
Behaviour of 99Tc in aqueous solutions in the presence of iron oxides and microorganisms
11
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/authorsrights
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Behaviour of 99Tc in aqueous solutions in the presence of iron oxides and microorganisms

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/authorsrights

Author's personal copy
Behaviour of 99Tc in aqueous solutions in the presence of iron oxidesand microorganisms
Rūta Druteikienė a,n, Benedikta Lukšienė a, Dalia Pečiulytė b, Kęstutis Mažeika a,Arūnas Gudelis a, Dalis Baltrūnas a
a Center for Physical Sciences and Technology, Savanorių ave. 231, LT-02300 Vilnius, Lithuaniab Nature Research Center, Akademijos str. 2, LT-08412 Vilnius, Lithuania
H I G H L I G H T S
� Abilities of wustite/magnetite and hematite to accumulate 99Tc(VII) were investigated.� The effect of microbial activity on the migration behaviour of technetium was tested.� Gradual sorption of [99Tc] TcO�
4 from aquatic solution onto wustite/magnetite (FeO/Fe3O4) under aerobic conditions was observed.� Specific microorganisms induced sorption of 99Tc(VII) by hematite.
a r t i c l e i n f o
Article history:Received 23 April 2013Received in revised form13 January 2014Accepted 18 February 2014Available online 25 February 2014
Keywords:TechnetiumWustite/magnetiteHematiteMicroorganism
a b s t r a c t
A set of experiments was performed to determine the factors that influence TcO�4 interaction with Fe-
bearing minerals and to explore the effect of microbial activity on the behaviour of Tc(VII) in solution, inthe presence of iron oxides under oxidizing medium. Gradual sorption of TcO�
4 (aq) onto wustite/magnetite was observed under alkaline conditions (pH 8–9). No pronounced effect of TcO�
4 (aq)interaction with hematite was observed in the investigating alkaline systems. At low pH values (2.7–4.5), TcO�
4 retention on hematite increases, suggesting that the process is dependent on pH. Sorption of99Tc (VII) onto hematite at pH 7.6–8.0 was achieved because of the presence of specific microorganisms.
& 2014 Elsevier Ltd. All rights reserved.
1. Introduction
The long-lived radionuclides produced by nuclear accidentsand in the operative fuel cycle represent a major concern to thepublic if they enter the environment (Liljenzin, 2002; Choppin,2005; Jaisi et al., 2009; Buechele et al., 2012). To assess thepotential of these radionuclides to cause harm to humans, theirgeochemical and biological behaviour must be evaluated. Thefission product 99Tc is presently considered as a key mobileradionuclide for the disposal of spent fuel and high-level radio-active waste (Begg et al., 2007). Nuclear installations generate arelatively large amount of 99Tc (half-life �2.1�105 years), whichoriginates from a 6% yield from the fission of 235U in nuclear powerplants (Maset et al., 2006). Technetium has been introduced intothe environment because of authorized and accidental discharges.For example, about 140 PBq of 99Tc was produced in nuclear power
plants worldwide from 1976 to 2006, and it was determined thatuntil 2010, nearly 10 GBq of 99Tc was released to the environment(Shi et al., 2012). Because most of 99Tc remains in the nuclear fuel,it is one of the most important radionuclides with respect to themanagement of spent fuel and radioactive waste. Techetium-99 isrelevant in geochemical systems of both intermediate and high-level nuclear waste repositories; it is present in aquifers, themarine environment and rivers (Standring et al., 2002; Keith-Roach et al., 2003; Frederickson et al., 2004; Burke et al., 2006; Shiet al., 2012). Therefore, 99Tc mobility is of major interest inenvironmental science, and is an important concern for the safedisposal of radioactive waste.
Most of the studies investigating geochemical processes andtheir impact on the transformation and dynamics of technetiumhave demonstrated that the Tc behaviour is complex, dependingon a series of factors such as electron transfer reactions (redox),biogenic factors, and microbial activity (Tagami and Uchida, 1996;Istok et al., 2004; Abdelouas et al., 2005; Peretyazhko et al., 2012).Technetium is a redox sensitive radionuclide and its migrationbehaviour primarily depends on its oxidation state, which in turn
Contents lists available at ScienceDirect
journal homepage: www.elsevier.com/locate/apradiso
Applied Radiation and Isotopes
http://dx.doi.org/10.1016/j.apradiso.2014.02.0200969-8043 & 2014 Elsevier Ltd. All rights reserved.
n Corresponding author. Tel.: þ370 5 2661648; fax: þ370 5 2602317.E-mail address: [email protected] (R. Druteikienė).
Applied Radiation and Isotopes 89 (2014) 85–94

Author's personal copy
depends on redox conditions of the surrounding environment (Lieserand Bauscher, 1987; Lloyd and Macaskie, 1996; Wildung et al., 2000;Abdelouas et al., 2005; Begg et al., 2007; Zachara et al., 2007). Underoxic conditions, technetium forms the highly soluble pertechnetateanion, TcO�
4 , which weakly sorbs onto mineral surfaces (Lieser andBauscher, 1987). However, the reduction and oxidation conditions ofgeochemical surroundings can bring about an alteration of thetechnetium oxidation state. In anoxic surroundings, the pertechne-tate anion TcO�
4 is reduced to the lower valence form of Tc(IV),which through hydrolysis forms a sparingly soluble solid phase (e.g.,TcO2 nH2O) which can be sorbed to mineral phases or organicfractions (Maes et al., 2004; Burke et al., 2005). A number ofexperiments have been conducted in order to assess the redoxbehaviour of technetium in natural sediments containing sediment-associated Fe and Mn oxide, metal-reducing bacteria, organic co-contaminants, or dissolved iron and iron minerals (Fredericksonet al., 2004; Maset et al., 2006; Peretyazhko et al., 2009). Reductionof TcO�
4 under iron- and nitrate-reducing conditions, as well asunder sulphate-reducing conditions, was observed (Wharton et al.,2000; Istok et al., 2004; Burke et al., 2005; Abdelouas et al., 2005).
The treatment of high-level radioactive waste and managementof the nuclear waste repositories are relevant to the technetiumchemistry. The main problem is the presence of different techne-tium species in heterogeneous waste (Lukens et al., 2002). Radi-olysis of pertechnetate has been examined in the high-levelnuclear waste tanks at the Savannah River and Hanford Sites(Lukens et al., 2001, 2002). The results show that the radiolysis ofTcO�
4 in highly alkaline solution in the presence of O scavengers isone more mechanism of reducing technetium by NO3
2–
. Radiolysisof TcO�
4 in alkaline solutions containing organic compounds,causes the formation of both insoluble radiolysis product TcO2
nH2O, and the soluble, lower-valent Tc complexes.Interaction between technetium and microorganisms plays an
important role in the environmental fate of the radionuclide witha multitude of mechanisms affecting transformations of its oxida-tion state (Marshall et al., 2009; Mohapatra et al., 2010; Borshet al., 2010). Studies of Tc behaviour in biologically active, organic-rich soil show Tc(VII) reduction in the presence of anaerobicmetal- and sulphate-reducing bacteria. However, nitrifying bac-teria, growing under aerobic/anaerobic reducing conditions, do notaffect the reduction of Tc(VII) to Tc(IV). Organic matter is adetermining factor in the immobilization of Tc(IV) in soil becauseof its ability to form strong complex (Abdelouas et al., 2005). Thefate of Tc(VII) introduced into organic-rich environments wasstudied using X-ray absorption fine structure spectroscopy (Maeset al., 2004). The authors reported the formation of small Tc(IV)polymer oxides, which might aggregate into larger units that couldinteract with dissolved/immobile humic substances, suggesting apotentially new model for metal-humate complexation in organic-rich environments. A study of the effects of progressive anoxia onthe solubility of Tc in sediments has demonstrated that theremoval of Tc(VII) from water to estuarine sediments is condi-tioned by reduction of Tc(VII) to Tc(IV) during the developmentof microbial anoxia, and coincides with Fe(III) reduction in thepresence of sulphate-reducing bacteria (Burke et al., 2005). Thereduction of Tc(VII) by dissimilation in microorganisms underneutral, acidic and alkaline conditions was extensively studied(Lyalikova and Khizhnyak, 1996; Lloyd et al., 1997, 1999, 2000;Wildung et al., 2000). The biotransformation of redox sensitiveradionuclides is the complex process, achieved by the directenzymatic interaction at the metal/microorganism interface orby abiotic reaction of the radionuclide with microbial reductionproducts (Mohapatra et al., 2010; Law et al., 2010).
In summary, different studies have reported that the potentialeither for migration or retention of 99Tc at contaminated sitesdepends on the geochemical behaviour of 99Tc determined by its
chemical speciation, the physical characteristics of the environ-ment (mineralogy, pH, complexing agents, redox conditions), andon the microbial activity. However, most of the studies have beenrelated to the investigation of technetium migration/retention inanaerobic reducing environments. The behaviour of technetiumunder aerobic conditions remains poorly characterized, regardlessof a broad range of direct and indirect processes formed in theaerobic environment, where technetium can undergo chemicallyor biologically controlled redox changes.
The goal of the present study was to investigate the behaviourof 99Tc(VII) in the heterogeneous water system containing well-defined amounts of hematite and magnetite with emphasis ontransformation of the oxidation state through microbial mediatedprocesses under aerobic conditions. A set of experiments wasperformed to determine the factors (mineralogy, pH, incubationtime, microorganisms) that influenced TcO�
4 (aq) sorption ontoiron oxides, and to explore the capability of microorganisms toparticipate in metal/radionuclide mobility processes, paying spe-cial attention to their involvement in redox processes.
