
Identifying and structurally characterizing CD1b in Aotus nancymaae owl monkeys
Upload
independentCategory
view
0download
0
PRIMATES, 33(2): 239-245, April 1992 239
Aotus vociferans x Aotus nancymai: Sympatry Without Chromosomal Hybridation
JULIO CESAR PIECZARKA, REGINA MARIA DE SOUZA BARROS, CLEUSA YOSHIKO NAGAMACH1
Universidade Federal do Pard
RAUL RODRIGUES, and ALVARO ESPINEL Universidad Nacional de Col6mbia
ABSTRACT. In a study relating geographic distribution of Aotus populations with karyotype and coat pattern, HERSHKOVlTZ (1983) suggested that this genus comprises nine species. A. vociferans (2n = 46) is distributed along the left bank of the Maranon river, and A. nancymai (2n = 54) along the right bank of the same river. In the present study we analyzed the karyotypes of 11 A. vociferans and 11 A. nancymai specimens using G- and C-banding and NOR-staining. Of these animals, five A. vociferans were sympatric with five A. nancymai in Isla Aramaza, Colombia; one A. vociferans was sympatric with three A. nancymai in Yahuma, Peru; one A. vociferans was sympatric with three A. nancymai in Terezinha I, Brazil. No signs of hybridation between the taxa was observed at any time, with all A. vociferans having 2n = 46 and all A. nancymai 2n = 54, as previously reported in the litera- ture. These data suggest that A. vociferans and A. nancymai should indeed be distinct species.
Key Words: Aotus; Cytogenetic; Sympatry; Taxonomy; Cebidae.
I N T R O D U C T I O N
Cytogenetic studies have shown that the genus Aotus has many different karyotypes (BRUMBACK, 1974; DE BOER, 1974; KOIFFMANN &; SALDANHA, 1974; MA et al., 1976; MUDRY DE PARGAMENT et al., 1984; MA et al., 1985; GIRALDO et al., 1986; PIECZARKA & NAGAMACHI, 1988). The diploid number ranges from 46 chromosomes (MA et al., 1976) to 58 (GIRALDO et al., 1986). On the basis of coat characteristics, differences in karyotype and geographic distribution, HERSHKOVITZ (1983) classified Aotus in nine species. The Aotus animals living along the left bank of the Maranon-Solimoes-Amazonas river have a gray neck and have been organized into four species: A. trivirgatus, A. vociferans, A. brumbacki, and A. lemurinus (A. l. lemurinus and A. 1. griseimembra). The animals living on the right bank have a red neck and have been organized into five species: A. infulatus, A. nigriceps, A. nancymai, A. miconax, and A. azarae (,4. a. azarae and A. a. bolivensis). Two populations having the same karyotype but different coat patterns are considered to be subspecies. I f a population has a different coat pattern with the other population but its karyotype is still unknown, it is considered to be a distinct species. After the classifica- tion of HERSHKOVITZ (1983) was published, PIECZARKA and NAGAMACHI (1988) described the karyotype of A. infulatus and did not obtain sufficient evidence to consider this taxon as a distinct species.
Despite the strong correlation between coat pattern, karyotype, and geographic distribu- tion, more concrete evidence is needed to validate or invalidate this classification. An in- teresting approach is to try to find regions of contact between populations of different taxa,

240 J.C. PIECZARKA et al.
Fig. 1. G-banding pattern of A. vociferans. 2n =46.
and to analyze the animals living in these regions in order to determine the possible occur- rence of hybridation. A. vociferans, which lives along the left bank of the Maranon river, and A. nancymai, which lives on the right bank, have considerably differentiated karyo- types, thus representing material that could provide significant information in this respect, specially if signs of hybridation are investigated by studying the karyotypes of sympatric specimens.
In the present study we analyzed the karyotypes of Aotus vociferans and A. nancymai specimens from regions in which sympatry occurs, near Leticia, Colombia. Cytotaxonomic considerations are also presented.
