Islet transplantation versus insulin therapy in patients with type ...

Apoptosis in the Pancreatic Islet Cells of the NeonatalRat Is Associated with a Reduced Expression ofInsulin-Like Growth Factor II that May Act as a SurvivalFactor*
J. PETRIK, E. ARANY, T. J. MCDONALD, AND D. J. HILL
Lawson Research Institute, St. Joseph’s Health Center, London, Ontario, Canada N6A 4V2; and theDepartments of Physiology (J.P., D.J.H.), Medicine (E.A., D.J.H.), Biochemistry (T.J.M.),Pharmacology and Toxicology (T.J.M.), and Pediatrics (J.P., D.J.H.), University of Western Ontario,and London Health Sciences Center (T.J.M.), London, Ontario, Canada N6A 5A5
ABSTRACTIslet cell ontogeny will define adult b-cell mass and will consist of
a balance of islet cell birth and death. We have investigated theontogeny of factors that may be related to developmental apoptosis inthe islets, insulin-like growth factor II (IGF-II) and inducible nitricoxide synthase (iNOS), in pancreata of young Wistar rats. Pancreatawere collected from rats of 21 days gestation to 29 days postnatal age.In situ hybridization and immunohistochemistry showed that IGF-IIwas expressed and present in fetal and neonatal islet cells, but de-clined rapidly 2 weeks after birth. Little IGF-I was associated withfetal or postnatal islets. Apoptosis in islet cells was visualized bymolecular histochemistry for DNA breakage in tissue sections. Apo-ptosis was low in the fetus, but increased in incidence postnatally sothat 13% of islet cells were undergoing apoptosis on postnatal day 14,with the incidence declining thereafter. Immunohistochemistry foriNOS showed that it was expressed within b-cells and was most
abundant 12 days after birth. When islets were isolated from ratpancreata 20–22 days after birth, islet cell viability, DNA syntheticrate, and insulin release were reduced after incubation with inter-leukin-1b, tumor necrosis factor, or interferon-g. An increased rate ofislet cell survival was found after simultaneous incubation with IGF-Ior -II. Cytokine-mediated islet cell death involved the induction ofapoptosis. Islets isolated from neonatal rats were not killed afterexposure to these cytokines at the same concentrations, but cytokine-induced cell death was seen when neonatal islets were incubated witha neutralizing antibody against IGF-II. These experiments show thata peak of islet cell apoptosis that is maximal in the rat pancreas 14days after birth is temporally associated with a fall in the islet cellexpression of IGF-II. IGF-II was shown to function as an islet survivalfactor in vitro. The induction of islet cell apoptosis in vivo may involvean increased expression of iNOS within b-cells. (Endocrinology 139:2994–3004, 1998)
IN THE RAT fetus, the cellular area immunostained forinsulin increases 2-fold over 2 days just before term due
to both b-cell replication and recruitment, and maturation ofundifferentiated b-cell precursors (1, 2). Endocrine cells de-velop from duct-like cells in the embryo, fetus, and neonateand form primitive islets in the mesenchyme adjacent to theducts. Final differentiation into glucagon-, somatostatin-,pancreatic polypeptide-, or insulin-expressing cells probablydepends on the expression of transcription factors such asPdx-1 (3) and on the actions of local peptide growth factorswithin the surrounding mesenchyme (4). The populationgrowth rate of all islet cells, including b-cells, slowed bypostnatal days 3 and 4 and continued to decline thereafter (2).The rate of mitosis in adult pancreatic b-cells is normally low(3% replication rate of b-cells/day) (5).
The change from a fetal type of b-cell population capableof expansion, to an adult b-cell population that is not mayoccur within a programed ontological pathway that extends
into postnatal life. The newborn rat made diabetic with strep-tozotocin has extensive b-cell destruction, but at 14 days afterbirth demonstrates normal glycemia, with considerable mi-totic activity apparent in the pancreatic ductal epitheliumfrom which the precursor b-cells derive (6). Similar b-cellrenewal occurs after alloxan treatment of the young rabbit(7). Pancreatectomy (90%) of the young rat is followed byregeneration of both exocrine and endocrine tissues (8). Newb-cells are derived by both neogenesis and cell replicationwithin the remaining islets. Endocrine regeneration is alsoseen in association with duct epithelial cell proliferation intransgenic mice with b-cells expressing interferon-g (IFNg)(9). In these mice, b-cell destruction leading to IDDM can bematched by new islets growing ectopically into duct lumens.Islet regeneration occurs in the pancreas of young diabeticpatients, and in some cases, islet neogenesis is associatedwith centroacinar and ductular cells, leading to the formationof large islets consisting primarily of b-cells (10). Islet cellregeneration in the pancreas of recent-onset IDDM patientsalso occurs, particularly in infants and small children (11, 12).Recently, it has been revealed that the ontogeny of islet cellsin early life involves a balance between b-cell replication andneogenesis, and programed b-cell death. A transient wave ofapoptosis occurs in neonatal rat islets between 1–2 weeks ofage (5, 13). However, b-cell mass is not altered appreciablyat this time, suggesting that a new population of b-cells
Received November 6, 1997.Address all correspondence and requests for reprints to: Dr. D. J.
Hill, Lawson Research Institute, St. Joseph’s Health Center, 268 Gros-venor Street, London, Ontario, Canada N6A 4V2. E-mail: [email protected].
* This work was supported by the Juvenile Diabetes Foundation, theCanadian Diabetes Association, and the Medical Research Council ofCanada.
0013-7227/98/$03.00/0 Vol. 139, No. 6Endocrinology Printed in U.S.A.Copyright © 1998 by The Endocrine Society
2994

compensates for the loss. A similar episode of b-cell apo-ptosis has recently been described in the human fetal pan-creas in third trimester (14). Although the cellular triggersinvolved in the initiation of developmental b-cell apoptosisare not known, the apoptosis that mediates b-cell destructionin response to cytokine action during autoimmune diabetesinvolves an increased intracellular concentration of nitricoxide (NO) and increased expression of inducible nitric oxidesynthase (iNOS) (15).
The mechanisms controlling islet cell neogenesis or apo-ptosis during development are largely unknown, but thesemay be linked to the relative expression of peptide growthfactors within the developing pancreas. There is considerablecircumstantial evidence that the insulin-like growth factors(IGFs) are major contributors to b-cell growth, maturation,and function and are expressed by b-cells throughout life.We previously reported that IGF-II messenger RNA (mRNA)is most abundant in the pancreas of the fetal rat and declinesduring the neonatal period (16). Conversely, IGF-I mRNAlevels were low, but detectable, in fetal life and rose to adultlevels within 2 weeks after birth. Others have reported thatIGF-I mRNA abundance peaked transiently shortly afterbirth and then declined sharply (17), a pattern found by usfor IGF-II, but not IGF-I, mRNA. Using in situ hybridizationand immunohistochemistry, IGF-I and -II mRNAs and pep-tides were shown to be present within islet cells throughoutlife, including b-cells (18). Similarly, in the midtrimester hu-man fetus, IGF peptides and IGF-binding protein-1 (IGFBP-1), -2, and -3 were localized by immunohistochemistry to theislets of Langerhans and b-cells (19). We and others showedthat isolated islets from the human (20) or rat fetus (21, 22)or neonate (23, 24) release both IGF-I and IGF-II, that exog-enous IGF-I or -II promotes increased islet cell DNA syn-thesis (21, 22, 25), and that isolated a- and b-cells from ratislets contain the high affinity type 1 IGF signaling receptor(26), as do pancreatic b-cell lines (27). We also showed thatisolated adult rat islets enriched in b-cells release IGF-I (28),whereas others have demonstrated a retention of IGF-IImRNA expression within the b-cells of the pancreas in bothrat and man (29, 30), and high levels of IGF-II are expressedby the rat b-cell line, INS-1 (31).
The transient wave of developmental apoptosis responsi-ble for a reduction in b-cell number after 1–2 weeks of post-natal life in the rat (13) coincides temporally with our dem-onstration of a diminished pancreatic expression of IGF-II atthis time, whereas pancreatic expression of IGF-I has not yetachieved adult values (16). A nadir in total IGF availabilitymay therefore exist in pancreas when apoptosis is transientlyhigh. IGFs inhibit apoptosis in mammary carcinoma cells,cerebellar granule neurons, ovarian preovulatory follicles,human erythroid colony-forming cells, and hematopoieticcells (32–35). However, IGF-I inhibits iNOS induction insome tissues (36) and may interfere with cytokine-stimulatedNO synthesis. The ability of IGF-I to limit neuronal damageelicited by experimental hypoxia-ischemia suggests an abil-ity of IGF-I to limit free radical generation (37).
In this study we have examined the precise anatomicalpattern of expression of IGF-I and -II in the pancreas of therat during early life, the relationship between developmentalb-cell apoptosis and the presence of iNOS, and present func-
tional evidence that IGFs can influence islet b-cell survival inthe neonatal rat.
Materials and MethodsAnimals
Pregnant Wistar rats (Charles River, Montreal, Canada) were killedat day 21 of gestation or were allowed to deliver and the offspring werekilled on postnatal days 4, 6, 7, 12, 14, 16, 18, 22, or 29. Rat fetuses weredecapitated with scissors, whereas the pregnant females and all rats atpostnatal ages were killed by CO2 asphyxiation. Pregnant and sucklingrats were allowed free access to food and water. All procedures wereperformed with ethical approval of the animal care committee of theUniversity of Western Ontario. After death, the pancreas was immedi-ately removed from each animal and placed in 5 ml sterile, ice-coldHanks’ Buffered Salts Solution, pH 7.5 (HBSS; Life Technologies, Bur-lington, Canada) if islets were to be isolated. If pancreata were to be fixedfor histology, they were placed in ice-cold fixative (0.2% glutaraldehydeand 4% paraformaldehyde buffered with 70 mm phosphate buffer, pH7.4) for 16 h at 4 C, followed by four washes at 4 C in PBS over a 48-hperiod. Fixed tissues were dehydrated in 70% (vol/vol) ethanol andembedded in paraffin.
