
Association of the p85 regulatory subunit of phosphatidylinositol 3-kinase with an essential...
10
1993 82: 3530-3538 TC He, H Zhuang, N Jiang, MD Waterfield and DM Wojchowski kinase with an essential erythropoietin receptor subdomain Association of the p85 regulatory subunit of phosphatidylinositol 3- http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requests Information about reproducing this article in parts or in its entirety may be found online at: http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprints Information about ordering reprints may be found online at: http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtml Information about subscriptions and ASH membership may be found online at: reserved. Copyright 2011 by The American Society of Hematology; all rights 900, Washington DC 20036. weekly by the American Society of Hematology, 2021 L St, NW, Suite Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published For personal use only. by guest on July 14, 2011. bloodjournal.hematologylibrary.org From
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Association of the p85 regulatory subunit of phosphatidylinositol 3-kinase with an essential...

1993 82: 3530-3538
TC He, H Zhuang, N Jiang, MD Waterfield and DM Wojchowski kinase with an essential erythropoietin receptor subdomainAssociation of the p85 regulatory subunit of phosphatidylinositol 3-
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
reserved.Copyright 2011 by The American Society of Hematology; all rights900, Washington DC 20036.weekly by the American Society of Hematology, 2021 L St, NW, Suite Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published
For personal use only. by guest on July 14, 2011. bloodjournal.hematologylibrary.orgFrom

RAPID COMMUNICATION
Association of the p85 Regulatory Subunit of Phosphatidylinositol 3-Kinase With an Essential Erythropoietin Receptor Subdomain
By Tong-Chuan He, Hongming Zhuang, Ning Jiang, Michael D. Waterfield, and Don M. Wojchowski
Using an active, HA1 epitope-tagged form of the murine erythropoietin (EPO) receptor and via direct coimmunopre- cipitation, the p85 regulatory subunit of phosphatidyl inosi- tol-3 kinase (p85/P13-K) is shown to associate with the EPO receptor in transfected FDC-P1 cell lines. Coimmuno- precipitation of p85 with epitope-tagged EPO receptors was observed initially in FDC-HER cells labeled metaboli- cally with [32P]orthophosphate, and association of these factors was confirmed by Western analyses of receptor immunoprecipitates using p85 antiserum. Interestingly, this association occurred in the absence of ligand, and ex- posure of FDC-HER cells to EPO did not detectably affect levels of receptor-associated p85 or overall levels of p85 phosphorylation. However, EPO was observed to stimu-
URING RED BLOOD CELL development, pro- D erythroblasts in marrow, spleen, and fetal liver (col- ony forming units-erythroid [CFU-e]) require exposure to the glycoprotein hormone, erythropoietin (EPO), for sur- vival, proliferation, and terminal differentiation.’-’ Insight into mechanisms involved in proliferative signalling has been provided, in part, through the recent cloning and analy- sis of the murine and human EPO receptors.495 Linked pat- tern sequence analyses originally of receptors for growth hormone, prolactin, interleukin-6 (IL-6), and IL-26 showed that the EPO receptor is related structurally to these, and an expanding superfamily of cytokine receptors (eg, receptors for IL-3, IL-4, IL-5, IL-7, granulocyte-macrophage colony- stimulating factor [GM-CSF], granulocyte-CSF [G-CSF], leukocyte-inhibitory factor [LIF], and ciliary neurotrophic factor [CNTF]).7 Conserved structure exists primarily within extracellular domains and includes four codistrib- uted cysteine residues, a membrane proximal WSXWS mo- tif, and paired sevenfold p-strand predicted ligand-binding domains. Limited homology within membrane-proximal cytosolic regions also has been recognized among the recep-
From the Department of Molecular and Cell Biology and the Center for Gene Regulation, The Pennsylvania State University, University Park, PA; and the Ludwig Institute for Cancer Research and Department ofBiochemistry and Molecular Biology, University College, WIPBBT, London, UK.
Submitted September 2, 1993; accepted September 28, 1993. Supported by National Institutes of Health Grants No.
R29DK40242 and ROI-HL44491 to D.M. W. Address reprint requests to Don D. Wojchowski, MD, Molecular
and Cell Biology and the Center for Gene Regulation, 408 S Frear Laboratory, The Pennsylvania State University, University Park, PA 16802.
The publication costs ofthis article were defayed in part by page charge payment. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. section I734 solely to indicate this fact. 0 1993 by The American Society of Hematology. 0006-4971/93/8212-0034$3.00/0
lated the rapid formation of phosphatidylinositol 32P-phos- phate in FDC-HER and FDC-ER cells. Through baculovirus- mediated expression of epitope-tagged EPO receptor forms in SF9 cells, domains for p85 association were mapped. Analyses of receptor forms with cytosolic trunca- tions and deletions delineated a candidate subdomain for p85 binding to an essential extended box-2 region (P329- E374; including a putative motif for SH2 binding, Y343LVL). These findings extend a mechanistic alignment between the EPO receptor and protein tyrosine kinase-encoding re- ceptors that likewise activate P13-K, and expand the im- portance of further defining pathways to P13-K activation. 0 1993 by The American Society of Hematology.
tors for EPO, IL-6 (gpl30), IL-2 (p-receptor), IL-3, G-CSF, IL-7, and IL-4 and is restricted to conserved 8- to 12-amino acid essential box- 1 and box-2 subdomains and intervening residue^.^^^ Within the EPO receptor, a subdomain immedi- ately distal to the box 2 domain also has been shown to be essential for mitogenic signalling.10%” Functional subdo- mains have been delineated largely through studies of re- constituted mitogenic activity of mutated EPO receptor forms in transfected myeloid and lymphoid cell lines.” These studies also show that a delimited membrane-proxi- mal 90-amino acid cytosolic receptor domain (including the above box motifs) mediates EPO-induced pr~liferation,’~,~’ whereas mitogenic signalling is modulated negatively by a distal carboxyl terminal
With regard to mechanisms of proliferative signalling, early studies using isolated proerythroblasts have associated EPO action with increases in intracellular calcium, cyclic AMP, and arachidonic acid.” However, little progress has been made in identifying direct targets in these pathways, and no recognizable catalytic domains are encoded within the cloned EPO receptor. More recently, EPO has been shown to rapidly activate protein tyrosine phosphoryla- tion,I5 and EPO-induced tyrosine phosphorylation of raf- 1,16 p120-GAP,I7 the EPO receptor per se,” and additional cellular protein^'^.*^ has been demonstrated. The latter pro- teins include a molecular weight (Mr) 97,000/ 100,000 cyto- solic factor,”*’ which also is tyrosine phosphorylated in re- sponse to IL-3 and GM-CSF,” as well as an Mr 130,000 potential protein tyrosine kinase (PTK) that associates with EPO-receptor complexes.” Very recent studies by Witth- uhn et al’’ indicate that this Mr 130,000 PTK corresponds to JAK2, a member of the Janus family of PTKsZ4 that is shown in vitro to associate with the EPO receptor and to be activated by EPO. Thus, the above work aligns the EPO receptor mechanistically with growth factor receptors that directly encode ligand-activatable PTKs (eg, kit, fms, trk, EGF and PDGF receptor family,25 as well as the class I1 cytokine receptor for interferon a lp that recently has been shown to activate the nonreceptor PTK, TYK-2.26 In the
3530 Blood, Vol 82, No 12 (December 15), 1993: pp 3530-3538
For personal use only. by guest on July 14, 2011. bloodjournal.hematologylibrary.orgFrom

€PO RECEPTOR INTERACTION WITH P85/P13-KINASE 3531
present study, the apparent overlap in mechanisms of prolif- erative signal transduction for these receptors versus the EPO receptor is extended through the observed constitutive association between an epitope-tagged form of the murine EPO receptor with the regulatory subunit of a second kinase that is known to associate with at least certain PTK-encod- ing receptors, ie, phosphatidylinositol-3 kinase (P13-K).27,28 Using recombinant factors expressed in insect SF9 cells, the essential extended box-2 cytosolic subdomain of the EPO receptorlo*’’ is also delineated as a candidate site for p85 binding.
