
SPEKTROSKOPI RESONANSI MAGNET INTI (NMR = NUCLEAR MAGNETIC RESONANCE)
Upload
independentCategory
view
1download
0
REVIEW ARTICLE
Application of magnetic resonance imaging in zoology
Alexander Ziegler • Martin Kunth •
Susanne Mueller • Christian Bock • Rolf Pohmann •
Leif Schroder • Cornelius Faber • Gonzalo Giribet
Received: 26 July 2011 / Revised: 21 September 2011 / Accepted: 22 September 2011 / Published online: 13 October 2011! Springer-Verlag 2011
Abstract Magnetic resonance imaging (MRI) is a non-invasive imaging technique that today constitutes one of
the main pillars of preclinical and clinical imaging. MRI’s
capacity to depict soft tissue in whole specimens ex vivo aswell as in vivo, achievable voxel resolutions well below
(100 lm)3, and the absence of ionizing radiation have
resulted in the broad application of this technique both inhuman diagnostics and studies involving small animal
model organisms. Unfortunately, MRI systems are expen-
sive devices and have so far only sporadically been used toresolve questions in zoology and in particular in zoomor-
phology. However, the results from two recent studies
involving systematic scanning of representative speciesfrom a vertebrate group (fishes) as well as an invertebrate
taxon (sea urchins) suggest that MRI could in fact be used
more widely in zoology. Using novel image data derivedfrom representative species of numerous higher metazoan
clades in combination with a comprehensive literature
survey, we review and evaluate the potential of MRI forsystematic taxon scanning. According to our results,
numerous animal groups are suitable for systematic MRI
scanning, among them various cnidarian and arthropodtaxa, brachiopods, various molluscan taxa, echinoderms, as
well as all vertebrate clades. However, various phyla in
their entirety cannot be considered suitable for thisapproach mainly due to their small size (e.g., Kinorhyncha)
or their unfavorable shape (e.g., Nematomorpha), while
other taxa are prone to produce artifacts associated eitherwith their biology (e.g., Echiura) or their anatomy (e.g.,
Polyplacophora). In order to initiate further uses of MRI in
zoology, we outline the principles underlying variousapplications of this technique such as the use of contrast
agents, in vivo MRI, functional MRI, as well as magneticCommunicated by T. Bartolomaeus.
A. Ziegler (&) ! G. GiribetMuseum of Comparative Zoology, Department of Organismicand Evolutionary Biology, Harvard University,26 Oxford Street, Cambridge, MA 02138, USAe-mail: [email protected]
G. Giribete-mail: [email protected]
M. Kunth ! L. SchroderLeibniz-Institut fur Molekulare Pharmakologie,Robert-Rossle-Strasse 10, 13125 Berlin, Germanye-mail: [email protected]
L. Schrodere-mail: [email protected]
S. MuellerCentrum fur Schlaganfallforschung, Charite-UniversitatsmedizinBerlin, Chariteplatz 1, 10117 Berlin, Germanye-mail: [email protected]
C. BockAlfred-Wegener-Institut fur Polar- und Meeresforschung,Am Handelshafen 12, 27570 Bremerhaven, Germanye-mail: [email protected]
R. PohmannMax-Planck-Institut fur Biologische Kybernetik,Spemannstr. 41, 72076 Tubingen, Germanye-mail: [email protected]
C. FaberInstitut fur Klinische Radiologie, Universitatsklinikum Munster,Albert-Schweitzer-Campus 1, 48149 Munster, Germany
123
Zoomorphology (2011) 130:227–254
DOI 10.1007/s00435-011-0138-8

resonance spectroscopy. Finally, we discuss how future
technical developments might shape the use of MRI for the
study of zoological specimens.
Keywords MRI ! High-throughput ! Noninvasive !Metazoa ! Three-dimensional ! NMR
Abbreviations2D Two-dimensional
3D Three-dimensionalBBB Blood–brain barrier
BOLD Blood oxygenation level-dependent
CA Contrast agentcLSM Confocal laser scanning microscopy
CSI Chemical shift imaging
CT Computed tomographyDTI Diffusion tensor imaging
DWI Diffusion-weighted imaging
FLASH Fast low-angle shotFMNH Field Museum of Natural History
fMRI Functional magnetic resonance imagingFOV Field of view
FR RARE factor
FSPGR Fast spoiled gradient echoMEMRI Manganese-enhanced magnetic resonance
imaging
MR Magnetic resonanceMRI Magnetic resonance imaging
MRS Magnetic resonance spectroscopy
NA Average numberNMR Nuclear magnetic resonance
OPT Optical projection tomography
PET Positron emission tomographyRARE Rapid acquisition with relaxation enhancement
SE Spin echo
SIO Scripps Institution of OceanographySNR Signal-to-noise ratio
TA Acquisition time
TE Echo timeTR Repetition time
TSE Turbo spin echo
lCT Micro-computed tomographyZMB Zoologisches Museum Berlin
ZMH Zoologisches Museum Hamburg
Introduction
Magnetic resonance imaging (MRI), a noninvasive imag-
ing technique based on the principle of nuclear magnetic
resonance (NMR), constitutes one of the main pillars ofpreclinical and clinical imaging (Baker 2010; Walter et al.
2010). The importance of MRI is based on its capacity to
depict hard and in particular soft structures at relatively
high resolutions (Tyszka et al. 2005), but without the useof ionizing radiation as would be the case in other
noninvasive imaging techniques such as computed
tomography (CT) or positron emission tomography (PET).Although various elements such as fluorine, carbon,
helium, or phosphorous can be detected using MRI,
hydrogen is by far the most frequent element to be targetedas it is abundantly present in all biological samples and
even in fossils (Mietchen et al. 2005, 2008a). Today’sbroad application of MRI in life sciences is reflected in the
recent publication of various books that summarize and
extend the current knowledge on specific MRI applicationssuch as, for example, magnetic resonance neuroimaging
(Modo and Bulte 2011), small animal imaging (Kiessling
et al. 2011), or in vivo NMR imaging (Schroder andFaber 2011).
Undoubtedly, MRI scanning of metazoans other than
humans focuses on model organisms. In the last two dec-ades, numerous animal species have become suitable
model organisms for MRI studies, both in vivo as well as
ex vivo. These organisms encompass almost exclusivelyvertebrate taxa, in particular the mouse (Mus musculusLinnaeus, 1758), rat (Rattus norvegicus (Berkenhout,
1769)), zebra finch (Taeniopygia guttata (Vieillot, 1817)),pig (Sus scrofa Linnaeus, 1758), zebra fish (Danio rerio(Hamilton, 1822)), as well as various smaller primate
species such as the rhesus macaque (Macaca mulatta(Zimmermann, 1780)), the capuchin monkey (Cebuscapucinus (Linnaeus, 1758)), and the marmoset monkey
(Callithrix jacchus (Linnaeus, 1758)). However, sometraditional invertebrate model organisms such as the fruit
fly (Drosophila melanogasterMeigen, 1830) and the honey
bee (Apis mellifera Linnaeus, 1758) have recently beenadded to this list (Haddad et al. 2004; Null et al. 2008).
Apart from its use in model organisms, the application
of MRI in studies with a primarily zoological focus datesback to the late 1980s when the first anatomical MRI
images of invertebrate specimens were published (Gassner
and Lohmann 1987; Lohmann and Gassner 1987; Conneret al. 1988). However, it took several years before
improved hardware and software permitted the successful
scanning of small animals at resolutions that would allow adifferentiation of anatomical structures in organisms of
only a few centimeters in size (Hart et al. 2003).
Due to the capacity of MRI to depict in particular softtissue with excellent contrast (Benveniste and Blackband
2006), MRI has seen application also in two other fields
involving the study of zoological specimens: veterinaryradiology and food science. For diagnostic purposes, MRI
is frequently used on animal patients such as dogs, cats,
and horses (Assheuer and Sager 1997; Gavin and Bagley2010; Elliott and Skerritt 2010; Murray 2011), but exotic
228 Zoomorphology (2011) 130:227–254
123

pets are frequently scanned as well (Straub and Jurina
2001; Valente et al. 2006). In contrast to veterinary radi-ology, the use of magnetic resonance (MR) techniques in
food science is aimed at noninvasively assessing the
quality of preprocessed as well as processed food intendedfor human consumption. In the course of these studies,
primarily fish species such as the Atlantic herring (Clupeaharengus Linnaeus, 1758), Atlantic salmon (Salmo salarLinnaeus, 1758), Atlantic cod (Gadus morhua Linnaeus,
1758), and steelhead trout (Salmo gairdneri Richardson,1836) have been scanned (Webb et al. 2001; Belton et al.
2003, 2005; Farhat et al. 2007; Gujonsdottir et al. 2009;
Renou et al. 2011). As small animal scanners have becomemore widely distributed, smaller culinary resources such as
oysters (Crassostrea gigas (Thunberg, 1793) and Ostreaedulis Linnaeus, 1758) are being routinely investigatednow as well (Davenel et al. 2006, 2010; Pouvreau et al.
2006).
Despite a growing number of successful studies based onthe application of MRI in zoology, this technology cannot
yet be considered a mainstream tool in studies on non-
model animal organisms. The principal reason for this iscertainly the perceived limited access to scanning systems
in combination with often prohibitive costs of operation.
However, despite these obstacles, an increasing number ofMR studies with a strictly zoological background are being
undertaken. Even major logistic efforts such as the sys-
tematic scanning of representative species from two animalgroups, namely actinopterygiids and other fishes (The
Digital Fish Library, http://www.digitalfishlibrary.org) as
well as echinoids (Ziegler et al. 2008; Ziegler in press) havebeen performed using MRI, demonstrating that large num-
bers of specimens can be analyzed with this technique.
More importantly, these exemplary high-throughput pro-jects were performed by tapping into a vast, yet largely
under-utilized resource, the worldwide collections of alco-
hol-preserved museum specimens. MRI’s capacity to non-invasively provide spatial information of whole specimen
internal anatomy was key to the large-scale use of museum
material (Corfield et al. 2008; Chanet et al. 2009) andresulted in a broad taxon sampling, which would have been
impossible to achieve based on freshly collected material
alone.As the gathered MR image data are digital by nature,
advanced 3D visualization and modeling software can be
used to convey complex morphological structures in amore plastic manner (Walter et al. 2010, Boistel et al.
2011), for example by embedding fully interactive, labeled
3D models of zoological specimens into the publicationitself (Ziegler et al. 2010b, c, 2011). In addition, the
simultaneous display of several interactive datasets on a
computer screen has become possible, for example byusing the ImageJ (NIH, http://rsb.info.nih.gov/ij) Volume
Viewer plugin, thereby permitting real-time comparison of
structures between different species. This improved andaccelerated access to morphological data certainly influ-
ences the way in which anatomical information will be
processed in the future (Budd and Olsson 2007; Schmidt-Rhaesa 2009; Giribet 2010).
One of the aims of this review is to assess the general
suitability of higher metazoan clades for their systematicscanning by MRI. To this end, we provide—in many cases
for the first time—MRI data on numerous phyla and listthose species that have previously been shown to be suit-
able candidates. In order to maximize taxon coverage, this
study centers primarily on the use of freshly fixed andmuseum specimens. Furthermore, we outline the principles
of various MR applications and provide examples for their
use in zoology. These applications include the use ofcontrast agents, in vivo MRI, functional MRI, as well as
MR spectroscopy. In addition, we provide a section on
practical and logistic considerations as well as a section onartifacts that may occur during the study of zoological
specimens. We conclude by providing an outlook on how
technical improvements could shape MRI’s further use inthe study of zoological specimens, hoping that the reader
becomes inspired by the breadth of possible MR applica-
tions in zoology.
Some practical and logistic considerations
Clinical (i.e., human) MRI scanners can primarily be found
in hospitals and neurobiology research institutions, whilepreclinical (i.e., small animal) MRI systems are predomi-
nantly located in neurobiological, pharmacological, and
developmental research institutes. Figure 1 depicts a rangeof clinical and preclinical scanners with corresponding coils
(the coil surrounds the sample and receives the radio fre-
quency signal emitted after excitation of the targeted nucleiwithin the sample). Clinical (Fig. 1a) as well as preclinical
(Fig. 1c, e, i, j) scanners have been sold in the thousands
over the last decades. Some institutions have pooled theirMRI systems in a dedicated imaging facility on a fee-based
access scheme, whereas other facilities may provide access
free of charge, often after approval of a project proposal.Since MRI technology is complex, support from trained
MRI staff will be necessary in most instances, at least in the
starting phase of an imaging project. Depending on the typeof magnet used and the imaging equipment available (coils,
gradient systems, etc.), millimeter- to meter-sized speci-
mens can be imaged either in part or whole. Typically, thecoils used in preclinical scanners provide enough room to fit
entire mice or rats (Fig. 1f), whereas those found in clinical
scanners can accommodate human-sized samples or partsthereof (Fig. 1b). Some MRI facilities have the capacity to
Zoomorphology (2011) 130:227–254 229
123

custom-build coils in order to provide the best possible
signal detection for a given sample (Fig. 1k). In case of
over-sized specimens, it might be valuable to look for early(and hence usually smaller) developmental stages.
While living specimens are usually scanned in their
natural medium (i.e., in air, fresh, or saltwater), ex vivostudies require specimen preparation prior to scanning,
mostly by placing the specimen in distilled water (Ziegler
and Mueller 2011) or in a hydrogen-free liquid such asFluorinert (van der Linden et al. 2009). Although the var-
ious fixation protocols used in zoology may influence
scanning results, MRI has been successfully performed onBouin-, formalin-, and ethanol-fixed material (Chanet et al.
2009). Depending on the sample and the scanning protocol,
specimens can be scanned in water, but also in formalin orsometimes even directly in 70% ethanol. Unfortunately, no
study has yet systematically addressed the effects of
varying fixation protocols on scanning results across taxa.In addition, as MRI has the capacity to modulate tissue
contrast depending on the chosen protocol, the choice of
scanning parameters is crucial for the success of the
experiment. However, only a handful of studies provide
visual examples of the effect of differing protocol usage onother than model organisms (Brouwer et al. 1992; Ziegler
et al. 2010a; Ziegler and Mueller 2011). This means that
each taxon will usually require protocol optimizationbefore systematic scanning is initiated.
The final voxel resolution achievable depends on a
number of factors, in particular the size of the specimen,the field of view (FOV), magnet strength, gradient strength,
and the type of protocol. On preclinical scanners, voxel
resolutions of (15 lm)3–(100 lm)3 are typically obtain-able, while clinical scanners provide about (90 lm)3–
(1 mm)3 voxel resolution. Higher in-plane (i.e., x- and
y-plane) resolutions may be achievable using 2D imagingprotocols, but this comes at the expense of resolution in the
z-plane, usually with a factor of 4–8. The selected resolu-
tion and protocol in turn define the duration of the scan. Forexample, 2D single-slice scans can be performed at con-
siderably high resolutions in less than 1 min, while 3D
a b c d
e f g h
k
i
j l m
Fig. 1 Overview of MRI hardware used in the course of this study.MRI scanners can be divided into clinical (i.e., human) andpreclinical (i.e., small animal) systems, with the latter being eitherbased on horizontal or vertical bore magnets. a Human 3 T MRIscanner (Philips Achieva) and b a corresponding head coil for adults.c Small animal 4.7 T MRI scanner (Bruker Biospec) including acustom-built swim tunnel for studies on marine organisms (d).e Small animal 7 T MRI scanner (Bruker Pharmascan) for studies onmodel organisms with (f) corresponding rat and mouse coils (3.8 cm
and 1.8 cm internal diameter, respectively). g Small animal 9.4 TNMR scanner (Bruker Avance) equipped for imaging and h acorresponding double-resonant 1H/129Xe birdcage coil (3 cm internaldiameter). i Small animal 9.4 T (Bruker Biospec) and j 16.4 T (BrukerBiospec) MRI scanners. k Custom-built surface coil with1.8 9 2.4 cm dimensions as used on the 16.4 T scanning systemshown in (j). l Small animal 17.6 T NMR scanner (Bruker Avance)equipped for imaging and m a corresponding coil with 5 mm internaldiameter
230 Zoomorphology (2011) 130:227–254
123

