
in the matter of eric h. bornstein an attorney at law - DRB ...
Upload
independentCategory
view
1download
0
Immunogenetics (2000) 51 :219–230 Q Springer-Verlag 2000
ORIGINAL PAPER
J. Javier Nino-Vasquez 7 Denise VogelRaul Rodriguez 7 Alberto MorenoManuel Elkin Patarroyo 7 Gerd PluschkeClaudia A. Daubenberger
Sequence and diversity of DRB genes of Aotus nancymaae,
a primate model for human malaria parasites
Received: 10 August 1999 / Revised: 11 October 1999
The nucleotide sequence data reported in this paper have beensubmitted to the EMBL/GenBank nucleotide sequencedatabases and have been assigned the accession numbersAF129792 to AF129808, AF132755 to AF132770, and AF169485to AF169487
J. Javier Nino-Vasquez (Y) 7 Raul RodriguezAlberto MorenoManuel Elkin PatarroyoInstituto de Inmunologia, Hospital San Juan de Dios,Universidad Nacional de Colombia, Avenida 1 No 10-01,Santafe de Bogota, D.C., Colombiae-mail: javinino6hotmail.comTel.: c57-133-71154Fax: c57-128-03999
Denise Vogel 7 Gerd Pluschke 7 Claudia A. DaubenbergerSwiss Tropicial Institute, Socinstrasse 57, CH-4002 Basel,Switzerland
Abstract The New World primate Aotus nancymaae issusceptible to infection with the human malaria para-site Plasmodium falciparum and Plasmodium vivax andhas therefore been recommended by the World HealthOrganization as a model for evaluation of malaria vac-cine candidates. We present here a first step in the mo-lecular characterization of the major histocompatibilitycomplex (MHC) class II DRB genes of Aotus nancy-maae (owl monkey or night monkey) by nucleotide se-quence analysis of the polymorphic exon 2 segments. Ina group of 15 nonrelated animals captivated in the wild,34 MHC DRB alleles could be identified. Six allelic li-neages were detected, two of them having human coun-terparts, while two other lineages have not been de-scribed in any other New World monkey species stud-ied. As in the common marmoset, the diversity of DRBalleles appears to have arisen largely by point muta-tions in the b-pleated sheets and by frequent exchangeof fixed sequence motifs in the a-helical portion. Pairsof alleles differing only at amino acid position b86 byan exchange of valine to glycine are present in Aotus,as in humans. Essential amino acid residues contribut-ing to MHC DR peptide binding pockets number 1 and
4 are conserved or semiconserved between HLA-DRand Aona-DRB molecules, indicating a capacity to bindsimilar peptide repertoires. These results support fullyour using Aotus monkeys as an animal model for evalu-ation of future subunit vaccine candidates.
Key words Aotus nancymaae 7 MHC class II DRmolecules 7 Allelic lineages 7 Polymorphism 7Peptide binding
Introduction
Proteins encoded within the class II region of the majorhistocompatibility complex (MHC), the HLA-D regionin humans, form polymorphic, noncovalently associatedab heterodimers expressed on the surface of special-ized antigen presenting cells. Three major loci in theHLA-D region have been identified, designated HLA-DR, -DQ, and -DP (Kappes and Strominger 1988).HLA-DR polymorphism appears to be responsible forvariations in the immune response of different individ-uals to antigens and may contribute to differences inthe susceptibility to infectious diseases and autoim-mune disorders (Hill 1998; Todd et al. 1988). Polymor-phism of HLA-DR is confined to a large extent to exon2 of the DRB gene, while the DRA gene is conserved(Brown et al. 1988; Fremont et al. 1996). The peptide-binding region (PBR) of the HLA-DR is formed as aninterchain dimer of the a1 and b1 domains, and differ-ences in sequences among alleles are clustered to re-gions of hypervariability in the b1 domains (Kaufmanet al. 1984). Recent elucidation of the three-dimension-al structure of HLA-DR has identified subregions orpockets in the PBR which influence the binding of pre-sented peptides and the subsequent recognition by Tcells (Stern et al. 1994a). Many of the residues liningthese pockets are highly polymorphic, probably deter-mining the different peptide binding specificities ofHLA-DR alleles (Brown et al. 1988, 1993; Chicz et al.1993; Stern et al. 1994).

