
Binding characteristics and regulatory mechanisms of the transcription factors controlling...
13
Binding Characteristics and Regulatory Mechanisms of the Transcription Factors Controlling Oleate-responsive Genes in Saccharomyces cerevisiae * Received for publication, October 3, 2007, and in revised form, February 5, 2008 Published, JBC Papers in Press, February 19, 2008, DOI 10.1074/jbc.M708215200 Igor V. Karpichev 1 , Jorge M. Durand-Heredia, Yi Luo 2 , and Gillian M. Small 3 From the Department of Biology, City College of the City University of New York, New York, New York 10031 Transcriptional activation of many genes involved in peroxi- some-related functions is regulated by the Oaf1p, Pip2p, and Adr1p transcription factors in Saccharomyces cerevisiae. We have analyzed the in vivo binding characteristics of Oaf1p-Pip2p and found that this complex is recruited to its target oleate- response element (ORE) under all growth conditions tested. In addition, this complex also binds to ORE-containing genes that do not appear to be regulated by these proteins, as well as to some genes lacking conventional OREs. The recruitment of the Oaf1p-Pip2p complex was greatly increased upon glucose dere- pression, possibly due to Oaf1p phosphorylation with only mod- erate increases upon oleate induction. Thus, this complex may receive a nutritional cue while it is already bound to DNA, sug- gesting that, in addition to the increase in Oaf1p-Pip2p binding, other mechanism(s) such as enhanced Adr1p association may drive the expression of highly inducible fatty acid-responsive genes. Adr1p binds to target genes in an oleate-dependent fash- ion and is involved in Oaf1p-Pip2p binding. In turn, the Oaf1p- Pip2p complex appears to be important for Adr1p binding to a subset of oleate-responsive genes. Adr1p is a positive regulator of ORE-containing genes, but it also acts as a negative factor in expression of some of these genes. Finally, we have also shown that Adr1p is directly involved in mediating oleate induction of Oaf1p-Pip2p target genes. The yeast Saccharomyces cerevisiae is able to use a wide vari- ety of carbon sources to maintain vegetative growth. When fer- mentable sugars, such as glucose, are supplied into culture media, genes involved in utilization of less favorable nonfer- mentable carbon sources are turned off via a mechanism called glucose repression (1). A switch from glucose to a nonferment- able carbon source triggers transcriptional activation of these genes through another pathway, termed glucose derepression (2). Some of these glucose-derepressible genes, however, are able to undergo further induction upon receiving additional nutritional signals. For example, the genes encoding -oxida- tion enzymes become significantly induced when oleate is included in glycerol-containing growth media; the addition of oleate also leads to peroxisome proliferation (3). The promoters of these genes share a common sequence motif termed oleate- response element (ORE) 4 (4). This sequence is sufficient to act as an in vitro binding target for a trans-acting factor(s) respon- sible for oleate induction (4). We and others (5–7) identified two highly homologous Zn 2 Cyc 6 transcription factors, Oaf1p and Pip2p, which mediate oleate-dependent transcriptional activation of ORE-containing genes. Both of these proteins are required to trigger oleate-dependent induction and can be recruited by synthetic OREs in in vitro binding assays. Based on this experimental evidence, and by analogy with mammalian steroid hormone receptors (8 –10), a model in which Oaf1p and Pip2p bind to OREs of target genes in the form of a het- erodimer, presumably upon receiving an oleate nutritional sig- nal, has been proposed (5–7). Oaf1p and Pip2p play different roles in oleate induction. Although both of these proteins possess ligand binding domains, only Oaf1p binds free fatty acids (11) and is capable of sensing a fatty acid nutritional signal (12). The different regulation of genes encoding Oaf1p and Pip2p probably reflects different roles of these proteins in oleate response, whereas OAF1 is constantly expressed under all growth con- ditions, PIP2 is tightly controlled by an autoregulation mechanism (5, 6, 13, 14). In an earlier study we conducted a global search for ORE- containing genes and identified 40 genes that have oleate response-like elements in their promoter regions (5). We dem- onstrated that more than 20 genes, encoding proteins with var- ious subcellular locations, are regulated by the Oaf1p-Pip2p transcription factors. These include a number of highly induc- ible genes, as well as some genes that are only moderately induced by oleate. Subsequently, several genome-wide searches for ORE-containing and oleate-inducible genes were carried out, and most of the yeast genes that are significantly up-regu- lated by fatty acids have been identified (14 –16). Interestingly, some of the Oaf1p-Pip2p-regulated genes lack a consensus ORE (5, 14). The minimal ORE is now defined as CGGN 3 TN(A/ R)N 8 –12 CCG (17). * This work was supported by American Heart Association Grant 0350364N, NIGMS Grant SO6 GM-08168 from the National Institutes of Health, and National Institutes of Health Grant RCMI RR 03060. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 1 Present address: Centre of Bioengineering, Russian Academy of Sciences, Moscow 117984, Russia. 2 Present address: Cardiovascular, Metabolic, and Endocrine Diseases Depart- ment, Pfizer Inc., Groton, CT 06385. 3 To whom correspondence should be addressed: 535 East 80th St., New York, NY 10075. Tel.: 212-794-5417; Fax: 212-794-5378; E-mail: Gillian.small@ mail.cuny.edu. 4 The abbreviations used are: ORE, oleate-response element; ChIP, chromatin immunoprecipitation; RT, reverse transcription; CIP, calf intestinal phos- phatase; UAS, upstream activation sequence; HA, hemagglutinin; IP, immunoprecipitation. THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 283, NO. 16, pp. 10264 –10275, April 18, 2008 © 2008 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. 10264 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 16 • APRIL 18, 2008 by guest on June 10, 2016 http://www.jbc.org/ Downloaded from
Transcript of Binding characteristics and regulatory mechanisms of the transcription factors controlling...

Binding Characteristics and Regulatory Mechanisms of theTranscription Factors Controlling Oleate-responsive Genes inSaccharomyces cerevisiae*
Received for publication, October 3, 2007, and in revised form, February 5, 2008 Published, JBC Papers in Press, February 19, 2008, DOI 10.1074/jbc.M708215200
Igor V. Karpichev1, Jorge M. Durand-Heredia, Yi Luo2, and Gillian M. Small3
From the Department of Biology, City College of the City University of New York, New York, New York 10031
Transcriptional activation of many genes involved in peroxi-some-related functions is regulated by the Oaf1p, Pip2p, andAdr1p transcription factors in Saccharomyces cerevisiae. Wehave analyzed the in vivobinding characteristics ofOaf1p-Pip2pand found that this complex is recruited to its target oleate-response element (ORE) under all growth conditions tested. Inaddition, this complex also binds to ORE-containing genes thatdo not appear to be regulated by these proteins, as well as tosome genes lacking conventional OREs. The recruitment of theOaf1p-Pip2p complex was greatly increased upon glucose dere-pression, possibly due toOaf1pphosphorylationwith onlymod-erate increases upon oleate induction. Thus, this complex mayreceive a nutritional cue while it is already bound to DNA, sug-gesting that, in addition to the increase inOaf1p-Pip2p binding,other mechanism(s) such as enhanced Adr1p association maydrive the expression of highly inducible fatty acid-responsivegenes. Adr1p binds to target genes in an oleate-dependent fash-ion and is involved in Oaf1p-Pip2p binding. In turn, the Oaf1p-Pip2p complex appears to be important for Adr1p binding to asubset of oleate-responsive genes. Adr1p is a positive regulatorof ORE-containing genes, but it also acts as a negative factor inexpression of some of these genes. Finally, we have also shownthat Adr1p is directly involved in mediating oleate induction ofOaf1p-Pip2p target genes.
The yeast Saccharomyces cerevisiae is able to use a wide vari-ety of carbon sources tomaintain vegetative growth.When fer-mentable sugars, such as glucose, are supplied into culturemedia, genes involved in utilization of less favorable nonfer-mentable carbon sources are turned off via a mechanism calledglucose repression (1). A switch from glucose to a nonferment-able carbon source triggers transcriptional activation of thesegenes through another pathway, termed glucose derepression(2). Some of these glucose-derepressible genes, however, are
able to undergo further induction upon receiving additionalnutritional signals. For example, the genes encoding �-oxida-tion enzymes become significantly induced when oleate isincluded in glycerol-containing growth media; the addition ofoleate also leads to peroxisomeproliferation (3). The promotersof these genes share a common sequence motif termed oleate-response element (ORE)4 (4). This sequence is sufficient to actas an in vitro binding target for a trans-acting factor(s) respon-sible for oleate induction (4). We and others (5–7) identifiedtwo highly homologous Zn2Cyc6 transcription factors, Oaf1pand Pip2p, which mediate oleate-dependent transcriptionalactivation of ORE-containing genes. Both of these proteins arerequired to trigger oleate-dependent induction and can berecruited by synthetic OREs in in vitro binding assays. Based onthis experimental evidence, and by analogy with mammaliansteroid hormone receptors (8–10), amodel inwhichOaf1p andPip2p bind to OREs of target genes in the form of a het-erodimer, presumably upon receiving an oleate nutritional sig-nal, has been proposed (5–7).Oaf1p and Pip2p play different roles in oleate induction.
Although both of these proteins possess ligand bindingdomains, only Oaf1p binds free fatty acids (11) and is capableof sensing a fatty acid nutritional signal (12). The differentregulation of genes encoding Oaf1p and Pip2p probablyreflects different roles of these proteins in oleate response,whereas OAF1 is constantly expressed under all growth con-ditions, PIP2 is tightly controlled by an autoregulationmechanism (5, 6, 13, 14).In an earlier study we conducted a global search for ORE-
containing genes and identified 40 genes that have oleateresponse-like elements in their promoter regions (5). We dem-onstrated that more than 20 genes, encoding proteins with var-ious subcellular locations, are regulated by the Oaf1p-Pip2ptranscription factors. These include a number of highly induc-ible genes, as well as some genes that are only moderatelyinduced by oleate. Subsequently, several genome-wide searchesfor ORE-containing and oleate-inducible genes were carriedout, and most of the yeast genes that are significantly up-regu-lated by fatty acids have been identified (14–16). Interestingly,some of the Oaf1p-Pip2p-regulated genes lack a consensusORE (5, 14). TheminimalORE is nowdefined asCGGN3TN(A/R)N8–12CCG (17).
* This work was supported by American Heart Association Grant 0350364N,NIGMS Grant SO6 GM-08168 from the National Institutes of Health, andNational Institutes of Health Grant RCMI RR 03060. The costs of publicationof this article were defrayed in part by the payment of page charges. Thisarticle must therefore be hereby marked “advertisement” in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
1 Present address: Centre of Bioengineering, Russian Academy of Sciences,Moscow 117984, Russia.
2 Present address: Cardiovascular, Metabolic, and Endocrine Diseases Depart-ment, Pfizer Inc., Groton, CT 06385.
3 To whom correspondence should be addressed: 535 East 80th St., New York,NY 10075. Tel.: 212-794-5417; Fax: 212-794-5378; E-mail: [email protected].
