
Barrosso Ruiz et al., 2014. Boquete de Zafarraya cave: A Neanderthal site in southern Iberia. In: R....
10
SOUTHERN MEDITERRANEAN COAST, GUADALQUIVIR RIVER AND BETIC INTRAMONTANE BASINS 463 habitat and the procurement area. Inhabitants of Cueva del Angel made an extraordinary use of small tools as flakes, also typologically diverse si- descrapers were common. In the future, further dating of levels with lithic industry on fluvial ter- races as well as Acheulean levels in caves, will help to ascertain the relationship between both con- texts. At the moment, the reference sites that can be compared with Cueva del Angel are Galería in Atapuerca (Carbonell et al., 1999), Cova del Bo- lomor (Fernández Peris, 2007) and Galeria Pesada (Marks et al., 2012). Regarding the chronology, the rejection of dates used so far prevent the establishment of a precise chronological framework of the Cueva del Angel, but there is no doubt that the analysis cur- rently being done will age considerably the date of 120 ky years available so far. Boquete de Zafarraya cave: A Neanderthal site in southern Iberia Cecilio Barroso *, Miguel Caparrós **, Deborah Barsky ***, Anne Marie Moigne ****, Antonio Monclova * Introduction Boquete de Zafarraya cave (hereinafter C.B.Z.) is in the Alcaucín municipality, Málaga Province (southern Spain). It is 1020 m asl, fac- ing S-E at the foot of a cliff, with a steep slope below it. The coordinates are: 36º 56` 58``N – 4º 7`40`` W. This cave in Sierra de Alhama overlooks an extremely irregular landscape of limestone rang- es and very narrow valleys boxed in between them. Scarcely a few dozen metres away, howev- er, the landscape opens onto the mountain pass, Boquete de Zafarraya, which at 900 m asl con- nects the Mediterranean coast with inland Anda- lucia (Fig. 1). This landscape mosaic is completed by the 22 km² Zafarraya polje, formed from an intra-montane depression of karst and tectonic origin filled with basically Quaternary sediments, powerfully contrasting with its surroundings by its extensive subhorizontal landscape (Barroso, C. et al., 2006). The prehistoric Boquete de Zafarraya cave site was discovered by a member of this team (C.B.R.) in 1979. During the first stage of the archaeological excavations (1981-1983), led by C. Barroso and F. Medina Lara, we dis- covered levels containing Mousterian indus- try, a rich and varied quaternary wildlife and two quaternary Neanderthal fossils (Zaf. 1) and (Zaf. 2) (Barroso et al., 1983; Barroso et al., 1984.). The second archaeological stage at the site continued from 1990 to 1994. B.Z.C. is the last vestige of a cave which col- lapsed and whose morphology has been heav- ily affected by powerful slope erosion. At the mouth, a large arch-shaped porch rises 30 me- tres, measuring approx. 10 metres at the base. On the north wall there is a narrow 22 m long gallery, barely 2 m. in the widest sections, devel- oped from a fault plane. The cave is in stratified pisolitic and pseudo-oolitic limestone. This gal- lery is the source of the quaternary deposit. The infill was preserved by the carbonation processes inside which cemented much of it, preventing the intense erosion which took place through- out the area, heavily affected by the steep slope. Neanderthal fossils B.Z.C. is perhaps one of Iberia’s most impor- tant sites of Homo neanderthalensis fossils (Bar- * Fundación Instituto de Investigación de Prehistoria y Evolución Humana. Plaza del Coso 21, 14900 Lucena (Córdoba, Es- paña) email: [email protected] ** Département de Préhistorie, Muséum National d’Histoire Naturelle, 75013 París, France *** Departement d’història i història de l’Art. Universitat Rovira i Virgili. **** Muséum National d’Histoire Naturelle. Centre Européen de Recherches Préhistoriques de Tautavel.
Transcript of Barrosso Ruiz et al., 2014. Boquete de Zafarraya cave: A Neanderthal site in southern Iberia. In: R....

SOUTHERN MEDITERRANEAN COAST, GUADALQUIVIR RIVER AND BETIC INTRAMONTANE BASINS 463
habitat and the procurement area. Inhabitants of Cueva del Angel made an extraordinary use of small tools as flakes, also typologically diverse si-descrapers were common. In the future, further dating of levels with lithic industry on fluvial ter-races as well as Acheulean levels in caves, will help to ascertain the relationship between both con-texts. At the moment, the reference sites that can be compared with Cueva del Angel are Galería in
Atapuerca (Carbonell et al., 1999), Cova del Bo-lomor (Fernández Peris, 2007) and Galeria Pesada (Marks et al., 2012).
Regarding the chronology, the rejection of dates used so far prevent the establishment of a precise chronological framework of the Cueva del Angel, but there is no doubt that the analysis cur-rently being done will age considerably the date of 120 ky years available so far.
Boquete de Zafarraya cave:
A Neanderthal site in southern Iberia
Cecilio Barroso *, Miguel Caparrós **, Deborah Barsky ***, Anne Marie
Moigne ****, Antonio Monclova *
Introduction
Boquete de Zafarraya cave (hereinafter C.B.Z.) is in the Alcaucín municipality, Málaga Province (southern Spain). It is 1020 m asl, fac-ing S-E at the foot of a cliff, with a steep slope below it. The coordinates are: 36º 56` 58``N – 4º 7`40`` W.