220
A prominent and highly conserved hydrophobicpocket, pocket P1 accommodates the primary anchorresidue of the peptide bound. Therefore, this aminoacid has the relative position 1 in the peptide sequence.Pocket P1 appears to be a major determinant of pep-tide binding to HLA-DR (Hammer et al. 1993). PocketP4 is described as a further major determinant of pep-tide binding (Doherty et al. 1998; Fu et al. 1995; Wu-cherpfennig and Strominger 1995). It could be shownthat in particular the polymorphic residues at positionsb70, b71, and b74 contributing to pocket P4 stronglyinfluence the peptide-binding specificity (Stern et al.1994). On the basis of physicochemical properties ofthe amino acid residues at b70, b71, and b74 and theirinfluence on T-cell recognition, several groups of DRrestrictive supermotif patterns (RSP) have been identif-ied (Ou et al. 1998a). This allows the categorization ofHLA-DR alleles on the basis of function instead ofwhole sequence homologies. Peptides carrying a nega-tive charge at position 4 favorably bind to DR mole-cules with a positively charged RSP, but not to DRmolecules with a negatively charged RSP (Fu et al.1995; Ou et al. 1998; Smith et al. 1998).The contemporary living primates can be classified
into New World monkeys (Platyrrhini) and Old Worldsimians (Catarrhini). The New World monkeys and theOld World simians radiated about 58 million years ago(Ciochon and Chiarelli 1980). Several species of NewWorld monkeys are currently employed in biomedicalresearch and one important representative is Aotusspp, which can sustain in a predictable way the develop-ment of asexual forms of the two major human malariaparasites, Plasmodium falciparum and Plasmodium vi-vax (Gysin 1998). In 1988 the World Health Organiza-tion (WHO) recommended Aotus spp. as an experi-mental model for P. falciparum blood stage infectionsand it is currently used for the evaluation of candidatemalaria vaccines (Gramzinski et al. 1997). We compareimmunogenetic properties of human and Aotus mon-keys to further substantiate the suitability of this modelfor the evaluation of vaccine candidates. After charac-terization of the TCRVA and TCRVB repertoire(Favre et al. 1998; Vecino et al. 1999) we describe herethe sequence diversity of exon 2 of MHC DRB Aotusnancymaae (MhcAona-DRB) genes.
Materials and methods
Nomenclature
Official designations for Aotus DRB alleles were obtained fromR.E. Bontrop and Natasja G. de Groot (Biomedical Primate Re-search Centre-TNO, Rijswijk, The Netherlands). They are basedupon shared sequence motifs in the first stretch of base pairs,phylogenetic analysis, and comparison with sequences that arefound in other New World monkeys (Klein et al. 1990). Aona-DRB alleles that show similarity to human equivalents are de-picted by identical lineage numbers. The last two digits are arbi-trary and reflect the order in which the alleles were found. Aona-DRB alleles with no apparent human equivalent are designated
by a workshop number like Aona-DRB*Wn. The workshop defi-nition indicates that it has not yet been established whether thesealleles are encoded by a separate locus or belong to a novel li-neage for which its human equivalent has not been detected(Bontrop et al. 1999).
Animals
Animals caught in the Colombian Amazon Area close to Leticiawere selected at random to ensure the presence of a representa-tive repertoire of different alleles. Leucocytes from 15 healthy A.nancymaae monkeys were obtained either by density gradientseparation of peripheral blood obtained by venous punction or byspleenectomy as described (Garraud et al. 1994). A lymphoblas-toid cell line established in vitro by Epstein-Barr viral transforma-tion from monkey 3026 was used as source of genomic DNA andtotal RNA (Chizzolini et al. 1991).