4 The abbreviations used are: ORE, oleate-response element; ChIP, chromatinimmunoprecipitation; RT, reverse transcription; CIP, calf intestinal phos-phatase; UAS, upstream activation sequence; HA, hemagglutinin; IP,immunoprecipitation.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 283, NO. 16, pp. 10264 –10275, April 18, 2008© 2008 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
10264 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 16 • APRIL 18, 2008
by guest on June 10, 2016http://w
ww
.jbc.org/D
ownloaded from

In addition to theORE, genes encoding peroxisomal proteinsoften contain a binding site for a sensor of less favored carbonsources, the C2H2 zinc-finger protein Adr1p (18–21). Thissequence, referred to as type 1 upstream activation sequence(UAS1), is defined as CYCCR(A/T/GN4–36(T/A/C)YGGRGand is often found in close proximity to, or overlapping with, anORE in the Oaf1p-Pip2p-regulated genes (17, 21, 22). Adr1p,originally identified as a regulator of the glucose-repressiblegene ADH2 (23, 24), was later shown to be required for growthon oleic acid as a sole carbon source (18, 25). Adr1p regulatesperoxisome-related target genes in two ways, directly via bind-ing to the resident UAS1 sequences, and also through mediat-ing expression of PIP2 (18–21). Adr1p target genes include thekey peroxisome-proliferation peroxin gene PEX11, and thusperoxisome proliferation is also under Adr1p control (19). Thebinding of Adr1p to peroxisome-related genes has been dem-onstrated both in vitro with synthetic UAS1 targets underoleate induction (18–21) and in vivo using chromatin immu-noprecipitation (ChIP) assays in the absence of glucose (26–28). Recently, Adr1p-regulated genes and Adr1p-binding siteshave been identified using genome-wide global analyses; how-ever, there have been no such analyses performed under oleateinduction conditions (26–31).In recent years, chromatin immunoprecipitation in combi-
nation with quantitative PCR has become a powerful andwidely used tool for detecting the associations of regulatoryproteinswith specificDNAsequences in vivo (32, 33). Althougha number of genome-wide location analyses using ChIP fol-lowed by DNA microarray (ChIP on chip) assays have beencompleted to date (27, 29, 30, 34), none of them focused specif-ically on Oaf1p-Pip2p interactions with their target sequences,and there have been no such studies addressing regulationunder fatty acid-inducing conditions. In this study, we haveexamined the binding characteristics of Oaf1p and Pip2p totheir target sequences under three different growth conditions,including oleate induction, in vivo. We confirmed that Oaf1prequires Pip2p for binding and that these proteins act as a het-erodimer on their binding sites under all growth conditionstested. We also demonstrated that, in contrast to the currentview, the amount of Oaf1p-Pip2p complex bound to the ORE issignificantly increased upon glucose derepression and
increases onlymoderately upon oleate induction. Furthermore,we provide evidence that Oaf1p undergoes phosphorylationwhen cells are grown on glycerol- or oleate-containing media,and this post-translational modification may cause anincreased Oaf1p-Pip2p binding under these conditions. Thus,Oaf1pmost likely receives a fatty acid nutritional signal while itis already bound to DNA, therefore receiving such a signal doesnot cause the initial binding of Oaf1p-Pip2p to the ORE, as waspreviously thought. These data also suggest that the increase inOaf1p-Pip2p binding upon a shift from glycerol to oleate is notsufficient to achieve maximal induction of highly inducibleoleate-responsive genes, and therefore there must be an addi-tional mechanism(s) involved in up-regulation of these genes.We have also demonstrated that Adr1p binds to its targetsequences under derepression and induction growth condi-tions, and this binding appears to be oleate-dependent. Ourfindings indicate that Adr1p is involved in Oaf1p-Pip2p bind-ing, and this complex, in turn, stimulates Adr1p recruitment tothe promoters of a subset of genes. In addition, we determinedthat Adr1p is not only required for achieving maximal expres-sion levels, but it also contributes to oleate induction of theOaf1p-Pip2p target genes. Finally, we have shown that Adr1p,being a positive regulator of oleate induction of peroxisome-related genes, plays an additional role in maintaining lowexpression levels of some oleate-responsive genes under glu-cose derepression conditions (negative regulation).
EXPERIMENTAL PROCEDURES
Yeast Strains and Growth Conditions
The yeast strains used in this study are listed in Table 1. Yeaststrains were grown in YPD, YPG, or YPGOmedia as describedpreviously (5, 6, 35).
Epitope Tagging
OAF1-HA3—The nine-amino acid epitope of influenza virushemagglutinin (HA) was used for C-terminal tagging theOAF1gene product in our previous studies (5). To create a plasmidthat contains both Oaf1p-HA3 and the URA3 selectablemarker, a DNA fragment encoding the OAF1-HA3 allele wassubcloned into pRS306 (36) to create pRS306OAF-1HA3. This
TABLE 1S. cerevisiae strains used in this study
Strain Genotype Ref.W3031A MATa leu2 ura3 trp1 ade2 his3 51OAF1-HA3 MATa leu2 ura3 �pRS306-URA3-OAF1-HA3� trp1 ade2 his3 oaf1::HIS3 This studyPIP2-HA3 MATa leu2 pip2::LEU2 ura3 trp1 �pRS304-TRP1-PIP2-HA3� his3 This studyPIP2-HA3/OAF1-myc13 MATa leu2 pip2::LEU2 ura3 trp1 �pRS304-TRP1-PIP2-HA3� his3 oaf1:: �OAF1-13Myc-TADH1-HIS5� This study��OAF1-HA3 MATa leu2 pip2::LEU2 ura3 �pRS306-URA3-OAF1-HA3� trp1 his3 oaf1::HIS3 This study��PIP2-HA3 MATa leu2 pip2::LEU2 ura3 trp1 �pRS304-TRP1-PIP2-HA3� his3 oaf1::HIS This studyADR1-myc13 MATa leu2 ura3 trp1 ade2 his3 adr1:: �ADR1-13Myc-TADH1-HIS5� This study�ADR1 MATa leu2 ura3 trp1 adr1::13Myc-TADH1-TRP1 ade2 his3 This studyPIP2-HA3/�ADR1 MATa leu2 pip2::LEU2 ura3 trp1 �pRS304-TRP1-PIP2-HA3� his3 adr1::13Myc-TADH1-HIS5 This study�PIP2/ADR1-myc13 MATa leu2 pip2::LEU2 ura3 trp1 adr1:: �ADR1-13Myc-TADH1-TRP1� his3 This study��1�P2/ADR1-myc13 MATa leu2 pip2::LEU2 ura3 trp1 adr1:: �ADR1-13Myc-TADH1-TRP1� his3 oaf1::HIS3 This studyOAF1-HA3/ADR1-myc13 MATa leu2 ura3 �pRS306-URA3-OAF1-HA3� trp1 adr1:: �ADR1-13Myc-TADH1-TRP1� ade2 his3 oaf1::HIS3 This studyOAF1-HA3/�ADR1 MATa leu2 ura3 �pRS306-URA3-OAF1-HA3� trp1 adr1::13Myc-TADH1-TRP1 ade2 his3 oaf1::HIS3 This study�OAF1/ADR1-myc13 MATa leu2 ura3 trp1 oaf1::13Myc-TADH1-TRP1 ade2 his3 adr1:: �ADR1-13Myc-TADH1-HIS5� This studyPIP2-HA3/OAF1-myc13/�ADR1 MATa leu2 pip2::LEU2 ura3 adr1�::URA3 trp1 �pRS304-TRP1-PIP2-HA3�
his3 oaf1:: �OAF1-13Myc-TADH1-HIS5�This study
MCY2692OAF1-HA3 MATa leu2 ura3 �pRS306-URA3-OAF1-HA3� snf1-K84R This study��1�P2 MATa leu2 pip2::LEU2 ura3 trp1 his3 oaf1::HIS3 5
Transcriptional Control of Oleate Response in Yeast
APRIL 18, 2008 • VOLUME 283 • NUMBER 16 JOURNAL OF BIOLOGICAL CHEMISTRY 10265
by guest on June 10, 2016http://w
ww
.jbc.org/D
ownloaded from

plasmidwas then linearized and integrated into theURA3 locusof the appropriate recipient yeast cells.PIP2-HA3—Three tandem copies of the HA epitope were
added to the C terminus of the PIP2 gene product directlyupstream from the stop codon, using a combination of PCR andsubcloning techniques. The resulting DNA fragment encodingPIP2-HA3 was subcloned into pRS304 to create pRS304PIP-2HA3. This plasmidwas then linearized and integrated into theTRP1 locus of our pip2� recipient strain.OAF1-myc13 and ADR1-myc13—Thirteen copies of the
10-amino acid human c-Myc epitope were integrated intothe OAF1 or ADR1 chromosomal loci using pF6a-13Myc-TRP1 or pF6a-13MycHis3-MX6 heterologous modules forPCR-based gene targeting (37).
Gene Disruption by PCR-based Gene Targeting
PCR-based gene targeting approach was also used to disruptthe OAF1 gene in the ADR1-myc13 strain using pF6a-13Myc-TRP1 as a template. Similarly, the ADR1 gene was disrupted inour wild-type strain (see Table 1). To disrupt the ADR1 gene inthe PIP2-HA3/OAF1-myc13 strain, aURA3 template was usedinstead of the pF6-based cassettes. The correct integration ofthe cassettes was verified by diagnostic PCR.
ChIP
The procedures described below for single and sequentialChIPs are modifications of the methods of Aparicio et al. (32)andGeisberg et al. (38), respectively. All buffers and stock solu-tion recipes were taken from original protocols, except wheredescribed below.Lysate Preparation—Yeast cultures were grown in 200 ml of
YPD, YPG, or YPGO aswas described previously (5). Cells weresubsequently cross-linked in the media, harvested, and washedas described (32). The fixed cells were resuspended in 6–10 mlof FAPI buffer (FA buffer supplemented with complete EDTA-free protease inhibitor mixture (Roche Applied Science) and 1mM phenylmethylsulfonyl fluoride). 1-ml aliquots of this sus-pension were placed in 2-ml Eppendorf tubes, and �1 ml ofglass beads (diameter � 0.5 mm) was added to every sample.The suspensions were incubated, with shaking, for 1.5–2 h at4 °C. The cell lysates were collected together and centrifuged asdescribed (32). The resultant pellets, containing cross-linkedchromatin, were resuspended in 4 ml of FAPI buffer, and thevolume was brought to 5 ml if required. The suspension wassonicated on ice 20 times for 30 s using continuous pulse with1-min intervals at a power output of �15% using aW-385 son-icator (Heat Systems-Ultrasonics, Farmingdale, NY). Followingsonication, the lysate was split into 2-ml Eppendorf tubes andcentrifuged in a refrigerated microcentrifuge at maximumspeed for 30 min. The recovered supernatants were pooled andpassed through a 0.22-�m syringe filter. A 100-�l sample wasthen taken to confirmDNA fragmentation (32), and the volumeof the remaining lysate was adjusted to 5 ml with FAPI buffer.The lysatewas divided into 500-�l aliquots placed in Eppendorftubes, frozen in liquid nitrogen, and stored at �80 °C. Beforeusing the lysate in ChIP assays, it was titrated to determine theamount of respective antibody required to precipitate between50 and 90% of the antigen in question as described (32).
Single ChIP Assay—For an individual single ChIP, 250–500�l of the lysate was taken into an Eppendorf tube, and the vol-ume was brought to 1 ml with FAPI buffer. A 50-�l sample wastaken to determine the amount of DNA present in the assay(input sample), and a 30-�l sample was taken to determine theamount of antigen in the assay (T-sample). These samples wereprocessed as described (32). Upon addition of antibody, thetube was incubated at 4 °C for 2 h on anOrbitron Rotator. 50�lof protein G-agarose beads (Roche Applied Science) were thenadded, and the incubation was continued overnight. The sam-ple was washed, eluted with 100 �l of the appropriate peptideelution buffer, and reverse cross-linked as described (32). DNAwas then purified using phenol/chloroform extraction followedby ethanol precipitation with GlycoBlue carrier (Ambion, Aus-tin, TX) as described (32) and was then dissolved in 40–100 �lof TE as required. A 30-�l supernatant sample (S-sample) wastaken before washing and run along with the T-sample onto an8% SDS-PAGE followed by ECLWestern blotting to determinethe percentage of the antigen precipitated by ChIP. The inputsamples were reverse cross-linked as described (32), purifiedusing a QIAquick PCR purification kit (Qiagen, Valencia, CA)according to the manufacturer’s instructions, and eluted in 200�l. Both ChIP and input samples were stored at �20 °C.Sequential ChIPAssay—Sequential ChIPswere performed in
both the forward and reverse direction. For this purpose, 1–2ml of cross-linked lysates was taken for each direction. 50-�linput samples were processed as described above. The firstChIP samples were set up and then carried out as describedabove for single chips. After elution with the correspondingpeptide, 10 �l of the first ChIP eluates was taken into a PCRtube containing 90 �l of TE, 100 �l of ChIP elution buffer, and20 �l of Pronase. The samples were subjected to reverse cross-linking and then purified as described (38). The samples wereresuspended in 20–100 �l of TE and stored at �20 °C as “firstChIP.” To set up second ChIPs, the remaining 90 �l of the firstChIP eluate weremixedwith 600�l of FABPR buffer (FA buffercontaining 5 mg/ml bovine serum albumin, 25 �g/ml � phageDNA (NewEnglandBiolabs, Beverly,MA)) and 50�g/mlE. colitRNA (Roche Applied Science), and second antibody (50% ofthe amount used above) was added to the samples. The secondChIP samples were incubated for 2 h and then 50 �l of proteinG-agarose beads were added, and incubation was continuedovernight. The samples were washed, eluted, and purified aswas described above for the single ChIPs. The purified samples,“second ChIP,” were stored at �20 °C.