MATERIALS AND METHODS
Cell lines and mitogenic assays. Murine myeloid IL-3/GM- CSF-dependent FDC-P 1 cells;9 FDC-ER cells,” and FDC-HER cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum, mol/L 2-mercaptoethanol, and 3% conditioned medium from WEHI-3B cells3’ Proliferation assays were performed in 96-well plates (2 X lo4 cells; 0.1 mL) using purified recombinant human EPO (Amgen Biologicals, Thousand Oaks, CA). Cells were labeled for 2 hours at 48 hours of culture with [’Hlthymidine (10 pCi/mL).
Construction of epitope-tagged and mutated EPO receptor forms. In part to improve immunoprecipitation of EPO receptors from transfected cell lines, an epitope (YDVPDYAS) derived from influenza virus hemagglutinin” was used to tag the murine EPO receptor at the amino terminus. A cDNA encoding this epitope (and a signal peptide for secretion) was prepared by polymerase chain reaction (PCR) from an epitope-tagged form of EPO’, using the primers S’-ACGGGTACCATGGGGGTGCACGAATG-3’ and 5‘-ATGGTCGACGCTGGCGTAGTCTGGGA-3’. This amplified fragment was cloned into Kpn I and Sal I sites of pBluescript-IIKS+ (Stratagene, La Jolla, CA). PCR primers (5’-TCGGTCGACGCA- CCTTCACCCAGCCT-3’ and 5’-TCCCCAGCCTGAGTCACT- GCCCAGCG-3’) were then used to amplify the amino terminus (A1-G3,’) of murine EPO receptor cDNA. This amplified fragment was cloned into the above construct at Sal I and BamHI sites, yield- ing a construct encoding the HA1 epitope, two intervening residues (V and D), and the N-terminus ofthe mature receptor. A full-length construct (pBHER) was completed by subcloning the BamHI to Not I fragment of the EPO receptor into the above construct, and this tagged EPO receptor cDNA was cloned into 5’ Kpn I and 3’Not I sites of a modified pXM4 expression vector (pXM-HER). Trun- cated epitope-tagged EPO receptor (HER) forms were constructed by inserting a 14-bp Xba I stop linker (Pharmacia, Piscataway, NJ) into the blunt-ended Bgl I1 site of pXM-HER (HER-K256) and in- serting a cassette with stop codons in each reading frame (5’-AGC- TAGTCTAGACTAGC-3’ and 5’-GGCCGCTAGTCTAGACT-3’) at Hind111 and Not I sites of pXM-HER (HER-E374). A receptor form with an in-frame deletion of the box 2 cytosolic domain (HERAB X 2) was constructed by ligating the BamHI and Apa I sites of the above construct using the oligo 5’-GATCGGCC-3’ (re- sulting in the insertion of a single arginine residue). All constructs were confirmed by restriction analyses and DNA sequencing.
HAI-tagged EPO receptor cDNAs (42 pg) were cotransfected with a G4 18-resistance plasmid (pM5-neo, 8 pg) into FDC-PI cells ( IO7 cells/mL in phosphate-buffered saline [PBS]) by high-voltage electroporation (562 V/cm, 950 p F geneZAPPER, IBI, New Haven, CT). Transfected cells were selected in G418 at 1 mg/mL (Geneticin; GIBCO BRL, Grand Island, NY) and, subse- quently, in medium lacking conditioned medium from WEHI-3 cells and containing EPO (5 U/mL). Sublines were established by dilutional cloning.
Gene transfer.
Receptor immunoprecipitation, metabolic labeling, and Western blotting. Exponentially growing FDC-P 1 -derived cell lines typi- cally were washed three times in DMEM and were starved in DMEM plus 1% fetal calfserum for 12 hours before EPO induction. In induction experiments, starved cells (1 X IO7 cells/mL) were incubated with 50 nmd/L Na3V04 at 37°C for 15 minutes, exposed to EPO (10 to 50 U/mL) for 30 minutes, collected by microcentrifu- gation, and lysed in 200 pL of 1% Triton X-100,50 mmol/L NaCl, 30 mmol/L Na4P207, 50 mmol/L NaF, 0.1 mmol/L Na3V04, 10 mmol/L Tris, pH 7.6, 0.5 pg/mL leupeptin, 0.7 pg/mL pepstatin, 50 pg/mL phenymethylsulfonyl fluoride, and 0.1% bovine serum albumin (lysis buffer). In immunoprecipitations, lysates were preexposed to protein A-sepharose (Sigma Chemicals, St Louis, MO) and then incubated with monoclonal antibody (MoAb) 12CA5 (20 pg; Berkeley Antibody Co, Berkeley, CA) for 2 hours at 4°C. Immune complexes then were absorbed using protein A-se- pharose (45 minutes at 4°C) and were eluted in sample buffer for sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS- PAGE). Western blotting of immune complexes was performed using antiserum to p85 of P13K33 or to EPO receptor peptides.34 Detection was by NBT/BICP using alkaline phosphatase-conju- gated second antibodies (Promega, Madison, WI).
Alternatively, starved cells were incubated in phosphate-free DMEM (Sigma Chemicals) supplemented with 1% fetal calf serum for 30 minutes and labeled metabolically for 2.5 hours in this me- dium containing 0.5 mCi/mL [32P]orthophosphate (ICN, Irvine, CA). Incubations with orthovanadate and EPO were performed as described above. 32P-labeled proteins from Triton X- 100 lysates were precipitated using MoAb 12CA5, antiphosphotyrosine MoAb PY20 (ICN), or antiserum to p85 (PI3-K) plus protein A-sepharose. 32P-phosphoproteins were eluted as above, and analyzed by SDS- PAGE and autoradiography.
Baculovirus constructs and expression of p85 and EPO receptor forms in SF9 cells. cDNA constructs encoding HER, HER-E374, HER-K256, and HERAB X 2 were subcloned into the baculovirus transfer vector pVL1393 (Invitrogen, San Diego, CA) at Kpn I and Not I sites. These plasmids then were cotransfected into insect SF9 cells35 together with linear, replication-defective baculovirus DNA (Pharmingen, San Diego, CA). Recombinant viruses were collected from transfected cells at 72 hours postinfection and were amplified by repeated infections. Expression levels were assessed by Western blotting using either antiserum to receptor peptides or MoAb 12CA5. The baculovirus vector for expression of p85 was as de- scribed previously.” In studies of association of p85 with HAI- tagged EPO receptor forms, SF9 cells were coinfected with cognate baculovirus vectors, collected 45 hours postinfection, lysed, and analyzed by immunoprecipitation (MoAb 12CA5) and Western blotting as described above.
