
Between-taxon matching of common and rare species richness patterns
23
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/281909981 Between-taxon matching of common and rare species richness patterns ARTICLE · DECEMBER 2015 DOI: 10.1111/geb.12372 READS 62 3 AUTHORS, INCLUDING: Carl Reddin University of Nantes 4 PUBLICATIONS 1 CITATION SEE PROFILE All in-text references underlined in blue are linked to publications on ResearchGate, letting you access and read them immediately. Available from: Carl Reddin Retrieved on: 04 April 2016
-
Upload
mfn-berlin -
Category
Documents
-
view
1 -
download
0
Transcript of Between-taxon matching of common and rare species richness patterns

Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/281909981
Between-taxonmatchingofcommonandrarespeciesrichnesspatterns
ARTICLE·DECEMBER2015
DOI:10.1111/geb.12372
READS
62
3AUTHORS,INCLUDING:
CarlReddin
UniversityofNantes
4PUBLICATIONS1CITATION
SEEPROFILE
Allin-textreferencesunderlinedinbluearelinkedtopublicationsonResearchGate,
lettingyouaccessandreadthemimmediately.
Availablefrom:CarlReddin
Retrievedon:04April2016

1
Between taxa matching of common and rare species richness patterns 1
Reddin C.J.1*, J.H. Bothwell2, J.J. Lennon1 2
Addresses: 1School of Biological Sciences, Queen's University Belfast, 97 Lisburn Road, Belfast BT9 3
7BL; 2School of Biological and Biomedical Sciences, Durham University, South Road, Durham, DH1 4
3LE. 5
Email: Carl Reddin, [email protected]; John Bothwell, [email protected]; Jack Lennon, 6
Key words: Species richness, rarity, surrogacy, indicator, marine, spatial, intertidal ecology 8
Running-title: Common and rare species distributions between taxa 9
*Corresponding author: [email protected] 10
Word count: Abstract, 295; Main text, approx. 4310 words (excluding figure legends). 11
Six figures, one table, Supporting Information: Appendix S1, S2 12
Number of references: 50 13
This is the pre-peer reviewed version of the following article: Reddin, C. 14
J., Bothwell, J. H., & Lennon, J. J. (2015). Between‐taxon matching of 15
common and rare species richness patterns. Global Ecology and 16
Biogeography, which has been published in final form here. 17
18

2
ABSTRACT 19
Aim. The contribution of rare and common species to overall species richness patterns within taxa 20
typically shows that common species explain most of the variation, but rare species sometimes 21
contribute more than expected for their range sizes. Given their importance within taxa, we might 22
hypothesise common species to drive richness pattern similarity between taxa, albeit for different 23
reasons. We studied three marine assemblages focusing on how common and rare species, arranged 24
along a continuous spectrum of range size, match each other. We expected two consumer taxa 25
(molluscs and crustaceans) to match most and additionally, a general signal of stronger common-26
common matches relative to rare-rare or rare-common matches between taxa. 27
Location. UK intertidal zone. 28
Methods. We used high resolution marine datasets for UK intertidal macroalgae, molluscs and 29
crustaceans each with > 400 species. We adapted current methods for within-taxon analyses where 30
rarity and commonness are treated as a continuous spectrum. Pattern-matching was determined by 31
spatial cross-correlation; strength and significance was estimated within and between taxa. 32
Results. Within taxa, common species drove richness patterns as expected, but information-scaled 33
patterns favoured rare species. Between-taxa, our hypothesis that the consumer pair would match 34
most was not supported. Small sub-assemblages (< 60 species) of common species produced the 35
maximum correlation, regardless of taxa pairing, as expected. Pattern-matching between rare 36
species was more idiosyncratic, with maximum correlation occurring between small sub-37
assemblages in only one case. Cross-correlations between common and rare species of different taxa 38
were consistently weak or absent. 39
Main conclusions. Internal structure of richness patterns of these marine taxa does not differ from 40
typical patterns observed in terrestrial taxa. Between taxa, the stronger correlations among common 41
species contrasted with the weak or absent correlation of rare species hints at a coordinated 42
decoupling of processes driving common and rare species richness patterns. 43

3
INTRODUCTION 44
Species richness is distributed non-randomly at multiple spatial (and temporal) scales but different 45
taxa usually do not show strong agreements over the location of richness hotspots (Prendergast et 46
al., 1993a; Reid, 1998; Myers et al., 2000). More generally, studies of similarity among the spatial 47
patterns of richness from different sympatric taxa typically document weak to moderate positive 48
correlations (Wolters et al. 2006; Rodrigues & Brooks, 2007). Such pattern matches are important 49
from the practical standpoint of using more easily recorded taxa as indicators for the more difficult 50
(e.g. Hess et al., 2006). They are also of considerable interest in that the degree to which one taxon’s 51
richness pattern matches another is potentially informative about the ecological and ultimate 52
evolutionary processes producing variation in species richness, a major feature of biodiversity (e.g. 53
Faith, 1992; Cairns et al., 2008). In this paper we present an analysis of richness patterns of three 54
major marine taxa, focusing on how the commonness and rarity of their constituent species 55
contributes to the matching of richness patterns between taxa. 56
It has been known for a long time that ecological communities are typically characterised by few 57
common (widespread) species and many rare (restricted range) species (Grinnell, 1922; Preston, 58
1948; Gaston, 1994; Pachepsky et al., 2001). Species richness analyses usually ignore information on 59
which species are contributing to richness variation (but see Jetz & Rahbek, 2002; Lennon et al., 60
2004). Common species have been found to drive richness patterns within several taxa (Machair 61
grassland vegetation: Lennon et al., 2011; neotropical palms: Kreft et al., 2006; Mexican mammal 62
species: Vásquez & Gaston, 2004; South African and British bird species: Jetz & Rahbek, 2002, 63
Lennon et al., 2004, Evans et al., 2005; and Swiss birds, butterflies and vascular plants: Pearman & 64
Weber, 2007). Much of this might arguably be thought of as obvious: most distribution records are 65
from common species, so species richness patterns are inclined to reflect patterns of common 66
species primarily (Jetz & Rahbek, 2002; Lennon et al., 2004). However, this is apparently obvious 67
only in light of the observation that common species have range sizes that are farther away from 68
complete coverage than the rare species are close to complete absence; the closer a species’ 69

