
Impacts of operating conditions and solution chemistry on osmotic membrane structure and performance
Upload
independentCategory
view
1download
0
Beyond the ionic and osmotic response to salinityin Chenopodium quinoa: functional elementsof successful halophytism
Francesco OrsiniA, Mattia AccorsiA, Giorgio GianquintoA, Giovanni DinelliA,Fabiana AntognoniB, Karina B. Ruiz CarrascoC, Enrique A. MartinezD,E,Mohammad AlnayefA, Ilaria MarottiA, Sara BosiA and Stefania BiondiA,F
ADipartimento di Scienze e Tecnologie Agroambientali, (DiSTA), Università di Bologna, viale Fanin 44,40127 Bologna, Italy.
BDipartimentodiBiologia EvoluzionisticaSperimentale,Università diBologna, via Irnerio42,40126Bologna, Italy.CDipartimento di Colture Arboree, Università di Bologna, viale Fanin 44, 40127 Bologna, Italy.DCentro de Estudios Avanzados en Zonas Aridas (CEAZA), Av. Raúl Bitrán s/n, La Serena, Chile.EApplied Biology and Ecology, Universidad Catolica del Norte, Coquimbo, Chile.FCorresponding author. Email: [email protected]
Abstract. Chenopodium quinoaWilld. (quinoa) is a halophyte for which some parameters linked to salt tolerance havebeen investigated separately in different genotypes and under different growth conditions. In this study, severalmorphological and metabolic responses were analysed in parallel after exposure to salinity. In vitro seed germinationwas initially delayedbya150mMNaCl treatment but eventually reached the same level as the control (0mMNaCl),whereasseedling root growth was enhanced; both parameters were moderately inhibited (~35–50%) by 300mMNaCl. In pot grownplants, plant size was reduced by increasing salinity (0–750mM NaCl). Transpiration and stomatal conductance weredecreased at the highest salinity levels tested, consistent with reduced stomatal density and size. The density of epidermalbladder cells (EBCs) on the leaf surface remained unaffected up to 600mMNaCl. Tissue contents of Na+ and Cl– increaseddramatically with salt treatment, but resulted in only a 50% increase in Na+ from 150 to 750mM NaCl. Internal K+ wasunaffected up to 450mMNaCl but increased at the highest salinity levels tested. Excretion through sequestration into EBCswas limited (generally �20%) for all ions. A modest dose-dependent proline accumulation, and concomitant reduction intotal polyamines and putrescine efflux occurred in NaCl-treated plants. Results confirm the importance of inorganic ions forosmotic adjustment, the plant’s ability to maintain K+ levels and the involvement of putrescine efflux in maintaining ionicbalance under high salinity conditions. Conversely, ion excretion and proline appear to play a minor role. Taken togetherthese results indicate which parameters could be used for future comparison among different genotypes.
Additional keywords: ion homeostasis, osmoprotectant, salt glands.
Introduction
Salinity is among themost widespread and relevant constraints inirrigated agriculture, affecting up to 7%and about one-third of theworld’s total and irrigated land respectively (Flowers andColmer2008). Thus, salt tolerance is an agronomically important trait thatis receiving increasing attention among scientists worldwide.Since salt tolerance is a multigenic trait, transgenic approachesbased on plant transformation with candidate genes have hadscant success. Tolerance to abiotic stresses is, indeed, a complextrait both from a genetic and a physiological point of view andsearching for genotypes with particularly high resistance canbenefit breeding programmes. In order to address this issue, it isdeemed useful to investigate the basic mechanisms that conferresistance to salt stress in tolerant species or ecotypes, in order to
identify specific traits associated with salt tolerance that may beselected in established crop plants.
Plants exposed to salt stress have to function despite severalconstraints: accumulation of toxic ions (especially Na+ and Cl–),a decrease in the external soil water potential (including aphysiological drought within the stressed tissues), oxidativestress and cytosolic K+ depletion (Munns 2002; Demidchiket al. 2010). Accumulation of ‘compatible solutes’, such asglycinebetaine and proline, is one of the best knownmechanisms for osmotic adjustment in plants exposed to salt ordrought stress, including halophytes (Flowers and Colmer 2008).The accumulation, biosynthesis, degradation, translocation androle of proline during salinity stress has been investigatedthoroughly (Kavi Kishor et al. 2005). In addition to its role as an
CSIRO PUBLISHING
www.publish.csiro.au/journals/fpb Functional Plant Biology, 2011, 38, 818–831
� CSIRO 2011 10.1071/FP11088 1445-4408/11/100818

osmolyte necessary for cell expansion under stress conditions,proline also acts as both as an osmoprotectant – protectingsubcellular structures and macromolecules – and as a signalmolecule (Matysik et al. 2002; Szabados and Savouré 2010).
Sequestration of the absorbed ions into specialised trichomes,generally known as salt glands or bladders, at the leaf and stemsurface often appears to be an efficient strategy contributing tosalinity resistance in some xero-halophytic species (Agarie et al.2007; Ben Hassine et al. 2009). These structures are involved incompartmentalising salt, thereby excluding it from the other leaftissues, in particular, from the underlying photosyntheticallyactive mesophyll. In chenopods, these salt glands are knownas epidermal bladder cells (EBCs) (Liphschitz andWaisel 1982).In a comparative study of an inland and a coastal populationof Atriplex halimus L., Ben Hassine et al. (2009) showed thatexogenous abscisic acid (ABA) improved NaCl resistance in thedrought tolerant inland population through an increase in Na+
excretion through the leaf bladders. These authors also providedevidence suggesting that polyamines (PAs) are involved in thesalt excretion process.
Further, PAs, of which putrescine (Put), spermidine (Spd)and spermine (Spm) are the most common in higher plants, arealiphatic polycations regarded as plant growth regulatorsinvolved in both development (Kusano et al. 2008) and stressresponses (Alcazar et al. 2010). There is a substantial amount ofevidence supporting the idea that PAs exert a protective functionduring stress (e.g. scavenging of reactive oxygen species,membrane stabilisation and cell wall stiffening); in salt stressthey also seem to play an important role as regulators of ionchannels (Kusano et al. 2008).
Quinoa (Chenopodium quinoa Willd.) is a grain like cropwhose cultivation started in the Andean regions more than7000 years ago (Jacobsen et al. 2003). It belongs toAmaranthaceae (formerly Chenopodiaceae), a family knownto comprise the highest proportion of halophytic genera. Theplant has been selected by the UN Food and AgricultureOrganisation (FAO) as one of the crops destined to offerfood security in this century, both for its excellent nutritionalfeatures and for its remarkable tolerance against several abioticstresses such as frost (Rosa et al. 2009), salinity (Delatorre-Herrera and Pinto 2009) and drought (Bosque-Sanchez et al.2003).
Indeed, the scientific interest towards quinoa has increasedexponentially in recent years and several authors have proposedits use as a model halophytic plant for unravelling the basicmechanisms involved in salt tolerance.Todate, several genotypeshave been investigated independently, exposed to differentsalinity levels but under different, therefore, not comparableconditions. Results range from germination delay (cv. Sajama,Ruffino et al. 2010), adaptation of physiological functions suchas photosynthesis (cv. Kankolla, Jensen et al. 2000), changes instomatal conductance (cv. Real, Bosque-Sanchez et al. 2003),regulation of plant water status (cv. Kankolla, Jensen et al.2000), accumulation of osmoprotectants (cv. Sajama, Ruffinoet al. 2010) and ion partitioning (cv.AndeanHybrid,Wilson et al.2002; cv.Amarilla,Roja,Hueque andPucura, Delatorre-Herreraand Pinto 2009; cv. 5206, Hariadi et al. 2011). However, acomprehensive study that simultaneously investigates theinfluence of salinity on the different elements is lacking.
Improving quinoa productivity is a key issue for this importantAndean staple crop and the success of breeding programmeswill rely heavily on germplasm evaluation.Most previous studieson salt or drought stress in quinoa have been performed onBolivian or Peruvian cultivars, but a single comparativeanalysis between genotypes exhibiting different levels ofsalinity tolerance is yet to be conducted. In Chile, quinoa hasbeen characterised as genetically diverse (Fuentes et al. 2009),but the eco-physiological features of the different coastal andhighland landraces from northern and southern provenancesare only now being investigated. The present study aimed toassess how several salt tolerance features vary in a Chileanaccession of quinoa (BO78) belonging to one of the twoecotypes so far identified in Chile, namely, the coastal (sealevel) ecotype (as opposed to the highland ‘salares’ or salt flatsecotype) (Fuentes et al. 2009).
The purpose of our present investigationwas to checkwhethersome of the parameters so far associated with salt stress in othergenotypes of quinoa also respond to salinity in BO78, anaccession originating from a relatively southern part of Chilethat has been, therefore, less subject to severe drought and saltstress than the highland (Altiplano) genotypes of Peru andBolivia. Given the paucity or lack of information about somefundamental physiological responses to abiotic stress inchenopods, we also analysed these in order to verify whether,in addition to other factors (e.g. stomatal conductance, ionaccumulation) they participate in the salt stress response ofquinoa. In particular, we checked the hypotheses that prolineaccumulation does not play a fundamental role in osmoticadjustment under high salinity and that PAs contribute to ionhomeostasis by becoming depleted rather than accumulated.The ultimate purpose was to define the parameters that mostrespond to salinity in order to use them for further studies aimedat comparing salt/drought tolerance in different Chileangenotypes, an issue that has only been recently and partiallyaddressed (Delatorre-Herrera and Pinto 2009). Results may alsohelp identify those factors that confer a complex, multifacetedtrait such as salt tolerance to some species (Flowers et al. 2010).
Materials and methodsSeed germination and root elongation assaysQuinoa (Chenopodium quinoa Willd.) seeds used in this studywere provided by the National Seed Bank of Chile, managed byINIA-Intihuasi at Vicuña, and kindly donated by IngridVonBaer(AGROGEN,Vilcún,Chile). The seedswere stored at�18�Cand10% RH according to the standard protocol for long-termconservation of seed banks. The studied BO78 seed-accessionoriginates from Collipulli (37�570S, 72�250W) in the Araucaníaregion of southern Chile. This accession has a significant geneticdistance from accessions of central and northern Chile (Fuenteset al. 2009).