RNA Purification
Total yeast RNA was purified according to a hot phenolextraction procedure as described (5). mRNA was preparedfrom total yeast RNA using Oligotex milk as specified by themanufacturer (Qiagen, Valencia, CA). Before use in the realtime RT-PCR assays, both yeast total andmRNAwere liberatedfrom DNA using the DNA-freeTM kit (Ambion Inc., Austin,TX) according to the manufacturer’s instructions.
Quantitatative Real Time PCR
Real time PCR was performed using the LightCycler 2.0Instrument (Roche Applied Science) and LightCycler software
Transcriptional Control of Oleate Response in Yeast
10266 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 16 • APRIL 18, 2008
by guest on June 10, 2016http://w
ww
.jbc.org/D
ownloaded from

version 4.0. The primers were designed using LightCyclerProbe Design 2 software based on the sequences of the specificgenes downloaded from the yeast genome data base. All oligo-nucleotide sequences are available upon request. The amplifi-cation programs were designed according to manufacturer’srecommendations. 10-�l reactions were set up in 20-�l Light-Cycler capillaries (Roche Applied Science). DNA (ChIPs andinputs) samples were amplified using either LightCycler Fast-Start DNA MasterPLUS SYBR Green I kit (Roche Applied Sci-ence) or homemade 2� SYBR Green Taq mix (32). The singleChIP data were processed using Microsoft Office Excel soft-ware and the following formula: Chip% � (2Cp input�Cp chip) �Fd � 100, where Chip% is the ChIP efficiency expressed in per-cent when compared with total DNA; Cp chip and Cp input arecrossing points (threshold cycles) for ChIP and input samples,respectively; and Fd is the dilution factor. The relative occu-pancy was calculated as the fold enrichment of the gene-spe-cific DNA over a reference gene DNA (GPD1 or POX1). Forsequential ChIPs, C values, or efficiencies of co-occupancy, inpercent, were calculated according to the following formula:C � 100(AB-A)/(A � B � A), whereA, B, andAB represent thefold enrichments for the first, second, and sequential ChIP,respectively (38). For real time RT-PCR, the one-step Quanti-Test Multiplex RT-PCR NR kit (Qiagen) was used. The one-step reactions were set up in 10 �l, and the RT-PCR programswere designed according to the manufacturer’s recommenda-tions. The relative mRNA expression (R) for every gene wascalculated using the formula R � 2Cp actin �Cp gene taking theACT1 gene as a reference.
Phosphatase Treatment of Oaf1p-HA3
Yeast strains were grown on YPD, YPG, or YPGOmedia, andcell extracts were prepared as described previously (35). Forphosphatase treatment of Oaf1p-HA3 expressed in wild-typecells (strain OAF1-HA3), cell extracts containing 10 �g of totalprotein were incubated at 37 °C for 1 h in the presence of 20units of calf intestinal phosphatase (CIP, New England Biolabs)in a total volume of 10–15�l. Phosphatase inhibitormixture (5mM sodium phosphate, pH 7.5, 10 mM sodium pyrophosphate,and 5 mM EDTA) and/or 0.2% SDS was added to the samples ifrequired. Control sample was treated identically, but CIP andphosphatase inhibitors were not included in the reaction mix-ture. The treated samples were then separated on a 7% SDS-PAGE followed by immunoblotting with 12CA5 anti-HA anti-body. To analyze the migration pattern of Oaf1p-HA3expressed in snf1 mutant cells under glucose, glycerol, andoleate growth conditions, cell lysates containing 50 �g of totalprotein were separated on a 7% SDS-PAGE followed by immu-noblotting with 12CA5 anti-HA antibody. Densitometric anal-ysiswas carried out to identify the exact position of each proteinband using ImageQuant software (GE Healthcare).
Antibodies
Rabbit polyclonal affinity purified HA and c-Myc antibodieswere obtained from Bethyl Laboratories (Montgomery, TX);mouse monoclonal anti-HA antibody (clone 12CA5), ratmonoclonal anti-HA antibody (clone 3F10), anti-HA-peroxi-dase high affinity and mouse monoclonal anti-c-Myc antibody
(clone 9E10), and anti-(c-Myc)-peroxidase were obtained fromRoche Applied Science; anti-rabbit IgG (whole molecule) per-oxidase conjugate was obtained from Sigma.
RESULTS
Choosing a System to Study Oaf1p and Pip2p Binding in Vivo—The binding of the Oaf1p-Pip2p complex to a synthetic POX1ORE in vitro is strongly regulated by the carbon source suppliedin the growth media. In our previous studies we observed aweak band shift using protein extracts prepared from glucose-grown cells (39). The signal was increased upon derepression(glycerol media) and reached its highest levels under oleate-growth conditions. One interpretation of these results is thatthe induction of the fatty acid-responsive genes in the presenceof oleate is because of an increase in the amount ofOaf1p-Pip2pcomplex binding to the target OREs in vivo. As a first steptoward testing this hypothesis, we measured the binding ofOaf1p and/or Pip2p to the target promoters in vivo, using chro-matin immunoprecipitation followed by real time PCR analysisof the immunoprecipitated DNA. Using a single ChIP assay,Pip2p-HA3 binding was highest when cells were grown onoleate for most of the genes analyzed. However, Oaf1p-HA3binding was highest under glycerol growth conditions whencompared with oleate (data not shown). It seems likely that thisdifference in Oaf1p-HA3 and Pip2p-HA3 binding levels inthese experiments is an artifact caused by HA-tagging Oaf1p.To overcome this potential problem, we created an additionalyeast strain that co-expresses a functional Pip2p-HA3 allele andaMyc-tagged Oaf1p allele (Oaf1p-myc13). UsingWestern blotanalysis with a polyclonal antibody to the acyl-CoA oxidaseprotein, we found that these co-expressing cells are able toinduce POX1 expression to a level similar to that seen for ourwild-type cells grown on oleate medium, indicating that thedouble-tagged (Oaf1p-myc13)-(Pip2p-HA3) complex is func-tional. We tested lysates prepared from this co-expressingstrain in single ChIP pilot experiments and found that the bind-ing patterns for both Oaf1p and Pip2p were similar to that seenfor Pip2p in previous experiments with a Pip2p-HA3 single-expressing strain (see below).In Vivo Binding of theOaf1p and Pip2p Transcription Factors
to the Promoters of the Target Genes—Table 2 lists 14 genes thatare potential targets for Oaf1p and Pip2p. These genes can beseparated into four different categories according to the modeof regulation by fatty acids. Type I genes are highly induced byunsaturated fatty acids such as oleate. These genes contain aconsensus ORE and are regulated by the Oaf1p and Pip2p tran-scription factors. Type II genes are also induced by oleate underthe regulation of Oaf1p and Pip2p, but to a lesser extent (2–4-fold). Type III genes contain a consensus ORE; however, thesegenes are repressed by unsaturated fatty acids while they areinduced by saturated fatty acids such as myristate. Finally, typeIV genes contain an ORE, but it appears to be nonfunctionalbecause they are not regulated by fatty acids. We also note thatthere are three genes (PXA1, YLR287c, and YMR018w) thatlack an obvious ORE in their promoters but nevertheless areup-regulated by oleate. Previously, we have shown that PXA1 isinduced 2-fold by oleate (5), whereas two other genes (YLR287c
Transcriptional Control of Oleate Response in Yeast
APRIL 18, 2008 • VOLUME 283 • NUMBER 16 JOURNAL OF BIOLOGICAL CHEMISTRY 10267
by guest on June 10, 2016http://w
ww
.jbc.org/D
ownloaded from
andYMR018w)were selected as candidates in a globalmicroar-ray expression search for oleate-inducible genes (14).To confirm thatOaf1p and Pip2p bind to the promoter of the
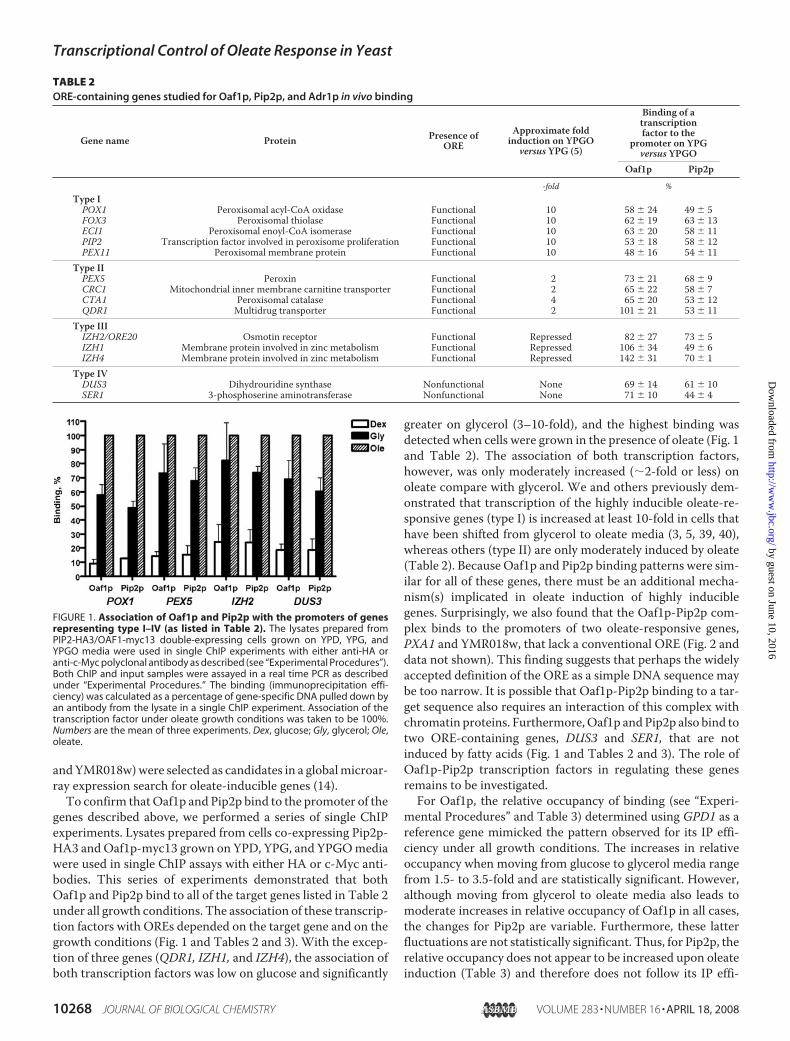
genes described above, we performed a series of single ChIPexperiments. Lysates prepared from cells co-expressing Pip2p-HA3 andOaf1p-myc13 grown on YPD, YPG, and YPGOmediawere used in single ChIP assays with either HA or c-Myc anti-bodies. This series of experiments demonstrated that bothOaf1p and Pip2p bind to all of the target genes listed in Table 2under all growth conditions. The association of these transcrip-tion factors with OREs depended on the target gene and on thegrowth conditions (Fig. 1 and Tables 2 and 3). With the excep-tion of three genes (QDR1, IZH1, and IZH4), the association ofboth transcription factors was low on glucose and significantly
greater on glycerol (3–10-fold), and the highest binding wasdetected when cells were grown in the presence of oleate (Fig. 1and Table 2). The association of both transcription factors,however, was only moderately increased (�2-fold or less) onoleate compare with glycerol. We and others previously dem-onstrated that transcription of the highly inducible oleate-re-sponsive genes (type I) is increased at least 10-fold in cells thathave been shifted from glycerol to oleate media (3, 5, 39, 40),whereas others (type II) are only moderately induced by oleate(Table 2). Because Oaf1p and Pip2p binding patterns were sim-ilar for all of these genes, there must be an additional mecha-nism(s) implicated in oleate induction of highly induciblegenes. Surprisingly, we also found that the Oaf1p-Pip2p com-plex binds to the promoters of two oleate-responsive genes,PXA1 and YMR018w, that lack a conventional ORE (Fig. 2 anddata not shown). This finding suggests that perhaps the widelyaccepted definition of the ORE as a simple DNA sequence maybe too narrow. It is possible that Oaf1p-Pip2p binding to a tar-get sequence also requires an interaction of this complex withchromatin proteins. Furthermore,Oaf1p andPip2p also bind totwo ORE-containing genes, DUS3 and SER1, that are notinduced by fatty acids (Fig. 1 and Tables 2 and 3). The role ofOaf1p-Pip2p transcription factors in regulating these genesremains to be investigated.For Oaf1p, the relative occupancy of binding (see “Experi-
mental Procedures” and Table 3) determined using GPD1 as areference gene mimicked the pattern observed for its IP effi-ciency under all growth conditions. The increases in relativeoccupancy when moving from glucose to glycerol media rangefrom 1.5- to 3.5-fold and are statistically significant. However,although moving from glycerol to oleate media also leads tomoderate increases in relative occupancy of Oaf1p in all cases,the changes for Pip2p are variable. Furthermore, these latterfluctuations are not statistically significant. Thus, for Pip2p, therelative occupancy does not appear to be increased upon oleateinduction (Table 3) and therefore does not follow its IP effi-
FIGURE 1. Association of Oaf1p and Pip2p with the promoters of genesrepresenting type I–IV (as listed in Table 2). The lysates prepared fromPIP2-HA3/OAF1-myc13 double-expressing cells grown on YPD, YPG, andYPGO media were used in single ChIP experiments with either anti-HA oranti-c-Myc polyclonal antibody as described (see “Experimental Procedures”).Both ChIP and input samples were assayed in a real time PCR as describedunder “Experimental Procedures.” The binding (immunoprecipitation effi-ciency) was calculated as a percentage of gene-specific DNA pulled down byan antibody from the lysate in a single ChIP experiment. Association of thetranscription factor under oleate growth conditions was taken to be 100%.Numbers are the mean of three experiments. Dex, glucose; Gly, glycerol; Ole,oleate.