4
distribution is to 50% coverage the greater its potential contribution to species richness patterns. For 70
within-taxon analyses of richness, this effect can be taken into account by weighting species for their 71
information content, or more loosely, as it is used here, the binomial variance (Lennon et al., 2004; 72
also Šizling et al., 2009). Once thus rescaled, in within-taxon analyses rare species typically tend to 73
have a stronger association with full richness patterns than do the commoner species (perhaps 74
excepting the rarest few species; Lennon et al., 2011; Heegaard et al., 2013; but see Vasquez & 75
Gaston, 2004). These studies support the notion that rare species may generally help identify species 76
richness hotspots. 77
Richness pattern structural analyses have hitherto been almost entirely limited to within rather than 78
between taxa. An exception is Pearman & Weber (2007) where species richness patterns of plants, 79
butterflies and birds were split into quartiles of common and rare species, ‘most common’ and ‘red-80
listed’ groups. Spatially explicit correlations within the groups between different taxa pairs showed 81
that only the most common group was significantly correlated among all three taxa. These results 82
suggest that common species may dominate between-taxa relationships; however, caution is 83
needed in that quartiles of ranked species (Gaston, 1994) or other groups may not fit natural 84
transitions from common to rare species, if indeed they exist (e.g. see Siqueira et al., 2012). This lack 85
of a universally applicable cut-off method follows inevitably from the lack of a universally useful way 86
of defining rare species (Gaston, 1994). 87
There is some evidence that common and rare species, either defined by their abundance or range 88
size, differ in their ecology (e.g. Cornwell & Ackerly, 2010). Lennon et al. (2011) used a spatial 89
regression model on subsets of grassland plants and found that common species richness patterns 90
were better explained by simple environmental variables. Similarly, Evans et al. (2005) found that 91
common species contribute more to the species-energy relationship in British birds. We might 92
therefore hypothesise that common species, having apparently stronger relationships with simple 93
environmental variables, might lean towards being environmental generalists, supported 94

5
additionally by their very commonness. These and other studies (Beville & Louda, 1999; Kunin & 95
Gaston, 2007; Matias et al., 2012), have suggested that the environmental requirements of rare 96
species may be more idiosyncratic, and therefore harder to predict. 97
There is a bias towards the terrestrial realm in this field and macroecology in general (Field et al., 98
2009; Tittensor et al., 2010). Studies of richness pattern relationships among taxa were found by 99
Wolters et al. (2006) to be concentrated on forests and grasslands (70% of all), and none on marine 100
systems. Marine and terrestrial systems have traditionally been described as fundamentally 101
different, but recent discussions propose that there may be as much difference within realms as 102
between them in many ways (Dawson & Hamner, 2008; Webb, 2012). More recently, global species 103
richness patterns of coastal taxa have been described as “remarkably congruent” (Tittensor et al., 104
2010). Here we focus on the UK intertidal, which is well sampled and covers a large array of 105
environmental variation (e.g. substrate, wave exposure; Blight et al., 2009). We used three 106
taxonomic assemblages: (1) macroalgae, a speciose group of primary producers defined mainly by 107
size, but typically containing the phyla Chlorophyta, Rhodophyta and Ochrophyta; (2) molluscs (the 108
consumer phylum Mollusca; see, for example, Gladstone, 2002 for indicator relationships with 109
macroalgae and molluscs); and (3) crustaceans (consumer subphylum Crustacea). 110
The most obvious null hypothesis is that all taxa are either equally correlated or uncorrelated with 111
each other. If this null hypothesis is rejected, as perhaps seems likely given previous studies, we can 112
proceed with predictions concerning the degree to which particular taxa are expected to show 113
stronger or weaker associations. For the three taxa considered here, taking a broad trophic 114
perspective we might expect associations between the two consumers groups (molluscs and 115
crustaceans) to be stronger than each is with the primary producers (macroalgae), because of shared 116
heterotrophic requirements of the former two phyla in contrast with the autotrophic requirements 117
of the latter (e.g. Wright et al., 1993). Furthermore, we expected the two producer-consumer pairs 118
to differ: crustaceans v. macroalgae to be strongest and molluscs v. macroalgae to be weakest. This 119

6
is what we would expect if we accept that crustaceans might be, on average, more dependent on 120
the habitat structure provided by macroalgae (Bégin et al., 2004), due to the smaller adult sizes and 121
softer and more permeable exoskeletons of most species, although there are many exceptions for 122
both points. 123
Superimposed on this expected pattern of between-taxa associations is the signal from different 124
parts of the common-rare spectrum within each taxon. For reasons discussed above, we expect the 125
within-taxon associations to favour a match of the common species to the full assemblage, all else 126
being equal and for purely statistical rather than ecological reasons. But when it comes to cross-127
taxon associations, this statistical tendency no longer holds. However, we expect richness patterns 128
from common species to match better between taxa, too. This is because, we suggest, that common 129
species distribution patterns and their associated richness patterns are defined more by where these 130
species are absent than by where they are present. We also suggest that the absences are subject to 131
relatively simple and more consistent causes across species; they are caused by common 132
environmental conditions inimical to species presence such as harsh environments hostile to life in 133
general. In contrast we expect the rare species patterns to match less well because these are 134
defined more by presence than absence (at least compared to the common species) and the reasons 135
for rare species presence we expect to be more environmentally idiosyncratic (Lennon et al., 2011). 136
METHODS 137
Dataset compilation 138
The database was collated in 2007 from the National Biodiversity Network (NBN) Gateway 139
(http://data.nbn.org.uk/) for intertidal macroalgae, molluscs and crustaceans (UK only; Shetlands 140
and other localities excluded due to data inconsistencies; See Fig. 1 and Appendix S1 in Supporting 141
Information). Several procedures were employed to maximise methodological, temporal and 142
identification quality, including objectively removing the oldest tail in the temporal distribution of 143
records, merging records of synonymous taxa (including removing subspecies and species 144