Seeds were briefly surface sterilised in 70% (v/v) ethanol,followed by a 10% (v/v) commercial bleach solution (6% activechlorine) for 5min, then rinsed with sterile distilled water4–5 times. They were then sown on 12� 12 cm Petri dishescontaining sterile half-strength Murashige and Skoog (MS)medium supplemented with 0.5% sucrose (Verslues et al.2006) and 0.8% (w/v) Phytagel (Sigma-Aldrich, Milan, Italy)
Functional response to salinity in quinoa Functional Plant Biology 819

and with 0, 150 or 300mM NaCl (Sigma-Aldrich). Preliminaryexperiments performed at differentNaCl concentrations (75, 150,300 and 450mM) showed that the highest concentrationcaused a total inhibition of germination, so only the twohighest sublethal doses were used. Plates, containing 10–12seeds each (five plates per treatment), were stratified at 4�C for3 days to synchronise germination, then transferred to growthchambers at 21� 2�C under a 16/8 h light/dark photoperiod anda light intensity of 50mmolm2 s–1. The number of germinatedseeds was assessed every 1 to 3 days over a period of 15 days.Seeds were considered germinated when the radicle was at least0.5 cm long. For the root elongation test, seeds were surfacesterilised, sown (12–15 seeds per dish) in 12� 12 cm Petri disheson top of a cellophanemembrane (Bio-Rad,Milan, Italy) overlaidon agarised MS medium without NaCl and stratified for 3 daysas described above. Petri dishes (five per treatment) were thenplaced vertically in the growth chamber. After 2 to 3 days, thecellophane membrane was used to transfer germinated seedlingsto new Petri dishes containing 0, 150 or 300mMNaCl. The newroot growth occurring after transfer was measured at regularintervals over a period of 7 days using a digital caliper.
Growth conditions and NaCl treatments
Seeds of BO78 were surface sterilised as described above andsown in sand-filled trayswhere theywere irrigatedwithwater andallowed to germinate; seedlings were thenmoved into sand-filledpots. Plantswere placed in agrowth chamber (IBI,Bologna, Italy)with a photosynthetic photon flux of 500mmolm–2 s–1 from coolwhite fluorescent bulbs and a 16/8 h light/dark photoperiod. Dayand night temperatures were set at 22 and 19�C respectively.Plants were irrigated with Hoagland’s nutrient solution everysecond day until the start of the salt treatments.
In all plants, excluding those grown at 0mM NaCl, salttreatments were started 30 days after sowing (DAS), whenplants had four to six leaf pairs, by incremental increases of150mM in the irrigation water every 3 days until the finalconcentrations of 150, 300, 450, 600 and 750mM NaCl werereached. The highest salinity treatmentwas reached after 15 days.The salt treatment was maintained by adding NaCl to theirrigation water (once a week to Hoagland solution) every timethe plants were irrigated. The saline solution drained freelyfrom the pots thereby preventing concentration of salts beyondtreatment levels (Lazarus et al. 2011). Twenty plants pertreatment were then kept under saline conditions for a further35-day period. Commercial sea salt (Cl–,16.4mol kg–1; Na+,12.3mol kg–1; Ca2+, 2.5mol kg–1; K+, 0.44mol kg–1; Mg2+,1.3mol kg–1; SO4
2–, 0.07mol kg–1) was used for salinitytreatments as in previous studies on salinity tolerance (Maggioet al. 2005).
For polyamine determinations, seedlings of C. quinoa BO78were transferred from sand to perlite added with Hoagland’snutrient solution and maintained in semi-hydroponic culture for2weeks, after which time theywere transferred to liquidmedium.Each seedlingwas placed in 5mL of the sameHoagland solution,containing (or not) 300mM NaCl (Sigma-Aldrich) and kept ina growth cabinet (16/8 h light/dark photoperiod, 21� 2�C,500mmolm–2 s–1) for 3 days. At sampling time, each seedling
was separately frozen in liquid nitrogen and stored at�80�Cuntiluse. The liquid medium was also collected and frozen.
Growth measurements
Plant height was measured during the course of the experimentand at the end of the experiment. Plants were then collected formeasurement of FW and DW of shoots (leaves plus stems) androots (separately).
Leaf gas exchangeLeaf transpiration (E), stomatal conductance (gs) and netphotosynthesis (A) were measured 15 days after salinisation onthree completely unfolded leaves of nine plants per treatment.Measurements of leaf gas exchange were performed on attachedleaf samples using a CIRAS-2 (PPSystem, Hitchin, UK) infraredgas analyser (closed system) with a Parkinson’s AutomaticUniversal Leaf Cuvette (PAR 1000mmolm–2 s–1, 26�C, CO2
13.63mmol L�1 and 300 cm3min–1 flow rate) equipped with18-mm diameter, 2.5-cm2 area cuvette inserts.
Stomatal size and density and salt bladder density
Micromorphological observations were conducted with abrightfield light microscope (Meiji Techno Co. Ltd, Saitama,Japan) on three 1-cm2 portions per leaf (excised from areasbetween the veins), from six leaves per plant: three young(third to fifth from the tip) and three mature (the lowest threenon-senescing leaves from the base) and two plants per treatment.Stomatal features were determined in all treatments, whereasbladder density was measured on plants grown at 0, 150, 300 and600mM NaCl. Stomatal and EBC density was evaluated fromdigital photos of the abaxial and adaxial surfaces. The stomatallengthwas determinedbyusing Image J software (Abramoff et al.2004) on 10 randomly chosen stomata per leaf portion. Thestomatal index (SI) was calculated according to the originalformula by Salisbury (1927):
SIð%Þ ¼ NS=ðEC þ NSÞ � 100; ð1Þwhere NS is the number of stomata and EC is the number ofepidermal cells.
Quantification of internal and excreted ionsAt the end of the experiment, nine plants per treatment wereharvested and divided into three portions: young leaves, matureleaves (as described above) and stems (excluding theinflorescences, if present). Measurements for each treatmentwere performed on pooled samples consisting of materialcollected from three plants. After FW determination, leaveswere submerged in 20mL of ultra-pure water and gentlyrubbed with a paintbrush, as described by Freitas and Breckle(1993) and recently applied byBenHassine et al. (2009), in orderto collect the salt fraction contained in the EBCs (henceforthdefined as ‘excreted’). After brushing, the leaf surface wasobserved under a stereomicroscope to check that all EBCs hadbeen removed and ruptured. The washing solution was stored at�20�Cuntil ion analysis, thewashed leaves and stemswere oven-dried at 65�C. After DW determination, anions and cations wereextracted from dry material of each plant portion according tothe procedure proposed by Cataldi et al. (2003). A capillary
820 Functional Plant Biology F. Orsini et al.
electrophoresis system (Beckman P/ACE 5500, PegasusScientific, Rockville, MD, USA) was used for thequantification of major anions (Cl�, SO4
2�, NO3�, NO2
�) andcations (Na+,K+,Ca2+,Mg2+) by the external standard calibrationmethod with linear regression according to the method describedby Dinelli et al. (1998). Both cations and anions were analysedin conventional 50-cm long (from injection point to detector)untreated fused silica capillaries (75mm internal diameter,Beckman) at a constant temperature of 25�C. The indirectdetection wavelength was 220 nm. The applied voltage was�20 and 20 kV for anion and cation separation respectively.The electrolyte buffer employed for the determination of theanions was 1.8mM potassium dichromate, 34mM boric acid,14mM sodium borate and 1mM diethylenetriamine. Theelectrolyte buffer for K+, Na+, Ca2+ and Mg2+ determinationwas 40mM citric acid and 23mM imidazole. Repeatability wasdetermined by analysis of leaf samples from at least five replicatemeasurements at different concentrations. Finally, the quality ofanalytical data was checked by cation–anion balance (Balestriniet al. 2000). Ion concentration of extruded and internal/tissuefractions was expressed as ion microequivalents g–1 DW.
Proline content
Tissue proline concentration was estimated following themethod by Bates et al. (1973) with slight modifications.Briefly, ~50mg of frozen seedling tissue (leaves, stems androots separately) was crushed in 1.2mL of 3% sulfosalicylicacid and the homogenate centrifuged at 16 000g at roomtemperature for 20min. A 0.5-mL aliquot of the supernatantwas made up to 1mL with water to which was added 2mLninhydrin reagent [2.5% ninhydrin in glacial acetic acid-distilledwater-85% orthophosphoric acid (6 : 3 : 1)]. The reactionmixtures were kept in a water bath at 90�C for 1 h for colourdevelopment. Test tubes were then cooled in an ice bath and2mL toluene added to separate the chromophore. Absorbance ofthe toluene phase was read by a spectrophotometer at 525 nm;proline concentration was calculated by comparing sampleabsorbancies with the standard proline curve.
Extraction and quantification of polyaminesPlant tissues (~200mg FW) were extracted with 10 volumes of4% perchloric acid (PCA); the homogenate was kept for 1 h onice then centrifuged at 15 000g for 30min. Aliquots of thesupernatants and standard solutions of putrescine (Put),spermidine (Spd) and spermine (Spm) were derivatised withdansylchloride as described by Scaramagli et al. (1999).Dansylated derivatives were extracted with toluene, taken todryness and resuspended in acetonitrile before HPLC analysis.The media from NaCl treated and untreated seedlings were takento dryness in a Rotavapor (Buchi Italia, Assago, Italy), and thedry residue resuspended in a small volume of 4% PCA. Thesuspension was then dansylated and processed as describedabove. PAs were separated and quantified by HPLC (Jasco,Tokyo, Japan) with a reverse phase C18 column (SpherisorbODS2, 5-mm particle diameter, 4.6� 250mm, Waters,Wexford, Ireland) and a programmed acetonitrile-water stepgradient, as previously described (Scaramagli et al. 1999).
Analyses (three replicates) were conducted on four biologicalreplicates.
Statistical analysis
Data were analysed by one-way ANOVA and means werecompared with Fisher’s Least Significance Difference (LSD)test where indicated.
Results
Germination, growth responses and physiological features
Only a small (7%), not statistically significant, reduction ingermination was observed at 15 DAS when seeds were sownon agar plates supplemented with 150mM NaCl, whereas asubstantial decrease (35% < at 0mM NaCl, P < 0.05) wasrecorded on plates containing 300mM NaCl (Fig. 1a). Whenseedlings germinated on medium lacking NaCl were transferredto plates with NaCl, the greatest root elongation was observed inthe presence of 150mM NaCl (1.5-fold more than 0mM NaCl,
120 0 mM 150 mM 300 mM
100
80
60
40
20
0
60
50
40
30
20
10
0
0 4 8
Days
(a)
(b)
Roo
t elo
ngat
ion
(mm
)S
eed
germ
inat
ion
(%)
Days
0 1 4 6 8
12 16
Fig. 1. (a) Seed germination and (b) root elongation in quinoa seedlingsgrown on agar plates containing 0, 150 and 300mM NaCl. Root elongationwas determined for seedlings germinated on control medium and thentransferred to media containing 0, 150 or 300mM NaCl. Values aremeans� s.e. (n= 30).
