
Intermolecular Interactions of the p85alpha Regulatory Subunit of Phosphatidylinositol 3-Kinase
12
Waterfield Panayotou, Paul C. Driscoll and Michael D. Das, Matthew J. Bottomley, George Ailsa G. Harpur, Meredith J. Layton, Pamela 3-Kinase Regulatory Subunit of Phosphatidylinositol α Intermolecular Interactions of the p85 STRUCTURE: PROTEIN CHEMISTRY AND doi: 10.1074/jbc.274.18.12323 1999, 274:12323-12332. J. Biol. Chem. http://www.jbc.org/content/274/18/12323 Access the most updated version of this article at . JBC Affinity Sites Find articles, minireviews, Reflections and Classics on similar topics on the Alerts: When a correction for this article is posted • When this article is cited • to choose from all of JBC's e-mail alerts Click here http://www.jbc.org/content/274/18/12323.full.html#ref-list-1 This article cites 41 references, 16 of which can be accessed free at at University of Melbourne (CAUL) on June 10, 2013 http://www.jbc.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Intermolecular Interactions of the p85alpha Regulatory Subunit of Phosphatidylinositol 3-Kinase

WaterfieldPanayotou, Paul C. Driscoll and Michael D.Das, Matthew J. Bottomley, George Ailsa G. Harpur, Meredith J. Layton, Pamela 3-KinaseRegulatory Subunit of Phosphatidylinositol
αIntermolecular Interactions of the p85STRUCTURE:PROTEIN CHEMISTRY AND
doi: 10.1074/jbc.274.18.123231999, 274:12323-12332.J. Biol. Chem.
http://www.jbc.org/content/274/18/12323Access the most updated version of this article at
.JBC Affinity SitesFind articles, minireviews, Reflections and Classics on similar topics on the
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/274/18/12323.full.html#ref-list-1
This article cites 41 references, 16 of which can be accessed free at
at University of Melbourne (CAUL) on June 10, 2013http://www.jbc.org/Downloaded from

Intermolecular Interactions of the p85a Regulatory Subunit ofPhosphatidylinositol 3-Kinase*
(Received for publication, January 25, 1999)
Ailsa G. Harpur‡§¶, Meredith J. Layton‡¶i, Pamela Das‡, Matthew J. Bottomley**‡‡,George Panayotou§§, Paul C. Driscoll**, and Michael D. Waterfield‡**
From the ‡Ludwig Institute for Cancer Research, 91 Riding House St., London W1P 8BT, the **Department ofBiochemistry and Molecular Biology, University College London, Gower St., London WC1E 6BT, United Kingdom,and the §§Institute of Molecular Oncology, Biomedical Science Research Center “A. Fleming,” Vari 16672, Greece
The regulatory subunit of phosphatidylinositol 3-ki-nase, p85, contains a number of well defined domainsinvolved in protein-protein interactions, including anSH3 domain and two SH2 domains. In order to investi-gate in detail the nature of the interactions of thesedomains with each other and with other binding part-ners, a series of deletion and point mutants was con-structed, and their binding characteristics and appar-ent molecular masses under native conditions wereanalyzed. The SH3 domain and the first proline-richmotif bound each other, and variants of p85 containingthe SH3 and BH domains and the first proline-rich motifwere dimeric. Analysis of the apparent molecular massof the deletion mutants indicated that each of thesedomains contributed residues to the dimerization inter-face, and competition experiments revealed that therewere intermolecular SH3 domain-proline-rich motif in-teractions and BH-BH domain interactions mediatingdimerization of p85a both in vitro and in vivo. Binding ofSH2 domain ligands did not affect the dimeric state ofp85a. Recently, roles for the p85 subunit have been pos-tulated that do not involve the catalytic subunit, and ifp85 exists on its own we propose that it would bedimeric.
The Class IA phosphatidylinositol 3-kinases (PI3K)1 are het-erodimeric enzymes with a p110 catalytic subunit and a p85regulatory subunit (1). The p85 subunit is a multidomain pro-tein comprising an amino-terminal SH3 domain, a BCR homol-ogy (BH) domain which has homology to the GTPase activatingprotein domain of the Break-point Cluster Region protein (2)and Rho subfamily GTPases, and two SH2 domains separated
by an inter-SH2 domain through which p85 binds the catalyticsubunit (3). The BH domain is flanked by two proline-richmotifs. To date, five isoforms of p85 have been identified. p85ahas been cloned from bovine (4), human (5), and mouse (6)cDNA libraries, whereas only bovine p85b has been identified(4). Two splice variants of p85a, termed p55 and p50, have beenidentified in the human (7), the rat (8, 9), and the mouse (10).p55a lacks the SH3 and BH domains and the first proline-richmotif (PRM1) but retains the second proline-rich motif (PRM2)and has an amino-terminal extension of 34 amino acids. Inp50a, this extension comprises only 6 residues. To date, nosplice variants of p85b have been identified. A variant knownas p55g or p55PIK has been cloned from bovine2 and human (11)cDNA libraries and is homologous to p55a, but no higher mo-lecular mass isoforms of this protein have yet been identified.
PI3K has been implicated in a wide range of signaling path-ways including those regulating proliferation and cell migra-tion (12). The modular domains in p85a possess intrinsic sig-naling functions, in that they bind numerous intracellularligands and mediate the formation of multiprotein complexes.The proline-rich motifs bind the SH3 domains of the Src familytyrosine kinases, Lyn and Fyn (13), whereas the small G pro-teins Rac (14, 15) and Cdc42 (16) are potential PI3K regulatorsor effectors that bind PI3K, presumably via the BH domain.The SH2 domains of p85 bind phosphotyrosine (pY)-containingsequences from a range of receptor tyrosine kinases and dock-ing proteins such as insulin receptor substrate 1 (17–19).
The roles of the various isoforms of the adaptor subunits ofthe Class IA PI3Ks are as yet undefined. Some isoforms, espe-cially the truncated isoforms p55a and p50a, have been shownto have restricted tissue distributions (9) compared with the85-kDa isoforms. In addition, the truncated isoforms areclearly unable to interact with proline-rich motif- or SH3 do-main-containing proteins or with small G proteins, and it hasbeen shown that there is some selectivity in the recruitment ofp85 isoforms by receptor tyrosine kinases (20). However, it isnot yet understood whether the large number of isoforms ofboth subunits of PI3K represents a functional redundancy orreflects a form of signaling specificity. We have therefore un-dertaken a detailed study of the potential interactions of theindividual domains of the p85 protein in order to elucidate theirroles within the adaptor subunit as a whole.
MATERIALS AND METHODS
Peptides—Peptides were synthesized by Zinsser Analytic or Alta Bio-sciences and were of the following sequences: P1, KISPPTPKPRPPR-PLPVAPGPS; P2, WNERQQPAPALPPKPPKPT; Tyr-740/Tyr-751, GG-pYMDMSKDESVDpYVPML; Tyr-740, GGpYMDMSKDESVDYVPML;Tyr-751, GGYMDMSKDESVDpYVPML (where pY represents a phospho-tyrosine residue).
* The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby marked“advertisement” in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.
§ Current address and to whom correspondence should be addressed:Cell Biophysics Laboratory, Imperial Cancer Research Fund, 44 Lin-coln’s Inn Fields, London WC2A 3PX, United Kingdom. Tel.: 44-171-269-3054; Fax: 44-171-269-3094; E-mail: [email protected].
¶ Supported by C. J. Martin Fellowships from the National Healthand Medical Research Council, Australia. Both authors contributedequally to this work.
i Current address: The Ludwig Institute for Cancer Research, PORoyal Melbourne Hospital, Parkville, 3050, Australia.
‡‡ Supported by a Wellcome Trust Prize Studentship. Current ad-dress: Structural Biology Programme, EMBL, Meyerhofstrasse, 1, Post-fach 10.2209, Heidelberg D-69126, Germany.