This cave in Sierra de Alhama overlooks an extremely irregular landscape of limestone rang-es and very narrow valleys boxed in between them. Scarcely a few dozen metres away, howev-er, the landscape opens onto the mountain pass, Boquete de Zafarraya, which at 900 m asl con-nects the Mediterranean coast with inland Anda-lucia (Fig. 1). This landscape mosaic is completed by the 22 km² Zafarraya polje, formed from an intra-montane depression of karst and tectonic origin filled with basically Quaternary sediments, powerfully contrasting with its surroundings by its extensive subhorizontal landscape (Barroso, C. et al., 2006).
The prehistoric Boquete de Zafarraya cave site was discovered by a member of this team (C.B.R.) in 1979. During the first stage of the archaeological excavations (1981-1983),
led by C. Barroso and F. Medina Lara, we dis-covered levels containing Mousterian indus-try, a rich and varied quaternary wildlife and two quaternary Neanderthal fossils (Zaf. 1) and (Zaf. 2) (Barroso et al., 1983; Barroso et al., 1984.). The second archaeological stage at the site continued from 1990 to 1994. B.Z.C. is the last vestige of a cave which col-lapsed and whose morphology has been heav-ily affected by powerful slope erosion. At the mouth, a large arch-shaped porch rises 30 me-tres, measuring approx. 10 metres at the base. On the north wall there is a narrow 22 m long gallery, barely 2 m. in the widest sections, devel-oped from a fault plane. The cave is in stratified pisolitic and pseudo-oolitic limestone. This gal-lery is the source of the quaternary deposit. The infill was preserved by the carbonation processes inside which cemented much of it, preventing the intense erosion which took place through-out the area, heavily affected by the steep slope.
Neanderthal fossils
B.Z.C. is perhaps one of Iberia’s most impor-tant sites of Homo neanderthalensis fossils (Bar-
* Fundación Instituto de Investigación de Prehistoria y Evolución Humana. Plaza del Coso 21, 14900 Lucena (Córdoba, Es-paña) email: [email protected]
** Département de Préhistorie, Muséum National d’Histoire Naturelle, 75013 París, France*** Departement d’història i història de l’Art. Universitat Rovira i Virgili.**** Muséum National d’Histoire Naturelle. Centre Européen de Recherches Préhistoriques de Tautavel.

PLEISTOCENE AND HOLOCENE HUNTER-GATHERERS IN IBERIA AND THE GIBRALTAR STRAIT:
THE CURRENT ARCHAEOLOGICAL RECORD464
roso et al., 2006 b). There are at least 9 indi-viduals (2 children, aged 14 months to 12 years and 7 adults, four almost 20 years old and two aged 25 and 30, and one undetermined). There is a notably poor representation of infants and a heavy adult mortality. The most surprising feature, however, is the “significant” presence of Neanderthal bones (16 fossils) in a cave which was visited sporadically by these hominids, who spent very short periods there, computed as a few days each time at the most judging by spa-tial analysis of the lithic industry and the frag-ments of animal bones bearing obvious signs of anthropic manipulation. Anatomically, the fos-sils include:
Cranial elements: 2 mandibles (Zaf. 2 and Zaf.4-5-18) (Fig. 2) and 5 separate teeth (Zaf. 12 Zaf. 16, Zaf. 20, Zaf. 23 and Zaf. 24).
Mandible Zaf 2, discovered in 1983, was ini-tially attributed to a Homo sapiens neandertha-lensis male aged between 25 and 30. The jawbone
is practically intact, with 13 teeth preserved. This jawbone was studied in depth by Maria Antoi-nette de Lumley, who established the phyloge-netic position of Zafarraya Man in the group of so-called “classic” Neanderthals. Zafarraya retains some features described for the Ante-Neander-thals, particularly amongst those discovered in Banyoles and Montmaurin, although no features permit their connection to modern populations. Mandible Zaf. 4-5-18, discovered in 1990 and 1992, is broken into 3 pieces, with evidence of fire exposure, related to the presence of a Mous-terian hearth in the quadrat Q-18. All three frag-ments are from an adult mandible. The propor-tions and morphology are comparable to Zaf. 2.
Upper limbs: 1 scapula (Zaf. 6), 1 humerus (Zaf. 22).
The specimen discovered in 1990 is the lower half of the glenoid cavity of a right scapula. Its blackish surface evidences exposure to fire and, like other human remains, is associated with the
Figure 1. Neanderthal Capra pirinaica hunting scene in Boquete Zafarraya (author: Antonio Monclova Bohorquez).

SOUTHERN MEDITERRANEAN COAST, GUADALQUIVIR RIVER AND BETIC INTRAMONTANE BASINS 465
hearth in quadriculate Q-18. Comparison with the Krapina scapulae shows it is amongst the variations attributed to males.
Humerus Zaf. 22 was discovered in 1992 at the rear of the gallery on a disturbed superficial level, and thus lacks stratigraphic context. Mor-phological and metric analysis has attributed this fossil to the Neanderthal group, although certain parameters such as maximum length, minimum perimeter and poor robustness are also present in modern humans. Morphologically, the humer-us matches the Neanderthals from Cova Tossal de la Font and Grotte d’ Hortus. The proportions indicate that this was a fairly robust and une-quivocally female Neanderthal.
Lower limbs: 3 femurs (Zaf. 1-15 and 16), 1 tibia (Zaf. 27) 1 coxis (Zaf.17) and 1 foot phal-ange (Zaf. 3).