7
morphotypes), and checking that species lists agreed with accepted taxonomic names using the 145
online tool “Match Taxa” (WoRMS Editorial Board, 2013), and objectively removing records 146
suspected to be inaccurate (full details in Appendix S1). The regional species pool was n = 500, n = 147
428 and n = 427 for macroalgae, molluscs and crustaceans, respectively (listed in Appendix S2). 148
Correlations among rare and common species: within- and between-taxa 149
The first part of our analyses focused on within-taxon structure in macroalgae, molluscs and 150
crustaceans. We adopted the method of Lennon et al. (2004; 2011) whereby the full spectrum of 151
range sizes is used, treating rarity and commonness as a continuum. In this approach, a sequence of 152
patterns arises as each species is added in turn. The species are added in order of range size defined 153
as the number of 10 km cells occupied, either from common to rare, beginning with the most 154
widespread species e.g. Fucus vesiculosus for macroalgae, or rare to common, beginning with the 155
most range-restricted species. This approach created two sets of richness patterns (with one set 156
beginning with the most common, the other with the rarest) each pattern of which was then 157
correlated with the pattern for total species richness for the taxon. 158
The second and main part of our analyses focused on between-taxa structure. The sets of richness 159
patterns created for the within-taxon analysis were used to estimate cross-correlations of the spatial 160
distributions of pairs of sub-assemblages of the macroalgae, molluscs and crustaceans. We 161
represented the correlations on orthogonal axes in a 2D space, with coordinates given by the sub-162
assemblage sizes in the two series of richness patterns. Four combinations of these sub-assemblage 163
series were possible for each pair of taxa compared. Where range size ranks were tied, most 164
frequently among rare species, ties were randomised, ranked and correlation values taken as the 165
median value of many repetitions (103 for within-taxon analyses, and 2000 for cross-correlations 166
between taxa). Statistical significance of within-taxon richness correlation coefficients could not be 167
assessed conventionally because the sub-assemblages are subsets of the full assemblage. Instead 168

8
significance was estimated by plotting the median and 95 % confidence intervals of a randomisation 169
null model, given by a random accumulation of species permuted 105 times. 170
The within-taxon richness structure is presented as in Lennon et al. (2004), where the correlations 171
were plotted against sub-assemblage size for the unscaled case, and for the scaled case against the 172
sub-assemblage sum of species’ expected binomial variance p (1 – p), where p is range size as a 173
proportion of total study area. The between-taxa richness composition correlation matrices were 174
plotted similarly, both unscaled and scaled. The scaled plots included a correction to account for 175
different taxa full assemblage sizes, defined by their total binomial variance. In total eight plots were 176
produced per pairwise comparison of taxa (four unscaled, four scaled). Regions of the correlation 177
matrices where values were not statistically significant were highlighted (p > 0.05; 102 iterations, 178
number limited by computational constraints), with their degrees of freedom adjusted to account 179
for spatial autocorrelation (Dutilleul et al., 1993, available in R package ‘SpatialPack’, Vallejos et al., 180
2013). All analyses were performed in the R statistics package (R Development Core Team, 2008). 181
RESULTS 182
The majority of species from all three taxa were rare (Fig. 2) although some species from each were 183
present across the majority of the study extent (e.g. Fucus vesiculosus Linnaeus, 1753, Mytilus edulis 184
Linnaeus, 1758, and Semibalanus balanoides (Linnaeus, 1767); the most widespread species for 185
macroalgae, molluscs and crustaceans, respectively). Full assemblage correlations were highest 186
between macroalgae and crustaceans (r = 0.679, d.f. = 180.6, p < 0.0001), followed by molluscs and 187
crustaceans (r = 0.579, d.f. = 211.8, p < 0.0001), and finally macroalgae and molluscs (r = 0.514, d.f. = 188
225.3, p < 0.0001). 189
Within-taxon structure 190
Within-taxa species richness was found to be strongly determined by common species for all three 191
groups (Fig. 3a-c). When the contribution of individual species was adjusted for their binomial 192

9
variance (Fig. 3d-f), rare species were more strongly associated with the full species richness than 193
common species for all three taxa. 194
Between-taxa structure 195
We classified the correlation surface plots into three general patterns to aid discussion of the results 196
(Table 1). A peak in correlation strength situated closer to the origin (few species used, e.g. Fig. 4a) 197
than to the complete assemblages point (top right corner) is a Small Assemblage Peak (SAP). The 198
opposite trend to a SAP, with a peak in correlation strength situated closer to the full assemblages 199
point (top right, e.g. Fig. 5d), is a Large Assemblage Peak (LAP). We observed a third major pattern of 200
a gradient sloping from one side to the other or top to bottom (e.g. Fig. 4f), indicating that the value 201
of the cross-correlation coefficient is relatively unaffected by one of the two compared assemblages. 202
Because this pattern was consistently observed for rare v. common comparisons (i.e. rare to 203
common series v. common to rare series), we called this the Rare-Common Trend (RCT). 204
Cross-correlations between common sub-assemblages consistently displayed a SAP (panel (a) in 205
Figures 4 - 6), where a peak in correlation strength was reached, not immediately, but with fewer 206
than 60 species. When species’ cumulative contribution to the sub-assemblage was scaled for range 207
size (panel (e) in Figures 4 - 6) the effect was an emphasis of the SAP. Maximum correlations 208
between taxa sub-assemblage pairs were the strongest between common sub-assemblages, where 209
values approached r = 0.8 (all p < 0.0001). Although these were consistently stronger than the 210
correlations between the full taxonomic assemblages, maximum strength between common sub-211
assemblages of taxa pairs was not always proportional to the respective full assemblage correlation 212
strength between taxa pairs (Table 1). The SAP generally did not fall on the y = x line but was 213
displaced towards macroalgae in both comparisons involving them (e.g. Fig. 4a), demonstrating that 214
common consumer sub-assemblages (molluscs or crustaceans) tend to be present alongside 215
relatively larger sub-assemblages of common macroalgae (rather than common sub-assemblages of 216
both being similar sized). This effect held after scaling for range size and taxon assemblage size. 217