1 The abbreviations used are: PI3K, phosphatidylinositol 3-kinases; BH,BCR homology; PRM, proline-rich motif; pY, phosphotyrosine; HP-SEC,high performance-size exclusion chromatography; DTT, dithiothreitol; GST,glutathione S-transferase; PAGE, polyacrylamide gel electrophoresis; SE-AUC, sedimentation equilibrium-analytical ultracentrifugation. 2 F. Pages and M. D. Waterfield, unpublished results.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 274, No. 18, Issue of April 30, pp. 12323–12332, 1999© 1999 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org 12323 at University of Melbourne (CAUL) on June 10, 2013http://www.jbc.org/Downloaded from

Construction, Expression, and Purification of Recombinant Pro-teins—Subcloning of cDNAs encoding p85a and p85b into the pAcC4baculovirus transfer vector has been previously described (21). p55g isthe bovine homologue of human p55PIK (11) and rat p55g (8). p49a is thebovine homologue of rat p50a (9) but with the unique 6-residue amino-terminal sequence deleted. cDNAs encoding p110a (22), p85aDSH3,p85aDBH, p85aDPRM1, p85aDPRM2, p85aDPRM1:PRM2, and p55g(Table I) were subcloned into the baculovirus transfer vector, pVL1393(Invitrogen). A sequence encoding a hexa-histidine tag was added to the39 end of the p49a cDNA, which was then subcloned into the baculovirustransfer vector, pBlueBac4 (Invitrogen). cDNAs encoding p85aSH3-BH-SH2, p85aSH3-BH, p85aSH3-PRM1, p85aSH3, p85acSH2, and p85aBH(Table I) were subcloned into the bacterial expression vector pGEX-2T(Amersham Pharmacia Biotech). Myc epitope-tagged (23) p85a, hexa-histidine-tagged p49a and p55g were also subcloned into pMT-SM (24)for expression in mammalian cells.
All cDNAs in pGEX-2T were expressed as glutathione S-transferasefusion proteins and purified by glutathione affinity chromatographyaccording to the manufacturer’s instructions. cDNAs in the baculovirustransfer plasmid pVL1393 were co-transfected into Sf9 cells with Bacu-loGold-linearized baculovirus DNA (PharMingen), whereas those in thetransfer plasmid pBlueBac4 were co-transfected into Sf9 cells withBac-N-Blue baculovirus DNA (Invitrogen). The recombinant baculovi-rus was plaque-purified and amplified as described previously (25).Exponentially growing Sf9 cells at a density of 1.5–2 3 106/ml wereinfected with recombinant baculoviruses at an multiplicity of infectionbetween 10 and 20 and harvested 2–3 days post-infection. SV40-trans-formed monkey kidney cells (Cos7) were maintained in Dulbecco’s mod-ified Eagle’s medium supplemented with 10% fetal calf serum, at 37 °Cand 10% CO2. Transient transfections were performed using DEAE-Dextran (Sigma). Cells (50% confluent) were transfected with 15 mg ofplasmid DNA per 15-cm tissue culture dish and harvested 48 hpost-transfection.
Sf9 cells expressing recombinant p85 variants were lysed in TritonX-100 lysis buffer (50 mM Tris, pH 8.0, 150 mM NaCl, 1% Triton X-100,5 mM dithiothreitol (DTT), and a range of protease inhibitors (50 mg/ml4-(2-aminoethyl)-benzenesulphenylfluoride hydrochloride, 16 mg/mlbenzamidine, 10 mg/ml 1,10-phenanthroline, 10 mg/ml aprotinin, 10mg/ml leupeptin, and 5 mg/ml pepstatin A)). p85 variants were purifiedby affinity chromatography using a phosphotyrosine-affinity columnthat was synthesized by immobilizing 2 mg of phosphotyrosine (Sigma)per ml of Actigel (Sterogene) according to the manufacturer’s instruc-tions. Briefly, up to 50 ml of cell lysate was applied to a 14 3 1-cmcolumn of phosphotyrosine-Actigel equilibrated in Buffer A (20 mM Tris,pH 8.0, containing 5 mM DTT) and eluted with 50 ml of Buffer Afollowed by 50 ml of Buffer A containing 150 mM NaCl and 50 ml ofBuffer A containing 2 M NaCl. Fractions of 2.5 ml were collected at aflow rate of 2.5 ml/min and exchanged into 3.5 ml of Buffer A usingpre-packed Sephadex G-25 columns (PD10, Amersham Pharmacia Bio-tech). If a second purification step was required, fractions containingp85 variants were pooled and applied to a 0.46 3 10-cm POROS 20 HQcolumn (Perspective Biosystems) equilibrated in Buffer A. Elution wascarried out using 17 ml of Buffer A followed by a 75-ml linear gradientto 1 M NaCl in the same buffer. Fractions of 2.5 ml were collected at aflow rate of 2.5 ml/min.
High Performance-Size Exclusion Chromatography (HP-SEC)—Transfected or wild type Cos7 cells were harvested by trypsinizationand Dounce-homogenized in hypotonic lysis buffer (5 mM Tris, pH 7.5,2.5 mM KCl, 1 mM DTT, 1 mM EDTA, 1 mM phenylmethylsufonylfluoride, 10 mM leupeptin, 10 mM pepstatin A, 1 mM 1,10-phenanthro-line, 1 mM sodium orthovanadate). The cytoplasmic fraction was clari-fied by centrifugation at 100,000 3 g at 4 °C for 45 min and filteredthrough a 0.22-mm membrane (Ultrafree-MC, Millipore). Aliquots of200 ml or less of molecular mass standards (Bio-Rad), recombinantproteins, or cell lysates were applied to a Superose 12/30 HR column(Amersham Pharmacia Biotech) equilibrated in TBS (20 mM Tris, pH8.0, 150 mM NaCl). Samples were eluted isocratically with TBS at a flowrate of 0.3 ml/min and 0.3- or 0.15-ml fractions were collected wherenecessary.
Sedimentation Equilibrium-Analytical Ulracentrifugation (SE-AUC)—All SE-AUC experiments were carried out using an Optima XL-A ana-lytical ultracentrifuge (Beckman) equipped with absorbance optics andan An60Ti rotor. Protein samples were buffer exchanged into SE-AUCbuffer (50 mM Tris, pH 7.4, 50 mM NaCl, 7 mM b-mercaptoethanol, and0.02% NaN3) using either pre-packed Sephadex G-25 columns (NAP5,Amersham Pharmacia Biotech) or by dialysis overnight. Three samplesof 110 ml of protein at approximately 0.6, 0.3 and 0.15 mg/ml wereanalyzed, and the reference cells contained 125 ml of SE-AUC buffer.
Experiments were performed at 4 °C at an optimized range of speedsbetween 4000 and 28,000 rpm. Equilibrium data were collected at 280nm in step scan mode with a radial increment of 0.001 cm between datapoints. Five readings were averaged each scan, from which a base-linescan taken at 360 nm was subtracted in order to correct for opticalimperfections. Readings were taken at 8-h intervals until no differencecould be detected between consecutive scans. The equilibrium distri-butions from three different loading concentrations and up to threerotor speeds were analyzed both individually and simultaneously usingthe Nonlin curve-fitting algorithm supplied with the ultracentrifuge(Beckman).
Binding Studies Using an Optical Biosensor—The procedure formeasuring interactions between domains and peptides using the BIA-core biosensor (Biacore AB) has been previously described (26). Briefly,biotinylated peptides were captured on immobilized avidin, and thensample was injected in running buffer (20 mM Hepes, pH 7.4, 150 mM
NaCl, 3.4 mM EDTA, 4 mM DTT, 0.005% Tween 20), and the response atequilibrium was recorded. For competition experiments, the response atequilibrium was compared with that obtained upon preincubation of theprotein or isolated domain solution with increasing amounts of freepeptide. In order to calculate IC50 values, the results were plotted asresonance units versus peptide concentration, and the curve obtainedwas fitted with the equation R 5 (Rmax/1 1 (C/IC50)P), where R is theresponse, Rmax is the response obtained in the absence of competitor, Cis the concentration of competitor, and P is the Hill coefficient.