2. Experimental
2.1. Materials and methods
Batch experiments were conducted in the laboratory to exam-ine the selective adsorption characteristics of 99Tc(VII) underambient conditions. In each experiment, synthetic powdered ironoxides (hematite and wustite/magnetite) and/or microorganismsisolated from groundwater and waterlogged soils were used.
The powders of synthetic iron oxides were purchased from AOREACHIM (Russia); since the compositions of iron oxides were notclear, Mössbauer spectroscopy was used for verification of the ironoxides.
The particle sizes of iron oxides were determined by SEMHelios NanoLab 650 (FEI).
The solutions and iron oxide (Fe2O3 and Fe3O4/FeO) powdersuspensions were prepared in deionised water (TKA LAB MICROsystem, conductivity 0.055 μS/cm; TOCo10 ppb).
Technetium-99 was purchased from National Physical Labora-tory (UK) in 0.1 mol dm�3 NH4OH aqueous solution. For the batchexperiments TcO�
4 (3.5�10�6 mol dm�3) solutions were pre-pared by diluting aliquots of the stock solution.
All batch experiments were performed in triplicate. Allreagents used were of analytical grade.
2.1.1. Microorganisms2.1.1.1. Isolation. Cultures of bacteria and fungi were isolated fromsamples of groundwater and waterlogged soil. For isolation of themicroorganisms, groundwater samples were serially diluted by fourorders of magnitude. Aliquots (0.1 mL) of the undiluted water andeach dilution were spread onto appropriate media. Undiluted watersamples were used for isolation of actinomycetes. The isolation offungi and bacteria was performed using diluted and undilutedwater samples. Ten grams of each soil sample were mixed with95 mL of sterile (0.1 M, pH 7.0) phosphate buffer in 150 mL conicalflasks and were shaken for 10 min at 150 rpm. Soil suspensionswere decanted and serial dilutions by six orders of magnitude wereprepared. Different plating media were used for the isolation ofmicroorganisms. Samples for the fungi isolation were plated eitheronto the 2% malt extract agar (MEA, Liofilchem, Italy) or a potatodextrose agar (PDA, Liofilchem, Italy) in Petri dishes (9 cmdiameter); bacteria were isolated on a nutrient agar (NA,Liofilchem, Italy) and actinomycetes on a Starch-casein agar (SCA,Liofilchem, Italy). Nystatin (50 μg mL�1) was added to avoid fungalcontamination in the actinomycetes cultures; chloramphenicol
R. Druteikienė et al. / Applied Radiation and Isotopes 89 (2014) 85–9486

Author's personal copy
(60 μg mL�1) was added to avoid bacterial growth in the fungicultures. Fungi, bacteria and actinomycetes were incubated at2572 1C, 3272 1C and 2872 1C, respectively, in the dark. Afterincubation, well separated colonies were counted (morphologicallydifferent colonies) and the percentage of occurrence was analysedto determine potential strains for further application in theinvestigation of 99Tc relationships with soil and watermicroorganisms. Dominant strains were maintained on nutrientagar slants at 4 1C.
2.1.1.2. Identification. Fungi from the primary cultures were sub-cultured onto a malt extract agar (MEA, Liofilchem, Italy) and apotato dextrose agar (PDA, Liofilchem, Italy), a corn meal agar (CMA,Liofilchem, Italy), and Czapek-Dox agar (CDA; Liofilchem, Italy) andwere incubated for up to 21 days at 2571 1C. Morphologicallydifferent colonies of bacteria and actinobacteria were sub-culturedon the fresh NA and SCA plates, respectively. Bacteria were identifiedand attributed to the species level on the basis of their physiologicalcharacteristics, using the Gram staining reaction and otherbiochemical tests (Holt and Bergey, 1994). Morphology of the fungiwas analysed both by observing colonies under a stereomicroscope(Nikon SMZ 745Te; �10 magnification) and by observing preparedslides under a light microscope (Motic B1 Series; �400 and �1000magnification). The fungi were identified based on the morphologicalcharacteristics and taxonomic keys described by Ellis and Ellis (1997),Pitt (1985), Kiffer and Morelet (2000), Domsh et al. (2007).
Velvety isolates of the actinomycetes were examined to deter-mine their Gram-positive morphology and formation of longfilamentous cells. Their microstructure was observed by the coverslip method; mycelium structure, colour, as well as arrangement ofconidiophores and arthrospores on the mycelium, were observedmicroscopically (Motic B1 series) using an oil immersion (�100)objective. Visually determined structures were compared withthose described by Claus and Berkeley (1986), to confirm speciesof the isolates.
After identification, separate strains of the microbial species,sub-cultured on the nutrient media slants, were maintained in theCulture Collection of the Nature Research Centre (Vilnius, Lithuania).
The Gram-positive actinobacterium (actinomycete) (Streptomy-cete coelicolor strain DPA-Tc3), three Gram-negative bacteria (Bacil-lus mycoides DPB-Tc2, Rhodococcus rubber DPB-Tc4, Pseudomonas
aeruginosa DPK-TcX), and three fungi (Aspergillus niger Tiegh. DPF-Tc7, Penicillium simplicissimum (Oudem.) Thom DPF-Tc6, Spicaria sp.DPB-Tc) dominating in the groundwater as well as the Gram-negative bacterium Arthrobacter globiformis strain DPB-Tc11, gram-positive bacterium Cellulomonas cellulans DPB-8, and fungus Fusar-ium oxysporum Schltdl. strain DPF-Tc3 dominating in the water-logged soil were applied for the investigation of Tc behaviour in thepresence of iron oxides. In addition to the dominance in naturalhabitats, specific microbial strains were chosen for further investi-gation based on their dependence on oxygen, their ability to reducenitrate, form H2S, produce organic acid and to tolerate different pHvalues, as observed during the earlier investigation (Lukšienė et al.,2012). Chosen microbial strains were sub-cultured for 4–7 days at25–32 1C in different media (depending on the microorganism):bacteria in a nutrient broth (NB; Liofilchem, Italy), actinomycete – ina starch casein medium (SC, without agar), and fungi – in maltextract broth (MEB; Liofilchem, Italy). After incubation by aeratingon a shaker at 140 rpm, biomass of the microorganisms wasseparated by centrifugation (bacteria and actinomycetes) or byfiltration (fungi), and then it was washed with deionised sterilewater.
2.1.2. Iron oxidesPowders of synthetic iron oxides (AO REACHIM, Russia) were
used for 99Tc(VII) sorption experiments. Based on SEM analysis,particle sizes of wustite/magnetite were in the range of 2�10�6–
2�10�5 m and those of hematite ranged between 2�10�6 and1�10�5 m. The SEM images of both minerals are presented inFig. 1.
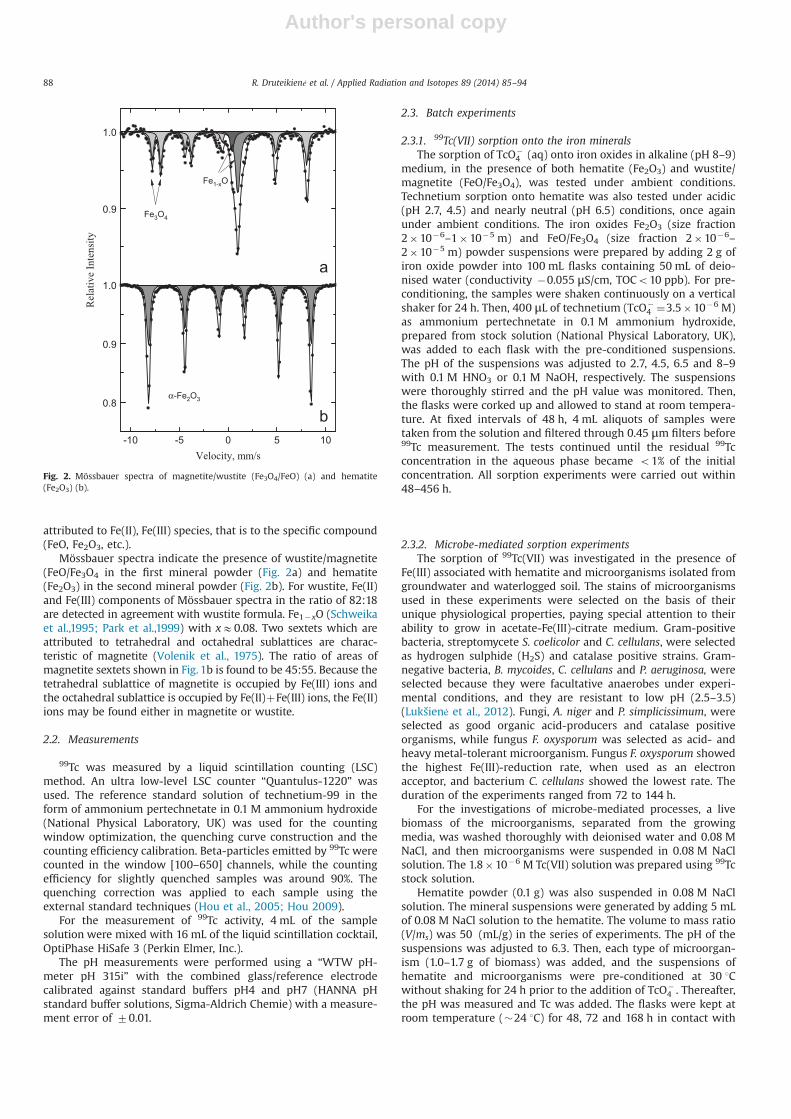
The compositions of two powdered iron oxides were verifiedusing Mössbauer spectroscopy. Mössbauer spectra of solid samples(Fig. 2) were measured in transmission geometry using 57Co (Rh)source and a Mössbauer spectrometer (Wissenschaftliche Elektro-nik GmbH) at room temperature. For the measurements, ironoxides powders were put into the plastic capsule (with thicknessof E30 mg/cm2).The spectra were fitted using WinNormos soft-ware decomposing to sextets, doublet and singlet subspectraand determining hyperfine parameters: the hyperfine field B,the isomer shift δ, the quadrupole splitting Δ and a relative area.According to the hyperfine parameters the area of a spectrum is
Fig. 1. Scanning electron microscope images of hematite (Fe2O3) ((a) and (b)); and wustite/magnetite (FeO/Fe3O4) ((c) and (d)).