10
In no cases were common sub-assemblages strongly correlated with rare sub-assemblages (i.e. no 218
SAP patterns, panels (b) and (c) in Fig. 4 - 6), or vice versa. Instead, RCT patterns showed that rare 219
sub-assemblage distributions matched the distribution patterns of full assemblages between taxa. 220
Peak correlation strength varied idiosyncratically relative to the full assemblage correlation between 221
taxa, with the strongest peak between macroalgal full assemblages and rare molluscan sub-222
assemblages (> 10 composite singleton species, r > 0.6, p < 0.0001). However, caution is warranted 223
for the interpretation of correlations including small sub-assemblages, which are likely to have non-224
Gaussian richness distributions. In any case, scaling for range size (panels (f) and (g) in Fig. 4 - 6) 225
clarified the RCT pattern and emphasised the relationship between full assemblages and rare sub-226
assemblages. 227
Zero or very weak correlations (r < 0.2) were found between rare sub-assemblages in two out of 228
three cases (panels (d) in Fig. 4 – 5, consumer taxa against macroalgae) following a LAP pattern. 229
Molluscs and crustaceans (Fig. 6d) demonstrated the only case of a SAP pattern for the rare v. rare 230
case, with small sub-assemblages of rare species being moderately correlated (> 14 singleton 231
species, r > 0.4, maximum r = 0.64, p < 0.0001, between sub-assemblages of 150 molluscan species, 232
range sizes 1 – 4 cells, and 270 crustacean species, range sizes 1 – 5 cells). However, scaling for range 233
size reduced the difference between taxa pairs, de-emphasising the peak between molluscs and 234
crustaceans, and amplifying the LAP patterns (panels (h) in Fig 4 – 6). Correlation strengths were 235
consistently stronger between rare v. rare sub-assemblage pairs than between common v. rare sub-236
assemblage pairs (range size scaled panels (h) v. (f-g) in Fig 4 – 6). 237
DISCUSSION 238
We found that common species drive richness pattern similarity between taxa. This shows a parallel 239
with common species driving richness patterns within taxa, although the underlying reasons for this 240
parallel may not be the same. From comparison of the three taxa we found strongly matching 241
distributions (SAP type) between common species sub-assemblages and weakly matching 242

11
distributions (LAP type) between rare species sub-assemblages. This conclusion was further 243
emphasised by scaling species by their range size. This scaling also reduced the prominence of the 244
single observation of a SAP pattern between rare sub-assemblages of molluscs and crustaceans, 245
suggesting this feature may be of minor significance. 246
Our specific hypotheses of relative correlation strength between taxa full assemblage patterns were 247
not supported, with the strongest correlation observed between crustaceans and macroalgae, rather 248
than crustaceans and molluscs as predicted. Instead of trophic similarities (autotrophy v. 249
heterotrophy) underlying taxa species richness associations (Wolters et al., 2006), a stronger 250
dependence of crustaceans upon the habitat structure provided by macroalgae may be responsible 251
for the strong association between crustacean and macroalgal richness. Habitat structure provided 252
by macroalgae, especially canopy formers, can provide shelter from predation and desiccation stress 253
(Bégin et al., 2004), potentially facilitating populations of many crustacean species which might 254
benefit more from this cover than molluscs. 255
The observed SAP patterns showed maximum cross-correlations to be much stronger than the full-256
assemblage correlations. This demonstrates that, at least in the cases studied here, common species 257
reinforce, and are central to, the congruency between full assemblage richness patterns of different 258
taxa, allowing the use of taxonomic ‘surrogates’ or ‘indicators’ (e.g. Hess et al., 2006). Maximum SAP 259
cross-correlations were not proportional to, and therefore not predictable by, the correlation 260
strengths between full assemblages of the taxa pairs. Instead, the addition of rarer species, reducing 261
the cross-correlation strength, was still important in determining the extent of full assemblage 262
similarity. We found correlations between full assemblages that were substantially higher (e.g. rmin = 263
0.51, rmax = 0.68) than those reported in meta-analyses (e.g. r = 0.37 from Wolters et al. 2006, and a 264
‘Species Accumulation Index’ = 0.41, interpretable similarly to an r value, from Rodrigues & Brooks, 265
2007). Pearman & Weber (2007), despite a negative correlation (r = -0.19, p = 0.25) between bird 266
and butterfly full assemblages, also found that their most common species sub-assemblage was the 267

12
only one that was significantly correlated among all three of their taxa, with correlation strengths 268
exceeding those of full assemblages. Coastal marine taxa may show relatively strong congruency 269
(Tittensor et al., 2010) and our study’s constraint to the intertidal habitat may have increased the 270
likelihood of environmental drivers similarly affecting the distributions of species from the different 271
taxa. After splitting their datasets into high and low altitude subsets, Pearman & Weber (2007) 272
concluded that (within-taxa) correlation strength could be affected by the range of habitat 273
heterogeneity covered by the data (hence also scale and extent; Hess et al., 2006). For instance, if 274
our study extended offshore, where waters increased in depth (increasing habitat heterogeneity), 275
we might have observed a decrease in between-taxa correlation between macroalgae and the 276
consumer taxa as macroalgal species distributions are limited to the photic zone while many 277
consumer species distributions do not directly need light to survive. These examples illustrate the 278
potential effect of environment on the observed between-taxa relationships, which vary between 279
studies (Hess et al., 2006; Wolters et al. 2006; Rodrigues & Brooks, 2007). 280
The displacement in the precise location of the SAP relative to the line y = x towards larger 281
macroalgal common sub-assemblages suggests that between-taxa sub-assemblage associations need 282
not be balanced by numbers of species, nor sub-assemblage relative commonness. Depending on 283
the degree of skewness of the taxon species occupancy distributions, between-taxa associations may 284
peak upon inclusion of less common species from one taxon, such as the ‘frequent’ and ‘occasional’ 285
species we observed for macroalgae. 286
Rare sub-assemblages were not completely uncorrelated between taxa, and generally showed 287
stronger cross-correlations than those between rare and common sub-assemblages. These weak 288
correlations suggest that rare species of different taxa do have a slight tendency to be found 289
together. Traditional means of defining species ‘hotspots’ often indicate little overlap between taxa 290
(Prendergast et al., 1993a; Reid, 1998; Orme et al., 2005), most likely because reasons for 291
occurrence of different rare species from different taxa are inconsistent. The within-taxon analyses 292