Functional response to salinity in quinoa Functional Plant Biology 821
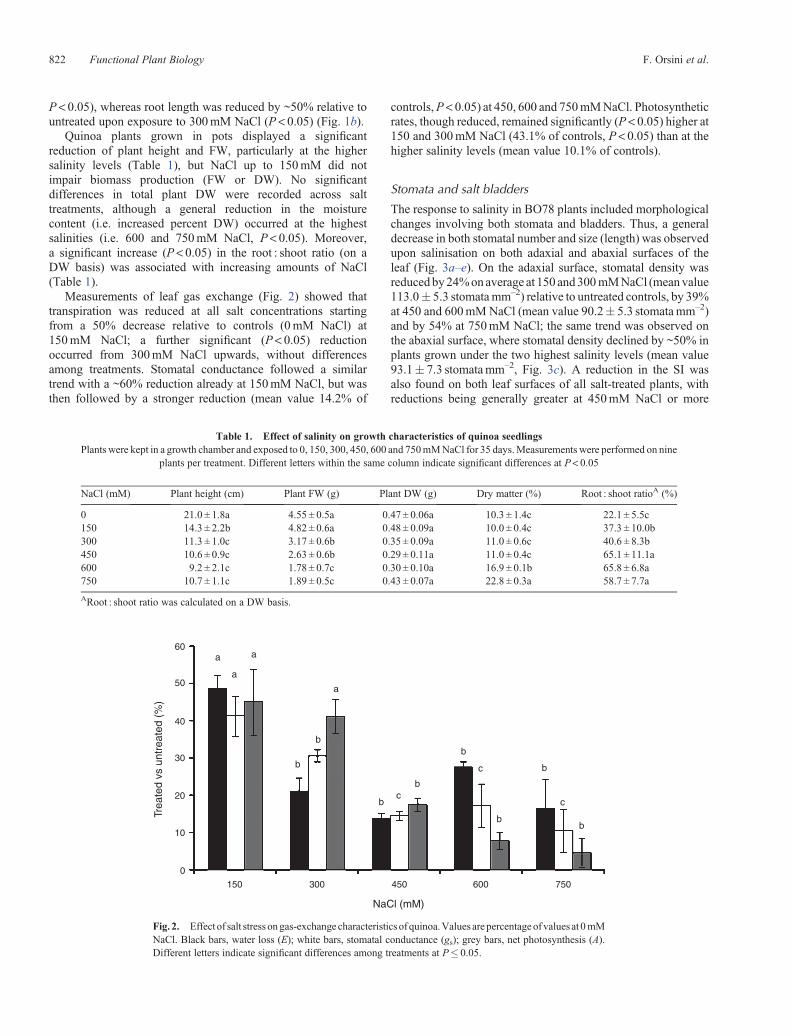
P < 0.05), whereas root length was reduced by ~50% relative tountreated upon exposure to 300mM NaCl (P < 0.05) (Fig. 1b).
Quinoa plants grown in pots displayed a significantreduction of plant height and FW, particularly at the highersalinity levels (Table 1), but NaCl up to 150mM did notimpair biomass production (FW or DW). No significantdifferences in total plant DW were recorded across salttreatments, although a general reduction in the moisturecontent (i.e. increased percent DW) occurred at the highestsalinities (i.e. 600 and 750mM NaCl, P< 0.05). Moreover,a significant increase (P < 0.05) in the root : shoot ratio (on aDW basis) was associated with increasing amounts of NaCl(Table 1).
Measurements of leaf gas exchange (Fig. 2) showed thattranspiration was reduced at all salt concentrations startingfrom a 50% decrease relative to controls (0mM NaCl) at150mM NaCl; a further significant (P < 0.05) reductionoccurred from 300mM NaCl upwards, without differencesamong treatments. Stomatal conductance followed a similartrend with a ~60% reduction already at 150mM NaCl, but wasthen followed by a stronger reduction (mean value 14.2% of
controls,P < 0.05) at 450, 600 and 750mMNaCl. Photosyntheticrates, though reduced, remained significantly (P < 0.05) higher at150 and 300mM NaCl (43.1% of controls, P < 0.05) than at thehigher salinity levels (mean value 10.1% of controls).
Stomata and salt bladders
The response to salinity in BO78 plants included morphologicalchanges involving both stomata and bladders. Thus, a generaldecrease in both stomatal number and size (length) was observedupon salinisation on both adaxial and abaxial surfaces of theleaf (Fig. 3a–e). On the adaxial surface, stomatal density wasreducedby24%onaverage at 150 and300mMNaCl (meanvalue113.0� 5.3 stomatamm–2) relative to untreated controls, by 39%at 450 and 600mMNaCl (mean value 90.2� 5.3 stomatamm–2)and by 54% at 750mM NaCl; the same trend was observed onthe abaxial surface, where stomatal density declined by ~50% inplants grown under the two highest salinity levels (mean value93.1� 7.3 stomatamm–2, Fig. 3c). A reduction in the SI wasalso found on both leaf surfaces of all salt-treated plants, withreductions being generally greater at 450mM NaCl or more
60a
a
a
b
b
b
b
c
b
b
c
b
c
a
b
50
Trea
ted
vs u
ntre
ated
(%
)
40
30
20
10
0
150 300 450
NaCl (mM)
600 750
Fig. 2. Effect of salt stress ongas-exchange characteristics of quinoa.Values arepercentageof values at 0mMNaCl. Black bars, water loss (E); white bars, stomatal conductance (gs); grey bars, net photosynthesis (A).Different letters indicate significant differences among treatments at P� 0.05.
Table 1. Effect of salinity on growth characteristics of quinoa seedlingsPlants were kept in a growth chamber and exposed to 0, 150, 300, 450, 600 and 750mMNaCl for 35 days.Measurements were performed on nine
plants per treatment. Different letters within the same column indicate significant differences at P< 0.05
NaCl (mM) Plant height (cm) Plant FW (g) Plant DW (g) Dry matter (%) Root : shoot ratioA (%)
0 21.0 ± 1.8a 4.55 ± 0.5a 0.47 ± 0.06a 10.3 ± 1.4c 22.1 ± 5.5c150 14.3 ± 2.2b 4.82 ± 0.6a 0.48 ± 0.09a 10.0 ± 0.4c 37.3 ± 10.0b300 11.3 ± 1.0c 3.17 ± 0.6b 0.35 ± 0.09a 11.0 ± 0.6c 40.6 ± 8.3b450 10.6 ± 0.9c 2.63 ± 0.6b 0.29 ± 0.11a 11.0 ± 0.4c 65.1 ± 11.1a600 9.2 ± 2.1c 1.78 ± 0.7c 0.30 ± 0.10a 16.9 ± 0.1b 65.8 ± 6.8a750 10.7 ± 1.1c 1.89 ± 0.5c 0.43 ± 0.07a 22.8 ± 0.3a 58.7 ± 7.7a
ARoot : shoot ratio was calculated on a DW basis.
822 Functional Plant Biology F. Orsini et al.
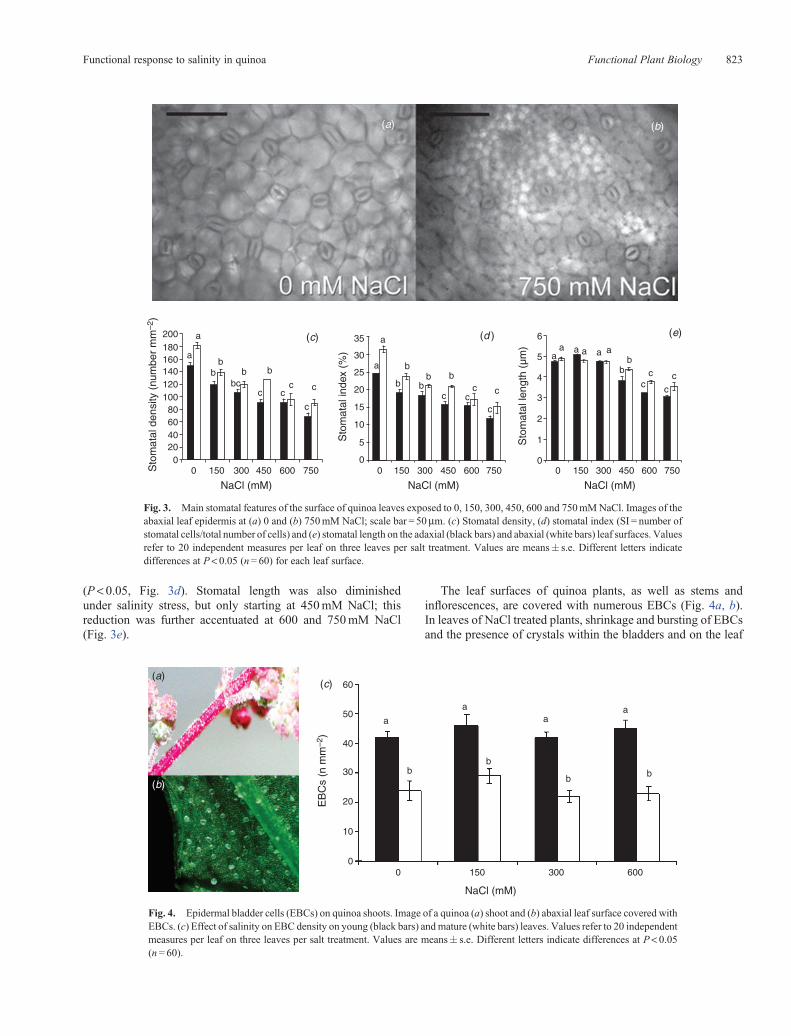
(P < 0.05, Fig. 3d). Stomatal length was also diminishedunder salinity stress, but only starting at 450mM NaCl; thisreduction was further accentuated at 600 and 750mM NaCl(Fig. 3e).