isotropic scans of an entire sample are usually performed
overnight.Isotropic datasets in particular are suitable for 3D
reconstruction. However, in most cases, MRI image data
display too many varying gray-scale levels for simplethreshold-based rendering and will therefore require manual
segmentation efforts. Unfortunately, the fast, automated
modeling that can be performed on humans and selectedmodel organisms is based on machine-learning algorithms
that have been trained using thousands of exemplary data-sets—this amount of indispensable data is usually not
available in comparative zoological studies of non-model
organisms. But since future developments in computervisualization and pattern recognition might solve the cur-
rent problem of laborious manual 3D reconstruction, it is of
even greater importance to deposit the raw datasets andmake them accessible to the public. A number of web-based
voxel repositories are currently being developed, such as
the Digital Fish Library (http://www.digitalfishlibrary.org),the Digital Morphology Library (http://www.digimorph.org),
and the Morphological Database (http://www.morphdbase.de).
Most of the sea urchin MRI datasets, for example, areavailable for inspection and download at MorphDBase.
However, the zoological community as well as its major
funding sources has not yet agreed on a standardized pro-cess for voxel data deposition (see Ziegler et al. 2010b and
Rowe and Frank 2011 for discussion).
Suitability of higher metazoan taxa for systematic MRIscanning
Species from various metazoan clades have been scanned
in the past. Table 1 lists those species for which anatomicalMRI (as well as other NMR) data with varying degrees of
detail and quality are currently available. In addition to a
literature survey, we provide image data for numerousmetazoan taxa from Ctenophora to Vertebrata (Figs. 2, 3,
4, 5, 6, 7, 8, 9), some of which have never been analyzed
before using MRI. However, the images provided hererepresent only a single virtual 2D slice out of a multitude of
slices that were gathered for each specimen scanned in the
course of this study. These images therefore only convey arestricted impression of the structural complexity of the
specimens and more can be learned by applying the above-
mentioned interactive dataset manipulation software.Of the 65 higher metazoan clades listed here (Table 1),
fifteen were judged as unsuitable right away because of the
minute size (e.g., Kinorhyncha, Micrognathozoa, Tardi-grada) or the unfavorable shape (e.g., Nematomorpha) of
their species. Apart from size and shape considerations, the
problem of artifact-causing materials located inside speci-mens has to be taken into consideration as well. As outlined
in the following section, various artifacts can impede suc-
cessful imaging of otherwise suitable taxa (e.g., Echiura,Sipuncula, Polyplacophora). Here, we give an estimate of a
clade’s general suitability for systematic MRI scanning as
previously performed on actinopterygiids and other fishes(The Digital Fish Library) as well as echinoids (Ziegler
et al. 2008; Ziegler in press). However, based on a number
of unforeseeable factors, each individual species understudy may prove to be suitable or not. Our conclusions are
primarily based on the use of freshly fixed and museummaterial, but as outlined in one of the following sections,
further aspects have to be taken into consideration when
studying organisms in vivo using MRI.Unfortunately, MRI currently does not permit to gather
meaningful anatomical data from specimens smaller than
about 1 mm. This void in whole specimen imaging can befilled using other imaging techniques such as micro-com-
puted tomography (lCT), confocal laser scanning micros-
copy (cLSM), or optical projection tomography (OPT)(Walter et al. 2010; Boistel et al. 2011). However, in
contrast to MRI, soft tissue imaging using these techniques
may require considerable specimen preparation, forexample through staining (Metscher 2009). Nonetheless,
the zoologist nowadays has a continuum of imaging
modalities at hand that permits the imaging of wholezoological specimens of varying sizes. In this context, MRI
clearly serves the size spectrum of millimeter- to meter-
sized specimens.
Potential artifacts occurring during MRI of zoologicalspecimens
The techniques used to encode spatial information in MRIcan lead to undesirable effects that alter image information
or, even worse, may appear as faulty anatomical detail. If
possible, these artifacts should be avoided or at least becorrectly identified. Many of the artifacts occurring in MR
images are due to imperfections of the experimental pro-
tocol or the hardware employed. For instance, interferencewith radio frequency signals from other electronic devices
(e.g., when the door to the scanner room is left open) can
cause stripes in the image(s). Faulty positioning or sizing ofthe FOV can lead to aliasing, i.e., the appearance of signal
from outside of the targeted region within the image.
However, some artifacts are caused by intrinsic propertiesof the hardware used or the sample itself.
A major source of artifacts is the movement of the
sample, whether in vivo or ex vivo. An MR image isgenerated by a large number of single scans that are
combined to form the final dataset. Any movement of the
sample during this series of scans causes blurring of theimage(s): it is important to note that both the sample and
Zoomorphology (2011) 130:227–254 231
123

Table 1 Suitability of metazoan taxa for systematic scanning usingMRI. Taxon information includes the clade name, its vernacularname, the number of described or known species, and their size range(based primarily on Westheide and Rieger 2007, 2010). The species
analyzed so far using MRI (and other NMR techniques) are listed inalphabetical order within each taxon. Systematic arrangement ofclades was based on Dunn et al. (2008) and Edgecombe et al. (2011)
Taxon information Species studied so far using MRI (and otherNMR techniques)
Suitability for MRI
Ctenophora, comb jellies, *80 spp.,few mm to few m
Pleurobrachia pileus (this study): wholespecimen, ex vivo, Fig. 2a
Suitable, but many species toosmall
Placozoa, 1 sp. (many undescribed),few mm
– Unsuitable, too small
Porifera, sponges, *10.000 spp., few mmto few m
Axinella sp. (this study): whole specimen, exvivo, Fig. 2b; Lubomirskia baicalensis(Muller et al. 2006): whole specimen, exvivo; Suberites domuncula (Bringmannet al. 1999): whole specimen, in vivo;unknown species (Lee et al. 2010): wholespecimen, in vivo
Suitable, but many species toosmall
Cnidaria, anemones, jellyfishes, coralsand others (incl. Myxozoa), *10.550spp., several lm to few m
Actinia sp. (Ziegler and Mueller 2011, thisstudy): whole specimen, ex vivo, Fig. 2c;Aurelia aurita (this study): wholespecimen, ex vivo; Cyanea capillata(Blackband and Stoskopf 1990): wholespecimen, in vivo
Suitable, but some species toosmall or too large
‘‘Mesozoa’’, orthonectids andrhombozoans, *107 spp., few mm
– Unsuitable, too small
Xenoturbellida, 1 sp., few cm – Presumably unsuitable
Acoela, *380 spp., several lm to severalmm
– Unsuitable, too small
Nemertodermatida, 11 spp., several lm toseveral mm
– Unsuitable, too small
Chaetognatha, arrow worms, *150 spp.,several mm to several cm
– Some larger species likelysuitable, many too small
Nematoda, roundworms,[15.000 spp.,several lm to few m
Ascaris sp. (this study): wholespecimen, ex vivo, Fig. 2d
Some large forms suitable, butmost species too small
Nematomorpha, horsehair worms, *320spp., few cm to few m
– Unsuitable, too thin
Tardigrada, water bears, *1.030 spp.,several lm to few mm
– Unsuitable, too small
Onychophora, velvet worms, *200 spp.,few mm to several cm
Ooperipatellus sp. (this study): wholespecimen, ex vivo, Fig. 3a
Some larger forms suitable, butmany too small
Arthropoda (Chelicerata—Crustacea)
Chelicerata, arachnids and horseshoecrabs, *100.000 spp., several lm toseveral cm
Acanthoscurria geniculata (Lauridsen et al.2011): whole specimen, in vivo;Aphonopelma californica (Pohlmann et al.2007): opisthosoma, in vivo; Nephila sp.(this study): whole specimen, ex vivo,Fig. 3b
Many large forms suitable, butmany smaller species not
Pycnogonida, sea spiders, *1.000 spp.,few mm to several cm
Pycnogonum sp., (this study): wholespecimen, ex vivo, Fig. 3c
Only some large taxa suitable,body usually too small
Myriapoda, centipedes, millipedes,symphylans, and pauropods, *16.000spp., few mm to several cm
Scolopendra sp. (this study): proximal part,ex vivo, Fig. 3d
Many suitable, but many too small
232 Zoomorphology (2011) 130:227–254
123

Table 1 continued
Taxon information Species studied so far using MRI (and otherNMR techniques)
Suitability for MRI
Hexapoda, springtails, proturans,diplurans, and insects, *1.000.000spp., several lm to several cm
Apis mellifera (Tomanek et al. 1996, Haddadet al. 2004): whole specimen and head, exvivo; Ascioplaga mimeta (Hornschemeyeret al. 2006): caput and thorax, ex vivo;Bombyx mori (Mapelli et al. 1997): larva, invivo; Canthon cyanellus cyanellus (Favilaet al. 2004): whole specimen, in vivo;Coccinella septempunctata (Chudek et al.1996, Geoghegan et al. 2000): wholespecimen, in vivo; Dinocampus coccinellae(Chudek et al. 1996, Geoghegan et al.2000): whole specimen, in vivo;Dinoponera quadriceps (Fresneau et al.1991, Hart et al. 2003): whole specimen, invivo; Drosophila melanogaster (Fresneauet al. 1991, Null et al. 2008, this study):whole specimen, in and ex vivo; Dytiscusmarginalis (Wecker et al. 2002): wholespecimen, ex vivo; Epiblema scudderiana(Mietchen et al. 2008b): whole specimen, invivo; Eurosta solidaginis (Mietchen et al.2008b): whole specimen, in vivo;Graphiphora augur (Goodman et al. 1995):whole specimen, in vivo; Manduca sexta(Conner et al. 1988, Michaelis et al. 2005,Watanabe et al. 2006, Hallock 2008): headand larva, in vivo; Periplaneta sp. (Zieglerand Mueller 2011, this study): proximal anddistal part, ex vivo, Fig. 3e; Periplanetafuliginosa (Takahashi et al. 1989): wholespecimen, in vivo; Pieris brassicae(Goodman et al. 1995): whole specimen, invivo; Plodia interpunctella (Chudek et al.1996): whole specimen, in vivo; Prionuscoriarius (this study): larva, ex vivo;Sarcophaga bullata (Jasanoff and Sun2002, Jasanoff 2005): head, in vivo;Sarcophaga peregrina (Price et al. 1999):whole specimen, in vivo; Schistocercagregaria (Gassner and Lohmann 1987):whole specimen, in vivo; Solenopsis invicta(Slowik et al. 1997): whole specimen, exvivo; Spodoptera litura (Skibbe et al.1995), thorax, ex vivo; Venturia cavescens(Chudek et al. 1996): whole specimen, invivo; Vespula vulgaris (Hart et al. 2003):whole specimen, in vivo
Many species suitable, but manymore too small
Crustacea, shrimps, crabs, lobsters, andbarnacles, *50.000 spp., several lm tofew m
Callinectes sapidus (Brouwer et al. 1992):whole specimen, in vivo; Cancer pagurus(Fernandez et al. 2000): whole specimen, invivo; Cherax destructor (Brinkley et al.2005): cephalothorax, in vivo; Majasquinado (Fernandez et al. 2000, Bock et al.2001a): cephalothorax, in vivo;Pachygrapsus marmoratus (Ziegler andMueller 2011, this study): whole specimen,ex vivo, Fig. 3f; Procambarus clarkia(Herberholz et al. 2004): whole specimen,in vivo
Many species are suitable, butmany will be too small
Zoomorphology (2011) 130:227–254 233
123

Table 1 continued
Taxon information Species studied so far using MRI (and otherNMR techniques)
Suitability for MRI
Priapulida, penis worms, *20 spp.,several lm to several cm
Priapulopsis bicaudatus (this study): wholespecimen, ex vivo, Fig. 2e
Some large species suitable, butmany too small
Loricifera, *200 spp. (mostundescribed), several lm
– Unsuitable, too small
Kinorhyncha, mud dragons,[150 spp.,several lm to few mm
– Unsuitable, too small
Bryozoa, moss animals,[4.500 spp.,several lm to mm
Dendrobeania sp. (this study): colony, exvivo, Fig. 4a
Single organism too small, but thecolony can be suitable
Entoprocta,*150 spp., several lm to fewmm
– Unsuitable, too small
Cycliophora, cycliophorans, 2 spp.,several lm
– Unsuitable, too small
Annelida (Echiura—Clitellata)
Echiura, spoon worms, *150 spp., fewmm to few m
Thalassema thalassemum (this study): wholespecimen, ex vivo, Fig. 4f
Some species suitable, but mostare too small
Sipuncula, peanut worms, *150 spp.,few cm to several cm
Sipunculus sp. (this study): whole specimen,ex vivo, Fig. 4e
Some larger forms suitable, butmany species too small
Polychaeta, bristle worms, *9.000 spp.,several lm to few m
Nereis sp. (Ziegler and Mueller 2011, thisstudy): proximal part, ex vivo, Fig. 4g
Some taxa suitable, but manytoo small
Clitellata, earthworms and leeches,[4.000 spp., few mm to few m
Hirudo medicinalis (this study): proximalpart, ex vivo, Fig. 4h
Some taxa suitable, but manytoo small and too thin
Mollusca (Solenogastres—Cephalopoda)
Solenogastres, *250 spp., few mm toseveral cm
– Some species likely suitable, butmost are too small
Caudofoveata, *70 spp., few mm to fewcm
– Some species likely suitable, butmany are too small
Monoplacophora, 25 spp., several lm tofew cm
– Some taxa likely suitable, but mostare too small
Polyplacophora, chitons, *900 spp., fewmm to several cm
Acanthochitona sp. (this study): wholespecimen, ex vivo, Fig. 5a; 22 furtherspecies were scanned by us in 2009, alldisplaying strong susceptibility artifactssurrounding the ferromagnetic radula
Unsuitable because of magnetite-bearing radula
Gastropoda, snails and slugs, *100.000spp., several lm to several cm
Achatina sp. (Cooper 2011): whole specimen,in vivo; Aplysia californica (Hsu et al.1996, Grant et al. 2000, Neustadter et al.2002, Novakovic et al. 2006): buccal massand single neuron, in and ex vivo;Dendronotus sp. (Ziegler and Mueller 2011,this study): whole specimen, ex vivo,Fig. 5b
Many species likely suitable, butmany are too small
Scaphopoda, tusk shells, *600 spp., fewmm to few cm
– Some taxa suitable, but many aretoo small
234 Zoomorphology (2011) 130:227–254
123

