
APPLIED GENETICS AND MOLECULAR BIOTECHNOLOGY Application of a combined approach involving classical...
10
APPLIED GENETICS AND MOLECULAR BIOTECHNOLOGY Application of a combined approach involving classical random mutagenesis and metabolic engineering to enhance FK506 production in Streptomyces sp. RM7011 SangJoon Mo & Sung-Kwon Lee & Ying-Yu Jin & Chung-Hun Oh & Joo-Won Suh Received: 30 May 2012 / Revised: 30 August 2012 / Accepted: 4 September 2012 / Published online: 2 October 2012 # Springer-Verlag Berlin Heidelberg 2012 Abstract FK506 production by a mutant strain (Streptomyces sp. RM7011) induced by N-methyl-N′-nitro-N-nitrosoguani- dine and ultraviolet mutagenesis was improved by 11.63-fold (94.24 mg/l) compared to that of the wild-type strain. Among three different metabolic pathways involved in the biosynthesis of methylmalonyl-CoA, only expression of propionyl-CoA carboxylase (PCC) pathway led to a 1.75-fold and 2.5-fold increase in FK506 production and the methylmalonyl-CoA pool, respectively, compared to those of the RM7011 strain. Lipase activity of the high FK506 producer mutant increased in direct proportion to the increase in FK506 yield, from low detection level up to 43.1 U/ml (12.6-fold). The level of specific FK506 production and lipase activity was improved by enhancing the supply of lipase inducers. This improvement was approximately 1.88-fold (71.5 mg/g) with the supplemen- tation of 5 mM Tween 80, which is the probable effective stimulator in lipase production, to the R2YE medium. When 5 mM vinyl propionate was added as a precursor for PCC pathway to R2YE medium, the specific production of FK506 increased approximately 1.9-fold (71.61 mg/g) compared to that under the non-supplemented condition. Moreover, in the presence of 5 mM Tween 80, the specific FK506 production was approximately 2.2-fold (157.44 mg/g) higher than that when only vinyl propionate was added to the R2YE medium. In particular, PCC expression in Streptomyces sp. RM7011 (RM7011/pSJ1003) together with vinyl propionate feeding resulted in an increase in the FK506 titer to as much as 1.6- fold (251.9 mg/g) compared with that in RM7011/pSE34 in R2YE medium with 5 mM Tween 80 supplementation, indi- cating that the vinyl propionate is more catabolized to propio- nate by stimulated lipase activity on Tween 80, that propionyl- CoA yielded from propionate generates methylmalonyl-CoA, and that the PCC pathway plays a key role in increasing the methylmalonyl-CoA pool for FK506 biosynthesis in RM7011 strain. Overall, these results show that a combined approach involving classical random mutation and metabolic engineer- ing can be applied to supply the limiting factor for FK506 biosynthesis, and vinyl propionate could be successfully used as a precursor of important methylmalonyl-CoA building blocks. Keywords FK506 . Methylmalonyl-CoA . Metabolic engineering . Propionyl-CoA carboxylase . Lipase activity . Vinyl propionate Introduction FK506, a 23-membered macrocyclic polyketide, was discov- ered in the culture broth of the soil bacterium Streptomyces tsukubaensis in 1987. It has become a clinically important immunosuppressant drug used following solid organ trans- plantation (Kino et al. 1987a, b). Biosynthesis of FK506 is initiated by polyketide synthase (PKS)-catalyzed condensa- tion of an unusual shikimic acid-derived starter unit 4,5-dihy- droxycyclohex-1-enecarboxylic acid and ten extender units, namely, two malonyl-CoA, five methylmalonyl-CoA, two methoxymalonyl-ACP, and a unique allylmalonyl-CoA SangJoon Mo and Sung-Kwon Lee contributed equally to the work. S. Mo : S.-K. Lee : Y.-Y. Jin : J.-W. Suh (*) Division of Bioscience and Bioinformatics, Myongji University, Youngin 449-7282, South Korea e-mail: [email protected] C.-H. Oh Department of Oral Physiology, College of Dentistry, Dankook University, Cheonan 330-714, South Korea Appl Microbiol Biotechnol (2013) 97:3053–3062 DOI 10.1007/s00253-012-4413-5
Transcript of APPLIED GENETICS AND MOLECULAR BIOTECHNOLOGY Application of a combined approach involving classical...

APPLIED GENETICS AND MOLECULAR BIOTECHNOLOGY
Application of a combined approach involving classicalrandom mutagenesis and metabolic engineering to enhanceFK506 production in Streptomyces sp. RM7011
SangJoon Mo & Sung-Kwon Lee & Ying-Yu Jin &
Chung-Hun Oh & Joo-Won Suh
Received: 30 May 2012 /Revised: 30 August 2012 /Accepted: 4 September 2012 /Published online: 2 October 2012# Springer-Verlag Berlin Heidelberg 2012
Abstract FK506 production by a mutant strain (Streptomycessp. RM7011) induced by N-methyl-N′-nitro-N-nitrosoguani-dine and ultraviolet mutagenesis was improved by 11.63-fold(94.24 mg/l) compared to that of the wild-type strain. Amongthree different metabolic pathways involved in the biosynthesisof methylmalonyl-CoA, only expression of propionyl-CoAcarboxylase (PCC) pathway led to a 1.75-fold and 2.5-foldincrease in FK506 production and the methylmalonyl-CoApool, respectively, compared to those of the RM7011 strain.Lipase activity of the high FK506 producer mutant increasedin direct proportion to the increase in FK506 yield, from lowdetection level up to 43.1 U/ml (12.6-fold). The level ofspecific FK506 production and lipase activity was improvedby enhancing the supply of lipase inducers. This improvementwas approximately 1.88-fold (71.5 mg/g) with the supplemen-tation of 5 mM Tween 80, which is the probable effectivestimulator in lipase production, to the R2YE medium. When5 mM vinyl propionate was added as a precursor for PCCpathway to R2YE medium, the specific production of FK506increased approximately 1.9-fold (71.61 mg/g) compared tothat under the non-supplemented condition. Moreover, in thepresence of 5 mM Tween 80, the specific FK506 productionwas approximately 2.2-fold (157.44 mg/g) higher than thatwhen only vinyl propionate was added to the R2YE medium.
In particular, PCC expression in Streptomyces sp. RM7011(RM7011/pSJ1003) together with vinyl propionate feedingresulted in an increase in the FK506 titer to as much as 1.6-fold (251.9 mg/g) compared with that in RM7011/pSE34 inR2YE medium with 5 mM Tween 80 supplementation, indi-cating that the vinyl propionate is more catabolized to propio-nate by stimulated lipase activity on Tween 80, that propionyl-CoA yielded from propionate generates methylmalonyl-CoA,and that the PCC pathway plays a key role in increasing themethylmalonyl-CoA pool for FK506 biosynthesis in RM7011strain. Overall, these results show that a combined approachinvolving classical random mutation and metabolic engineer-ing can be applied to supply the limiting factor for FK506biosynthesis, and vinyl propionate could be successfully usedas a precursor of important methylmalonyl-CoA buildingblocks.
Keywords FK506 .Methylmalonyl-CoA .Metabolicengineering . Propionyl-CoA carboxylase . Lipase activity .
Vinyl propionate
Introduction
FK506, a 23-membered macrocyclic polyketide, was discov-ered in the culture broth of the soil bacterium Streptomycestsukubaensis in 1987. It has become a clinically importantimmunosuppressant drug used following solid organ trans-plantation (Kino et al. 1987a, b). Biosynthesis of FK506 isinitiated by polyketide synthase (PKS)-catalyzed condensa-tion of an unusual shikimic acid-derived starter unit 4,5-dihy-droxycyclohex-1-enecarboxylic acid and ten extender units,namely, two malonyl-CoA, five methylmalonyl-CoA, twomethoxymalonyl-ACP, and a unique allylmalonyl-CoA
SangJoon Mo and Sung-Kwon Lee contributed equally to the work.