MATERIAL AND METHODS
Twenty-two specimens of the genus Aotus were investigated. Of these, 11 (7 males and 4 females) were considered to belong to the species A. vociferans, and 11 (4 males and 7 females) to the species A. nancymai according to the phenetic key for Aotus species and subspecies proposed by HERSHKOVITZ (1983). All animals were collected in the vicinity of Leticia, Colombia (70~ 4~ Five A. vociferans were sympatric with five

Sympatry Between A. vociferans and A. nancymai 241
Fig. 2. C-banding pattern of A. vociferans. 2n=46.
A. nancymai in Isla Aramaza, Colombia; one A. vociferans was sympatric with three A. nancymai in Yahuma, Peru; one A. vociferans was sympatric with three A. nancymai in Terezinha I, Brazil. The other animals were from nearby sites and did not present sympatry.
Blood (3 ml) was collected from each animal by femoral puncture using heparinized syringes. Chromosomes were obtained by the culture method of MOORHEAD et al. (1960). G-, C-banding, and NOR-staining were obtained using the techniques described by SCHERES (1972), SUMNER (1972), and HOWELL and BLACK (1980). Karyotypes were ar- ranged according to MA et al. (1976).
RESULTS AND DISCUSSION
All A. vociferans specimens had 2n=46. Thirteen autosome pairs are bi-arm and the other nine pairs are acrocentric. Chromosome X is a submetacentric similar to the X detect- ed in most New World monkeys. Chromosome Y is very small and acrocentric. Figure 1 shows the G-banding pattern of this species. The C-banding pattern of A. vociferans is presented in Figure 2.
The karyotype of A. nancymai had 2n = 54, with 14 hi-arm autosomic pairs and 12 acrocentric autosome pairs. Chromosome X is similar to that of A. vociferans. The Y chro- mosome is acrocentric. MA et al. (1976) described it as a metacentric. Variation in mor- phology of the Y chromosome is a common feature in New World monkeys (NAGAMACHI
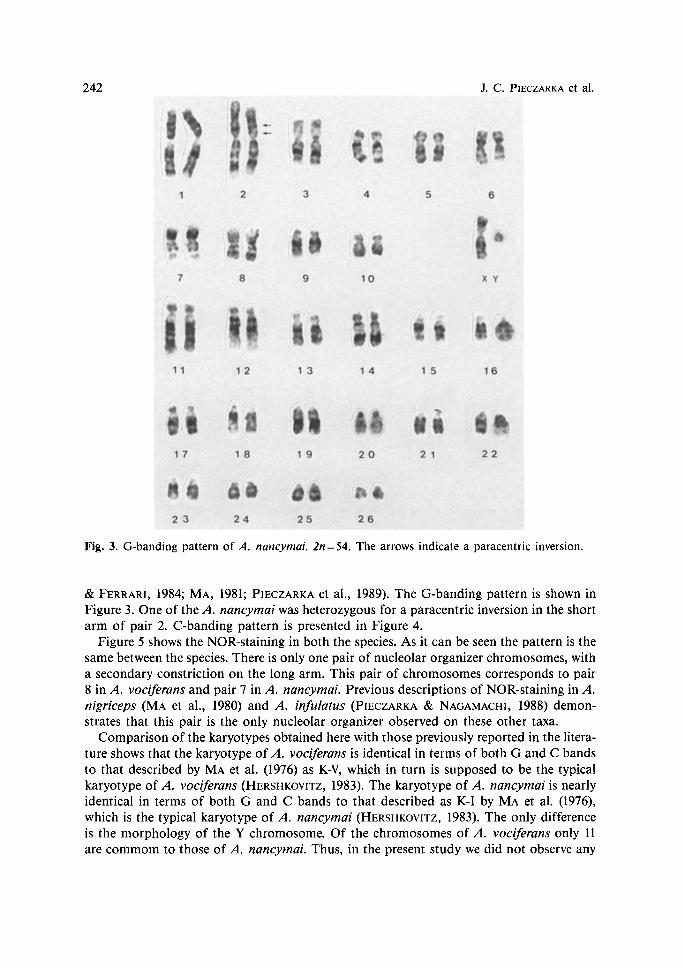
242 J.C. PIECZARKA et al.
Fig. 3. G-banding pattern of A. nancymai. 2n = 54. The arrows indicate a paracentric inversion.
