinositol 3-kinase-Cdc42/Rac1 pathway.Cell. Mol. Life Sci. 62, 1167–1175.
Kang, H.K., Suh, J.H., Lee, J.J., Yoon, S.H., Hyun, J.W., Choi, S.W., Choi, J.Y., Ryu, K.H.,Chung, M.H., 2003. Induction of the differentiation of HL-60 promyelocyticleukemia cells by L-ascorbic acid. Free Radic. Res. 37, 773–779.
Kim, M.S., Park, M.J., Kim, S.J., Lee, C.H., Yoo, H., Shin, S.H., Song, E.S., Lee, S.H., 2005.Emodin suppresses hyaluronic acid-induced MMP-9 secretion and invasion ofglioma cells. Int. J. Oncol. 27, 839–846.
Kim, Y.M., Lee, C.H., Kim, H.G., Lee, H.S., 2004. Anthraquinones isolated from Cassiatora (Leguminosae) seed show an antifungal property against phytopathogenicfungi. J. Agric. Food Chem. 52, 6096–6100.
Kumar, A., Dhawan, S., Aggarwal, B.B., 1998. Emodin (3-methyl-1, 6, 8-trihydroxyanthraquinone) inhibits TNF-induced NF-kappaB activation,IkappaB degradation, and expression of cell surface adhesion proteins inhuman vascular endothelial cells. Oncogene 17, 913–918.
Kuo, P.L., Lin, T.C., Lin, C.C., 2002. The antiproliferative activity of aloe-emodin isthrough p53-dependent and p21-dependent apoptotic pathway in humanhepatoma cell lines. Life Sci. 71, 1879–1892.
Lee, H.Z., Hsu, S.L., Liu, M.C., Wu, C.H., 2001. Effects and mechanisms of aloe-emodinon cell death in human lung squamous cell carcinoma. Eur. J. Pharmacol. 431,287–295.
Lee, H.Z., Lin, C.J., Yang, W.H., Leung, W.C., Chang, S.P., 2006. Aloe-emodin inducedDNA damage through generation of reactive oxygen species in human lungcarcinoma cells. Cancer Lett. 239, 55–63.
Lee, H.Z., Wu, C.H., Chang, S.P., 2005. Release of nucleophosmin from the nucleus:Involvement in aloe-emodin-induced human lung non small carcinoma cellapoptosis. Int. J. Cancer 113, 971–976.
Lin, J.G., Chen, G.W., Li, T.M., Chouh, S.T., Tan, T.W., Chung, J.G., 2006. Aloe-emodininduces apoptosis in T24 human bladder cancer cells through the p53dependent apoptotic pathway. J. Urol. 175, 343–347.
Maksimovic-Ivanic, D., Mijatovi, S., Harhaji, L., Miljkovic, D., Dabideen, D., FanCheng, K., Mangano, K., Malaponte, G., Al-Abed, Y., Libra, M., Garotta, G.,Nicoletti, F., Stosic-Grujicic, S., 2008. Anticancer properties of the novel nitricoxide-donating compound (S, R)-3-phenyl-4,5-dihydro-5-isoxazole acetic acid-nitric oxide in vitro and in vivo. Mol. Cancer Ther. 7, 510–520.
Mijatovic, S., Maksimovic-Ivanic, D., Mojic, M., Timotijevic, G., Miljkovic, D.,Mangano, K., Donia, M., Di Cataldo, A., Al-Abed, Y., Cheng, K.F., Stosic-Grujicic,S., Nicoletti, F., 2011. Cytotoxic and immune-sensitizing properties of nitricoxide-modified Saquinavir in iNOS-positive human melanoma cells. J. Cell.Physiol. 226, 1803–1812.
Mijatovic, S., Maksimovic-Ivanic, D., Radovic, J., Miljkovic, D., Kaludjerovic, G.N.,Sabo, T.J., Trajkovic, V., 2005a. Aloe emodin decreases the ERK-dependentanticancer activity of cisplatin. Cell. Mol. Life Sci. 62, 1275–1282.
Mijatovic, S., Maksimovic-Ivanic, D., Radovic, J., Miljkovic, Dj., Harhaji, Lj, Vuckovic,O., Stosic-Grujicic, S., Mostarica Stojkovic, M., Trajkovic, V., 2005b. Anti-gliomaaction of aloe emodin: the role of ERK inhibition. Cell. Mol. Life Sci. 62, 589–598.
Mijatovic, S., Maksimovic-Ivanic, D., Radovic, J., Popadic, D., Momcilovic, M., Harhaji,Lj., Miljkovic, D., Trajkovic, V., 2004. Aloe-emodin prevents cytokine-inducedtumor cell death: the inhibition of auto-toxic nitric oxide release as a potentialmechanism. Cell. Mol. Life Sci. 61, 1805–1815.