Islet isolation
The islet isolation technique was modified from that of Hellerstromet al. (38). The pancreata were placed in sterile, ice-cold HBSS, pH 7.4,containing penicillin (100 U/ml), streptomycin (0.1 mg/ml), and fun-gizone (0.25 mg/ml; Life Technologies). After removal of gut and spleenremnants, the pancreata were transferred to small, sterile glass vials(;10 pancreata/vial for fetal or neonatal islets, 5 pancreata/vial forolder animals up to 22 days, and a single pancreas for the adult) andwashed with ice-cold HBSS. HBSS was removed from each of the vialsand replaced with HBSS containing 2 mg/ml collagenase (type V; SigmaChemical Co., St. Louis, MO). The vials were shaken for 4–5 min in awater bath (200 cycles/min) at 37 C for tissues from fetal and neonatalanimals up to 6 days age and for 5–8 min for tissues from older rats. Thedigestion was terminated by the addition of 15 ml ice-cold HBSS to eachvial. The digest was dispersed by pipette, and tissue was collected bycentrifugation after washing with ice-cold HBSS. The washed digest wasresuspended in 5 ml tissue culture medium, RPMI 1640, pH 7.4, con-taining 11.1 mm glucose and 25 mm HEPES (Life Technologies) andsupplemented with antibiotics (100 U/ml penicillin and 0.1 mg/mlstreptomycin), fungizone (0.25 mg/ml), and 10% heat-inactivated, virus-and mycoplasma-free FBS (Life Technologies). The digest was distrib-uted in 1-ml portions onto tissue culture petri dishes (100 mm; Nunc,Toronto, Canada) containing 14 ml of an identical tissue culture me-dium. The culture plates were incubated for 48–72 h at 37 C in ahumidified atmosphere of 5% CO2 in air. Islets were counted by eyeunder a dissecting microscope, detached from the culture dish, har-vested using an automatic pipette, and transferred to nontissue culturegrade petri dishes (100 mm; Falcon, Lincoln Park, NJ) that did not permitcell attachment for the experimental period. A typical yield of islets from10 pancreata was approximately 1500.
Islet culture
At the beginning of the treatment period, the islets were distributedin equal batches (60–80 islets/plate) onto nontissue culture grade petridishes (50 mm; Falcon) containing 1 ml tissue culture medium. Culturemedium consisted of glucose-free DMEM (Imperial Laboratories, An-dover, UK), pH 7.4, containing antibiotics and fungizone (as describedabove) and supplemented with 2 mm glutamine (Life Technologies) and8.7 mm glucose. Medium was further supplemented as required withrecombinant human IGF-I or IGF-II (100 ng/ml; GroPep, Adelaide,Australia), a monoclonal antibody against rat IGF-II (20 mg/ml; AmanoInternational Enzyme Co., Troy, VA), recombinant human interleu-kin-1b (IL-1b; 2.5 ng/ml), tumor necrosis factor-a (TNFa; 10 ng/ml),IFNg (10 ng/ml; R&D Systems, Minneapolis, MN), or rabbit antihumanplacental lactogen (Dimension Laboratories, Mississauga, Canada),alone or in combination. Where IGF-I or -II and cytokines or IGF-IIantiserum and cytokines were present together, the IGF or antiserum
IGF-II AND ISLET CELL APOPTOSIS 2995

was added 5 min before the cytokine and allowed to equilibrate with theislets at 37 C. Islets were incubated for 24, 48, or 72 h. In some exper-iments, [methyl-3H]thymidine (20 Ci/mm; ICN Biomedicals, Irvine, CA)at 5 mCi/ml was added for the final 24 h of culture. At the end of thetreatment period, conditioned medium was removed, and the islets werewashed in PBS (Life Technologies) and assessed for either viability orDNA synthetic rate, as described below. Separate experiments wererequired for each type of analysis.
In some experiments islets were plated into 8-well chamber slides(Lab-Tek, Nalge-Nunc International, Naperville, IL; 20 islets/well) andallowed to attach to the glass slides overnight in a humidified incubatorat 37 C, gassed as described above, before the addition of IGF-I or -II orcytokines, alone and in combination. After 48 h, the culture medium wasremoved, and the islets were washed in PBS. Islets attached duringculture to chamber slides were dehydrated through ascending ethanolconcentrations (50%, 70%, 90%, and 100%), and air-dried. Slides werestored with desiccant until histological processing.
Assessment of islet viability
To assess islet cell viability after incubation in test culture medium,the islets from each plate were resuspended in 0.5 ml PBS containing 5mg/ml trypan blue (Sigma). Trypan blue solution was filtered throughan ultramembrane (0.2 mm; Gelman Science, Ann Arbor, IL) to removeparticulates before use. All islets were examined immediately under adissecting microscope, and any islet containing one or more cells thathad taken up trypan blue was considered nonviable. At the time ofviability assessment, the recovery of islets was 92 6 5% (mean 6sd) ofthose initially added to each culture dish, and this did not differ betweencontrol cultures or those that had contained cytokines. The within-batchcoefficient of variation on assessment of islet viability after repeatedmeasures was 3%.
Estimation of islet DNA content and synthetic rate
Islets were suspended in PBS (500 ml) and solubilized by ultrasoni-cation. DNA was precipitated with 1 ml ice-cold 10% trichloroacetic acidand solubilized by overnight incubation with 1 m sodium hydroxide.After neutralization with 1 m HCl, the DNA content of islets was mea-sured by fluorometry using Hoechst fluorochrome 33258 (AldrichChemical Co., Milwaukee, WI) with an excitation wavelength of 375 nmand an emission wavelength of 458 nm. Calf thymus DNA (1.5–24mg/ml; Sigma) was used for calibration. To measure the rate of DNAsynthesis, [3H]thymidine incorporation was measured by liquid scin-tillation counting. Two 50-ml aliquots of homogenized islets were re-moved, and the DNA was precipitated in 500 ml ice-cold 5% trichloro-acetic acid. The precipitates were collected by filtration through a glass-fiber disk (2.5 cm; Whatman GF/A, Whatman International, Maidstone,UK). Any remaining free isotope was removed by washing with distilledwater. The radioactivity on the filters was determined by liquid scin-tillation counting (LS 5000 TD b-counter, Beckman, Palo Alto, CA). Theincorporation of [3H]thymidine was expressed as disintegrations permin/mg DNA.
Insulin release
The insulin content of conditioned culture medium was measured byRIA using Wright antiserum in a modification of the method of Halesand Randle (39) as modified by Herbert et al. (40) and described by uspreviously (41). Rat insulin (Novo Nordisk, Mississauga, Canada) wasused for the standard curve. The within-assay coefficient of variationwas 6.5%, and the between-assay coefficient of variation was 9%. Theminimum level of detection was 2 fmol/ml. There was no detectablecross-reactivity in the insulin assay with IGF-I or -II.
Immunohistochemistry
Histological sections of pancreas (5 mm) were cut from paraffin blockswith a rotary microtome and mounted on poly-l-lysine-coated glassmicroscope slides. Immunohistochemistry was performed to localizeIGF-I or -II, insulin, or iNOS within islets by a modified avidin-biotinperoxidase method (42). Sections were deparaffinized in xylene, rehy-drated in a descending ethanol series (100%, 90%, and 70%, vol/vol), and
washed in PBS before incubation in 1% (vol/vol) hydrogen peroxide toblock endogenous peroxide activity, followed by a 15-min incubation in5% BSA in PBS to reduce nonspecific binding. Slides were incubated for48 h at 4 C in a humidified chamber with either rabbit antihuman IGF-Ior IGF-II (1:2000 dilution; GroPep), guinea pig antiinsulin antibody(1:1000 dilution; provided by Dr. T. J. McDonald), or mouse anti-iNOSantiserum (1:50 dilution; Transduction Laboratories, Lexington, KY) in0.01 m PBS (pH 7.5) containing 2% (wt/vol) BSA and 0.01% (wt/vol)sodium azide (100 ml/slide). All subsequent incubations were per-formed at room temperature. Biotinylated goat antirabbit IgG (1:500),goat antimouse IgG (1:100), or mouse antiguinea pig IgG (1:100) (VectorLaboratories, Burlingame, CA) were diluted in the same buffer andapplied for 2 h, then the slides were washed in PBS and incubated withavidin and biotinylated horseradish peroxidase for 1 h. After washingin PBS and 50 mm Tris-HCl, pH 7.5, peptide immunoreactivity waslocalized by incubation in fresh 1.89 mm diaminobenzidine with 0.03%(vol/vol) hydrogen peroxide for 2 min, and the reaction was quenchedin excess 50 mm Tris-HCl, pH 7.5. Tissue sections were counterstainedwith Carazzi’s hematoxylin, dehydrated in an ascending ethanol series,then cleared with xylene and mounted under glass coverslips. To es-tablish the specificity of staining, the primary antisera to IGF-I or -II werepreabsorbed overnight at 4 C with 100 nm homologous ligand beforeapplication to the sections. In each case, staining was abolished. Furthercontrols included substitution of primary antisera with nonimmuneserum and omission of the secondary antiserum.