TABLE 2ORE-containing genes studied for Oaf1p, Pip2p, and Adr1p in vivo binding
Gene name Protein Presence ofORE
Approximate foldinduction on YPGO
versus YPG (5)
Binding of atranscriptionfactor to the
promoter on YPGversus YPGO
Oaf1p Pip2p-fold %
Type IPOX1 Peroxisomal acyl-CoA oxidase Functional 10 58 24 49 5FOX3 Peroxisomal thiolase Functional 10 62 19 63 13ECI1 Peroxisomal enoyl-CoA isomerase Functional 10 63 20 58 11PIP2 Transcription factor involved in peroxisome proliferation Functional 10 53 18 58 12PEX11 Peroxisomal membrane protein Functional 10 48 16 54 11
Type IIPEX5 Peroxin Functional 2 73 21 68 9CRC1 Mitochondrial inner membrane carnitine transporter Functional 2 65 22 58 7CTA1 Peroxisomal catalase Functional 4 65 20 53 12QDR1 Multidrug transporter Functional 2 101 21 53 11
Type IIIIZH2/ORE20 Osmotin receptor Functional Repressed 82 27 73 5IZH1 Membrane protein involved in zinc metabolism Functional Repressed 106 34 49 6IZH4 Membrane protein involved in zinc metabolism Functional Repressed 142 31 70 1
Type IVDUS3 Dihydrouridine synthase Nonfunctional None 69 14 61 10SER1 3-phosphoserine aminotransferase Nonfunctional None 71 10 44 4
Transcriptional Control of Oleate Response in Yeast
10268 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 16 • APRIL 18, 2008
by guest on June 10, 2016http://w
ww
.jbc.org/D
ownloaded from

ciency pattern. Because Oaf1p and Pip2p IP efficiency patternsappear to be similar for an individual gene, the observed differ-ence in relative occupancy patterns of these proteins is mostlikely caused by variations in the experimental backgroundwhen a nonbindingGPD1 locus is used as a reference gene (32).To reduce the potential for experimental error, we deter-
mined the relative occupancy of Oaf1p and Pip2p using an
Oaf1p-Pip2p-binding gene (POX1) as a reference gene, ratherthan genes that do not recruit these transcription factors. Formost of the genes studied, the relative occupancy to an individ-ual gene was similar for both transcription factors on a givengrowth condition (glycerol or oleate), when expressed in POX1units (Fig. 2). These findings indicate thatOaf1p andPip2p bindto target genes in an equimolar ratio, and strongly support amodel in whichOaf1p and Pip2p act as a heterodimer to triggeroleate-inducible transcription. Thus, it appears that the differ-ence in patterns for Oaf1p and Pip2p relative occupancy seenpreviously (Table 3) is most likely because of the backgroundvariations for these two proteins whenGPD1 is used as a refer-ence gene. In contrast, we also observed that the relative occu-pancy of Oaf1pwas greater than that of Pip2p forQDR1,CTA1,and IZH genes in glycerol growth conditions (Fig. 2B). Thissuggests that Oaf1p alone, most likely in the form of ahomodimer, may be recruited to the promoter of a subset oftarget genes.The binding of both Oaf1p and Pip2p under a given growth
condition was variable among the genes investigated (Fig. 2).We have found no correlation between the groups of genes andthe relative occupancies of both proteins under a specificgrowth condition. For example, theDUS3 gene is not regulatedby the Oaf1p-Pip2p complex; however, both transcription fac-tors bind to this gene, and their associationwith theDUS3OREwas almost three times higher than the association withORE ofthe highly inducible POX1 promoter (Fig. 2).Regulated Phosphorylation of Oaf1p—Phosphorylation has
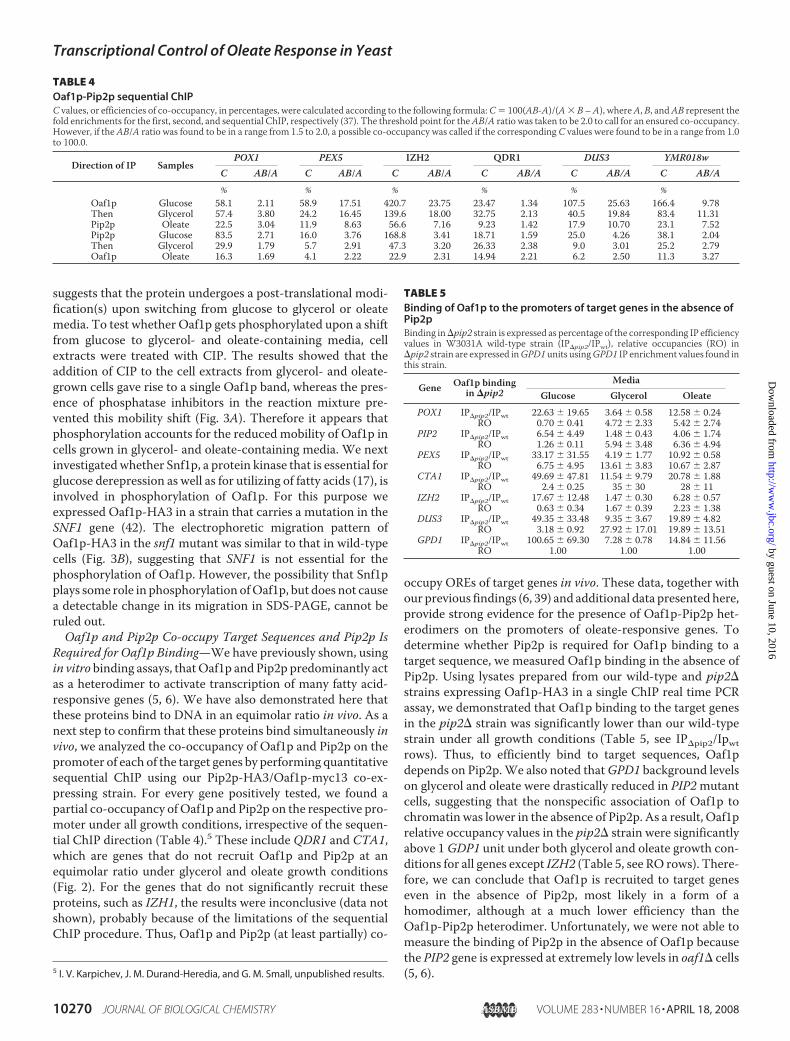
been shown to be required for activation of yeast transcriptionfactors, such as Gal4p (41). Because expression ofOAF1 is con-stitutive, irrespective of the growth conditions, we hypothe-sized that Oaf1p may be phosphorylated, and this modificationmay increase Oaf1p-Pip2p binding upon glucose derepressionand oleate induction. Immunoblot analysis showed that Oaf1p-HA3 expressed in glycerol-grown cells appeared to migrateslightly slower thanwhen this proteinwas expressed in glucose-grown cells (Fig. 3A). Upon switching fromglycerol to oleate, anadditional increase in Oaf1p-HA3 mobility was seen (Fig. 3A).Decreased mobility of Oaf1p-HA3 in glycerol- and oleate-grown cells compared with that seen in glucose-grown cells
FIGURE 2. Comparison of the relative occupancies of the Oaf1p and Pip2ptranscription factors on the promoters of target genes under glycerol (A)and oleate (B) growth conditions. Relative occupancies were calculated asdescribed under “Experimental Procedures” and Table 3 using the POX1 geneas a reference. Numbers are the mean of three experiments.
FIGURE 3. Oaf1p is phosphorylated upon glucose derepression andoleate induction. A, phosphatase treatment of Oaf1p-HA3. Reactions werecarried out with cell extracts from glucose-grown cells (lanes 1, 4, 7, and 10),glycerol-grown cells (lanes 2, 5, 8, and 11), and oleate-grown cells (lanes 3, 6, 9,and 12). The addition of CIP, phosphatase inhibitor mixture and 0.2% SDS isindicated below each lane. Numbers I–III show exact positions of each form ofOaf1p-HA3. B, migration pattern of Oaf1p-HA3 expressed in the snf1 mutantcells grown on glucose-containing media (lane 1), glycerol-containing media(lane 2), and oleate-containing media (lane 3).
TABLE 3Relative occupancies of Oaf1p and Pip2p on target genesThe relative occupancies were calculated as the fold enrichments of a given genomicregion over the GPD1 background according to the formula RO � IPgene/IPGPD1,where IPgene and IPGPD1 are the immunoprecipitation efficiencies for a given targetgene and theGPD1 gene, and RO is the relative occupancy expressed inGPD1 units,respectively.