Polymerase chain reaction, cloning, and sequencing of exon 2 ofAona-DRB genes
Genomic DNA was isolated either from leucocytes or cell line3026 using the NucleoSpin CcT kit (Macherey-Nagel AG, Oen-singen, Switzerland) according to the manufacturer’s protocol.MHC DRB gene-specific primers employed to amplify exon 2were sense (5b CCGGATCCTTCGTGTCCCCACAGCACG 3b)and anti-sense (5b TCGCCGCTGCACTGTGAAG 3b) as de-scribed (Tiercy et al. 1990). Amplifications were performed withthe following profile: 2 min 95 7C; 33! (30 s 96 7C, 30 s 55 7C, 30 s72 7C); 7 min 72 7C; soak at 4 7C. From monkeys 9191, 9200, 11145,11190, 11192, and cell line 3026 the second exon of HLA-DRBwas also amplified from cDNA. Therefore, total RNA was iso-lated from PBMC using the NucleoSpin RNA kit (Macherey-Na-gel AG) according to the manufacturer’s protocol. After reversetranscription using Superscript and oligo(dT)12–18 primer (Gibco-BRL Life Technologies, Basel, Switzerland), amplifications wereperformed with primers sense (5b GGTGCTGAGCTCCCGAC-CTGGC3b) and anti-sense (5b AGGTTGTGGTGCTGCAGGG-C3b) binding to the sequence encoding the signal peptide or ami-no acid positions 107 to 114 in exon 3, respectively. This differentset of primers was employed to reduce the possibility that someDRB loci could not be amplified with the first set of primers. Po-lymerase chain reaction (PCR) cycling conditions were as follows:2 min 95 7C; 33! (30 s 96 7C, 30 s 55 7C, 30 s 72 7C); 7 min 72 7C;soak at 4 7C. PCR products were purified using a PCR productpurification kit (Roche Molecular Biochemicals, Rotkreuz, Swit-zerland) according to the manufacturer’s protocol and cloned intothe pGEM5 T-vector (Promega, Catalys AG, Wallisellen, Swit-zerland). After isolation of plasmids using the NucleoSpin kit(Macherey-Nagel AG), double-stranded plasmid DNA was se-quenced and analyzed employing an ABI PRISM 310 geneticanalyzer (Perkin Elmer, Foster City, Calif.) using the ABIPRISM Sequencing Analysis 3.3 and MT Navigator 1.0.2.b3 soft-ware. The reported alleles represent at least three identical clonesthat were obtained after independent amplifications from thesame animal or at least two clones derived from two or more dif-ferent animals.
Phylogenetic analysis
Phylogenetic analysis was performed employing the PHYLIP3.572 package available under http://bioweb.pasteur.fr. The phy-logenetic tree was constructed according to the neighbor-joiningmethod (Saitou and Nei 1987). In accordance with the proposednomenclature for MHC in nonhuman species (Klein et al. 1990),DRB alleles from Callithrix jacchus are referred to as Caja-DRB,Cebus apella as Ceap-DRB, Callicebus moloch as Camo-DRB,and Saimiri sciureus as Sasc-DRB.

221
Results
Polymorphism of Aona-DRB
The MHC DRB exon 2 from a panel of randomly chos-en A. nancymaae monkeys was amplified by PCR fromcDNA and/or genomic DNA. A total of 239 sequenceswere obtained which identified 34 different alleles. Thederived exon 2 Aona-DRB nucleotide sequences aregiven in Fig. 1. The deduced amino acid sequences areshown in Fig. 2. None of the 34 sequences display fea-tures that would suggest they are pseudogenes. TheAona-DRB sequences lack the characteristic motifs atthe 3b end of exon 2 (amino acid position 84 to 92,namely EILDRFLVP, EISERFLVP, or QILERFLVP)described in other Platyrrhini sequences (Antunes et al.1998; Trtkova et al. 1993).The identified Aona-DRB alleles can be divided into