Immunoprecipitations and Western Blotting—Transfected Cos7 cellswere lysed on ice in Nonidet P-40 lysis buffer (1% Nonidet P-40, 20 mM
Tris, pH 7.4, 50 mM NaCl, 50 mM NaF, 1 mM EDTA, 500 mM sodiumorthovanadate, 2 mM phenylmethylsulfonyl fluoride, 1 mg/ml leupeptin,and 1 mM aprotinin), and cell debris and nuclei were removed bycentrifugation at 10,000 3 g for 10 min at 4 °C.
Immunoprecipitations from Cos7 cell lysates were performed for 2 husing p85a monoclonal antibodies (U9, U13, and U14 (27)) or an anti-Myc epitope monoclonal antibody (23) immobilized using proteinG-Sepharose Fast Flow (Amersham Pharmacia Biotech) or a metal-chelate affinity matrix (Talon, CLONTECH). Precipitated proteinswere analyzed by sodium dodecyl sulfate-polyacrylamide gel electro-phoresis (SDS-PAGE), Western blotting onto polyvinylidene difluo-ride membrane (Gelman Sciences), probing with an appropriatemouse monoclonal antibody, and detection of bound horseradish perox-idase-conjugated goat anti-mouse antibody (Bio-Rad) using enhancedchemiluminescence (ECL, Amersham Pharmacia Biotech).
Protein in fractions from HP-SEC was precipitated using 10% tri-chloroacetic acid and analyzed by SDS-PAGE and Western blotting asdescribed above. Proteins were precipitated from Sf9 cell lysates usingGST fusion proteins captured on glutathione-Sepharose CL 4B (Amer-
TABLE IAmino acid sequence specifications for p85a deletion and
substitution mutantsThe residues derived from the amino acid sequence of full-length
p85a (1–742) that comprise each mutant are listed. Single amino acidsubstitutions are denoted as P96A, indicating that the proline at posi-tion 96 was mutated to arginine. At the bottom is a schematic repre-sentation of the domain structure of p85a. The positions of the desig-nated domain boundaries are numbered according to the sequence ofp85a.
Construct Amino acid specification
p85a 1–742p85aDSH3 86–742p85aDPRM1 1–742,P96A,P99Ap85aDPRM2 1–742,P308A,P311Ap85aDPRM1:PRM2 1–742,P96A,P99A,P308A,P311Ap85aDBH 1–133,303–742p49a 338–742p85aSH3-BH-SH2 1–446p85aSH3-BH 1–339p85aBH 105–302p85aSH3-PRM1 1–106p85aSH3 1–86p85a cSH2 614–724
Interactions of Regulatory Subunit of PI 3-Kinase12324
at University of Melbourne (CAUL) on June 10, 2013http://www.jbc.org/Downloaded from

sham Pharmacia Biotech) and analyzed for lipid kinase assay as de-scribed below.
Phosphatidylinositol 3-Kinase Assays—PI3K assays of glutathione-Sepharose CL 4B-precipitated proteins were carried out essentially asdescribed previously (28). Lipid kinase assays contained 2 mM MgCl2, 1mM ATP, 20 mCi of [g-32P]ATP, and 200 mg/ml phosphatidylinositol.Extracted phospholipids were analyzed by thin layer chromatographyin 65% 1-propanol, 0.7 M acetic acid, 50 mM phosphoric acid.
RESULTS
Interactions of the SH3 Domain and the Proline-rich Motifsof p85a—It has previously been reported that both proline-richmotifs in p85a conform to the consensus ligand for the SH3
domain of p85a (29). In order to determine which of the twoproline-rich motifs was the preferred ligand for the isolatedp85a SH3 domain (p85aSH3), we compared its ability to bindto peptides derived from the sequences of the first (P1) orsecond (P2) proline-rich motifs of p85a, using an optical bio-sensor (Fig. 1A). Binding of p85aSH3 to immobilized P1 wasobserved in this system, producing a response of greater than400 resonance units (Fig. 1A), whereas little or no p85aSH3injected at the same concentration bound to P2 (Fig. 1A), sug-gesting that P1 is the preferred ligand for p85aSH3.
In contrast, wild type p85a, which contains the SH3 domain,did not bind immobilized P1 to a significant extent (Fig. 1B).The small amount of binding observed was equivalent to thatobserved for binding of the same concentration of p85a to asurface on which P1 was not immobilized (data not shown). Amutant of p85a, in which two proline residues in PRM1 thathave been shown to be important for p85a SH3 domain binding(30) were changed to alanine (p85aDPRM1), was able to bindP1; however, a variant of p85a with mutations at the equiva-lent residues in PRM2 (p85aDPRM2) was not. A variant ofp85a in which both proline-rich motifs contained mutations(p85aDPRM1:PRM2) bound to P1 to a similar extent asp85aDPRM1 (Fig. 1B). Mutations in PRM1 therefore allowedp85aSH3 to bind exogenous P1 peptide, whereas the SH3 do-main in the context of wild type p85a could not bind to exoge-nous P1. Mutations in PRM2 did not affect binding to P1peptide, confirming that the first proline-rich motif was thepreferred ligand for the p85a SH3 domain. Thus, mutation ofkey residues in PRM1 of p85a not only affected this site butalso the binding characteristics of the SH3 domain. Thesemutations increased the ability of the SH3 domain to bindexogenous ligands, indicating that in wild type p85a the SH3domain interacts with PRM1 and that when this interaction isdisrupted the SH3 domain is free to bind exogenous peptide.
Similarly, deletion of the SH3 domain of p85a would beexpected to free PRM1 and allow it to bind exogenous SH3domains. When wild type p85a was co-expressed with the p110catalytic subunit in Sf9 cells, a p85aSH3 GST fusion protein(GST-p85aSH3) immobilized on glutathione-Sepharose CL4Bwas poorly able to co-precipitate PI3K lipid kinase activity fromSf9 cell lysates. However, a monoclonal antibody directedagainst the SH2 domain of p85a was able to immunoprecipitatePI3K activity; thus both p85a and p110a were expressed inthese cells. The same antibody was able to immunoprecipitatePI3K activity from lysates of Sf9 cells co-infected with p110a
and a mutant of p85a in which the SH3 domain had beendeleted (p85aDSH3). Immobilized GST-p85aSH3 was able toco-precipitate the p110a-p85aDSH3 complex to a much greaterextent than the p110a-p85a complex (Fig. 1C); thus deletion ofthe SH3 domain increased the ability of an exogenous SH3domain to bind PRM1, confirming that the SH3 domain andPRM1 are bound to each other in both p85a and the p110a-p85a complex.
Intermolecular Interactions of p85 Isoforms—Examination ofpurified, recombinant p85a and p85b by HP-SEC under nativeconditions showed that they had apparent molecular massesthat were approximately double (162 6 14 and 151 6 12 kDa,respectively) that expected from their amino acid sequences(Fig. 2, A and B). In contrast, the apparent molecular masses ofthe truncated p85a isoforms, p55g and p49a (82 6 4 kDa and52 6 2 kDa), were much closer to their predicted molecularmasses (55 and 49 kDa; Fig. 2, A and B).