Gene Transcriptionfactor
MediaGlucose Glycerol Oleate
POX1 Oaf1p 2.24 0.35 7.75 1.76 12.25 4.51Pip2p 4.38 1.96 12.13 3.83 14.57 2.34
PEX5 Oaf1p 11.82 2.02 25.79 1.65 37.59 7.90Pip2p 14.99 12.06 43.33 14.61 36.88 4.77
IZH2 Oaf1p 5.09 0.92 7.46 0.10 9.57 0.92Pip2p 6.75 4.95 13.61 3.83 10.67 2.87
DUS3 Oaf1p 14.52 5.51 22.31 3.19 34.58 8.89Pip2p 16.59 13.79 35.79 12.25 33.69 1.57
GPD1 Oaf1p 1.00 1.00 1.00Pip2p 1.00 1.00 1.00
Transcriptional Control of Oleate Response in Yeast
APRIL 18, 2008 • VOLUME 283 • NUMBER 16 JOURNAL OF BIOLOGICAL CHEMISTRY 10269
by guest on June 10, 2016http://w
ww
.jbc.org/D
ownloaded from
suggests that the protein undergoes a post-translational modi-fication(s) upon switching from glucose to glycerol or oleatemedia. To test whether Oaf1p gets phosphorylated upon a shiftfrom glucose to glycerol- and oleate-containing media, cellextracts were treated with CIP. The results showed that theaddition of CIP to the cell extracts from glycerol- and oleate-grown cells gave rise to a single Oaf1p band, whereas the pres-ence of phosphatase inhibitors in the reaction mixture pre-vented this mobility shift (Fig. 3A). Therefore it appears thatphosphorylation accounts for the reduced mobility of Oaf1p incells grown in glycerol- and oleate-containing media. We nextinvestigatedwhether Snf1p, a protein kinase that is essential forglucose derepression as well as for utilizing of fatty acids (17), isinvolved in phosphorylation of Oaf1p. For this purpose weexpressed Oaf1p-HA3 in a strain that carries a mutation in theSNF1 gene (42). The electrophoretic migration pattern ofOaf1p-HA3 in the snf1mutant was similar to that in wild-typecells (Fig. 3B), suggesting that SNF1 is not essential for thephosphorylation of Oaf1p. However, the possibility that Snf1pplays some role in phosphorylation ofOaf1p, but does not causea detectable change in its migration in SDS-PAGE, cannot beruled out.Oaf1p and Pip2p Co-occupy Target Sequences and Pip2p Is
Required for Oaf1p Binding—We have previously shown, usingin vitro binding assays, thatOaf1p and Pip2p predominantly actas a heterodimer to activate transcription of many fatty acid-responsive genes (5, 6). We have also demonstrated here thatthese proteins bind to DNA in an equimolar ratio in vivo. As anext step to confirm that these proteins bind simultaneously invivo, we analyzed the co-occupancy of Oaf1p and Pip2p on thepromoter of each of the target genes by performing quantitativesequential ChIP using our Pip2p-HA3/Oaf1p-myc13 co-ex-pressing strain. For every gene positively tested, we found apartial co-occupancy ofOaf1p and Pip2p on the respective pro-moter under all growth conditions, irrespective of the sequen-tial ChIP direction (Table 4).5 These include QDR1 and CTA1,which are genes that do not recruit Oaf1p and Pip2p at anequimolar ratio under glycerol and oleate growth conditions(Fig. 2). For the genes that do not significantly recruit theseproteins, such as IZH1, the results were inconclusive (data notshown), probably because of the limitations of the sequentialChIP procedure. Thus, Oaf1p and Pip2p (at least partially) co-
occupy OREs of target genes in vivo. These data, together withour previous findings (6, 39) and additional data presentedhere,provide strong evidence for the presence of Oaf1p-Pip2p het-erodimers on the promoters of oleate-responsive genes. Todetermine whether Pip2p is required for Oaf1p binding to atarget sequence, we measured Oaf1p binding in the absence ofPip2p. Using lysates prepared from our wild-type and pip2�strains expressing Oaf1p-HA3 in a single ChIP real time PCRassay, we demonstrated that Oaf1p binding to the target genesin the pip2� strain was significantly lower than our wild-typestrain under all growth conditions (Table 5, see IP�pip2/Ipwtrows). Thus, to efficiently bind to target sequences, Oaf1pdepends on Pip2p.We also noted thatGPD1 background levelson glycerol and oleate were drastically reduced in PIP2mutantcells, suggesting that the nonspecific association of Oaf1p tochromatin was lower in the absence of Pip2p. As a result, Oaf1prelative occupancy values in the pip2� strain were significantlyabove 1GDP1 unit under both glycerol and oleate growth con-ditions for all genes except IZH2 (Table 5, see RO rows). There-fore, we can conclude that Oaf1p is recruited to target geneseven in the absence of Pip2p, most likely in a form of ahomodimer, although at a much lower efficiency than theOaf1p-Pip2p heterodimer. Unfortunately, we were not able tomeasure the binding of Pip2p in the absence of Oaf1p becausethe PIP2 gene is expressed at extremely low levels in oaf1� cells(5, 6).5 I. V. Karpichev, J. M. Durand-Heredia, and G. M. Small, unpublished results.
TABLE 4Oaf1p-Pip2p sequential ChIPC values, or efficiencies of co-occupancy, in percentages, were calculated according to the following formula:C� 100(AB-A)/(A� B –A), whereA, B, andAB represent thefold enrichments for the first, second, and sequential ChIP, respectively (37). The threshold point for theAB/A ratio was taken to be 2.0 to call for an ensured co-occupancy.However, if theAB/A ratio was found to be in a range from 1.5 to 2.0, a possible co-occupancy was called if the correspondingC values were found to be in a range from 1.0to 100.0.
Direction of IP SamplesPOX1 PEX5 IZH2 QDR1 DUS3 YMR018w
C AB/A C AB/A C AB/A C AB/A C AB/A C AB/A% % % % % %
Oaf1p Glucose 58.1 2.11 58.9 17.51 420.7 23.75 23.47 1.34 107.5 25.63 166.4 9.78Then Glycerol 57.4 3.80 24.2 16.45 139.6 18.00 32.75 2.13 40.5 19.84 83.4 11.31Pip2p Oleate 22.5 3.04 11.9 8.63 56.6 7.16 9.23 1.42 17.9 10.70 23.1 7.52Pip2p Glucose 83.5 2.71 16.0 3.76 168.8 3.41 18.71 1.59 25.0 4.26 38.1 2.04Then Glycerol 29.9 1.79 5.7 2.91 47.3 3.20 26.33 2.38 9.0 3.01 25.2 2.79Oaf1p Oleate 16.3 1.69 4.1 2.22 22.9 2.31 14.94 2.21 6.2 2.50 11.3 3.27
TABLE 5Binding of Oaf1p to the promoters of target genes in the absence ofPip2pBinding in�pip2 strain is expressed as percentage of the corresponding IP efficiencyvalues in W3031A wild-type strain (IP�pip2/IPwt), relative occupancies (RO) in�pip2 strain are expressed inGPD1units usingGPD1 IP enrichment values found inthis strain.
Gene Oaf1p bindingin �pip2
MediaGlucose Glycerol Oleate
POX1 IP�pip2/IPwt 22.63 19.65 3.64 0.58 12.58 0.24RO 0.70 0.41 4.72 2.33 5.42 2.74
PIP2 IP�pip2/IPwt 6.54 4.49 1.48 0.43 4.06 1.74RO 1.26 0.11 5.94 3.48 6.36 4.94
PEX5 IP�pip2/IPwt 33.17 31.55 4.19 1.77 10.92 0.58RO 6.75 4.95 13.61 3.83 10.67 2.87
CTA1 IP�pip2/IPwt 49.69 47.81 11.54 9.79 20.78 1.88RO 2.4 0.25 35 30 28 11
IZH2 IP�pip2/IPwt 17.67 12.48 1.47 0.30 6.28 0.57RO 0.63 0.34 1.67 0.39 2.23 1.38
DUS3 IP�pip2/IPwt 49.35 33.48 9.35 3.67 19.89 4.82RO 3.18 0.92 27.92 17.01 19.89 13.51
GPD1 IP�pip2/IPwt 100.65 69.30 7.28 0.78 14.84 11.56RO 1.00 1.00 1.00
Transcriptional Control of Oleate Response in Yeast
10270 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 16 • APRIL 18, 2008
by guest on June 10, 2016http://w
ww
.jbc.org/D
ownloaded from

Adr1p Binds to the Promoters of Oaf1p and Pip2p TargetGenes under Derepression and Induction Conditions—Thepleiotropic activator Adr1p is required for oleate-dependentinduction of a number of genes involved in peroxisome func-tion (18–20, 26). To confirm that this transcription factor bindsto Oaf1p-Pip2p target genes in vivo, we created a strain thatexpresses a c-Myc-tagged allele of Adr1p (Adr1p-myc13). Weperformed a series of single ChIP experiments with lysates pre-pared from these cells and found that Adr1p binds to the type Iand type II genes listed in Table 2, with the exception of QDR1and PEX5 (Fig. 4 and Table 6). ForQDR1 and PEX5, the relativeoccupancy ranged up to 2 GPD1 units, suggesting that weakAdr1p binding to the promoters of these genes might havetaken place under oleate growth conditions. We did not detectbinding to genes belonging to the IZH family, and we onlyfound weak association (relative occupancy 2.5 GPD1 units orless) with genes harboring a nonfunctional ORE (DUS3 andSER1) (Table 6). Interestingly, Adr1p binding appears to beoleate-responsive for the majority of genes tested in this seriesof experiments.Is the Binding of Adr1p and the Oaf1p-Pip2p Complex
Dependent on One Another?—Adr1p binds to a sequence,referred to as UAS1, in target genes (14, 22). The UAS is oftenfound in close proximity or overlaps with the ORE of Oaf1p-Pip2p target genes (17, 19–21, 26). To investigate the relation-ship between Oaf1p, Pip2p, and Adr1p on the promoters oftarget genes, we first analyzed the binding of Oaf1p and Pip2pin anADR1 deletion strain. Single ChIP assays were carried outwith lysates prepared from Oaf1p-myc13/Pip2p-HA3 co-ex-pressing strain that lacked Adr1p (Table 1). We found thatthere was a decrease in Oaf1p binding to the majority of type Iand type II genes listed in Table 2, in the ADR1 deletion strain(Fig. 5A). However, for other genes, Oaf1p binding was eitherunaffected or enhanced (i.e. for PIP2) (Fig. 5B). For genes thatweakly bind Adr1p, or do not bind this transcription factor atall, Oaf1p binding remained unchanged (Fig. 5C). We alsonoted that binding of Pip2p was decreased for each gene ana-lyzed. For the PEX11 gene, the binding of bothOaf1p and Pip2p
was greatly decreased (�5-fold).These results indicate that theOaf1p-Pip2p complex is dependenton Adr1p for binding to targetgenes.We next investigatedwhether the
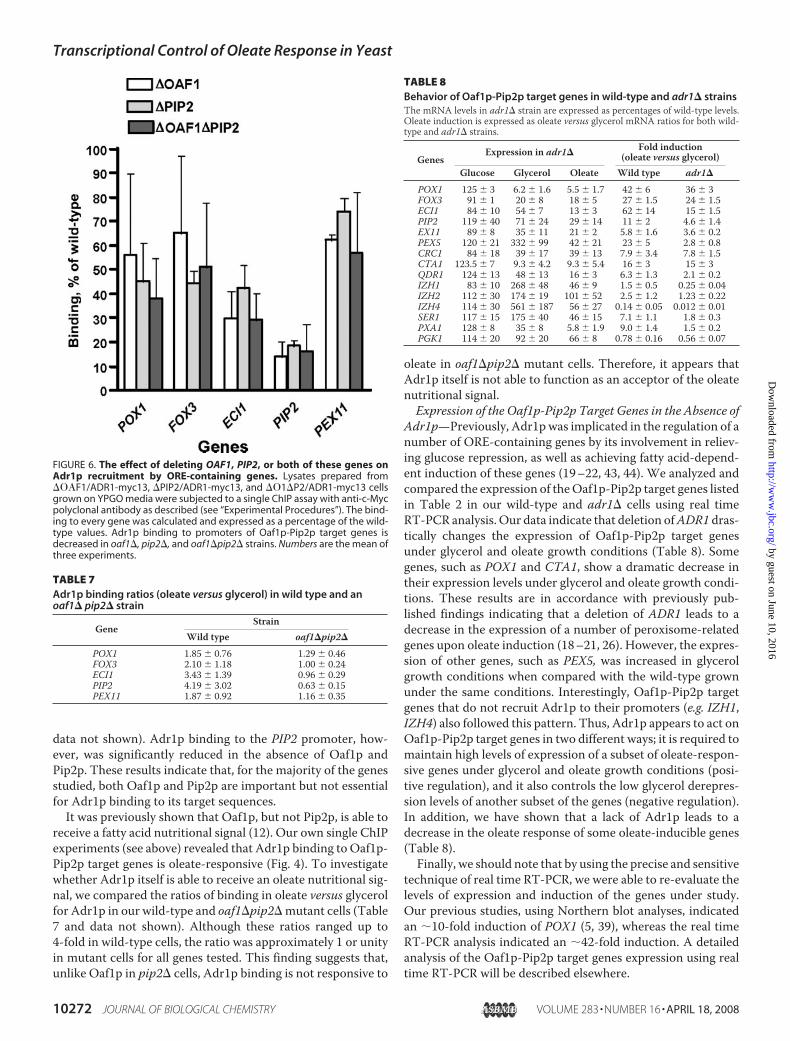
Oaf1p-Pip2p complex is importantfor Adr1p binding using the sameapproach. Three additional Adr1p-myc13-expressing strains that lackeither Oaf1p or Pip2p or both ofthese proteins were created for thispurpose (Table 1). These experi-ments showed that Adr1p bindingto target genes was moderatelyreduced compared with our “wild-type”Adr1p-myc13-expressing strainunder both glycerol and oleategrowth conditions for all genes withthe exception of PIP2 (Fig. 6 and
FIGURE 4. Association of Adr1p with promoters of Oaf1p-Pip2p targetgenes. Lysates were prepared from ADR1-myc13 cells grown on YPD, YPG,and YPGO media and used in single ChIP experiments with anti-c-Myc poly-clonal antibody as described (see “Experimental Procedures”). Adr1p binding(IP efficiency) was calculated as a percentage of gene-specific DNA pulleddown by c-Myc antibody from the lysate in a single-ChIP experiment. Bindingof Adr1p under oleate growth conditions was taken to be 100%. Numbers arethe mean of three experiments. Dex, glucose; Gly, glycerol; Ole, oleate.