Table 1 continued
Taxon information Species studied so far using MRI (and otherNMR techniques)
Suitability for MRI
Bivalvia, bivalves, *15.000 spp., severallm to few m
Anodonta sp. (Synthesys NA F Pilot Study,http://www.synthesys.info/na_f.htm):whole specimen, in and ex vivo; Arcopsisadamsi FMNH 3 (this study): whole spec-imen, ex vivo; Argopecten purpuratus (vonBrand et al. 2009): whole specimen, invivo; Barbatia cancelaria FMNH 348 (thisstudy): whole specimen, ex vivo; Brachi-dontes exustus FMNH 243 (this study):whole specimen, ex vivo; Cerastodermaedule (this study): whole specimen, ex vivo,Fig. 5c; Corbicula fluminea FMNH 242(this study): whole specimen, ex vivo;Crassostrea gigas (Davenel et al. 2006,Pouvreau et al. 2006): whole specimen, invivo; Crassostrea virginica (Lannig et al.2008, Lee et al. 2010): whole specimen, invivo; Cyrenoida floridana FMNH 6 (thisstudy): whole specimen, ex vivo; Elliptiocomplanata (Holliman et al. 2008): wholespecimen, in vivo; Gemma gemma FMNH357 (this study): whole specimen, ex vivo;Histella arctica FMNH 264 (this study):whole specimen, ex vivo; Margaritiferaauricularia (Synthesys NA F Pilot Study,http://www.synthesys.info/na_f.htm):whole specimen, in and ex vivo; Mytilusedulis (this study): whole specimen, exvivo; Ostrea edulis (Davenel et al. 2010):whole specimen, in vivo; Solemya velumFMNH 358 (this study): whole specimen,ex vivo; Teredo clappi FMNH 2 (thisstudy): whole specimen, ex vivo; Yoldialimatula FMNH 359 (this study): wholespecimen, ex vivo
Many species are suitable, butsome will be too small
Cephalopoda, cephalopods, *1.000 spp.,few cm to several m
Loligo pealeii (Mooney et al. 2010): wholespecimen, in vivo; Lolliguncula brevis(Gozansky et al. 2003): receptor organ, exvivo; Sepia sp. (this study): wholespecimen, ex vivo, Fig. 5d; Sepia officinalis(Quast et al. 2001): brain, ex vivo
Mostly suitable, but a few taxa willbe too small
Nemertea, ribbon worms, *1.100 spp.,few mm to several m
Lineus sp. (this study): whole specimen, exvivo, Fig. 4c; Dinonemertes cf.investigatoris (this study): median part, exvivo
Some large forms suitable, butmany species too small
Brachiopoda, lamp shells, *380 spp.,few mm to few cm
Terebratalia transversa (this study): wholespecimen, ex vivo, Fig. 4d
Suitable, some species will be toosmall
Phoronida, horseshoe worms, *14 spp.,few mm to several cm
– Some taxa likely suitable, butsome species too small
Gastrotricha, hairy backs,[500 spp.,several lm to few mm
– Unsuitable, too small
Platyhelminthes, flatworms, cestodes,trematodes, and others,[22.120 spp.,few mm to several m
Fasciola hepatica (this study): wholespecimen, ex vivo, Fig. 4b
Some species suitable, but manytoo small
Gnathostomulida, jaw worms,\100 spp.,several lm to few mm
– Unsuitable, too small
Micrognathozoa, 1 sp., several lm – Unsuitable, too small
Zoomorphology (2011) 130:227–254 235
123

Table 1 continued
Taxon information Species studied so far using MRI (and otherNMR techniques)
Suitability for MRI
Rotifera (incl. Acanthocephala), wheelanimals and thorny-headed worms,*3.100 spp., several lm to few cm
– Typical rotifers are too small, butsome acanthocephalans may besuitable
Myzostomida, myzostomids, *140 spp.,few mm
– Unsuitable, too small
Echinodermata (Crinoidea—Echinoidea)
Crinoidea, feather stars and sea lilies,*620 spp., few cm to few m
Antedon mediterranea ZMH E6859 (Zieglerin press, this study): whole specimen, exvivo, Fig. 6a
Partly suitable, but body will betoo small in many species
Asteroidea, sea stars, *1.600 spp., fewcm to few m
Acanthaster planci (this study): wholespecimen, in and ex vivo, Fig. 6b; Asteriasrubens (this study): whole specimen, exvivo; Asterina gibbosa ZMH E1195(Ziegler in press, this study): wholespecimen, ex vivo
Suitable, but some species will betoo small
Ophiuroidea, brittle stars, *2.000 spp.,few mm to several cm
Ophiocoma nigra ZMH E2025 (Ziegler inpress, this study): whole specimen, ex vivo,Fig. 6c
Suitable, but some taxa will be toosmall
Holothuroidea, sea cucumbers, *1.200spp., few mm to few m
Aslia lefevrei (Ziegler in press, this study):whole specimen, ex vivo, Fig. 6d;Echinopsolus acanthocola (this study):whole specimen, ex vivo
Suitable, but some will be toosmall
Echinoidea, sea urchins, sand dollars,and heart urchins, *950 spp., few mmto several cm
94 Species: Ziegler (in press) provides a fulllist; Clypeaster rosaceus ZMB Ech 2520(this study): whole specimen, ex vivo,Fig. 6e
Suitable, but some taxa showstrong artifacts (sediment)
Hemichordata, acorn worms andpterobranchs, *101 spp., few mm tofew m
Harrimania kupferi ZMB Ent 1579 (thisstudy): whole specimen, ex vivo, Fig. 6f
Many suitable, but some will betoo small
Urochordata, tunicates, salps, and others,*2.120 spp., few mm to several m
Ciona intestinalis (Ziegler and Mueller 2011,this study): whole specimen, ex vivo,Fig. 6g
Many suitable, but some will betoo small or too large
Cephalochordata, lancelets, 29 spp., fewcm
Branchiostoma lanceolatum (this study):whole specimen, ex vivo, Fig. 7a
Suitable, some are too small
Vertebrata (Myxini—Mammalia)
Myxini, hagfish, *60 spp., several cm tofew m
Eptatretus stoutii (The Digital Fish Library):whole specimen, ex vivo, Fig. 7b
Suitable
Petromyzontida, lampreys, 42 spp.,several cm to few m
Petromyzon marinus (The Digital FishLibrary): whole specimen, ex vivo, Fig. 7c
Suitable
Chondrichthyes, cartilaginous fishes,[1.100 spp., several cm to several m
46 species: The Digital Fish Library;Chiloscyllium arabicum (Blackband andStoskopf 1990): torso, in vivo; Isurusoxyrinchus (Perry et al. 2007): torso, exvivo; Lamna ditropis (Perry et al. 2007):torso, ex vivo; Squalus acanthias (thisstudy): head, ex vivo; Triakis semifasciata(this study): whole specimen, ex vivo,Fig. 7d
Suitable
Sarcopterygii, lobe-finned fishes, 8 spp.,few m
4 species: The Digital Fish Library;Latimeria chalumnae (The Digital FishLibrary): head and torso, ex vivo, Fig. 7e
Suitable
236 Zoomorphology (2011) 130:227–254
123

Table 1 continued
Taxon information Species studied so far using MRI (and otherNMR techniques)
Suitability for MRI
Actinopterygii, ray-finned fishes,*30.000 spp., several mm to few m
222 species: The Digital Fish Library;Acanthostracion quadricornis (Boistel et al.2011): whole specimen, ex vivo; Agonuscataphractus (this study): head, ex vivo,Fig. 7f; Alloclinus holderi (Rowe and Frank2011): whole, ex vivo; Clupea harengus(Veliyulin et al. 2007): thorax, ex vivo;Crassius sp. (Blackband and Stoskopf1990): torso, in vivo; Cyprinus carpio (Vander Linden et al. 2004, Van den Burg et al.2006, Chanet et al. 2009): whole specimenand head, in and ex vivo; Danio rerio(Kabli et al. 2006): whole specimen, in andex vivo; Gadus morhua (Bock et al. 2002,Lannig et al. 2004): whole specimen, invivo; Hyporhamphus australis (Butcheret al. 2009): whole specimen, in and exvivo; Lampris guttatus (Runcie et al. 2009):brain, ex vivo; Monopterus albus(Rasmussen et al. 2010): proximal part, exvivo; Pachycara brachycephalum (Market al. 2002, Van der Linden et al. 2004):whole specimen, in vivo; Porichthysnotatus (Forbes et al. 2006): wholespecimen, in vivo; Salmo gairdneri (Nottet al. 1999): head, ex vivo; Salmo salar(Veliyulin et al. 2006): thorax, ex vivo;Sebaster sp. (Rogers et al. 2008): wholespecimen, ex vivo; Toxotes jaculatrix(Chanet et al. 2009): whole specimen, exvivo; Zoarces viviparus (Bock et al. 2001b,2002): whole specimen, in vivo
Suitable, but some species will betoo small
Amphibia, amphibians, *6.300 spp., fewmm to few m
Bombina orientalis (this study): wholespecimen, ex vivo, Fig. 8a; Rana sp. (thisstudy): whole specimen, ex vivo; Ranaesculenta (Sbarbati et al. 1992): head, invivo; Rana sylvatica (Rubinsky et al.1994a): whole specimen, in vivo; Xenopuslaevis (Lee et al. 2007): embryo, in vivo
Suitable, but some species will betoo small
Testudines, turtles, *280 spp., few cm tofew m
Caretta caretta (Valente et al. 2006): wholespecimen, in and ex vivo; Chrysemys pictamarginata (Rubinsky et al. 1994b): wholespecimen, in vivo; Geochelone pardalispardalis (Raiti and Haramati 1997): wholespecimen, in vivo; Kinixys sp. (Straub andJurina 2001): whole specimen, in vivo;Testudo graeca, T. hermanni, T. horsfieldii(Straub and Jurina 2001): whole specimen,in vivo; Trachemys scripta (Blackband andStoskopf 1990, Straub and Jurina 2001,Stecyk et al. 2009, Rasmussen et al. 2010,Lauridsen et al. 2011): whole specimen andheart, in and ex vivo; Trachemys scriptaelegans (this study): head, ex vivo, Fig. 8b;unknown species (Kuoni et al. 1993): wholespecimen, in and ex vivo
Suitable
Sphenodontida, tuataras, 2 spp., severalcm
– Presumably suitable
Zoomorphology (2011) 130:227–254 237
123

the coil surrounding the sample are immobile for the
duration of the scan. Movement artifacts are especiallyproblematic for in vivo MRI, where body motion has to be
suppressed by tight fixation and/or the use of anesthesia. It
is, however, not possible to completely avoid motion arti-facts in body regions that display movement due to, for
example, heart beat or breathing. In these cases, special
measures like synchronizing the image acquisition withmovement or the use of additional navigator scans to detect
motion are necessary. Even when scanning samples postmortem, care has to be taken to keep the sample from
moving, for example caused by its flotation in the sur-
rounding fluid or by scanner vibration caused by the gra-dient system. Low-melting agarose can be used to embed
tiny or fragile specimens for scanning in order to avoid this
type of artifacts. In addition, glass or plastic rods can beemployed to immobilize fixed specimens within the sample
container (Ziegler and Mueller 2011).
Another major source of artifacts are local variations of
magnetic susceptibility within the sample, which maycause distortions of the highly homogeneous magnetic field
required for MRI. These distortions may lead to image
deformations or even to dropouts that may extend muchfurther than the actual size of the artifact-causing sub-
stance. The most common reason for this type of artifact is
air within or surrounding the sample, but even strongersusceptibility artifacts are caused by ferro- or paramagnetic
materials. In zoological specimens, these materials may bepresent in the form of buckshot, complex mineralized
structures, or ingested magnetic sediments (Elliott and
Skerritt 2010; Gavin and Bagley 2010). For ex vivo spec-imen scanning, immersion of the entire sample in distilled
water, agarose, or any fluid with susceptibility close to that
of the tissue can help to avoid the susceptibility variationsat the tissue/air interfaces. Air bubbles inside the sample
should, if possible, be removed by degassing (Ziegler and
Table 1 continued
Taxon information Species studied so far using MRI (and otherNMR techniques)
Suitability for MRI
Squamata, scaled reptiles,[6.000 spp.,few cm to several m
Euleptes sp. (this study): frontal part, ex vivo,Fig. 8c
Suitable, some species may be toolong
Crocodilia, crocodilians, 24 spp., few mto several m
Alligator mississippiensis (this study): wholespecimen, ex vivo, Fig. 8d
Suitable, but some species may betoo large
Aves, birds,[10.000 spp., few cm tofew m
Apteryx australis, A. haastii, A. mantelli, A.rowi (Corfield et al. 2008): brain, ex vivo;Ciconia ciconia (Berthold et al. 2001):thorax, in vivo; Serinus canaria(Tindemans et al. 2003): brain, in vivo;Sylvia borin (Czisch et al. 2001):reproductive organs, in vivo; Taeniopygiaguttata (Poirier et al. 2008, this study):head, ex vivo, Fig. 8e
Suitable, but some species may betoo large
Mammalia, mammals, 5.416 spp., few cmto several m
Numerous mammalian model organisms aswell as pets have been scanned using MRI;Daubentonia madagascariensis (Kaufmanet al. 2005): brain, ex vivo; Delphinusdelphis (Marino et al. 2002): brain, ex vivo;Homo sapiens (Ziegler et al. 2011): head, invivo, Fig. 9c; Kogia simus (Marino et al.2003b): brain, ex vivo; Macaca mulatta(Saleem et al. 2002): brain, in vivo; Musmusculus (this study): whole specimen, exvivo, Fig. 9a; Orcinus orca (Marino et al.2004): brain, ex vivo; Oryctolaguscuniculus (Wang et al. 2005): brain, exvivo; Papio papio (Liu et al. 2008): womb,in vivo; Phocoena phocoena (Marino et al.2003a): brain, ex vivo; Platanista gangetica(Endo et al. 1999): torso, ex vivo; Susscrofa (Rasmussen et al. 2010, this study):heart and head, ex and in vivo, Fig. 9b;Tursiops truncatus (Marino et al. 2001,Ridgway et al. 2006): head and brain, inand ex vivo; various primate species(Hopkins et al. 2000): brain, in vivo
Suitable, but some species may betoo large
238 Zoomorphology (2011) 130:227–254
123