S. Mo : S.-K. Lee :Y.-Y. Jin : J.-W. Suh (*)Division of Bioscience and Bioinformatics, Myongji University,Youngin 449-7282, South Koreae-mail: [email protected]
C.-H. OhDepartment of Oral Physiology, College of Dentistry,Dankook University,Cheonan 330-714, South Korea
Appl Microbiol Biotechnol (2013) 97:3053–3062DOI 10.1007/s00253-012-4413-5

extender unit are incorporated into nascent polyketide chain.The linear polyketide chain is followed by incorporation oflysine-derived pipecolic acid and a cyclization step, mediatedby FkbP peptide synthetase, resulting in the earliest macro-lactone intermediate of FK506. Finally, post-PKS processingreactions are catalyzed by a specific methyl transferase andoxidoreductase resulting in FK506 (Motamedi et al. 1997,1998; Mo et al. 2009, 2011).
Recently, other promising biological activities, such asneuroprotective and neuroregenerative activities, werereported, creating significant interest in the further develop-ment of FK506 for different medicinal uses (Sierra-Paredeset al. 2008; Gold 2000). As a consequence of its pharmaco-logical importance and broad applicability, considerabletime and effort has been expended enhancing FK506 yieldand developing more efficient and economical processes forindustrial production (Kumar et al. 2007). Classical technol-ogies have been used to enhance FK506 titers in industrialstrains, such as ultraviolet (UV) irradiation (Kim and Park2007) and sequential adaptation techniques (Jung et al.2009). Although the classic strain improvement technologyis methodologically simple, it is an extremely effective wayto enhance productivity of fermentation processes (Baltz1986a, b, 1999), and many scientists have used this meth-odology to generate industrial strains despite the develop-ment of genetic manipulation skills (Xu et al. 2005; Khajiqet al. 2009; Chen et al. 2009). However, this classic strainimprovement method is a time- and labor-intensive process,suggesting the use of a more rational method involvingmetabolic engineering (Chen et al. 2010).
Metabolic engineering of biochemical pathways inmicroorganisms through molecular biology techniques is apowerful tool to increase productivity of primary and sec-ondary metabolites (Nielsen 2001; Cameron et al. 1993).Successfully applying metabolic engineering dependsheavily on (1) genome sequence information of the micro-organism, (2) understanding the biosynthesis of the targetmolecule and its related metabolic fluxes, and (3) establish-ing molecular biology methodologies to perform the desiredgenetic manipulations in the producers (Rokem et al. 2007;Chen et al. 2010). Enhanced production of microbial sec-ondary metabolites is achieved through metabolic engineer-ing by developing an understanding of the biosyntheticmachinery and regulatory networks of secondary metabo-lites (Borodian et al. 2005; Hertweck 2009) and developingnew genetic manipulation strategies, e.g., polymerase chainreaction (PCR)-targeting system (Gust et al. 2003), inStreptomyces. Since recombinant DNA technology has beendeveloped, several reports have used a method, called the“reverse engineering techniques,” to identify the molecularbasis of successful mutate-and-screen industrial strains or toexplain the cause of high productivity in successfully devel-oped strains by transposition mutagenesis (Reeves et al.
2004, 2006). Unfortunately, reverse engineering is difficultto apply in existing high-producing industrial strains be-cause the relevant genes in these industrial strains are notmarked and cannot be identified. Moreover, even if genome-sequencing techniques accomplish remarkable develop-ments, no simple method is useful to distinguish advanta-geous mutations from silent or detrimental genes between ahighly improved industrial strain and its wild-type progen-itor (Reeves et al. 2004). Here, we report a successfulcombined approach of classical random mutagenesis andrational metabolic engineering for a detailed analysis ofprecursors limiting the productivity of a target polyketide.We also provide the most effective precursor supply path-way that supports the maximum FK506 titer in Streptomycessp. RM7011.
Materials and methods
Bacterial strains, plasmids, culture condition, and genemanipulation
The Escherichia coli strain DH5α and the E. coli–Streptomyces vector pSE34 were used as a host for thecloning and expression experiments according to standardmolecular biology procedures (Kieser et al. 2000; Sambrookand Russell 2001; Smirnova and Reynolds 2001). Litmus28(New England Biolabs) was used for subcloning, Streptomycessp. KCCM 11116P spores were produced on ISP4 agar plates(Shirling et al. 1966), and a seed culture was prepared in R2YEbroth (Kieser et al. 2000). FK506 production was carriedout by inoculating a baffled 250-ml flask containing 50 mlR2YE medium at pH 7.2 with 500 μl of a seed culturesuspension and then incubating the suspension on an orbitalshaker (180 rpm) for 6 days at 28 °C. Saccharopolysporaerythraea, Streptomyces coelicolor A3(2), and Salmonellatyphimurium LT2 ATCC 29631 were cultured in SCM,YEME, and nutrient broths, respectively, to isolate genomicDNA (Horswill et al. 1999; Kieser et al. 2000; Reeves et al.2007). DNA extraction and manipulation and transformationof E. coli and Streptomyces were performed according tostandard protocols (Sambrook and Russell 2001; Kieser etal. 2000). Restriction enzymes andmolecular biology reagentswere purchased from New England Biolabs and RocheDiagnostics. PCR products were purified using theQIAquick PCR Purification kit (Qiagen), and restriction frag-ments were purified from agarose gel using the QIAquick GelPurification kit (Qiagen). Luria–Bertani (LB), SOB, and SOCliquidmedia were used to growE. coli (Sambrook and Russell2001). Ampicillin (100 μg/ml) and thiostrepton (20 μg/ml),all from Sigma-Aldrich, were added to growth media asrequired. At the beginning of cultivation, lipase inducers, suchas olive oil, soybean oil, corn oil, and Tween 80 from Sigma-
3054 Appl Microbiol Biotechnol (2013) 97:3053–3062

Aldrich, were added to the R2YEmedium at concentrations of5 g/l. Additionally, Tween 80 was added at various concen-trations to R2YE medium at the beginning of cultivation todetermine the optimal initial Tween 80 concentration forefficient FK506 production.
Screening of the FK506-producing strain
Bacterial strains isolated from a soil sample collected in Koreawere screened for antifungal activity with Aspergillus nigerATCC6275 on YPD agar medium. More than 10,000 micro-organisms with antifungal activity were re-screened for im-munosuppressive activity using the T cell proliferation assay,as described previously (Kino et al. 1987b). MJM7001 strain,which showed the higher levels of FK506 production com-pared with others which were FK506-producing strain among200 strains that inhibited T cell proliferation, was chosen forfurther study, and its FK506 production was confirmed bio-chemically by detecting FK506 on high-performance liquidchromatography (HPLC) and HPLC-electrospray ionization-tandem mass spectrometry (ESI-MS/MS), according to themethod of Mo et al. (2009, 2011). The MJM7001 strain wastentatively identified as Streptomyces sp. (Locci et al. 1989)and was deposited in the strain collection of the KoreanCulture Center of Microorganisms under accession numberKCCM 11116P. Strains were stored in a liquid nitrogen stock-er, and working stocks were maintained on Bennett's mediumslants (Iwasa et al. 1970).
Development of the FK506-producing strain by mutagenictreatment
Spore suspensions of Streptomyces sp. KCCM 11116P (107
spores ml−1) were treated with N-methyl-N′-nitro-N-nitro-soguanidine (NTG; 0.5 mgl−1) in 50 mM Tris and maleicacid buffer, pH 9.0, for 10, 20, 40, and 60 min (Hopwood etal. 1985). For the second round mutation, spore suspension(50 μl) was mixed with 10 ml sterile distilled water and givethe power of UV stratalinker at a distance of 15 cm for 20,40, 60, 80, 100, 120, 140, and 160 s from the UV lamp (l0300). All manipulations were carried out in a dark room toavoid any possible photoreaction (Delić et al. 1970).Survival ratios were calculated for each treatment by spread-ing the NTG-treated and UV-exposed spore suspension(killing ratio, 99 %) and parent strain on regeneration agarmedium plates that were incubated at 28 °C for 5 days.