Immunoprecipitations of EPO receptors were performed in FDC-HER cells using MoAb 12CA5 or anti- EPO receptor peptide antibodies as described above. After incuba- tions with protein A-sepharose, gels were washed three times with each of the following buffers: PBS, pH 7.4; 50 mmol/L LiC1, 100 mmol/L Tris-HC1, pH 7.4; 100 mmol/L NaCl, 1 mmol/L EDTA, 10 mmol/L Tris-HC1, pH 7.4. Kinase assays were performed essen- tially according to Auger et al.36 Briefly, washed gels were resus- pended in 10 mmol/L MgCl,, 25 mmol/L HEPES, pH 7.4. Phos- phatidylinositol (Avanti Polar Lipids, Alabaster, AL) micelles (prepared by sonication in 25 mmol/L HEPES, pH 7.4, 1 mmol/L EDTA) were then added at a final concentration of 0.2 mg/mL (total volume, 50 pL). Kinase reactions were initiated by addition of 10 pCi of [y3,P]ATP (4,500 Ci/mmol; NEN, Boston, MA) and were incubated at 25°C for 20 minutes. Reactions were terminated by the addition of 0.02 mL of 8 N HCI and 0.12 mL of a 1 : 1 mixture
PZ3-kinase assays.
For personal use only. by guest on July 14, 2011. bloodjournal.hematologylibrary.orgFrom
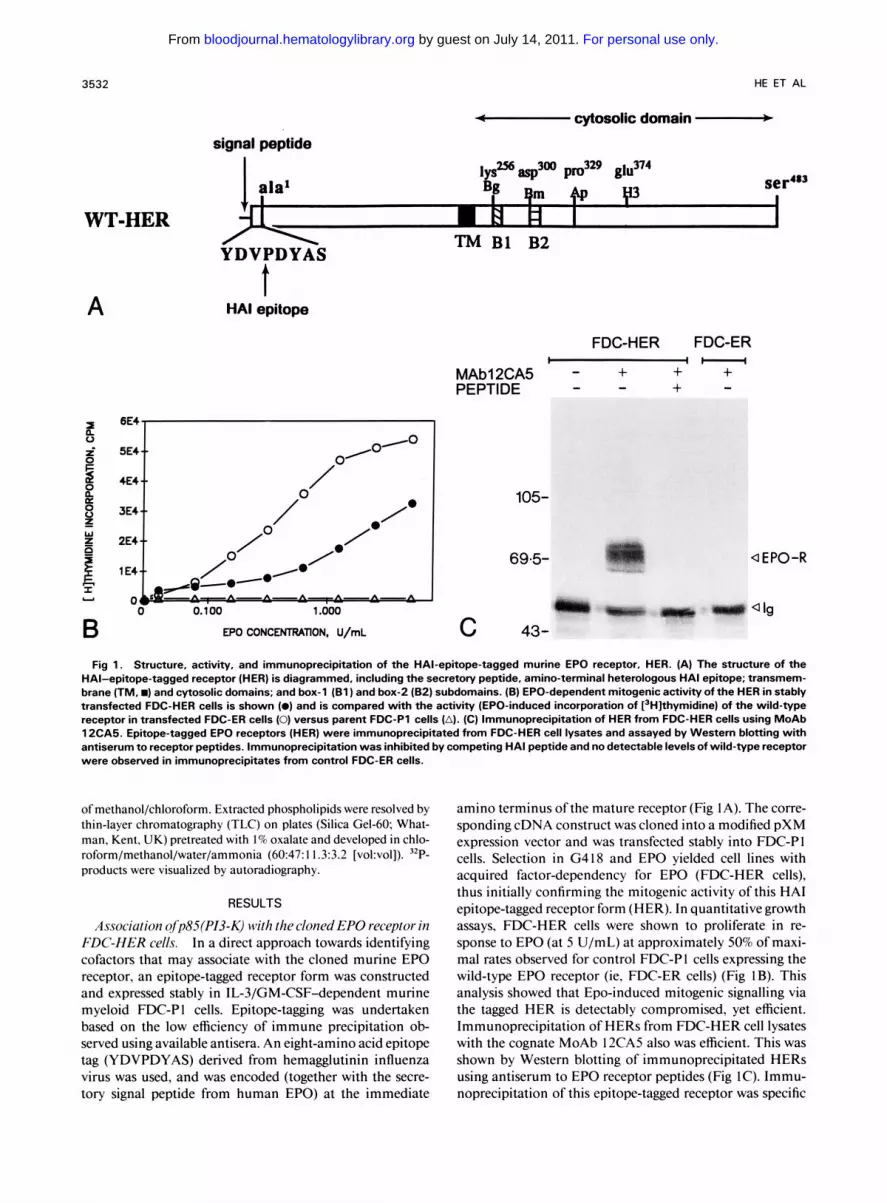
3532 HE ET AL - cytosolic domain
signal peptide
WT-HER YDVPDYAS TM B1 B2
A
x a 0
i 0 z x a z z
0 0
W
I Y
B
BE4
5E4
4€4
3E4
2E4
1 E4
t HA1 epitope
EPO CONCENTRATION, U/mL
FDC-HER FDC-ER
+ + + +
I 1 -
- - MAbl2CA5 - PEPTIDE -
105-
69.5-
c 43-
Q EPO-R
Q lg
Fig 1. Structure, activity, and immunoprecipitation of the HAl-epitope-tagged murine €PO receptor, HER. (A) The structure of the HAl-epitope-tagged receptor (HER) is diagrammed, including the secretory peptide, amino-terminal heterologous HA1 epitope; transmem- brane (TM. m) and cytosolic domains; and box-1 (Bl) and box-2 (82) subdomains. (B) €PO-dependent mitogenic activity of the HER in stably transfected FDC-HER cells is shown (0) and is compared with the activity (EPO-induced incorporation of [jHIthymidine) of the wild-type receptor in transfected FDC-ER cells (0) versus parent FDC-P1 cells (A). (C) Immunoprecipitation of HER from FDC-HER cells using MoAb 12CA5. Epitope-tagged EPO receptors (HER) were immunoprecipitated from FDC-HER cell lysates and assayed by Western blotting with antiserum to receptor peptides. Immunoprecipitation was inhibited by competing HA1 peptide and no detectable levels of wild-type receptor were observed in immunoprecipitates from control FDC-ER cells.
o f methanol/chloroform. Extracted phospholipids were resolved by thin-layer chromatography (TLC) on plates (Silica Gel-60; What- man, Kent, UK) pretreated with I % oxalate and developed in chlo- roform/methanol/water/ammonia (6047: I I .3:3.2 [vol:vol]). "P- products were visualized by autoradiography.