There are several mechanisms by which a protein undernative conditions can have an apparent molecular mass differ-ent from that predicted from its amino acid sequence. HP-SECmeasures the hydrodynamic volume of a protein, which is de-
FIG. 1. Interactions of the p85a SH3 domain and the proline-rich motifs of p85a. A, comparison of binding of p85aSH3 to immo-bilized peptides corresponding to the first proline-rich motif, P1 (O), orthe second proline-rich motif, P2 (- - -), from p85a using an opticalbiosensor. B, comparison of binding of equal concentrations of p85a(–z z z–) and p85a variants with point mutations in the first,p85aDPRM1 (O), second, p85aDPRM2 (- - -) or both, p85aDPRM1:PRM2 (– –) proline-rich motifs to immobilized P1 peptide. C, precipita-tion of phosphatidylinositol kinase activity associated with p110a/p85aor p110a/p85aDSH3 from Sf9 cell lysates using GST-p85aSH3 or anti-bodies to p85a.
Interactions of Regulatory Subunit of PI 3-Kinase 12325
at University of Melbourne (CAUL) on June 10, 2013http://www.jbc.org/Downloaded from

fined as the spherical volume occupied by that protein as ittumbles rapidly in solution. Non-spherical proteins often elutewith an abnormally high apparent molecular mass comparedwith the globular proteins used to calibrate the column. Pro-teins can also interact nonspecifically with the column matrix,impeding their progress through the column and resulting in alater elution time, leading to an underestimate of the molecularmass of a protein. High apparent molecular masses may alsoresult from dimerization or oligomerization of the protein; thuswe investigated whether the higher than expected molecularmass of p85a was due to dimerization by determining themolecular masses of p85a and p49a under different conditions.
HP-SEC was carried out in buffers containing either 5 mM
DTT or 8 M urea in order to determine whether the putativep85a dimeric interaction could be disrupted. The apparentmolecular masses of p85a and p55g were unaltered in thepresence of 5 mM DTT (Fig. 2B), suggesting that dimerization ofp85a was not due to the presence of an intermolecular disulfidebond. In contrast, p85a had an apparent molecular mass of89 6 5 kDa in the presence of 8 M urea. High concentrations ofurea disrupt non-covalent, but not covalent, bonds, indicatingthat p85a forms a dimer via a non-covalent interaction. Theapparent molecular mass of p49a was relatively unaffected inthe presence of 8 M urea however (Fig. 2B), suggesting that it ismonomeric under native conditions.
Sedimentation Equilibrium-Analytical Ultracentrifugation(SE-AUC) was employed to determine the apparent molecularmasses of a number of p85a variants, as the equilibrium dis-tribution of a solute in a gravitational field is dependent on themolecular mass of the solute, but independent of its shape,whereas HP-SEC is dependent on both mass and shape. Anal-ysis of the equilibrium distribution of the concentration ofprotein with respect to radial position in the centrifuge dem-onstrated the occurrence of self-association in some samples.Self-association was defined as a lack of adherence to theLamm equation, which describes the distribution of ideal, non-associating solute particles in a gravitational field. The occur-rence of self-association manifests as a non-random distribu-tion of residuals to the fit of the experimental (absorbanceversus radius) data to a derivative of the Lamm equation.
The equilibrium distribution of p85a did not fit that of anideal, non-associating solute, as the residuals were non-random(Fig. 3D). The apparent molecular mass of p85a was dependenton protein concentration and ranged between 80 and 110 kDa(Fig. 3D), suggesting that p85a exists as an equilibrium ofmonomers and dimers under these conditions. In contrast,p49a was monomeric under the conditions examined. The re-siduals of the fit for an ideal, non-associating solute were dis-tributed randomly around zero, and p49a had an apparentmolecular mass of approximately 47 kDa (Fig. 3C). Thus, p85a,
FIG. 2. Analysis of naturally occur-ring p85a isoforms by HP-SEC. A,high performance size-exclusion chroma-tography of recombinant p85a (O), p85b(– –), p55g (- - -) or p49a(–zzz–). The Super-ose 12/30 column was equilibrated andrun in TBS at 0.3 ml/min. The retentiontimes of the molecular mass standardsare indicated (in kDa), as is the void vol-ume (Vo). B, summary of data from HP-SEC of recombinant p85 isoforms andp85a deletion mutants and a diagram-matic representation of the domains ineach p85 isoform is indicated. The pre-dicted molecular mass for each isoform (inkDa) was calculated from its amino acidsequence. Molecular masses determinedby HP-SEC were calculated from the re-tention time (min) of the maximum heightof the peak for each recombinant protein.The retention time was converted to anapparent molecular mass by comparisonto a plot of log molecular mass (kDa) ver-sus the retention time (min) of the maxi-mum height of the peak for a range ofstandard globular proteins chromato-graphed using the same buffer conditions.Apparent molecular masses are denotedas the mean of at least three measure-ments 6 1 S.D. Where only one measure-ment was taken, an approximate molecu-lar mass is indicated. For HP-SEC oftransfected Cos7 cytosolic extracts, arange of apparent molecular masses iscalculated by converting the fractionnumbers that are positive for the appro-priate band by Western blotting into re-tention time (min) and therefore into ap-parent molecular mass (kDa) as describedabove.
Interactions of Regulatory Subunit of PI 3-Kinase12326
at University of Melbourne (CAUL) on June 10, 2013http://www.jbc.org/Downloaded from

which was apparently dimeric by HP-SEC, fitted a model forself-association, although the equilibrium dissociation constantfor p85a dimer formation (estimated from the plot of apparentmolecular mass versus protein concentration; Fig. 3D) was inthe micromolar range, which is a lower affinity than that ex-pected for a protein that was determined to be constitutivelydimeric by HP-SEC. Given that p49a was shown to be mono-meric using both techniques, this suggested that the dimeriza-tion of p85 was mediated by domains that are not present inp49a.
An Intermolecular Interaction between the SH3 Domain andPRM1 Contributes to the Dimerization of p85a—The observa-tion that p85a and p85b were dimeric under native conditions,but p55g and p49a were not, suggested that the amino-termi-nal half of the higher molecular mass regulatory subunits wasinvolved in intermolecular interactions that caused them todimerize. Given that the SH3 domain and the first proline-richmotif reside in this region of the protein, and interact with eachother (Fig. 1), we investigated whether this interaction wasinvolved in the dimerization of p85a.
FIG. 3. Sedimentation equilibrium-analytical ultracentrifugation analy-sis of domains of p85a. The data (E) arefitted to a curve describing the concentra-tion distribution of single, ideal speciesusing the Nonlin curve-fitting algorithm(Beckman). A plot of the residuals for thisfit is shown in the upper panel. The de-pendence of apparent molecular mass onprotein concentration is shown in thelower panel for p85aBH (A), p85aSH3-PRM1 (B), p49a (C), and p85a (D).
Interactions of Regulatory Subunit of PI 3-Kinase 12327
at University of Melbourne (CAUL) on June 10, 2013http://www.jbc.org/Downloaded from
Indeed, expression of the amino-terminal half of p85a on itsown, either with or without one of the SH2 domains (p85aSH3-BH-SH2 and p85aSH3-BH; Fig. 2B), resulted in proteins withapparent molecular masses by HP-SEC (101 6 10 and 53 6 4kDa, respectively) that were still approximately double thosepredicted (50 and 38 kDa, respectively). This confirms that theamino-terminal half of p85a contains all the amino acid resi-dues required for dimerization.
When the SH3 domain of p85 was deleted, the apparentmolecular mass of the resulting protein (p85aDSH3) deter-mined by HP-SEC under native conditions was 139 6 7 kDa,which was approximately double that predicted from its aminoacid sequence (74 kDa) (Fig. 2B). This suggested that removalof the SH3 domain did not disrupt the dimer interface to asignificant extent. Similarly, mutation of two proline residues(Pro-96 and Pro-99) in the first proline-rich motif to alanine(p85aDPRM1, Fig. 2B) did not convert p85a to a monomericspecies. Deletion of the remaining domain within the amino-terminal portion of p85a, the BH domain (p85aDBH), also didnot disrupt dimerization, as its molecular mass by HP-SEC(145 6 11 kDa) was still approximately double that predicted(64 kDa). In contrast, deletion of both the SH3 and BH do-mains, as is the case for both p55g and p49a, had already beenshown to result in a monomeric protein. Therefore, the dimer-ization interface resides in the amino-terminal half of the mol-ecule, either in the SH3 or BH domains or the interveningregions.