& FERRARI, 1984; MA, 1981; PIECZARKA et al., 1989). The G-banding pattern is shown in Figure 3. One of the A. nancymai was heterozygous for a paracentric inversion in the short arm of pair 2. C-banding pattern is presented in Figure 4.
Figure 5 shows the NOR-staining in both the species. As it can be seen the pattern is the same between the species. There is only one pair of nucleolar organizer chromosomes, with a secondary constriction on the long arm. This pair of chromosomes corresponds to pair 8 in A. vociferans and pair 7 in A. nancymai. Previous descriptions of NOR-staining in A. nigriceps (MA et al., 1980) and A. infulatus (PIECZARKA ~; NAGAMACHI, 1988) demon- strates that this pair is the only nucleolar organizer observed on these other taxa.
Comparison of the karyotypes obtained here with those previously reported in the litera- ture shows that the karyotype of A. vociferans is identical in terms of both G and C bands to that described by MA et al. 0976) as K-V, which in turn is supposed to be the typical karyotype of A. vociferans (HERSHrOVITZ, 1983). The karyotype of A. nancymai is nearly identical in terms of both G and C bands to that described as K-I by MA et al. (1976), which is the typical karyotype of A. nancymai (HERSHKOVITZ, 1983). The only difference is the morphology of the Y chromosome. Of the chromosomes of A. vociferans only 11 are commom to those of A. nancymai. Thus, in the present study we did not observe any

Sympatry Between A. vociferans and A. nancymai 243
Fig. 4. C-banding pattern of A. nancymak 2n=54.
typical A. vociferans chromosome in A. nancymai or vice versa, showing the absence of any sign of hybridation between these two taxa.
HERSHKOVlTZ (1983) emphasized the need to look for situations of sympatry between different taxa since the subdivision of Aotus genus into species based primarily on karyo- types may seem tenuous. Our sample is particularly interesting from this viewpoint because it consists of a taxon representing each Aotus group, with marked karyotypic differences, located at a site were the meanders of the Maranon river permit passive specimen dispersal from one side of the river to the other. Despite the old rule which says that " the absence of proof does not prove the absence of a phenomenon," the chromosomal differences be- tween the taxa, A. vociferans and A. nancymai, clearly indicate that there is very high the chance that the hybridation did not occur. Evidence of reduced fertility has been reported for Aotus pairs with different karyotypes held in captivity (ELLIOT et al., 1976; CIOMANEC & CAMPBELL, 1977; SIMPSON & JONES, 1982). These laboratory results, taken together with our field observations support the subdivision of the genus Aotus into more than one species.
Acknowledgments. We thank Profs. HORACIO SCHNEIDER and MARIA IRACILDA SAMPAIO for collect- ing the material, and Ms. MARIA DA GLORIA SANTOS and Mr. FRANCISCO MENDES BORGES for techni- cal assistance. This research was supported by FINEE CNPq, PIG, and ELETRONORTE.

244 J .C . PIECZARKA et al.
Fig. 5. NOR-staining on a. Aotus vociferans and b. A. nancymai.
R E F E R E N C E S
BRUMBACK, R. A., 1974. A third species of the owl monkey (Aotus). J. Hered., 65:321 -323. CIOMANEC, J. C. • A. K. CAMPBELL, 1977. Breeding the owl monkey (Aotus trivirgatus) in a labora-
tory environment. Lab. Anita. Sci., 27: 512-517. DE BOER, L. E. M., 1974. Cytotaxonomy of the Platyrrhini (Primates). Genen. Phaenenm., 17"
1-115. ELLIOT, M. E., P. H. SEHGAL, ~s L. V. CHALIFOUX, 1976. Management and breeding of Aotus trivir-
gatus. Lab. Anirn. Sci., 26: 1037-1040. GIRALDO, A., M. L. BUENO, E. SILVA, J. RAMIREZ, J. UMANA, & C. ESPINAL, 1986. Estudio
Citogenetico de 288 Aotus Colombianos. Biomedica, 6 (1-2) : 5 - 1 3 . HERSHKOVITZ, P., 1983. Two new species of night monkeys, genus Aotus (Cebidae, Platyrrhini): a
preliminary report on Aotus taxonomy. Amer. J. Primatol., 4: 209-243.