RESULTS
Association ofp85(PI3-K) with thecloned EPO receptor in FDC-HER cells. In a direct approach towards identifying cofactors that may associate with the cloned murine EPO receptor, an epitope-tagged receptor form was constructed and expressed stably in IL-3/GM-CSF-dependent murine myeloid FDC-P I cells. Epitope-tagging was undertaken based on the low efficiency of immune precipitation ob- served using available antisera. An eight-amino acid epitope tag (YDVPDYAS) derived from hemagglutinin influenza virus was used, and was encoded (together with the secre- tory signal peptide from human EPO) at the immediate
amino terminus ofthe mature receptor (Fig IA). The corre- sponding cDNA construct was cloned into a modified pXM expression vector and was transfected stably into FDC-PI cells. Selection in G418 and EPO yielded cell lines with acquired factor-dependency for EPO (FDC-HER cells), thus initially confirming the mitogenic activity of this HA1 epitope-tagged receptor form (HER). In quantitative growth assays, FDC-HER cells were shown to proliferate in re- sponse to EPO (at 5 U/mL) at approximately 50% of maxi- mal rates observed for control FDC-PI cells expressing the wild-type EPO receptor (ie, FDC-ER cells) (Fig IB). This analysis showed that Epo-induced mitogenic signalling via the tagged HER is detectably compromised, yet efficient. Immunoprecipitation of HERS from FDC-HER cell lysates with the cognate MoAb 12CA5 also was efficient. This was shown by Western blotting of immunoprecipitated HERS using antiserum to EPO receptor peptides (Fig IC). Immu- noprecipitation of this epitope-tagged receptor was specific
For personal use only. by guest on July 14, 2011. bloodjournal.hematologylibrary.orgFrom
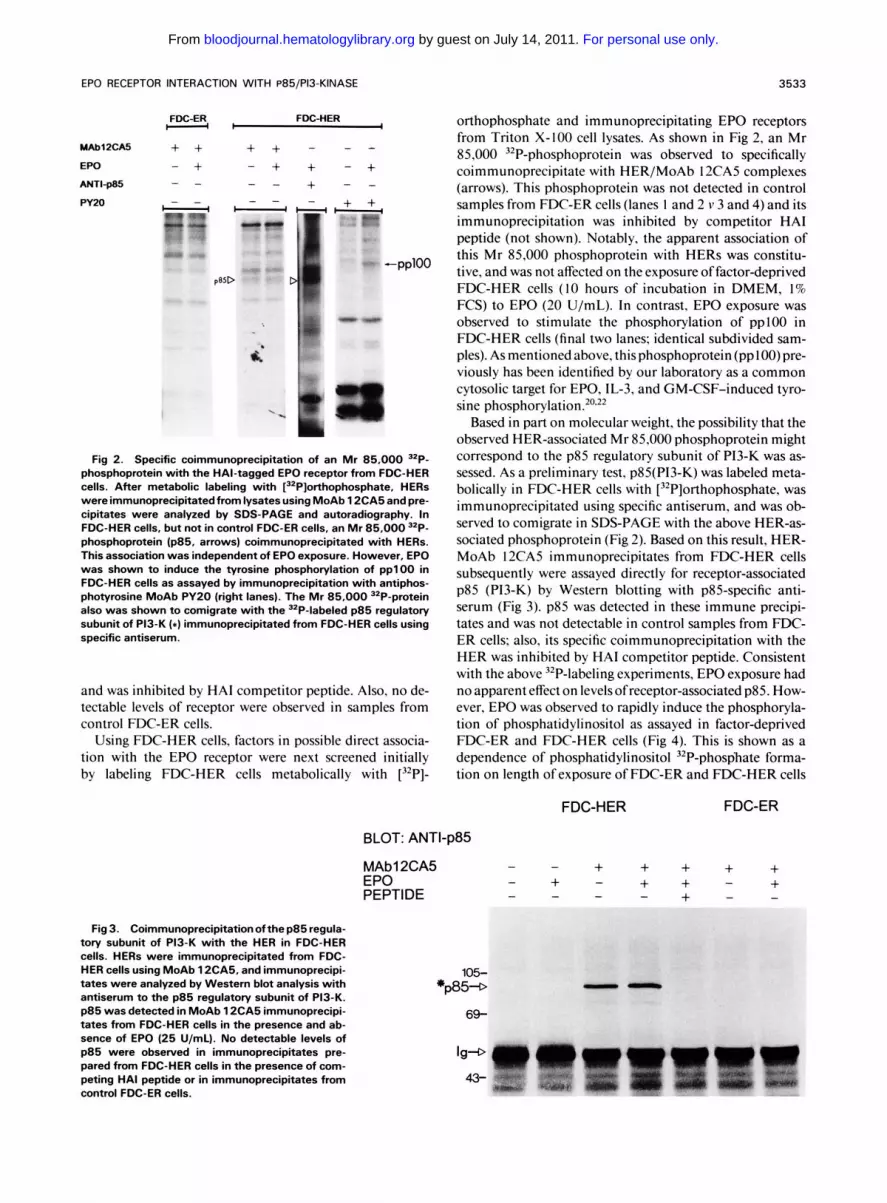
€PO RECEPTOR INTERACTION WITH P85/P13-KINASE 3533
FDC-ER FDC-HER H C I
MAb12CM + + + + - - - EPO - + - + + - +
+ - - NIP85 + + w20
- - - - - - - - - - 1 . -4
-pplOO
p8SD -- .I Fig 2. Specific coimmunoprecipitation of an Mr 85,000 "P-
phosphoprotein with the HAl-tagged EPO receptor from FDC-HER cells. After metabolic labeling with [32P]orthophosphate, HERS were immunoprecipitatedfrom lysates using MoAb 1 2CA5 and pre- cipitates were analyzed by SDS-PAGE and autoradiography. In FDC-HER cells, but not in control FDC-ER cells, an Mr 85,000 32P- phosphoprotein (p85, arrows) coimmunoprecipitated with HERs. This association was independent of EPO exposure. However, EPO was shown to induce the tyrosine phosphorylation of pp l00 in FDC-HER cells as assayed by immunoprecipitation with antiphos- photyrosine MoAb PY20 (right lanes). The Mr 85,000 32P-protein also was shown to comigrate with the 32P-labeled p85 regulatory subunit of P13-K ( 8 ) immunoprecipitated from FDC-HER cells using specific antiserum.
and was inhibited by HA1 competitor peptide. Also, no de- tectable levels of receptor were observed in samples from control FDC-ER cells.
Using FDC-HER cells, factors in possible direct associa- tion with the EPO receptor were next screened initially by labeling FDC-HER- cells metabolically
Fig 3. Coimmunoprecipitation of the p85 regula- tory subunit of P13-K with the HER in FDC-HER cells. HERs were immunoprecipitated from FDC- HER cells using MoAb 12CA5, and immunoprecipi- tates were analyzed by Western blot analysis with antiserum to the p85 regulatory subunit of P13-K. p85 was detected in MoAb 12CA5 immunoprecipi- tates from FDC-HER cells in the presence and ab- sence of EPO (25 U/mL). No detectable levels of p85 were observed in immunoprecipitates pre- pared from FDC-HER cells in the presence of com- peting HA1 peptide or in immunoprecipitates from control FDC-ER cells.