Femur Zaf. 1 was found during the 1982 digs. Only the upper half of a right femur was preserved, in which part of the upper epiphysis and the up-per half of the 225 mm long diaphysis can be ap-preciated. The femoral head and trochanters were broken. The initial study (see note) attributes this to a Neanderthal adult male aged 30-49 years. It has a relatively strong size ratios for a Neanderthal, measuring 159 cm to 162cm in height. The medial bone fragmentation and the large splinter extracted from the diaphysis are clearly anthropogenic. Met-ric and morphological analysis concludes that Zaf. 1 is distinct from Neanderthal femurs in the lack of a vertical incurvation. On the other hand, the cortical thickness, antero-posterior flatness of the shaft and the lack of a posterior pilaster are comparable to Neanderthals. The less pronounced muscular relief than other fossils suggests that these were less ro-bust individuals than the classic Neanderthals pop-ulations in southwest France and the Middle East.
Femurs Zaf. 15 and 26 (Fig.2).
Found heavily fragmented and burned in the Q-18 hearth during the 1990-1992 excavations. Zaf. 15 preserves the shaft of a left human femur. Both the anterior and posterior faces have been reconstructed from 11 fragments. Zaf. 26 retains part of the diaphysis of a right femur, recon-structed from 9 fragments. Its robustness is quite different from Zaf.1: Zaf. 15 and 26 have slender shafts and weaker anteroposterior and transver-sal diameters than Zaf.1. The major difference in strength between the three femurs could reflect
sexual dimorphism, with FAZ. 1 belonging to a slim male individual while the other two femurs are from two females.
Zaf. 27, a right human tibia (Fig. 2), consists of 10 burned fragments found in the Q-18 hearth. The lower half of the shaft and a fragment of the upper half have been preserved. It seems to be from a graciles individual, whose overall proportions sug-gest an adult female, with the diameter in the mid-section of the shaft close to the Ferrassie 2 individu-als, attributed to a female, and smaller than those of the Chapelle-aux-Saints tibia, attributed to a male.
The phalanx of the right foot (Zaf. 3), lo-cated during the 1983 dig, is incomplete, al-
Figure 2. Neanderthal fossils in Boquete de Zafarraya cave. From Left to right A: Mandible Zafarraya 2 (photo. Cecilio Barroso Ruíz), B: Burnt mandible 4-5-18, C: Burnt Femur Zafarraya 26, D: Burnt Tibia Zafarraya 27, E: Burnt femur Zafarraya 15 (B, C. D and E: photos. Rafael López Gómez.

PLEISTOCENE AND HOLOCENE HUNTER-GATHERERS IN IBERIA AND THE GIBRALTAR STRAIT:
THE CURRENT ARCHAEOLOGICAL RECORD466
though it may be attributed to a proximal phalanx of the second toe of the right foot. Its morphological and metric characteristics sug-gest that this may be Neanderthal phalanx. It belongs to an adult, as the overall dimensions of the head are comparable to the homologous Shanidar 8 phalanx, attributed to a female. The coxis (Zaf.17) was located during the 1992 dig in quadrat Q-18, on the edge of the pit that shaped the Mousterian hearth. It is well preserved and permits the examination of the symphysis face, the horizontal and descendant branches of the pubis. The break at the branches is old and covered with concretion. It is attrib-uted to a male individual estimated to have been 20-21 years old.
Thorax: 1 rib (Zaf. 19).
This human right rib appeared in the 1992 dig in the context of the Q-18 hearth. It is from a right C7, two-thirds of which are well preserved. It is characterized by its thickness, slight helical torsion and faces angled obliquely at the base, which probably gave the bottom of the rib cage an open form, contrary to modern men. It is at-tributed to an adult Neanderthal male.
Mousterian industry
A rich and stone tool industry has been col-lected from all seven distinct Archeostratigraph-ical Units (AU) attributed to the Mousterian (Barroso et al 2006 c.) The global assemblage comprises 813 items; mostly knapped in flint as well as some quartzose-sandstone. Lithic raw materials were collected from primary and sec-ondary sources not exceeding 12 km away. Out-crops situated some 11 km away provided the finest flint and were preferred to other nearby sources. The assemblage is well preserved overall but some flakes display post-depositional edge damage. Patina attributed to fire exposure is ob-served on only 17 items.
The lithic type distribution per-AU varies significantly according to occupation intensity and the extension of excavated surfaces but no noteworthy technological or typological differ-ences are detected within the sequence. Knap-ping waste (L> 2 cm) dominates the assemblage and the average flake/core ratio is 23. This core
deficiency is to be put into relation with the pau-city of small-sized flakes and flakes with residual cortex, all suggesting that much of the knapping and shaping occurred off-site.
Many of the 539 knapped items (flakes, blades, points) were obtained by Levallois production methods (194 pcs., Levallois technological in-dex excluding pseudo-Levallois points= 36%). No important variation in the Levallois index is observed within the sequence: the entire assem-blage is Levallois dominant. A ratio of 1 Leval-lois product for every 2 to 3 standard products is constant regardless of the raw material used. Levallois flakes are numerically prevalent (161 pcs.) compared to blades (25 pcs.) and points (8 pcs.).
Recurrent centripetal is the most common Levallois method attested from flakes and cores, although lineal, unipolar recurrent and bipolar methods have also been identified. A few sym-metrical preferential flakes reflect a final phase of recurrent centripetal Levallois schemes with dorsal surfaces bearing traces of their core’s pre-pared convex surface. The Levallois flakes often present prepared, facetted platforms (LevFI = 48,6, FI = 16,6).