R. Druteikienė et al. / Applied Radiation and Isotopes 89 (2014) 85–94 87
Author's personal copy
attributed to Fe(II), Fe(III) species, that is to the specific compound(FeO, Fe2O3, etc.).
Mössbauer spectra indicate the presence of wustite/magnetite(FeO/Fe3O4 in the first mineral powder (Fig. 2a) and hematite(Fe2O3) in the second mineral powder (Fig. 2b). For wustite, Fe(II)and Fe(III) components of Mössbauer spectra in the ratio of 82:18are detected in agreement with wustite formula. Fe1�xO (Schweikaet al.,1995; Park et al.,1999) with xE0.08. Two sextets which areattributed to tetrahedral and octahedral sublattices are charac-teristic of magnetite (Volenik et al., 1975). The ratio of areas ofmagnetite sextets shown in Fig. 1b is found to be 45:55. Because thetetrahedral sublattice of magnetite is occupied by Fe(III) ions andthe octahedral sublattice is occupied by Fe(II)þFe(III) ions, the Fe(II)ions may be found either in magnetite or wustite.
2.2. Measurements
99Tc was measured by a liquid scintillation counting (LSC)method. An ultra low-level LSC counter “Quantulus-1220” wasused. The reference standard solution of technetium-99 in theform of ammonium pertechnetate in 0.1 M ammonium hydroxide(National Physical Laboratory, UK) was used for the countingwindow optimization, the quenching curve construction and thecounting efficiency calibration. Beta-particles emitted by 99Tc werecounted in the window [100–650] channels, while the countingefficiency for slightly quenched samples was around 90%. Thequenching correction was applied to each sample using theexternal standard techniques (Hou et al., 2005; Hou 2009).
For the measurement of 99Tc activity, 4 mL of the samplesolution were mixed with 16 mL of the liquid scintillation cocktail,OptiPhase HiSafe 3 (Perkin Elmer, Inc.).
The pH measurements were performed using a “WTW pH-meter pH 315i” with the combined glass/reference electrodecalibrated against standard buffers pH4 and pH7 (HANNA pHstandard buffer solutions, Sigma-Aldrich Chemie) with a measure-ment error of 70.01.
2.3. Batch experiments
2.3.1. 99Tc(VII) sorption onto the iron mineralsThe sorption of TcO�
4 (aq) onto iron oxides in alkaline (pH 8–9)medium, in the presence of both hematite (Fe2O3) and wustite/magnetite (FeO/Fe3O4), was tested under ambient conditions.Technetium sorption onto hematite was also tested under acidic(pH 2.7, 4.5) and nearly neutral (pH 6.5) conditions, once againunder ambient conditions. The iron oxides Fe2O3 (size fraction2�10�6–1�10�5 m) and FeO/Fe3O4 (size fraction 2�10�6–
2�10�5 m) powder suspensions were prepared by adding 2 g ofiron oxide powder into 100 mL flasks containing 50 mL of deio-nised water (conductivity �0.055 μS/cm, TOCo10 ppb). For pre-conditioning, the samples were shaken continuously on a verticalshaker for 24 h. Then, 400 μL of technetium (TcO�
4 ¼3.5�10�6 M)as ammonium pertechnetate in 0.1 M ammonium hydroxide,prepared from stock solution (National Physical Laboratory, UK),was added to each flask with the pre-conditioned suspensions.The pH of the suspensions was adjusted to 2.7, 4.5, 6.5 and 8–9with 0.1 M HNO3 or 0.1 M NaOH, respectively. The suspensionswere thoroughly stirred and the pH value was monitored. Then,the flasks were corked up and allowed to stand at room tempera-ture. At fixed intervals of 48 h, 4 mL aliquots of samples weretaken from the solution and filtered through 0.45 μm filters before99Tc measurement. The tests continued until the residual 99Tcconcentration in the aqueous phase became o1% of the initialconcentration. All sorption experiments were carried out within48–456 h.
2.3.2. Microbe-mediated sorption experimentsThe sorption of 99Tc(VII) was investigated in the presence of
Fe(III) associated with hematite and microorganisms isolated fromgroundwater and waterlogged soil. The stains of microorganismsused in these experiments were selected on the basis of theirunique physiological properties, paying special attention to theirability to grow in acetate-Fe(III)-citrate medium. Gram-positivebacteria, streptomycete S. coelicolor and C. cellulans, were selectedas hydrogen sulphide (H2S) and catalase positive strains. Gram-negative bacteria, B. mycoides, C. cellulans and P. aeruginosa, wereselected because they were facultative anaerobes under experi-mental conditions, and they are resistant to low pH (2.5–3.5)(Lukšienė et al., 2012). Fungi, A. niger and P. simplicissimum, wereselected as good organic acid-producers and catalase positiveorganisms, while fungus F. oxysporum was selected as acid- andheavy metal-tolerant microorganism. Fungus F. oxysporum showedthe highest Fe(III)-reduction rate, when used as an electronacceptor, and bacterium C. cellulans showed the lowest rate. Theduration of the experiments ranged from 72 to 144 h.
For the investigations of microbe-mediated processes, a livebiomass of the microorganisms, separated from the growingmedia, was washed thoroughly with deionised water and 0.08 MNaCl, and then microorganisms were suspended in 0.08 M NaClsolution. The 1.8�10�6 M Tc(VII) solution was prepared using 99Tcstock solution.
Hematite powder (0.1 g) was also suspended in 0.08 M NaClsolution. The mineral suspensions were generated by adding 5 mLof 0.08 M NaCl solution to the hematite. The volume to mass ratio(V/ms) was 50 (mL/g) in the series of experiments. The pH of thesuspensions was adjusted to 6.3. Then, each type of microorgan-ism (1.0–1.7 g of biomass) was added, and the suspensions ofhematite and microorganisms were pre-conditioned at 30 1Cwithout shaking for 24 h prior to the addition of TcO�
4 . Thereafter,the pH was measured and Tc was added. The flasks were kept atroom temperature (�24 1C) for 48, 72 and 168 h in contact with
-10 -5 0 5 10
0.8
0.9
1.0
Velocity, mm/s
α-Fe2O3
0.9
1.0
Rel
ativ
e In
tens
ity
Fe3O4
Fe1-xO
Fig. 2. Mössbauer spectra of magnetite/wustite (Fe3O4/FeO) (a) and hematite(Fe2O3) (b).
R. Druteikienė et al. / Applied Radiation and Isotopes 89 (2014) 85–9488
Author's personal copy
the air. Prior to Tc analysis, the suspensions were filtered through0.45 μm filters to remove bacteria cells.
To test the ability of the microorganisms alone to accumulatetechnetium (KD), the batch experiment with selected microorgan-isms were carried out in the absence of hematite. The micro-organism biomass was separated from the growing media, washedwith deionised water and then microorganisms were suspended in0.08 M NaCl solution. An appropriate amount of TcO�
4 was addedto obtain the 1.8�10�6 M 99Tc(VII) solution. The pH of thesolutions were adjusted to 7. After 48 h, the solution was filteredand 99Tc was measured in an aliquot of solution by LSC.
2.4. KD calculation
Distribution coefficients (KD) were calculated to describe theadsorption behaviour of Tc in the experimental systems in thepresence of iron oxides and microorganisms. The KD is defined as
KD ¼ ðAi–AeÞ=Ae � V=m;
where Ai is the initial activity of 99Tc in solution, and Ae is theresidual activity of 99Tc in solution after the sorption; V is thevolume of the solution and m is the mass of the solid (Vejsada,2006).