13
show that rare species distributions tend to more strongly associate with full assemblage 293
distributions after taking into account binomial variance (Fig. 3; Lennon et al., 2004; 2011; Šizling et 294
al., 2009). This pattern may arise due to the nestedness of species distributions (characteristic of 295
metacommunities and mutualistic networks; Almeida-Neto et al., 2008), where rare species’ 296
occurrences typically tend to fall on top of those of common species (Šizling et al., 2009; Heegaard et 297
al., 2013). A parallel observation in our study saw relatively high cross-correlations between the 298
distributions of rare sub-assemblages and full assemblages of different taxa (RCT patterns), although 299
these cross-correlations did not achieve the strength of within-taxon correlations between 300
distributions of similarly sized rare sub-assemblages and full assemblages. 301
We observed only one instance of a strong cross-correlation (SAP) between rare sub-assemblages, 302
for molluscs and crustaceans. We first explored the possibility that this might emerge because of 303
sampling effort. Working with NBN data on intertidal macroalgae and molluscs, Blight et al. (2009) 304
detected that cells occupied by university marine stations topped the ranks of raw species richness 305
for both. However we observed rare macroalgal sub-assemblages to be distributed dissimilarly to 306
molluscan sub-assemblages (either rare or common). It is possible that experts in the consumer taxa 307
and experts in macroalgae may not frequent the same areas. Upon examination of the geographical 308
distribution of molluscan and crustacean sub-assemblages at this peak of cross-correlation, some of 309
the peaks of co-occurrence fell close to marine stations (possibly well-frequented study sites), but 310
this did not account for all large peaks, nor the numerous minor sites of co-occurrence that were 311
spread across the country. Therefore the cross-correlation peak between molluscan and crustacean 312
rare sub-assemblages does not appear to be solely due to proximity to marine stations. Potential 313
ecological explanations include rare habitat types important for both consumer taxa, but not for 314
macroalgae (intertidal caves for example). Macroalgae require sunlight, mostly inorganic chemicals, 315
and are essentially bound to solid substrate, whilst the consumer taxa are generally not restricted by 316
these conditions (though there are exceptions, such as sessile barnacles in Crustacea). Molluscs and 317
crustaceans can be diverse in sedimentary habitats (e.g. Ellingsen, 2002), whilst macroalgae are 318

14
more restricted to solid substrata (again, there are exceptions). In our case this between-taxa 319
clustering of rare species was likely to be partly attributable to some sites of high sampling effort. 320
Overlap and clustering of UK rare benthic species’ distributions (10 km grain) has been found 321
previously by Sanderson (1996), using a small sample of rare species with reliable distributions (data-322
deficient species were removed). In south-east Australia, although from a small sample size (15 323
sites), Gladstone (2002) recorded a strong correlation (r = 0.93, non-spatial p < 0.01) between 324
molluscan species and species of all taxa, both richnesses weighted to emphasise rare species. Rare 325
species will always be sensitive to sampling effort (e.g. difficult to detect species; Boulinier et al., 326
1998), and are frequently regarded as ‘noise’ and recommended for removal prior to analysis (e.g. 327
Clarke & Green, 1988; Cottenie & De Meester 2003), despite their potential functional importance 328
(Emmerson et al., 2001; Lyons et al., 2005). 329
While our common-to-rare sequences always began with the same commonest (few) species, the 330
rare-to-common sequences did not start with any particular rarest species because there were many 331
such candidates, and so each cross-correlation resulted from a bootstrap over many permutations of 332
randomly ranking the tied ranks (singletons, doubletons, and so forth). This is important because the 333
identity of the commonest species is likely to be ecologically significant (e.g. in species interactions 334
and ecosystem functioning; O’Connor & Crowe, 2005; Gaston, 2010), but the identity of singleton 335
species may not always be important (notwithstanding chance or transient occurrences; Novotný & 336
Basset, 2000; Magurran & Henderson, 2003). 337
Common species have been suggested as responsible for the majority of certain ecosystem functions 338
and services (Walker, 1992; Gaston, 2010) and indeed it would be surprising if this were not the 339
case. Our conclusions suggest that the processes that limit the range of common species may be 340
generally consistent across taxa, provided that certain conditions are maintained (for example, that 341
taxa share the same broadly defined habitat). Studies that have associated patterns of common sub-342
assemblages with environmental variables have suggested that common species are environmental 343

15
generalists (Evans et al., 2005; Lennon et al., 2011) that share an aversion to particularly hostile 344
habitats (e.g. low energy habitats; Evans et al., 2005). 345
Most studies of the contribution of common and rare species to full richness patterns have been 346
from birds (e.g. Lennon et al., 2004; Evans et al., 2005; Pearman & Weber, 2007), which are usually 347
well sampled and have large accessible datasets (e.g. Boulinier et al., 1998). Our study from a marine 348
perspective covering both primary producers (macroalgae) and consumers (molluscs and 349
crustaceans) widens the potential generality of these patterns across realms (Dawson & Hamner, 350
2008; Webb, 2012). In our analysis all three marine intertidal taxa supported the emerging general 351
trend that, typically (i.e. with right-skewed species’ occupancy distributions, Heegaard et al., 2013), 352
common species drive species richness patterns (Jetz & Rahbek 2002, Lennon et al., 2004), but when 353
species’ contributions are corrected for their range size it is typically the rare species that are most 354
associated with overall richness patterns (Lennon et al., 2004; Šizling et al., 2009). Atypical datasets 355
(in which species occupancy distributions are not right-skewed) may still present complications 356
(Šizling et al., 2009; Heegaard et al., 2013), which is an important potential limit to generalisation. 357
358
ACKNOWLEDGEMENTS. CJR was funded by the Department of Employment and Learning 359
Northern Ireland as part of his PhD thesis. We thank dataset providers British Phycological Society, 360
Natural Resources Wales, Conchological Society of Great Britain & Ireland, English Nature, Joint 361
Nature Conservation Committee, Marine Biological Association, Centre for Environmental Data and 362
Recording, Marine Conservation Society, and Scottish Natural Heritage. 363
364
REFERENCES 365
Almeida‐Neto, M., Guimaraes, P., Guimarães, P.R., Loyola, R.D. & Ulrich, W. (2008) A consistent metric for 366 nestedness analysis in ecological systems: reconciling concept and measurement. Oikos, 117, 1227-367 1239. 368