Screening study for the FK506 hyper-producing mutantusing the A. niger bioassay and HPLC analysis
Colonies of Streptomyces sp. KCCM 11116P treated withNTG and exposed to UV irradiation were grown on ISP4medium to obtain spores. Then, the spores were spread on
Bennett's agar plate to yield 50–80 colonies per plate. After5 days incubation at 28 °C, the agar-grown colonies weregouged out to the same size with a sterilized cork borer.Agar plug-grown colonies were transferred to YPD agar in-oculated with 106 cells ml−1 of A. niger (Kumar et al. 2000).After 5 days incubation at 28 °C, colonies capable of produc-ing wide inhibition zones were selected, cultured, and ana-lyzed as described previously (Mo et al. 2009). Additionally,FK506 production of the strains was finally confirmed byHPLC and HPLC-ESI-MS/MS analyses according to themethod of Mo et al. (2009, 2011). An authentic FK506standard (Sigma-Aldrich) was used to construct aFK506 calibration curve for the HPLC analysis. Thelevel of FK506 production reported is the average oftwo series of duplicate separate cultivations and extrac-tions. The detailed procedure for the agar plug screeningtechnique is illustrated in Fig. 1.
Overexpression of the methylmalonyl-CoA formationpathway in Streptomyces sp. RM7011
Overexpression experiments with plasmids expressing threedifferent methylmalonyl-CoA formation pathways were car-ried out. The genes encoding the enzyme involved in themethylmalonyl-CoA mutase (MCM) pathway (mutAB),malonyl/methylmalonyl-CoA and methylmalonyl-CoA epim-erase (matB), and propionyl-CoA synthetase gene (prpE) andpropionyl-CoA carboxylase (PCC) pathway (accAI, pccB)were cloned into streptomycetes expression plasmid pSE34to generate pSJ1001, pSJ1002, and pSJ1003, respectively.pSJ1002 and pSJ1003 was constructed in the same way,except pSJ1001, as described previously (Mo et al. 2009).The pSJ1001 was constructed using the following method: themutA gene (coding for the small subunit of MCM) wasobtained as a BglII–XbaI PCR fragment from S. erythraeaNRRL2338 genomic DNA and inserted into Litmus28i. Theoligonucleotide primer set used to amplify mutA was 5′-GTATTAAGATCTCTGTCCGGCGCGACAAGCGTC-3′(forward) and 5′-CACTCTAGACGAGCCGGCCAG-3′(reverse) containing the BglII and XbaI sites (underlined),respectively. The mutB gene (coding for the large subunit ofMCM) was obtained as an XbaI–HindIII PCR fragment fromthe S. erythraea NRRL2338 genomic DNA and inserted intothe same site of Litmus 28i containing the mutA gene. Theprimer set used to amplify mutB was 5′-ATCTTATCTAGAGTGACCGCCCACGAG-3 ′ (forward) and 5′-CATTCTAAGCTTTCAGCCCGCGCTCGACG-3′ (reverse) con-taining the XbaI and HindIII sites (underlined), respectively.The BglII–HindIII fragment of themutA andmutB genes wereobtained from the resulting Litmus28i, and ligated to theBamHI–HindIII sites of pSE34, generating pSJ1001.Protoplasts of Streptomyces sp. RM7011 were prepared fol-lowing standard methods (Kieser et al. 2000) and transformed
Appl Microbiol Biotechnol (2013) 97:3053–3062 3055

with pSJ1001, pSJ1002, and pSJ1003. The resultingthiostrepton-resistant transformants were obtained, and theirrespective FK506 production levels were evaluated as de-scribed above.
HPLC-ESI-MS/MS analyses of the intracellularmethylmalonyl-CoA pools
Streptomyces sp. RM7011 and RM7011/pSJ1003 were cul-tured in R2YE medium, supplemented with 20 μg/ml thio-strepton when required, and 5 ml of culture broth was takeneach day for 4 days. The mycelia were collected, washedtwice with cold distilled water, and the wet biomass wasmeasured according to the method of Mo et al. (2009). Cellpellets were suspended in 300-μl 15 % trichloroacetic acidsolution and lysed by vortexing for 3 min at 4 °C with 150-μl glass beads. Lysates were centrifuged again for 10 min at13,000 rpm and 4 °C. The supernatant was passed through anOASIS HLB SPE cartridge (OASIS HLB 3 cm3/60 mg) undera vacuum, as reported previously (Park et al. 2007). Analysisof the intracellular propionyl-CoAs and methylmalonyl-CoAs were performed according to the method of Park etal. (2007). The experiments were carried out in two indepen-dent replicates.
Lipase activity
Sonicated culture broth (10 μl) was added to 2.99 ml Tris–HCl buffer (pH 8.0) containing 30 μM of para-nitrophenylbutyrate (p-NPB) as a substrate, according to the modifiedmethod of Lu et al. (2009). The reactions were followedspectrophotometrically at 410 nm. Lipase activity was de-termined according to a para-nitrophenol (p-NP) standardcurve measured under the same conditions. Lipase activitywas expressed in units. One unit was the amount of lipasethat liberated 1 mmol of p-NP per minute. Lipase activitywas measured in whole broth, the cell-free supernatant, andthe pellet sample to determine the cellular location of thelipase activity. The whole broth and pellet sample weresubjected to sonication using Soniprep 150 (Sanyo) at an8-μm setting for 2×10 min. All assays were performed intwo duplicate series.
RM7011 feeding study of the propionyl-CoA precursor
RM7011 seed cultures were grown in R2YE medium for48 h at 28 °C and then used as 1 % inoculums for 50-mlliquid culture of the same medium. Sodium propionate andvinyl propionate (Sigma-Aldrich) were added to a final
Spore suspention of Streptomyces sp. KCCM 11116P
UV/NTG mutation
Incubate at 28°C for 4-5 days on BN agar Agar plug
Bioassy with A. nigeron PDA agar in square dish plate
Liquid culture ofactive mutant strain
Extraction and analysis
HPLC analysis of FK506 production
Strain stabilization
Cell stock
Cork borer
Colonies on BN agar
Incubate agar plug with on PDA agar in square dish plate
Control
Select for sub-culture & future work up in shake flask
Fig. 1 Flow charts for the modified agar plug method and Aspergillus niger bioassay
3056 Appl Microbiol Biotechnol (2013) 97:3053–3062

concentration of 5 mM at 48 h. Another RM7011 strain wasgrown in R2YE medium containing 5 mM Tween 80 tostimulate lipase production, and then sodium propionateand vinyl propionate were added at 48 h as described above.The production cultures were grown for 5 days at 28 °C asdescribed above. Duplicate cell-free broths were collected inthe flask by vacuum filtration, and then extracted with ethylacetate for HPLC analysis according to the method of Mo etal. (2009). RM7011/pSE34 and RM7011/pSJ1003 were fedwith 5 mM vinyl propionate in R2YE medium containingthiostrepton (20 μg/ml) supplemented with 5 mM Tween 80to investigate the effect of PCC pathway overexpression.Duplicated samples (50 ml) of fermentation broth werecollected to measure FK506 production every 24 h for 6 daysafter the main culture inoculation and extracted as describedpreviously (Mo et al. 2009).
Results
Isolation of FK506-producing strain and selection of FK506high producers by random mutagenesis
FK506 production by the 200 strains that inhibited T cellproliferation was analyzed qualitatively by HPLC andHPLC-ESI-MS/MS. As a result, the MJM7001 strainshowed the highest FK506 production among all FK506-producing strains. The MJM7001 strain was identified as aStreptomyces sp. by 16S rDNA analysis, and it was depos-ited with the designation Streptomyces sp. KCCM 11116P inthe strain collection of the Korean Culture Center ofMicroorganisms.
Streptomyces sp. KCCM 11116P wild-type strain sporeswere treated with NTG and UV to obtain mutants withincreased FK506. FK506 production of individual mutagen-esis colonies was assayed using the agar plug method withA. niger ATCC6275 (Kumar et al. 2000; Kim and Park2007), and the colonies with the largest growth inhibitionzone diameters were analyzed again by HPLC. One colonywith the largest growth inhibition zone diameter was select-ed from 3,641 mutagenized colonies with NTG and desig-nated as RM1753. The RM1753 mutant strain produced anapproximate 2.56-fold increase (20.8 mg/l) in FK506 yieldcompared with that in the wild-type strain (8.1 mg/l;Table 1). The second round of mutations and selectionswas performed using RM1753 strain. RM7011 mutantstrain, which was selected among 3,508 mutagenized colo-nies with UV light, showed the highest FK506 production,reaching 94.24 mg/l in comparison with the RM1753 strainthat produced 20.8 mg/l (Table 1). Over 7,100 mutatedcolonies with NTG and UV were screened using this screenmethod, and the RM7011 mutant strain resulted in approx-imately 11.63-fold (94.24 mg/l) increase in the FK506 yield
compared to that of the wild-type strain (8.1 mg/l; Table 1).However, the two mutant strains, RM1753 and RM7011,were not distinct when compared to the wild-type strain, andthe three strains (wild type, RM1753, and RM7011) showedparallel growth rates and final cell densities (data notshown), indicating that random NTG and UV mutagenesisdid not increase cell density.