Among the 23 cores unearthed at the site, only 4 are clearly of Levallois conception: 3 centrip-etal recurrent and 1 unipolar recurrent. A further three cores present convex extraction surfaces analogous to Levallois-type production strategies. Alternative core reduction modes appear non-standardized, mainly because the matrices are exhausted. These are uni, bi and multi-direction-al and polyhedral. Most cores are knapped from flint nodules rather than flakes and the assem-blage includes only three double ventral flakes and two cores on flakes.
Blades were mostly made from the finest flint and their overall index is quite low (BI = 8,1%). Elongated supports were rarely obtained by methods other than Levallois (NLevBI =5,5%, LevBI= 12,8%). Use of a soft percussion instru-ment is only sporadically recognized. The BDZ industry is further characterized by a high fre-quency of core-edge flakes (Fr.: éclat débordant) and pseudo-Levallois points, both typically ob-tained during different phases of Levallois and discoid core surface convexity maintenance.

SOUTHERN MEDITERRANEAN COAST, GUADALQUIVIR RIVER AND BETIC INTRAMONTANE BASINS 467
Nearly one quarter of the flakes > 2 cm are retouched into various tool types. While no se-lectivity of supports is evidenced, the Levallois typological index is to be considered elevated (41,4 %). The retouched toolkit includes: Mous-terian points, scrapers (42%), some notched tools (30 %), trimmed and truncated instruments, as well as some Upper Paleolithic types. Quina re-touch is rare and there are no handaxes and only one chopper-like tool.
Given the techno-typological features out-lined above, the stone industries from the Bo-quete de Zafarraya Mousterian levels are as-cribed to a Levallois dominant typical Mousterian.
Large animal vertebrates
The faunal remains found at the Zafarraya cave site (Barroso et al., 2006) provide informa-tion about various aspects of the Neanderthal lifestyle. First, the list of fauna matches a fresh episode in the middle of the last glaciation. The abundance of Cuon alpinus and Panthera pardus (Fig. 3) is unusual at European sites, permitting its comparison with caves in the Basque Coun-try. The vast majority of the sites where panther (245 remains, 10 individuals) or dhole are well represented have been dated in the Hengelo epi-sode. Ursus arctos (Fig 3), Crocuta crocuta, Felis silvestris, Lynx pardina and Vulpes vulpes form the classic range of Upper Pleistocene carni-vores. The herbivores do not point to a specific biochronology except for the apparently robust horse, attributed to Equus c. germanicus which became extinct 30,000 years ago. A substantial majority is mountain fauna, particularly goats (78% of the identified remains) and fallow deer, which are associated with their usual predators: panther, lynx and dhole. The frequent meso– to supra-Mediterranean environment has been de-scribed on the basis of palynological analysis as cold and very dry for a herbaceous steppe stra-tum with a few pines. The sedimentological se-quence dated between 49 and 38 ky corresponds to several alternations of more or less moist cli-mates with a homogeneous vegetation through-out the Mousterian occupation of the cave. The microfauna also describes a colder climate than today at 1000 m asl. The cave provided shelter for these large mammals on several occasions. The 46 archaeo-stratigraphic units defined from
the sedimentary sequence, along with evidence of humans and carnivores, clearly show an alter-nation in the occupations.
The remains of aurochs, deer, wild boar and horse indicate that hominids frequented the Za-farraya polje moorland where these animals were present. Some of the rodents and small birds also came from these moist wooded areas.
The faunal remains from the different levels suggest a concomitant presence between carni-vores and Neanderthals, with several possible scenarios. There may have been mutual periodic exclusions in which Neanderthals were replaced
Figure 3. Carnivore fossils in Boquete de Zafarraya cave. From left to right o A: Panthera Pardus mandible, B: Linx pardina ulna, C: Panthera pardus maxillary, D: Panthera Pardus mandible, Ursus arctos ulna (Photos: Rafael López Gómez).

PLEISTOCENE AND HOLOCENE HUNTER-GATHERERS IN IBERIA AND THE GIBRALTAR STRAIT:
THE CURRENT ARCHAEOLOGICAL RECORD468
by carnivores or vice-versa as the cave occupants. Another scenario is that these two types of preda-tors were not mutually exclusive and may have coexisted, or perhaps the presence of carnivores might even be a direct consequence of the hu-mans. The analysis of different interventions by panthers, lynx and dholes suggests a more intense involvement by the former, which is also consist-ent with the number of individuals at all levels. The presence of dholes on the upper levels of the infill is more sporadic and their remains show greater pressure by predators. Skeletal represen-tation of goats indicates that the carcasses were brought to the site, although the bias is not uni-form in all levels. The abundant remains of this species (2667 items in the stratigraphy) show all stages of impact by carnivores on virtually every part of the skeleton. However, carnivore marks are associated with evidence of anthropological activity. Evidence of carnivore activity is particu-larly notable on the bones of other carnivores such as dholes, but also on the remains of a hyena and also a bear discovered on the low levels of the in-fill, on the “levels containing panther”. Deer and wild boar were probably prey for large carnivores. Human activity is visible on many goat items. Like the large carnivores, Neanderthals frequented the cave when hunting goats in this area. The high nu-merical presence of juveniles indicates the hunting season, which ran from spring to summer. Goat butchering is attested by the numerous cut marks on skulls, phalanges, axial skeletons and long bones. Deliberate fracturing of the jaws and long bones is quite common. Other species such as deer, from a much broader area, were also carried intact to the cave, and similarly have numerous marks and de-liberate, systematic breakage. In this case, evidence of human activity predominates over the marks of small carnivores, which are superimposed on the cut marks. The carcasses of large herbivores such as aurochs, horses and wild asses (Equus hydrun-tinus) are quite incomplete. They may have been dismembered previously, with the most nutritious parts brought to the site. Evidence of carnivore impact on these remains is generally weak.