3. Results
3.1. Sorption of Tc(VII) onto Fe-bearing minerals
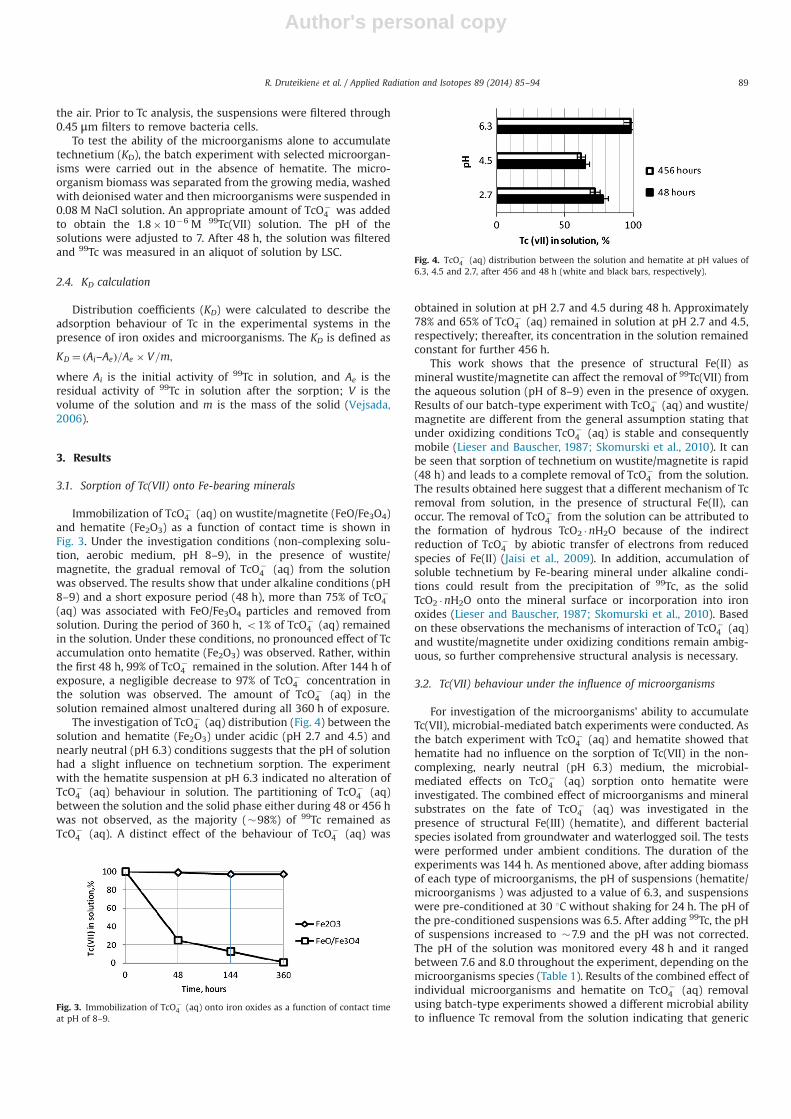
Immobilization of TcO�4 (aq) on wustite/magnetite (FeO/Fe3O4)
and hematite (Fe2O3) as a function of contact time is shown inFig. 3. Under the investigation conditions (non-complexing solu-tion, aerobic medium, pH 8–9), in the presence of wustite/magnetite, the gradual removal of TcO�
4 (aq) from the solutionwas observed. The results show that under alkaline conditions (pH8–9) and a short exposure period (48 h), more than 75% of TcO�
4(aq) was associated with FeO/Fe3O4 particles and removed fromsolution. During the period of 360 h, o1% of TcO�
4 (aq) remainedin the solution. Under these conditions, no pronounced effect of Tcaccumulation onto hematite (Fe2O3) was observed. Rather, withinthe first 48 h, 99% of TcO�
4 remained in the solution. After 144 h ofexposure, a negligible decrease to 97% of TcO�
4 concentration inthe solution was observed. The amount of TcO�
4 (aq) in thesolution remained almost unaltered during all 360 h of exposure.
The investigation of TcO�4 (aq) distribution (Fig. 4) between the
solution and hematite (Fe2O3) under acidic (pH 2.7 and 4.5) andnearly neutral (pH 6.3) conditions suggests that the pH of solutionhad a slight influence on technetium sorption. The experimentwith the hematite suspension at pH 6.3 indicated no alteration ofTcO�
4 (aq) behaviour in solution. The partitioning of TcO�4 (aq)
between the solution and the solid phase either during 48 or 456 hwas not observed, as the majority (�98%) of 99Tc remained asTcO�
4 (aq). A distinct effect of the behaviour of TcO�4 (aq) was
obtained in solution at pH 2.7 and 4.5 during 48 h. Approximately78% and 65% of TcO�
4 (aq) remained in solution at pH 2.7 and 4.5,respectively; thereafter, its concentration in the solution remainedconstant for further 456 h.
This work shows that the presence of structural Fe(II) asmineral wustite/magnetite can affect the removal of 99Tc(VII) fromthe aqueous solution (pH of 8–9) even in the presence of oxygen.Results of our batch-type experiment with TcO�
4 (aq) and wustite/magnetite are different from the general assumption stating thatunder oxidizing conditions TcO�
4 (aq) is stable and consequentlymobile (Lieser and Bauscher, 1987; Skomurski et al., 2010). It canbe seen that sorption of technetium on wustite/magnetite is rapid(48 h) and leads to a complete removal of TcO�
4 from the solution.The results obtained here suggest that a different mechanism of Tcremoval from solution, in the presence of structural Fe(II), canoccur. The removal of TcO�
4 from the solution can be attributed tothe formation of hydrous TcO2 �nH2O because of the indirectreduction of TcO�
4 by abiotic transfer of electrons from reducedspecies of Fe(II) (Jaisi et al., 2009). In addition, accumulation ofsoluble technetium by Fe-bearing mineral under alkaline condi-tions could result from the precipitation of 99Tc, as the solidTcO2 �nH2O onto the mineral surface or incorporation into ironoxides (Lieser and Bauscher, 1987; Skomurski et al., 2010). Basedon these observations the mechanisms of interaction of TcO�
4 (aq)and wustite/magnetite under oxidizing conditions remain ambig-uous, so further comprehensive structural analysis is necessary.
3.2. Tc(VII) behaviour under the influence of microorganisms
For investigation of the microorganisms' ability to accumulateTc(VII), microbial-mediated batch experiments were conducted. Asthe batch experiment with TcO�
4 (aq) and hematite showed thathematite had no influence on the sorption of Tc(VII) in the non-complexing, nearly neutral (pH 6.3) medium, the microbial-mediated effects on TcO�
4 (aq) sorption onto hematite wereinvestigated. The combined effect of microorganisms and mineralsubstrates on the fate of TcO�
4 (aq) was investigated in thepresence of structural Fe(III) (hematite), and different bacterialspecies isolated from groundwater and waterlogged soil. The testswere performed under ambient conditions. The duration of theexperiments was 144 h. As mentioned above, after adding biomassof each type of microorganisms, the pH of suspensions (hematite/microorganisms ) was adjusted to a value of 6.3, and suspensionswere pre-conditioned at 30 1C without shaking for 24 h. The pH ofthe pre-conditioned suspensions was 6.5. After adding 99Tc, the pHof suspensions increased to �7.9 and the pH was not corrected.The pH of the solution was monitored every 48 h and it rangedbetween 7.6 and 8.0 throughout the experiment, depending on themicroorganisms species (Table 1). Results of the combined effect ofindividual microorganisms and hematite on TcO�
4 (aq) removalusing batch-type experiments showed a different microbial abilityto influence Tc removal from the solution indicating that generic
Fig. 3. Immobilization of TcO�4 (aq) onto iron oxides as a function of contact time
at pH of 8–9.
Fig. 4. TcO�4 (aq) distribution between the solution and hematite at pH values of
6.3, 4.5 and 2.7, after 456 and 48 h (white and black bars, respectively).
R. Druteikienė et al. / Applied Radiation and Isotopes 89 (2014) 85–94 89
Author's personal copy
peculiarities of the microorganisms are very important in theseprocesses. Although two of the strains – bacterium C. cellulans andfungus F. oxysporum – were obtained from the same waterloggedsoil, the first of them did not influence the immobilisation of Tc(VII) in hematite, while the latter, on the contrary, increased Tcremoval (up to 85%) from the experimental solution comparedwith that in the system without microorganisms (Table 1).
Differences in the removal of TcO�4 (aq) from solution by
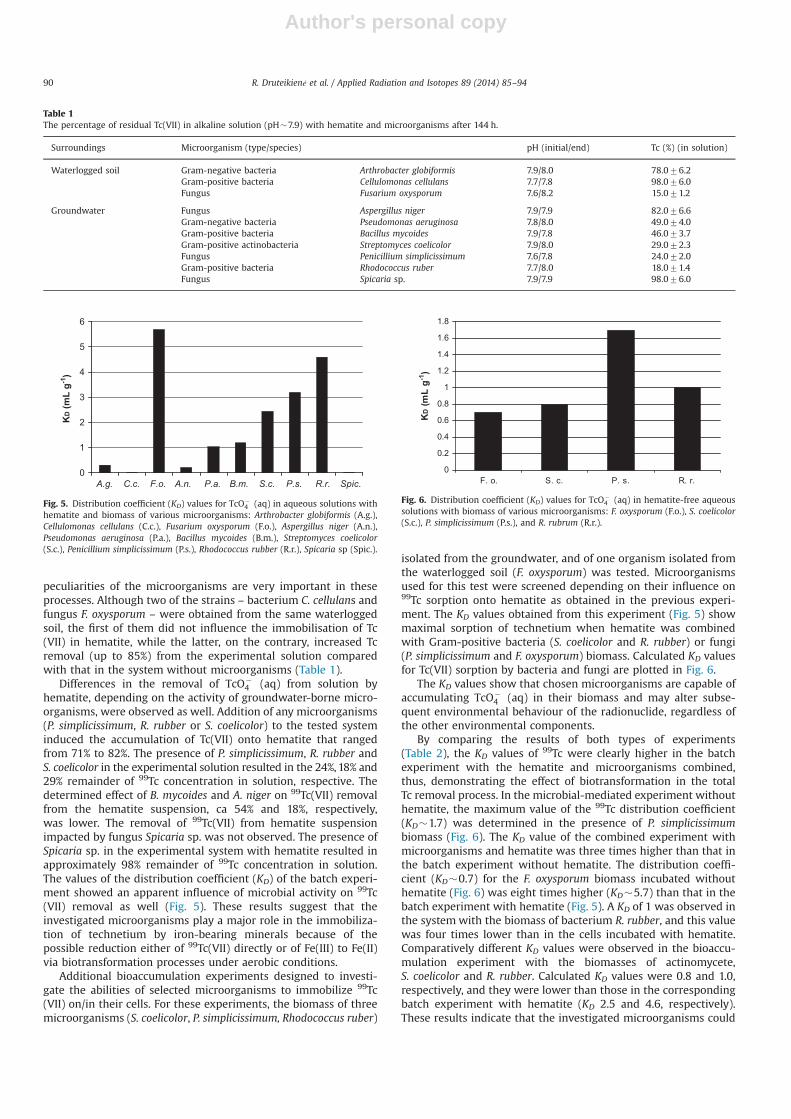
hematite, depending on the activity of groundwater-borne micro-organisms, were observed as well. Addition of any microorganisms(P. simplicissimum, R. rubber or S. coelicolor) to the tested systeminduced the accumulation of Tc(VII) onto hematite that rangedfrom 71% to 82%. The presence of P. simplicissimum, R. rubber andS. coelicolor in the experimental solution resulted in the 24%, 18% and29% remainder of 99Tc concentration in solution, respective. Thedetermined effect of B. mycoides and A. niger on 99Tc(VII) removalfrom the hematite suspension, ca 54% and 18%, respectively,was lower. The removal of 99Tc(VII) from hematite suspensionimpacted by fungus Spicaria sp. was not observed. The presence ofSpicaria sp. in the experimental system with hematite resulted inapproximately 98% remainder of 99Tc concentration in solution.The values of the distribution coefficient (KD) of the batch experi-ment showed an apparent influence of microbial activity on 99Tc(VII) removal as well (Fig. 5). These results suggest that theinvestigated microorganisms play a major role in the immobiliza-tion of technetium by iron-bearing minerals because of thepossible reduction either of 99Tc(VII) directly or of Fe(III) to Fe(II)via biotransformation processes under aerobic conditions.