A construct comprising just the SH3 domain and PRM1(p85aSH3-PRM1, Fig. 2B) had an apparent molecular mass byHP-SEC (23 6 2 kDa) approximately double that predicted (12kDa), suggesting it was dimeric. Like p85a, the equilibriumsedimentation characteristics of p85aSH3-PRM1 were consist-ent with that of a protein undergoing self-association (Fig. 3B).The residuals of the fit for an ideal, non-associating solute werenon-random, and the apparent molecular mass of p85aSH3-PRM1 was dependent on protein concentration and ranged
between 16 and 22 kDa, with an equilibrium dissociation con-stant for dimer formation in the micromolar range (Fig. 3B). Incontrast, the SH3 domain alone was unable to dimerize, as ithad a similar apparent molecular mass by HP-SEC to thatpredicted from its amino acid sequence (13 6 1 and 9.7 kDa,respectively).
The apparent molecular mass of the isolated BH domain wasdetermined by HP-SEC (32 6 2 kDa) and was approximately1.5 times that predicted (22 kDa). The p85aBH also appearedto self-associate by SE-AUC, although to a lesser degree thanfor p85aSH3-PRM1 (Fig. 3A). The estimated equilibrium dis-sociation constant for dimer formation was in the millimolarrange.
The interaction surface for dimerization therefore does notreside in a single domain of the amino terminus of p85a, asneither the SH3 nor BH domain alone was observed to dimerizefully. Moreover, deletion of either domain or mutation of PRM1did not generate monomeric p85a (Fig. 2B). The interactionsurface involved in dimerization must therefore involve resi-dues that are widely distributed in the primary structure ofp85a, so that no single domain self-associates to a significantdegree, but are positioned in three-dimensional space in thefolded protein so that they cooperate to form a single bindingsurface. In contrast, p85aSH3-PRM1 was able to self-associate,and the estimated equilibrium dissociation constants fordimerization of p85aSH3-PRM1 and p85a were similar, sug-gesting that SH3-PRM1 binding was intermolecular and con-tributed to dimerization. However, the contribution of the BHdomain to dimerization of full-length p85a did not allow us todiscriminate between an inter- or intramolecular SH3-PRM1interactions in the whole protein.
If the interaction between the p85a SH3 domain and the firstproline-rich motif was intermolecular and contributed todimerization, exogenous P1 peptide should compete for thisinteraction and result in the formation of monomeric p85a. Theaffinities for self-association as determined by SE-AUC were
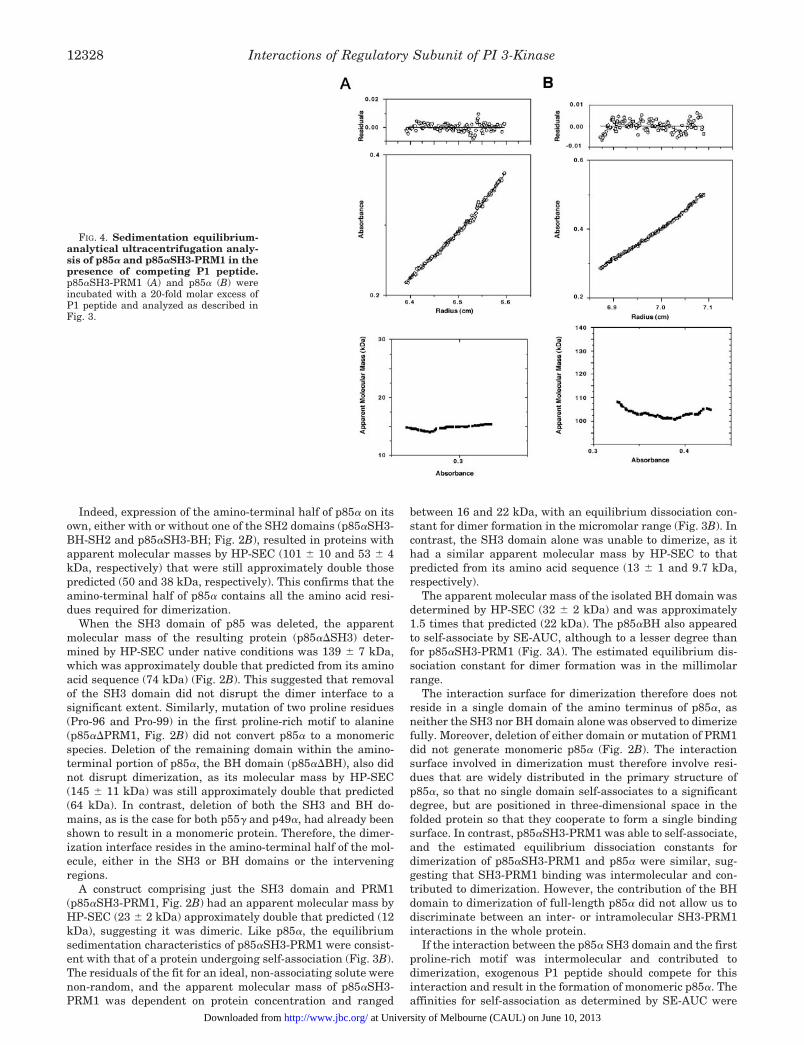
FIG. 4. Sedimentation equilibrium-analytical ultracentrifugation analy-sis of p85a and p85aSH3-PRM1 in thepresence of competing P1 peptide.p85aSH3-PRM1 (A) and p85a (B) wereincubated with a 20-fold molar excess ofP1 peptide and analyzed as described inFig. 3.
Interactions of Regulatory Subunit of PI 3-Kinase12328
at University of Melbourne (CAUL) on June 10, 2013http://www.jbc.org/Downloaded from

lower than the apparent affinities suggested by the molecularmasses of the p85a variants as determined by HP-SEC, thuscompetition experiments were performed using SE-AUC. In thepresence of a 20-fold molar excess of P1 peptide, both p85aSH3-PRM1 and wild type p85a behaved as ideal, non-associatingsolutes, and their apparent molecular masses were not depend-ent on protein concentration (Fig. 4, A and B), suggesting thatP1 peptide competed for binding in a site involved in dimerformation. However, the apparent molecular mass of p85abound to P1 peptide was greater than expected, suggesting thateither a small degree of self-association was mediated by theBH domain or that the high concentration of P1 in the analytesolution increased its viscosity, resulting in an artificially highestimation of apparent molecular mass. An intermolecular in-teraction between the p85a SH3 domain and the p85a PRM1therefore seems to contribute to p85a dimerization, althoughthe BH domain also contributes residues to this interface.
Dimerization of p85a in Vivo—In order to confirm whetherp85a dimerization observed in vitro also occurred in vivo, p85
isoforms were transfected into Cos7 cells and Western blotanalysis of HP-SEC fractions of whole cell lysates revealedimmunoreactive bands in fractions covering a relatively broadrange of molecular masses. p85a was detected in fractions36–38, which corresponded to a molecular mass range of 93–176 kDa (Figs. 5A and 2B). Although this was a broad range,the majority of the p85a eluted at a time that corresponded toa molecular mass greater than the monomeric molecular massfor p85a (83 kDa), and there was a population at a dimericmolecular mass (176 kDa). p85a therefore is dimeric both as ahomogeneous, purified recombinant protein and within thecontext of a cellular environment, when it is at a relatively lowconcentration and in a protein-rich environment, which areconditions that would be expected to reduce nonspecificself-association.