Overexpression of the methylmalonyl-CoA formationpathways in mutant RM7011
Methylmalonyl-CoA is a limiting factor for FK506 produc-tion in Streptomyces clavuligerus CKD1119, and overex-pression of MCM pathway leads to a significant increase inFK506 production in R2YE medium (Mo et al. 2009).Based on this result, we investigated the effect of the afore-mentioned three pathways that affect methylmalonyl-CoAlevel and FK506 production by heterologous expression ofthe MCM, MatB, and PCC pathways in the RM7011 strain(Fig. 2a). Interestingly, only the PCC pathway produced a1.75-fold (164.92 mg/l) increase in the FK506 titers amongthe three above-mentioned pathways in RM7011 strain(Fig. 2a). In contrast, heterologous expression of themutAB genes or matB gene in RM7011 strain failed toenhance FK506 production. Furthermore, providing succi-nate or methylmalonate to the culture broth did not affectFK506 production in RM7011/pSJ1001 or RM7011/pSJ1002 (data not shown). This result corresponds to theobservation that neither the MCM nor the MatB pathwaysincreased FK506 production in the RM7011 strain.
To corroborate these results, the RM7011 and RM7011/pSJ1003 strains were analyzed for their intracellularmethylmalonyl-CoA levels. Cells from the RM7011 andRM7011/pSJ1003 strains were harvested at day 2 to analyzethe CoA-ester, when the difference in their FK506 produc-tion had become clear. The heterologous expression of thePCC pathway in RM7011 strain yielded a 2.5-fold increasein methylmalonyl-CoA level compared to the levels present
Table 1 Comparative result of FK506 titers after random mutation ofStreptomyces sp. KCCM 11116P
Strain FK506 productivity(mg/L)c
Relative productivity(fold)
Streptomyces sp.KCCM 11116P
8.1±0.91 1.0
RM1753a 20.8±2.1 2.56
RM7011b 94.24±4.7 11.63
a A strain of Streptomyces sp. KCCM 11116P mutated by NTGtreatmentb A strain of Streptomyces sp. RM1753 mutated by UV lightc Values shown represent the means plus and minus standard deviationof two sets of replicate measurements
Appl Microbiol Biotechnol (2013) 97:3053–3062 3057
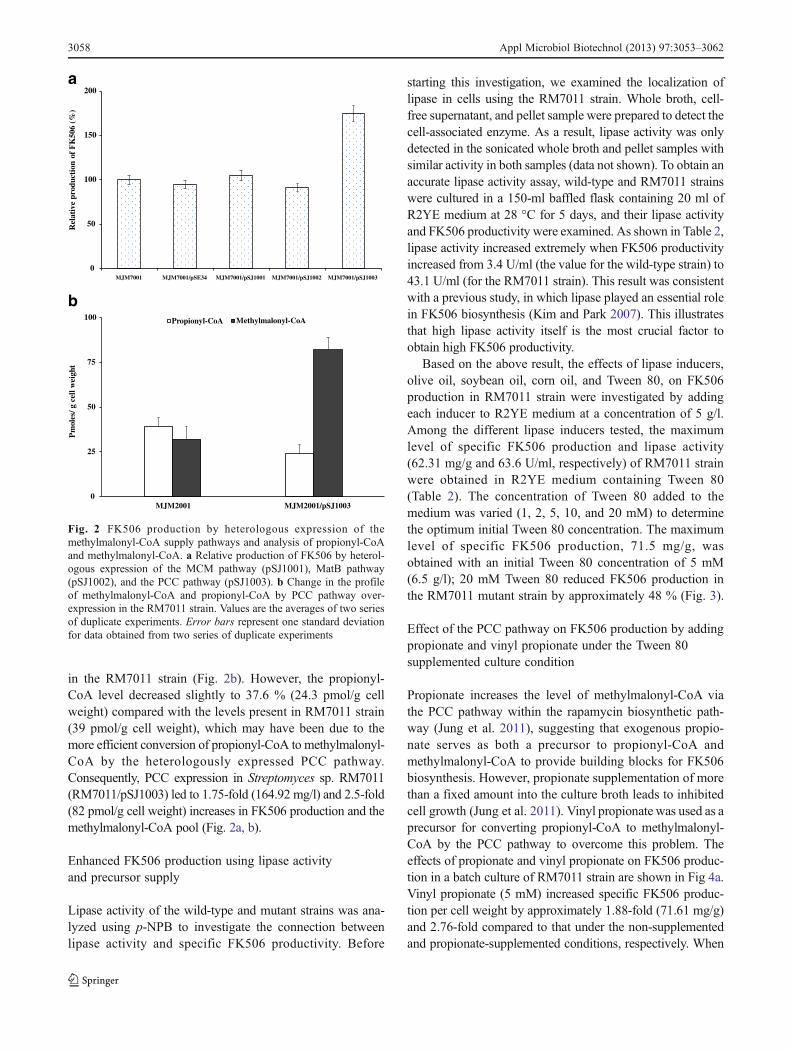
in the RM7011 strain (Fig. 2b). However, the propionyl-CoA level decreased slightly to 37.6 % (24.3 pmol/g cellweight) compared with the levels present in RM7011 strain(39 pmol/g cell weight), which may have been due to themore efficient conversion of propionyl-CoA to methylmalonyl-CoA by the heterologously expressed PCC pathway.Consequently, PCC expression in Streptomyces sp. RM7011(RM7011/pSJ1003) led to 1.75-fold (164.92 mg/l) and 2.5-fold(82 pmol/g cell weight) increases in FK506 production and themethylmalonyl-CoA pool (Fig. 2a, b).
Enhanced FK506 production using lipase activityand precursor supply
Lipase activity of the wild-type and mutant strains was ana-lyzed using p-NPB to investigate the connection betweenlipase activity and specific FK506 productivity. Before
starting this investigation, we examined the localization oflipase in cells using the RM7011 strain. Whole broth, cell-free supernatant, and pellet sample were prepared to detect thecell-associated enzyme. As a result, lipase activity was onlydetected in the sonicated whole broth and pellet samples withsimilar activity in both samples (data not shown). To obtain anaccurate lipase activity assay, wild-type and RM7011 strainswere cultured in a 150-ml baffled flask containing 20 ml ofR2YE medium at 28 °C for 5 days, and their lipase activityand FK506 productivity were examined. As shown in Table 2,lipase activity increased extremely when FK506 productivityincreased from 3.4 U/ml (the value for the wild-type strain) to43.1 U/ml (for the RM7011 strain). This result was consistentwith a previous study, in which lipase played an essential rolein FK506 biosynthesis (Kim and Park 2007). This illustratesthat high lipase activity itself is the most crucial factor toobtain high FK506 productivity.