The leaf surfaces of quinoa plants, as well as stems andinflorescences, are covered with numerous EBCs (Fig. 4a, b).In leaves of NaCl treated plants, shrinkage and bursting of EBCsand the presence of crystals within the bladders and on the leaf
200 (c) (d ) (e)
a
a
bb
b b
c cc
a
a
b
b
bb b
c cc
c
c
aa a a a a
bb
cc
cc
c
c
bc
35 6
5
4
3
2
1
0
30
25
20
15
10
5
0
180160140120100806040200
0 150 300 450 600 750 0 150 300 450 600 750 0 150 300 450
NaCl (mM)NaCl (mM)NaCl (mM)
Sto
mat
al in
dex
(%)
Sto
mat
al le
ngth
(µm
)
Sto
mat
al d
ensi
ty (
num
ber
mm
–2)
600 750
(a) (b)
Fig. 3. Main stomatal features of the surface of quinoa leaves exposed to 0, 150, 300, 450, 600 and 750mMNaCl. Images of theabaxial leaf epidermis at (a) 0 and (b) 750mM NaCl; scale bar = 50mm. (c) Stomatal density, (d) stomatal index (SI = number ofstomatal cells/total number of cells) and (e) stomatal length on the adaxial (black bars) and abaxial (white bars) leaf surfaces. Valuesrefer to 20 independent measures per leaf on three leaves per salt treatment. Values are means� s.e. Different letters indicatedifferences at P< 0.05 (n= 60) for each leaf surface.
60(a)
(b)
(c)
a
aa
a
bb
b b
50
40
30
20
10
00 150
NaCl (mM)
EB
Cs
(n m
m–2
)
300 600
Fig. 4. Epidermal bladder cells (EBCs) on quinoa shoots. Image of a quinoa (a) shoot and (b) abaxial leaf surface covered withEBCs. (c) Effect of salinity on EBC density on young (black bars) andmature (white bars) leaves. Values refer to 20 independentmeasures per leaf on three leaves per salt treatment. Values are means� s.e. Different letters indicate differences at P< 0.05(n= 60).
Functional response to salinity in quinoa Functional Plant Biology 823
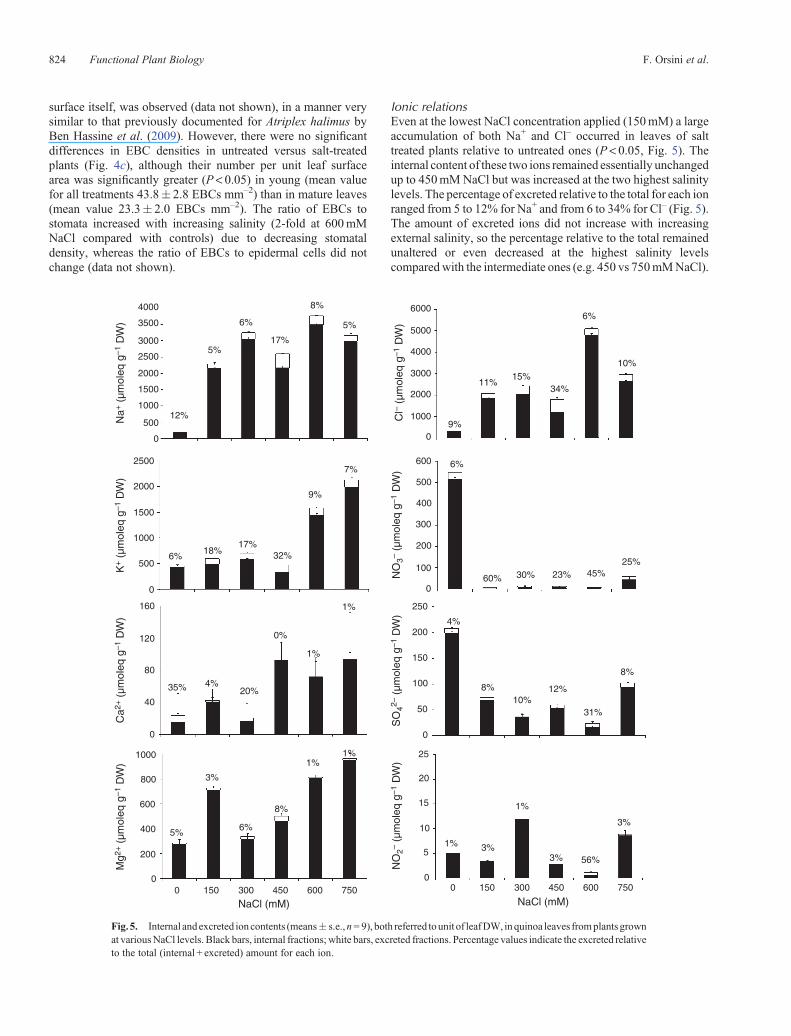
surface itself, was observed (data not shown), in a manner verysimilar to that previously documented for Atriplex halimus byBen Hassine et al. (2009). However, there were no significantdifferences in EBC densities in untreated versus salt-treatedplants (Fig. 4c), although their number per unit leaf surfacearea was significantly greater (P < 0.05) in young (mean valuefor all treatments 43.8� 2.8 EBCs mm–2) than in mature leaves(mean value 23.3� 2.0 EBCs mm–2). The ratio of EBCs tostomata increased with increasing salinity (2-fold at 600mMNaCl compared with controls) due to decreasing stomataldensity, whereas the ratio of EBCs to epidermal cells did notchange (data not shown).
Ionic relationsEven at the lowest NaCl concentration applied (150mM) a largeaccumulation of both Na+ and Cl– occurred in leaves of salttreated plants relative to untreated ones (P < 0.05, Fig. 5). Theinternal content of these two ions remained essentially unchangedup to 450mMNaCl but was increased at the two highest salinitylevels. The percentage of excreted relative to the total for each ionranged from 5 to 12% for Na+ and from 6 to 34% for Cl– (Fig. 5).The amount of excreted ions did not increase with increasingexternal salinity, so the percentage relative to the total remainedunaltered or even decreased at the highest salinity levelscomparedwith the intermediate ones (e.g. 450 vs 750mMNaCl).
4000 6000
5000
4000
3000
2000
1000
0
600
500
400
300
200
250
200
150
100
50
0
25
20
15
10
5
00 150 300
NaCl (mM)NaCl (mM)
450 600 75000
150 300 450 600 750
100
0
3500
3000
Na+
(µm
oleq
g–1
DW
)
Cl–
(µm
oleq
g–1
DW
)N
O3–
(µm
oleq
g–1
DW
)N
O2–
(µm
oleq
g–1
DW
)S
O42–
(µm
oleq
g–1
DW
)
K+ (
µmol
eq g
–1 D
W)
Ca2
+ (
µmol
eq g
–1 D
W)
Mg2
+ (
µmol
eq g
–1 D
W)
2500
2000
1500
12%
5%
6%
17%
8%
5%
9%
11%15%
34%
6%
10%
25%45%23%30%60%
6%
4%
8%10%
12%
31%
8%
3%
56%3%
1%
3%1%5%
3%
35% 4%20%
0%
1%
1%
6%18%
17%32%
9%
7%
6%
8%
1%1%
1000
500
0
2500
2000
1500
1000
500
0
160
120
80
40
0
1000
800
600
400
200
Fig.5. Internal andexcreted ioncontents (means� s.e.,n= 9), both referred tounit of leafDW, inquinoa leaves fromplantsgrownat variousNaCl levels. Black bars, internal fractions; white bars, excreted fractions. Percentage values indicate the excreted relativeto the total (internal + excreted) amount for each ion.
824 Functional Plant Biology F. Orsini et al.

Significant increases (P< 0.05) in internal leaf K+ contentoccurred only when the highest NaCl treatments were supplied;indeed, the internal amount of this cation increased by ~3- and4-fold at 600 and 750mMNaCl, respectively, relative to controls(Fig. 5). In NaCl treated plants, the proportion of excreted K+
relative to the total also increased significantly (P < 0.05)compared with untreated plants, especially at moderate salinity(from150 to 450mMNaCl)where it reached 20–30%,whereas athigher NaCl concentrations the percentage decreased to <10%(Fig. 5).
NaCl application also influenced the accumulation of theother measured ions, although bivalent cations and anionsfollowed opposite trends. Thus, an increase was observed forCa2+, especially at NaCl doses of 450mM or above (P < 0.05,Fig. 5). Similarly, a significant (P< 0.05) increase in Mg2+ leaftissue contents was found in treatments with the highest salinitylevels (Fig. 5).A reduction (P< 0.05) in anions (namelyNO3
– andSO4
2–) was observed under saline treatments, with NO3–
displaying the strongest (on average 9-fold) decrease at allsalinity levels, starting from 150mM NaCl (Fig. 5). NO2
–, thenegligible amount of which is usually adopted as a control ofthe reliability of the analysis, was not significantly affected bysalinity. Further, at the lowest internal content of this anion(observed at 600mM NaCl) an extremely high amount (56%over the total) of it was extruded, suggesting that a minimumthreshold of excretion would be maintained anyway.