Additional bioaccumulation experiments designed to investi-gate the abilities of selected microorganisms to immobilize 99Tc(VII) on/in their cells. For these experiments, the biomass of threemicroorganisms (S. coelicolor, P. simplicissimum, Rhodococcus ruber)
isolated from the groundwater, and of one organism isolated fromthe waterlogged soil (F. oxysporum) was tested. Microorganismsused for this test were screened depending on their influence on99Tc sorption onto hematite as obtained in the previous experi-ment. The KD values obtained from this experiment (Fig. 5) showmaximal sorption of technetium when hematite was combinedwith Gram-positive bacteria (S. coelicolor and R. rubber) or fungi(P. simplicissimum and F. oxysporum) biomass. Calculated KD valuesfor Tc(VII) sorption by bacteria and fungi are plotted in Fig. 6.
The KD values show that chosen microorganisms are capable ofaccumulating TcO�
4 (aq) in their biomass and may alter subse-quent environmental behaviour of the radionuclide, regardless ofthe other environmental components.
By comparing the results of both types of experiments(Table 2), the KD values of 99Tc were clearly higher in the batchexperiment with the hematite and microorganisms combined,thus, demonstrating the effect of biotransformation in the totalTc removal process. In the microbial-mediated experiment withouthematite, the maximum value of the 99Tc distribution coefficient(KD�1.7) was determined in the presence of P. simplicissimumbiomass (Fig. 6). The KD value of the combined experiment withmicroorganisms and hematite was three times higher than that inthe batch experiment without hematite. The distribution coeffi-cient (KD�0.7) for the F. oxysporum biomass incubated withouthematite (Fig. 6) was eight times higher (KD�5.7) than that in thebatch experiment with hematite (Fig. 5). A KD of 1 was observed inthe system with the biomass of bacterium R. rubber, and this valuewas four times lower than in the cells incubated with hematite.Comparatively different KD values were observed in the bioaccu-mulation experiment with the biomasses of actinomycete,S. coelicolor and R. rubber. Calculated KD values were 0.8 and 1.0,respectively, and they were lower than those in the correspondingbatch experiment with hematite (KD 2.5 and 4.6, respectively).These results indicate that the investigated microorganisms could
Table 1The percentage of residual Tc(VII) in alkaline solution (pH�7.9) with hematite and microorganisms after 144 h.
Surroundings Microorganism (type/species) pH (initial/end) Tc (%) (in solution)
Waterlogged soil Gram-negative bacteria Arthrobacter globiformis 7.9/8.0 78.076.2Gram-positive bacteria Cellulomonas cellulans 7.7/7.8 98.076.0Fungus Fusarium oxysporum 7.6/8.2 15.071.2
Groundwater Fungus Aspergillus niger 7.9/7.9 82.076.6Gram-negative bacteria Pseudomonas aeruginosa 7.8/8.0 49.074.0Gram-positive bacteria Bacillus mycoides 7.9/7.8 46.073.7Gram-positive actinobacteria Streptomyces coelicolor 7.9/8.0 29.072.3Fungus Penicillium simplicissimum 7.6/7.8 24.072.0Gram-positive bacteria Rhodococcus ruber 7.7/8.0 18.071.4Fungus Spicaria sp. 7.9/7.9 98.076.0
0
1
2
3
4
5
6
A.g. C.c. F.o. A.n. P.a. B.m. S.c. P.s. R.r. Spic.
KD (m
L g-1
)
Fig. 5. Distribution coefficient (KD) values for TcO�4 (aq) in aqueous solutions with
hematite and biomass of various microorganisms: Arthrobacter globiformis (A.g.),Cellulomonas cellulans (C.c.), Fusarium oxysporum (F.o.), Aspergillus niger (A.n.),Pseudomonas aeruginosa (P.a.), Bacillus mycoides (B.m.), Streptomyces coelicolor(S.c.), Penicillium simplicissimum (P.s.), Rhodococcus rubber (R.r.), Spicaria sp (Spic.).
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
F. o. S. c. P. s. R. r.
KD (m
L g-1
)
Fig. 6. Distribution coefficient (KD) values for TcO�4 (aq) in hematite-free aqueous
solutions with biomass of various microorganisms: F. oxysporum (F.o.), S. coelicolor(S.c.), P. simplicissimum (P.s.), and R. rubrum (R.r.).
R. Druteikienė et al. / Applied Radiation and Isotopes 89 (2014) 85–9490

Author's personal copy
stimulate the processes of technetium removal from solution inthe presence of Fe(III)-containing mineral surfaces.
4. Discussion
Many studies have shown that Fe(II), when in the sorbed ormineral state can be a strong reductant for 99Tc(VII) under anoxic/reducing conditions (Cui and Eriksen, 1996; Peretyazhko et al., 2009;Jaisi et al., 2009). At present, it is known that abiotic reduction ofsoluble TcO�
4 by both Fe(II)aq and Fe(II)ads can bring about theformation of relatively insoluble, amorphous TcO2 �nH2O at pH above3 (Lieser and Bauscher, 1987). Furthermore, TcO�
4 undergoes stage-by-stage one-, two- or three-electron reductions process leading tothe formation of different aqueous species (e.g., TcO2þ , TcO (OH)þ ,TcO2(s), TcO4) in non-complexing aqueous solutions under reducingconditions (Heller-Grossman et al., 1981; Warwick et al., 2007). Onthe other hand, the structural incorporation of Tc species into ironoxides is possible (Peretyazhko et al., 2009; Skomurski et al., 2010;Um et al., 2012). The origin of Tc(IV) species is possible in oxidizingenvironments with high pH values in the presence of a γ radiationsource and specific organic compounds. Lukens et al. (2002) showedthat Tc can exist as some soluble Tc(IV) phase in Hanford tank wastesunder these conditions.
Our research shows that ferrous iron Fe(II) associated with ironoxide wustite/magnetite strongly affects the behaviour of 99Tc(VII)in solution under aerobic/oxidizing conditions. Various pathwaysfor Tc removal from solution under the investigated conditions arepossible. The removal of TcO�
4 from solution can be attributed tothe formation of hydrous TcO2 because of the indirect reduction ofTc(VII) by abiotic transfer of electrons from reduced species ofFe(II). On the other hand, rapid accumulation of TcO�
4 by thisFe-bearing mineral could result from the precipitation of Tc ontothe wustite/magnetite surface. Taking into consideration the non-complexing medium of the experimental system, and accessibilityof O2, we assumed that Fe(II) subsequently reduced pertechnetate(TcO�
4 ) to TcO2þ which eventually co-precipitated with theFe(OH)3(s) formed as a consequence of the oxidation of Fe(II) toFe(III) under these experimental conditions.
However, a conclusive evaluation of TcO�4 (aq) interaction with
the Fe(II)-bearing minerals could be done after further analysis ofpost-reaction solids.
Experiments with TcO�4 (aq) and the Fe(III)-bearing mineral
hematite showed limited to no Tc removal from solution underaerobic/oxidizing conditions. This result is in agreement withliterature data stating that the affinity of TcO�
4 for Fe-bearingminerals is weak under most conditions (Ambe et al., 1996;Liu and Fan, 2005). The quantum-mechanical method used bySkomurski et al. (2010) to evaluate mechanisms for possiblestructural incorporation of Tc into the hematite structure suggeststhat the incorporation of TcO�
4 in the hematite lattice is not
feasible because of the energetic inadequacy and structural dis-tortion required of the coordination environment.