Mueller, S.O., Stopper, H., Dekant, W., 1998. Biotransformation of theanthraquinones emodin and chrysophanol by cytochrome P450 enzymes.Bioactivation to genotoxic metabolites. Drug Metab. Dispos. 26, 540–546.
Pecere, T., Gazzola, M.V., Mucignat, C., Parolin, C., Vecchia, F.D., Cavaggioni, A., Basso,G., Diaspro, A., Salvato, B., Carli, M., Palu, G., 2000. Aloe-emodin is a new type ofanticancer agent with selective activity against neuroectodermal tumors.Cancer Res. 60, 2800–2804.
Russo, A.E., Torrisi, E., Bevelacqua, Y., Perrotta, R., Libra, M., McCubrey, J.A.,Spandidos, D.A., Stivala, F., Malaponte, G., 2009. Melanoma: molecularpathogenesis and emerging target therapies. Int. J. Oncol. 34, 1481–1489.
Steinbeck, M.J., Kim, J.K., Trudeau, M.J., Hauschka, P.V., Karnovsky, M.J., 1998.Involvement of hydrogen peroxide in the differentiation of clonal HD-11EMcells into osteoclast-like cells. J. Cell. Physiol. 176, 574–587.
Su, Y.T., Chang, H.L., Shyue, S.K., Hsu, S.L., 2005. Emodin induces apoptosis in humanlung adenocarcinoma cells through a reactive oxygen species-dependentmitochondrial signaling pathway. Biochem. Pharmacol. 70, 229–241.
Tabolacci, C., Lentini, A., Mattioli, P., Provenzano, B., Oliverio, S., Carlomosti, F.,Beninati, S., 2010. Antitumor properties of aloe-emodin and induction oftransglutaminase 2 activity in B16–F10 melanoma cells. Life Sci. 87, 316–324.
Tang, C.H., Grimm, E.A., 2004. Depletion of endogenous nitric oxide enhancescisplatin-induced apoptosis in a p53-dependent manner in melanoma cell lines.J. Biol. Chem. 279, 288–298.
Tokumoto, Y.M., Durand, B., Raff, M.C., 1999. An analysis of the early events whenoligodendrocyte precursor cells are triggered to differentiate by thyroidhormone, retinoic acid, or PDGF withdrawal. Dev. Biol. 213, 327–339.
Ullmannová, V., Stöckbauer, P., Hradcová, M., Soucek, J., Haskovec, C., 2003.Relationship between cyclin D1 and p21(Waf1/Cip1) during differentiation ofhuman myeloid leukemia cell lines. Leuk. Res. 27, 1115–1123.
Wasserman, L., Avigad, S., Beery, E., Nordenberg, J., Fenig, E., 2002. The effect of aloeemodin on the proliferation of a new merkel carcinoma cell line. Am. J.Dermatopathol. 24, 17–22.
Wu, X., Hasinoff, B.B., 2005. The antitumor anthracyclines doxorubicin anddaunorubicin do not inhibit cell growth through the formation of iron-mediated reactive oxygen species. Anticancer Drugs 16, 93–99.
Xiao, Q., Luo, Z., Pepe, A.E., Margariti, A., Zeng, L., Xu, Q., 2009. Embryonic stem celldifferentiation into smooth muscle cells is mediated by Nox4-produced H2O2.Am. J. Physiol. Cell. Physiol. 296, C711–C723.
Yagi, T., Yamauchi, K., Kuwano, S., 1997. The synergistic purgative action of aloe-emodin anthrone and rhein anthrone in mice. synergism in large intestinalpropulsion and water secretion. J. Pharm. Pharmacol. 49, 22–25.
Yang, J., Li, H., Chen, Y.Y., Wang, X.J., Shi, G.Y., Hu, Q.S., Kang, X.L., Lu, Y., Tang, X.M.,Guo, Q.S., Yi, J., 2004. Anthraquinones sensitize tumor cells to arseniccytotoxicity in vitro and in vivo via reactive oxygen species-mediated dualregulation of apoptosis. Free. Radic. Biol. Med. 37, 2027–2041.
Yeh, F.T., Wu, C.H., Lee, H.Z., 2003. Signalling pathway for aloe-emodin-inducedapoptosis in human H460 lung nonsmall carcinoma cell. Int. J. Cancer 106, 26–33.
Zhang, L., Lau, Y.K., Xia, W., Hortobagyi, G.N., Hung, M.C., 1999. Tyrosine kinaseinhibitor emodin suppresses growth of HER-2/neu-overexpressing breastcancer cells in athymic mice and sensitizes these cells to the inhibitory effectof paclitaxel. Clin. Cancer Res. 5, 343–353.
J. Radovic et al. / Food and Chemical Toxicology 50 (2012) 3181–3189 3189




