The fossil association on the archaeological levels of Boquete de Zafarraya cave is indicative of frequent occupation by Neanderthals in sum-mer, at the same time as carnivores exploited the cave. The mortality of juvenile deer and carni-vores coincides with that of juvenile capridae,
clear evidence of intervention by carnivores and humans at the same time. The considerable an-thropic presence at the Zafarraya site, evidenced by the stone tools found in every archaeo-stratigraphic unit, seems to have posed no im-pediment to panthers frequenting this cave. The abundance of goats and rabbits in the environs must have proved a particular challenge for both groups.
Palaeoecology
The Zafarraya cave environment was gener-ally inhabited by rock-dwelling species of large mammals directly related to the morphology of this landscape, as well as other species more char-acteristic of woodlands, areas with abundant wa-ter, the polje, sheltered zones and mountain are-as with lush vegetation, all propitiated by higher moisture levels than the current local conditions. The best represented carnivore species in Zafar-raya cave are dholes and leopards (Fig. 3), both rare at most European Pleistocene sites (all the more so in caves at this altitude). The dhole, typ-ical of the warmest periods of the Eemian inter-stadial, is robust, closer to the currently species found in semi-desert and mountain landscapes, where it hunts in groups. The leopard occu-pied several environments including forests and abrupt areas such as Zafarraya where herbivores hunted medium-small sized herbivores. Hyena and fox, more robust than the Mediterranean fox, were scarce while the lynx was slightly more numerous. Bears, lynx and weasel were quite ro-bust, indicating a temperate climate. Of all the carnivore taxa, only brown bear, bobcat and lynx are characteristic of forest habitats, while the rest were more or less ubiquitous (Barroso et al., 2006e; Monclova et al., 2012.).
The mountain goat is the most abundant herbi-vore in Zafarraya Cave, probably a preferential prey for Neanderthals, dholes and leopards. Relatively gracile, this goat is different from the larger pre-Würm species and the Mediterranean goats associ-ated with Mousterian sites. The continuous presence of deer indicates that it was hunted during summer in the polje woods near the cave. Chamois seek win-ter refuge in forests when the peaks and pastures are covered with snow, but in Zafarraya they ap-pear in more benign periods, probably because the

SOUTHERN MEDITERRANEAN COAST, GUADALQUIVIR RIVER AND BETIC INTRAMONTANE BASINS 469
moister climate at the time reduced the snow cover in the higher areas. The auroch appears frequently throughout the Pleistocene on the Iberian Peninsula. Its minor presence in Zafarraya could be associated with flooded areas in the cave near the polje. Wild boar are usually quite rare in the Pleistocene, and while scarce in Zafarraya, they show a preference for forested and, more particularly, wet areas. Horses, also rare, are represented by the robust Equus cabal-lus germanicus, known in Western Europe during the first half of the last glaciation. Its relative absence could be related to the site’s elevation. The wild ass, frequent since the last interglacial and typical of the Mediterranean region since the beginning of the Middle Pleistocene, could survive in rugged areas like Zafarraya (Monclova et al., 2012).
Given that the few mammal taxa at Zafarraya reduce the representativity of the cenograms, in some of them data from several archaeo-strati-graphic units have been used to clarify the site’s environmental and climatic evolution. On the ba-sis of the preferences shown by different taxa for open/closed, wet/dry environments, these ceno-grams point to a moist climate which only under-went minor variations throughout the stratigraph-ic sequence. The lack of a dividing line between the data on the large and small body mass of all Zafarraya mammals indicates that the habitat was closed. When carnivores (average sized species) are removed from the analysis, a dividing line does appear between macro– and micro-mammals, indicative of the presence of open spaces on the edge of the forests in a temperate and permanent-ly moist climate.
Evidence of a closed environment not far from the rocky cave landscape, in a climate tending to be moist, matches the polje. The various biotopes around Zafarraya formed a landscape mosaic in which different forest fauna taxa coexisted with other inhabitants of more open and even more humid environments such as streams and wet-lands, which only retreated to the polje in the driest periods. Through the successive archaeo-stratigraphic units in Zafarraya, an abundance of hypsodont animals alternated with braquidonts, an indication of mountain + grassland or forested environments, respectively.
The climatogram drafted to compare Zafar-raya’s major archaeo-stratigraphic units clusters species by their ecological affinities (giving pri-
ority to the moisture gradient). It illustrates the climate trend through the sedimentary infill of the cave as the fauna composition changed in the stratigraphic sequence. Some results are dif-ficult to interpret. While the goat/deer or bison/deer ratio reflects variations in the intensity of the cold at many Mousterian sites, the altitude of the Zafarraya site hinders this pattern, and thus the continued presence of goats does not neces-sarily indicate variations in the intensity of the cold but rather oscillations linked to the presence or absence of other species. Similarly, although the presence of horses might indicate alternat-ing colder or drier climates, here they are not only rare but their remains are associated with archaeo-stratigraphic units with highly uneven moisture levels according to other records. In un-gulates, neither chamois nor aurochs show vari-ations related to the frequency of the presence of deer in the stratigraphic sequence, while the presence of wild boars, usually associated with in-creased moisture, does match the increase in deer and chamois, also in a moister period (Monclova et al., 2012). Finally, variations in the proportions of the macromammals through the Zafarraya stratigraphic sequence seem to confirm a moist, temperate climate, cooler than today, with a wet-ter initial phase when the forests spread, but gen-erally characterized by Mediterranean conditions. The various Neanderthals occupations occurred during the more or less fresh and more or less temperate periods, always at the start of or during the summer, when leopards and dholes arrived.