An investigation of TcO�4 (aq) interaction with hematite under
the influence of microorganisms has shown that microorganismsplay a major role in Fe(III) reactivity toward Tc(VII). The biotrans-formation of radionuclides achieved either by reduction of radio-nuclides by the microorganisms and their indirect abioticreduction by various reactive forms of iron or other reductantslikely produced by the microorganisms. Many microorganisms areable to enzymatically reduce radionuclides for use as the energysource (Mohapatra et al., 2010). As it is known, bacteria exhibitdifferent metabolic types (Holt and Bergey, 1994). Bacterial meta-bolism is classified on the basis of three major criteria: the kind ofenergy used for growth, the source of carbon, and the electrondonors used for growth (Xu, 2006). In addition, microbial abilitiesdepend not only on the species but also on the strain and thehabitat from which it was isolated. An obvious difference in theability of bacterial strains, isolated from the waterlogged soil andthe groundwater, to remove TcO�
4 (aq) from solution was deter-mined. Water-borne bacteria were significantly (with statisticalsignificance Po0.002) more effective in TcO�
4 (aq) binding withthe biomass than soil-borne bacteria. However, the differencesbetween two soil-borne bacteria, and among the four water-bornebacteria were also determined. These findings confirm thatbacterial activity in the investigated systems depends on theenvironmental conditions, from which bacterium was isolated, aswell as bacterium taxonomy. This research showed that in thepresence of Gram-negative bacteria P. aeruginosa, approximately51% of TcO�
4 (aq) was removed from the experimental solutionand sorbed onto hematite. P. aeruginosa is classified as an aerobicorganism and is considered by many researchers to be a facultativeanaerobe, as it is well adapted to proliferate under conditions ofpartial or total oxygen depletion (Palmer et al., 2007). P. aeruginosatends to form biofilms, which are complex bacterial communitiesthat adhere to a variety of surfaces. Biofilms can lead to retentionof dissolved heavy metals at sorption sites of the extra cellularpolymeric substance, cell cytoplasm, cell walls and cell mem-branes (McLean et al., 1996; Pollmann et al., 2006). Krawczyk-Baersch et al. (2008) showed that the presence of aqueous 238U(VI)in the biofilm microenvironment could lead to an increase ofrespiratory activity in the most metabolically active biofilm zone,and that oxygen consumption may induce 238U reduction and leadto the removal of 238U as 238U(IV) solids from solution.
In the present investigation a biofilm formation of glass flaskswas observed throughout the incubation period of P. aeruginosa,B. mycoides and S. coelicolor cultures.
Investigations into the ability of hematite to accumulate TcO�4
(aq) in the presence of water-borne eubacteria showed thatB. mycoides, R. rubber and actinobacteria, S. coelicolor induced�54%, 82%, 71% removal of TcO�
4 (aq) from solution, respectively.The results suggest that the biofilm formation may play a role in
Table 2Summary of microorganism-mediated accumulation (KD) experiment (with/without hematite).
Surroundings Microorganism (type/species) KD (Tc/Fe2O3/biomass) KD (Tc/biomass)
Waterlogged soil Gram-negative bacteria A. globiformis 0.3 –a
Gram-positive bacteria C. cellulans 0.02 –
Fungus F. oxysporum 5.7 0.7
Groundwater Fungus A. niger 0.2 –
Gram-negative bacteria P. aeruginosa 1.0 –
Gram-positive bacteria B. mycoides 1.2 –
Gram-positive actinobacteria S. coelicolor 2.5 0.8Fungus P. simplicissimum 3.2 1.7Gram-positive bacteria Rhodococcus ruber 4.6 1.0Fungus Spicaria sp. 0.02 –
a Not measured for these systems.
R. Druteikienė et al. / Applied Radiation and Isotopes 89 (2014) 85–94 91

Author's personal copy
this process. Eubacteria are present in most habitats and grow insoil, water, and also in radioactive waste (Fredrickson et al., 2004).Gram-positive B. mycoides is an obligate aerobe, meaning that itscells can undergo morphological changes when starved of nutri-ents. For instance, bacterial cells can migrate to each other andbehave as aggregates (Kaiser, 2004), or form biofilms on surfaces(Donlan, 2002). In these circumstances, bacteria perform separatetasks of a multicellular community.
Differences in the ability of tested fungi to participate in TcO�4
(aq) immobilisation in the presence of hematite were determined.The bacterium, C. cellulans, and fungus Spicaria sp. practically hadno effect on the sorption of TcO�
4 (aq) onto hematite under theaerobic experimental conditions, since approximately only 2% of99Tc was removed from the solution. Significantly lower amountsof TcO�
4 (aq) (from 15% to 29%; Po0.01) remained in solutionswhen the biomass of bacterium, R. rubber, actinobacterium, S.coelicolor, or each of two fungi (F. oxysporum, and P. simplicissi-mum) was added. TcO�
4 (aq) was removed from solution at similarrates, with concomitant sorption of 15% to 29% of TcO�
4 (aq) fromsolution in 144 h. Almost half (46–49%) of the initial TcO�
4 (aq)amount was removed from solution by both Gram-positive bac-terium, B. mycoides, and Gram-negative bacterium, P. aeruginosa.These findings show a tenuous ability of C. cellulanas and Spicariasp. to bind the TcO�
4 (aq) with their biomass. We suppose, that thefungi F. oxysporum and P. simplicissimum, as well as Gram-positivebacterium, R. rubber, and actinomycete, S. coelicolor, were able tostimulate interaction between TcO�
4 (aq) and hematite via reduc-tion of TcO�
4 (aq) or its accumulation in the biomass of micro-organisms. In comparison, only 12% of the TcO�
4 (aq) was removedfrom solution when A. niger was added. Gram-negative bacterium,A. globiformis, was slightly more effective; approximately 22% ofTcO�
4 (aq) remained in biomass.These hematite/microorganism batch experiments show that
the removal of TcO�4 from solution was facilitated by the activity of
microorganisms. One mechanisms is biological reduction of struc-tural Fe(III) to Fe(II), which may occur under these experimentalconditions. Then, the reduction of TcO�
4 (aq) by Fe(II) is expected.However, further comprehensive structural and chemical analysesof the reaction products are necessary to obtain the knowledge ofTc removal mechanisms.
The ability of a great variety of common bacteria (Bacillaceae,Pseudomonadaceae), as well as anaerobic, nitrogen-fixing bacteria,to reduce structural Fe(III) to soluble Fe(II) has been observed(Balashova and Zavarzin, 1980; Johnson and McGinness, 1991;Lovley, 1991; Lloyd and Macaskie, 1996; Lloyd et al., 1997, 1999;Wildung et al., 2000; Martin, 2005). Biological reduction of iron byfungi was shown by several authors (Ottow and von Klopotek,1969; Jalal et al., 1987; Leong and Winkelmann, 1998; De Luca andWood, 2000; Kosman, 2003). Woolfolk and Whiteley (1962)showed that the cell-free extract of Gram-negative bacterium,Vellionella atypica, reduced U(VI) along with a variety of othermetals. Although the reduction of Tc(VII) has been demonstratedfor a range of organisms (Lloyd and Macaskie, 1996; Lloyd et al.,1997, 1999; Wildung et al., 2000), under anaerobic conditions,very scarce information about Tc(VII) reduction under aerobicconditions is available (Istok et al., 2004; Burke et al., 2005). Inaddition, Martin (2005) showed that the dissolution of hematiteoccurred by ligand-promoted, proton-promoted, reductive andsynergistic pathways. Reductants also rapidly accelerate iron oxidedissolution; examples of reductants are ascorbic acid, oxalic acid,hydrogen sulphate, and phenols (Stone and Morgan, 1984; Stone,1987a, 1987b; Martin, 2005). Fungus A. niger, which was used inthis study, produces oxalic acid which could serve as Tc(VII)reductant, or citric acid which could be involved in the reactionswith the hematite (Sayer and Gadd, 1997). Fe(II)-oxalato aqueouscomplex is a strong reductant, which rapidly reduces iron oxide
through reductive dissolution. Oxalate secretion by fungi offersmany advantages for their growth and colonization of substrates.Gram-positive bacteria produce succinic acid as well (Song andLee, 2006). Three of the four Gram-positive bacteria used in thepresent investigation could be assigned to a group of active metal-reductants. Fungi Fusarium sp. (Foster, 1949) and P. simplicissimum(Gallmetzer et al., 2002) are known to produce and secretesuccinic acid. P. simplicissimum and F. oxysporum were effectivein the 99Tc reduction and sorption processes (Gallmetzer et al.,2002). R. ruber synthesises red pigment may also participate in theenzymatic reduction of Tc (Lloyd et al., 1997). Many fungi (Müllerand Raumond, 1984; Gadd and White, 1990) and bacteria (Sayyedet al., 2005) secrete specific hydroxamate siderophores whenshort of iron. In the present investigation, two fungal isolates,F. oxysporum and P. simplicissimum, showed effectiveness in TcO�
4(aq) removing from solution under aerobic conditions. Furtherinvestigation is needed to confirm if these fungi are direct TcO�
4(aq) or Fe(III) reducers, or if they serve as intermediates betweenTcO�
4 (aq) and Fe(III) in the sorption process.
5. Conclusions
The presence of structural Fe(II) associated with wustite/mag-netite in non-complexing aqueous solution (pH 8–9) under aerobicconditions can affect the removal of TcO�
4 (aq) from solution, thuslimiting its mobility in oxidizing environments. The results showthat under alkaline conditions (pH 8–9), after a short exposureperiod (48 h), more than 75% of TcO�
4 (aq) was associated withFeO/Fe3O4 particles and removed from solution. Structural Fe(III)associated with hematite was not able to immobilize TcO�
4 (aq)under similar alkaline conditions. However, low pH (2.7–4.5)increases TcO�
4 (aq) retention on hematite, which suggests thatthe process is dependent on pH.
Retention of TcO�4 (aq) onto hematite is achieved because of
the presence of specific microorganisms isolated fromwaterloggedsoil and groundwater. B. mycoides, R. rubber, actinobacteria,S. coelicolor and fungi F. oxysporum and P. simplicissimum inducedthe removal of TcO�
4 (aq) from solution in the presence of hematite.This research showed that in the presence of Gram-negative bacteriaP. aeruginosa, approximately 51% of TcO�
4 (aq) was removed fromthe experimental solution. The bacterium, C. cellulans, and fungusSpicaria sp. practically had no effect on sorption of TcO�
4 ontohematite under the aerobic experimental conditions.
An obvious difference between the bacterial strains, isolated fromthe waterlogged soil and the groundwater, in their ability toinfluence TcO�
4 (aq) behaviour in solution was determined. Water-borne bacteria were more effective in TcO�
4 (aq) immobilizationwith the biomass than soil-borne bacteria. The differences betweentwo soil-borne bacteria, and among four water-borne bacteria werealso determined. These findings have confirmed that the bacterialactivity in the investigated processes really depends on the environ-mental conditions as well as on the bacterium taxonomy.