Mueller 2011). During in vivo measurements, the presence
of air-filled structures is usually unavoidable and can onlybe treated by adapting the sequence so as to minimize their
negative effects, although some image distortions or
dropouts may remain. Unfortunately, susceptibility arti-facts cannot always be avoided, and several examples are
presented in some of the figures in this article. These
include artifacts caused by air bubbles within (Figs. 2b, 4d,8a) or surrounding the sample (Figs. 2b, 3c, 6g) as well as
ferromagnetic inclusions (Figs. 2e, 4e, f, 5a, c, 6f). Theseinclusions are potentially present in all sediment-ingesting
animals (e.g., Echiura, Sipuncula, some Echinoidea, some
Holothuroidea, some Bivalvia, and some Gastropoda), butalso in taxa that feed on sediment-eaters (e.g., Crustacea,
Vertebrata). Ways to tackle this problem may be to starve
living specimens, or, in the case of museum material, to
systematically look for specimens that display the least orno artifacts. In some instances, a surgical removal of the
artifact-causing structure, for example in the case of the
strongly ferromagnetic radula found in Polyplacophora(Fig. 5a), might be an option.
Darker regions within the image dataset may also be
caused by variations of the fields of the radio frequencycoils. In particular, the use of small surface coils for signal
reception can lead to inhomogeneous image intensity(Fig. 8b, e). In addition, examining relatively large samples
at higher field strengths (i.e., 7 T and above) can lead to
strong inhomogeneities of the transmit field. These inho-mogeneities appear when the wavelength of the MR signal
is smaller than the object size, leading to strong variations
a b c
d e
1 cm
1 cm 1 cm
1 cm
5 mm
cr
ph
me te
bd
mf
bmph
ep
ut
incu
prin
tr
ca
ac
ba
lm
ep
cu
rm
ep
Fig. 2 Results from MRI scans of various metazoan taxa fromCtenophora to Priapulida. a Horizontal section through an entirecomb jelly (Pleurobrachia pileus (O.F. Muller, 1776), Ctenophora).Note the massive shrinkage artifact caused by too rapid rehydration.b Vertical section through an entire sponge (Axinella sp., Porifera).The black spots within and surrounding the specimen are presumablysusceptibility artifacts caused by tiny air bubbles. c Vertical sectionthrough an entire sea anemone (Actinia sp., Cnidaria). Note themovement artifacts (caused by scanner vibration) indicated by theslight blur occurring around the tentacles. d Horizontal sectionthrough the frontal and caudal areas of a large roundworm (Ascaris
sp., Nematoda). e Horizontal section through an entire penis worm(Priapulopsis bicaudatus (Danielssen, 1868), Priapulida). The darkareas are susceptibility artifacts caused by ferromagnetic sedimentingested by the animal. Please refer to Table 2 for an overview ofscanning parameters, while Table 1 provides assessments of thesuitability of metazoan taxa for systematic MRI. ac aboral canal, babase, bd basal disc, bm basilar muscle, ca caudal appendage, cr combrow, cu cuticle, ep epidermis, in intestine, lm longitudinal retractormuscle, me mesohyl, mf mesenterial filament, ph pharynx, prproboscis, rm retractor muscle, te tentacle, tr trunk ring, ut uterus
Zoomorphology (2011) 130:227–254 239
123

in image intensity and contrast (van de Moortele et al.
2005). Specialized hardware and transmit technologies are
necessary to compensate for these effects.Further relevant artifacts are caused by chemical shift
differences, especially between water and fat. Here, fatty
regions are shifted in one dimension relative to the watersignal. Often, this type of artifact can be avoided by
experimentally suppressing the fat signal, usually resulting
in longer scan times. Finally, so-called Gibbs ringingconsists of shifted repetitions of sharp edges in the image.
This is an inherent effect of the spatial encoding technique
and can best be removed by retrospective multiplication ofthe raw data with a filter function. However, while this
effectively removes the artifact, it also somewhat degrades
the spatial resolution of the dataset.
Applying contrast agents in MRI of zoologicalspecimens
For MRI of humans and other animals, a variety of contrastagents (CA) that modulate tissue contrast are available.
Most of these substances comprise molecules with para-
magnetic properties that either increase (T1 agents) ordecrease (T2 agents) the local MRI signal in their vicinity.
T1 agents often contain paramagnetic Gd3? (gadolinium) or
Mn2? (manganese) ions, while T2 agents are usuallycomposed of iron oxide nanoparticles (Shapiro et al. 2004).
Common to all CAs is their property of having a specific
influence on a given tissue, which originates from proper-ties of those molecules that bind the surroundings of
paramagnetic cores. These molecular properties change the
a
5 mm
db
e f1 cm
1 cm
1 cm
1 cm
c
1 cm
lm
op
bl
ss
lm
tr
pr
in
mu ph
lm
fb
in
hclm
cs
he gi
le
hd
ps
pe mu
an
le
an
os
Fig. 3 Results from MRI scans of various panarthropod taxa fromOnychophora to Crustacea. a Vertical section through an entire velvetworm (Ooperipatellus sp., Onychophora). This specimen showsstrong susceptibility artifacts presumably related to an unsuitablefixation protocol. b Horizontal section through an entire spider(Nephila sp., Chelicerata). Note the strong signal originating from thepaired book lungs. c Virtual horizontal section through body and legsof a sea spider (Pycnogonum sp., Pycnogonida). This specimen showsstrong artifacts presumably related to air bubbles attached to thecuticula. d Horizontal section through the anterior segments of a
centipede (Scolopendra sp., Myriapoda). e Horizontal section throughthe anterior half of a cockroach (Periplaneta sp., Hexapoda).f Horizontal section through the carapace of a crab (Pachygrapsusmarmoratus (Fabricius, 1787), Crustacea). Please refer to Table 2 foran overview of scanning parameters, while Table 1 provides assess-ments of the suitability of metazoan taxa for large-scale MRI. anantenna, bl book lung, cs cardiac stomach, fb fat body, gi gills, hchindgut cecum, hd head, he heart, in intestine, le leg, lm leg muscle,mu musculature, op opisthosoma, os ostium, pe pericardium, phpharynx, pr proboscis, ps prosoma, ss sucking stomach, tr trunk
240 Zoomorphology (2011) 130:227–254
123

relative contrast of tissues and make organs distinguishable
in MR images (Weinmann et al. 2003).There are CAs that do not penetrate the blood–brain
barrier (BBB) and may therefore be specifically used to
detect blood leakage in patients with a brain tumor or otherpathologies. Some CAs may remain intravascular and can
thus provide contrast to perform angiography (Fig. 8d),
perfusion measurements, or estimations of vessel densityand size, while other CAs specifically enter neurons via
calcium channels and allow visualizing neuronal tracts.CAs used in molecular MRI either target specific mole-
cules or can be activated by enzymes, a property that can
be exploited to image gene expression (Louie et al. 2000).However, the most common use of CAs is currently their
systemic application in order to achieve visualization of
pathological conditions or a better delineation of morpho-
logical detail.Such systemic CAs may also be applied ex vivo when
studying fixed specimens. Staining protocols have been
published for a number of species and organs such as therat fetus (Petiet et al. 2007), the mouse brain (Kim et al.
2009), or whole zebra fish specimens (Ullmann et al.
2010). However, the specific protocols in general dependon both the CAs used and the sample under investigation
and must therefore be optimized individually. When usingGd-based T1 agents, CA concentrations of 2–10 mM are a
good starting point. Specimens should be incubated in CA-
containing buffer for at least 2 h per millimeter samplethickness (personal observation), and imaging should be
performed in the same solution to avoid dilution and
1 cm
a b c d
e f g h1 cm 1 cm 1 cm
1 cm 1 cm 1 cm 5 mm
os
vsut
vgin
in
epde
loes
go bi
pe
ne
cu
in
vm
in
epin
pr as
ep
lm
cr
ph
pa
bv
brma
sm
gu
tr
ca
Fig. 4 Results from MRI scans of various metazoan taxa fromBryozoa to Clitellata. a Vertical section through a colony of mossanimals (Dendrobeania sp., Bryozoa). The three-dimensional orien-tation of the sample did not permit to obtain a well-aligned virtualsection. b Horizontal section through the proximal part of a flatworm(Fasciola hepatica Linnaeus, 1758, Platyhelminthes). c Sectionthrough an entire ribbon worm (Lineus sp., Nemertea). The knot-like habitus of this specimen did not permit to obtain a symmetricalvirtual section. d Horizontal section through an entire lamp shell(Terebratalia transversa (Sowerby, 1846), Brachiopoda). The twoblack spots inside the animal are susceptibility artifacts originatingfrom trapped air bubbles. e Vertical section through an entire peanutworm (Sipunculus sp., Sipuncula). Particularly, the proximal areas aredisfigured by susceptibility artifacts originating from ingested sedi-ment. f Horizontal section through an entire spoon worm (Thalassema
thalassemum (Pallas, 1774), Echiura). The dark areas within thespecimen are susceptibility artifacts derived from ingested sediment.g Horizontal section through the anterior part of a bristle worm(Nereis sp., Polychaeta). h Horizontal section through the proximalpart of a leech (Hirudo medicinalis Linnaeus, 1758, Clitellata). Pleaserefer to Table 2 for an overview of scanning parameters, whileTable 1 provides assessments of the suitability of metazoan taxa forlarge-scale MRI. as anterior sucker, bi blind intestine, br singlebryozoan, bv blood vessel, ca cecum, cr crop, cu cuticle, de dermis, epepidermis, es esophagus, go gonad, gu gutter, in intestine, lmlongitudinal muscle, lo lophophore, ma mantle, ne nephridium, osoral sucker, pa parapodium, pe pedicle, ph pharynx, pr proboscis, smspindle muscle, tr trunk, ut uterus, vg vitelline gland, vm ventralretractor muscle, vs ventral sucker
Zoomorphology (2011) 130:227–254 241
123

therefore loss of contrast over acquisition time. Table 2
lists those figures that present specimens scanned after
application of a systemic CA.It is important to note that in certain specimens, contrast
between soft tissues is not always increased when CAs are
applied (Ziegler and Mueller 2011). However, contrast willalways increase significantly between soft tissue and cal-
cified or mineralized structures, such as components of an
endo- or exoskeleton. The effect of CAs in soft tissue isthat the relevant time constant T1 may be reduced to about
1% of its original value (Petiet et al. 2007), potentially
resulting in a signal increase of two orders of magnitude.To take full advantage of this gain, appropriate imaging
sequences have to be used since losses from the concom-
itant reduction in the time constant T2 have to be mini-mized. In particular, gradient echo sequences with very
short echo and repetition times (e.g., FLASH) allow full
exploitation of this gain in contrast, while coincidentally
reducing imaging times up to two orders of magnitude.
Shortened scan times can then either be used to increase
throughput in systematic morphological studies (Zhanget al. 2010), or, alternatively, to increase spatial resolution
for individual specimen imaging. The gain in signal-to-
noise ratio (SNR) may consequently allow improvement inisotropic resolution by a factor of two, resulting in a
reduction in voxel size by a factor of eight. Thus, MRI of
fixed specimens can always benefit from the use of T1 CAs.
In vivo MRI of zoological specimens
Apart from its usage on fixed specimens, MRI can also be
used to study living animals. Besides taking live images offunctioning organs, this approach minimizes misinterpre-
tations due to structural changes following fixation and
preservation (Natt and Frahm 2005). A number of
1 cm
a
1 cm 1 cm
1 cm
b
dc
angi
fo
ma
bpst
bm
go
fo
fo
pm amve ey te
cu
ma
dgol
pg
vm
vmpf
ma
in
lp sh
go
Fig. 5 Results from MRI scans of various molluscan taxa fromPolyplacophora to Cephalopoda. a Horizontal section through anentire chiton (Acanthochitona sp., Polyplacophora). The strongsusceptibility artifact in the proximal part is caused by the magnetite-and cobalt-bearing radula. b Vertical section through an entirenudibranch (Dendronotus sp., Gastropoda). c Vertical section throughan entire bivalve (Cerastoderma edule (Linnaeus, 1758), Bivalvia).The slight susceptibility artifacts are caused by ingested para- orferromagnetic particles within the digestive tract. d Horizontal section
through an entire squid (Sepia sp., Cephalopoda). Please refer toTable 2 for an overview of scanning parameters, while Table 1provides assessments of the suitability of metazoan taxa for large-scale MRI. am anterior adductor muscle, an anus, bm buccal mass, bpbranchial plume, cu cuttlebone, dg digestive cecum, ey eye, fo foot, gigills, go gonad, in intestine, lp labial palp, ma mantle, ol optic lobe, pfposterior foot retractor muscle, pg pallial groove, pm posterioradductor muscle, sh systemic heart, st stomach, te tentacle, veventricle, vm visceral mass
242 Zoomorphology (2011) 130:227–254
123

prerequisites are necessary in order to obtain in vivo MRI
data with a similar resolution as would be the case withMRI of preserved samples (Benveniste and Blackband
2002).
In general, a satisfactory resolution for morphologicalMRI can only be achieved by using comparatively long
scanning times. Unfortunately, body movement and natural
organ function in living animals will almost certainly leadto imaging artifacts. Therefore, such samples need to be
mechanically fixed, chemically relaxed, or even fullyanesthetized. In the case of crustaceans or bivalves, for
example, mechanical fixation can easily be achieved by
attaching the carapace or the shell to the experimentalchamber. Such a setup was successfully applied under
defined physiological conditions for several days inside a
seawater flow-through chamber (Fig. 1d) located withinthe MRI scanner (Bock et al. 2001a; Lannig et al. 2008). In
these studies, an in-plane resolution of (200 lm)2–
(500 lm)2 was typically achieved, permitting the differ-entiation of various internal organs such as leg muscles,
hepatopancreas, gonads, gills, and heart in the Europeanspider crab Maja squinado (Herbst, 1788) as well as fast
d gfe
b ca
5 mm 10 cm 1 cm
1 cm 1 cm1 cm1 cm
ri
in
am
pc
cs
ve
bu
mo
bt
inrm st
in
lm
pb
es
go
tr
copr
in
asma
tu
bp
ra
go
dm
os
Fig. 6 Results from MRI scans of various basal deuterostome taxafrom Crinoidea to Tunicata. a Vertical section through the central partof a feather star (Antedon mediterranea (Lamarck, 1816), ZMHE6859, Crinoidea). In order to achieve a higher local resolution, thearms of the specimen were not scanned in their entirety. b Horizontalsection through an entire starfish (Acanthaster planci (Linnaeus,1758), Asteroidea). c Horizontal section through the central part of abrittle star (Ophiocoma nigra O.F. Muller, 1789, ZMH E2025,Ophiuroidea). This specimen had been slightly damaged duringpreparation. d Vertical section through an entire sea cucumber (Aslialefevrei (Barrois, 1882), Holothuroidea). e Horizontal section throughan entire sand dollar (Clypeaster rosaceus (Linnaeus, 1758), ZMBEch 2520, Echinoidea). The sediment inside the gut of this specimenis only slightly ferromagnetic and therefore causes almost no artifacts.f Vertical section through an entire acorn worm (Harrimania kupferi(von Willemoes-Suhm, 1871), ZMB Ent 1579, Enteropneusta). The
internals of this animal are filled with artifact-causing sediment.g Vertical section through an entire tunicate (Ciona intestinalis(Linnaeus, 1767), Tunicata). The tunica of the animal traps tinyartifact-inducing air bubbles. Accession numbers refer to theZoologisches Museum Berlin (ZMB) and the Zoologisches MuseumHamburg (ZMH). Please refer to Table 2 for an overview of scanningparameters, while Table 1 provides assessments of the suitability ofmetazoan taxa for large-scale MRI. Image (b) courtesy of Robert Sigland Hannes Imhof (Munchen, Germany). am ampulla, as atrialsiphon, bp branchial pharynx, bt branchial tentacle, bu bursa, cocollar, cs cardiac stomach, dm dilator muscle, es esophagus, go gonad,in intestine, lm lantern muscle, ma mantle, mo mouth, os oral siphon,pb pillar bridge, pc pyloric cecum, pr proboscis, ra radial canal, riring canal, rm retractor muscle, st stomach, tr trunk, tu tunica, vevertebra
Zoomorphology (2011) 130:227–254 243
123