FIGURE 5. The effect of deleting ADR1 on Oaf1p and Pip2p recruitment by target genes. Lysatesprepared from the �ADR1(PIP2-HA3/OAF1-myc13) strain grown on oleate were used in single ChIP exper-iments as described in Fig. 1. The binding of Oaf1p and Pip2p to each individual gene was calculated asdescribed in Fig. 1 and expressed as a percentage of the wild-type value (PIP2-HA3/OAF1-myc13 co-expressing strain). A, reduced recruitment of Oaf1p and Pip2p. B, unchanged or increased recruitment ofOaf1p and Pip2p. C, unchanged recruitment of Oaf1p and reduced recruitment of Pip2p. Numbers are themean of three experiments.
TABLE 6Relative occupancy of Adr1p on the promoters of Oaf1p-Pip2ptarget genesSee Table 3 for definition of GPD1 units.
GeneMedia
Glucose Glycerol OleatePOX1 1.04 0.16 3.49 2.14 5.64 2.64FOX3 1.36 0.40 2.74 1.48 4.80 1.57ECI1 1.03 0.34 1.36 0.61 4.06 0.60PIP2 1.55 1.31 2.66 2.60 7.45 3.75PEX11 0.90 0.16 1.43 0.64 2.31 0.62PEX5 0.95 0.18 1.04 0.58 1.87 0.52CRC1 0.92 0.78 5.02 2.00 5.01 0.87CTA1 1.05 0.32 4.68 2.41 7.10 1.34QDR1 1.15 0.26 1.08 0.44 1.32 0.67PXA1 1.27 0.15 8.98 5.08 7.05 1.20IZH1 1.00 0.16 0.62 0.11 0.70 0.30IZH2 0.95 0.46 0.42 0.24 0.61 0.24IZH4 0.95 0.16 0.83 0.19 0.73 0.21DUS3 1.49 0.18 1.30 0.32 1.60 0.37SER1 1.59 0.32 1.71 0.64 1.65 0.90GPD1 1.00 1.00 1.00
Transcriptional Control of Oleate Response in Yeast
APRIL 18, 2008 • VOLUME 283 • NUMBER 16 JOURNAL OF BIOLOGICAL CHEMISTRY 10271
by guest on June 10, 2016http://w
ww
.jbc.org/D
ownloaded from
data not shown). Adr1p binding to the PIP2 promoter, how-ever, was significantly reduced in the absence of Oaf1p andPip2p. These results indicate that, for the majority of the genesstudied, both Oaf1p and Pip2p are important but not essentialfor Adr1p binding to its target sequences.It was previously shown that Oaf1p, but not Pip2p, is able to
receive a fatty acid nutritional signal (12). Our own single ChIPexperiments (see above) revealed that Adr1p binding to Oaf1p-Pip2p target genes is oleate-responsive (Fig. 4). To investigatewhether Adr1p itself is able to receive an oleate nutritional sig-nal, we compared the ratios of binding in oleate versus glycerolfor Adr1p in our wild-type and oaf1�pip2�mutant cells (Table7 and data not shown). Although these ratios ranged up to4-fold in wild-type cells, the ratio was approximately 1 or unityin mutant cells for all genes tested. This finding suggests that,unlike Oaf1p in pip2� cells, Adr1p binding is not responsive to
oleate in oaf1�pip2� mutant cells. Therefore, it appears thatAdr1p itself is not able to function as an acceptor of the oleatenutritional signal.Expression of the Oaf1p-Pip2p Target Genes in the Absence of
Adr1p—Previously, Adr1pwas implicated in the regulation of anumber of ORE-containing genes by its involvement in reliev-ing glucose repression, as well as achieving fatty acid-depend-ent induction of these genes (19–22, 43, 44). We analyzed andcompared the expression of theOaf1p-Pip2p target genes listedin Table 2 in our wild-type and adr1� cells using real timeRT-PCR analysis. Our data indicate that deletion ofADR1 dras-tically changes the expression of Oaf1p-Pip2p target genesunder glycerol and oleate growth conditions (Table 8). Somegenes, such as POX1 and CTA1, show a dramatic decrease intheir expression levels under glycerol and oleate growth condi-tions. These results are in accordance with previously pub-lished findings indicating that a deletion of ADR1 leads to adecrease in the expression of a number of peroxisome-relatedgenes upon oleate induction (18–21, 26). However, the expres-sion of other genes, such as PEX5, was increased in glycerolgrowth conditions when compared with the wild-type grownunder the same conditions. Interestingly, Oaf1p-Pip2p targetgenes that do not recruit Adr1p to their promoters (e.g. IZH1,IZH4) also followed this pattern. Thus, Adr1p appears to act onOaf1p-Pip2p target genes in two different ways; it is required tomaintain high levels of expression of a subset of oleate-respon-sive genes under glycerol and oleate growth conditions (posi-tive regulation), and it also controls the low glycerol derepres-sion levels of another subset of the genes (negative regulation).In addition, we have shown that a lack of Adr1p leads to adecrease in the oleate response of some oleate-inducible genes(Table 8).Finally, we should note that by using the precise and sensitive
technique of real time RT-PCR, we were able to re-evaluate thelevels of expression and induction of the genes under study.Our previous studies, using Northern blot analyses, indicatedan �10-fold induction of POX1 (5, 39), whereas the real timeRT-PCR analysis indicated an �42-fold induction. A detailedanalysis of the Oaf1p-Pip2p target genes expression using realtime RT-PCR will be described elsewhere.
FIGURE 6. The effect of deleting OAF1, PIP2, or both of these genes onAdr1p recruitment by ORE-containing genes. Lysates prepared from��F1/ADR1-myc13, �PIP2/ADR1-myc13, and ��1�P2/ADR1-myc13 cellsgrown on YPGO media were subjected to a single ChIP assay with anti-c-Mycpolyclonal antibody as described (see “Experimental Procedures”). The bind-ing to every gene was calculated and expressed as a percentage of the wild-type values. Adr1p binding to promoters of Oaf1p-Pip2p target genes isdecreased in oaf1�, pip2�, and oaf1�pip2� strains. Numbers are the mean ofthree experiments.
TABLE 7Adr1p binding ratios (oleate versus glycerol) in wild type and anoaf1� pip2� strain
GeneStrain
Wild type oaf1�pip2�
POX1 1.85 0.76 1.29 0.46FOX3 2.10 1.18 1.00 0.24ECI1 3.43 1.39 0.96 0.29PIP2 4.19 3.02 0.63 0.15PEX11 1.87 0.92 1.16 0.35
TABLE 8Behavior of Oaf1p-Pip2p target genes in wild-type and adr1� strainsThe mRNA levels in adr1� strain are expressed as percentages of wild-type levels.Oleate induction is expressed as oleate versus glycerol mRNA ratios for both wild-type and adr1� strains.