with [32Pj-
orthophosphate and immunoprecipitating EPO receptors from Triton X-100 cell lysates. As shown in Fig 2, an Mr 85,000 32P-phosphoprotein was observed to specifically coimmunoprecipitate with HER/MoAb 12CA5 complexes (arrows). This phosphoprotein was not detected in control samples from FDC-ER cells (lanes I and 2 v 3 and 4) and its immunoprecipitation was inhibited by competitor HA1 peptide (not shown). Notably, the apparent association of this Mr 85,000 phosphoprotein with HERs was constitu- tive, and was not affected on the exposure of factor-deprived FDC-HER cells ( I O hours of incubation in DMEM, 1% FCS) to EPO (20 U/mL). In contrast, EPO exposure was observed to stimulate the phosphorylation of ppl00 in FDC-HER cells (final two lanes; identical subdivided sam- ples). As mentioned above, thisphosphoprotein(ppl00)pre- viously has been identified by our laboratory as a common cytosolic target for EPO, IL-3, and GM-CSF-induced tyro- sine phosphorylation.20.22
Based in part on molecular weight, the possibility that the observed HER-associated Mr 85,000 phosphoprotein might correspond to the p85 regulatory subunit of P13-K was as- sessed. As a preliminary test, p85(P13-K) was labeled meta- bolically in FDC-HER cells with [32P]orthophosphate, was immunoprecipitated using specific antiserum, and was ob- served to comigrate in SDS-PAGE with the above HER-as- sociated phosphoprotein (Fig 2). Based on this result, HER- MoAb 12CA5 immunoprecipitates from FDC-HER cells subsequently were assayed directly for receptor-associated p85 (P13-K) by Western blotting with p85-specific anti- serum (Fig 3). p85 was detected in these immune precipi- tates and was not detectable in control samples from FDC- ER cells: also, its specific coimmunoprecipitation with the HER was inhibited by HA1 competitor peptide. Consistent with the above 32P-labeling experiments, EPO exposure had no apparent effect on levels of receptor-associated p85. How- ever, EPO was observed to rapidly induce the phosphoryla- tion of phosphatidylinositol as assayed in factor-deprived FDC-ER and FDC-HER cells (Fig 4). This is shown as a dependence of phosphatidylinositol 32P-phosphate forma- tion on length of exposure of FDC-ER and FDC-HER cells
FDC-HER FDC-ER
BLOT: ANTI-p85
MAbl2CA5 - - EPO - + - PEPTIDE - - - - + -
+ + + + + + + - +
-
105- *@54
69-
For personal use only. by guest on July 14, 2011. bloodjournal.hematologylibrary.orgFrom

A FDC-HER C FDC-ER
aPP2P
aOri
0 5 5 10 15 20min 1
(-1 12CA5
B
3 x
bQ a'
(v M A
v) 0 g
d 4 z
a
n
v) 0 I
~PPP
aOr i
0 5 10 15 20
D
3 1 x
x a' cv
rc1 A 0 t v) 0 Z - i E I a v) 0 I a
EPO EXPOSURE, MINUTES EPO EXPOSURE, MINUTES
Fig 4. EPO-induced formation of phosphatidyl inositol =P-phosphate in FDC-HER cells. Factor-deprived FDC-HER cells (A and B) and control FDC-ER cells (C and D) were exposed to EPO (20 U/mL) for increasing intervals, and receptor-associated p85/P13-K was isolated by immunoprecipitation with MoAb 12CA5 or antiserum to EPO receptor peptides, respectively. P13-K activity then was assayed via incuba- tion with phosphatidylinositol and [yJ2P]ATP. Phosphatidyl inositol 32P-phosphate was analyzed by TLC (see Materials and Methods) and assayed quantitatively by laser densitometry.
For personal use only. by guest on July 14, 2011. bloodjournal.hematologylibrary.orgFrom
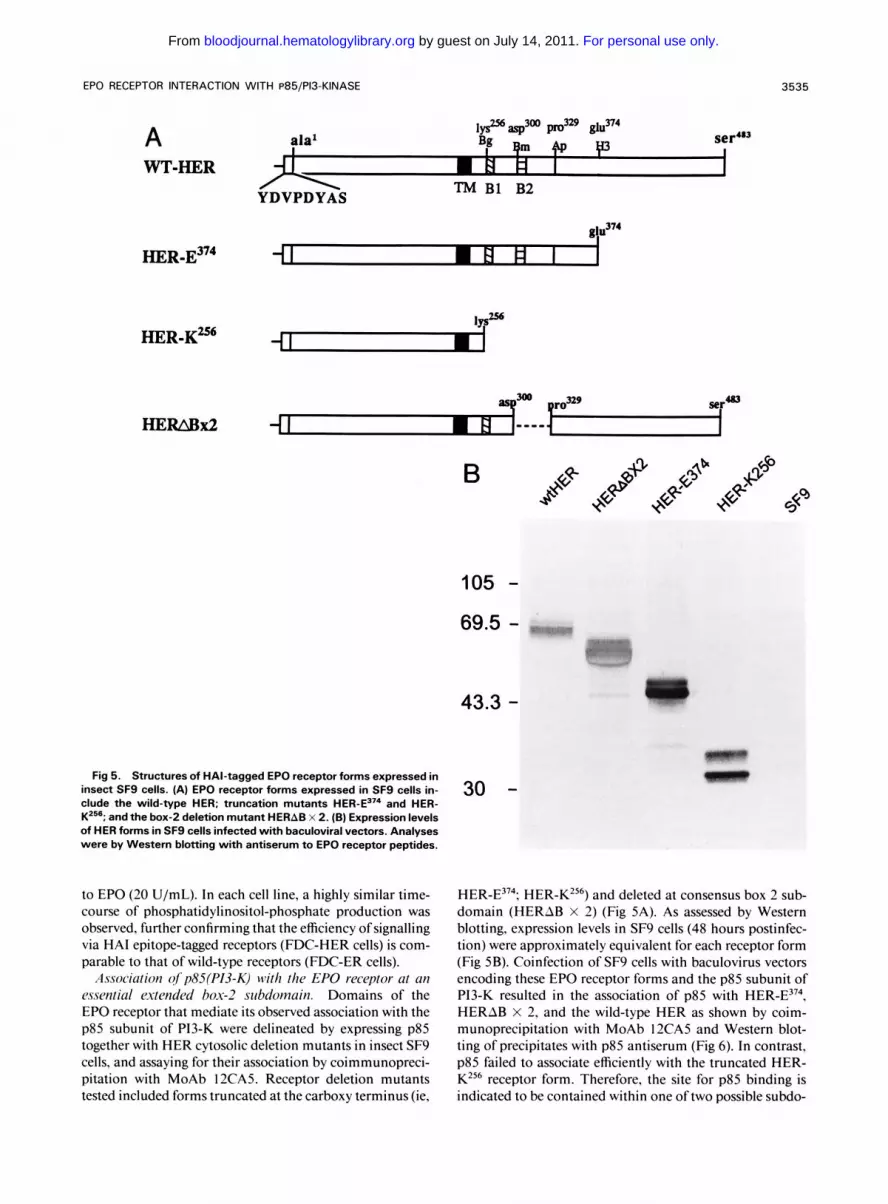
EPO RECEPTOR INTERACTION WITH P85/P13-KINASE 3535
P sera' lyS2Tbasp300 pro329 glu37' A ala1 qm +p
WT-HER W H H I 1 TM B1 B2 YDVPDYAS
HER-E374 i I . E l # I
105 - 69.5 -
Fig 5. Structures of HAl-tagged EPO receptor forms expressed in insect SFS cells. (A) EPO receptor forms expressed in SF9 cells in- clude the wild-type HER; truncation mutants and HER- K2"; and the box-2 deletion mutant HERAB x 2. (B) Expression levels of HER forms in SF9 cells infected with baculoviral vectors. Analyses were by Westem blotting with antiserum to EPO receptor peptides.
to EPO (20 U/mL). In each cell line, a highly similar time- course of phosphatidylinositol-phosphate production was observed, further confirming that the efficiency of signalling via HA1 epitope-tagged receptors (FDC-HER cells) is com- parable to that of wild-type receptors (FDC-ER cells).