Sympatry Between A. vociferans and A. nancymai 245
HOWELL, W. M. & D. A. BLACK, 1980. Controlled silver-staining of nucleolar organizer regions with protective colloidal developer: a 1-step method. Experientia, 36: 1014.
KOIFFMANN, C. P. & P. H. SALDANHA, 1974. Cytogenetic of brazilian monkeys. J. Human Evol., 3: 275 - 282.
MA, N. S. E, 1981. Chromosome evolution in the owl monkey, Aotus. Amer. J. Phys. Anthropol., 54: 293- 303.
- - , R. AQUINO, & W. E. COLLINS, 1985. Two new karyotypes in the Peruvian Owl Monkey (Aotus trivirgatus). Amer. J. Primatol., 9" 333- 341.
- - , T. C. JONES, A. MILLER, L. M. MORGAN, & E. ADAMS, 1976. Chromosome polymorphism and banding patterns in the owl monkey (Aotus). Lab. Anim. Sci., 26 (6): 1022-1036.
- - , B. M. RENQUIST, R. HALL, P. K. SEHGAL, Y. SIMEONE, & T. C. JONES, 1980. X X / " X O " sex determination system in a population of Peruvian owl monkey, Aotus. J. Hered., 71: 336- 342.
MOORHEAD, D. S., P. C. NOWELL, W. J. MELLMAN, D. M. BATTIPS, & D. A. HUNGERFORD, 1960. Chromosomes preparations of leukocytes cultured from human peripheral blood. Exp. Cell Res., 20:613 - 615.
MUDRV DE PAR~AMENT, M. D., O. J. COLLILAS, & S. B. DE SALUM, 1984. The Aotus from Northern Argentina. Primates, 25: 530-537.
NAGAMACHI, C. Y. & I. FERRARI, 1984. Cytogenetic studies of Callithrix jacchus (Callithricidae, Platyrrhini) from two different sites in Brazil. I. Morphologic variability of chromosome Y. Brazil J. Genet., 7 (3): 497-507.
PIECZARKA, J. C. & C. Y. NAGAMACH|, 1988. Cytogenetic Studies of Aotus from Eastern Amazonia: Y/Autosome Rearrangement. Amer. J. Primatol., 14: 255- 263.
, - - , & R. M. S. BARROS, 1989. The karyotype of Ateles paniscus paniscus (Cebidae, Primates): 2n=32. Brazil J. Genet., 12 (3): 543-551.
SCHERES, M. J. C., 1972. Identification of two Robertsonian translocations with a Giemsa banding technique. Humangenetik, 15: 253 - 256.
SIMPSON, J. S. & A. C. JONES, 1982. Hybrid production in owl monkeys (Aotus trivirgatus). Lab. Anim., 16: 71-72.
SUMNER, A. T., 1972. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res., 75: 304-306.
- - Received October 2, 1991; Accepted December 16, 1991
Authors' Names and Addresses: JuLlO CESAR PIECZARKA, REGINA MARIA DE SOUZA BARROS, and CLEUSA YOSHIKO NAGAMACH~, Departamento de Gendtica, Centro de Ci~ncias Biol6gicas, Universidade Federal do Pard, Beldm, Pard, Brasil; RAUL RODRIGUES and ALVARO ESPINEL, Universidad Nacional de Coldmbia, Bogotd, ColOmbia.
Copyright © 2022 FDOKUMEN




