Dual staining for iNOS and insulin was performed by first perform-ing immunohistochemistry for insulin as described above, using dia-minobenzidine as the chromagen. Before counterstaining and dehydra-tion, the sections were subjected to immunohistochemistry for iNOS asdescribed, except that alkaline phosphatase (blue) was used as thechromagen. Alkaline phosphatase substrate kit III was obtained fromVector Laboratories. Antimouse alkaline phosphatase conjugate (Sigma)was applied to each section for 1 h at room temperature, sections werewashed, and alkaline phosphatase substrate was applied for 20 min.Sections were washed and counterstained with Mayer’s hemalum.
Visualization of apoptosis
Immunocytochemistry was performed to localize apoptotic nucleiwithin either tissue sections or isolated islets (43) using the Apoptag insitu apoptosis detection kit (Oncor, Gaithersburg, MD). Islets attachedduring culture to chamber slides were rehydrated through descendingethanol concentrations (100%, 90%, and 70%) and incubated in PBS for5 min. Staining was performed according to the manufacturer’s protocolafter incubation with proteinase K (20 mg/ml; Boehringer Mannheim,Dorval, Canada) for 15 min, washing in distilled water, and the quench-ing of endogenous peroxidase by incubation in 2% (vol/vol) hydrogenperoxide in PBS for 5 min. Color was generated with diaminobenzidine[1.89 mm activated with 0.03% (vol/vol) hydrogen peroxide for 2 min],and the tissue was counterstained with methyl green for 1 min. Sectionswere dehydrated in alcohols, cleared in xylene, and mounted withPermount (Fisher Scientific, Fairlawn, NJ) under glass coverslips. Dualstaining for insulin was performed by incubation with antiinsulin an-tiserum after detection of apoptosis and before dehydration. In this casethe chromagen used for insulin immunoreactivity was alkaline phos-phatase (as described above).
Northern blot analysis
Total RNA was extracted from isolated islets as previously described(16). Before hybridization, the integrity and relative amounts of RNAfrom each islet batch were assessed by size separation on urea-agarosegels by analytical electrophoresis, followed by ethidium bromide stain-ing. Islets in which ribosomal RNA showed degradation were not usedfor experiments.
For Northern blot hybridization, between 15–20 mg total RNA weredenatured for 15 min at 65 C in 6% (vol/vol) deionized formaldehyde(Fluka Chemicals, Georgetown, Canada) and 50% (vol/vol) deionizedformamide (Fluka) in 1 3 hybridization buffer [20 mm MOPS (mor-pholinopropanesulfonic acid), 5 mm sodium acetate, and 5 mm EDTA,pH 7.0] and separated by size on 1% agarose gels containing 6% deion-ized formaldehyde. The separated RNA was then transferred to Zeta-Probe membranes (Bio-Rad, Richmond, CA) by the capillary transfer
2996 IGF-II AND ISLET CELL APOPTOSIS Endo • 1998Vol 139 • No 6

technique (44). The blots were prehybridized for 2 h in hybridizationbuffer containing 1 3 SSPE (150 mm NaCl, 10 mm sodium phosphatemonobasic, and 1 mm EDTA), 7% (wt/vol) SDS, 100 mg/ml salmonsperm DNA, and 50% deionized formamide. The blots were thenhybridized in the same buffer at 42 C overnight in a shaking waterbath with 2 3 106 cpm/ml radiolabeled complementary DNA (cDNA)probe for IGF-I or -II or with 1 3 106 cpm/ml for a cDNA encoding18S ribosomal RNA. After hybridization, the blots were washed suc-cessively with 1 3 SSC (standard saline citrate; 0.75 m NaCl and 0.075m sodium citrate) with 0.1% SDS (wt/vol) once for 30 min at 22 C,followed by 30 min at 42 C. Two final washes were performed, eachfor 30 min, at 42 C in 0.1 3 SSC with 0.1% (wt/vol) SDS. Blots wereair-dried and exposed to x-ray film (Kodak XAR-5, Eastman Kodak,Rochester, NY) at 270 C with intensifying screens for up to 10 daysbefore developing. Blots were hybridized consecutively with up tothree different cDNAs. Between consecutive hybridizations, the blotswere washed with 0.01 3 SSC with 0.5% (wt/vol) SDS at 90 C for 1 hwith two changes of washing solution to remove the previous cDNAprobe.
Radiolabeling of cDNA probes with [a-32P]deoxy-CTP (ICN Bio-medicals, Irvine, CA) was carried out by random priming using aPharmacia oligolabeling kit (Pharmacia LKB Biotechnology, Uppsala,Sweden) to specific activities of 1–2 3 109 dpm/mg. Separation ofradiolabeled cDNA from unincorporated [32P]deoxy-CTP was carriedout using Pharmacia NICK columns containing Sephadex G-50 (Phar-macia). cDNA probes used for hybridization of Northern blots andfor in situ hybridization were provided by the following investigators.A 500-bp rat IGF-I cDNA in pGEM Blue (Promega, Madison, WI)encoding exon 3 and part of exon 4 of the recombinant IGF-I gene wasprovided by Dr. L. Murphy, University of Manitoba (Winnipeg, Can-ada) (45). A 807-bp mouse IGF-II in pGEM 4z was provided by Dr.G. Bell, University of Chicago (Chicago, IL). A rat 18S ribosomal RNAcDNA was provided by Dr. D. Denhardt, Rutgers University (Pis-cataway, NJ). The latter was used to determine equality of RNAloading and transfer.
In situ hybridization
Sections of pancreas were deparaffinized in xylene, rehydrated in adescending ethanol series (100%, 90%, and 70%, vol/vol), and washedin PBS, then permeabilized with 10 mg/ml proteinase K (BoehringerMannheim) in 0.1 m Tris-HCl (pH 8.0) and 50 mm EDTA at 37 C for 30min. After rinsing in PBS, sections were dipped in 0.1 m triethanolamine-acetic anhydride for 10 min at room temperature, rinsed again in PBS,then dehydrated in a graded ethanol series (70%, 90%, and 100%, vol/vol), air-dried, and stored at 270 C until hybridization. Sections wereprehybridized by incubation at 45 C for 16 h in humidified chambers in100 ml 1 3 hybridization buffer [50% deionized formamide, 0.3 m NaCl,20 mm Tris-HCl (pH 8.0), 1 mm EDTA, 1 3 Denhardt’s solution, 500mg/ml yeast transfer RNA, 100 mg/ml denatured salmon sperm DNA,10% (wt/vol) dextran sulfate, 0.1% SDS, and 100 mm dithiothreitol(DTT)]. Radiolabeled complementary RNA (cRNA) probes (106 cpm)were diluted with 25 ml water, denatured at 80 C for 2 min, and madeup to the required volume for each slide with 1.3 3 hybridization buffer.Each slide was incubated with cRNA probe under glass coverslips in ahumidified chamber at 50 C for 16 h, the coverslips were removed bysoaking the slides in 10 mm DTT in 2 3 SSC, and after a further incu-bation at 55 C for 10 min in 1 3 hybridization buffer, the sections weretreated with 20 mg/ml ribonuclease A (RNase A), 1 U/ml RNase T1 in0.5 m NaCl, 10 mm Tris-HCl (pH 8.0), and 1 mm EDTA at 37 C for 30min. Sections were washed as follows: twice for 30 min at room tem-perature in 2 3 SSC, twice for 30 min at 55 C, then twice for 15 min atroom temperature in 0.1 3 SSC. To screen the extent of RNA hybrid-ization, slides were subjected to autoradiography with Kodak XAR filmafter dehydration in an ascending ethanol series (70%, 90%, and 100%,vol/vol) and air-dried. Kodak NTB-3 photoemulsion diluted 1:1 withwater was applied subsequently to all sections and exposed for up to 14days at 4 C, then slides were developed in Kodak D19, rinsed in water,and fixed in Kodafix. Sections were counterstained with Carazzi’s he-matoxylin and dehydrated in an ascending ethanol series (70%, 90%, and100%, vol/vol) with a final wash in xylene, and coverslips were appliedwith Permount (Fisher Scientific, Jersey City, NJ). Slides used for in situhybridization were viewed under dark- and lightfield microscopy.
The restriction enzymes (BRL, Burlington, Canada) and RNA poly-merases (Promega) used to linearize the plasmids containing these cD-NAs and to generate 35S-radiolabeled riboprobes are as follows: anti-sense rat IGF-I, HindIII/T7; sense rat IGF-I, PvuII/SP6; antisense mouseIGF-II, HindIII/TSP6; and sense mouse IGF-II, EcoRI/T7. RadiolabeledcRNA probes were synthesized by adding the following to a sterilemicrofuge tube: 4 ml 5 3 transcription buffer [200 mm Tris-HCl (pH 7.5),30 mm MgCl2, 10 mm spermidine, and 50 mm NaCl]; 2 ml 100 mm DTT;0.8 ml (20 U) RNase inhibitor; 4 ml 2.5 mm GTP, ATP, UTP, or CTP; 2.4ml 100 mm CTP or UTP; 1 ml (200 ng) linearized riboprobe DNA; 5 mla-thio-[35S]UTP (400 Ci/mmol; New England Nuclear, Billerica, MA),0.3 ml sterile water, and 1 ml (5–10 U) SP6, T3, or T7 RNA polymerase.Incubation was performed at 37 C for 1 h. To remove template DNA, 1ml deoxyribonuclease I containing no RNase activity (1 U/ml; BRL,Gaithersburg, MD) was added, and the reaction was further incubatedat 37 C for 15 min. Yeast transfer RNA was added (50 mg), and thereaction volume was increased to 450 ml with sterile water, then ex-tracted in sequence with equal volumes of phenol-chloroform and chlo-
FIG. 1. Immunohistochemical localization of apoptosing cells (ar-rows) using molecular histochemistry in a pancreatic islet from a14-day-old rat. The number of condensed, apoptotic cells is greatestat this time. Magnification bar 5 10 mm.