Association ofp85(P13-K) with the EPO receptor at an essential mended box-2 subdomain. Domains of the EPO receptor that mediate its observed association with the p85 subunit of P13-K were delineated by expressing p85 together with HER cytosolic deletion mutants in insect SF9 cells, and assaying for their association by coimmunopreci- pitation with MoAb 12CA5. Receptor deletion mutants tested included forms truncated at the carboxy terminus (ie,
43.3 -
30 -
HER-E374; HER-KZ56) and deleted at consensus box 2 sub- domain (HERAB X 2) (Fig 5A). As assessed by Western blotting, expression levels in SF9 cells (48 hours postinfec- tion) were approximately equivalent for each receptor form (Fig 5B). Coinfection of SF9 cells with baculovirus vectors encoding these EPO receptor forms and the p85 subunit of PI3-K resulted in the association of p85 with HER-E374, HERAB X 2, and the wild-type HER as shown by coim- munoprecipitation with MoAb 12CA5 and Western blot- ting of precipitates with p85 antiserum (Fig 6). In contrast, p85 failed to associate efficiently with the truncated HER- K256 receptor form. Therefore, the site for p85 binding is indicated to be contained within one oftwo possible subdo-
For personal use only. by guest on July 14, 2011. bloodjournal.hematologylibrary.orgFrom

3536 HE ET AL
BLOT: ANTI-p85 IP: MAb12CA5 Fig 6. Coimmunoprecipitation of baculovirus-
expressed p85 and' HAi-tagged EPO receptor BV-HER: WT WT BX2 BX2 K256 K256 E374 E374 forms. p85 (P13-K) and HER forms (see Fig 5) were BV-p85: - + - + - + - + coexpressed in SF9 cells and receptors were im-
p85D -
mains ofthe EPO receptor: the box I-containing membrane proximal subdomain K256-D300: or the extended box 2 sub- domain P329-E374. For several PTK-encoding receptors, in- cluding EGF and PDGF, association with p85 previously has been shown to occur via amino and carboxy terminal SH2 domains within ~85.'~.'' Although no canonical sites for SH2 binding occur in the EPO receptor, a Y343LVL mo- tif exists within the extended box-2 subdomain. This motif corresponds closely in sequence to SH2 binding sites identi- fied previously in ErbB2 (YLGL)39 and basic FGF receptors (YLEL),40 and therefore is suggested to comprise a site for direct association ofthe EPO receptor with p85. Also, West- ern blotting of baculovirus-expressed receptor forms (in- cluding HER-E374) with the antiphosphotyrosine antibody PY20 indicated that Y343 is phosphorylated in SF9 cells. Alternatively, it is possible that association of p85 with the EPO receptor might occur through a domain of p85 that is distinct from the terminal SH2 domains or may depend on an intermediating accessory factor.
DISCUSSION
Using an active, epitope-tagged form of the murine EPO receptor, the present study directly demonstrates the associa- tion of the p85 regulatory subunit of P13-K with this recep- tor in FDC-HER cells. Moreover, this association is ob- served to occur in factor-deprived cells: no increases in levels of receptor-associated p85 are observed on exposure to EPO: and no modulation in overall levels of phosphory- lation of receptor-associated p85 is apparent. However. EPO is shown to rapidly induce the formation ofphosphati- dylinositol '*P-phosphate in FDC-HER and FDC-ER cells. These observations show convincingly that the association of p85 with the EPO receptor in FDC-HER cells is constitu- tive and raise important questions regarding mechanisms of PI3-K activation and the possible role of PI3-K in EPO- stimulated cell proliferation.
With regard to mechanisms for activation of P13-K in FDC-HER cells, by analogy, to P13-K activation in PTK- encoding receptor systems (eg, EGF and PDGF recep t~rs) . '~ .~ ' these are thought to include ligand-induced bind- ing of p85 via SH2 motifs to phosphotyrosine residues within activated receptors. Subsequently, the pl I O catalytic subunit42 of P13-K then may gain access to membrane phos- phatidylinositol s~bstrates .~~ In contrast, in the EPO recep- tor system, p85 presently is shown to preassemble constitu-
munoprecipitated from Triton X-1 00 lysates using MoAb 12CA5. p85 that bound to these receptor forms was assayed by coimmunoprecipitation with MoAb 12CA5 and Western blotting of precipitates using specific antiserum to p85. Controls included cells infected only with baculoviruses encoding HER forms ([ - 1 lanes v SF9-p85). The position of p85 is indexed in the left margin (arrow). Mr stan- dard (Mr X 10') are indexed in the right margin.
-105
-69'5
tively with receptor in the absence of ligand. thus indicating that alternative mechanisms may exist for EPO-induced P13-K activation. Most recently. EPO has been shown to activate JAK2" (a directly associated Janus-type PTK)24 and has been shown previously to stimulate 'the tyrosine phosphorylation of gap120,'7 raf-1.I6 pp100,'5*2G22 ~ ~ 1 3 0 . ' ~ and the EPO receptor per se." Consequently, one possible mechanism for P13-K activation in FDC-HER cells is the direct, induced (tyrosine) phosphorylation of receptor-asso- ciated p85 (plIO/PI3-K). Previously, this has been pro- posed as a possible mechanism for P13-K activation in IL-2 receptor system4' and presently is being assessed in the EPO receptor system. Alternatively. because EPO-induced tyro- sine phosphorylation of the EPO receptor is primarily at distal carboxyl-terminal residues," induced receptor phos- phorylation also might contribute allosterically to p85/ pl IO/P13-K activation. Consistent with this model is the recent observation that activated p85/P13-K complexes can be retrieved in association with tyrosine phosphorylated forms of the EPO receptor from Ba/F3 cells using biotiny- lated EPO." However, in this latter study, immunoprecipi- tation of EPO receptor complexes was not possible using available antisera: therefore, preassembly of receptor with p85 could not be addressed. Interestingly. the issue of li- gand-induced versus apparent constitutive assembly of EPO receptor complexes also may extend to a consideration of nonreceptor PTK activation. As mentioned above, the EPO receptor recently has been shown in vitro to associate in the absence of ligand with the cytosolic PTK, JAK2.2' Using the HAI-tagged murine EPO receptor and antiserum to JAK2, we likewise have observed receptor-JAK2 asso- ciation (ie, coimmunoprecipitation) in FDC-HER cells (T-C.H. and D.M.W. unpublished data). Notably, JAKZ also has been shown to associate with the receptor for growth hormone (GH)?' However, in the GH system, li- gand binding apparently is required both for GH receptor- JAK2 association as well as for JAKZ activation. Thus, these observations, together with the present work, under- line the likehood that discrete molecular mechanisms and/ or cofactors for P13-K regulation may exist among various growth factor receptor systems.