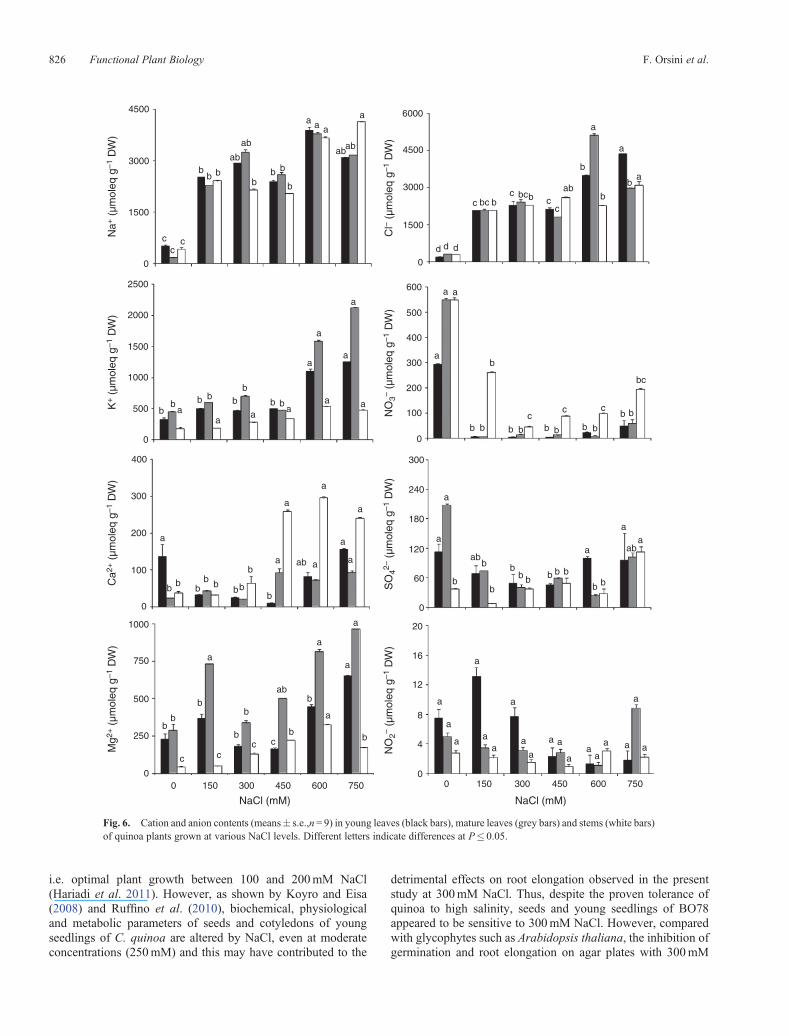
Ion contents were also analysed separately in the differentparts of the shoot (young leaves, mature leaves and stems). Na+
accumulation was similar in all of the three parts; after a dramaticincrease at 150mM NaCl (P < 0.05), it remained constant upto 450mM NaCl and then increased slightly (Fig. 6). Cl– alsoincreased to the same extent in all plant parts upon exposureto moderate salinity (150mM, P < 0.05), remained constant upto 450mM, then increased further, especially in leaves, at thehighest concentrations of NaCl (Fig. 6). K+ increased (P < 0.05)more inmature leaves than inyoungoneswhenNaClwas applied,especially at the highest salinity level, whereas changes werenegligible in stems (Fig. 6). In the stems of salt treated plants, K+
contents did not increase markedly, despite the strongaccumulation of Na+, so a strongly diminished K+ : Na+ ratio(0.35 in untreated vs 0.11 with 750mM NaCl) was observed(data not shown), whereas in young leaves the ratio fell by lessthan 50% (0.66 in untreated vs 0.37 with 750mM NaCl). UponNaCl treatment, the greatest accumulation of Ca2+ occurredin stems, whereas Mg2+ was mainly found in mature leaves.The salt induced reduction of NO3
– was greater (P< 0.05) inboth young and mature leaves than in stems, whereas SO4
– wasstrongly reduced in mature leaves and increased in stems(P < 0.05).
As shown in Fig. 7, both excreted (Fig. 7a) and internallyaccumulated (Fig. 7b) Cl– and Na+ were, starting from 150mMNaCl application, the most abundant ions (relative to all thosedetermined), followed byK+. Cl– dominated in the excreted ionicfraction, whereas Na+ was the most abundant ion accumulatedinside the plant. NO3
– was the next most abundant, even thoughboth its internal and excreted fractions declined dramatically inthe presence of NaCl (Fig. 7a, b). Other ions were found in muchsmaller amounts, so their relative values were, in quantitativeterms, negligible (data not shown).
Changes in proline and polyamine levels
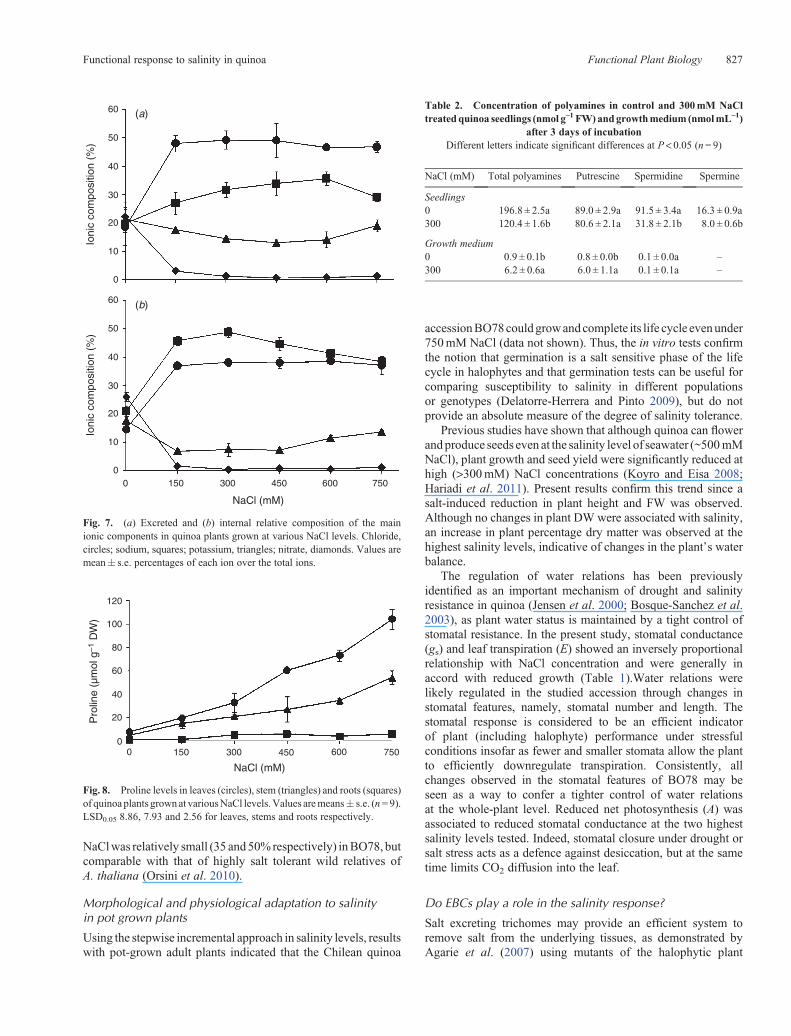
Proline concentration in the different plant organs increasedsignificantly (P < 0.05) with salinisation, especially in leavesand stems, starting from 450mM NaCl (Fig. 8). At the highestconcentrations of NaCl (600 and 750mM), the increase wasgreatest in the leaves (~10-fold relative to 0mM NaCl).Proline was detectable also in roots, albeit at very lowconcentrations, but showed a significant increase (P< 0.05)only at 750mM NaCl.
Quinoa seedlings had very similar amounts of Put andSpd, and Spm was the least abundant of the three PAs(Table 2). After being exposed to 300mM NaCl for 3 days,all three PAs declined (P < 0.05) compared with untreatedplants; Spd and Spm showed the greatest reduction (~65 and50% respectively). Indeed, a ~40% decrease in total PAs(Put + Spd + Spm) was observed in NaCl treated seedlings.In the medium of seedlings exposed to NaCl, there was a large(~6.8-fold) increase in the amount of excreted Put comparedwith the NaCl free medium, but no appreciable differences in theconcentration of Spd (Spm was below the detection limit)(Table 2).
Discussion
Evaluating the early developmental response to highsalinity in BO78
Delayed or diminished seed germination in response to NaCl hasbeen reported for several halophytes, including quinoa (Koyroand Eisa 2008; Delatorre-Herrera and Pinto 2009; Ruffino et al.2010) and has been viewed as a protective strategy to ensuremaximum survival (Orsini et al. 2010). Indeed, salt toleranceis often a developmental stage specific phenomenon (Flowersand Colmer 2008). Since rapid screening at the germination orseedling stage is less laborious, less time consuming and cheaperbut still reliable (Dasgan et al. 2002), it may be considered usefulfor a preliminary evaluation of salt tolerance.
Results from the present study showed that low–moderatesalinity (150mM NaCl) caused an initial delay (up to day 2)in germination of BO78. However, from then on this wasthe same as in the absence of NaCl, thus, confirming thehalophytic behaviour of quinoa. In contrast, when 300mMNaCl was supplied, germination was further delayed and inthe long term, moderately inhibited relative to controls. Intheir evaluation of seed germination under strongly salineconditions (400mM NaCl) in four quinoa selections fromnorthern (Altiplano) and southern (non-saline soils, higherprecipitation) Chile, Delatorre-Herrera and Pinto (2009)registered a highly variable provenance dependent responseranging from 53 to 90% inhibition.
Root elongation tests are generally regarded as being highlyinformative in the evaluation of salt tolerance (Verslues et al.2006). The measure of root elongation in BO78 upon transfer ofseedlings to salt-containing plates, therefore, separated from theeffect on germination, showed that at 150mMNaCl, root growthwas enhanced. Halophytic dicots are likely to increase theirgrowth under mildly saline conditions and this is particularlytrue for their belowground organs (Orsini et al. 2010). Althoughpublished data for in vitro root elongation tests with quinoa arenot available, results from adult plants confirm this behaviour,
Functional response to salinity in quinoa Functional Plant Biology 825
i.e. optimal plant growth between 100 and 200mM NaCl(Hariadi et al. 2011). However, as shown by Koyro and Eisa(2008) and Ruffino et al. (2010), biochemical, physiologicaland metabolic parameters of seeds and cotyledons of youngseedlings of C. quinoa are altered by NaCl, even at moderateconcentrations (250mM) and this may have contributed to the
detrimental effects on root elongation observed in the presentstudy at 300mM NaCl. Thus, despite the proven tolerance ofquinoa to high salinity, seeds and young seedlings of BO78appeared to be sensitive to 300mM NaCl. However, comparedwith glycophytes such as Arabidopsis thaliana, the inhibition ofgermination and root elongation on agar plates with 300mM
4500 6000
d
a
a a
b
a
a
a
aa
a
bb
a
a
a
a
a
bb
aa
aa
a
a
a
a
a
cc
c
bb
ab
ab
a a a
a
abab
bb
b b
b
ab b bb
b b
a
a
aaab
b bb
b bbb
b
b
a
a
a
a
abb
b
bb
bb
c cc c
a
a
a
aa
a
a
a
a a
aa
a
a
a
aa
b
b
b
bb b b b b
b b
abab
b
b
b b b b b b
b b
bc
cc c
d d
c bcbc
bbc
cc
ab
b
b
b
a
a
a
4500
3000
1500
0
600
500
400
300
200
300
240
180
120
60
20
16
12
8
4
00 150 300 450 600 7500 150 300 450
NaCl (mM) NaCl (mM)
600 750
0
100
0
3000
Na+
(µm
oleq
g–1
DW
)K
+ (
µmol
eq g
–1 D
W)
Ca2
+ (
µmol
eq g
–1 D
W)
Mg2
+ (
µmol
eq g
–1 D
W)
NO
2– (µ
mol
eq g
–1 D
W)
SO
42– (
µmol
eq g
–1 D
W)
NO
3– (µ
mol
eq g
–1 D
W)
Cl–
(µm
oleq
g–1
DW
)
1500
0
2500
2000
1500
1000
500
0
400
300
200
100
0
1000
750
500
250
0
Fig. 6. Cation and anion contents (means� s.e.,n= 9) in young leaves (black bars), mature leaves (grey bars) and stems (white bars)of quinoa plants grown at various NaCl levels. Different letters indicate differences at P� 0.05.