Taphonomy
The taphonomic and zooarchaeological analy-sis of Zafarraya cave gave rise to a proposed model in which the Neanderthal occupants of the cave became specialist hunters of goats, the most abun-dant species in the record (Barroso and de Lumley, H. 2006). However, the wide diversity of herbi-vore prey that inhabited the environment and the parallel presence –and continued occupation– of carnivores and Neanderthals prevent this model from fully reflecting the interaction between the two. Their overlap in the cave may have occasion-ally led to competition for food resources in the environs. The interactions between Neanderthals, carnivores and herbivorous prey has been the sub-ject of a recent study (Caparros et al., 2012).

PLEISTOCENE AND HOLOCENE HUNTER-GATHERERS IN IBERIA AND THE GIBRALTAR STRAIT:
THE CURRENT ARCHAEOLOGICAL RECORD470
There are three possible scenarios: 1) Neander-thals predominated over carnivores and excluded them from the surrounding ecological system, or vice versa. 2) Once the Neanderthal and carnivore niches overlapped, one excluded the other from the immediate ecological environment. 3) Once the two niches overlapped, neither was able to exclude the other and they coexisted. In order to determine which of the three was most likely, we studied the overlapping Neanderthal/carnivore occupation of the cave from the perspective of their interaction with the herbivore prey items and also their competition for Capra, the most abundant resource.
Using the zooarchaeological analysis and the known ecology of the taxa which fed in the vi-cinity of the cave, a path analysis (Wright, 1968) was applied to the faunal data to define the like-ly fauna acquisition strategies by Neanderthals and carnivores. Initially, the applicable scenario seems to be a complete overlap of the Nean-derthal and carnivore niches, both cohabiting the cave environment. The analysis shows that: 1) Although the continued presence of Capra made it a prey for both groups, Panthera was the main predator, alternating with the Neander-thals in the cave occupation. 2) Rupicapra was overwhelmingly hunted by Panthera and rarely by Neanderthals. 3) Neanderthals were the main accumulators of Cervus, hunted at some distance from the cave and probably scavenged by the carnivores. 4) Large herbivores such as horses and aurochs were hunted by Neander-thals and secondarily by carnivores.
In the vicinity of the cave, Panthera was clearly more efficient than the Neanderthals, who were excluded when this carnivore was present (Capra and Rupicapra remain in the record). The large herbivores that roamed the Zafarraya polje probably offered good opportu-nities for cooperative hunting by Neanderthals (accumulation of Cervus and others). If there was indeed coexistence, as evidenced by scav-enging, it must have been during short periods of random or seasonal occupation.
Dating
Following the discovery of the B.Z.C. site, pre-liminary studies were presented in several publi-
cations, where the age of the Mousterian levels were estimated to be more recent than other Middle Palaeolithic sites in the region. This first estimate was based on a biostratigraphic study which compared rodent populations found in the Zafarraya sediments and those in Cariguela. This latter site was considered contemporary to “Würm II”, permitting the inference that Zafar-raya could be the “Würm II-III” or “Würm III” in-terstadial. In 1989, Barroso et al., forwarded the open hypotheses that the Zafarraya fossils might represent Europe’s last Neanderthal populations. The first series of radiocarbon datings from the Mousterien levels of B.Z.C. were presented in 1995 (Hublin et al., 1995). Two methods were used, C-14 (based on the acid fraction and the collagen) and the U-Th method, using herbi-vore bone samples. The ages for C-14 (colla-gen) ranged from 29.8 + 0.6 ky (Gif-9140-II) and higher levels of 31.8 + 0.55 ky (Gif/LSM-9140-I) level 8, while the acid fraction, for the same samples gave 23.6 + 1 + 0.1 ky and 22.0 ky. The U-Th ages are stratigraphically consist-ent, ranging from 25.1 + 1.3 ky on level I-3 to 31.7 + 3 ky on level I-8. Level D, the location of one of the Neanderthal fossils (Zaf. 2), was dated by U-Th at 33.4 + 0.2 ky. These absolute dates showed that between 30 ky and 33 ky BP, groups of Neanderthals survived the pressure of A.M.H. in a sort of “refuge” at the southern extreme of Iberia. The hypothesis of the “last Neander-thals” in Zafarraya thus seemed to be confirmed at this site, which gave rise to the paradigm. A new set of absolute datings began almost a decade later using several methods (Michel et al., 2003, Michel et al., 2006). C-14 dat-ings (AMS) of 20 charcoal samples by ORAU yielded quite mixed results because the collec-tion methodology for the 1996 samples was not excessively rigorous. Samples from stratigraph-ic sections in grids which were never dug, the source of the problem were: Q-9, P-9, R-9, P-20 and R-1. These samples, with the exception of R-16, yielded aberrant dates.
The charcoal samples located and extracted from the stratigraphic sections under controlled conditions during the dig were: P10395, Oxa-9001, P10397, P10398, Oxa-9002 and P10400. This methodological control makes these sam-ples highly reliable with respect to their spatial location and stratigraphic context.