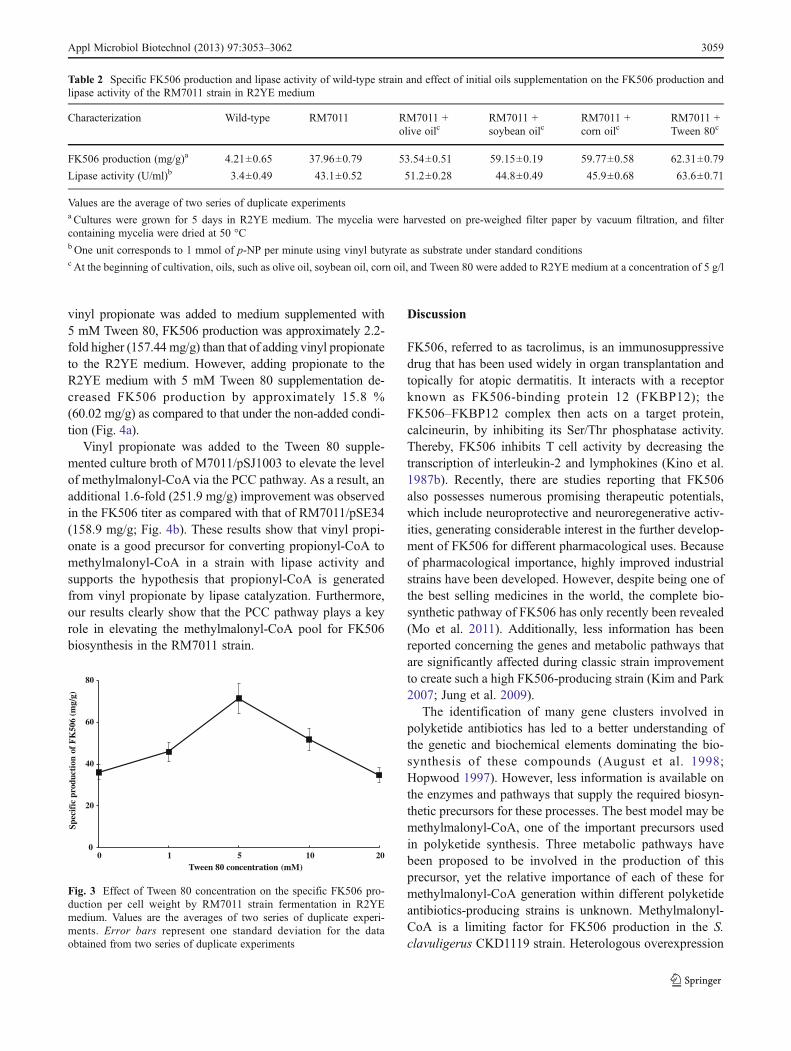
Based on the above result, the effects of lipase inducers,olive oil, soybean oil, corn oil, and Tween 80, on FK506production in RM7011 strain were investigated by addingeach inducer to R2YE medium at a concentration of 5 g/l.Among the different lipase inducers tested, the maximumlevel of specific FK506 production and lipase activity(62.31 mg/g and 63.6 U/ml, respectively) of RM7011 strainwere obtained in R2YE medium containing Tween 80(Table 2). The concentration of Tween 80 added to themedium was varied (1, 2, 5, 10, and 20 mM) to determinethe optimum initial Tween 80 concentration. The maximumlevel of specific FK506 production, 71.5 mg/g, wasobtained with an initial Tween 80 concentration of 5 mM(6.5 g/l); 20 mM Tween 80 reduced FK506 production inthe RM7011 mutant strain by approximately 48 % (Fig. 3).
Effect of the PCC pathway on FK506 production by addingpropionate and vinyl propionate under the Tween 80supplemented culture condition
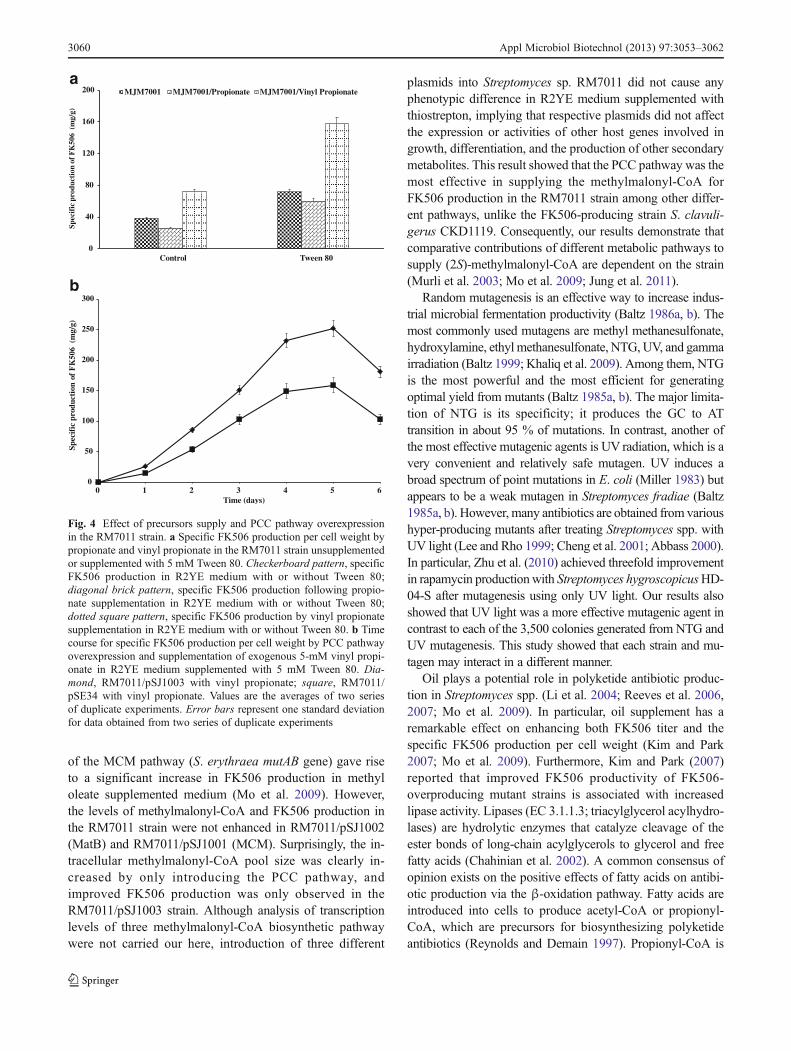
Propionate increases the level of methylmalonyl-CoA viathe PCC pathway within the rapamycin biosynthetic path-way (Jung et al. 2011), suggesting that exogenous propio-nate serves as both a precursor to propionyl-CoA andmethylmalonyl-CoA to provide building blocks for FK506biosynthesis. However, propionate supplementation of morethan a fixed amount into the culture broth leads to inhibitedcell growth (Jung et al. 2011). Vinyl propionate was used as aprecursor for converting propionyl-CoA to methylmalonyl-CoA by the PCC pathway to overcome this problem. Theeffects of propionate and vinyl propionate on FK506 produc-tion in a batch culture of RM7011 strain are shown in Fig 4a.Vinyl propionate (5 mM) increased specific FK506 produc-tion per cell weight by approximately 1.88-fold (71.61 mg/g)and 2.76-fold compared to that under the non-supplementedand propionate-supplemented conditions, respectively. When
0
50
100
150
200
MJM7001 MJM7001/pSE34 MJM7001/pSJ1001 MJM7001/pSJ1002 MJM7001/pSJ1003
Rel
ativ
e pr
oduc
tion
of
FK
506
(%)
0
25
50
75
100
MJM2001 MJM2001/pSJ1003
Pm
oles
/ g c
ell w
eigh
t
Propionyl-CoA Methylmalonyl-CoA
a
b
Fig. 2 FK506 production by heterologous expression of themethylmalonyl-CoA supply pathways and analysis of propionyl-CoAand methylmalonyl-CoA. a Relative production of FK506 by heterol-ogous expression of the MCM pathway (pSJ1001), MatB pathway(pSJ1002), and the PCC pathway (pSJ1003). b Change in the profileof methylmalonyl-CoA and propionyl-CoA by PCC pathway over-expression in the RM7011 strain. Values are the averages of two seriesof duplicate experiments. Error bars represent one standard deviationfor data obtained from two series of duplicate experiments
3058 Appl Microbiol Biotechnol (2013) 97:3053–3062
vinyl propionate was added to medium supplemented with5 mM Tween 80, FK506 production was approximately 2.2-fold higher (157.44 mg/g) than that of adding vinyl propionateto the R2YE medium. However, adding propionate to theR2YE medium with 5 mM Tween 80 supplementation de-creased FK506 production by approximately 15.8 %(60.02 mg/g) as compared to that under the non-added condi-tion (Fig. 4a).