TABLE 1. Incidence of apoptotic cells in rat pancreatic islets(percentage of islet cells demonstrating apoptotic nuclei) inanimals from 21 days gestation until adulthood
Age % Islet cells
Fetal21 days 1.99 6 0.25
Postnatal4 days 5.90 6 1.236 days 6.46 6 0.6112 days 9.17 6 0.4714 days 13.18 6 3.4018 days 8.14 6 1.2522 days 2.36 6 0.91
Adult 0.23 6 0.04
Figures represent the mean 6 SEM (n 5 3–4).
IGF-II AND ISLET CELL APOPTOSIS 2997
roform-isoamyl alcohol. After ethanol precipitation with 0.1 vol 3 msodium acetate (pH 5.2) and 2.5 vol cold ethanol, RNA was pelleted bycentrifugation, washed in 70% (vol/vol) ethanol, dried, and resus-pended in 500 ml 0.2% (wt/vol) SDS, 2 mm EDTA, 0.5 m sodium acetate.35S-Labeled riboprobes were precipitated with ethanol as describedabove, dried, and dissolved in 200 ml 20 mm DTT. The quality and sizeof radiolabeled cRNA transcripts were determined by urea-agarose gelelectrophoresis of 10,000 cpm cRNA probe and autoradiography. Toimprove cRNA probe penetration of tissue sections, limited alkalinehydrolysis was used to reduce transcript size to about 150 bases.
Morphometric and statistical analysis
Morphometric analysis was performed using a Zeiss transmitted lightmicroscope and Incident-light fluorescence Axioskope (Carl Zeiss Can-ada, Ltd., North York, Ontario, Canada) at a magnification of 3250.Analyses were performed with Northern Eclipse version 2.0 morpho-metric analysis software (Empix Imaging Co., Mississauga, Canada).The percentage of islet cells immunopositive for iNOS, IGF-I, or IGF-IIor demonstrating apoptotic nuclei was calculated at each age from threesections of each pancreas, representing both head and tail regions. Sec-tions chosen contained at least five islets, and pancreata from at least fiveanimals were examined for each age.
Islet cultures were performed with three or four replicate cultureplates for each variable within an individual experiment derived froma pool of islets. Each experiment was repeated from three to five timesusing pools of islets derived from separate animals. Representativeexperiments are shown as the mean 6 sem. Differences between meanvalues for variables within individual experiments were compared forstatistical difference by ANOVA. For histological analyses of the pres-ence of IGF-I or -II, at least five separate pancreata were examined, andrepresentative views are shown.
Results
Sections of pancreas were stained for the presence of ap-optotic cells determined by DNA breakage to establish theontogeny of islet cell apoptosis. Apoptotic nuclei within con-densed islet cells with little cytoplasm were seen at all agesexamined (Fig. 1). The prevalence of apoptotic nuclei in en-docrine islet cells of fetuses at 21 days gestation was less than2%, but this increased postnatally to 9% on postnatal day 12
and was maximal at 13% on postnatal day 14 (Table 1). At 22days postnatal age, the incidence of islet cell apoptosis haddecreased to 2%. The majority of apoptotic cells were locatedcentrally within the islets, suggesting that they were b-cells.However, dual staining immunohistochemistry for both in-sulin and apoptosis failed to confirm this, as staining forinsulin could not be conclusively demonstrated within thecondensed, apoptosing cells. The occurrence of islet cell ap-optosis was a feature of 85% of the islets examined on post-natal day 14.
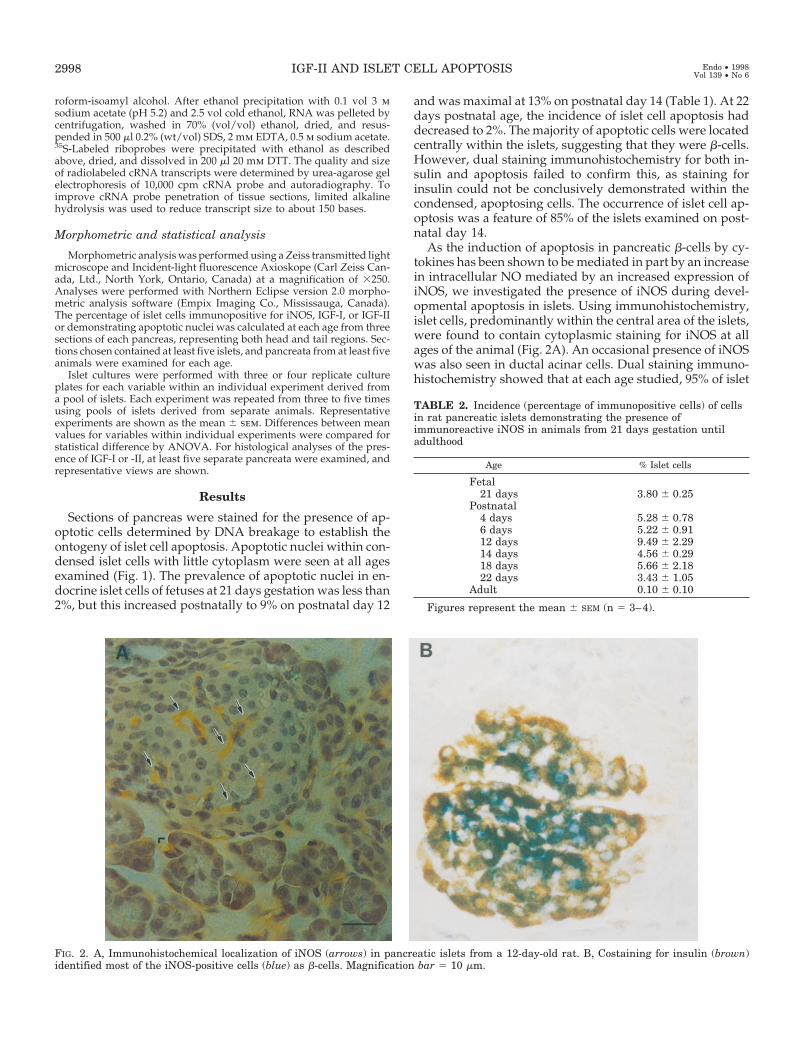
As the induction of apoptosis in pancreatic b-cells by cy-tokines has been shown to be mediated in part by an increasein intracellular NO mediated by an increased expression ofiNOS, we investigated the presence of iNOS during devel-opmental apoptosis in islets. Using immunohistochemistry,islet cells, predominantly within the central area of the islets,were found to contain cytoplasmic staining for iNOS at allages of the animal (Fig. 2A). An occasional presence of iNOSwas also seen in ductal acinar cells. Dual staining immuno-histochemistry showed that at each age studied, 95% of islet
FIG. 2. A, Immunohistochemical localization of iNOS (arrows) in pancreatic islets from a 12-day-old rat. B, Costaining for insulin (brown)identified most of the iNOS-positive cells (blue) as b-cells. Magnification bar 5 10 mm.
TABLE 2. Incidence (percentage of immunopositive cells) of cellsin rat pancreatic islets demonstrating the presence ofimmunoreactive iNOS in animals from 21 days gestation untiladulthood
Age % Islet cells
Fetal21 days 3.80 6 0.25
Postnatal4 days 5.28 6 0.786 days 5.22 6 0.9112 days 9.49 6 2.2914 days 4.56 6 0.2918 days 5.66 6 2.1822 days 3.43 6 1.05
Adult 0.10 6 0.10
Figures represent the mean 6 SEM (n 5 3–4).
2998 IGF-II AND ISLET CELL APOPTOSIS Endo • 1998Vol 139 • No 6

cells immunopositive for iNOS also contained insulin (Fig.2B). When pancreata from various ages of rat were com-pared, the percentage of islet cells containing immunoreac-tive iNOS was greatest at almost 10% on postnatal day 12 anddeclined thereafter to negligible levels by weaning (Table 2).Immunoreactive iNOS could not be demonstrated in apo-ptotic islet cells, but the compacted nature of these cellsrendered any cytoplasmic localization of ligands unlikely.
IGF-I and -II have been shown to prevent the experimentalinduction of apoptosis in a variety of cell types (32–35). Wehave previously demonstrated a fall in the whole pancreaticexpression of IGF-II in neonatal life (16), suggesting that thismay be functionally related to the developmental apoptosisof neonatal islets. To determine the anatomical location andontogeny of IGF-I and -II expression, we localized thesemRNAs in pancreatic sections using in situ hybridization andthe distribution of the peptides by immunohistochemistry. Insitu hybridization for IGF-II mRNA showed a relatively highsignal intensity within islet endocrine cells on postnatal day7 (Fig. 3A) in both the central and peripheral regions of theislets. Hybridization signal was also present in the ductalepithelial cells, but with relatively less abundance than inislets. Hybridization with a sense strand IGF-II cDNAshowed little signal, demonstrating its specificity in pancre-
atic tissues (not shown). A similar hybridization pattern wasseen on postnatal days 4 and 6. By postnatal day 29, nospecific hybridization signal for IGF-II mRNA remainedwithin pancreatic islets (Fig. 3, B and D). Immunohistochem-istry showed a widespread distribution of IGF-II peptide inall islet cells in late gestation and postnatal day 7, but this wasless intense on postnatal days 12 and 14 and was absent bypostnatal day 22 (Fig. 4). IGF-II was also visualized in aminority of ductal epithelial cells in early life. Conversely, nospecific hybridization signal for IGF-I mRNA was seen in thefetal or neonatal pancreas (Fig. 3C), and only a low level ofmRNA was detected in islets after weaning (Fig. 3D). How-ever, on postnatal day 29, IGF-I mRNA was seen withinductal epithelial cells. After immunohistochemistry for IGF-Ipeptide, little immunoreactivity was seen in the fetal or neo-natal islets or acinar tissue (Fig. 4). By postnatal day 22,immunoreactivity for IGF-I was distributed throughout theislet cells, but staining was not intense. Additionally, immu-noreactivity was located in the ductal epithelial cells. Thesestudies confirmed that IGF-II expression in the pancreasdeclined substantially in neonatal life and showed that themajor sites of IGF-II expression were the pancreatic islets.