16
Bégin, C., Johnson, L.E. & Himmelman, J.H. (2004) Macroalgal canopies: distribution and diversity of associated 369 invertebrates and effects on the recruitment and growth of mussels. Marine Ecology Progress Series, 370 271, 121-32. 371
Blight, A.J., Allcock, L.A., Maggs, C.A. & Johnson, M.P. (2009) Intertidal molluscan and algal species richness 372 around the UK coast. Marine Ecology Progress Series, 396, 235-243. 373
Boulinier, T., Nichols, J.D., Sauer, J.R., Hines, J.E. & Pollock, K.H. (1998) Estimating species richness: The 374 importance of heterogeneity in species detectability. Ecology, 79, 1018–1028. 375
Cairns, D.K., Gaston, A.J. & Huettmann, F. (2008) Endothermy, ectothermy and the global structure of marine 376 vertebrate communities. Marine Ecology Progress Series, 356, 239-250. 377
Clarke, K.R. & Green, R.H. (1988) Statistical design and analysis for a" biological effects" study. Marine Ecology 378 Progress Series, 46, 213-226. 379
Cornwell, W.K. & Ackerly, D.D. (2010) A link between plant traits and abundance: evidence from coastal 380 California woody plants. Journal of Ecology, 98, 814-821. 381
Cottenie, K. & De Meester, L. (2003) Connectivity and cladoceran species richness in a metacommunity of 382 shallow lakes. Freshwater Biology, 48, 823-832. 383
Dawson, M.N. & Hamner, W.M. (2008) A biophysical perspective on dispersal and the geography of evolution 384 in marine and terrestrial systems. Journal of the Royal Society Interface, 5, 135-150. 385
Dutilleul, P., Clifford, P., Richardson, S. & Hemon, D. (1993) Modifying the t test for assessing the correlation 386 between two spatial processes. Biometrics, 49, 305-314. 387
Ellingsen, K.E. (2002) Soft-sediment benthic biodiversity on the continental shelf in relation to environmental 388 variability. Marine Ecology Progress Series, 232, 15-27. 389
Emmerson, M.C., Solan, M., Emes, C., Paterson, D.M. & Raffaelli, D. (2001) Consistent patterns and the 390 idiosyncratic effects of biodiversity in marine ecosystems. Nature, 411, 73-77. 391
Evans, K.L., Greenwood, J.J. & Gaston, K.J. (2005) Relative contribution of abundant and rare species to 392 species–energy relationships. Biology letters, 1, 87-90. 393
Faith, D. P. (1992) Conservation evaluation and phylogenetic diversity. Biological Conservation, 61, 1-10. 394
Field, R., Hawkins, B.A., Cornell, H.V., Currie, D.J., Diniz‐Filho, J.A. F., Guégan, J.F., Kaufman, D.M., Kerr, J.T., 395 Mittelbach, G.G., Oberdorff, T., O’Brien, E.M. & Turner, J.R. (2009) Spatial species‐richness gradients 396 across scales: a meta‐analysis. Journal of Biogeography, 36, 132-147. 397
Gaston, K.J. (2010) Valuing common species. Science, 327, 154-155. 398
Gaston, K.J. (1994) Rarity, pp. 2. Chapman & Hall, London. 399
Gladstone, W. (2002) The potential value of indicator groups in the selection of marine reserves. Biological 400 Conservation, 104, 211-220. 401
Grinnell, J. (1922) The role of the" accidental". The Auk, 39, 373-380. 402
Heegaard, E., Gjerde, I. & Sætersdal, M. (2013) Contribution of rare and common species to richness patterns 403 at local scales. Ecography, 36, 937–946, 404
Hess, G.R., Bartel, R.A., Leidner, A.K., Rosenfeld, K.M., Rubino, M.J., Snider, S.B. & Ricketts, T.H. (2006). 405 Effectiveness of biodiversity indicators varies with extent, grain, and region. Biological Conservation, 406 132, 448-457. 407

17
Hourigan, T.F., Timothy, C.T. & Reese, E.S. (1988) Coral reef fishes as indicators of environmental stress in coral 408 reefs. In Marine Organisms as Indicators, pp. 107-135. Springer, New York. 409
Jetz, W. & Rahbek, C. (2002) Geographic range size and determinants of avian species richness. Science, 297, 410 1548-1551. 411
Kreft, H., Sommer, J.H. & Barthlott, W. (2006) The significance of geographic range size for spatial diversity 412 patterns in Neotropical palms. Ecography, 29, 21-30. 413
Lennon, J.J., Koleff, P., Greenwood, J.J. & Gaston, K.J. (2004) Contribution of rarity and commonness to 414 patterns of species richness. Ecology Letters, 7, 81-87. 415
Lennon, J.J., Beale, C.M., Reid, C.L., Kent, M. & Pakeman, R.J. (2011) Are richness patterns of common and rare 416 species equally well explained by environmental variables? Ecography, 34, 529-539. 417
Lyons, K.G., Brigham, C.A., Traut, B.H. & Schwartz, M.W. (2005) Rare species and ecosystem functioning. 418 Conservation Biology, 19, 1019-1024. 419
Magurran, A.E. & Henderson, P.A. (2003) Explaining the excess of rare species in natural species abundance 420 distributions. Nature, 422, 714-716. 421
Matias, M.G., Chapman, M.G., Underwood, A.J. & O'Connor, N.E. (2012) Increasing density of rare species of 422 intertidal gastropods: tests of competitive ability compared with common species. Marine Ecology 423 Progress Series, 453, 107-116. 424
Myers, N., Mittermeier, R.A., Mittermeier, C.G., Da Fonseca, G.A. & Kent, J. (2000) Biodiversity hotspots for 425 conservation priorities. Nature, 403, 853-858. 426
Novotný, V. & Basset, Y. (2000) Rare species in communities of tropical insect herbivores: pondering the 427 mystery of singletons. Oikos, 89, 564-572. 428
O'Connor, N.E. & Crowe, T.P. (2005) Biodiversity loss and ecosystem functioning: distinguishing between 429 number and identity of species. Ecology, 86, 1783-1796. 430
Orme, C.D.L., Davies, R.G., Burgess, M., Eigenbrod, F., Pickup, N., Olson, V.A., Webster, A.J., Ding, T., 431 Rasmussen, P.C., Ridgely, R.S., Stattersfield, A.J., Bennett, P.M., Blackburn, T.M., Gaston, K.J. & 432 Owens, I.P. (2005) Global hotspots of species richness are not congruent with endemism or threat. 433 Nature, 436, 1016-1019. 434
Pachepsky, E., Crawford, J.W., Bown, J.L. & Squire, G. (2001) Towards a general theory of biodiversity. Nature, 435 410, 923-926. 436
Pearman, P.B. & Weber, D. (2007) Common species determine richness patterns in biodiversity indicator taxa. 437 Biological Conservation, 138, 109-119. 438
Prendergast, J.R., Quinn, R.M., Lawton, J.H., Eversham, B. C. & Gibbons, D. W. (1993a) Rare species, the 439 coincidence of diversity hotspots and conservation strategies. Nature, 365, 335-337. 440
Prendergast, J., Wood, S., Lawton, J. & Eversham, B. (1993b) Correcting for variation in recording effort in 441 analyses of diversity hotspots. Biodiversity Letters, 1, 39-53. 442
Preston, F.W. (1948) The commonness, and rarity, of species. Ecology, 29, 254-283. 443
R Development Core Team (2008) R: A language and environment for statistical computing. R Foundation for 444 Statistical Computing, Vienna, Austria. http://www.R-project.org 445
Reid, W.V. (1998) Biodiversity hotspots. Trends in Ecology & Evolution, 13, 275-280. 446
Rodrigues, A.S.L. & Brooks, T.M. (2007) Shortcuts for Biodiversity Conservation Planning: The Effectiveness of 447 Surrogates. Annual Review of Ecology, Evolution, and Systematics, 38, 713–37. 448