Using recombinant factors expressed in SF9 cells, a puta- tive EPO receptor cytosolic subdomain for p85 binding (ie, P329-E374) also presently is delineated. Located within this domain, and specifically within an extended box 2 region
For personal use only. by guest on July 14, 2011. bloodjournal.hematologylibrary.orgFrom

EPO RECEPTOR INTERACTION WITH P85/P13-KINASE 3537
(which has been shown previously to be essential for EPO induced mitogenesis),'0,'' is a single putative motif for SH2 binding (ie, Y343LVL). Because in essentially all systems de- fined to date, binding of p85 to growth factor receptor com- plexes is via amino- and/or carboxyl-terminal p85 SH2 do- mains, the Y343LVL motif ofthe EPO receptor is considered to comprise a likely site for p85 association. This sequence approximates SH2 binding motifs that occur within the EGF3' and basic FGF receptors,40 for example, with variabil- ity observed within third position (V, D, and G, respec- tively). However, in this context, it is noteworthy that no consensus motifs for SH2 binding occur within either the EPO receptor or, for example, the related 0-receptor for IL-2 that likewise has been shown to bind p85 and to medi- ate P13-K activation.43 Thus, as further shown by the recent identification of a novel motif for p85 binding in the recep- tor for hepatocyte growth as yet incompletely de- fined diversity apparently exists within functional motifs for p85 (SH2) binding. In the EPO receptor system, functional assessment of the YLVL motif as a site for p85 association will derive from the ongoing construction and expression of point-mutated receptor forms in permissive, factor-depen- dent hematopoietic cell lines.
P13-K is known to catalyze (via a recently cloned pl10 the phosphorylation of the glycosyl moiety of
phosphatidylinositol at the D-3 position. Although the ac- tion of derived phosphoinositol lipids in signalling is pres- ently undefined, a role for P13-K in proliferative signalling nonetheless is indicated by recent studies in several sys- t e m ~ . ~ ~ As a recent and important example, in the PDGF system receptor forms that activate PI3-K yet fail to activate PLC-7 and GAP/ras nonetheless efficiently support ligand- dependent mitogenesis.47 Based on this observation, it is of significant interest to consider whether distinct receptor do- mains likewise might be involved in PI3-K versus GAP/ras activation; and whether EPO-activation of P13-K might de- pend on activation of a nonreceptor PTK (ie, JAK2). The present work establishes an immediately useful model for addressing these specific questions. Ligand-dependent acti- vation of p85(PI3-K) also recently has been shown to be activated in three alternative class I cytokine receptor sys- tems: the IL-7 receptor in pre-B cells48; the IL-4 receptor in FDC-P2 cells49; and the IL-2 p receptor in Baf-BO3 cells.50 In FDC-P2 cells, an association of the IL-4 receptor with p85(PI3-K) was shown. Also, in Baf-BO3 cells, IL-2-depen- dent activation of P13-K was deficient for an IL-20 receptor deletional mutant lacking a serine-rich domain that is essen- tial for mitogenic signalling, yet apparently dispensable for nonreceptor PTK a~tivation.~' These findings, together with the present work, underline the importance of further defining molecular mechanisms for PI3-K activation via the EPO receptor and related class I cytokine receptor systems, including the possible requirement in this pathway for acti- vated, nonreceptor PTKs (eg, JAK- 1 and JAK-2).24
ACKNOWLEDGMENT
The authors thank Amgen Biologicals for the generous provision of recombinant human EPO used in these studies and Mardi Hock-
REFERENCES
I . Gregory CL, Eaves A C Human marrow cells capable of eryth- ropoietic differentiation in vitor: Definition of three erythroid col- ony responses. Blood 49355, 1977
2. Koury MJ, Boudurant MC: Maintenance by erythropoietin of viability and maturation of murine erythroid precursor cells. J Cell Physiol 137:65, 1988
3. Dai CH, Krantz SB, Zsebo KM: Human burst-forming units- erythroid need direct interaction with stem cell factor for further development. Blood 78:2493, 199 1
4. DAndrea AD, Lodish HF, Wong GG: Expression cloning of the murine erythropoietin receptor. Cell 57:277, 1989
5. Jones SS, DAndrea AD, Haines LL, Wong GG: Human eryth- ropoietin receptor: Cloning, expression, and biological characteriza- tion. Blood 76:31, 1990
6. Bazan J F A novel family of growth factor receptors: A com- mon binding domain in the growth hormone, prolactin, erythropoi- etin, and IL-6 receptors, and the p75 IL-2 receptor p chain. Bio- chem Biophys Res Commun 164:788, 1989
7. Bazan J F Structural design and molecular evolution of a cy- tokine receptor superfamily. Proc Natl Acad Sci USA 87:6934, 1990
8. Miura 0, Cleveland JL, Ihle JN: Inactivation of erythropoie- tin receptor function by point mutations in a region having homol- ogy with other cytokine receptors. Mol Cell Biol 13:1788, 1993
9. DAndrea AD, Fasman GD, Lodish HF: Erythropoietin re- ceptor and interleukin-2 receptor p chain: A new receptor family. Cell 56:1023, 1989
10. Quelle DE, Wojchowski DM: Localized cytosolic domains of the erythropoietin receptor regulate growth signaling and down- modulate responsiveness to granulocyte-marcophage colony-stimu- lating factor. Proc Natl Acad Sci USA 88:480 I , 199 I
11. Chiba T, Kishi A, Sugiyama M, Amanuma H, Machide M, Magata Y, Todokoro K: Functionally essential cytoplasmic do- main of the erythropoietin receptor. Biochem Biophys Res Com- mun 186:1236, 1992
12. Wojchowski DM, He T-C: Signal transduction in theerythro- poietin receptor system. Stem Cells 11:38 1, 1993
13. DAndrea AD, Yoshimura A, Youssoufia H, Zon LI, Koo J-W, Lodish H F The cytoplasmic of the erythropoietin receptor contains nonoverlapping positive and negative growth-regulatory domains. Mol Cell Biol 1 1 : 1980, 199 1
14. De La Chapelle A, Trakelin A-L, Juvonen E: Truncated erythropoietin receptor causes dominantly inherited benign human erythrocytosis. Proc Natl Acad Sci USA 90:4495, 1993
15. Quelle FW, Wojchowski DM: Proliferative action oferythro- poietin is associated with rapid protein tyrosine phosphorylation in responsive B6SUt.EP cells. J Biol Chem 266:609, 1991
16. Carroll MP, Spivak JL, McMahon M, Weich N, Rapp UF, May S: Erythropoietin induces raf- 1 activation and raf- 1 is required for erythropoietin-mediated proliferation. J Biol Chem 266: 14964, 1991
17. Torti M, Marti KB, Altshuler D, Yamamoto K, Lapetina EG: Erythropoietin induces p2 lras activation and p12OGAP tyro- sine phosphorylation in human erythroleukemia cells. J Biol Chem 26723293, 1992
18. Miura 0, DAndrea A, Kabat D, Ihle J N Induction of tyro- sine phosphorylation by erythropoitin receptor correlates with mi- togenesis. Mol Cell Biol l 1:4895, 1991