We performed experiments with rat islets in vitro to de-termine whether either IGF-I or IGF-II could protect islet cells
FIG. 3. In situ hybridization to visualize IGF-II (A and B) or IGF-I (C and D) mRNAs in sections of rat pancreas from animals 7 days (A andC) or 29 days (B and D) of age. Is, Islets; E, exocrine tissue; Mes, mesenchymal connective tissue. Magnification bar 5 10 mm.
IGF-II AND ISLET CELL APOPTOSIS 2999

from cell death induced by exposure to cytokines, which hasbeen demonstrated previously to involve the initiation ofapoptosis (15). Islets isolated from rats on postnatal days20–22 days or from adult female rats were incubated in thepresence of single concentrations of IL-1b (2.5 ng/ml), TNFa(10 ng/ml), or IFNg (10 ng/ml), with or without IGF-I or -II(100 ng/ml), for 24 or 48 h. After 24-h incubation, the meanviability of islets from rats 20–22 days of age incubated in thepresence of each cytokine alone was significantly lower thanthat in control cultures for TNFa and IFNg, but not for IL-1b.After incubation for 48 h, cell viability in control cultures was90 6 3% (mean 6 sem; three separate experiments), dem-onstrating that the islet isolation and culture were associatedwith a low incidence of cell death. However, in the presenceof IL-1b, TNFa, or IFNg, each cytokine alone caused a 60–90% significant reduction in islet cell viability after 48 h(Table 3). Coincubation with either IGF-I or IGF-II signifi-cantly reduced cell mortality in the presence of IL-1b orTNFa, but not in the presence of IFNg. Analysis of the DNA
synthetic rate showed that under control conditions therewas incorporation of [3H]thymidine into islet cells of rats at20–22 days postnatal age, suggesting a potential for mito-genesis (Table 4). Exposure to IL-1b, TNFa, or IFNg aloneeach caused a significant reduction in [3H]thymidine incor-poration per mg DNA. Coincubation with either IGF-I or -II(100 ng/ml) significantly reversed the decrease in radiothy-midine incorporation seen with IL-1b or TNFa, but not withIFNg. Similar results were found when [3H]thymidine in-corporation was expressed per number of islets (not shown).The release of insulin into conditioned culture medium wassubstantially reduced in response to each of the cytokinesalone, and this was not reversed by the presence of IGF-I or-II (Table 5). Islets isolated from adult rats had similarlyreduced viability in response to cytokines, which was re-versed by coincubation with IGFs (not shown).
Islets from rats 20–22 days of age were grown in chamberslides and exposed to IL-1b for 48 h, with or without IGF-Ior -II (100 ng/ml). Islets were then fixed, and apoptotic nuclei
FIG. 4. Immunohistochemistry to visualize immunoreactive IGF-II (A and B) or IGF-I (C and D) in sections of rat pancreas from animals 7days (A and C) or 22 days (B and D) of age. Is, Islets; E, exocrine tissue; Mes, mesenchymal connective tissue. Magnification bar 5 10 mm.
3000 IGF-II AND ISLET CELL APOPTOSIS Endo • 1998Vol 139 • No 6
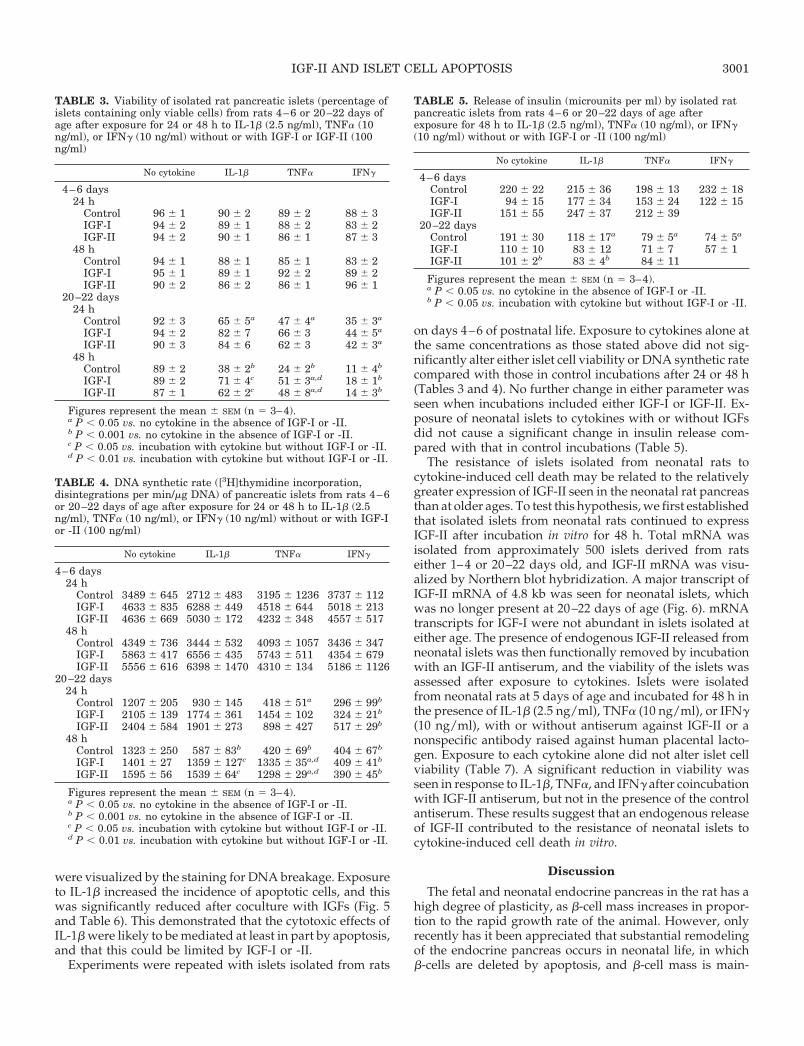
were visualized by the staining for DNA breakage. Exposureto IL-1b increased the incidence of apoptotic cells, and thiswas significantly reduced after coculture with IGFs (Fig. 5and Table 6). This demonstrated that the cytotoxic effects ofIL-1b were likely to be mediated at least in part by apoptosis,and that this could be limited by IGF-I or -II.
Experiments were repeated with islets isolated from rats
on days 4–6 of postnatal life. Exposure to cytokines alone atthe same concentrations as those stated above did not sig-nificantly alter either islet cell viability or DNA synthetic ratecompared with those in control incubations after 24 or 48 h(Tables 3 and 4). No further change in either parameter wasseen when incubations included either IGF-I or IGF-II. Ex-posure of neonatal islets to cytokines with or without IGFsdid not cause a significant change in insulin release com-pared with that in control incubations (Table 5).
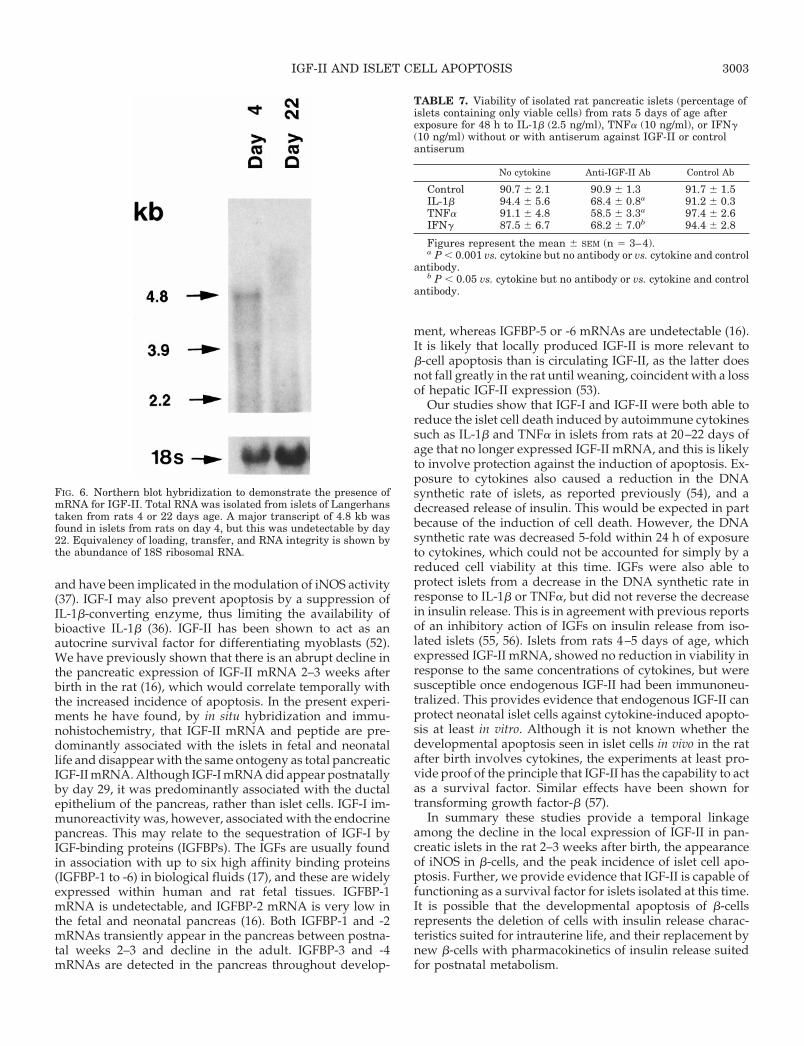
The resistance of islets isolated from neonatal rats tocytokine-induced cell death may be related to the relativelygreater expression of IGF-II seen in the neonatal rat pancreasthan at older ages. To test this hypothesis, we first establishedthat isolated islets from neonatal rats continued to expressIGF-II after incubation in vitro for 48 h. Total mRNA wasisolated from approximately 500 islets derived from ratseither 1–4 or 20–22 days old, and IGF-II mRNA was visu-alized by Northern blot hybridization. A major transcript ofIGF-II mRNA of 4.8 kb was seen for neonatal islets, whichwas no longer present at 20–22 days of age (Fig. 6). mRNAtranscripts for IGF-I were not abundant in islets isolated ateither age. The presence of endogenous IGF-II released fromneonatal islets was then functionally removed by incubationwith an IGF-II antiserum, and the viability of the islets wasassessed after exposure to cytokines. Islets were isolatedfrom neonatal rats at 5 days of age and incubated for 48 h inthe presence of IL-1b (2.5 ng/ml), TNFa (10 ng/ml), or IFNg(10 ng/ml), with or without antiserum against IGF-II or anonspecific antibody raised against human placental lacto-gen. Exposure to each cytokine alone did not alter islet cellviability (Table 7). A significant reduction in viability wasseen in response to IL-1b, TNFa, and IFNg after coincubationwith IGF-II antiserum, but not in the presence of the controlantiserum. These results suggest that an endogenous releaseof IGF-II contributed to the resistance of neonatal islets tocytokine-induced cell death in vitro.