Vinyl propionate was added to the Tween 80 supple-mented culture broth of M7011/pSJ1003 to elevate the levelof methylmalonyl-CoA via the PCC pathway. As a result, anadditional 1.6-fold (251.9 mg/g) improvement was observedin the FK506 titer as compared with that of RM7011/pSE34(158.9 mg/g; Fig. 4b). These results show that vinyl propi-onate is a good precursor for converting propionyl-CoA tomethylmalonyl-CoA in a strain with lipase activity andsupports the hypothesis that propionyl-CoA is generatedfrom vinyl propionate by lipase catalyzation. Furthermore,our results clearly show that the PCC pathway plays a keyrole in elevating the methylmalonyl-CoA pool for FK506biosynthesis in the RM7011 strain.
Discussion
FK506, referred to as tacrolimus, is an immunosuppressivedrug that has been used widely in organ transplantation andtopically for atopic dermatitis. It interacts with a receptorknown as FK506-binding protein 12 (FKBP12); theFK506–FKBP12 complex then acts on a target protein,calcineurin, by inhibiting its Ser/Thr phosphatase activity.Thereby, FK506 inhibits T cell activity by decreasing thetranscription of interleukin-2 and lymphokines (Kino et al.1987b). Recently, there are studies reporting that FK506also possesses numerous promising therapeutic potentials,which include neuroprotective and neuroregenerative activ-ities, generating considerable interest in the further develop-ment of FK506 for different pharmacological uses. Becauseof pharmacological importance, highly improved industrialstrains have been developed. However, despite being one ofthe best selling medicines in the world, the complete bio-synthetic pathway of FK506 has only recently been revealed(Mo et al. 2011). Additionally, less information has beenreported concerning the genes and metabolic pathways thatare significantly affected during classic strain improvementto create such a high FK506-producing strain (Kim and Park2007; Jung et al. 2009).
The identification of many gene clusters involved inpolyketide antibiotics has led to a better understanding ofthe genetic and biochemical elements dominating the bio-synthesis of these compounds (August et al. 1998;Hopwood 1997). However, less information is available onthe enzymes and pathways that supply the required biosyn-thetic precursors for these processes. The best model may bemethylmalonyl-CoA, one of the important precursors usedin polyketide synthesis. Three metabolic pathways havebeen proposed to be involved in the production of thisprecursor, yet the relative importance of each of these formethylmalonyl-CoA generation within different polyketideantibiotics-producing strains is unknown. Methylmalonyl-CoA is a limiting factor for FK506 production in the S.clavuligerus CKD1119 strain. Heterologous overexpression
Table 2 Specific FK506 production and lipase activity of wild-type strain and effect of initial oils supplementation on the FK506 production andlipase activity of the RM7011 strain in R2YE medium
Characterization Wild-type RM7011 RM7011 +olive oilc
RM7011 +soybean oilc
RM7011 +corn oilc
RM7011 +Tween 80c
FK506 production (mg/g)a 4.21±0.65 37.96±0.79 53.54±0.51 59.15±0.19 59.77±0.58 62.31±0.79
Lipase activity (U/ml)b 3.4±0.49 43.1±0.52 51.2±0.28 44.8±0.49 45.9±0.68 63.6±0.71
Values are the average of two series of duplicate experimentsa Cultures were grown for 5 days in R2YE medium. The mycelia were harvested on pre-weighed filter paper by vacuum filtration, and filtercontaining mycelia were dried at 50 °CbOne unit corresponds to 1 mmol of p-NP per minute using vinyl butyrate as substrate under standard conditionsc At the beginning of cultivation, oils, such as olive oil, soybean oil, corn oil, and Tween 80 were added to R2YE medium at a concentration of 5 g/l
0
20
40
60
80
0 1 5 10 20
Spec
ific
pro
duct
ion
of F
K50
6 (m
g/g)
Tween 80 concentration (mM)
Fig. 3 Effect of Tween 80 concentration on the specific FK506 pro-duction per cell weight by RM7011 strain fermentation in R2YEmedium. Values are the averages of two series of duplicate experi-ments. Error bars represent one standard deviation for the dataobtained from two series of duplicate experiments
Appl Microbiol Biotechnol (2013) 97:3053–3062 3059
of the MCM pathway (S. erythraea mutAB gene) gave riseto a significant increase in FK506 production in methyloleate supplemented medium (Mo et al. 2009). However,the levels of methylmalonyl-CoA and FK506 production inthe RM7011 strain were not enhanced in RM7011/pSJ1002(MatB) and RM7011/pSJ1001 (MCM). Surprisingly, the in-tracellular methylmalonyl-CoA pool size was clearly in-creased by only introducing the PCC pathway, andimproved FK506 production was only observed in theRM7011/pSJ1003 strain. Although analysis of transcriptionlevels of three methylmalonyl-CoA biosynthetic pathwaywere not carried our here, introduction of three different
plasmids into Streptomyces sp. RM7011 did not cause anyphenotypic difference in R2YE medium supplemented withthiostrepton, implying that respective plasmids did not affectthe expression or activities of other host genes involved ingrowth, differentiation, and the production of other secondarymetabolites. This result showed that the PCC pathway was themost effective in supplying the methylmalonyl-CoA forFK506 production in the RM7011 strain among other differ-ent pathways, unlike the FK506-producing strain S. clavuli-gerus CKD1119. Consequently, our results demonstrate thatcomparative contributions of different metabolic pathways tosupply (2S)-methylmalonyl-CoA are dependent on the strain(Murli et al. 2003; Mo et al. 2009; Jung et al. 2011).
Random mutagenesis is an effective way to increase indus-trial microbial fermentation productivity (Baltz 1986a, b). Themost commonly used mutagens are methyl methanesulfonate,hydroxylamine, ethyl methanesulfonate, NTG, UV, and gammairradiation (Baltz 1999; Khaliq et al. 2009). Among them, NTGis the most powerful and the most efficient for generatingoptimal yield from mutants (Baltz 1985a, b). The major limita-tion of NTG is its specificity; it produces the GC to ATtransition in about 95 % of mutations. In contrast, another ofthe most effective mutagenic agents is UV radiation, which is avery convenient and relatively safe mutagen. UV induces abroad spectrum of point mutations in E. coli (Miller 1983) butappears to be a weak mutagen in Streptomyces fradiae (Baltz1985a, b). However, many antibiotics are obtained from varioushyper-producing mutants after treating Streptomyces spp. withUV light (Lee and Rho 1999; Cheng et al. 2001; Abbass 2000).In particular, Zhu et al. (2010) achieved threefold improvementin rapamycin productionwith Streptomyces hygroscopicusHD-04-S after mutagenesis using only UV light. Our results alsoshowed that UV light was a more effective mutagenic agent incontrast to each of the 3,500 colonies generated from NTG andUV mutagenesis. This study showed that each strain and mu-tagen may interact in a different manner.