GenesExpression in adr1�
Fold induction(oleate versus glycerol)
Glucose Glycerol Oleate Wild type adr1�
POX1 125 3 6.2 1.6 5.5 1.7 42 6 36 3FOX3 91 1 20 8 18 5 27 1.5 24 1.5ECI1 84 10 54 7 13 3 62 14 15 1.5PIP2 119 40 71 24 29 14 11 2 4.6 1.4EX11 89 8 35 11 21 2 5.8 1.6 3.6 0.2PEX5 120 21 332 99 42 21 23 5 2.8 0.8CRC1 84 18 39 17 39 13 7.9 3.4 7.8 1.5CTA1 123.5 7 9.3 4.2 9.3 5.4 16 3 15 3QDR1 124 13 48 13 16 3 6.3 1.3 2.1 0.2IZH1 83 10 268 48 46 9 1.5 0.5 0.25 0.04IZH2 112 30 174 19 101 52 2.5 1.2 1.23 0.22IZH4 114 30 561 187 56 27 0.14 0.05 0.012 0.01SER1 117 15 175 40 46 15 7.1 1.1 1.8 0.3PXA1 128 8 35 8 5.8 1.9 9.0 1.4 1.5 0.2PGK1 114 20 92 20 66 8 0.78 0.16 0.56 0.07
Transcriptional Control of Oleate Response in Yeast
10272 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 16 • APRIL 18, 2008
by guest on June 10, 2016http://w
ww
.jbc.org/D
ownloaded from

DISCUSSION
In this study we analyzed the binding of the Oaf1p-Pip2ptranscription factors to promoters of target genes in vivo. Usingquantitative single ChIP assays on a single gene basis, we haveshown that bothOaf1p andPip2p bind to their target sequencesunder glucose, glycerol, and oleate growth conditions in vivo,although this binding was much lower on glucose when com-pared with glycerol or oleate. This should not come as a sur-prise, because Oaf1p and Pip2p are constantly localized to thenucleus (17), and the OREs to which they bind usually reside innucleosome-free regions and remain accessible to them irre-spective of the growth conditions (45).The patterns of binding for both Oaf1p and Pip2p were sim-
ilar for the majority of the genes tested (with only three excep-tions, see “Results”). The pattern of binding mimicked that ofmRNA expression for genes that are only moderately inducedby oleate, but not for highly inducible genes (5). Many genesthat are under control of the Oaf1p-Pip2p complex are regu-lated by two distinct mechanisms, glucose repression andoleate induction.We have shown here that a shift from glucoseto glycerol media causes a large increase in binding of theOaf1p-Pip2p complex to its response element, with only amod-erate further increase in the amount of binding when cells areshifted to inducing conditions (Fig. 1). What could trigger thisincrease in binding upon glycerol derepression?We have dem-onstrated thatOaf1p undergoes phosphorylation in cells grownon glycerol or oleate media and that Snf1p kinase does notappear to be essential for this phosphorylation. Based on thesefindings, and by analogy with other yeast transcription factors(41), we hypothesize that this post-translational modificationcauses the increase in the Oaf1p-Pip2p binding under glucosederepression. Upon receiving a fatty acid nutritional cue, Oaf1pappears to undergo an additional phosphorylation and that, inturn, might further increase Oaf1p-Pip2p binding to anORE. Itis also possible that this additional phosphorylation causes aconformational change in the Oaf1p-Pip2p complex that leadsto unmasking its transcriptional activity. It remains unclearwhether Pip2p, the Oaf1p counterpart, is similarly phosphoryl-ated and what role this post-translational modification mayplay in regulating transcriptional activity of the Oaf1p-Pip2pcomplex upon glucose derepression and oleate induction. Apossible role of Adr1p phosphorylation is discussed below. Thenature of a putative protein kinase(s) involved in the phospho-rylation of both Oaf1p and Pip2p remains to be determined.In addition to the genes that contain obvious OREs in their
promoters, we were able to detect Oaf1p-Pip2p binding to twogenes that lack a clearly defined ORE. The minimal consensusORE is defined as CGGN3TN(A/R)N8–12CCG (17), but per-haps this definition is too restrictive. A genome-wide search forall Oaf1p-Pip2p-binding sites is required to define a consensusORE that is physiologically meaningful. Furthermore, it is pos-sible that interaction of the Oaf1p-Pip2p transcription factorswith a target DNA sequence is more complex than simple pro-tein-DNA binding, and it may also involve interaction of thiscomplex with proteins associated with nucleosome-free OREs(45). In this study we also found that some OREs previouslydefined to be nonfunctional (5) appear to recruit the Oaf1p-
Pip2p complex. In this case it is possible that Pip2p, rather thanOaf1p, plays a role as an acceptor of a nutritional signal becauseboth of these proteins have ligand binding domains (11), butonly Oaf1p binds free fatty acids. The nature of any addi-tional signal, as well as the role of Oaf1p-Pip2p in the regu-lation of genes harboring “nonfunctional” OREs, has yet tobe determined.We and others previously postulated that the Oaf1p-Pip2p
complex acts in a formof a heterodimer to regulate target genes(6, 7). The mammalian transcription factors, peroxisome pro-liferator-activated receptor � and retinoic acid receptor �,dimerize and subsequently bind to peroxisome proliferator-re-sponse elements of genes encoding peroxisomal proteins onlyupon ligand binding (8–10). By analogy, it has also been pro-posed that the Oaf1p-Pip2p complex binds to OREs uponreceiving a nutritional signal (5–7), although the possibility ofOaf1p and Pip2p being permanently bound to OREs was alsoraised (17, 41). We show here that Oaf1p and Pip2p co-occupythe promoter regions of target genes in vivo, that they bind tomost of the target sequences at an equimolar ratio, and that thebinding of one protein appears to be dependent on the other. Inaddition, our results indicate that target genes preferablyrecruit the Oaf1p-Pip2p heterodimer rather than an Oaf1p-Oaf1p homodimer. These findings strongly support the currentview that the Oaf1p-Pip2p transcription factors act predomi-nantly as a heterodimer on target genes. It appears, however,that homodimers of Oaf1p may play an additional role inregulating certain genes such as QDR1, CTA1, and the IZHgenes. This is in accordance with our previous finding thatOaf1p is capable of mediating partial oleate-dependentinduction of CTA1 and QDR1 as well as maintaining sub-wild-type levels of IZH2 expression under glucose repressionin the absence of Pip2p (5). It is possible that both Oaf1p-Oaf1p homodimers and Oaf1p-Pip2p heterodimers controlthe expression of these genes.Our findings that the Oaf1p-Pip2p complex is able to bind to
DNA without receiving an oleate nutritional signal raises thequestion as to whether both forms (DNA-bound and DNA-free) of this complex are capable of receiving this signal. Othertranscription activators, such as Put3p and Leu3p, althoughalways bound to a promoter are controlled by direct interactionwith lowmolecular weight effectors (41, 46–49). In light of therecent identification of theOaf1p ligand-binding domain that iscapable of binding free fatty acids (11), it now seems likely thattranscriptional activity of the Oaf1p-Pip2p complex is con-trolled by a similar mechanism, rather than by the peroxisomeproliferator-activated receptor �-retinoic acid receptor �model of regulation.In addition to theOaf1p-Pip2p complex, Adr1p is involved in
the regulation ofmany peroxisome-related genes via binding toits resident target UAS1 sequences in their promoters (18–21).This binding was previously demonstrated in vivo using ChIPassays (26–28); however, none of these studies were performedunder oleate induction conditions, and there have been no suchstudies focused on Adr1p interactions with the Oaf1p-Pip2pcomplex.We showhere thatAdr1p binds in vivo to the promot-ers of Oaf1p-Pip2p target genes (with the exception of the IZHgenes) under glucose derepression and oleate induction condi-
Transcriptional Control of Oleate Response in Yeast
APRIL 18, 2008 • VOLUME 283 • NUMBER 16 JOURNAL OF BIOLOGICAL CHEMISTRY 10273
by guest on June 10, 2016http://w
ww
.jbc.org/D
ownloaded from

tions. Because the expression of peroxisome-related genesappears to be impaired inADR1mutant cells (17), we also askedwhether Adr1p acts on these genes by mediating the Oaf1p-Pip2p binding to their OREs. Indeed, the association of Oaf1pand Pip2p with OREs was lower in the adr1� mutant cells thenin our wild-type cells for most of the genes studied. It is note-worthy that binding of these proteins to PEX11 was mostreduced in adr1� cells. Because Pex11p is a key regulator ofperoxisome proliferation, this supports the involvement ofAdr1p in controlling this process (19).As discussed above, Adr1p generally acts as a positive regu-
lator of Oaf1p-Pip2p binding. However, the association ofOaf1p to the PIP2 promoter was increased in ADR1 mutantcells, suggesting that Adr1p regulates Oaf1p-Pip2p binding tothis promoter in a negative fashion. BecausePIP2plays a uniqueregulatory role among fatty acid-responsive genes, it is perhapsnot surprising that Oaf1p-Pip2p binding to the PIP2 promoteris also regulated in a distinctive fashion.We also analyzed the importance of the Oaf1p-Pip2p com-
plex for Adr1p binding and found that this complex is impor-tant but not essential. Our results also show that Adr1p bindingto thePIP2OREappears to be under the stringent control of theOaf1p-Pip2p complex. It is interesting to note that Adr1p bindsto the majority of Oaf1p-Pip2p target genes in an oleate-de-pendent fashion in wild-type cells, but it loses its ability torespond to oleate in oaf1�pip2� mutant cells. Thus, it appearsthat this protein is not capable of receiving an oleate nutritionalsignal independently, but rather it receives such a signalthrough theOaf1p-Pip2p complex. The issue of whetherAdr1preceives an oleate nutritional signal directly (i.e. by physicallyinteracting with the Oaf1p-Pip2p complex) or indirectly (i.e. byinteracting with another intermediate factor(s)) remains to beaddressed.Adr1p positively regulates the expression of Oaf1p-Pip2p
target genes, although the level of regulation is variable (17).Our findings indicate that Adr1p is required for achieving themaximum expression of these genes when oleate is supplied inthe growthmedia. This raises the possibility that, in addition tocatabolite derepression, Adr1p might also be involved in theoleate induction of Oaf1p-Pip2p target genes. Interestingly,when ADR1 is disrupted, the response to oleate is reduced in anumber of these genes, whereas others respond normally.Together, these results suggest that Adr1p is directly involvedin oleate induction of at least some target genes, includinghighly inducible genes such as ECI1 and PIP2. Because anincrease in Adr1p binding occurs under oleate growth condi-tions, it is reasonable to speculate that such an increase contrib-utes to the up-regulation of Oaf1p-Pip2p target genes seenupon induction. However, the oleate induction is not com-pletely abolished in the adr1� strain; therefore, there must beadditional mechanism(s) involved in this up-regulation. Onepossibility might be the recruitment of chromatin remodelingfactors by the Oaf1p-Pip2p complex and/or by Adr1p uponreceiving a nutritional cue.The mechanism controlling Adr1p binding upon switching
from glycerol- to oleate-containing media remains unclear.Based on the finding that the Snf1p kinase is required for Adr1pto bind to DNA, it has been suggested that initial binding of
Adr1p to UAS1 under derepression conditions is because of itsphosphorylation by the Snf1p kinase (see Ref. 17 and referencestherein). It is possible that Adr1p undergoes additional phos-phorylation under oleate induction, and such phosphorylationis accountable for the increase in its binding. It is also clear thatthe regulatory role of Adr1p is complex and variable for differ-ent genes. For example, the expression of PEX5 is increased incells lacking Adr1p when cells are grown in the presence ofglycerol, but it decreased when the cells are switched to mediacontaining oleate. Thus, Adr1p appears to act as a positive ornegative regulator, depending on the environmental condi-tions. It is likely that the positive regulation includes the stim-ulation of Oaf1p-Pip2p binding to a target sequence by Adr1punder both glucose repression and oleate induction. However,the mechanism by which Adr1p negatively regulates Oaf1p-Pip2p target genes remains to be investigated.The results obtained during the course of this study, together
with previous findings from our group and others, lead us topropose amodel ofOaf1p-Pip2p andAdr1p action that inducesthe expression of oleate-responsive genes. When cells aregrown in the presence of glucose, Pip2p is expressed atextremely low levels, whereas Oaf1p is constitutively expressedat higher levels. Under these conditions, Pip2p is most likelypresent exclusively in the formof heterodimerswithOaf1p, andthese heterodimers bind only weakly to the ORE. Under thesame glucose repression conditions, Adr1p is not able to bind toits target sequence UAS1. Together this results in a very lowlevel of expression of oleate-responsive genes in this nutritionalenvironment. Upon derepression, Oaf1p and perhaps Pip2pundergo phosphorylation, and this post-translationalmodifica-tion greatly increases Oaf1p-Pip2p binding to the ORE and ini-tiates a higher expression of PIP2 by an autoregulatory mecha-nism. Adr1p also binds to UAS1 under derepressionconditions, probably because of its phosphorylation. This bind-ing increases Oaf1p-Pip2p association with the ORE whereasthis complex, in turn, positively influences Adr1p recruitmentto UAS1. The presence of both Oaf1p-Pip2p and Adr1p on thepromoters of oleate-responsive genes turns on the transcrip-tional machinery and allows it to drive the expression of targetgenes to the moderate derepression levels. When oleate is sup-plied into glycerol-containingmedia,Oaf1p receives a fatty acidnutritional signal and appears to undergo additional phospho-rylation. This causes a further moderate increase in Oaf1p-Pip2p binding to OREs. The oleate nutritional cue turns theDNA-bound Oaf1p-Pip2p heterodimer to the active transcrip-tional state, probably via a conformational change because of itsphosphorylation. It is possible that Adr1p also undergoes addi-tional phosphorylation upon receiving a fatty acid nutritionalsignal from the Oaf1p-Pip2p complex, and this phosphoryla-tion causes an increase in its binding. All of these eventsincrease the level of expression of the target genes, includingPIP2, and trigger the autoregulatory mechanism to achievemaximal transcription under oleate induction conditions.While this manuscript was under review, Smith et al. (50)
published a report describing a systematic analysis of theresponse to fatty acids in yeast, using a combination of genome-wide chromatin localization and expression analyses. In theirstudy, the authors investigated a regulatory network of four
Transcriptional Control of Oleate Response in Yeast
10274 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 16 • APRIL 18, 2008
by guest on June 10, 2016http://w
ww
.jbc.org/D
ownloaded from

transcription factors, Oaf1p, Pip2p, Adr1p, and an additionalnegative regulator of the oleate response Ykr064wp, namedOaf3p (50). The authors demonstrated that this regulatory net-work expanded, and there was more combinatorial control, inthe presence of oleate. These results are in accordance with ourdata that demonstrate an increase in Oaf1p-Pip2p associationwith target genes, as well as new interactions of Adr1p withtarget sequences, upon switching fromglucose to oleate growthconditions.We should note, however, that the systematic anal-ysis of Smith et al. (50) did not detect some well establishedinteractions such as the association of the Oaf1p-Pip2p com-plex with the POX1 ORE or Adr1p with the POX1 UAS1 (5, 6,19, 39, 42). Clearly, additional studies are necessary to charac-terize transcriptional regulatory networks of yeast response tofatty acids and the details of the precise mechanism by which afatty acid nutritional signal causes transcriptional activation oftarget genes.