18
Siqueira, T., Bini, L.M., Roque, F.O., Marques Couceiro, S.R., Trivinho‐Strixino, S. & Cottenie, K. (2012) Common 449 and rare species respond to similar niche processes in macroinvertebrate metacommunities. 450 Ecography, 35, 183-192. 451
Šizling, A.L., Šizlingová, E., Storch, D., Reif, J. & Gaston, K.J. (2009) Rarity, commonness, and the contribution of 452 individual species to species richness patterns. The American Naturalist, 174, 82-93. 453
Tittensor, D.P., Mora, C., Jetz, W., Lotze, H.K., Ricard, D., Berghe, E.V. & Worm, B. (2010) Global patterns and 454 predictors of marine biodiversity across taxa. Nature, 466, 1098-1101. 455
Vallejos, R., Osorio, F., & Cuevas, F. (2013) SpatialPack - An R package for computing spatial association 456 between two stochastic processes defined on the plane. Submitted. 457
Vázquez, L.B. & Gaston, K.J. (2004) Rarity, commonness, and patterns of species richness: the mammals of 458 Mexico. Global Ecology and Biogeography, 13, 535-542. 459
Walker, B.H. (1992) Biodiversity and ecological redundancy. Conservation Biology, 6, 18-23. 460
Webb, T.J. (2012). Marine and terrestrial ecology: unifying concepts, revealing differences. Trends in ecology & 461 evolution, 27, 535-541. 462
Wolters, V., Bengtsson, J. & Zaitsev, A.S. (2006) Relationship among the species richness of different taxa. 463 Ecology, 87, 1886-1895. 464
WoRMS Editorial Board (2013) World Register of Marine Species. Available from 465 http://www.marinespecies.org at VLIZ. Accessed 2013-11-28 466
Wright, D.H., Currie, D.J. & Maurer, B.A. (1993) Energy supply and patterns of species richness on local and 467 regional scales. In: Species diversity in ecological communities: historical and geographical 468 perspectives. (ed. by R.E. Ricklefs & D. Schluter), pp. 66-74. Chicago University Press, Chicago. 469
470
BIOSKETCH 471
Carl Reddin is quantitative marine ecologist with an interest in macroecological methods and their 472
use to extend our understanding of biodiversity patterns across realms. 473
John Bothwell works on seaweed, primarily on their evolution and systematics, but also on their 474
bioenergy potential and ecological importance along the UK’s coastlines. 475
Jack Lennon's main research interests include understanding how environmental constraints and 476
processes act on the spatial and temporal distribution of species to create and maintain emergent 477
diversity patterns. 478
479