Oil plays a potential role in polyketide antibiotic produc-tion in Streptomyces spp. (Li et al. 2004; Reeves et al. 2006,2007; Mo et al. 2009). In particular, oil supplement has aremarkable effect on enhancing both FK506 titer and thespecific FK506 production per cell weight (Kim and Park2007; Mo et al. 2009). Furthermore, Kim and Park (2007)reported that improved FK506 productivity of FK506-overproducing mutant strains is associated with increasedlipase activity. Lipases (EC 3.1.1.3; triacylglycerol acylhydro-lases) are hydrolytic enzymes that catalyze cleavage of theester bonds of long-chain acylglycerols to glycerol and freefatty acids (Chahinian et al. 2002). A common consensus ofopinion exists on the positive effects of fatty acids on antibi-otic production via the β-oxidation pathway. Fatty acids areintroduced into cells to produce acetyl-CoA or propionyl-CoA, which are precursors for biosynthesizing polyketideantibiotics (Reynolds and Demain 1997). Propionyl-CoA is
0
40
80
120
160
200
Control Tween 80
Spec
ific
pro
duct
ion
of F
K50
6 (
mg/
g)
MJM7001 MJM7001/Propionate MJM7001/Vinyl Propionate
0
50
100
150
200
250
300
0 1 2 3 4 5 6
Spec
ific
pro
duct
ion
of F
K50
6 (
mg/
g)
Time (days)
a
b
Fig. 4 Effect of precursors supply and PCC pathway overexpressionin the RM7011 strain. a Specific FK506 production per cell weight bypropionate and vinyl propionate in the RM7011 strain unsupplementedor supplemented with 5 mM Tween 80. Checkerboard pattern, specificFK506 production in R2YE medium with or without Tween 80;diagonal brick pattern, specific FK506 production following propio-nate supplementation in R2YE medium with or without Tween 80;dotted square pattern, specific FK506 production by vinyl propionatesupplementation in R2YE medium with or without Tween 80. b Timecourse for specific FK506 production per cell weight by PCC pathwayoverexpression and supplementation of exogenous 5-mM vinyl propi-onate in R2YE medium supplemented with 5 mM Tween 80. Dia-mond, RM7011/pSJ1003 with vinyl propionate; square, RM7011/pSE34 with vinyl propionate. Values are the averages of two seriesof duplicate experiments. Error bars represent one standard deviationfor data obtained from two series of duplicate experiments
3060 Appl Microbiol Biotechnol (2013) 97:3053–3062

an intermediate product produced through a variety of path-ways including CoA ligation of propionate and catabolism ofodd chain fatty acids. It has been demonstrated that propionyl-CoA provides the starter unit, and (2S)-methylmalonyl-CoAprovides the extender units required for PKS-catalyzed forma-tion of polyketide antibiotics (Xue et al. 1998; Marsden et al.1994; Bao et al. 1999; Jung et al. 2011). As methylmalonyl-CoA is a limiting precursor of FK506 and rapamycin biosyn-thesis, supporting its metabolism could have an impact onFK506 and rapamycin production. Yoon and colleaguesreported that the MCM and PCC pathways are engineeredinto S. clavuligerus CKD1119 and S. hygroscopicus UN2-2for enhanced FK506 and rapamycin productions (Mo et al.2009; Jung et al. 2011). Their UV2/PCC strain was grown inmedium supplemented with 1 mM propionate for thepropionyl-CoA-dependent PCC pathway, and UV2/PCCstrain successfully increased methylmalonyl-CoA pooland rapamycin production. However, adding more than1 mM suppressed cell growth of UV2/PCC strain, con-sequently, decreased rapamycin production. We usedvinyl propionate as a precursor to convert propionyl-CoA to methylmalonyl-CoA by the PCC pathway inthe RM7011 strain with lipase activity to overcome thisproblem. The results from both the supplementation ofvinyl propionate and induction of lipase activity indicate thatvinyl propionate is a good precursor of methylmalonyl-CoAvia propionyl-CoA in an RM7011 strain with lipase activity.Additionally, stimulating lipase production by Tween 80 andoverexpressing the PCC pathway by plasmid-based expres-sion dramatically increased the specific FK506 productionlevel per cell weight. These results demonstrate that the PCCpathway is the most effective supply route for methylmalonyl-CoA to increase FK506 production in the RM7011 strain,contrary to previous results (Mo et al. 2009). In addition, vinylpropionate supplementation in culture broth promoted theenhancement of methylmalonyl-CoA through the PCC path-way without a significant suppression in cell mass. Thisresult also shows that short-chain esters such as vinylacetate, vinyl propionate, and vinyl butyrate can serveas limiting precursors for polyketide antibiotics biosyn-thesis in strains with lipase activity. In cases of strainswithout lipase activity, we suggest that short-chainesters can be used as a precursor by heterologous ex-pression of the lipase gene using expression or integra-tion plasmids. The strategy described herein suggests anew mode of precursor supply involving short-chain estersand lipase activity, even though more detailed mechanismsneed to be examined through additional experiments, such asan incorporation analysis using a radioisotope substrate.
Acknowledgement This work was supported by a grant from theNext-Generation BioGreen 21 Program (No. PJ008093 andPJ008333), Rural Development Administration, Republic of Korea.
References
Abbass RN (2000) Genetical studies for rifamycin antibiotic over-production in Amycolatopseis mediteranei. Dissertation. AinShams University, Cairo
August PR, Tang L, Yoon YJ, Ning S, Müller R, Yu TW, Taylor M,Hoffmann D, Kim CG, Zhang X, Hutchinson CR, Floss HG(1998) Biosynthesis of the ansamycin antibiotic rifamycin: deduc-tions from the molecular analysis of the rif biosynthetic genecluster of Amycolatopsis mediterranei S699. Chem Biol 5:69–79
Baltz RH (1986a) Mutagenesis in Streptomyces. In: Demain AL,Soloman NA (eds) Manual of industrial microbiology and bio-technology. American Society for Microbiology, Washington,pp 184–190
Baltz RH (1986b) Mutagenesis in Streptomyces. In: Queener SW, DayLE (eds) The bacteria, vol 9, Antibiotics-producing Streptomyces.Academic, New York, pp 61–93
Baltz RH (1999) Mutagenesis. In: Flickinger MC, Drew SW (eds)Encyclopedia of bioprocess technology: Fermentation, biocataly-sis, and separation. Wiley, New York, pp 1819–1822
Baltz RH, Stonesifer J (1985a) Adaptive response and enhancement ofN-methyl-N'-nitro-N-nitrosoguanidine mutagenesis by chloram-phenicol in Streptomyces fradiae. J Bacteriol 164:944–946
Baltz RH, Stonesifer J (1985b) Mutagenic and error-free DNA repair inStreptomyces. Mol Gen Genet 200:351–355
Bao W, Sheldon PJ, Wendt-Pienkowski E, Hutchinson CR (1999) TheStreptomyces peucetius dpsC gene determines the choice of starterunit in biosynthesis of the daunorubicin polyketide. J Bacteriol181:4690–4695
Borodina I, Krabben P, Nielsen J (2005) Genome-scale analysis ofStreptomyces coelicolor A3(2) metabolism. Genome Res 15:820–829
Cameron DC, Tong IT (1993) Cellular and metabolic engineering.Appl Biochem Biotechnol 38:105–140
Chahinian H, Nini L, Boitard E, Dubès JP, Comeau LC, Sarda L (2002)Distinction between esterases and lipases: a kinetic study withvinyl esters and TAG. Lipids 37:653–662
Chen X, Wei P, Fan L, Yang D, Zhu X, Shen W, Xu Z, Cen P (2009)Generation of high-yield rapamycin-producing strains throughprotoplasts-related techniques. Appl Microbiol Biotechnol83:507–512
Chen Y, Smanski MJ, Shen B (2010) Improvement of secondarymetabolite production in Streptomyces by manipulating pathwayregulation. Appl Microbiol Biotechnol 86:19–25
Cheng YR, Huang J, Qiang H, Lin WL, Demain AL (2001)Mutagenesis of the rapamycin producer Streptomyces hygrosco-picus FC904. J Antibiot (Tokyo) 54:967–972
Delić V, Hopwood DA, Friend EJ (1970) Mutagenesis by N-methyl-N'-nitro-N-nitrosoguanidine (NTG) in Streptomyces coelicolor.Mutat Res 9:167–182