SOUTHERN MEDITERRANEAN COAST, GUADALQUIVIR RIVER AND BETIC INTRAMONTANE BASINS 471
Eight faunal remains were chosen for C-14 (AMS) dating, although half the samples were rejected due to lack of collagen. The samples at depths from 168 cm to –171 cm yielded three ages ranging from (OxA 8411) 26.36 + 0.44 ky (Z6d) to (OxA 8999) 33.3 + 1.2 ky (Z8os). A sample from –223 cm yielded an incongruous dating, (OxA 9000) 30.65 + 0.65 ky (Z69os). U-Th alpha spectrometry was applied to 25 herbivore teeth. The results were not conclu-sive, as six ranged chronologically from 29,300 + 1.4-1.3 (Z4cem) to 39,700 + 2.2 (Z126os) at depths between –133 cm and –239 cm. Eight of the samples were between 40.9 + 1.3 ky (Z2os) and 60 + 3.7-3.6 ky (Z205os) at similar depths.
The U-Th thermal ionization mass spectrom-etry (TIMS) was applied to 9 samples, two of which were invalidated. The remaining 7 showed a range between 16.88 + 0.535 ky (Z6d.a) at 171cm and 53,345 + 5,025 (Z696e) at –133 cm. At the same depth, alpha spectrometry dated the –170 cm samples at 31.6 + 203 and for –133cm, at 32.8 + 0.201 –0.2 ky. The obvious contra-dictions prevent conclusive results from being reached.
The ESR (EU, early uptake) and ESR (LU, linear uptake) method was used with 6 horse and goat teeth. The result was (Z62) 24 + 0.3 ky
(EU) and 24 + 0.4 ky (LU) at –165cm, while the oldest (Z126) 44 + 0.5 ky (EU) and 45 + 0.6 ky was from –178 cm. The weighted mean of the most consistent results yielded by this method, i.e. above 30 ky, is 35 + 0.3 to 39 ky + 0.3 ky. The age of Neanderthal mandible Zaf. 2 could not be defined by high resolution gamma spec-trometry direct dating due to its low uranium content. The imprecise conclusion was that it must be at least 42 ky old.
The final conclusion was that the results showed a chronological range from 30 ky to 45 ky, thus leaving the possible late age of Neander-thals in southern Spain unresolved.
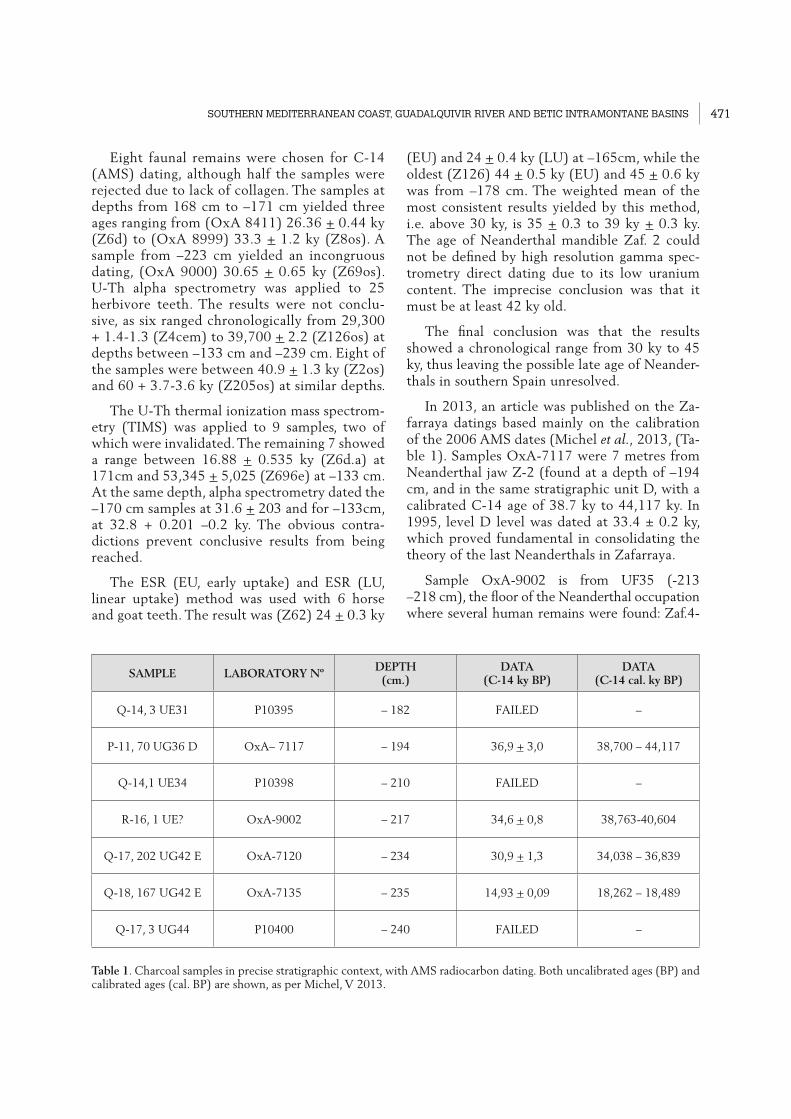
In 2013, an article was published on the Za-farraya datings based mainly on the calibration of the 2006 AMS dates (Michel et al., 2013, (Ta-ble 1). Samples OxA-7117 were 7 metres from Neanderthal jaw Z-2 (found at a depth of –194 cm, and in the same stratigraphic unit D, with a calibrated C-14 age of 38.7 ky to 44,117 ky. In 1995, level D level was dated at 33.4 ± 0.2 ky, which proved fundamental in consolidating the theory of the last Neanderthals in Zafarraya.