and catch muscle, pericardium, gills, and heart in the
Eastern oyster Crassostrea virginica (Gmelin, 1791).As an alternative to permanent mechanical fixation,
cold- or freeze-tolerant ectotherms like insects, amphibi-
ans, or reptiles can be immobilized and thus more easilystudied by exposing them to temperatures near or even
below their freezing point, likely in combination with non-
permanent mechanical fixation. Such approaches have beensuccessfully applied to the study of anuran (Rubinsky et al.
1994a), chelonian (Rubinsky et al. 1994b; Stecyk et al.2009), hexapod (Hart et al. 2003), and aranean (Pohlmann
et al. 2007; Lauridsen et al. 2011) species. For example, the
following morphological features could be depicted in thecase of chelonians: carapace, gall bladder, gut, lungs, liver,
plastron, and ventricle (Stecyk et al. 2009).
For in vivo MRI of organisms that possess delicate skinsand where mechanical fixation of the body or any other form
of restraining is not desirable, as for example in the case of
cephalopods, amphibians, and mammals, adequate anes-thetics need to be applied instead. Typical anesthetics for
water breathers are ethanol and MS 222 (tricaine methane-sulfonate). For many invertebrates, MgCl2 (magnesium
1 cm
b
10 cm
1 cm
1 cm
d
c
a phno
mu
gs ey
brto
mu
br
mu
no
li
ey
mu
gi
he
hd
oo
10 cm 1 cm
fe
ey
mo
no
eygi
brst
sb
Fig. 7 Results from MRI scans of deuterostome taxa from Cepha-lochordata to Actinopterygii. a Vertical section through the proximalpart of a lancelet (Branchiostoma lanceolatum (Pallas, 1774),Cephalochordata). b Vertical section through an entire hagfish(Eptatretus stoutii (Lockington, 1878), SIO 87-125, Myxini). c Ver-tical section through the frontal part of a lamprey (Petromyzonmarinus Linnaeus, 1758, SIO 74-134, Petromyzontida). d Horizontalsection through an entire leopard shark (Triakis semifasciata Giard,1855, Chondrichthyes). e Vertical section through the head of acoelacanth (Latimeria chalumnae Smith, 1939, SIO 75-347,
Sarcopterygii). f Horizontal section through the head of a scorpaeniidfish (Agonus cataphractus Linnaeus, 1758, Actinopterygii). Acces-sion numbers refer to the Scripps Institution of Oceanography (SIO),La Jolla. Please refer to Table 2 for an overview of scanningparameters, while Table 1 provides estimates of the suitability ofmetazoan taxa for large-scale MRI. Images (b, c, and e) courtesy ofLawrence R. Frank (San Diego, CA, USA). br brain, ey eye, hd head,he heart, gi gill, gs gill slit, li liver, mo mouth, mu musculature, nonotochord, oo olfactory organ, ph pharynx, sb swim bladder, ststomach, to tooth
244 Zoomorphology (2011) 130:227–254
123
chloride) is an easy-to-handle, reversible muscle relaxant
(Cooper 2011)—concentrations of 2% in salt or freshwaterhave led to good initial results (personal observation). Vol-
atile narcotics like isoflurane and its derivatives are com-
monly applied for anesthesia in small mammals. However,the effects of all these anesthetics are time-dependent and
require accurate dosage as well as extensive physiological
monitoring of the animal under study. Furthermore, in par-ticular for smaller organisms, anesthesia might influence
routine metabolism and therefore may result in potentialmisinterpretations of in vivo conditions and regulatory
processes (Iwama et al. 1989).
In addition to anatomical information, in vivo MRI alsohas great potential to display dynamic information through
the use of angiography and other flow-sensitive MR tech-
niques that can be used to study the function of the heart orthe cardiovascular system both in vertebrates and inverte-
brates. For example, apart from morphological information
obtained on vessel architecture in M. squinado, MR angi-
ography was used to measure the ventilatory performancein this water breather (Bock et al. 2001a).
Functional MRI of zoological specimens
Functional MRI (fMRI) describes a set of techniques usedto visualize neuronal as well as muscular activity in vivo
(Ulmer and Jansen 2010). It is now a common tool in theimaging of human and small animal brain activity (van der
Linden et al. 2007, 2009). fMRI studies in animals may
provide insight into the principles of brain activity, thepathophysiology of brain functions, as well as brain func-
tional reorganization and plasticity. Currently, the most
commonly used methods in fMRI are the blood oxygena-tion level-dependent (BOLD) contrast imaging and the
manganese-enhanced MRI (MEMRI) techniques.
rca
10 cm
1 cm
1 cm
1 cm1 cm
b c
ed
ey
bcli
lu
fb
in
eymu
br
ey
he
br
lu
mu
heao
mu
to
ce
fe
beco
po
mu
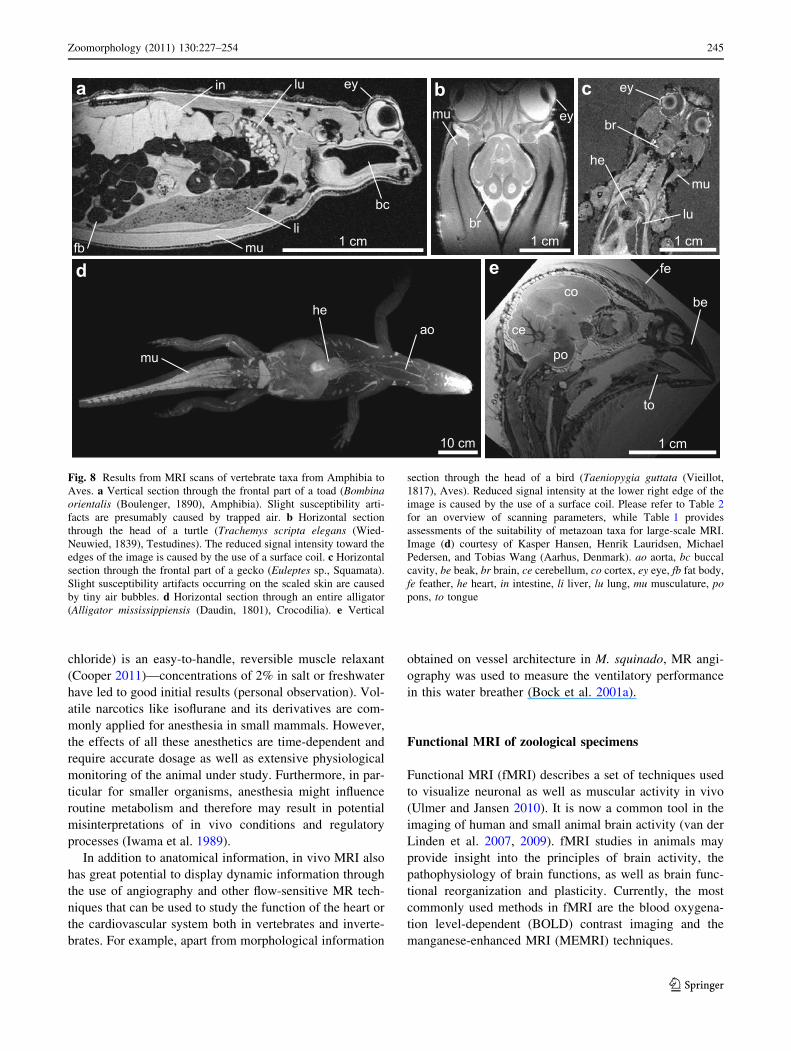
Fig. 8 Results from MRI scans of vertebrate taxa from Amphibia toAves. a Vertical section through the frontal part of a toad (Bombinaorientalis (Boulenger, 1890), Amphibia). Slight susceptibility arti-facts are presumably caused by trapped air. b Horizontal sectionthrough the head of a turtle (Trachemys scripta elegans (Wied-Neuwied, 1839), Testudines). The reduced signal intensity toward theedges of the image is caused by the use of a surface coil. c Horizontalsection through the frontal part of a gecko (Euleptes sp., Squamata).Slight susceptibility artifacts occurring on the scaled skin are causedby tiny air bubbles. d Horizontal section through an entire alligator(Alligator mississippiensis (Daudin, 1801), Crocodilia). e Vertical
section through the head of a bird (Taeniopygia guttata (Vieillot,1817), Aves). Reduced signal intensity at the lower right edge of theimage is caused by the use of a surface coil. Please refer to Table 2for an overview of scanning parameters, while Table 1 providesassessments of the suitability of metazoan taxa for large-scale MRI.Image (d) courtesy of Kasper Hansen, Henrik Lauridsen, MichaelPedersen, and Tobias Wang (Aarhus, Denmark). ao aorta, bc buccalcavity, be beak, br brain, ce cerebellum, co cortex, ey eye, fb fat body,fe feather, he heart, in intestine, li liver, lu lung, mu musculature, popons, to tongue
Zoomorphology (2011) 130:227–254 245
123

BOLD contrast imaging was first described by Ogawaet al. (1990) and reflects hemodynamic changes in response
to neuronal activity. Here, hemoglobin is used as an
endogenous contrast agent by relying on the difference inthe magnetic properties of oxyhemoglobin (diamagnetic)
and deoxyhemoglobin (paramagnetic). BOLD contrast
imaging takes advantage of the fact that paramagneticsubstances act as MR-detectable CAs, which induce a
signal loss in magnetic-susceptibility-sensitive MRI. Dur-
ing neuronal activity, both the metabolic rate as well as thedemand for oxygen rises, resulting in an increase in the
regional blood flow. This leads to the presence of more
oxygen than neurons can dissipate, in turn decreasing therate of deoxyhemoglobin in the activated area, and thereby
causing an increase in the MRI signal intensity. Unfortu-
nately, fMRI using BOLD cannot be applied to metazoansthat use hemocyanin instead of hemoglobin for oxygen
transport (Bock et al. 2007).
For event-related BOLD fMRI, an appropriate stimulus(whether somatosensory, visual, auditory, or olfactory) has
to be provided. In order to avoid motion artifacts as well as
stress for the animal under study, fMRI investigations onzoological specimens have to be performed under anes-
thesia. Here, it is important to choose fMRI-compatible
anesthetics like a-chloralose or medetomidine (at least invertebrates), which do not reduce the coupling between
functional activation and hemodynamic response (van der
Linden et al. 2007). As the changes in the MRI signalinduced by fluctuations in the blood oxygenation level
are relatively small, the stimulus detection phases and
subsequent recoveries have to be measured in an alternateand repetitive manner using ultra-fast T2
*-weighted
sequences. Due to the complexity of the procedure, fMRI
datasets have to be analyzed with dedicated statistical tools(Lazar 2008).
In contrast to the indirect approach using BOLD contrast
imaging, Lin and Koretsky (1997) and Silva et al. (2004)demonstrated that MEMRI enables direct imaging of neu-
ronal activity using T1-weighted MRI. Mn2? ions possess
an ionic radius similar to that of Ca2? (calcium) ions,which act as mediators for the release of neurotransmitters
and neuronal signal transduction in metazoans. Mn2? ions
can therefore enter these excitable cells through voltage-gated calcium channels as well. As Mn2? ions have para-
magnetic properties and shorten the T1 relaxation time of
hydrogen protons, their presence leads to increased signalintensity in T1-weighted MRI. In addition to activation-
induced imaging, MEMRI can also be used to trace specific
neuronal connections such as olfactory, visual, andsomatosensory pathways (Watanabe et al. 2006). Further-
more, manganese can act as a whole-brain CA after sys-
temic administration (Silva et al. 2004; van der Lindenet al. 2007).
One of the advantages of imaging with Mn2? ions is that,
once they accumulate within a cell, their excretion rate isrelatively low, in the order of weeks. This allows admin-
istration of the CA outside the scanner, while the animal is
being stimulated and still awake. Mn2? ions are delivered asa solution of MnCl2 (manganese chloride). Here, the cel-
lular toxicity of Mn2? ions and their compatible dose have
a b c
1 cm 5 cm 10 cm
ey
ve
br
lu
liin
ey
mu
co
lv
nonoey
ea
ea
coce
di
Fig. 9 Results fromMRI scans of various mammalian taxa. aVerticalsection through an entire juvenile mouse (Mus musculus Linnaeus,1758, Rodentia). b Horizontal section through the head of a mini pig(Sus scrofa Linnaeus, 1758, Artiodactyla). c Horizontal sectionthrough the head of a human (Homo sapiens Linnaeus, 1758,
Primates). Please refer to Table 2 for an overview of scanningparameters, while Table 1 provides assessments of the suitability ofmetazoan taxa for large-scale MRI. br brain, ce cerebellum, co cortex,di diaphragm, ea ear, ey eye, in intestine, li liver, lu lung, lv lateralventricle, mu musculature, no nose, ve vertebra
246 Zoomorphology (2011) 130:227–254
123

Tab
le2
MRIscanners
andscanning
parametersforspecim
ensused
inthisstud
yandpresentedin
Figs.2,3,4,5,
6,7,8,9
Figurewithinarticle
MRIscanning
system
Protocolparameters
Vox
elresolution
Con
trast
agent
Scann
ing
medium
Specimen
state
Figures
2b–
e,3b,
c,f,4b,
c,e,
f,h,
5a–d,
6d,
e,g,
7f,
8c,
9a
7TPharm
ascan70
/16AS(Bruker
Biospin
GmbH
,Ettling
en,
Germany)
FLASH,3D
,TR30
ms,TE6.7ms,NA12
,Matrix(384
)3,
FOV
(3.12cm
)3,TA*15
h(81lm
)3–
Distilled
water
Exvivo
Figures
2a,3a,d,
e,4a,d,
g,6a,
c,f,7a
9.4TBrukerAvance(Bruker
Biospin
GmbH
,Ettling
en,
Germany)
RARE,2D
,TR27
12.5
ms,TE11
.6ms,FR4,
NA3,
Matrix(600
)2,FOV
(3cm
)2,TA*
15min
509
509
200lm
Magnevist
Distilled
water
Exvivo
Figure6b
1.5TAchieva
(Philips
Healthcare,
Eindh
oven,The
Netherlands)
TSE,3D,T
R90
0ms,TE81
.2ms,NA10
,Matrix(102
4)2,
FOV
(21cm
)2,TA11
h23
min
(300
lm)3
–70
% Ethanol
Exvivo
Figure7b
7TBrukerBiospec
AvanceII
(BrukerBiospin
GmbH
,Ettling
en,Germany)
FLASH,3D
,TR22
.814
ms,TE11
.322
ms,NA3,
Matrix
1789
7809
466,
FOV
15.979
709
41.82cm
,TA
n/a
(90lm
)3ProHance
ProHance
Exvivo
Figure7c
7TBrukerBiospec
AvanceII
(BrukerBiospin
GmbH
,Ettling
en,Germany)
FLASH,3D
,TR21
.669
ms,TE10
.722
ms,NA3,
Matrix
1679
1024
917
8,FOV
159
709
15.97cm
,TA
n/a
(90lm
)3ProHance
ProHance
Exvivo
Figure7d
3TMagnetom
Trio(Siemens
Healthcare,
Erlangen,
Germany)
FLASH,3D
,TR15
ms,TE5ms,NA20
,Matrix
1712
928
89
256,
FOV
659
9.14
98.12
cm,TA
6h24
min
3809
3189
318lm
–Air
Exvivo
Figure7e
3TSigna
Exite
HDx(G
eneral
Electrics
Healthcare,
Pittsbu
rgh,
PA,USA)
FSPGR,3D
,TR10
.668
ms,TE3.41
6ms,FR4,
NA3,
Matrix51
29
5129
236,
FOV
399
399
18.88cm
,TAn/a
(761
lm)3
ProHance
ProHance
Exvivo
Figure8a
9.4TBrukerAvance(Bruker
Biospin
GmbH
,Ettling
en,
Germany)
SE,2D
,TR63
63ms,TE15
ms,NA64
,Matrix
1024
951
2,FOV
2.59
3.3cm
,TA*
58h
64.4
924
.49
110lm
–Fluorinert
Exvivo
Figure8b
16.4
TBrukerBiospec
(Magnex
Scientific,
Oxford,
UK)
RARE,2D
,TR40
00ms,TE8.2ms,NA1,
Matrix
3129
256,
FOV
3.12
92.56
cm,TA3min
12s
1009
1009
250lm
–Air
Invivo
Figure8d
1.5TMagnetom
Avanto(Siemens
Healthcare,
Erlangen,
Germany)
2D,TR14
ms,TE5.6ms,NA6,
Matrix26
09
320,
FOV
10.6
913
cm,TAn/a
40.6
940
.6lm
–Air
Invivo
Figure8e
9.4TBiospec
(BrukerBiospin
GmbH
,Ettling
en,Germany)
RARE,3D
,TR15
00ms,TE9.97
ms,NA8,
FR16
,Matrix28
89
2889
256,
FOV
2.59
92.59
92.30
cm,TA*
15h
(90lm
)3–
Distilled
water
Exvivo
Figure9b
3TAchieva
(Philips
Healthcare,
Eindh
oven,The
Netherlands)
TSE,2D
,TR25
00ms,TE80
ms,NA6,
Matrix
2909
290,
FOV
1.69
1.6cm
,TA6min
30s
2009
2009
2000
lm–
Air
Invivo
Figure9c
3TAchieva
(Philips
Healthcare,
Eindh
oven,The
Netherlands)
3DT1T
FE,3D
,TR6.9ms,TE3.4ms,NA2,
Matrix
2569
1539
80,FOV
2.56
92.04
91.6cm
,TA*
4min
5009
5009
1000
lm–
Air
Invivo
Zoomorphology (2011) 130:227–254 247
123