Acknowledgments—We gratefully acknowledge Alexandra Obregon,Vitaly Zyhadlo, and Juncheng Li for excellent technical assistance.Wealso thank Drs. Avrom Caplan and Gintaras Deikus for reading andhelpfully discussing the manuscript.
REFERENCES1. Gancedo, J. M. (1998)Microbiol. Mol. Biol. Rev. 62, 334–3612. Schuller, H. J. (2003) Curr. Genet. 43, 139–1603. Veenhuis, M., Mateblowski, M., Kunau, W. H., and Harder, W. (1987)
Yeast 3, 77–844. Einerhand, A. W. C., Kos, W. T., Distel, B., and Tabak, H. F. (1993) Eur.
J. Biochem. 314, 323–3315. Karpichev, I. V., and Small, G. M. (1998)Mol. Cell. Biol. 18, 6560–65706. Karpichev, I. V., Luo, Y., Marians, R. C., and Small, G.M. (1997)Mol. Cell.
Biol. 17, 69–807. Rottensteiner, H., Kal, A. J., Hamilton, B., and Tabak, H. F. (1997) Eur.
J. Biochem. 247, 776–7838. Issemann, I., and Green, S. (1990) Nature 347, 645–6509. Issemann, I., Prince, R. A., Tugwood, J. D., and Green, S. (1993) J. Mol.
Endocrinol. 11, 37–4710. Reddy, J. K., and Hashimoto, T. (2001) Annu. Rev. Nutr. 21, 193–23011. Phelps, C., Gburcik, V., Suslova, E., Dudek, P., Forafonov, F., Bot, N.,
MacLean, M., Fagan, R. J., and Picard, D. (2006) Proc. Natl. Acad. Sci.U. S. A. 103, 7077–7081
12. Baumgartner, U., Hamilton, B., Piskacek, M., Ruis, H., and Rottensteiner,H. (1999) J. Biol. Chem. 274, 22208–22216
13. Rottensteiner, H., Kal, A. J., Filpits, M., Binder, M., Hamilton, B., Tabak,H. F., and Ruis, H. (1996) EMBO J. 15, 2924–2934
14. Smith, J. J., Marelli, M., Christmas, R. H., Vizeacoumar, F. J., Dilworth,D. J., Ideker, T., Galitski, T., Dimitrov, K., Rachubinski, R. A., and Aitchi-son, J. (2002) J. Cell Biol. 158, 259–271
15. Kal, A. J., van Zonneveld, A. J., Benes, V., van den Berg, M., Koerkamp,M. G., Albermann, K., Strack, N., Ruijter, J. M., Richter, A., Dujon, B.,Ansorge, W., and Tabak, H. F. (1999)Mol. Biol. Cell 10, 1859–1872
16. Koerkamp, M. G., Rep, M., Bussemaker, H. J., Hardy, G. P., Mul, A., Piek-arska, K., Szigyarto, C. A., De Mattos, J. M., and Tabak, H. F. (2002)Mol.Biol. Cell 13, 2783–2794
17. Gurvitz, A., and Rottensteiner, H. (2006) Biochim. Biophys. Acta 1763,1392–1402
18. Simon, M., Adam, G., Rapatz, W., Spevak, W., and Ruis, H. (1991) Mol.Cell. Biol. 11, 699–704
19. Gurvitz, A., Hiltunenm, J. K., Erdmann, R., Hamilton, B., Hartig, A., Ruis,H., and Rottensteiner, H. (2001) J. Biol. Chem. 276, 31825–31830
20. Gurvitz, A., Wabnegger, L., Rottensteiner, H., Dawes, I. W., Hartig, A.,Ruis, H., and Hamilton, B. (2000)Mol. Cell. Biol. Res. Commun. 4, 81–89
21. Rottensteiner, H., Wabnegger, L., Erdmann, R., Hamilton, B., Ruis, H.,Hartig, A., and Gurvitz, A. (2003) J. Biol. Chem. 278, 27605–27611
22. Cheng, C., Kacherovsky, N., Dombek, K. M., Camier, S., Thukral, S. K.,Rhim, E., and Young, E. T. (1994)Mol. Cell. Biol. 14, 3842–3852
23. Ciriacy, M. (1975)Mol. Gen. Genet. 138, 157–16424. Denis, C. L., and Young, E. T. (1983)Mol. Cell. Biol. 3, 360–37025. Simon, M., Binder, M., Adam, G., Hartig, A., and Ruis, H. (1992) Yeast 8,
303–30926. Young, E. T., Dombek, K. M., Tachibana, C., and Ideker, T. (2003) J. Biol.
Chem. 278, 26146–2615827. Harbison, C. T., Gordon, D. B., Lee, T. I., Rinaldi, N. J., Macisaac, K. D.,
Danford, T. W., Hannett, N. M., Tagne, J. B., Reynolds, D. B., Yoo, J.,Jennings, E. G., Zeitlinger, J., Pokholok, D. K., Kellis, M., Rolfe, P. A.,Takusagawa, K. T., Lander, E. S., Gifford, D. K., Fraenkel, E., and Young, R.(2004) Nature 431, 99–104
28. Tachibana, C., Yoo, J. Y., Tagne, J. B., Kacherovsky, N., Lee, T. I., andYoung, E. (2005)Mol. Cell. Biol. 25, 2138–2146
29. Lee, T. I., Rinaldi, N. J., Robert, F., Odom, D. T., Bar-Joseph, Z., Gerber,G. K., Hannett, N. M., Harbison, C. T., Thompson, C. M., Simon, I.,Zeitlinger, J., Jennings, E. G.,Murray, H. L., Gordon, D. B., Ren, B.,Wyrick,J. J., Tagne, J. B., Volkert, T. L., Fraenkel, E., Gifford, D. K., andYoung, R. A.(2002) Science 298, 763–764
30. Ren, B., Robert, F., Wyrick, J. J., Aparicio, O., Jennings, E. G., Simon, I.,Zeitlinger, J., Schreiber, J., Hannett, N., Kanin, E., Volkert, T. L., Wilson,C. J., Bell, S. P., and Young, R. A. (2000) Science 290, 2306–2309
31. Gunji, W., Kai, T., Takahashi, Y., Maki, Y., Kurihara, W., Utsugi, T., Fuji-mori, F., and Murakami, Y. (2004) DNA Res. 11, 163–177
32. Aparicio, O., Geisberg, J. V., Sekinger, E., Yang, A., Moqtaderi, Z., andStruhl, K. (2005) inCurrent Protocols inMolecular Biology (Ausubel, F.M.,Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. G., Smith, J. A., andStruhl, K., eds) pp. 21.3.1–21.3.33, John Wiley & Sons, Inc., New York
33. Orlando, V. (2000) Trends Biochem. Sci. 25, 99–10434. Robert, F., Pokholok, D. K., Hannett, N. M., Rinaldi, N. J., Chandy, M.,
Rolfe, A.,Workman, J. L., Gifford, D. K., andYoung, R. (2004)Mol. Cell 16,199–209
35. Karpichev, I. V., Cornivelli, L., and Small, G. M. (2002) J. Biol. Chem. 277,19609–19617
36. Sikorski, R. S., and Hieter, P. (1989) Genetics 122, 19–2737. Bahler, J., Wu, J.-Q., Longtine, M., Shan, N. G., Amos, M., Steever, A. B.,
Wach, A., Philippsen, P., and Pringle, J. R. (1998) Yeast 14, 943–95138. Geisberg, J. V., and Struhl, K. (2005) in Current Protocols in Molecular
Biology (Ausubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman,J. G., Smith, J. A., and Struhl, K., eds) pp. 21.8.1–21.8.7, JohnWiley & Sons,Inc., New York
39. Luo, Y., Karpichev, I. V., Kohanski, R. A., and Small, G. M. (1996) J. Biol.Chem. 271, 12068–12075
40. Skoneczny, M., Chelstowska, A., and Rytka, J. (1988) Eur. J. Biochem. 174,297–302
41. MacPherson, S., Larochelle, M., and Turcotte, B. (2006) Microbiol. Mol.Biol. Rev. 70, 583–604
42. Luo, Y. (1997) Molecular Mechanisms Regulating Transcription of theGene Encoding Peroxisomal Acyl-CoA Oxidase in Saccharomyces cerevi-siae. Ph.D. dissertation, Mount Sinai School of Medicine, New York
43. Filipits, M., Simon, M. M., Rapatz, W., Hamilton, B., and Ruis, H. (1993)Gene (Amst.) 132, 49–55
44. Simon, M. M., Pavlik, P., Hartig, A., Binder, M., Ruis, H., Cook, W. J.,Denis, C. L., and Schanz, B. (1995)Mol. Gen. Genet. 249, 289–296
45. Agricola, E., Verdone, L., Xella, B., Di Mauro, E., and Caserta, M. (2004)Biochemistry 43, 8878–8884
46. Sellick, C. A., and Reece, R. (2005) Trends Biochem. Sci. 30, 405–41247. Sellick, C. A., and Reece, R. (2003) EMBO J. 22, 5147–515348. Kirkpatrick, C. R., and Schimmel, P. (1995)Mol. Cell. Biol. 15, 4021–403049. Sze, J. Y., Woontner, M., Jaehning, J. A., and Kohlhaw, G. (1992) Science
258, 1143–114550. Smith, J. J., Ramsey, S. A., Marelli, M., Marzolf, B., Hwang, D., Saleem,
R. A., Rachubinski, R. A., and Aitchison, J. D. (2007)Mol. Systems Biol. 3,1–14
51. Thomas, B. J., and Rothstein, R. (1989) Cell 56, 619–630
Transcriptional Control of Oleate Response in Yeast
APRIL 18, 2008 • VOLUME 283 • NUMBER 16 JOURNAL OF BIOLOGICAL CHEMISTRY 10275
by guest on June 10, 2016http://w
ww
.jbc.org/D
ownloaded from

Igor V. Karpichev, Jorge M. Durand-Heredia, Yi Luo and Gillian M. SmallSaccharomyces cerevisiaeControlling Oleate-responsive Genes in
Binding Characteristics and Regulatory Mechanisms of the Transcription Factors
doi: 10.1074/jbc.M708215200 originally published online February 19, 20082008, 283:10264-10275.J. Biol. Chem.
10.1074/jbc.M708215200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/283/16/10264.full.html#ref-list-1
This article cites 48 references, 25 of which can be accessed free at
by guest on June 10, 2016http://w
ww
.jbc.org/D
ownloaded from











![[123doc vn] - tim-hieu-nam-men-saccharomyces-cerevisiae](https://static.fdokumen.com/doc/165x107/6345cd51f474639c9b0502af/123doc-vn-tim-hieu-nam-men-saccharomyces-cerevisiae.jpg)