In contrast, p55g and p49a were detected in fractions thatcorresponded to molecular mass ranges of 40–61 and 26–49kDa, respectively, in this system (Figs. 5, B and C, and 2B). Thehighest molecular mass populations for both p55g and p49a
FIG. 5. p85a from transfected Cos7 cells behaves as a dimeric complex upon size-exclusion chromatography and co-precipitateswith p85aDBH but not with p49a. A–C, cytosolic extracts prepared from unstimulated Cos7 cells, transfected with p85a (A), p55g (B), or p49a(C), were subjected to size-exclusion chromatography. Aliquots of each fraction were analyzed by Western blotting with an appropriate mousemonoclonal antibody (lower strip of each panel A, B, and C). Absorbance at 220 nm (O) and the corresponding elution volumes of the molecularmass standards (kDa) are indicated, as is the void volume (Vo). The Superose-12 column was equilibrated and run in TBS at 0.3 ml/min, and 0.3-mlfractions were collected and analyzed by SDS-PAGE followed by Western blotting with the appropriate antibody. Unfractionated cytosolic extractswere analyzed in parallel (Lys) with the relevant antibody. D and E, Nonidet P-40 lysates were prepared from untreated Cos7 cells transfected withMyc epitope-tagged p85a and/or p85aDBH (D) or Myc epitope-tagged p85a and/or p49a (E). Myc epitope-tagged p85a was immunoprecipitatedusing an anti-Myc tag monoclonal antibody (9E10) and immunoblotted for co-precipitated p85aDBH or p49a using U14, a mouse monoclonalantibody directed against the SH2 domain of p85a which recognizes p85a, p85aDBH, or p49a. The relative migration positions of the variousisoforms on SDS-PAGE are indicated.
Interactions of Regulatory Subunit of PI 3-Kinase 12329
at University of Melbourne (CAUL) on June 10, 2013http://www.jbc.org/Downloaded from

were not greater than their predicted monomeric molecularmasses; therefore they are apparently monomeric within acellular context. Interestingly, p55g and p49a seemed to bindto other intracellular proteins from Cos7 cells and form highmolecular mass (.300 kDa) complexes. In addition, two vari-ants of p55g were observed, but the origin of these was notinvestigated further.
cDNAs encoding an amino-terminally Myc epitope-taggedversion of p85a, untagged p85aDBH, or hexa-histidine-taggedp49a were transiently transfected into Cos7 cells, alone or incombination. Precipitation with appropriate antibodies immo-bilized on protein G-Sepharose or affinity matrices showed thateach protein was expressed in Cos7 cells transfected with asingle cDNA (Fig. 5D, lanes 2 and 3 and Fig. 5E, lane 2). WhenMyc-tagged p85a and p85aDBH, a deletion mutant of p85 thatstill had the ability to dimerize in vitro (Fig. 2B), were co-transfected, immunoprecipitation with a monoclonal antibodydirected against the Myc epitope was able to co-immunopre-cipitate untagged p85aDBH (Fig. 5D, lane 1). The difference inmolecular mass between these two forms of p85a allowed themto be resolved by SDS-PAGE. In contrast, p49a, a deletionmutant of p85 that did not have the ability to dimerize in vitro(Fig. 2B), was unable to co-immunoprecipitate with Myc-taggedp85a (Fig. 5E, lane 1); therefore, forms of p85a that have theability to dimerize in vitro can form mixed heterodimers invivo, whereas monomeric forms cannot.
Binding of Phosphopeptides to the SH2 Domains Does NotAffect the Oligomerization States of p85a Variants—Binding ofa diphosphotyrosine-containing peptide that mimics an acti-vated platelet-derived growth factor b-R (Tyr-740/Tyr-751) tothe SH2 domains of p85a affects the oligomerization state of
the p110a-p85a complex3; therefore, we investigated the effectof Tyr-740/Tyr-751 binding on dimerization of p85a isoforms.The IC50 for competition of either Tyr-740/Tyr-751 or peptideswith the same amino acid sequence but with only one phospho-tyrosine residue (Tyr-740 or Tyr-751) with p85 variants forbinding to immobilized Tyr-751 was compared. The IC50 for thecompetition of Tyr-740, Tyr-751, or Tyr-740/Tyr-751 with theisolated carboxyl-terminal SH2 domain of p85a (p85a cSH2) forbinding to immobilized Tyr-751 was similar, although Tyr-740had a slightly lower affinity for p85a cSH2 compared with theother peptides (Fig. 6A). The binding characteristics of p85aand p49a in this system were very similar (Fig. 6, B and C).Both p85a and p49a had lower IC50 values for the competitionof Tyr-740/Tyr-751 for binding to Tyr-751 compared with eithermono-phosphopeptide, suggesting that both SH2 domains en-gage both phosphotyrosine residues within this peptide (Fig. 6,B and C).
The apparent molecular masses of p85a and p49a were de-termined at a range of concentrations by HP-SEC (Fig. 6D) andagreed with those previously described (Fig. 2B). There ap-peared to be a slight increase in apparent molecular mass ofp85a with increasing protein concentration (Fig. 6D), support-ing the idea that p85a can undergo self-association, whereasp49a cannot. Although Tyr-740/751 was shown to bind p85a orp49a with high affinity (Fig. 6, B and C), addition of Tyr-740/751 did not affect the apparent molecular mass of either p85aor p49a, regardless of protein concentration (Fig. 6D); therefore
3 Layton, M. J., Harpur, A. G., Panayotou, G., Bastiaens, P. I. H., andWaterfield, M. D. (1998) J. Biol. Chem. 273, 33379–33385.
FIG. 6. The role tyrosine phosphopeptide-binding in the dimerization of p85a. Inhibition of p85a cSH2 (A), p49a (B), or p85a (C) bindingto immobilized Tyr-751 by increasing concentrations of Tyr-740 (‚), Tyr-751 (E), and Tyr-740/Tyr-751 (●). D, apparent molecular mass of p85a(circles) and p49a (triangles) at a range of concentrations in the absence (open symbols) or presence (closed symbols) of a constant 10-fold molarexcess of Tyr-740/Tyr-751. Molecular masses were determined by HP-SEC and were calculated from the retention time (min) of the maximum peakheight of p85a or p49a. The retention time was converted to apparent molecular mass by comparison to a plot of log molecular mass (kDa) versusretention time (min) of the maximum height of the peak for a range of standard proteins (Bio-Rad) chromatographed using the same bufferconditions as in Fig. 2A.
Interactions of Regulatory Subunit of PI 3-Kinase12330
at University of Melbourne (CAUL) on June 10, 2013http://www.jbc.org/Downloaded from

the engagement of the SH2 domains of the regulatory subunitsof PI3K does not affect their oligomerization state.
DISCUSSION
The p85 subunit of PI3K may represent yet another intra-cellular signal transduction protein which utilizes dimerizationas a regulatory mechanism, in a similar manner to cell-surfacereceptors, which are often activated by ligand-induced dimer-ization or oligomerization (31). Several other intracellular sig-naling proteins, such as the STAT transcription factors (32)and c-Raf (33, 34), have also been shown to be regulated bydimerization.
In this study, a number of techniques have been used todemonstrate that the amino-terminal portion of the p85a sub-unit of PI3K binds to itself in an intermolecular manner (Fig.7), which leads to dimerization of p85a both in vitro and in vivo.Additionally, we determined that the isolated SH3 domain ofp85a binds only one of its endogenous proline-rich motifs,PRM1, and the same interaction occurs within the whole pro-tein. In order to determine whether these two interactions wererelated, we further investigated the binding properties of eachof the domains of p85a. However, it became clear that dimer-ization was not mediated by a single domain, and the involve-ment of the BH domain in dimerization made interpretation ofthis data more complicated. However, the ability of the P1peptide to disrupt both p85aSH3-PRM1 and p85a dimers (Fig.4) indicated that the SH3 domain of one p85a molecule bindsthe PRM1 of a second p85a (Fig. 7).