to be considered (Silva et al. 2004). For activity-induced
MEMRI in vertebrates, in some cases, an opening of theBBB is required, which can be achieved using a 25%
hypertonic solution of D-mannitol (Silva et al. 2004). If
manganese is administered in small doses, opening the BBBmay not be necessary. However, for neuronal tract tracing,
MnCl2 has to be introduced directly into the nervous tissue
using the olfactory, intravitreal, or retinal pathways (Linand Koretsky 1997), while systemic administration of
MnCl2 can be performed intraperitoneally, subcutaneously,or intramuscularly (Silva et al. 2004). Although the main
application of fMRI is in studies on vertebrate model
organisms and humans, fMRI has previously been per-formed on flies (Jasanoff and Sun 2002).
Magnetic resonance spectroscopy as well as non-protonand diffusion MRI of zoological specimens
In addition to MRI based on hydrogen nuclei, NMR tech-
niques can also be used for magnetic resonance spectros-
copy (MRS) as well as non-proton and diffusion MRI. Inprinciple, these applications allow for the study of physi-
ological function both ex vivo and in vivo.
MRS is used to retrieve information about the molecularenvironment of the element under investigation: due to the
influence of neighboring atoms, hydrogen nuclei in dif-
ferent molecules are subject to a slightly different magneticfield, which can lead to a change in their Larmor frequency,
a phenomenon known as chemical shift. This can be
employed to noninvasively analyze a broad range ofcomplex samples. As MRS in its most simple form
acquires data from within the entire sample, no spatial
information is given. To overcome this deficiency, local-ized spectroscopy methods have been developed and they
can be divided into single- and multi-voxel techniques. In
single-voxel localization, the main benefit is that themagnetic field can be optimized better than compared to
bigger volumes, which in turn further improves spectral
resolution. Various techniques are based on leaving thesignal unperturbed either inside or outside the voxel of
interest (Granot et al. 1986, Ordidge et al. 1986; Bottomley
1987; Frahm et al. 1989; Garwood and De la Barre 2001).However, one disadvantage of single-voxel localized
spectroscopy is its potential to miss important areas of the
sample. In addition, in order to fill a 2D matrix of spectraand to overlay these data with an anatomical image, a long
acquisition time is inevitable. Despite such drawbacks,
MRS techniques have been successfully applied to thestudy of numerous vertebrate and invertebrate species
(Lohmann and Ratcliffe 1988; Hart et al. 2003).
As an extension to MRS, chemical shift imaging (CSI)combines spectroscopic with spatial information, thereby
offering to obtain metabolic profiles from multiple spatial
positions within a sample. In the classical CSI method, onespectrum is acquired at each phase-encoding step
(Maudsley et al. 1983). However, classical CSI is still
relatively time-consuming, as the acquisition time increa-ses with the number of points in the spectral dimension, but
more importantly in the three spatial dimensions. Hence,
there always remains a trade-off between sufficient sensi-tivity due to averaging and short acquisition times (Poh-
mann et al. 1997). Still, versatile CSI applications arepossible ex vivo as well as in vivo, for example pH-metry
(Gallagher et al. 2008), biopsies of cerebral lesions while
avoiding healthy tissue (Hermann et al. 2008), or measur-ing the oxidative capacity among leg muscles in humans
(Forbes et al. 2009). Apart from applications in humans,
CSI has been successfully performed with marine species(Lee et al. 2010). While metabolites of interest may be
abundantly present in living specimens, their concentration
in fixed tissue may differ drastically. Many metabolites willlikely have been replaced by the fixative, whereas, for
example, a comparison between fat and water will still be
feasible.In addition to hydrogen MRS or MRI, other nuclei such
as 3He (helium), 13C (carbon), 17O (oxygen), 19F (fluorine),31P (phosphorus), and 129Xe (xenon), as long as they arepresent in sufficient concentration, can be used to probe the
sample. Such methods are referred to as non-proton or
x-nuclei MRI. Since hydrogen protons have a high naturalabundance in organic substances, there is also a strong
hydrogen background signal from all areas within biolog-
ical samples. This can make it beneficial to scan differentnuclei, as they are usually less abundant than hydrogen and
in turn also have a weaker background signal. In addition,
the polarization of some of these nuclei, in particular 3He,13C, and 129Xe can be drastically increased by hyperpo-
larization (Walker and Happer 1997; Golman et al. 2001;
Ardenkjar-Larsen et al. 2003; Schroder et al. 2006). Thiseffect may lead to an improvement in signal strength of
four orders of magnitude, which can be used to detect very
low concentrations. For instance, hyperpolarized 3He hasbeen successfully used to image void spaces inside verte-
brate lung tissue, something not feasible with hydrogen
MRI.In many biological objects, the movement of molecules
occurs along fibrous structures such as blood capillaries
and neural or muscular tissue. These structures can bemade visible by MR techniques such as diffusion-weighted
imaging (DWI) (Le Bihan et al. 1986) or diffusion tensor
imaging (DTI) (Basser et al. 1994). Diffusion measure-ments by NMR are based on signal loss, as random motion
of particles causes the MR signal to attenuate during the
scan. In particular, DTI techniques have found applicationon typical MRI model organisms such as monkeys
248 Zoomorphology (2011) 130:227–254
123

(Pierpaoli and Basser 1996), zebra finches (De Groof et al.
2006), and mice (Aggarwal et al. 2009), but also on humantissue (Jellison et al. 2004). Interestingly, DTI can in
principle be applied both in vivo as well as ex vivo.
Potential artifacts that may occur during diffusion MRI arediscussed by Le Bihan et al. (2006).
The future of MRI
MRI and NMR research have been a huge success story
over the last 60 years, with fundamental breakthroughs in
the field continuously increasing the range of applicationsin the life sciences. With such strong momentum, it is
expected that future discoveries will bring further
improvements. The major driving force in MRI is certainlythe domain of clinical imaging, with most hardware solu-
tions adapted to the size and specific morphology of
humans. Some promising recent innovations in humanMRI may also be relevant for the study of smaller animals,
whereas others mainly address problems that are not
encountered when studying objects significantly smallerthan humans.
In general, the physical limitations—in particular with
regard to resolution—of MRI strongly depend on the typeof application, as this determines the hardware used and the
protocol chosen. For example, dynamic studies in living
animals require adequate temporal resolution and thus limitthe time to acquire detailed spatial information. More
importantly, the smaller the specimen, the smaller the
signal-generating volume elements as higher resolutionswill be necessary. This potential conflict has recently been
termed the ‘‘Bermuda triangle of small animal MRI’’ (Ja-
kob 2011), and the understandable desire of zoomorphol-ogists for maximum achievable resolution is not easy to
meet. Glover and Mansfield (2002) quote the resolution
limit of MRI due to molecular diffusion in the case ofhydrogen protons to be approximately 8 lm. Movement or
flow artifacts in living organisms might cause even more
severe restrictions on resolution. Some of these problemscan be tackled by proper preparation and fixation of the
sample. However, manipulating the molecular dynamics of
a sample may also influence relaxation behavior of thetissue under study. Hence, physical limitations on the
microscopic level like, for example, molecular diffusion
and T2 relaxation can usually not be corrected, in particularwhen studying biological samples that cannot be manipu-
lated internally.
On the other hand, approaches involving hardwareimprovements, for example the use of phased array coils
(Blamire 2008) or cryo probes (Baltes et al. 2009), can
increase the available SNR by a factor of 2–3. Furtherprogress can be seen in the development of sequences with
ultra-short echo times. Such improved acquisition schemes
increase the available signal or even make tissues that wereinvisible with conventional techniques now detectable, for
example tendons and cortical bone (Blamire 2008).
Although these approaches do not bypass the fundamentallimit in resolution, they at least allow approaching it and
may enable a considerable reduction in image acquisition
time.In summary, the micrometer scale represents the current
physical limit for conventional MRI applications. Near-future developments will predominantly reduce acquisition
times rather than yield major benefits in spatial resolution.
The more distant future will show how much of the newideas from other specialized fields can be imported into life
sciences to further improve MRI and its multiple applica-
tions in zoology.
Conclusions
We are convinced that the morphological study of zoo-
logical specimens can considerably profit from an extendeduse of MRI. The potential of this technique for zoology lies
particularly in the capacity of MRI to reveal gross mor-
phology in situ, both in vivo and ex vivo. Furthermore,systematic scanning efforts will provide a boost to zoology
by triggering new research, especially so when MRI is used
in combination with advanced 3D modeling and visuali-zation techniques as well as dataset deposition in voxel
repositories (Giribet 2010; Ziegler et al. 2010b; Rowe and
Frank 2011). The density of adequate MRI scanners iscurrently high enough to provide the logistic framework for
such studies, although basic zoological research using MRI
systems can be impeded by the sometimes prohibitive costof scanner access. Nevertheless, previous comprehensive
studies on vertebrates as well as invertebrates have dem-
onstrated that there may be ways to tackle this problem.Numerous metazoan clades have been shown to be suitable
for systematic MRI analysis and we hope that the multitude
of further NMR applications will inspire the members ofthe zoological community to increase the use of these
exciting technologies in their field.
Acknowledgments We would like to thank Thomas Bartolomaeus(Bonn, Germany), Sven Gemballa (Tubingen, Germany), MatthiasGlaubrecht (Berlin, Germany), Alexander Gruhl (London, UnitedKingdom), Alexander Haas (Hamburg, Germany), Markus Koch(Bonn, Germany), Janina Lehrke (Bonn, Germany), Carsten Luter(Berlin, Germany), Christian Muller (Frankfurt, Germany), ThomasStach (Berlin, Germany), and Esther Ullrich-Luter (Berlin, Germany)for specimen supply. We also gratefully acknowledge editorial sup-port by Christopher Witte (Berlin, Germany). Bivalve research wassupported by AToL grants from the National Science Foundation,USA, to Rudiger Bieler (#0732854), Paula M. Mikkelsen (#0732860),and Gonzalo Giribet (#0732903). We are grateful to Thomas
Zoomorphology (2011) 130:227–254 249
123

Bartolomaeus, Doug Eernisse and one anonymous reviewer for theirsupportive comments on this manuscript.
References
Aggarwal M, Zhang J, Miller MI, Sidman RL, Mori S (2009)Magnetic resonance imaging and micro-computed tomographycombined atlas of developing and adult mouse brains forstereotaxic surgery. Neuroscience 162:1339–1350
Ardenkjar-Larsen JH, Fridlund B, Gram A, Hansson G, Hansson L,Lerche MH (2003) Increase in signal-to-noise ratio of[10, 000times in liquid-state NMR. Proc Natl Acad Sci USA100:10158–10163
Assheuer J, Sager M (1997) MRI and CT atlas of the dog. BlackwellScience, Berlin
Baker M (2010) The whole picture. Nature 463:977–980Baltes C, Radzwill N, Bosshard S, Marek D, Rudin M (2009) Micro
MRI of the mouse brain using a novel 400 MHz cryogenicquadrature RF probe. NMR Biomed 22:834–842
Basser PJ, Mattiello J, Le Bihan D (1994) Estimation of the effectiveself-diffusion tensor from the NMR spin echo. J Magn Reson103:247–254
Belton PS, Gil AM, Webb GA, Rutledge D (2003) Magneticresonance in food science: latest developments. RSC Publishing,Cambridge
Belton PS, Engelsen SB, Jakobsen HJ, Amin MHG, van den Berg F(2005) Magnetic resonance in food science: the multivariatechallenge. RSC Publishing, Cambridge
Benveniste H, Blackband SJ (2002) MR microscopy and highresolution small animal MRI: applications in neuroscienceresearch. Prog Neurobiol 67:393–420
Benveniste H, Blackband SJ (2006) Translational neuroscience andmagnetic-resonance microscopy. Lancet Neurol 5:536–544
Berthold P, Elverfeldt D, Fiedler W, Hennig J, Kaatz M, Querner U(2001) Magnetic resonance imaging and spectroscopy (MRI,MRS) of seasonal patterns in body composition: a methodolog-ical pilot study in white storks (Ciconia ciconia). J Ornithol142:63–72
Blackband SJ, Stoskopf MK (1990) In vivo nuclear magneticresonance imaging and spectroscopy of aquatic organisms.Magn Reson Imag 8:191–198
Blamire AM (2008) The technology of MRI–the next 10 years? Br JRadiol 81:601–617
Bock C, Frederich M, Wittig RM, Portner HO (2001a) Simultaneousobservations of haemolymph flow and ventilation in marinespider crabs at different temperatures: a flow weighted MRIstudy. Magn Reson Imag 19:1113–1124
Bock C, Sartoris FJ, Wittig RM, Portner HO (2001b) Temperaturedependent pH regulation in stenothermal Antarctic and eury-thermal temperate eelpout (Zoarcidae): an in vivo NMR study.Polar Biol 24:869–874
Bock C, Sartoris FJ, Portner HO (2002) In vivo MR spectroscopy andMR imaging on non-anaesthetized marine fish: techniques andfirst results. Magn Reson Imag 20:165–172
Bock C, Lurmann G, Portner HO (2007) Temperature and oxygen-ation dependence of haemoglobin and hemocyanin relaxationtimes at 9.4 T. Proc Int Soc Magn Reson Med 15:2749
Boistel R, Swoger J, Krzic U, Fernandez V, Gillet B, Reynaud EG(2011) The future of three-dimensional microscopic imagingin marine biology. Marine Ecol. doi:10.1111/j.1439-0485.2011.00442.x
Bottomley PA (1987) Spatial localization in NMR spectroscopy invivo. Ann NY Acad Sci 508:333–348
Bringmann G, Wolf K, Lanz T, Haase A, Hiort J, Proksch P, MullerWEG (1999) Direct demonstration of spatial water distributionin the sponge Suberites domunucla by in vivo NMR imaging.Mar Ecol Prog Ser 189:307–310
Brinkley CK, Kolodny NH, Kohler SJ, Sandeman DC, Beltz BS(2005) Magnetic resonance imaging at 9.4 T as a tool forstudying neural anatomy in non-vertebrates. J Neurosci Methods146:124–132
Brouwer M, Engel DW, Bonaventura J, Johnson GA (1992) In vivomagnetic resonance imaging of the blue crab, Callinectessapidus: effect of cadmium accumulation in tissues on protonrelaxation properties. J Exp Zool 263:32–40
Budd GE, Olsson L (2007) Editorial: a renaissance for evolutionarymorphology. Acta Zool (Stockholm) 88:1
Butcher PA, Broadhurst MK, Hall KC, Cullis BR, Nicoll RG (2009)Scale loss and mortality in angled-and-released Eastern seagarfish (Hyporhamphus australis). ICES J Marine Sci 67:522–529
Chanet B, Fusellier M, Baudet J, Madec S, Guintard C (2009) Noneed to open the jar: a comparative study of magnetic resonanceimaging results on fresh and alcohol preserved common carps(Cyprinus carpio (L. 1758), Cyprinidae, Teleostei). ComptesRendus de Biol 332:413–419
Chudek JA, Crook AME, Hubbard SF, Hunter G (1996) Nuclearmagnetic resonance microscopy of the development of theparasitoid wasp Venturia canescens within its host moth Plodiainterpunctella. Magn Reson Imag 14:679–686
Conner WE, Johnson GA, Coffer GP, Dittrich K (1988) Magneticresonance microscopy—in vivo sectioning of a developinginsect. Experientia 44:11–12
Cooper JE (2011) Anesthesia, analgesia, and euthanasia of inverte-brates. ILAR J 52:196–204
Corfield JR, Wild JM, Cowan BR, Parsons S, Kubke MF (2008) MRIof postmortem specimens of endangered species for comparativebrain anatomy. Nat Protoc 3:597–605
Czisch M, Coppack T, Berthold P, Auer DP (2001) In vivo magneticresonance imaging of the reproductive organs in a passerine birdspecies. J Avian Biol 32:278–281
Davenel A, Quellec S, Pouvreau S (2006) Noninvasive characteriza-tion of gonad maturation and determination of the sex of Pacificoysters by MRI. Magn Reson Imag 24:1103–1110
Davenel A, Gonzalez R, Suquet M, Quellec S, Robert R (2010)Individual monitoring of gonad development in the European flatoyster Ostrea edulis by in vivo magnetic resonance imaging.Aquaculture 307:165–169
De Groof G, Verhoye M, van Meir V, Tindemans I, Leemans A, vander Linden A (2006) In vivo diffusion tensor imaging (DTI) ofbrain subdivisions and vocal pathways in songbirds. Neuroimage29:754–763
Dunn CW, Hejnol A, Matus DQ, Pang K, Browne WE, Smith SA,Seaver E, Rouse GW, Obst M, Edgecombe GD, Sorensen MV,Haddock SHD, Schmidt-Rhaesa A, Okusu A, Kristensen RM,Wheeler WC, Martindale MQ, Giribet G (2008) Broad phylog-enomic sampling improves resolution of the animal tree of life.Nature 452:745–749
Edgecombe GD, Giribet G, Dunn CW, Hejnol A, Kristensen RM,Neves RC, Rouse GW, Worsaae K, Sorensen MV (2011)Higher-level metazoan relationships: recent progress andremaining questions. Org Divers Evol 11:151–172
Elliott I, Skerritt G (2010) Handbook of small animal MRI. Wiley-Blackwell, Oxford
Endo H, Yamagiwa D, Arishima K, Yamamoto M, Sasaki M, HayashiY, Kamiya T (1999) MRI examination of trachea and bronchi inthe Ganges river dolphin (Platanista gangetica). J Vet Med Sci61:1137–1141
250 Zoomorphology (2011) 130:227–254
123