Thus, we cannot yet use a classical explanation of the resultsobtained, because a genetic assignation to the genus must beinvestigated in future.
Acknowledgments
The research leading to these results has received funding fromthe European Atomic Energy Community Seventh FrameworkProgram [FP7/2007–2013] under grant agreement No. 212287,Collaborative Project Recosy and from the Lithuanian Agency forScience, Innovation and Technology (Grant no. 31V-6).
R. Druteikienė et al. / Applied Radiation and Isotopes 89 (2014) 85–9492

Author's personal copy
References
Abdelouas, A., Grambow, B., Fattahi, M., Andres, Y., Leclerc-Cessac, E., 2005.Microbial reduction of 99Tc in organic matter-rich soils. Sci. Total Environ336, 255–268.
Ambe, S., Iwamoto, M., Maeda, H., Ambe, F., 1996. Multitracer study on adsorptionof metal ions on α-Fe2O3. J. Radioanal. Nucl. Chem. 205 (2), 269–275.
Balashova, V.V., Zavarzin, G.A., 1980. Anaerobic reduction of ferric iron by hydrogenbacteria. Microbiology 48, 635–639.
Begg, J.D.C., Burke, I.T., Morris, K., 2007. The behaviour of technetium duringmicrobial reduction in amended soils from Dounreay. UK. Sci. Total Environ373, 297–304.
Borsh, T., Kretzschmar, R., Kappler, A., Van Cappellen, P., Ginder-Vogel, M., Voegelin,A., Campbell, K., 2010. Biogeochemical redox processes and their impact oncontaminant dynamics. Environ. Sci. Technol. 44, 15–23.
Buechele, A.C., McKeown, D.A., Lukens, W.W., Shuh, D.K., Pegg, I.L., 2012. Tc and Rebehavior in borosilicate waste glass vapor hydration tests II. J. Nucl. Mater 429,159–165.
Burke, I.T., Boothman, C., Lloyd, J.R., Livens, F.R., Morris, K., 2005. Effects onprogressive anoxia on the solubility of technetium in sediments. Environ. Sci.Technol. 39, 4109–4119.
Burke, I.T., Boothman, C., Lloyd, J.R., Mortimer, R.J.G., Livens, F.R., Charnok, J.M.,Mcbeth, J.M., Mortimer, R.J.G., Morris, K., 2006. Reoxidation behaviour oftechnetium, iron, and sulfur in estuarine sediments. Environ. Sci. Technol. 40,3529–3635.
Choppin, G.R., 2005. Actinide science: fundamental and environmental aspects.J. Nucl. Radiochem. Sci. 6 (1), 1–5.
Claus, D., Berkeley, C.K., 1986. Bergeys's Manual of Systematic Bacteriology. SneathPHA, William and Wilkins, Baltimore, pp. 1105-1139.
Cui, D., Eriksen, T.E., 1996. Reduction of pertechnetate in solution by ferrous iron insolution: influence of sorbed and precipitate Fe(II). Environ. Sci. Technol 30 (7),2259–2262.
De Luca, N.G., Wood, P.M., 2000. Iron uptake by fungi: contrasted mechanisms withinternal or external reduction. Adv. Microb. Physiol. 43, 39–74.
Donlan, R., 2002. Biofilms: microbial life on surfaces. Emerg. Infect. Dis. 8, 881–890.Domsh, K.H., Gams, W, Anderson, T-H., 2007. Compendium of Soil Fungi, second ed.
IHW-Verlag, EchingEllis, M.B., Ellis, J.P., 1997. Microfungi on Land Plants. An Identification Handbook.
The Richmond Publishing Co. Ltd, LondonFrederickson, J.K., Zachara, J.M., Kennedy, D.W., Kukadapu, R.K., McKinley, J.P.,
Heald, S.M., Liu, C., Plymale, A.E., 2004. Reduction of TcO4� by sediment
associated Fe(II). Geochim. Cosmochim. Acta 68, 3171–3187.Fredrickson, J.K., Zachara, J.M., Balkwill, D.L., Kennedy, D., Li, S.-M., Konstandarithes,
H., Daly, M., Romine, M., Brockman, F., 2004. Geomicrobiology of high-levelnuclear waste-contaminated vadose sediments at the Hanford site, Washingtonstate. Appl. Environ. Microbiol. 70 (7), 4230–5241.
Foster, J.W., 1949. Chemical Activities of Fungi. Academic Press, New YorkGadd, G.M., White, C., 1990. Biosorption of radionuclides by yeast and fungal
biomass. J. Chem. Technol. Biotechnol. 49, 331–343.Gallmetzer, M., Meraner, J., Burgstaller, W., 2002. Succinate synthesis and excretion
by Penicillium simplicissimum under aerobic and anaerobic conditions. Micro-biol. Lett 210, 221–225.
Heller-Grossman, L., Abrashkin, S., Shafferman, A., Davis, M., Taube, R., 1981.Tc-99m generators. Physico-chemical factors in the radiolytic reduction ofpertechnetate. Appl. Radiat. Isot 32, 501–506.
Holt, J.C., Bergey, D.H., 1994. Bergey's Manual of Determinative Bacteriology, 9th ed.Williams & Wilkins, Baltimore
Hou, X., Ostergaard, L.F., Nielsen, S.P., 2005. Determination of 63Ni and 55Fe innuclear waste samples using radiochemical separation and liquid scintillationcounting. Anal. Chim. Acta 535, 297–307.
Hou, X, 2009. Liquid Scintillation Counting for the Determination of Beta Emitters—Principle and Application. ⟨http://www.nks.org/download/seminar/2009_radworkshop/NKS-B_RadWorkshop_8.pdf⟩.
Istok, J.D., Senko, J.M., Krumholz, L.R., Watson, D., Bogle, M.A., Peacock, A., Chang, Y.J.,2004. In situ bioreduction of technetium and uranium in nitrate-contaminatedaquifer. Environ. Sci. Technol. 38, 468–475.
Jaisi, D.P., Dong, H., Plymale, A.E., Fredrickson, J.K., Zachara, J.M., Heald, S., Liu, C.,2009. Reduction and long-term immobilization of technetium by Fe(II) asso-ciated with clay mineral nontronite. Chem. Geol. 264, 127–138.
Jalal, M.A., Love, S.K., van der Helm, D., 1987. Siderophore mediated iron(III) uptakein Gliocladium virens. 2. Role of ferric mono- and dihydroxamates as irontransport agents. J. Inorg. Biochem. 29 (4), 259–267.
Johnson, D.B., McGinness, S., 1991. Ferric iron reduction by acidophilic hetero-trophic bacteria. Appl. Environ. Microbiol. 57, 207–211.
Kaiser, D., 2004. Signaling in mycobacteria. Ann. Rev. Microbiol. 58, 75–98.Keith-Roach, M.J., Morris, K., Dahlgaard, H., 2003. An investigation into technetium
binding in sediments. Mar. Chem. 81, 149–162.Kiffer, E., Morelet, M., 2000. The Deuteromycetes. Mitosporic Fungi. Classification
and Generic Keys. Science Publishers Inc, USAKosman, D.J., 2003. Molecular mechanism of iron uptake in fungi. Molec. Microbiol
47 (5), 1185–1197.Krawczyk-Baersch, E., Grossmann, K., Arnold, T., Hofmann, S., Wobus, A., 2008.
Influence of uranium (VI) on the metabolic activity of stable multispeciesbiofilms studied by oxygen microsensors and fluorescence microscopy. Geo-chim. Cosmochim. Acta 72, 5251–5265.
Law, G.T.W., Geissler, A., Lloyd, J.R., Livens, F.R., Boothman, C., Begg, J.D.C., Denecke,M.A., Rothe, J., Dardenne, K., Burke, I.T., Charnock, J.M., Morris, K., 2010.Geomicrobial redox cycling of the transuranic element neptunium. Environ.Sci. Technol. 44, 8924–8929.
Lieser, K.H, Bauscher, C., 1987. Technetium in the hydrosphere and in the geosphere: I.Chemistry of technetium and iron in natural waters and influence of the redoxpotential on the sorption of technetium. Radiochim. Acta 42, 205–213.
Liljenzin, J-O., 2002. Behavior of radionuclides in the environment. In: Chopin, G.,Liljenzin, J-O., Rydgerg, J. (Eds.), The Radiochemistry and Nuclear Chemistry,third ed. Butterworth-Heinemann, Oxford, p. 642
Liu, D.J., Fan, X.H., 2005. Adsorption behaviour of 99Tc on Fe, Fe2O3 and Fe3O4.J. Radioanal. Nucl. Chem. 264 (3), 691–698.
Leong, S.A., Winkelmann, G., 1998. Molecular biology of iron transport in fungi. In:Sigel, A., Sigel, H. (Eds.), Iron Transport and Storage in Microorganisms, Plants,and Animals. Marcel Dekker, New York, pp. 147–186
Lloyd, J.R., Macaskie, L.E., 1996. A novel phosphorimager-based technique formonitoring the microbial reduction of technetium. Appl. Environ. Microbiol.62, 578–582.
Lloyd, J.R., Ridley, J., Khizhnyak, T., Lyalikova, N.N., Macaskie, L.E., 1999. Reduction oftechnetium by Desulfovibrio desulfuricans: biocatalyst characterization and usein a flow through bioreactors. Appl. Environ. Microbiol. 65, 2691–2696.
Lloyd, J.R., Sole, V.A., Van Praagh, C.V.G., Lovely, D.R., 2000. Direct and Fe(II)-mediated reduction of technetium by the Fe(III)-reducting bacteria. Appl.Environ. Microbiol. 66, 3743–3749.