19
TABLES 480
Table 1. Maximum correlation between taxa (of any sub-assemblage comparison i.e. the peak value 481 in the correlation surfaces shown in Figure 4-6) and its coordinates on the unscaled correlation 482 surfaces, given as the proportion of each full assemblage represented by the sub-assemblages 483 correlated (Common, C; Rare, R). Also given is a classification of the cross-correlation plots according 484 to major surface trends (Small Assemblage Peak, SAP; Rare Common Trend, RCT; Large Assemblage 485 Peak, LAP). The correlation for the full taxon comparisons, in bold, is given as rtotal. Figure numbers 486 for the correlation surfaces are indicated. 487
Sub-assemblage pair C v. C C v. R R v. C R v. R
Figure a,e b,f c,g d,h
Type Type Type Type
Macroalgae v. molluscs 4 SAP RCT RCT LAP
Macroalgae v. crustaceans 5 SAP RCT RCT LAP
Molluscs v. crustaceans 6 SAP RCT RCT SAP
rtotal rmax rmax rmax rmax
Macroalgae v. molluscs 0.51 0.79 (.05, .02) 0.63 (1.0, .04) 0.53 (.21, 1.0) 0.51 (1.0, 1.0)
Macroalgae v. crustaceans 0.68 0.76 (.09, .04) 0.72 (.93, .27) 0.69 (.53, 1.0) 0.69 (.96, 1.0)
Molluscs v. crustaceans 0.58 0.74 (.05, .05) 0.60 (1.0, .09) 0.61 (.10, 1.0) 0.64 (.35, .63)
488
489
FIGURE LEGENDS 490
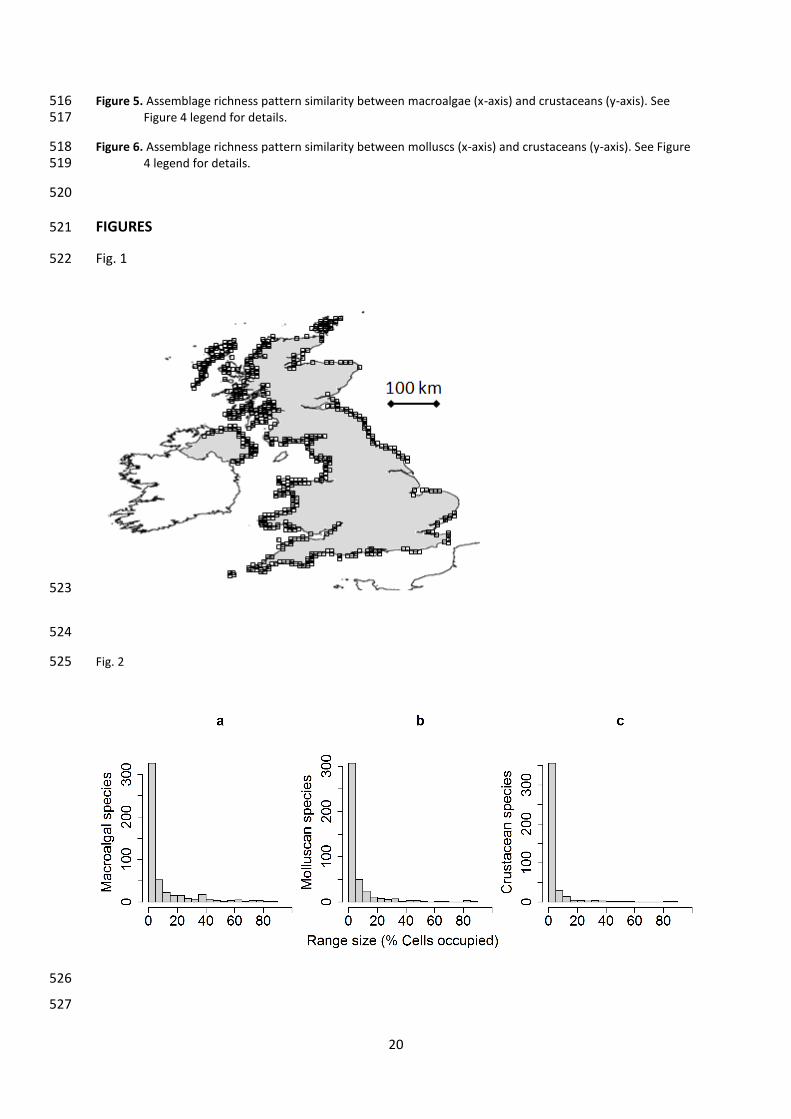
Figure 1. Coastline map of the UK showing the 10 km cells (n = 522) occupied by at least one record for each 491 taxon. Latitude min = 50°N, max = 59°N, approximately. 492
Figure 2. The range size of intertidal species, (a) macroalgae, (b) molluscs and (c) crustaceans, showing that 493 most species were rare (small range size). Total cell numbers were 522 for all taxa. 494
Figure 3. The correlation of sub-assemblages built from common to rare (thin solid line), or rare to common 495 (dashed line), with the full species richness for macroalgae (a & d), molluscs (b & e) or crustaceans (c 496 & f). The top row (a-c) shows sub-assemblages built species-by-species, from a single species to the 497 full species richness, while the bottom row (d-f) shows sub-assemblages built from cumulative 498 binomial variance (species contribute varying amounts depending upon their occupancy). The grey 499 region shows the null hypothesis of correlation with the full assemblage (95% confidence intervals of 500 the mean, while the thick solid line is the median, derived from 104 randomly permuted 501 accumulations of species from the species pool). 502
Figure 4. Assemblage richness pattern similarity between macroalgae (x-axis) and molluscs (y-axis). Axes 503 represent sub-assemblages, built species-by-species from the origin of the plot, beginning with: 504 commonest ‘x’ against commonest ‘y’ (panels a,e); commonest ‘x’ against rarest ‘y’ (panels b,f); rarest 505 ‘x’ against commonest ‘y’ (panels c,g); rarest ‘x’ against rarest ‘y’ (panels d,h); the diagonal line is y = 506 x, equal assemblage growth. Top row (panels a,b,c,d) represents unscaled sub-assemblages, 507 increasing by species number (i.e. each species contributes ‘1’ to the sub-assemblage), while bottom 508 row (panels e,f,g,h) represents scaled sub-assemblages, by increasing species binomial variance (i.e. 509 weighting by information content) scaled in proportion to the largest assemblage size; the smaller 510 assemblage was multiplied by the size difference between the two taxonomic assemblages, putting 511
the smaller assemblage on equal footing with the larger, 𝑝𝑎 (1 − 𝑝𝑎) (∑ 𝑝𝑏 (1−𝑝𝑏)
∑ 𝑝𝑎(1−𝑝𝑎)) , where pa and pb 512
were range sizes, as proportions of total study area, for the taxon of smallest and largest full 513 assemblage size, respectively. Delineated grey regions are regions where the correlations were not 514 significant (p > 95, corrected for spatial autocorrelation). 515
20
Figure 5. Assemblage richness pattern similarity between macroalgae (x-axis) and crustaceans (y-axis). See 516 Figure 4 legend for details. 517
Figure 6. Assemblage richness pattern similarity between molluscs (x-axis) and crustaceans (y-axis). See Figure 518 4 legend for details. 519
520
FIGURES 521
Fig. 1 522
523
524
Fig. 2 525
526
527

21
Fig. 3 528
529
530
Fig. 4 531
532
533

22
Fig. 5 534
535
Fig. 6 536
537





![Binder 200, Small families [Trematoda Taxon Notebooks]](https://static.fdokumen.com/doc/165x107/6324444cb104cba27a091035/binder-200-small-families-trematoda-taxon-notebooks.jpg)






![Binder 216, Terminology [Trematoda Taxon Notebooks]](https://static.fdokumen.com/doc/165x107/63338ab23108fad7760f19c8/binder-216-terminology-trematoda-taxon-notebooks.jpg)