Farhat IA, Belton PS, Webb GA (2007) Magnetic resonance in foodscience: from molecules to man. RSC Publishing, Cambridge
Favila ME, Fresneau D, Gonord P, Ruaud JP (2004) Nuclearmagnetic resonance microscopy of the internal structure of thecarrion rolling scarab Canthon cyanellus cyanellus (Scarabaei-dae: Scarabaeinae). Coleopt Bull 58:1125–1131
Fernandez M, Bock C, Portner HO (2000) The cost of being a caringmother: the ignored factor in the reproduction of marineinvertebrates. Ecol Lett 3:487–494
Forbes JG, Morris HD, Wang K (2006) Multimodal imaging of thesonic organ of Porichthys notatus, the singing midshipman fish.Magn Reson Imag 24:321–331
Forbes SC, Slade JM, Francis RM, Meyer RA (2009) Comparison ofoxidative capacity among leg muscles in humans using gated 31P2-D chemical shift imaging. NMR Biomed 22:1063–1071
Frahm J, Bruhn H, Gyngell ML, Merboldt KD, Hanicke W, Sauter R(1989) Localized high-resolution proton NMR spectroscopyusing stimulated echoes: initial applications to human brain invivo. Magn Reson Med 9:79–93
Fresneau D, Dantas de Araujo C, Kan SK, Gonord P, Jallon JM(1991) Premieres tentatives d’imagerie RMN sur des insectes.Actes des Colloques Insectes Sociaux 7:195–199
Gallagher FA, Kettunen MI, Day SE, Hu DE, Ardenkjaer-Larsen JH,Zandt R, Jensen PR, Karlsson M, Golman K, Lerche MH,Brindle KM (2008) Magnetic resonance imaging of pH in vivousing hyperpolarized 13C-labelled bicarbonate. Nature 453:940–943
Garwood M, De la Barre L (2001) The return of the frequency sweep:designing adiabatic pulses for contemporary NMR. J MagnReson 153:155–177
Gassner G, Lohmann JAB (1987) Combined proton NMR imagingand spectral analysis of locust embryonic development. ProcNatl Acad Sci USA 84:5297–5300
Gavin PR, Bagley RS (2010) Practical small animal MRI. Wiley-Blackwell, Oxford
Geoghegan IE, Chudek JA, Mackay RL, Lowe C, Moritz S, McNicolRJ, Birch ANE, Hunter G, Majerus MEN (2000) Study ofanatomical changes in Coccinella septempunctata (Coleoptera:Coccinellidae) induced by diet and by infection with the larva ofDinocampus coccinellae (Hymenoptera: Braconidae) usingmagnetic resonance microimaging. Eur J Entomol 97:457–461
Giribet G (2010) A new dimension in combining data? The use ofmorphology and phylogenomic data in metazoan systematics.Acta Zool (Stockholm) 91:11–19
Glover P, Mansfield P (2002) Limits to magnetic resonance micros-copy. Rep Prog Phys 65:1489–1511
Golman K, Axelsson O, Johannesson H, Mansson S, Olofsson C,Petersson JS (2001) Parahydrogen-induced polarization inimaging: subsecond 13C angiography. Magn Reson Med 46:1–5
Goodman BA, Gordon SC, Chudek JA, Hunter G, Woodford JAT(1995) Nuclear magnetic resonance microscopy as a non-invasive tool to study the development of lepidopteran pupae.J Insect Physiol 41:419–424
Gozansky EK, Ezell EL, Budelmann BU, Quast MJ (2003) Magneticresonance histology: in situ single cell imaging of receptor cellsin an invertebrate (Lolliguncula brevis, Cephalopoda) senseorgan. Magn Reson Imag 21:1019–1022
Granot J (1986) Selected volume excitation using stimulated echoes(VEST): application to spatially localized spectroscopy andimaging. J Magn Reson 70:488–492
Grant SC, Aiken NR, Plant HD, Gibbs S, Mareci TH, Webb AG,Blackband SJ (2000) NMR spectroscopy of single neurons.Magn Reson Med 44:19–22
Gujonsdottir M, Belton PS, Webb GA (2009) Magnetic resonance infood science: challenges in a challenging world. RSC Publishing,Cambridge
Haddad D, Schaupp F, Brandt R, Manz G, Menzel R, Haase A (2004)NMR imaging of the honeybee brain. J Insect Sci 4:7
Hallock KJ (2008) Magnetic resonance microscopy of flows andcompressions of the circulatory, respiratory, and digestivesystems in pupae of the tobacco hornworm, Manduca sexta.J Insect Sci 8:10
Hart AG, Bowtell RW, Kockenberger W, Wenseleers T, RatnieksFLW (2003) Magnetic resonance imaging in entomology: acritical review. J Insect Sci 3:5
Herberholz J, Mims CJ, Zhang X, Hu X, Edwards DH (2004)Anatomy of a live invertebrate revealed by manganese-enhancedmagnetic resonance imaging. J Exp Biol 207:4543–4550
Hermann EJ, Hattingen E, Krauss JK, Marquardt G, Pilatus U, FranzK, Setzer M, Gasser T, Tews DS, Zanella FE, Seifert V,Lanfermann H (2008) Stereotactic biopsy in gliomas guided by3-Tesla 1H-chemical-shift imaging of choline. Stereotact FunctNeurosurg 86:300–307
Holliman FM, Davis D, Bogan AE, Kwak TJ, Cope WG, Levine JF(2008) Magnetic resonance imaging of live freshwater mussels(Unionidae). Invertebr Biol 127:396–402
Hopkins WD, Pilcher DW, MacGregor L (2000) Sylvian fissureasymmetries in nonhuman primates revisited: a comparativeMRI study. Brain Behav Evol 56:293–299
Hornschemeyer T, Goebbels J, Weidemann G, Faber C, Haase A(2006) The head morphology of Ascioplaga mimeta (Coleoptera:Archostemata) and the phylogeny of Archostemata. Eur JEntomol 103:409–423
Hsu EW, Aiken NR, Blackband SJ (1996) Nuclear magneticresonance microscopy of single neurons under hypotonicperturbation. Am J Physiol 271:C1895–C1900
Iwama GK, McGeer JC, Pawluk MP (1989) The effects of five fishanaesthetics on acid-base balance, hematocrit, blood gases,cortisol, and adrenaline in rainbow trout. Can J Zool 67:2065–2073
Jakob P (2011) Small animal magnetic resonance imaging: basicprinciples, instrumentation and practical issue. In: Kiessling F,Pichler BJ, Hauff P (eds) Small animal imaging. Basics andpractical guide. Springer, Berlin, pp 151–164
Jasanoff A (2005) Functional MRI using molecular imaging agents.Trends Neurosci 28:120–126
Jasanoff A, Sun PZ (2002) In vivo magnetic resonance microscopy ofbrain structure in unanesthetized flies. J Magn Reson 158:79–85
Jellison BJ, Field AS, Medow J, Lazar M, Salamat MS, Alexander AL(2004) Diffusion tensor imaging of cerebral white matter: apictorial review of physics, fiber tract anatomy, and tumorimaging patterns. Am J Neuroradiol 25:356–369
Kabli S, Alia A, Spaink HP, Verbeek FJ, De Groot HJM (2006)Magnetic resonance microscopy of the adult zebrafish. Zebrafish3:431–439
Kaufman JA, Ahrens ET, Laidlaw DH, Zhang S, Allman JM (2005)Anatomical analysis of an Aye–Aye brain (Daubentonia mad-agascariensis, Primates: Prosimii) combining histology, struc-tural magnetic resonance imaging, and diffusion-tensor imaging.Anat Rec A 287:1026–1037
Kiessling F, Pichler BJ, Hauff P (2011) Small animal imaging. Basicsand practical guide. Springer, Berlin
Kim S, Pickup S, Hsu O, Poptani H (2009) Enhanced delineation ofwhite matter structures of the fixed mouse brain using Gd-DTPAin microscopic MRI. NMR Biomed 22:303–309
Kuoni W, Augustiny N, Rubel A (1993) Application of magneticresonance imaging in reptile medicine. MAGMA 1:61–63
Lannig G, Bock C, Sartoris FJ, Portner HO (2004) Oxygen limitationof thermal tolerance in cod, Gadus morhua L., studied bymagnetic resonance imaging and on-line venous oxygen mon-itoring. Am J Physiol Regul Integr Comp Physiol 287:R902–R910
Zoomorphology (2011) 130:227–254 251
123

Lannig G, Cherkasov AS, Portner HO, Bock C, Sokolova IM (2008)Cadmium-dependent oxygen limitation affects temperature tol-erance in eastern oysters (Crassostrea virginica Gmelin). Am JPhysiol Regul Integr Comp Physiol 294:1338–1346
Lauridsen H, Hansen K, Wang T, Agger P, Andersen JL, Knudsen PS,Rasmusseun AS, Uhrenholt L, Pedersen M (2011) Inside out:modern imaging techniques to reveal animal anatomy. PLoS One6:e17879
Lazar N (2008) The statistical analysis of functional MRI data.Springer, Berlin
Le Bihan D, Breton E, Lallemand D, Grenier P, Cabanis E, Laval-Jeantet M (1986) MR imaging of intravoxel incoherent motions:application to diffusion and perfusion in neurologic disorders.Radiology 161:401–407
Le Bihan D, Poupon C, Amadon A, Lethimonnier F (2006) Artifactsand pitfalls in diffusion MRI. J Magn Reson Imag 24:478–488
Lee SC, Mietchen D, Cho JH, Kim YS, Kim C, Hong KS, Lee C,Kang D, Lee W, Cheong C (2007) In vivo magnetic resonancemicroscopy of differentiation in Xenopus laevis embryos fromthe first cleavage onwards. Differentiation 75:84–92
Lee H, Tikunov A, Stoskopf MK, Macdonald JM (2010) Applicationsof chemical shift imaging to marine sciences. Marine Drugs8:2369–2383
Lin YJ, Koretsky AP (1997) Manganese ion enhances T1-weightedMRI during brain activation: an approach to direct imaging ofbrain function. Magn Reson Med 38:378–388
Liu F, Garland M, Duan Y, Stark RI, Xu D, Dong Z, Bansal R,Peterson BS, Kangarlu A (2008) Study of the development offetal baboon brain using magnetic resonance imaging at 3 Tesla.Neuroimage 40:148–159
Lohmann JAB, Gassner G (1987) Proton NMR in vivo imaging: atool for studying embryo development of small organisms. AnnNY Acad Sci 508:435–436
Lohmann JAB, Ratcliffe RG (1988) Prospects for NMR imagingin the study of biological morphogenesis. Experientia 44:666–672
Louie AY, Huber MM, Ahrens ET, Rothbacher U, Moats R, JacobsRE, Fraser SE, Meade TJ (2000) In vivo visualization of geneexpression using magnetic resonance imaging. Nat Biotechnol18:321–325
Mapelli M, Greco F, Gussoni M, Consonni R, Zetta L (1997)Application of NMR microscopy to the morphological study ofthe silkworm, Bombyx mori, during its metamorphosis. MagnReson Imag 15:693–700
Marino L, Sudheimer KD, Murphy TL, Davis KK, Pabst DA,McLellan WA, Rilling JK, Johnson JI (2001) Anatomy andthree-dimensional reconstructions of the brain of a bottlenosedolphin (Tursiops truncatus) from magnetic resonance images.Anat Rec 264:397–414
Marino L, Sudheimer KD, Pabst DA, McLellan WA, Filsoof D,Johnson JI (2002) Neuroanatomy of the common dolphin(Delphinus delphis) as revealed by magnetic resonance imaging(MRI). Anat Rec 268:411–429
Marino L, Sudheimer K, Sarko D, Sirpenski G, Johnson JI (2003a)Neuroanatomy of the harbor porpoise (Phocoena phocoena)from magnetic resonance images. J Morphol 257:308–347
Marino L, Sudheimer K, Pabst DA, McLellan WA, Johnson JI(2003b) Magnetic resonance images of the brain of a dwarfsperm whale (Kogia simus). J Anat 203:57–76
Marino L, Sherwood CC, Delman BN, Tang CY, Naidich TP, Hof PR(2004) Neuroanatomy of the killer whale (Orcinus orca) frommagnetic resonance images. Anat Rec A 281:1256–1263
Mark FC, Bock C, Portner HO (2002) Oxygen-limited thermaltolerance in Antarctic fish investigated by MRI and 31P-MRS.Am J Physiol Regul Integr Comp Physiol 283:R1254–R1262
Maudsley AA, Hilal SK, Perman WH, Simon HE (1983) Spatiallyresolved high resolution spectroscopy by ‘‘four-dimensional’’NMR. J Magn Reson 51:147–152
Metscher BD (2009) MicroCT for comparative morphology: simplestaining methods allow high-contrast 3D imaging of diverse non-mineralized animal tissues. BMC Physiol 9:11
Michaelis T, Watanabe T, Natt O, Boretius S, Frahm J, Utz S,Schachtner J (2005) In vivo 3D MRI of insect brain: cerebraldevelopment during metamorphosis of Manduca sexta. Neuro-image 24:596–602
Mietchen D, Keupp H, Manz B, Volke F (2005) Non-invasivediagnostics in fossils—magnetic resonance imaging of patho-logical belemnites. Biogeosciences 2:133–140
Mietchen D, Aberhan M, Manz B, Hampe O, Mohr B, Neumann C,Volke F (2008a) Three-dimensional magnetic resonance imagingof fossils across taxa. Biogeosciences 5:25–41
Mietchen D, Manz B, Volke F, Storey K (2008b) In vivo assessmentof cold adaptation in insect larvae by magnetic resonanceimaging and magnetic resonance spectroscopy. PLoS One12:e3826
Modo M, Bulte JWH (2011) Magnetic resonance neuroimaging.Methods and protocols. Springer, Berlin
Mooney TA, Hanlon RT, Christensen-Dalsgaard J, Madsen PT,Ketten DR, Nachtigall PE (2010) Sound detection by the longfinsquid (Loligo pealeii) studied with auditory evoked potentials:sensitivity to low-frequency particle motion and not pressure.J Exp Biol 213:3748–3759
Muller WEG, Kaluzhnaya OV, Belikov SI, Rothenberger M, Schro-der HC, Reiber A, Kaandorp JA, Manz B, Mietchen D, Volke F(2006) Magnetic resonance imaging of the siliceous skeleton ofthe demosponge Lubomirskia baicalensis. J Struct Biol153:31–41
Murray RC (2011) Equine MRI. Blackwell-Wiley, OxfordNatt O, Frahm J (2005) In vivo magnetic resonance imaging: insights
into structure and function of the central nervous system. MeasSci Technol 16:R17–R36
Neustadter DM, Drushel RF, Crago PE, Adams BW, Chiel HJ (2002)A kinematic model of swallowing in Aplysia californica basedon radula/odontophore kinematics and in vivo magnetic reso-nance images. J Exp Biol 205:3177–3206
Nott KP, Evans SD, Hall LD (1999) Quantitative magnetic resonanceimaging of fresh and frozen-thawed trout. Magn Reson Imag17:445–455
Novakovic VA, Sutton GP, Neustadter DM, Beer RD, Chiel HJ(2006) Mechanical reconfiguration mediates swallowing andrejection in Aplysia californica. J Comp Physiol A 192:857–870
Null B, Liu CW, Hedehus M, Conolly S, Davis RW (2008) High-resolution, in vivo magnetic resonance imaging of Drosophila at18.8 Tesla. PLoS One 3:e2817
Ogawa S, Lee TM, Kay AR, Tank DW (1990) Brain magneticresonance imaging with contrast dependent on blood oxygena-tion. Proc Natl Acad Sci USA 87:9868–9872
Ordidge RJ, Connelly A, Lohman JAB (1986) Image-selected in vivospectroscopy (ISIS)—a new technique for spatially selectiveNMR spectroscopy. J Magn Reson 66:283–294
Perry CN, Cartamil DP, Bernal D, Sepulveda CA, Theilmann RJ,Graham JB, Frank LR (2007) Quantification of red myotomalmuscle volume and geometry in the shorfin mako shark (Isurusoxyrinchus) and the salmon shark (Lamna ditropis) using T1-weighted magnetic resonance imaging. J Morphol 268:284–292
Petiet A, Hedlund L, Johnson GA (2007) Staining methods formagnetic resonance microscopy of the rat fetus. J Magn ResonImag 25:1192–1198
Pierpaoli C, Basser PJ (1996) Toward a quantitative assessment ofdiffusion anisotropy. Magn Reson Med 36:893–906
252 Zoomorphology (2011) 130:227–254
123