Discussion
The fetal and neonatal endocrine pancreas in the rat has ahigh degree of plasticity, as b-cell mass increases in propor-tion to the rapid growth rate of the animal. However, onlyrecently has it been appreciated that substantial remodelingof the endocrine pancreas occurs in neonatal life, in whichb-cells are deleted by apoptosis, and b-cell mass is main-
TABLE 3. Viability of isolated rat pancreatic islets (percentage ofislets containing only viable cells) from rats 4–6 or 20–22 days ofage after exposure for 24 or 48 h to IL-1b (2.5 ng/ml), TNFa (10ng/ml), or IFNg (10 ng/ml) without or with IGF-I or IGF-II (100ng/ml)
No cytokine IL-1b TNFa IFNg
4–6 days24 h
Control 96 6 1 90 6 2 89 6 2 88 6 3IGF-I 94 6 2 89 6 1 88 6 2 83 6 2IGF-II 94 6 2 90 6 1 86 6 1 87 6 3
48 hControl 94 6 1 88 6 1 85 6 1 83 6 2IGF-I 95 6 1 89 6 1 92 6 2 89 6 2IGF-II 90 6 2 86 6 2 86 6 1 96 6 1
20–22 days24 h
Control 92 6 3 65 6 5a 47 6 4a 35 6 3a
IGF-I 94 6 2 82 6 7 66 6 3 44 6 5a
IGF-II 90 6 3 84 6 6 62 6 3 42 6 3a
48 hControl 89 6 2 38 6 2b 24 6 2b 11 6 4b
IGF-I 89 6 2 71 6 4c 51 6 3a,d 18 6 1b
IGF-II 87 6 1 62 6 2c 48 6 8a,d 14 6 3b
Figures represent the mean 6 SEM (n 5 3–4).a P , 0.05 vs. no cytokine in the absence of IGF-I or -II.b P , 0.001 vs. no cytokine in the absence of IGF-I or -II.c P , 0.05 vs. incubation with cytokine but without IGF-I or -II.d P , 0.01 vs. incubation with cytokine but without IGF-I or -II.
TABLE 4. DNA synthetic rate ([3H]thymidine incorporation,disintegrations per min/mg DNA) of pancreatic islets from rats 4–6or 20–22 days of age after exposure for 24 or 48 h to IL-1b (2.5ng/ml), TNFa (10 ng/ml), or IFNg (10 ng/ml) without or with IGF-Ior -II (100 ng/ml)
No cytokine IL-1b TNFa IFNg
4–6 days24 h
Control 3489 6 645 2712 6 483 3195 6 1236 3737 6 112IGF-I 4633 6 835 6288 6 449 4518 6 644 5018 6 213IGF-II 4636 6 669 5030 6 172 4232 6 348 4557 6 517
48 hControl 4349 6 736 3444 6 532 4093 6 1057 3436 6 347IGF-I 5863 6 417 6556 6 435 5743 6 511 4354 6 679IGF-II 5556 6 616 6398 6 1470 4310 6 134 5186 6 1126
20–22 days24 h
Control 1207 6 205 930 6 145 418 6 51a 296 6 99b
IGF-I 2105 6 139 1774 6 361 1454 6 102 324 6 21b
IGF-II 2404 6 584 1901 6 273 898 6 427 517 6 29b
48 hControl 1323 6 250 587 6 83b 420 6 69b 404 6 67b
IGF-I 1401 6 27 1359 6 127c 1335 6 35a,d 409 6 41b
IGF-II 1595 6 56 1539 6 64c 1298 6 29a,d 390 6 45b
Figures represent the mean 6 SEM (n 5 3–4).a P , 0.05 vs. no cytokine in the absence of IGF-I or -II.b P , 0.001 vs. no cytokine in the absence of IGF-I or -II.c P , 0.05 vs. incubation with cytokine but without IGF-I or -II.d P , 0.01 vs. incubation with cytokine but without IGF-I or -II.
TABLE 5. Release of insulin (microunits per ml) by isolated ratpancreatic islets from rats 4–6 or 20–22 days of age afterexposure for 48 h to IL-1b (2.5 ng/ml), TNFa (10 ng/ml), or IFNg(10 ng/ml) without or with IGF-I or -II (100 ng/ml)
No cytokine IL-1b TNFa IFNg
4–6 daysControl 220 6 22 215 6 36 198 6 13 232 6 18IGF-I 94 6 15 177 6 34 153 6 24 122 6 15IGF-II 151 6 55 247 6 37 212 6 39
20–22 daysControl 191 6 30 118 6 17a 79 6 5a 74 6 5a
IGF-I 110 6 10 83 6 12 71 6 7 57 6 1IGF-II 101 6 2b 83 6 4b 84 6 11
Figures represent the mean 6 SEM (n 5 3–4).a P , 0.05 vs. no cytokine in the absence of IGF-I or -II.b P , 0.05 vs. incubation with cytokine but without IGF-I or -II.
IGF-II AND ISLET CELL APOPTOSIS 3001

tained predominantly by the generation of new islets fromthe ductal epithelium. The process of islet cell apoptosis wasfirst predicted on mathematical grounds (5) and was recentlyconfirmed by experimental observation (13). A similar phe-nomenon is seen in the rat postpartum (46), where the largeincrease in pancreatic b-cells that occurs during pregnancyis reduced by b-cell apoptosis postpartum. Clearly, the en-docrine pancreas has a much greater capacity for functionalplasticity than was previously thought, at least in rodents.Our studies confirm those of Scaglia et al. (13) in showing apeak of cell apoptosis in the rat islets of Langerhans 13–17days after birth. We were unable to positively identify theapoptotic cells as once being b-cells because they could notbe shown to contain immunoreactive insulin. This may wellhave been destroyed before the terminal events in apoptosis,DNA breakage, which was the detection criterium used here.The central location of most apoptotic cells within the isletsis consistent with them having once been functional b-cells.Apoptotic cells are rapidly removed from tissues in vivo (47).Our findings that by 14 days after birth over 13% of islet cellswere undergoing apoptosis may be an underestimate. As theincidence of apoptosis increases steadily from postnatal day
6 and begins to fall only after day 14, at least a third of theislet cells may be lost in this way in neonatal life.
NO production, driven by increased expression of iNOS,has been implicated in the autoimmune destruction inducedby cytokines such as IL-1b (14, 48). Other intracellular mech-anisms may also exist that lead to b-cell apoptosis, includingthe expression of fas (49) and the generation of oxygen freeradicals (50). In an attempt to identify possible intracellularmechanisms responsible for the induction of developmentalapoptosis in islets, we examined the distribution of iNOS.Immunohistochemistry showed the presence of iNOS in pan-creatic islets, with a mean of 10% of cells having immunopo-sitive cytoplasm on postnatal day 12. The peak incidence ofiNOS presence in islet cells was 2 days before the maximalincidence of apoptosis, suggesting a possible mechanisticrelationship. Dual staining for insulin and iNOS showed thatalmost all iNOS-positive cells were b-cells, located within thecentral areas of the islets. The appearance of iNOS and theoccurrence of apoptosis were present in most islets 12–14days after birth. The events that lead to an increased distri-bution of iNOS and increased incidence of apoptosis at thistime are not known. However, it is at this time in the cor-responding ontogeny of the mouse pancreas that infiltrationof the islets with T cells has been shown to occur (51). It ispossible that colonization of the islets with monocytes, mac-rophages, or T cells in early postnatal life may lead to risesin the local concentrations of cytokines such as IL-1b, leadingto a controlled b-cell deletion by apoptosis. However, it isalso possible that these events require the removal of a sur-vival factor for b-cells, and this may be the local availabilityof IGF-II.