Previous studies have also demonstrated that various frag-ments of p85a are dimeric. A fragment of p85a encompassingresidues 1–101 (similar to p85aSH3-PRM1 in this study) (35)and a fragment similar to p85aSH3-BH (36) have been shownto be dimers. Crystals of the BH domain of p85a have previ-ously been shown to contain two monomers per asymmetricunit (36); however, the hydrophobic dimerization interface wasvery small, involving only four residues of each monomer. Incomparison, the three-dimensional structure of the p85aSH3domain has been determined by both x-ray crystallography and
NMR, and no evidence for self-association of the isolated SH3domain of p85a has been described (30, 37).
The suggestion from the crystal structure of the BH domainthat the 4-residue interface represents a small portion of alarger interaction (36) is confirmed by the contribution of theBH domain to the overall dimeric interface of p85a (Fig. 2B).The size of the interface shown in the crystal structure sug-gests that the affinity of dimerization would be low. If mono-meric and dimeric BH domain existed in equilibrium, and theinterconversion rate was fast, the low affinity for self-associa-tion would result in only a small proportion of dimeric BHdomain and thus may explain the slightly higher average mo-lecular mass observed by both HP-SEC and SE-AUC (Figs. 2Band 3A).
Interactions between SH3 domains and proline-rich motifshave been reported to regulate several intracellular signalingmolecules. In p85a, this interaction not only participates in thedimerization interface but may also block the binding of exog-enous ligands for the p85a SH3 domain and PRM1. Stimula-tion of the B cell receptor has been shown to lead to the bindingof the SH3 domains of the Src family tyrosine kinases, Lyn andFyn, to the p85a PRM1 and the up-regulation PI3K activity(13). The endogenous, intermolecular interaction may thereforealso regulate the binding of exogenous ligands to the SH3domain and PRM1. A similar mechanism has been demon-strated in Itk, a member of the Tec family of cytoplasmictyrosine kinases (38), although the SH3 domain proline-richmotif interaction was intramolecular in this protein. Interac-tion of the SH3 domain with an adjacent proline-rich motif inItk prevented the binding of the SH3 domain to proline-richmotifs in Sam-68 and Grb2. Tyrosine phosphorylation withinthe SH3 domain of another Tec family tyrosine kinase, Btk,was shown to disrupt an intramolecular SH3 domain proline-rich motif interaction and release the Btk SH3 domain to allowit to recruit substrates of the Btk kinase domain (39). Intramo-lecular SH3 domain binding to proline-rich motifs may there-fore be a general mechanism for the regulation of SH3 domainfunction, although there seems to be a number of ways in whichthe SH3 domain can be released to bind its exogenous ligands.The interaction of the p85a SH3 domain and p85a PRM1 seemsto be a special case of this general mechanism in which anintermolecular SH3-PRM1 interaction occurs between the twounits of a dimer.
Recent evidence has suggested that the p85 subunit mayhave functions that are independent of the p110 catalytic sub-unit. A truncated form of p85a, lacking the carboxyl-terminalSH2 domain and 52 residues of the inter-SH2 region, wasidentified as a potential oncogene in a number of murine lym-phomas (40). Overexpression of this mutant protein (p65)caused transformation of NIH-3T3 fibroblasts, and p65 andv-Raf had synergistic transforming activities. Murine embry-onic fibroblasts in which the gene for p85a was deleted byhomologous recombination had a defect in a p53-mediated apo-ptotic pathway (41). Apoptosis caused by oxidative stress wasreduced in cells lacking p85a. Apoptosis was not inhibited bythe PI3K inhibitor, wortmannin, suggesting that the lipid ki-nase activity of the p110 subunit is not a requisite componentof the p53- and p85a-mediated pathways.
One of the best defined functions of the regulatory subunit ofPI3K is the down-regulation of p110-mediated lipid kinaseactivity (42, 43), suggesting that populations of PI3K compris-ing a complex of the p110 and p85 subunits or the p110 subunitalone would have different activities. A 100-kDa PI3K activityhas been detected in lysates of bovine thymus by size-exclusionchromatography (SEC) (44) suggesting that the p110 subunitmay exist on its own and implying that the p85 regulatory
FIG. 7. Proposed model of intermolecular interactions of thep85a regulatory subunit of phosphatidylinositol 3-kinase. Asindicated with the arrows, the N-terminal domains of p85a, the SH3,BH, and PRM1 domains, interact in an intermolecular manner leadingto the formation of a p85a dimer (as discussed in the text).
Interactions of Regulatory Subunit of PI 3-Kinase 12331
at University of Melbourne (CAUL) on June 10, 2013http://www.jbc.org/Downloaded from

subunit may also exist as a separate entity with a biologicalfunction distinct from its role in the regulation of p110 activity.Another potential role of a pool of dimeric p85 in vivo could beto bind and stabilize the p110 catalytic subunit as soon as it istranslated, as p110a has been shown to be prone to inactivationand degradation at 37 °C without the bound regulatory subunit(42).
The existence of forms of the regulatory subunit of PI3K,such as p49a or p55g, that do not have the potential for self-association may represent a mechanism for divergent signal-ing. PI3K-mediated signaling through the insulin receptor andinsulin receptor substrate 1 has been shown to preferentiallyutilize forms of the PI3K regulatory subunit that do not havethe potential for self-association (8, 20). As yet it is unclearwhat implication a lack of dimerization of some regulatorysubunit isoforms has for PI3K signaling. The use of protein-protein interactions of SH3 and BH domains and proline-richmotifs to regulate the nature and localization of PI3K activitywould not be an option for the lower molecular mass isoforms.The regulatory subunit of PI3K may therefore represent bothan adaptor for the catalytic subunit and a mediator of otherprotein-protein interactions that utilize these domains. Thisstudy has highlighted that these same domains are also impli-cated in self-association of the p85 protein, which adds a fur-ther layer to the potential complexity of the regulation of PI3Kactivity in cell signaling events.
Acknowledgments—We thank Akunna Akpan and Krishna Pitrolafor assistance with Sf9 cell culture, Professor I. D. Campbell and Dr. K.Drickamer (University of Oxford) for access to the analytical ultracen-trifuge, and Dr. R. Wallis for kind assistance in the implementation andinterpretation of the SE-AUC experiments.