Pohlmann A, Moller M, Decker H, Schreiber WG (2007) MRI oftarantulas: morphological and perfusion imaging. Magn ResonImag 25:129–135
Pohmann R, von Kienlin M, Haase A (1997) Theoretical evaluationand comparison of fast chemical shift imaging methods. J MagnReson 129:145–160
Poirier C, Vellema M, Verhoye M, van Meir V, Wild JM, BalthazartJ, van der Linden A (2008) A three-dimensional MRI atlas ofthe zebra finch brain in stereotaxic coordinates. Neuroimage41:1–6
Pouvreau S, Rambeau M, Cochard JC, Robert R (2006) Investigationof marine bivalve morphology by in vivo MR imaging: firstanatomical results of a promising technique. Aquaculture259:415–423
Price WS, Kobayashi A, Ide H, Natori S, Arata Y (1999) Visualizingthe postembryonic development of Sarcophaga peregrina (fleshfly) by NMR microscopy. Physiol Entomol 24:386–390
Quast MJ, Neumeister H, Ezell EL, Budelmann BU (2001) MRmicroscopy of cobalt-labeled nerve cells and pathways in aninvertebrate brain (Sepia officinalis, Cephalopoda). Magn ResonMed 45:575–579
Raiti P, Haramati N (1997) Magnetic resonance imaging andcomputerized tomography of a gravid leopard tortoise (Geoche-lone pardalis pardalis) with metabolic bone disease. J Zoo WildlMed 28:189–197
Rasmussen AS, Lauridsen H, Laustsen C, Jensen BG, Pedersen SF,Uhrenholt L, Boel LWT, Uldbjerg N, Wang T, Pedersen M(2010) High-resolution ex vivo magnetic resonance angiogra-phy: a feasibility study on biological and medical tissues. BMCPhysiol 10:3
Renou JP, Belton PS, Webb GA (2011) Magnetic resonance in foodscience: an exciting future. RSC Publishing, Cambridge
Ridgway S, Houser D, Finneran J, Carder D, Keogh M, van Bonn W,Smith C, Scadeng M, Dubowitz D, Mattrey R, Hoh C (2006)Functional imaging of dolphin brain metabolism and blood flow.J Exp Biol 209:2902–2910
Rogers BL, Lowe CG, Fernandez-Juricic E, Frank LR (2008)Utilizing magnetic resonance imaging (MRI) to assess theeffects of angling-induced barotrauma on rockfish (Sebastes).Can J Fish Aquac Sci 65:1245–1249
Rowe T, Frank LR (2011) The disappearing third dimension. Science331:712–714
Rubinsky B, Wong STS, Hong JS, Gilbert J, Roos M, Storey KB(1994a) 1H magnetic resonance imaging of freezing and thawingin freeze-tolerant frogs. Am J Physiol 266:R1771–R1777
Rubinsky B, Hong JS, Storey KB (1994b) Freeze tolerance in turtles:visual analysis by microscopy and magnetic resonance imaging.Am J Physiol 267:R1078–R1088
Runcie RM, Dewar H, Hawn DR, Frank LR, Dickson KA (2009)Evidence for cranial endothermy in the opah (Lampris guttatus).J Exp Biol 212:461–470
Saleem KS, Pauls JM, Augath M, Trinath T, Prause BA, HashikawaT, Logothetis NK (2002) Magnetic resonance imaging ofneuronal connections in the macaque monkey. Neuron 34:685–700
Sbarbati A, Leclerq F, Antonakis K, Osculati F (1992) Magneticresonance imaging of the saccular otolithic mass. J Anat181:369–732
Schmidt-Rhaesa A (2009) Morphology and deep metazoan phylog-eny. Zoomorphology 128:199–200
Schroder L, Faber C (2011) In vivo NMR imaging. Methods andprotocols. Methods in Molecular Biology 771. Humana Press,New York
Schroder L, Lowery TJ, Hilty C, Wemmer DE, Pines A (2006)Molecular imaging using a targeted magnetic resonance hyper-polarized biosensor. Science 314:446–449
Shapiro EM, Skrtic S, Sharer K, Hill JM, Dunbar CE, Koretsky AP(2004) MRI detection of single particles for cellular imaging.Proc Natl Acad Sci USA 30:10901–10906
Silva AC, Lee JH, Aoki I, Koretsky AP (2004) Manganese-enhancedmagnetic resonance imaging (MEMRI): methodological andpractical considerations. NMR Biomed 17:532–543
Skibbe U, Christeller JT, Eccles CD, Laing WA, Callaghan PT (1995)A method to distinguish between chemical shift and suscepti-bility effects in NMR micoscopy and its application to insectlarvae. Magn Reson Imag 13:471–479
Slowik TJ, Green BL, Thorvilson HG (1997) Detection of magnetismin the red imported fire ant (Solenopsis invicta) using magneticresonance imaging. Bioelectromagnetics 18:396–399
Stecyk JAW, Bock C, Overgaard J, Wang T, Farrell AP, Portner HO(2009) Correlation of cardiac performance with cellular ener-getic components in the oxygen-deprived turtle heart. Am JPhysiol Regul Integr Comp Physiol 297:756–768
Straub J, Jurina K (2001) Magnetic resonance imaging in chelonians.Semin Avian Exotic Pet Med 10:181–186
Synthesys Network Activity F Pilot Study ‘‘Report on the technicalparameters of MRI’’. http://www.synthesys.info/na_f.htm
Takahashi MK, Kato K, Matsushita K, Umeda M, Nishina M, Hori E,Takahashi M, Oshaka A (1989) 1H-MRI and in vivo 31P-MRSstudy of diseased state of smoky-brown cockroach infected withcockroach densovirus. Jpn J Sanit Zool 40:289–293
Tindemans I, Verhoye M, Balthazart J, van der Linden A (2003)In vivo dynamic ME-MRI reveals differential functionalresponses of RA- and area X-projecting neurons in the HVCof canaries exposed to conspecific song. Eur J Neurosci18:3352–3360
Tomanek B, Jasinski A, Sulek Z, Muszynska J, Kulnowski P,Kwiecinski S, Krzyzak A, Skorka T, Kibinski J (1996) Magneticresonance microscopy of internal structure of drone and queenhoney bees. J Apic Res 35:3–9
Tyszka JM, Fraser SE, Jacobs RE (2005) Magnetic resonancemicroscopy: recent advances and applications. Curr OpinBiotechnol 16:93–99
Ullmann JF, Cowin G, Kurniawan ND, Collin SP (2010) Magneticresonance histology of the adult zebrafish brain: optimization offixation and gadolinium contrast enhancement. NMR Biomed23:341–346
Ulmer S, Jansen O (2010) fMRI. Basics and clinical applications.Springer, Berlin
Valente ALS, Cuenca R, Zamora MA, Parga ML, Lavin S, Alegre F,Marco I (2006) Sectional anatomic and magnetic resonanceimaging of coelomic structures of loggerhead sea turtles. Am JVet Res 67:1347–1353
Van de Moortele PF, Akgun C, Adriany G, Moeller S, Ritter J,Collins CM, Smith MB, Vaughan JT, Ugurbil K (2005) B1destructive interferences and spatial phase patterns at 7 T with ahead transceiver array coil. Magn Reson Med 54:1503–1518
Van den Burg EH, Verhoye M, Peeters RR, Meek J, Flik G, van derLinden A (2006) Activation of a sensorimotor pathway inresponse to a water temperature drop in a teleost fish. J Exp Biol209:2015–2024
Van der Linden A, Verhoye M, Portner HO, Bock C (2004) Thestrengths of in vivo magnetic resonance imaging (MRI) to studyenvironmental adaptational physiology in fish. Magn ResonMater Phys Biol Med 17:236–248
Van der Linden A, van Camp N, Ramos-Cabrer P, Hoehn M (2007)Current status of functional MRI on small animals: application tophysiology, pathophysiology, and cognition. NMR Biomed20:522–545
Van der Linden A, van Meir V, Boumans T, Poirier C, Balthazart J(2009) MRI in small brains displaying extensive plasticity.Trends Neurosci 32:257–266
Zoomorphology (2011) 130:227–254 253
123

Veliyulin E, Borge A, Singstad T, Gribbestad I, Erikson U (2006)Post mortem studies of fish using magnetic resonance imaging.Mod Magn Reson 1:959–966
Veliyulin E, Felberg HS, Digre H, Martinez I (2007) Non-destructivenuclear magnetic resonance image study of belly bursting inherring (Clupea harengus). Food Chem 101:1545–1551
Von Brand E, Cisterna M, Merino G, Uribe E, Palma-Rojas C,Rosenblitt M, Albornoz JL (2009) Non-destructive method tostudy the internal anatomy of the Chilean scallop Argopectenpurpuratus. J Shellfish Res 28:325–327
Walker TG, Happer W (1997) Spin-exchange optical pumping ofnoble-gas nuclei. Rev Mod Phys 69:629–642
Walter T, Shattuck DW, Baldock R, Bastin ME, Carpenter AE, Duce S,Ellenberg J, Fraser A, Hamilton N, Pieper S, RaganMA, SchneiderJE, Tomancak P, Heriche JK (2010) Visualization of image datafrom cells to organisms. Nat Methods Suppl 7:S26–S41
Wang LC, Wan DP, Jung SM, Chen CC, Wong HF, Wan YL (2005)Magnetic resonance imaging findings in the brains of rabbitsinfected with Angiostrongylus cantonensis: a long-term investi-gation. J Parasitol 91:1237–1239
Watanabe T, Schachtner J, Krizan M, Boretius S, Frahm J, MichaelisT (2006) Manganese-enhanced 3D MRI of established anddisrupted synaptic activity in the developing insect brain in vivo.J Neurosci Methods 158:50–55
Webb GA, Belton PS, Gil AM, Delgadillo I (2001) Magneticresonance in food science: a view to the future. RSC Publishing,Cambridge
Wecker S, Hornschemeyer T, Hoehn M (2002) Investigation of insectmorphology by MRI: assessment of spatial and temporalresolution. Magn Reson Imag 20:105–111
Weinmann HJ, Ebert W, Misselwitz B, Schmitt-Willich H (2003)Tissue-specific MR contrast agents. Eur J Radiol 46:33–44
Westheide W, Rieger R (2007) Spezielle Zoologie, Teil 1: Einzellerund wirbellose Tiere. 2. Auflage. Spektrum AkademischerVerlag, Munchen
Westheide W, Rieger R (2010) Spezielle Zoologie, Teil 2: Wirbel-oder Schadeltiere. 2. Auflage. Spektrum Akademischer Verlag,Munchen
Zhang X, Schneider JE, Portnoy S, Bhattacharya S, Henkelman RM(2010) Comparative SNR for high-throughput mouse embryoMR microscopy. Magn Reson Med 63:1703–1707
Ziegler A (in press) Broad application of non-invasive imagingtechniques to echinoids and other echinoderm taxa. In: Proceed-ings of the 7th European Conference on Echinoderms. LectureNotes Proceedings. Springer, Berlin
Ziegler A, Mueller S (2011) Analysis of freshly fixed and museuminvertebrate specimens using high-resolution, high-throughputMRI. In: Schroder L, Faber C (eds) In vivo NMR imaging.Methods in Molecular Biology 771. Humana Press, New York,pp 633–651
Ziegler A, Faber C, Mueller S, Bartolomaeus T (2008) Systematiccomparison and reconstruction of sea urchin (Echinoidea)internal anatomy: a novel approach using magnetic resonanceimaging. BMC Biol 6:33
Ziegler A, Bartolomaeus T, Mueller S (2010a) Sea urchin (Echinoi-dea) anatomy revealed by magnetic resonance imaging and 3Dvisualization. In: Harris LG, Boettger SA, Walker CW, LesserMP (eds) Echinoderms: Durham. CRC Press, Boca Raton,pp 305–310
Ziegler A, Ogurreck M, Steinke T, Beckmann F, Prohaska S, ZieglerA (2010b) Opportunities and challenges for digital morphology.Biol Direct 5:45
Ziegler A, Mooi R, Rolet G, De Ridder C (2010c) Origin andevolutionary plasticity of the gastric caecum in sea urchins(Echinodermata: Echinoidea). BMC Evol Biol 10:313
Ziegler A, Mietchen D, Faber C, von Hausen W, Schobel C, SellererM, Ziegler A (2011) Effectively incorporating selected multi-media content into medical publications. BMC Med 9:17
254 Zoomorphology (2011) 130:227–254
123
Copyright © 2022 FDOKUMEN




