Sample OxA-9002 is from UF35 (-213 –218 cm), the floor of the Neanderthal occupation where several human remains were found: Zaf.4-
SAMPLE LABORATORY NºDEPTH(cm.)
DATA (C-14 ky BP)
DATA (C-14 cal. ky BP)
Q-14, 3 UE31 P10395 – 182 FAILED –
P-11, 70 UG36 D OxA– 7117 – 194 36,9 + 3,0 38,700 – 44,117
Q-14,1 UE34 P10398 – 210 FAILED –
R-16, 1 UE? OxA-9002 – 217 34,6 + 0,8 38,763-40,604
Q-17, 202 UG42 E OxA-7120 – 234 30,9 + 1,3 34,038 – 36,839
Q-18, 167 UG42 E OxA-7135 – 235 14,93 + 0,09 18,262 – 18,489
Q-17, 3 UG44 P10400 – 240 FAILED –
Table 1. Charcoal samples in precise stratigraphic context, with AMS radiocarbon dating. Both uncalibrated ages (BP) and calibrated ages (cal. BP) are shown, as per Michel, V 2013.

PLEISTOCENE AND HOLOCENE HUNTER-GATHERERS IN IBERIA AND THE GIBRALTAR STRAIT:
THE CURRENT ARCHAEOLOGICAL RECORD472
5-18, Zaf.15, Zaf.16, Zaf.27, Zaf.17 and Zaf.19. This level has been dated at 38,763 to 40,604 ky. Both this and the previous data seem quite reli-able, allowing the Zafarraya Neanderthal fossils to be dated between 38.7 ky and 44 ky.
The most recent publication on this issue (Wood et al., 2013), present results from Zafar-raya. The author applied extremely strict proto-cols such as ultrafiltration pretreatment (AF) of bone collagen to prevent contamination of the processed samples. Previously, the nitrogen con-tent (% N) of the selected bone sample is meas-ured to determine the collagen content, which is used to decide whether or not to radiocarbon date the sample.
Two stages were used in the Zafarraya analysis. In the first stage, bone samples previously dated by C-14 (AMS) at the ORAU (OxA-8024, OxA-8999, OxA-9000, OxA-8411) with results published in Michel et al., 2003 and Michel et al., 2006.) were chosen. Only sample Z8os, which was subject to the AF protocol, yielded a dating of 33.3 + 1.2 ky. The rest had a low % N and were rejected. In the second stage, 29 bone samples from the stratigraphy were analysed. Only two had an acceptable %N, ZAF2 (0.8) and ZAF5 (1.5). and 0.8 Another three sam-ples, ZAF3 (0.7) ZAF7 (0.6) and ZAF8 (0.6), were on the borderline. The AF protocol was applied to these samples, yielding results in ZAF2 (46.3 ± 2.5 ky) and ZAF7 (49.3 ky). The latter, almost on the
threshold of the method, showed an age greater than 46.7 ± 0.7 ky, i.e., roughly 13,000 years older.
The work by both Michel and Wood under-scores the age of the Mousterian levels at Zafar-raya, between 38.7 ky and 49.3 ky.
Only 6 out of 33 analysed samples contained a % N that would permit collagen to be obtained, and only three of these specimens could be dated, less than 10% of all the selected samples. This method-ology was also used at other sites along the Mediter-ranean coast: Les Cendres (3 samples), Mallaetes (2 samples), El Niño (23 samples), Quebrada (9 sam-ples), Sima de las Palomas (1 sample), El Salt (43 samples), Nerja (25 samples) and Gorham’s Cave (73 samples), all with negative results.
These newer results clearly question not only the chronological framework on which the evidence of the Middle Palaeolithic in the Spanish Mediterra-nean was based, but also the very existence of the last Neanderthals in their “refuge” in southern Iberia. This paradigm is thus now under serious challenge, and the existence of these late hominids is difficult to sustain at present. After Zafarraya, Gorham’s Cave was presented as further near-indisputable proof for the paradigm, however in the light of the data pro-vided above, this can no longer be sustained. Other challenges or scenarios must be investigated to ex-plain what seems to have been a “depopulation” of southern Iberia from 40 ky until the arrival of the Gravettian groups.
THE LOWER AND MIDDLE PLEISTOCENE
IN THE GUADIX-BAZA AND ORCE BASIN
Introduction and geological context
Robert Sala Ramos*
The Guadix-Baza region spans a large area in south-eastern Iberia. It was a vast intramontane endorheic basin between the Miocene and the end of the Middle Pleistocene, when regional tectonics led to the formation of the Guadiana Menor River which drained the former lake system. The first
hominins who settled in Western Europe occupied this area at a time when the Guadix sub-basin was crossed by a hydrological system, descending from Sierra Nevada and Sierra de Cazorla and flowing into a lake in the Baza sub-basin in the eastern part of the region.
This 3,000 km2 basin lies between the Internal and External Zones of the Betic Range. The Cenozoic infill was initially composed of marine
* Universitat Rovira i Virgili. Institut Catala de Paleoecologia Humana i Evolucio Social.




