826 Functional Plant Biology F. Orsini et al.
NaClwas relatively small (35 and50%respectively) inBO78, butcomparable with that of highly salt tolerant wild relatives ofA. thaliana (Orsini et al. 2010).
Morphological and physiological adaptation to salinityin pot grown plants
Using the stepwise incremental approach in salinity levels, resultswith pot-grown adult plants indicated that the Chilean quinoa
accessionBO78couldgrowandcomplete its life cycle evenunder750mM NaCl (data not shown). Thus, the in vitro tests confirmthe notion that germination is a salt sensitive phase of the lifecycle in halophytes and that germination tests can be useful forcomparing susceptibility to salinity in different populationsor genotypes (Delatorre-Herrera and Pinto 2009), but do notprovide an absolute measure of the degree of salinity tolerance.
Previous studies have shown that although quinoa can flowerandproduce seeds even at the salinity level of seawater (~500mMNaCl), plant growth and seed yield were significantly reduced athigh (>300mM) NaCl concentrations (Koyro and Eisa 2008;Hariadi et al. 2011). Present results confirm this trend since asalt-induced reduction in plant height and FW was observed.Although no changes in plant DW were associated with salinity,an increase in plant percentage dry matter was observed at thehighest salinity levels, indicative of changes in the plant’s waterbalance.
The regulation of water relations has been previouslyidentified as an important mechanism of drought and salinityresistance in quinoa (Jensen et al. 2000; Bosque-Sanchez et al.2003), as plant water status is maintained by a tight control ofstomatal resistance. In the present study, stomatal conductance(gs) and leaf transpiration (E) showed an inversely proportionalrelationship with NaCl concentration and were generally inaccord with reduced growth (Table 1).Water relations werelikely regulated in the studied accession through changes instomatal features, namely, stomatal number and length. Thestomatal response is considered to be an efficient indicatorof plant (including halophyte) performance under stressfulconditions insofar as fewer and smaller stomata allow the plantto efficiently downregulate transpiration. Consistently, allchanges observed in the stomatal features of BO78 may beseen as a way to confer a tighter control of water relationsat the whole-plant level. Reduced net photosynthesis (A) wasassociated to reduced stomatal conductance at the two highestsalinity levels tested. Indeed, stomatal closure under drought orsalt stress acts as a defence against desiccation, but at the sametime limits CO2 diffusion into the leaf.
Do EBCs play a role in the salinity response?
Salt excreting trichomes may provide an efficient system toremove salt from the underlying tissues, as demonstrated byAgarie et al. (2007) using mutants of the halophytic plant
60 (a)
(b)
50
40
Ioni
c co
mpo
sitio
n (%
)Io
nic
com
posi
tion
(%)
NaCl (mM)
30
20
10
0
60
50
40
30
20
10
00 150 300 450 600 750
Fig. 7. (a) Excreted and (b) internal relative composition of the mainionic components in quinoa plants grown at various NaCl levels. Chloride,circles; sodium, squares; potassium, triangles; nitrate, diamonds. Values aremean� s.e. percentages of each ion over the total ions.
120
100
80
60
40
20
00 150 300
NaCl (mM)
Pro
line
(µm
ol g
–1 D
W)
450 600 750
Fig. 8. Proline levels in leaves (circles), stem (triangles) and roots (squares)of quinoa plants grownat variousNaCl levels.Values aremeans� s.e. (n= 9).LSD0.05 8.86, 7.93 and 2.56 for leaves, stems and roots respectively.
Table 2. Concentration of polyamines in control and 300mM NaCltreatedquinoa seedlings (nmol g–1 FW) andgrowthmedium (nmolmL–1)
after 3 days of incubationDifferent letters indicate significant differences at P< 0.05 (n= 9)
NaCl (mM) Total polyamines Putrescine Spermidine Spermine
Seedlings0 196.8 ± 2.5a 89.0 ± 2.9a 91.5 ± 3.4a 16.3 ± 0.9a300 120.4 ± 1.6b 80.6 ± 2.1a 31.8 ± 2.1b 8.0 ± 0.6b
Growth medium0 0.9 ± 0.1b 0.8 ± 0.0b 0.1 ± 0.0a –
300 6.2 ± 0.6a 6.0 ± 1.1a 0.1 ± 0.1a –
Functional response to salinity in quinoa Functional Plant Biology 827

Mesembryanthemum crystallinum lacking EBCs. In the BO78accession of quinoa, EBCswere abundant in both control and salttreated plants. Differences in their density could be associatedto leaf age, as previously described by Adams et al. (1998) inM. crystallinum, but not to salt treatment. The fact that EBCdensity on the leaf surface did not increase in salt-treated plantscompared with controls may also explain why the amount ofexcreted ions did not increase even though the internalconcentrations did. This suggests that the excretion mechanismbased on salt glands had reached saturation at 150mM NaCl.Data regarding the number of salt bladders in relation to externalsalinity, or to the amount of internally accumulated or excretedions have not yet been published. Present data indicating that theirnumber did not increase with increasing salinity suggests thatfurther investigation is needed to test the hypothesis that EBCsplay a crucial role in conferring salt tolerance in chenopods.
Internal ion accumulation is a key feature in quinoasalt tolerance
In halophytes, salt tolerance is mainly based on inclusion ratherthan exclusion of salts. Thus, although some tolerant glycophytesrestrict ion movement to the shoots by limiting ion influx into theroot xylem, halophytes readily translocate and accumulate ionsin the aerial parts (Flowers and Colmer 2008). The accumulatedions (mainly Na+, Cl– and K+) are supposedly used for osmoticadjustment in aerial plant parts, thus, facilitatingwater uptake andtransport and presumably lowering the metabolic cost requiredfor the production of organic osmolytes such as proline, asconfirmed by present data (see below).
In a recent paper, Hariadi et al. (2011) quantified theimportance of inorganic ions for osmotic adjustment in quinoaplants exposed to a 0–500mM NaCl concentration range;indicating that most (up to 95% in mature leaves) of it reliedon the accumulation ofNa+, K+ andCl–, whereas the contributionof organic osmolytes was limited. Present results confirmthis notion, as shown by the fact that under most or all salttreatments Na+, Cl– and K+ accumulated several-fold comparedwith controls, becoming the three most represented ions inboth leaves and stems. In general, no differences wereobserved between young and mature leaves, although a greateraccumulation of K+ was observed in mature leaves at the highestsalinities (600–750mMNaCl), a feature also reported byHariadiet al. (2011).
NaCl-induced K+ efflux or impaired K+ uptake can be highlydetrimental (Demidchik et al. 2010). Thus, the regulation of K+
homeostasis is an important aspect of salt tolerance and the abilityto retain an optimal K+ : Na+ ratio is crucial for tolerance oradaptation to salt stress (Munns and Tester 2008). Present resultsconfirm the notion that the ability of quinoa to cope with saltstress includes mechanisms aimed at avoiding, or limiting, K+
deficiency, as indicated by the fact that plants exposed to the twohighest salinity levels exhibited a 3-fold higher concentration ofK+ than controls or plants exposed to lower NaCl concentrations.However, as previously reported by Hariadi et al. (2011), quinoaplants were unable to maintain a K+ : Na+ ratio equal to that ofcontrols (0mM NaCl) already at moderate NaCl concentrationsdue to strong accumulation of Na+. Under our conditions, theK+ : Na+ ratio declined (by ~4-fold and 10-fold in young and
mature leaves respectively) starting from 150mM NaCl.However, with increasing salinity levels the ratio did notdecline further (young leaves) and even increased (matureleaves) due to high K+ levels, confirming the hypothesis thatsalt tolerance is linked to limitation of K+ depletion even underhighly unfavourable conditions.
Given that the ion extraction method adopted in thepresent work did not extract structural K+, most of the free K+
measured presumably originated from the vacuole. Therefore, theaccumulation of this cation under salinity favours the hypothesisthat it has an important role in leaf osmotic adjustment understrongly saline conditions (Flowers andColmer 2008).Moreover,under our experimental conditions, at the low–medium NaClconcentration range, total K+ increased only slightly comparedwith controls, whereas the percentage of excreted K+ increasedseveral fold. Under the two highest salinity treatments, however,the fraction that was excreted declined to 7–9%, despite a strongincrease in tissue K+ levels. This suggests that under the highestsalt stress conditions, tissue K+ was accumulated rather thanexcreted, in order to contribute more efficiently to the osmoticadjustment of leaf cells.
The comprehension of ionic relations should not be limited tothese three elements, but indeed, needs to consider also the leastrepresented ones. For instance, experimental evidence suggeststhat Ca2+ has a function in salt tolerance. Externally suppliedCa2+ reduces the toxic effects of NaCl (Shabala et al. 2006),presumably by improving K+ : Na+ selectivity. Either low K+ orhigh Na+ tissue concentrations can trigger transient Ca2+
increases that activate stress signalling networks, leading tosalt adaptation (Luan et al. 2009). In BO78 plants exposed tothe highest NaCl concentrations, strongly increased Ca2+ contentwas observed and this was especially true in the stems, where,in contrast, no decreases in K+ were associated with salinity.Given its purported beneficial function under salt stress, thisaccumulation of Ca2+ may be viewed as a positive response.Similarly, the ability of BO78 plants to preserve ion homeostasiswas also reflected in the fact that Mg2+ changed only slightly oreven increased under all salinity levels ranging from 150 to750mM NaCl. Magnesium is deeply woven into cellularmetabolism and has many important roles in plants as it isa constituent of chlorophyll and ATP, as well as a cofactor formany enzymes.