Gold BG (2000) Neuroimmunophilin ligands: evaluation of their ther-apeutic potential for the treatment of neurological disorders.Expert Opin Investig Drugs 9:2331–2342
Gust B, Challis GL, Fowler K, Kieser T, Chater KF (2003) PCR-targeted Streptomyces gene replacement identifies a protein do-main needed for biosynthesis of the sesquiterpene soil odor geo-smin. Proc Natl Acad Sci U S A 100:1541–1546
Hertweck C (2009) The biosynthetic logic of polyketide diversity.Angew Chem Int Ed Engl 48:4688–4716
Hopwood DA (1997) Genetic contributions to understanding polyke-tide synthases. Chem Rev 97:2465–2498
Hopwood DA, Bibb MJ, Chater KF, Keiser T, Bruton CJ, Keiser HM,Lydiate DJ, Smith CP, Ward JM, Schrempf H (1985) Geneticmanipulation of Streptomyces: a laboratory manual. John InnesInstitute Foundation, Norwich
Appl Microbiol Biotechnol (2013) 97:3053–3062 3061

Horswill AR, Escalante-Semerena JC (1999) The prep gene ofSalmonella typhimurium LT2 encodes propionyl-CoA synthase.Microbiology 145:1381–1388
Iwasa T, Yamamoto H, Shibata M (1970) Studies on validamycins, newantibiotic, Streptomyces hygroscopicus var. limoneus nov. var., val-damycin producing organism. J Antibiot (Tokyo) 12:596–602
Jung S, Moon S, Lee K, Park YJ, Yoon S, Yoo YJ (2009) Straindevelopment of Streptomyces sp. for tacrolimus production usingsequential adaptation. J Ind Microbiol Biotechnol 36:1467–1471
Jung WS, Yoo YJ, Park JW, Park SR, Han AR, Ban YH, Kim EJ, KimE, Yoon YJ (2011) A combined approach of classical mutagenesisand rational metabolic engineering improves rapamycin biosyn-thesis and provides insights into methylmalonyl-CoA precursorsupply pathway in Streptomyces hygroscopicus ATCC 29253.Appl Microbiol Biotechnol 91:1389–1397
Khaliq S, Akhtar K, Afzal Ghauri M, Iqbal R, Mukhtar Khalid A,Muddassar M (2009) Change in colony morphology and kineticsof tylosin production after UV and gamma irradiation mutagenesisof Streptomyces fradiae NRRL-2702. Microbiol Res 164:469–477
Kieser T, Bibb MJ, Buttner MJ, Chater KF, Hopwood DA (2000)Practical Streptomyces genetics. John Innes Foundation, Norwich
Kim HS, Park YI (2007) Lipase activity and tacrolimus production inStreptomyces clavuligerus CKD 1119 mutant strains. J MicrobiolBiotechnol 17:1638–1644
Kino T, Hatanaka H, Hashimoto M, Nishiyama M, Goto T, Okuhara M,Kohsaka M, Aoki H, Imanaka H (1987a) FK-506, a novel immu-nosuppressant isolated from a Streptomyces. I. Fermentation, isola-tion, and physico-chemical and biological characteristics. J Antibiot(Tokyo) 40:1249–1255
Kino T, Hatanaka H, Miyata S, Inamura N, Nishiyama M, Yajima T,Goto T, Okuhara M, Kohsaka M, Aoki H, Ochiai T (1987b) FK-506, a novel immunosuppressant isolated from a Streptomyces. II.Immunosuppressive effect of FK-506 in vitro. J Antibiot (Tokyo)40:1256–1265
Kumar MS, Kumar PM, Sarnaik HM, Sadhukhan AK (2000) A rapidtechnique for screening of lovastatin-producing strains ofAspergillus terreus by agar plug and Neurospora crassa bioassay.J Microbiol Methods 40:99–104
Kumar P, Malviya HK, Maurya RK, Shukla A (2007) Fermentationprocesses for the preparation of tacrolimus. International publica-tion patent no. WO2007/039816 A2
Lee SH, Rho YT (1999) Improvement of tylosin fermentation by muta-tion and medium optimization. Lett Appl Microbiol 28:142–144
Li C, Florova G, Akopiants K, Reynolds KA (2004) Crotonyl-coenzyme A reductase provides methylmalonyl-CoA precursorsfor monensin biosynthesis by Streptomyces cinnamonensis in anoil-based extended fermentation. Microbiology 150:3463–3472
Locci R (1989) Streptomyces and related genera. In: Garrity G (ed)Bergey's manual of systematic bacteriology, vol 4. Williams &Wilkins Company, Baltimore, pp 2451–2508
Lu Y, Lin F, Wang X, Bie X, Sun H, Wuyundalai LZ (2009)Identification of bacteria producing a thermophilic lipase withpositional non-specificity and characterization of the lipase.Anal Microbiol 59:565–571
Marsden AF, Caffrey P, Aparicio JF, Loughran MS, Staunton J, LeadlayPF (1994) Stereospecific acyl transfers on the erythromycin-producing polyketide synthase. Science 263:378–380
Miller J (1983) Mutational specificity in bacteria. Annu Rev Genet17:215–238
Mo S, Ban YH, Park JW, Yoo YJ, Yoon YJ (2009) Enhanced FK506production in Streptomyces clavuligerus CKD1119 by engineeringthe supply of methylmalonyl-CoA precursor. J Ind MicrobiolBiotechnol 36:1473–1482
Mo S, Kim DH, Lee JH, Park JW, Basnet DB, Ban YH, Yoo YJ, ChenSW, Park SR, Choi EA, Kim E, Jin YY, Lee SK, Park JY, Liu Y,Lee MO, Lee KS, Kim SJ, Kim D, Park BC, Lee SG, Kwon HJ,Suh JW, Moore BS, Lim SK, Yoon YJ (2011) Biosynthesis of theallylmalonyl-CoA extender unit for the FK506 polyketide syn-thase proceeds through a dedicated polyketide synthase and facil-itates the mutasynthesis of analogues. J Am Chem Soc 133:976–985
Motamedi H, Shafiee A (1998) The biosynthetic gene cluster for themacrolactone ring of the immunosuppressant FK506. Eur JBiochem 256:528–534
Motamedi H, Cai SJ, Shafiee A, Elliston KO (1997) Structural orga-nization of a multifunctional polyketide synthase involved in thebiosynthesis of the macrolide immunosuppressant FK506. Eur JBiochem 244:74–80
Murli S, Kennedy J, Dayem LC, Carney JR, Kealey JT (2003)Metabolic engineering of Escherichia coli for improved 6-deoxyerythronolide B production. J Ind Microbiol Biotechnol30:500–509
Nielsen J (2001) Metabolic engineering. Appl Microbiol Biotechnol55:263–283
Park JW, Jung WS, Park SR, Park BC, Yoon YJ (2007) Analysis ofintracellular short organic acid-coenzyme A esters from actino-mycetes using liquid chromatography-electrospray ionization-mass spectrometry. J Mass Spectrom 42:1136–1147
Reeves AR, Cernota WH, Brikun IA, Wesley RK, Weber JM (2004)Engineering precursor flow for increased erythromycin produc-tion in Aeromicrobium erythreum. Metab Eng 6:300–312
Reeves AR, Brikun IA, Cernota WH, Leach BI, Gonzalez MC, WeberJM (2006) Effects of methylmalonyl-CoA mutase gene knockoutson erythromycin production in carbohydrate-based and oil-basedfermentations of Saccharopolyspora erythraea. J Ind MicrobiolBiotechnol 33:600–609
Reeves AR, Brikun IA, Cernota WH, Leach BI, Gonzalez MC, WeberJM (2007) Engineering of the methylmalonyl-CoA metabolitenode of Saccharopolyspora erythraea for increased erythromycinproduction. Metab Eng 9:293–303
Reynolds KA, Demain AL (1997) Rapamycin, FK506 and ascomycinrelated compounds. In: Strohl WR (ed) Biotechnology of anti-biotics, 2nd edn. Dekker, New York, p 497
Rokem JS, Lantz AE, Nielsen J (2007) Systems biology of antibioticproduction by microorganisms. Nat Prod Rep 24:1262–1287
Sambrook J, Russell DW (2001) Molecular cloning: a laboratorymanual, 3rd edn. Cold Spring Harbor, New York
Shirling EB, Gottlieb D (1966) Methods for characterization ofStreptomyces species. Int J Syst Bacteriol 16:313–340
Sierra-Paredes G, Sierra-Marcuño G (2008) Ascomycin and FK506:pharmacology and therapeutic potential as anticonvulsants andneuroprotectants. CNS Neurosci Ther 14:36–46
Smirnova N, Reynolds KA (2001) Branched-chain fatty acid biosyn-thesis in Escherichia coli. J Ind Microbiol Biotechnol 27:246–251
Xu ZN, Shen WH, Chen XY, Lin JP, Cen PL (2005) A high-throughputmethod for screening of rapamycin-producing strains ofStreptomyces hygroscopicus by cultivation in 96-well microtiterplates. Biotechnol Lett 27:1135–1140
Xue Y, Zhao L, Liu HW, Sherman DH (1998) A gene cluster formacrolide antibiotic biosynthesis in Streptomyces venezuelae:architecture of metabolic diversity. Proc Natl Acad Sci U S A95:12111–12116
Zhu X, Zhang W, Chen X, Wu H, Duan Y, Xu Z (2010) Generation ofhigh rapamycin producing strain via rational metabolic pathway-based mutagenesis and further titer improvement with fed-batchbioprocess optimization. Biotechnol Bioeng 107:506–515
3062 Appl Microbiol Biotechnol (2013) 97:3053–3062




