19. Yoshimura A, Lodish H F In vitro phosphorylation of the erythropoietin receptor and an associated protein, ppl30. Mol Cell Biol 12:706, 1992
20. Quelle FW, Quelle DE, Wojchowski DM: Interleukin 3, enberry for expert assistance in assembling this manuscript. granulocyte-marcophage colony-stimulating factor, and trans-
For personal use only. by guest on July 14, 2011. bloodjournal.hematologylibrary.orgFrom

3538 HE ET AL
fected erythropoietin receptors mediate tyrosine phosphorylation of a common cytosolic protein (pp100) in FDC-ER cells. J Biol Chem 267:17055, 1992
2 1. Linnekin D, Evans GA, DAndrea A, Farrar W: Association of the erythropoietin receptor with protein tyrosine kinase activity. Proc Natl Acad Sci USA 89:6237, 1992
22. Quelle FW, Egerton M, Samelson LE, Wojchowski DM: Cy- tokine-induced phosphorylation ofpplOO in FDC-PI cellsisat tyro- sine residues. Biochem Biophys Res Commun 188: 1040, 1992
23. Witthuhn BA, Quelle FW, Silennoinen 0, Yi T, Tang B, Miura 0, Ihle JN: JAK2 associates with the erythropoietin receptor and is tyrosine phosphorylated and activated following stimulation with erythropoietin. Cell 74:227, 1993
24. Wilks AF, Harpur AG, Kurban RR, Ralph SJ, Zurcher G, Ziemiecki A: Two novel protein-tyrosine kinases, each with a sec- ond phosphotransferase-related catalytic domain, define a new class of protein kinase. Mol Cell Biol 1 1:2057, 1991
25. Ullrich A, Schlessinger J: Signal transduction by receptors with tyrosine kinase activity. Cell 61:203, 1990
26. Valazquez L, Fellous M, Stark GR, Pellegrini A: A protein kinase in the interferon a//3 signaling pathway. Cell 70:3 13, 1992
27. Parker PJ, Waterfield MD: Phosphatidylinositol 3-kinase: A novel effector. Cell Growth Differ 3:747, 1992
28. Mayer BJ, Baltimore D Signalling through SH2 and SH3 domains. Trends Cell Biol 3:8, 1993
29. Dexter TM, Garland J, Scott D, Scolnick E, Metcalf D Growth of factor-dependent hemopoietic precursor cell lines. J Exp Med 152:1036, 1980
30. Ihle JN, Keller J, Greenberger JS, Henderson L, Yetter RA, Morse HC 111: Phenotypic characteristics of cell lines requiring in- terleukin 3 for growth. J Immunol 129:1377, 1982
3 I . Wilson IA, Niman HL, Houghten RA, Cherenson AR, Con- nolly ML, Lerner RA: The structure of an antigenic determinant in a protein. Cell 37:767, 1984
32. Quelle DE, Lynch KJ, Burkett-Smith RE, Weiss S, Whitford W, Wojchowski DM: Phosphorylable and epitope-tagged human erythropoietins: Utility and purification of native baculovirus-de- rived forms. Protein Expression Purification 3:46 I , 1992
33. Gout I, Dhand R, Panayotou G, Fry MJ, Holes I, Otsu M, Waterfield MD: Expression and characterization of the p85 subunit of the phosphatidylinositol 3-kinase complex and a related p850 protein by using the baculovirus expression system. Biochem J 288:395, 1992
34. Li J-P, DAndrea AD, Lodish HF, Baltimore D: Activation of cell growth by binding of Friend spleen focus-forming virus gp55 glycoprotein to the erythropoietin receptor. Nature 343:762, 1990
35. Summers MD, Smith GE: A Manual of Methods for Baculo- virus Vectors and Insect Cell Culture Procedures (ed 2). Texas Agri- cultural Experiment Station, TX, 1988, p 5
36. Auger KR, Serunian LA, Soltoff SP, Libby P, Cantley LC: PDGF-dependent tyrosine phosphorylation stimulates production of novel polyphosphoinositides in intact cells. Cell 57: 167, 1989
37. Mcglade CJ, Ellis C, Reedijk M, Anderson D. Mbamalu G, Reith AD, Panayotou G, End P, Bernstein A, Kazlauskas A, Water-
field MD, Pawson T: SH2 domains ofthe p85a subunit ofphospha- tidylinositol 3-kinase regulate binding to growth factor receptors. Mol Cell Biol 12:99 I , 1992
38. Hu P, Margolis B, Skolnik EY, Lammers R, Ullrich A, Schlessinger J: Interaction of phosphatidylinositol 3-kinase-asso- ciated p85 with epidermal growth factor and platelet-derived growth factor receptors. Mol Cell Biol I2:98 I , 1992
39. Zhou S, Shoelson SE, Chaudhuri M, Gish G, Pawson T, Haser W, King F, Roberts T, Ratnofsky S, Lechleider RJ, Nee1 BG, Birge RB, Chou MM, Hanafusa H, Schallhausen B, Cantley LC: SH2 domains recognize specific phosphopeptide sequences. Cell 72:767, 1993
40. Mohammadi M, Dionne CA, Li W, Spivak T, Honegger AM, Jaye M, Schlessinger J: Point mutation in FGF receptor elimi- nates phosphatidylinositol hydrolysis without affecting mitogene- sis. Nature 356:68 1, 1992
4 1. Ostu M, Hiles I, Gout 1, Fry MJ, Ruiz-Larrea F, Panayotou G, Thomposon A, Dhand R, Hsuan J, Totty N, Smith AD, Moryan SJ, Courtneidge SA, Parker PJ, Waterfield M D Characterization of two related p85 kd proteins containing SH2 and SH3 domains that bind to receptor tyrosine kinases and polyoma virus middle T anti- ger1/pp60'-'~ complexes, and associate with phosphatidylinositol 3-kinase. Cell 65:91, 1991
42. Hiles ID, Otsu M, Volinia S, Fry MJ, Gout I, Dhand R, Panayotou G, Totty NF, Hsuan J, Courtneidge SA, Parker PJ, Wa- terfield MD: Phosphatidylinositol 3-kinase: Structure and expres- sion of the I IO kd catalytic subunit. Cell 70:419, 1992
43. Merida I, Diez E, Gaulton GN: IL-2 binding activates a tyro- sine-phosphorylated phosphatidylinositol 3-kinase. J Immunol 147:2202, 1991
44. Damen JE, Mui A, Puil L, Pawson T, Krystal G: Phosphati- dylinositol 3-kinase associates, via its SH2 domains, with the acti- vated erythropoietin receptor. Blood 8 1:3204, I993
45. Argetsinger LS, Campell GS. Yang X, Silvennoinen 0, Ihle JN, Carter-Su C: Identification of JAK2 as a growth hormone re- ceptor-associated tyrosine kinase. Cell 74:237, 1993
46. Ponzetto C, Bardelli A, Maina F, Longati P, Panayotou G, Dhand R, Waterfield MD, Comoglio PM: A novel recognition mo- tif for phosphatidylinositol 3-kinase binding mediates its associa- tion with the hepatocyte growth factor/scatter factor receptor. Mol Cell Biol 13:4600. 1993
47. Vallas M, Kazlauskas A: Phospholipase C-y 1 and phosphati- dyl inositol 3-kinase are the down-stream mediators of the PDGF receptor's mitogenic signal. Cell 73:32 I , 1993
48. Dadi HK, Roifman CM: Interleukin-7 receptor mediates the activation of phosphatidylinositol 3-kinase in human B-cell precur- sors. Biochem Biophys Res Commun 192:459, 1993
49. Wang L-M, Keegan AD, Paul WE, Heidaran MD, Gutkind JS, Pierce JH: IL-4 activates a distinct signal transduction cascade from IL-3 in factor-dependent myeloid cells. EMBO J 11:4899, 1992
50. Merida 1, Williamson P, Kuziel WA, Greene WC, Gaulton GN: The serine-rich cytoplasmic domain of the interleukin-2 re- ceptor beta chain is essential for interleukine-2-dependent tyrosine protein kinase and phosphatidyl inositol 3-kinase activation. J Biol Chem 268:6765, 1993
For personal use only. by guest on July 14, 2011. bloodjournal.hematologylibrary.orgFrom




