Upon exposure to NaCl, anions are usually less representeddue to their reduced uptake as a consequence of the increased Cl–
concentration (Lohaus et al. 2000). For example, increasedconcentrations of Cl– compete with NO3
– uptake andaccumulation (Liu and Shelp 1996). In BO78, leaf and stemNO3
– and SO4– contents were reduced upon salinity, although no
relevant changes were observable among salinised treatments,suggesting that the plants may be subjected to some nutrientdeficiency.However, despite the reduced tissueNO3
– content andthe high percentage of this cation accumulated in EBCs, a featurepreviously documented in Atriplex littoralis (Freitas and Breckle1993), the amino acid proline was able to accumulate under saltstress.
In accord with Hariadi et al. (2011), results with BO78 showthat increasing the supplied NaCl concentration by five times(from150 to750mM) resulted in a~50%increase in leafNa+.Thefact that tissue Na+ contents did not increase significantly in
828 Functional Plant Biology F. Orsini et al.

response to increasing salinity suggests either a very strict controlof xylem loading or an efficient removal of this ion throughexcretion. Under our experimental conditions, ion eliminationby sequestration into ECBs appears to play a minor role underhigh salinity. Indeed, results showed that, with few exceptions,the excreted fraction of all ions were within the same low rangeof values (5–34%), irrespective of applied salinity levels. Inparticular, the excreted Na+ fraction did not exceeded 8% ofthe total. InM. crystallinum, when the leaves accumulated Na+ toconcentrations in excess of 600mM, EBCs accumulated less ofthis cation than the underlying mesophyll (Agarie et al. 2007).Owing to limited Na+ excretion, the K+ : Na+ ratio in BO78remained generally higher in the EBCs than inside the leaf,with decreases relative to controls of up to 17-fold inside theleaves andup to4-fold in the excreted fraction.This, togetherwiththeir unaltered density, indicates that ECBs contribute to limitingexcess salt accumulation, but may not represent amajor tolerancemechanism at very high salinities and that from 150mM NaClupwards, limitation of Na+ uptake may play a preponderant roleinstead.
Further, the two bivalent cations (Ca2+ and Mg2+) displayedthe lowest excretion rate, possibly in accord with their importantfunctions in plant cell physiology (energy metabolism andsignalling).
Finally, the mean (total anions) : (total cations) ratio that wasexcreted in both control and salt treated plants was ~1.0, whereasfor internal ion accumulation the ratio for untreated controls was0.90� 0.01 but upon salinisation the ratio fell to 0.65� 0.01(data not shown). This decline suggests that stress induced arelevant accumulation of other anionic species (i.e. organic acids,zwitterionic amino acids, proteins) not detected by the adoptedanalytical approach.
Proline accumulation may not be a key elementfor salt tolerance in quinoa
Upon salt stress, proline concentration increased in all plantorgans of the BO78 accession of quinoa, although to thegreatest extent in leaves, reaching at the most a concentrationof ~0.1mmol g–1 DW. Indeed, the reported levels for bothproline and glycinebetaine in quinoa plants subjected to saltstress have been regarded as being too low to contributesignificantly to cell osmotic adjustment (Ruffino et al. 2010).Extremely low concentrations (0.4–0.9mg g–1 FWcorrespondingto ~0.03–0.08mmol g–1 DW) of proline where also reported byAguilar et al. (2003) in Peruvian accessions of quinoa harvestedacross different environments. Thus, as also observed in thepresent study, the concentrations of proline accumulatedappear not to be sufficient to provide osmotic adjustment inquinoa. Chen et al. (2007) observed that in salt tolerant barleycultivars K+ was the main contributor to cytoplasmic osmolality,whereas in salt sensitive genotypes, glycinebetaine and prolinecompensated for reduced cytosolic K+ levels.
Finally, it has been proposed that osmolytes may contributeindirectly to osmotic adjustment by regulatingK+ transport acrossthe plasma membrane, thus, limiting NaCl induced efflux ofthis cation (Cuin and Shabala 2005, 2007). Present and earlier(Ruffino et al. 2010; Hariadi et al. 2011) observations of veryhigh K+ levels in quinoa under salt stress, together with limited
proline accumulation, are consistent with this notion. In the caseof quinoa, inorganic ions appear to be responsible for osmoticadjustment, so a role for proline as osmoprotectant in mitigatingNa+ toxicity warrants further investigation.
What role do polyamines play in salt stress response?
Despite their purported protective function against varioustypes of stress, there are cases, as presently observed inquinoa, in which PAs are depleted, rather than accumulated instressed tissues. Thus, although increases in PA titers induced bysalt stress have been reported, there is growing evidence thatthe response is more complex. In Brassica (Das et al. 1995) andtomato (Santa-Cruz et al. 1998), short term exposure to salinityinduced PA accumulation, whereas long term exposure led eitherto no changes or to a reduction of one or more PA. Further, Putaccumulation was higher in sensitive than in tolerant lines oftomato (Santa-Cruz et al. 1998).
PAs are cations at physiological pH, which implies that theymayalsoplayapart in ionhomeostasis. Since, inBO78plants, saltstress led to an increase in total Na+ and K+, free PAs may havedecreased in order to maintain ion balance. Indeed, an inverserelationship between PA and Na+ tissue concentrations in tomatoplants has been reported (Santa-Cruz et al. 1998). Moreover, thefact that apple shoots treated with 100mMNaCl contained moreK+ and less Put than those treated with 200mM NaCl providedfurther evidence for the role of PAs in maintaining ion balanceunder long term salt treatment (Liu et al. 2008). A similarconclusion was drawn by Janicka-Russak et al. (2010), whofound a decrease in PA levels in cucumber roots treated for24 h with 200mM NaCl.
The possibility that cellular PAs decline as a result ofexcretion, rather than altered biosynthesis or catabolism, hasreceived scant attention. Indeed, the question has beenaddressed only once in barley roots pre-incubated withradiolabelled Put (Tamai et al. 2000). Present results confirmthe notion that efflux does indeed contribute to lowering PA,especiallyPut, levels inquinoaplants exposed to salinity. Specificinteractions between PAs and the ion excretion process at theleaf salt bladder level have been recently proposed for Atriplex,where application of exogenous ABA improved salt stressresistance in a drought resistant population by enhancing Na+
excretion while inducing an increase in total PA content (BenHassine et al. 2009).
In conclusion, the BO78 accession of quinoa functionallyovercame salt stress even though this germplasm comes froma region where salt stress is not as strong as in the more northernlatitudes of Chile and in the Altiplano. The plant responseinvolved several adaptive strategies at the morphological(reduction of stomatal size and density), physiological(downregulation of water loss and stomatal conductance) andbiochemical (ion homeostasis and depletion of PAs) levels.Thus, these can be regarded as useful indicators of adaptationto salinity in this quinoa accession. Some of them confirmprevious knowledge regarding quinoa or halophytic dicots ingeneral, but others, such as EBC density and ion sequestrationcapacity and PAs, require further investigation to assess their rolein this and other chenopods. Experiments are underway tocompare Chilean accessions adapted to different environments
Functional response to salinity in quinoa Functional Plant Biology 829

in order to provide useful indications for emerging breedingprogrammes for this exceptionally stress resistant andnutritionally valuable pseudo cereal, representing a primaryfood security issue for the Andean region. Moreover, as withsome other halophytes (Flowers et al. 2010), quinoa has greatpotential as a worldwide multi-purpose crop (e.g. forage,revegetation of salt affected land and remediation of pollutedsoil).
Acknowledgements
Funding from the TWAS-ICGEBexchange programme ‘Tolerance strategiesof quinoa plants under salt stress’ (CRP.PB/CHI06–01) and from the EUIRSES programme (PIRSES-GA-2008–230862) is gratefully acknowledged.The research was also financed by University of Bologna funds (RFO).
References
AbramoffMD,Magelhaes PJ, RamSJ (2004) Image processingwith ImageJ.Biophotonics International 11, 36–42.
Adams P, Nelson DE, Yamada S, Chmara W, Jensen RG, Bohnert HJ,Griffiths H (1998) Growth and development of Mesembryanthemumcrystallinum (Aizoaceae). New Phytologist 138, 171–190. doi:10.1046/j.1469-8137.1998.00111.x
Agarie S, Shimoda T, Shimizu Y, Baumann K, Sunagawa H, Kondo A,Ueno O, Nakahara T, Nose A, Cushman JC (2007) Salt tolerance, saltaccumulation and ionic homeostasis in an epidermal bladder-cell-lessmutant of the common ice plant Mesembryanthemum crystallinum.Journal of Experimental Biology 58, 1957–1967.
Aguilar PC, Cutipa Z,Machaca E, LopezM, Jacobsen SE (2003)Variation ofproline content of quinoa (Chenopodium quinoa Willd.) in high beds(Waru Waru). Food Reviews International 19, 121–127. doi:10.1081/FRI-120018878
Alcázar R, Altabella T, Marco F, Bortolotti C, Reymond M, Koncz C,Carrasco P, Tiburcio AF (2010) Polyamines: molecules withregulatory functions in plant abiotic stress tolerance. Planta 231,1237–1249. doi:10.1007/s00425-010-1130-0
Balestrini R, Galli L, Tartari G (2000) Wet and dry atmospheric depositionat prealpine and alpine sites in northern Italy. Atmospheric Environment34, 1455–1470. doi:10.1016/S1352-2310(99)00404-5
Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free prolinein water stress studies. Plant and Soil 39, 205–207. doi:10.1007/BF00018060
Ben Hassine A, Ghanem ME, Bouzid S, Lutts S (2009) Abscisic acid hascontrasting effects on salt excretion and polyamine concentrations of aninland and a coastal population of the Mediterranean xero-halophytespecies Atriplex halimus. Annals of Botany 104, 925–936. doi:10.1093/aob/mcp174
Bosque-Sanchez H, Lemeur R, Van Damme P, Jacobsen SE (2003)Ecophysical analysis of drought and salinity stress of quinoa(Chenopodium quinoa Willd.). In ‘Quinoa – an Andean food group’.(Eds SE Jacobsen, A Mujica) pp. 111–119. (Marcel Dekker Inc.: NewYork)
Cataldi TRI, Margiotta G, Del Fiore A, Bufo SA (2003) Ionic content in plantextracts determined by ion chromatography with conductivity detection.Phytochemical Analysis 14, 176–183. doi:10.1002/pca.700
Chen Z, Cuin TA, Zhou M, Twomey A, Naidu BP, Shabala S (2007)Compatible solute accumulation and stress-mitigating effects in barleygenotypes contrasting in their salt tolerance. Journal of ExperimentalBotany 58, 4245–4255. doi:10.1093/jxb/erm284
Cuin TA, Shabala S (2005) Exogenously supplied compatible solutes rapidlyameliorate NaCl induced potassium efflux from barley root. Plant & CellPhysiology 46, 1924–1933. doi:10.1093/pcp/pci205
Cuin TA, Shabala S (2007) Potassium efflux channels mediate Arabidopsisroot responses to reactive oxygen species and the mitigating effect ofcompatible solutes.Plant,Cell&Environment30, 875–885. doi:10.1111/j.1365-3040.2007.01674.x
Das S, Bose A, GhoshB (1995) Effect of salt stress on polyaminemetabolismin Brassica campestris. Phytochemistry 39, 283–285. doi:10.1016/0031-9422(94)00920-O
Dasgan HY, Aktas H, Abak K, Cakmak I (2002) Determination of screeningtechniques to salinity tolerance in tomatoes and investigation of genotyperesponses. Plant Science 163, 695–703. doi:10.1016/S0168-9452(02)00091-2
Delatorre-Herrera J, Pinto M (2009) Importance of ionic and osmoticcomponents of salt stress on the germination of four Quinoa(Chenopodium quinoa Willd.) selections. Chilean Journal ofAgricultural Research 69, 477–485. doi:10.4067/S0718-58392009000400001
Demidchik V, Cuin TA, Svistunenko D, Smith SJ, Miller AJ, Shabala S,Sokolik A, Yurin V (2010) Arabidopsis root K+-efflux conductanceactivated by hydroxyl radicals: single-channel properties, genetic basisand involvement in stress-induced cell death. Journal of Cell Science123, 1468–1479. doi:10.1242/jcs.064352
Dinelli G, Di Martino E, Vicari A (1998) Multi-determination of mineralions in environmental water samples by capillary electrophoresis.Recent Research Developments in Agricultural and Food Chemistry 2,435–442.