REFERENCES
1. Zvelebil, M. J., MacDougall, L., Leevers, S., Volinia, S., Vanhaesebroeck, B.,Gout, I., Panayotou, G., Domin, J., Stein, R., Pages, F., Koga, H., Salim, K.,Linacre, J., Das, P., Panaretou, C., Wetzker, R., and Waterfield, M. (1996)Philos. Trans. R. Soc. Lond. Biol. Sci. 351, 217–223
2. Diekmann, D., Brill, S., Garrett, M. D., Totty, N., Hsuan, J., Monfries, C., Hall,C., Lim, L., and Hall, A. (1991) Nature 351, 400–402
3. Dhand, R., Hara, K., Hiles, I., Bax, B., Gout, I., Panayotou, G., Fry, M. J.,Yonezawa, K., Kasuga, M., and Waterfield, M. D. (1994) EMBO J. 13,511–521
4. Otsu, M., Hiles, I., Gout, I., Fry, M. J., Ruiz Larrea, F., Panayotou, G.,Thompson, A., Dhand, R., Hsuan, J., Totty, N., Smith, A. D., Morgan, S. J.,Courtneidge, S. A., Parker, P. J., and Waterfield, M. D. (1991) Cell 65,91–104
5. Skolnik, E. Y., Margolis, B., Mohammadi, M., Lowenstein, E., Fischer, R.,Drepps, A., Ullrich, A., and Schlessinger, J. (1991) Cell 65, 83–90
6. Escobedo, J. A., Navankasattusas, S., Kavanaugh, W. M., Milfay, D., Fried,V. A., and Williams, L. T. (1991) Cell 65, 75–82
7. Antonetti, D. A., Algenstaedt, P., and Kahn, C. R. (1996) Mol. Cell. Biol. 16,2195–2203
8. Inukai, K., Anai, M., Van Breda, E., Hosaka, T., Katagiri, H., Funaki, M.,Fukushima, Y., Ogihara, T., Yazaki, Y., Kikuchi, Oka, Y., and Asano, T.(1996) J. Biol. Chem. 271, 5317–5320
9. Inukai, K., Funaki, M., Ogihara, T., Katagiri, H., Kanda, A., Anai, M.,Fukushima, Y., Hosaka, T., Suzuki, M., Shin, B. C., Takata, K., Yazaki, Y.,Kikuchi, M., Oka, Y., and Asano, T. (1997) J. Biol. Chem. 272, 7873–7882
10. Fruman, D. A., Cantley, L. C., and Carpenter, C. L. (1996) Genomics 37,113–121
11. Pons, S., Asano, T., Glasheen, E., Miralpeix, M., Zhang, Y., Fisher, T. L.,
Myers, M. G., Jr., Sun, X. J., and White, M. F. (1995) Mol. Cell. Biol. 15,4453–4465
12. Vanhaesebroeck, B., Leevers, S. J., Panayotou, G., and Waterfield, M. D.(1997) Trends Biochem. Sci. 22, 267–272
13. Pleiman, C. M., Hertz, W. M., and Cambier, J. C. (1994) Science 263,1609–1612
14. Tolias, K. F., Cantley, L. C., and Carpenter, C. L. (1995) J. Biol. Chem. 270,17656–17659
15. Nobes, C. D., Hawkins, P., Stephens, L., and Hall, A. (1995) J. Cell Sci. 108,225–233
16. Zheng, Y., Bagrodia, S., and Cerione, R. A. (1994) J. Biol. Chem. 269,18727–18730
17. Songyang, Z., Shoelson, S. E., Chaudhuri, M., Gish, G., Pawson, T., Haser,W. G., King, F., Roberts, T., Ratnofsky, S., Lechleider, R. J., Neel, B. G.,Birge, R. B., Fajardo, J. E., Chou, M. M., Hanafusa, H., Schaffhausen, B.,and Cantley, L. C. (1993) Cell 72, 767–778
18. Panayotou, G., and Waterfield, M. D. (1993) BioEssays 15, 171–17719. Backer, J. M., Myers, M. G., Jr., Shoelson, S. E., Chin, D. J., Sun, X. J.,
Miralpeix, M., Hu, P., Margolis, B., Skolnik, E. Y., Schlessinger, J., andWhite, M. F. (1992) EMBO J. 11, 3469–3479
20. Shepherd, P. R., Nave, B. T., Rincon, J., Nolte, L. A., Bevan, A. P., Siddle, K.,Zierath, J. R., and Wallberg Henriksson, H. (1997) J. Biol. Chem. 272,19000–19007
21. Gout, I., Dhand, R., Panayotou, G., Fry, M. J., Hiles, I., Otsu, M., andWaterfield, M. D. (1992) Biochem. J. 288, 395–405
22. Hiles, I. D., Otsu, M., Volinia, S., Fry, M. J., Gout, I., Dhand, R., Panayotou, G.,Ruiz Larrea, F., Thompson, A., Totty, N. F., Hsuan, J. J., Courtneidge, S. A.,Parker, P. A., and Waterfield, M. D. (1992) Cell 70, 419–429
23. Evan, G. I., Lewis, G. K., Ramsay, G., and Bishop, J. M. (1985) Mol. Cell. Biol.5, 3610–3616
24. Kaufman, R. J., Davies, M. V., Pathak, V. K., and Hershey, J. W. (1989) Mol.Cell. Biol. 9, 946–958
25. O’Reilly, D. R., Miller, L. K., and Luckow, V. A. (1994) Baculovirus ExpressionVectors: A Laboratory Manual, Oxford University Press, New York
26. Panayotou, G., Gish, G., End, P., Truong, O., Gout, I., Dhand, R., Fry, M. J.,Hiles, I., Pawson, T., and Waterfield, M. D. (1993) Mol. Cell. Biol. 13,3567–3576
27. End, P., Gout, I., Fry, M. J., Panayotou, G., Dhand, R., Yonezawa, K., Kasuga,M., and Waterfield, M. D. (1993) J. Biol. Chem. 268, 10066–10075
28. Whitman, M., Kaplan, D. R., Schaffhausen, B., Cantley, L., and Roberts, T. M.(1985) Nature 315, 239–242
29. Rickles, R. J., Botfield, M. C., Weng, Z., Taylor, J. A., Green, O. M., Brugge,J. S., and Zoller, M. J. (1994) EMBO J. 13, 5598–5604
30. Yu, H., Chen, J. K., Feng, S., Dalgarno, D. C., Brauer, A. W., and Schreiber,S. L. (1994) Cell 76, 933–945
31. Schlessinger, J., and Ullrich, A. (1992) Neuron 9, 383–39132. Darnell, J. E., Jr., Kerr, I. M., and Stark, G. R. (1994) Science 264,1415–142133. Luo, Z., Tzivion, G., Belshaw, P. J., Vavvas, D., Marshall, M., and Avruch, J.
(1996) Nature 383, 181–18534. Farrar, M. A., Alberol, I., and Perlmutter, R. M. (1996) Nature 383, 178–18135. Chen, J. K., and Schreiber, S. L. (1994) Bioorg. Med. Chem. Lett. 4, 1755–176036. Musacchio, A., Cantley, L. C., and Harrison, S. C. (1996) Proc. Natl. Acad. Sci.
U. S. A. 93, 14373–1437837. Booker, G. W., Gout, I., Downing, A. K., Driscoll, P. C., Boyd, J., Waterfield,
M. D., and Campbell, I. D. (1993) Cell 73, 813–82238. Andreotti, A. H., Bunnell, S. C., Feng, S., Berg, L. J., and Schreiber, S. L.
(1997) Nature 385, 93–9739. Park, H., Wahl, M. I., Afar, D. E., Turck, C. W., Rawlings, D. J., Tam, C.,
Scharenberg, A. M., Kinet, J. P., and Witte, O. N. (1996) Immunity 4,515–525
40. Jimenez, C., Jones, D. R., Rodriguez Viciana, P., Gonzalez Garcia, A.,Leonardo, E., Wennstrom, S., von Kobbe, C., Toran, J. L., R-Borlado, L.,Calvo, V., Copin, S. G., Albar, J. P., Gaspar, M. L., Diez, E., Marcos, M. A.,Downward, J., Martinez, A. C., Merida, I., and Carrera, A. C. (1998) EMBOJ. 17, 743–753
41. Yin, Y., Terauchi, Y., Solomon, G. G., Aizawa, S., Rangarajan, P. N., Yazaki,Y., Kadowaki, T., and Barrett, J. C. (1998) Nature 391, 707–710
42. Yu, J., Zhang, Y., McIlroy, J., Rordorf Nikolic, T., Orr, G. A., and Backer, J. M.(1998) Mol. Cell. Biol. 18, 1379–1387
43. Kodaki, T., Woscholski, R., Hallberg, B., Rodriguez Viciana, P., Downward, J.,and Parker, P. J. (1994) Curr. Biol. 4, 798–806
44. Shibasaki, F., Fukui, Y., and Takenawa, T. (1993) Biochem. J. 289, 227–231
Interactions of Regulatory Subunit of PI 3-Kinase12332
at University of Melbourne (CAUL) on June 10, 2013http://www.jbc.org/Downloaded from

Citations http://www.jbc.org/content/274/18/12323#otherarticles
This article has been cited by 11 HighWire-hosted articles:
at University of Melbourne (CAUL) on June 10, 2013http://www.jbc.org/Downloaded from




