Exogenous IGFs have been demonstrated to prevent in-duced apoptosis in a variety of differing cell types (32–35)
FIG. 5. Demonstration of endocrinecell apoptosis (arrows) using molecularhistochemistry in islets isolated fromrats 20–22 days age in control medium(A), after incubation with IL-1b (2.5 ng/ml; B), or after incubation with IL-1band IGF-I (100 ng/ml; C).
TABLE 6. Incidence of islet cells containing apoptotic nuclei(percentage) in isolated rat pancreatic islets from rats 20–22 daysof age after incubation for 48 h with IL-1b (2.5 ng/ml) without orwith IGF-I or -II (100 ng/ml)
No cytokine IL-1b
Control 1.1 6 1.0 18.6 6 3.5a
IGF-I 2.3 6 1.5 5.1 6 1.6b
IGF-II 2.2 6 2.0 4.5 6 2.2b
Figures represent the mean 6 SEM (n 5 3–4).a P , 0.01 vs. no cytokine in the absence of IGF-I or -II.b P , 0.05 vs. incubation with cytokine but without IGF-I or -II.
3002 IGF-II AND ISLET CELL APOPTOSIS Endo • 1998Vol 139 • No 6
and have been implicated in the modulation of iNOS activity(37). IGF-I may also prevent apoptosis by a suppression ofIL-1b-converting enzyme, thus limiting the availability ofbioactive IL-1b (36). IGF-II has been shown to act as anautocrine survival factor for differentiating myoblasts (52).We have previously shown that there is an abrupt decline inthe pancreatic expression of IGF-II mRNA 2–3 weeks afterbirth in the rat (16), which would correlate temporally withthe increased incidence of apoptosis. In the present experi-ments he have found, by in situ hybridization and immu-nohistochemistry, that IGF-II mRNA and peptide are pre-dominantly associated with the islets in fetal and neonatallife and disappear with the same ontogeny as total pancreaticIGF-II mRNA. Although IGF-I mRNA did appear postnatallyby day 29, it was predominantly associated with the ductalepithelium of the pancreas, rather than islet cells. IGF-I im-munoreactivity was, however, associated with the endocrinepancreas. This may relate to the sequestration of IGF-I byIGF-binding proteins (IGFBPs). The IGFs are usually foundin association with up to six high affinity binding proteins(IGFBP-1 to -6) in biological fluids (17), and these are widelyexpressed within human and rat fetal tissues. IGFBP-1mRNA is undetectable, and IGFBP-2 mRNA is very low inthe fetal and neonatal pancreas (16). Both IGFBP-1 and -2mRNAs transiently appear in the pancreas between postna-tal weeks 2–3 and decline in the adult. IGFBP-3 and -4mRNAs are detected in the pancreas throughout develop-
ment, whereas IGFBP-5 or -6 mRNAs are undetectable (16).It is likely that locally produced IGF-II is more relevant tob-cell apoptosis than is circulating IGF-II, as the latter doesnot fall greatly in the rat until weaning, coincident with a lossof hepatic IGF-II expression (53).
Our studies show that IGF-I and IGF-II were both able toreduce the islet cell death induced by autoimmune cytokinessuch as IL-1b and TNFa in islets from rats at 20–22 days ofage that no longer expressed IGF-II mRNA, and this is likelyto involve protection against the induction of apoptosis. Ex-posure to cytokines also caused a reduction in the DNAsynthetic rate of islets, as reported previously (54), and adecreased release of insulin. This would be expected in partbecause of the induction of cell death. However, the DNAsynthetic rate was decreased 5-fold within 24 h of exposureto cytokines, which could not be accounted for simply by areduced cell viability at this time. IGFs were also able toprotect islets from a decrease in the DNA synthetic rate inresponse to IL-1b or TNFa, but did not reverse the decreasein insulin release. This is in agreement with previous reportsof an inhibitory action of IGFs on insulin release from iso-lated islets (55, 56). Islets from rats 4–5 days of age, whichexpressed IGF-II mRNA, showed no reduction in viability inresponse to the same concentrations of cytokines, but weresusceptible once endogenous IGF-II had been immunoneu-tralized. This provides evidence that endogenous IGF-II canprotect neonatal islet cells against cytokine-induced apopto-sis at least in vitro. Although it is not known whether thedevelopmental apoptosis seen in islet cells in vivo in the ratafter birth involves cytokines, the experiments at least pro-vide proof of the principle that IGF-II has the capability to actas a survival factor. Similar effects have been shown fortransforming growth factor-b (57).
In summary these studies provide a temporal linkageamong the decline in the local expression of IGF-II in pan-creatic islets in the rat 2–3 weeks after birth, the appearanceof iNOS in b-cells, and the peak incidence of islet cell apo-ptosis. Further, we provide evidence that IGF-II is capable offunctioning as a survival factor for islets isolated at this time.It is possible that the developmental apoptosis of b-cellsrepresents the deletion of cells with insulin release charac-teristics suited for intrauterine life, and their replacement bynew b-cells with pharmacokinetics of insulin release suitedfor postnatal metabolism.
FIG. 6. Northern blot hybridization to demonstrate the presence ofmRNA for IGF-II. Total RNA was isolated from islets of Langerhanstaken from rats 4 or 22 days age. A major transcript of 4.8 kb wasfound in islets from rats on day 4, but this was undetectable by day22. Equivalency of loading, transfer, and RNA integrity is shown bythe abundance of 18S ribosomal RNA.
TABLE 7. Viability of isolated rat pancreatic islets (percentage ofislets containing only viable cells) from rats 5 days of age afterexposure for 48 h to IL-1b (2.5 ng/ml), TNFa (10 ng/ml), or IFNg(10 ng/ml) without or with antiserum against IGF-II or controlantiserum
No cytokine Anti-IGF-II Ab Control Ab
Control 90.7 6 2.1 90.9 6 1.3 91.7 6 1.5IL-1b 94.4 6 5.6 68.4 6 0.8a 91.2 6 0.3TNFa 91.1 6 4.8 58.5 6 3.3a 97.4 6 2.6IFNg 87.5 6 6.7 68.2 6 7.0b 94.4 6 2.8
Figures represent the mean 6 SEM (n 5 3–4).a P , 0.001 vs. cytokine but no antibody or vs. cytokine and control
antibody.b P , 0.05 vs. cytokine but no antibody or vs. cytokine and control
antibody.
IGF-II AND ISLET CELL APOPTOSIS 3003

Acknowledgments
We are grateful to Dr. Joanna Hogg for assistance with in situ hy-bridization, and to Dr. Dianne Finegood for intellectual input.
References
1. Hill DJ, Hogg J 1991 Growth factor control of pancreatic b cell hyperplasia.In: Herington A (ed) Clinical Endocrinology and Metabolism. Bailliere Tindall,London, pp 689–698
2. Kaung HL 1994 Growth dynamics of pancreatic islet cell populations duringfetal and neonatal development of the rat. Dev Dyn 200:163–175
3. Fernandes A, King LC, Guz Y, Stein R, Wright CVE, Teitelman G 1997Differentiation of new insulin-producing cells is induced by injury to adultpancreatic islets. Endocrinology 138:1750–1762
4. Dudek RW, Lawrence IE 1988 Morphologic evidence of interaction betweenadult ductal epithelium of pancreas and fetal foregut mesenchyme. Diabetes37:891–900
5. Finegood DT, Scaglia L, Bonner-Weir S 1995 Dynamics of b cell mass in thegrowing rat pancreas. Diabetes 44:249–256
6. Cantenys D, Portha B, Dutrillaux MC, Hollande E, Roze C, Picon L 1981Histogenesis of the endocrine pancreas in newborn rats after destruction bystreptozotocin. Virchows Arch Cell Pathol 35:109–122
7. Bencosme SA 1955 Cytology of the islet cells in alloxan diabetic rabbits. Am JPathol 31:1149–1164
8. Brockenbrough JS, Weir GC, Bonner-Weir S 1988 Discordance of exocrineand endocrine growth after 90% pancreatectomy in rats. Diabetes 37:232–236
9. Gu D, Sarvetnick N 1993 Epithelial cell proliferation and islet neogenesis inIFN-g transgenic mice. Development 118:33–46
10. Cecil RL 1911 On hypertrophy and regeneration of the islands of Langerhans.J Exp Med 14:500–519
11. Gepts W, de Mey J 1978 Islet cell survival determined by morphology. Animmunocytochemical study of the islet of Langerhans in juvenile diabetesmellitus. Diabetes [Suppl 1] 27:251–261
12. Volk BW, Wellman KF 1985 The pancreas in idiopathic diabetes. In: Volk BW,Arquilla ER (eds) The Diabetic Pancreas, ed 2. Plenum Press, New York, p 353
13. Scaglia L, Cahill CJ, Finegood DT, Bonner-Weir S 1997 Apoptosis participatesin the remodeling of the endocrine pancreas in the neonatal rat. Endocrinology138:1736–1741
14. Tornehave D, Larsson L-I 1997 Presence of Bcl-Xl during development of thehuman fetal and rat neonatal endocrine pancreas: correlation to programmedcell death. Exp Clin Endocrinol Diabetes 105:A27