Flowers TJ, Colmer TD (2008) Salinity tolerance in halophytes. NewPhytologist 179, 945–963. doi:10.1111/j.1469-8137.2008.02531.x
Flowers TJ, Galal HK, Bromham L (2010) Evolution of halophytes: multipleorigins of salt tolerance in land plants. Functional Plant Biology 37,604–612. doi:10.1071/FP09269
Freitas H, Breckle S-W (1993) Accumulation of nitrate in bladder hairs ofAtriplex species. Plant Physiology and Biochemistry 31, 887–892.
Fuentes FF, Martinez EA, Hinrichsen PV, Jellen EN, Maughan PJ (2009)Assessment of genetic diversity patterns inChileanquinoa (Chenopodiumquinoa Willd.) germplasm using multiplex fluorescent microsatellitemarkers. Conservation Genetics 10, 369–377. doi:10.1007/s10592-008-9604-3
Hariadi Y, Marandon K, Tian Y, Jacobsen SE, Shabala S (2011) Ionic andosmotic relations in quinoa (Chenopodium quinoa Willd.) plants grownat various salinity levels. Journal of Experimental Botany 62, 185–193.doi:10.1093/jxb/erq257
Jacobsen SE, Mujica A, Jensen CR (2003) The resistance of quinoa(Chenopodium quinoa Willd.) to adverse abiotic factors. Food ReviewsInternational 19, 99–109. doi:10.1081/FRI-120018872
Janicka-Russak M, Kabala K, Miodzinska E, Klobus G (2010) The role ofpolyamines in the regulation of the plasma membrane and the tonoplastprotonpumpsunder salt stress. JournalofPlantPhysiology167, 261–269.doi:10.1016/j.jplph.2009.09.010
Jensen CR, Jacobsen SE, AndersenMN, NúñezN, Andersen SD, RasmussenL, Mogensen VO (2000) Leaf gas exchange and water relationcharacteristics of field quinoa (Chenopodium quinoa Willd.) duringsoil drying. European Journal of Agronomy 13, 11–25. doi:10.1016/S1161-0301(00)00055-1
Kavi Kishor PB, Sangam S, Amruth RN, Sri Laxmi P, NaiduKR, RaoKRSS,SreenathRaoReddyKJ, TheriappanP, SreenivasuluN (2005)Regulationof proline biosynthesis, degradation, uptake and transport in higher plants:its implications in plant growth and abiotic stress tolerance. CurrentScience 88, 424–438.
Koyro HW, Eisa SS (2008) Effect of salinity on composition, viability andgermination of seeds of Chenopodium quinoaWilld. Plant and Soil 302,79–90. doi:10.1007/s11104-007-9457-4
Kusano T, Berberich T, Tateda C, Takahashi Y (2008) Polyamines: essentialfactors for growth and survival. Planta 228, 367–381. doi:10.1007/s00425-008-0772-7
830 Functional Plant Biology F. Orsini et al.

Lazarus BE, Richards JH, Gordon PE, Oki LR, Barnes CS (2011) Plasticitytradeoffs in salt tolerance mechanisms among desert Distichlis spicatagenotypes.Functional Plant Biology 38, 187–198. doi:10.1071/FP10192
Liphschitz N, Waisel Y (1982) Adaptation of plants to saline environments:salt excretion and glandular structure. In ‘Tasks for vegetation science’.(Eds DN Sen, KS Rajpurohit) pp. 197–214. (Dr W Junk Publishers:The Hague, The Netherlands)
Liu L, Shelp BJ (1996) Impact of chloride on nitrate absorption andaccumulation by broccoli (Brassica oleracea var. italica). CanadianJournal of Plant Science 76, 367–377. doi:10.4141/cjps96-066
Liu J-H, Inoue H, Moriguchi T (2008) Salt stress-mediated changes in freepolyamine titers and expression of genes responsible for polyaminebiosynthesis of apple in vitro shoots. Environmental and ExperimentalBotany 62, 28–35. doi:10.1016/j.envexpbot.2007.07.002
Lohaus G, HussmannM, Pennewiss K, Schneider H, Zhu J-J, Sattelmacher B(2000) Solute balance of a maize (Zea mays L.) source leaf as affected bysalt treatment with special emphasis on phloem retranslocation and ionleaching. Journal of Experimental Botany 51, 1721–1732. doi:10.1093/jexbot/51.351.1721
Luan S, Lan W, Chul Lee S (2009) Potassium nutrition, sodium toxicity andcalciumsignalling: connections through theCBL-CIPKnetwork.CurrentOpinion in Plant Biology 12, 339–346. doi:10.1016/j.pbi.2009.05.003
MaggioA, PascaleSD,RuggieroC,BarbieriG (2005)Physiological responseof field grown cabbage to salinity and drought stress. European Journalof Agronomy 23, 57–67. doi:10.1016/j.eja.2004.09.004
Matysik J, Ali Bhalu B, Mohanty P (2002) Molecular mechanism ofquenching of reactive oxygen species by proline under water stress inplants. Current Science 82, 525–532.
Munns R (2002) Comparative physiology of salt and water stress. Plant, Cell& Environment 25, 239–250. doi:10.1046/j.0016-8025.2001.00808.x
Munns R, Tester M (2008) Mechanisms of salinity tolerance. Annual Reviewof Plant Biology 59, 651–681. doi:10.1146/annurev.arplant.59.032607.092911
OrsiniF,D’UrzoMP, InanG,SerraS,OhDH,MickelbartMV,ConsiglioF,LiX, Jeong JC, Yun DJ, Bohnert HJ, Bressan RA, Maggio A (2010)A comparative study of salt tolerance parameters in 11 wild relativesofArabidopsis thaliana. Journal of Experimental Botany 61, 3787–3798.doi:10.1093/jxb/erq188
Rosa M, Hilal M, Gonzalez JA, Prado FE (2009) Low-temperature effect onenzyme activities involved in sucrose–starch partitioning in salt-stressedand salt acclimated cotyledons of quinoa (Chenopodium quinoa Willd.)seedlings.Plant Physiology and Biochemistry 47, 300–307. doi:10.1016/j.plaphy.2008.12.001
Ruffino AMC, Rosa M, Hilal M, Gonzalez JA, Prado FE (2010) The roleof cotyledon metabolism in the establishment of quinoa (Chenopodiumquinoa) seedlings growing under salinity. Plant and Soil 326, 213–224.doi:10.1007/s11104-009-9999-8
Salisbury EJ (1927) On the causes and ecological significance of stomatalfrequency with special reference to the woodland flora. PhilosophicalTransactions of the Royal Society B 216, 848–852.
Santa-Cruz A, Perez-Alfocea F, Caro M, Acosta M (1998) Polyamines asshort term salt tolerance traits in tomato. Plant Science 138, 9–16.doi:10.1016/S0168-9452(98)00143-5
Scaramagli S, Franceschetti M, Michael AJ, Torrigiani P, Bagni N (1999)Polyamines and flowering: spermidine biosynthesis in the differentwhorls of developing flowers of Nicotiana tabacum L. PlantBiosystems 133, 229–237. doi:10.1080/11263509909381554
Shabala S,DemidchikV, Shabala L,CuinTA, Smith SJ,MillerAJ, Davis JM,Newman IA (2006) Extracellular calcium ameliorates NaCl induced K+
loss fromArabidopsis root and leaf cells by controlling plasmamembraneK+ permeable channels. Plant Physiology 141, 1653–1665. doi:10.1104/pp.106.082388
Szabados L, Savouré A (2010) Proline: a multifunctional amino acid. Trendsin Plant Science 15, 89–97. doi:10.1016/j.tplants.2009.11.009
Tamai T, Shimada Y, Sugimoto T, Shiraishi N, Oji Y (2000) Potassiumstimulates the efflux of putrescine in roots of barley seedlings. Journalof Plant Physiology 157, 619–626.
Verslues PE,AgarwalM,Katiyar-Agarwal S, Zhu J, Zhu J-K (2006)Methodsandconcepts inquantifying resistance todrought, salt and freezing, abioticstresses that affect plant water status. The Plant Journal 45, 523–539.doi:10.1111/j.1365-313X.2005.02593.x
Wilson C, Read JJ, Abo-Kassem E (2002) Effect of mixed salt salinity ongrowth and ion relations of a quinoa and a wheat variety. Journal of PlantNutrition 25, 2689–2704. doi:10.1081/PLN-120015532
Manuscript received 12 April 2011, accepted 23 July 2011
Functional response to salinity in quinoa Functional Plant Biology 831
http://www.publish.csiro.au/journals/fpb
Copyright © 2022 FDOKUMEN




