Lloyd, J.R., Cole, J.A., Macaskie, L.E., 1997. Reduction and removal of heptavalenttechnetium from solution by Escherichia coli. J. Bacteriol 179, 2014–2021.
Lukens, W.W.J., Bucher, J.J., Edelstein, N.M., Shuh, D.K., 2001. Radiolysis of TcO4� in
alkaline, nitrate solutions: reduction by NO32� . J. Phys. Chem. A 105, 9611–9615.
Lukens, W.W.J., Bucher, J.J., Edelstein, N.M., Shuh, D.K., 2002. Products of pertech-netate radiolysis in highly alkaline solution: structure of TcO2 � xH2O. Environ.Sci. Technol 36, 1124–1129.
Lukšienė, B., Druteikienė, R., Pečiulytė, D., Baltrūnas, D., Remeikis, V., Paškevičius, A.,2012. Effect of microorganisms on the plutonium oxidation states. Appl. Radiat.Isot 70, 442–449.
Lyalikova, N.N., Khizhnyak, T., 1996. Reduction of heptavalent technetium byacidophilic bacteria of the genus Thiobactillus. Microbiologia 65, 468–473.
Lovley, D.R., 1991. Dissimilatory Fe(III) and Mn(IV) reduction. Microbiol. Rev. 55,259–287.
McLean, R.J.C., Fortin, D., Brown, D.A., 1996. Microbial metal-binding mechanismsand their relation to nuclear waste disposal. Can. J. Microbiol. 42, 392–400.
Maes, A., Geraedts, K., Bruggeman, C., Vanchuysen, J., Rossberg, A., Henning, C.,2004. Evidence for the interaction of technetium colloids with humic sub-stances by X-ray absorption spectroscopy. Environ. Sci. Technol. 38, 2044–2051.
Marshall, M.J., Dohnalkova, A., Kennedy, D.W., Plymale, A.E., Thomas, S.H., Loffler, F.E., Sandorf, R.A., Zachara, J.M., Fredrickson, J., Beliaev, A.S., 2009. Electrondonor-dependent radionuclide reduction and nanoparticle formation by Anae-romyxobacter dehalogenans strain 2CP-C. Environ. Microbiol 11 (2), 534–543.
Maset, E.R., Sidhu, S.H., Fisher, A., Heydon, A., Worsfold, P.J., Cartwright, A.J., Keith-Roach, M.J., 2006. Effect of organic co-contaminants on technetium andrhenium speciation and solubility under reducing conditions. Environ. Sci.Technol. 40, 5472–5477.
Martin, S.T., 2005. Precipitation and dissolution of iron and manganese oxides. In:Grassian, V.H. (Ed.), Environmental Catalysis. CRC Press, Boca Raton, pp. 61–81
Mohapatra, B.R., Dinardo, O., Gould, W.D., Koren, D.W., 2010. Biochemical andgenomic facets on the dissimilatory reduction of radionuclides by microorgan-isms—a review. Mineral. Eng. 23, 591–599.
Müller, G., Raumond, K.N., 1984. Specificity and mechanism of ferrioxamine-mediated iron transfer in Streptomyces pilosus. J. Bacteriol. 160 (1), 304–312.
Ottow, J.C.G., von Klopotek, A., 1969. Anzymatic reduction of iron oxide by fungi.Appl. Microbiol 18 (1), 41–43.
Palmer, K.L., Brown, S.A., Whiteley, M., 2007. Membrane-bound nitrate reductase irrequired for anaerobic growth in cystic fibrosis sputum. J. Bacteriol. 189 (12),4449–4455.
Park, J-Ch., Kim, D., Lee, Ch-S., Kim, D-K., 1999. A new synthetic route to wüstitebull. Korean Chem. Soc 20 (9), 1005–1009.
Peretyazhko, T., Zachara, J.M., Heald, S.M., Jeon, B.H., Kukadapu, R.K., Liu, C., Moore,D., Resch, C.T., 2009. Heterogeneous reduction of Tc(VII) by Fe(II) at the solid–water interface. Geochim. Cosmochim. Acta 72, 1521–1539.
Peretyazhko, T.S., Zachara, J.M., Kukadapu, R.K., Heald, S.M., Kutnyakov, I.V., Resch,C.T., Arey, B.W., Wang, C.M., Kovarik, L., Phillips, J.L., Moore, D.A., 2012.Pertechnetate (TcO4
�) reduction by reactive ferrous iron in naturally anoxic,redox transition zone sediments from tha Hanford Site, USA. Geochim.Cosmochim. Acta 92, 48–66.
Pitt, J.I., 1985. A Laboratory Guide to Common Penicillium Species. CommonwealthScientific and Industrial Research organization. Division of Food ResearchLaboratory, NSW, Australia
Pollmann, K., Raff, J., Merroun, M., Fahmy, K., Selenska-Pobell, S., 2006. Metalbinding by bacteria from uranium mining waste piles and its technologicalapplications. Biotechnol. Adv. 24, 58–68.
Sayyed, R.Z., Badgujar, M.D., Sonawane, H.M., Mhaske, M.M., Chincholkar, S.B.,2005. Production of microbial iron chelators (siderophores) by fluorescentPseudomonads. Ind. J. Biotechnol. 4 (784-490).
Sayer, J.A., Gadd, G.M., 1997. Solubilization and transformation of insolubleinorganic metal compounds to insoluble metal oxalates by Aspergillus niger.Mycol. Res 101, 653–661.
R. Druteikienė et al. / Applied Radiation and Isotopes 89 (2014) 85–94 93

Author's personal copy
Skomurski, F.N., Rosso, K.M., Krupka, K.M., McGrail, B.P., 2010. Techne-tium incorporation into hematite (α-Fe2O3). Environ. Sci. Technol. 44,5855–5861.
Shi, K., Hou, X., Roos, P., Wu, W., 2012. Determination of technetium-99 inenvironmental samples: a review. Anal. Chim. Acta 709, 1–20.
Song, H., Lee, S.Y., 2006. Production of succinic acid by bacterial fermentation. Enz.Microb. Technol 39, 352–361.
Standring, W.J.F., Oughton, D.H.., Salbu, B., 2002. Potential remobilization of 137Cs,60Co, 99Tc, and 90Sr from contaminated Mayak sediments in river and estuaryenvironments. Environ. Sci. Technol. 36, 2330–2337.
Stone, A.T., Morgan, J.J., 1984. Reduction and dissolution of manganese (III) andmanganese (IV) oxides by organics: 2. Survey of the reactivity of organics.Environ. Sci. Technol. 18, 617–624.
Stone, A.T., 1987a. Reductive dissolution of manganese (III/IV) oxides by substitutephenols. Environ. Sci. Technol. 21, 979–988.
Stone, A.T., 1987b. Microbial metabolites and the reductive dissolution ofmanganese oxides: oxalate and pyruvate. Geochim. Cosmochim. Acta 51,919–925.
Schweika, W., Hoser, A., Martin, M., Carlsson, A.E., 1995. Defect structure of ferrousFe1�xO. Phys. Rev. B: Condens. Matter 51, 15771–15788.
Tagami, K., Uchida, S., 1996. Microbial role in the immobilization of technecium insoil under waterlogged conditions. Chemosphere 33, 217–225.
Um, W., Chang, H., Icenhover, J.P., Lukens, W.W., Serene, R.J., Qafoku, N., Kukkadapu,R.K., Westsik , J.H., 2012. Iron oxide waste form for stabilizing 99Tc. J. Nucl.Mater. 429 (1–3), 201–209.
Vejsada, J., 2006. The uncertainties associated with the application of batchtechnique for distribution coefficients determination—a case study of cesiumadsorption on four different bentonites. Appl. Radiat. Isot 64, 1538–1548.
Volenik, K., Seberini, M., Neid, J., 1975. A Mössbauer anc X-ray diffraction study ofnonstoichiometry in magnetite. Czech. J. Phys. B 25, 1063–1071.
Wildung, R.E., Gorby, Y.A., Krupka, K.M., Hess, N.J., Li, S.W., Plymale, A.E., McKinley,J.P., Fredrickson, J.K., 2000. Effect of electron donor and solution chemistry onproducts of dissimilatory reduction of technetium by Shewanella putrefaciens.Appl. Environ. Microbiol. 66, 2451–2460.
Warwick, P., Aldridge, S., Evans, N., Vines, S., 2007. The solubility of technetium (IV)at high pH. Radiochim. Acta 95, 709–716.
Wharton, M.J., Atkins, B., Charnock, J.M., Livens, F.R., Pattrick, R.A.D., Collison, D.,2000. An X-ray absorption spectroscopy study of the co-precipitation of Tc andRe with mackinawite (FeS). Appl. Geochem. 15, 347–354.
Woolfolk, C.A., Whiteley, H.R., 1962. Reduction of inorganic compounds withmolecular hydrogen by Micrococcus lactilyticus. J. Bacteriol. 84, 647–668.
Xu, J., 2006. Microbial ecology in the age of genomics and metagenomics: concepts,tools, and recent advances. Mol. Ecol. 15 (7), 1713–1731.
Zachara, J.M., Heald, S.M., Jeon, B.H., Kukkadapu, R.K., Liu, C., McKinley, J.P.,Dohnalkova, A.C., Moore, D.A., 2007. Reduction of pertechnetate [Tc(VII)] byaqueous Fe(II) and the nature of solid phase redox products. Geochim.Cosmochim. Acta 71, 2137–2157.
R. Druteikienė et al. / Applied Radiation and Isotopes 89 (2014) 85–9494




