15. Corbett JA, McDaniel ML 1995 Intraislet release of interleukin-1 inhibits b cellfunction by inducing b cell expression of inducible nitric oxide synthase. J ExpMed 181:559–568
16. Hogg J, Hill DJ, Han VKM 1994 The ontogeny of insulin-like growth factor(IGF) and IGF binding protein gene expression in the rat pancreas. J MolEndocrinol 13:49–58
17. Calvo EL, Bernatchez G, Pelletier G, Iovanna JL, Morisset J 1997 Down-regulation of IGF-I mRNA expression during postnatal pancreatic develop-ment and overexpression after subtotal pancreatectomy and acute pancreatitisin the rat pancreas. J Mol Endocrinol 18:233–242
18. Hill DJ, Hogg J 1992 Expression of insulin-like growth factors (IGFs) and theirbinding proteins (IGFBPs) during pancreatic development in rat, and modulationof IGF actions on rat islet DNA synthesis by IGF BPs. In: Vinik A (ed) PancreaticIslet Cell Regeneration and Growth. Plenum Press, New York, pp 113–120
19. Hill DJ, Clemmons DR 1992 Similar distribution of insulin-like growth factorbinding proteins-1, -2 and -3 in human fetal tissues. Growth Factors 6:315–326
20. Hill DJ, Frazer A, Swenne I, Wirdnam PK, Milner RDG 1987 Somatomedin-Cin the human fetal pancreas: cellular localization and release during organculture. Diabetes 36:465–471
21. Hogg J, Han VKM, Clemmons DR, Hill DJ 1993 Interactions of glucose, insulin-like growth factors (IGFs) and IGF binding proteins in the regulation of DNAsynthesis by isolated fetal rat islets of Langerhans. J Endocrinol 138:401–412
22. Swenne I, Hill DJ, Strain AJ, Milner RDG 1987 Growth hormone regulationof somatomedin-C/insulin-like growth factor I production and DNA repli-cation in fetal rat islets in tissue culture. Diabetes 36:288–294
23. Scharfmann R, Corvol M, Czernichow P 1989 Characterization of IGF I pro-duced by fetal rat pancreatic cells, Diabetes 38:686–690
24. Romanus JA, Rabinovitch A, Rechler MM 1985 Neonatal rat islet cell culturessynthesize insulin-like growth factor I. Diabetes 34:696–702
25. Rabinovitch A, Quigley C, Russel T, Patel Y, Mintz DH 1982 Insulin andmultiplication stimulating activity monolayer cultures. Diabetes 31:160–164
26. Van Schravendijk CF, Foriers A, Van Den Brande JL, Pipeleers DG 1987Evidence for the presence of type I insulin-like growth factor receptors on ratpancreatic A and B cells, Endocrinology 121:1784–1788
27. Fehmann HC, Jehle P, Markus U, Goke B 1996 Functional receptors forinsulin-like growth factors-I (IGF-I) and IGF-II on insulin-, glucagon-, andsomatostatin-producing cells. Metabolism 45:759–766
28. Swenne I, Hill DJ 1989 Growth hormone regulation of DNA replication, butnot insulin production is mediated by somatomedin-C/insulin-like growthfactor I in isolated pancreatic islets from adult rats. Diabetologia 32:191–197
29. Hoog A, Grimelius L, Falkmer S, Sara VR 1993 A high molecular weight IGF-2immunoreactive peptide (pro-IGF-2?) in the insulin cells of the islets of Lang-erhans in pancreas of man and rat. Regul Pept 47:275–283
30. Maake C, Reinecke M 1993 Immunohistochemical localization of insulin-likegrowth factor 1 and 2 in the endocrine pancreas of rat, dog, and man, and theircoexistance with classical islet hormones. Cell Tissue Res 273:249–259
31. Asfari M, Wei D, Noel M, Holthuizen PE, Czernichow P 1995 IGF-II geneexpression in a rat insulin-producing b-cell line (INS-1) is regulated by glucose.Diabetologia 38:927–935
32. Geier A, Haimshon M, Beery R, Lunenfeld B 1992 Insulin-like growth factor-Iinhibits cell death induced by cycloheximide in MCF-7 cells - a model systemfor analyzing control of cell death. In Vitro Cell Dev Biol Anim 28A:725–729
33. Galli C, Meucci O, Scorziello A, Werge TM, Calissano P, Schettini G 1995Apoptosis in cerebellar granule cells is blocked by high KCl, forskolin, andIGF-I through distinct mechanisms of action: the involvement of intracellularcalcium and RNA synthesis. J Neurosci 15:1172–1179
34. Chun SY, Billig H, Tilly JL, Furuta I, Tsafriri A, Hsueh AJW 1994 Gonad-otropin suppression of apoptosis in cultured preovulatory follicles: mediatoryrole of endogenous insulin-like growth factor I. Endocrinology 135:1845–1853
35. Muta K, Krantz SB 1993 Apoptosis of human erythroid colony-forming cellsis decreased by stem cell factor and insulin-like growth factor I as well aserythropooetin. J Cell Physiol 156:264–271
36. Jung Y, Miura M, Yuan J 1996 Suppression of IL-1b converting enzyme-mediated cell death by insulin-like growth factor. J Biol Chem 271:5112–5117
37. Guan J, Williams C, Gunning M, Mallard C, Gluckman P 1993 The effectsof IGF-I treatment after hypoxic-ischaemic brain injury in adult rats. J CerebrBlood Flow Metab 13:609–616
38. Hellerstrom C. Lewis NJ, Borg H, Johnson R, Freinkel N 1979 Method forlarge scale isolation of pancreatic islets by tissue culture of fetal rat pancreas.Diabetes 28:769–776
39. Hales CN, Randle PJ 1963 Immunoassay of insulin with insulin antibodyprecipitate. Biochem J 88:137–146
40. Herbert V, Lau K, Gottlieb CW, Bleicher SJ 1965 Coated charcoal immuno-assay of insulin. J Clin Endocrinol 25:1375–1384
41. Schnuerer EM, Rokaeus Å, Carlquist M, Bergman T, Dupre J, McDonald TJ1990 Rat and porcine galanin are equipotent in inhibiting insulin responses toglucose in the anesthetized rat. Pancreas 5:70–74
42. Hsu SM, Raine L, Fanger H 1981 Use of avidin-biotin peroxidase complex(ABC) in immunoperoxidase techniques: a comparison between ABC andunlabelled antibody (PAP) procedures. J Histochem Cytochem 29:577–580
43. Wijsman JH, Jonker RR, Keijzer R, Van de Velde CJ, Cornelisse CJ, VanDierendonck JH 1993 A new method to detect apoptosis in paraffin sections:in situ end labelling of fragmented DNA. J Histochem Cytochem 41:7–12
44. Southern EM 1975 Detection of specific sequences among DNA fragmentsseparated by gel electrophoresis. J Mol Biol 98:503–507
45. Murphy LJ, Tachibana K, Friesen HG 1988 Stimulation of hepatic insulin-likegrowth factor-I gene expression by ovine prolactin: evidence for intrinsicsomatogenic activity in the rat. Endocrinology 122:2027–2033
46. Scaglia L, Smith FE, Bonner-Weir S 1995 Apoptosis contributes to the invo-lution of b cell mass in the postpartum rat pancreas. Endocrinology136:5461–5468
47. Bursch W, Kleine L, Tenniswood M 1990 Biochemistry of cell death byapoptosis. Biochem Cell Biol 68:1071–1074
48. Kenato H, Fujii J, Seo HG, Suzuki K, Matsuoka T, Nakamura M, TatsumiH, Yamasaki Y, Kamada T, Taniguchi N 1995 Apoptotic cell death triggeredby nitric oxide in pancreatic b-cells. Diabetes 44:733–738
49. Yamada K, Takane-Gyotuku N, Yuan X, Ichikawa F, Inada C, Nonaka K 1996Mouse islet cell lysis mediated by interleukin-1-induced Fas. Diabetologia39:1306–1312
50. Rabinovitch A, Suarez WL, Thomas PD, Strynadka K, Simpson I 1992 Cy-totoxic effects of cytokines on rat islets: evidence for involvement of freeradicals and lipid peroxidation. Diabetologia 35:409–413
51. Yang Y, Charlton, B, Shimada A, dal Canto RD, Fathman G 1996 MonoclonalT cells identified in early NOD islet infiltrates. Immunity 4:189–194
52. Stewart CE, Rotwein P 1996 Insulin-like growth factor-II is an autocrinesurvival factor for differentiating myoblasts. J Biol Chem 271:11330–11338
53. Brown AL, Graham DE, Nissley SP, Hill DJ, Strain AJ, Rechler MM 1986Developmental regulation of insulin-like growth factor II mRNA in differentrat tissues. J Biol Chem 261:13144–13150
54. Mandrup-Poulsen T, Bendtzen K, Nerup J, Dinarello CA, Svenson M,Nielsen JH 1986 Affinity purified human interleukin 1 is cytotoxic to islet cellsof Langerhans. Diabetologia 29:63–67
55. Van Schravendijk CFH, Heylen L, Van Den Brande JL, Pipeleers DG 1990Direct effect of insulin and insulin-like growth factor-I on the secretory activityof rat pancreatic beta cells. Diabetologia 33:649–653
56. Hill DJ, Sedran RJ, Brenner SL, McDonald TJ 1997 Insulin-like growthfactor-I (IGF-I) has a dual effect on insulin release from isolated, perifusedadult rat islets of Langerhans. J Endocrinol 153:15–25
57. Cunningham JM, Mabley JG, Mwebe B, Kovani E, Green IC 1994 Suppres-sion of interleukin-1b-induced nitric oxide synthase activity by transforminggrowth factor-b in the insulin-secreting cell line RINm5F. J Endocrinol [Suppl]140:37
3004 IGF-II AND ISLET CELL APOPTOSIS Endo • 1998Vol 139 • No 6
Copyright © 2022 FDOKUMEN