two large groups – those resembling Catarrhini DRBsequences and those unique to Platyrrhini. Aona-DRB1*03 and Aona-DRB3*06 alleles belong to theformer group, and Aona-DRB*W13, Aona-DRB*W18,Aona-DRB*W29, and Aona-DRB*W30 to the latter.These findings indicate that the human species and theAotus monkeys share certain MHC DRB trans-speciesallelic lineages. A phylogenetic tree constructed for theexon 2 sequences by the neighbor-joining method (Sai-tou and Nei 1987) supports the notion that the Aotusalleles are closely related to each other. Furthermore,selected human alleles like allele HLA-DRB1*0801cluster together with Aona-DRB1*0301, Aona-DRB1*0302, and Aona-DRB1*0308 (Fig. 3). Sequencesof the related allelic lineage DRB1*03, derived fromCallithrix jacchus and Saimiri sciureus are located on adifferent branch (Fig. 3).Codons 7 to 54 encode a b-pleated sheet platform
structure underlying the antigen-binding groove, andcodons 60 to 80 encode a long a-helical loop that formsthe lateral boundary of the antigen-binding MHCgroove (Stern et al. 1994). In the Aona-DRB1*03 and-DRB3*06 lineages the contact residues in the b-pleated sheets of the peptide-binding site are highlyconserved and most of the variations seen at the codonsfor the contact residues within the b-pleated sheets canbe explained by point mutations. Six different fixedmotifs (YWNSQKDFLEERRALVDTYCR, YWNSQ-KDYVEQKRGQVDNYCR, YWNSQ KDYVEQKR-GRVDNYCR, YWNSQKDYVERKRGRVDNYCR,YFNSLKDFMEETRAAVDTYCK, YWNSLKERLE-YLRAAVDTYCR) encompassing amino acid posi-tions 60 to 80 were found in the DRB1*03 andDRB3*06 alleles. Most of them were found in both al-lelic lineages (Fig. 2).The Aona-DRB*W1801 and Aona-DRB*W1802 al-
leles share a stretch of amino acids at positions 60–80with Aona-DRBW2903. Aona-DRB3*2901, Aona-DRB3*2902, Aona-DRW*1301, and Aona-DRW*1302share motifs with the Aona-DRB1*03 and DRB3*06 li-neage members (Fig. 2).
Certain residues of exon 2 are important for the cor-rect structure and function of DR molecules (Kaufmanet al. 1984). All Aona-DRB alleles have cysteins at bothresidues 15 and 79 needed for the correct folding of DRmolecules (Kaufman et al. 1984). HLA-DR moleculeshave a tendency to form dimers and certain b-chain re-sidues are involved in this interaction. The amino acidresidues at the interface are at position 49, 50, 51, 52,and 55, which are extremely conserved in humans (al-anine, valine, threonine, glutamic acid, and arginine, re-spectively (Brown et al. 1993). The same amino acidscan be found at the corresponding positions of Aona-DRB alleles (Fig. 2).
Distribution of Aona-DRB alleles between individualanimals
The number of Aona-DRB alleles identified per animalvaried between 3 and 1213. In those monkeys wherefewer alleles were identified, a more extensive se-quence analysis of a larger panel of DNA clones mightresult in the identification of further allele sequences.In addition, some DRB alleles may not have been am-plified under the conditions used, which could explainthe heterogeneity in the number of DRB alleles per an-imal. Several animals appear to possess more than twoDRB1*03 and DRB3 genes, so that more than onegene of each locus could be borne by a single chromo-some (Table 1). The Aona-DRB1*0305 allele wasfound in all (11/15) animals from which a larger num-ber of DNA clones were analyzed. This allele may becontrolled by a separate DRB1 locus as described inanother New World monkey, the common marmoset(Antunes et al. 1998). However, detailed contig map-ping studies are needed to confirm these assumptions.
Amino acids contributing to Aona-DRB bindingpockets
The prominent hydrophobic pocket 1 accommodatesthe primary peptide anchor residue and is apart from aglycine/valine dimorphism at position b86 highly con-served among different HLA-DR molecules. As with tothis dimorphism at position b86 in human alleles, wecould identify three pairs of Aona-DRB alleles differ-ing at position b86 by glycine or valine, namely Aona-DRB1*0304/0311, Aona-DRB1*0305/0312, and Aona-DRB3*0603/0604. Unlike in humans, position b86 ofAona-DRB can also be occupied by the hydrophobicamino acids alanine and phenylalanine.HLA-DR molecules can be grouped apart, accord-
ing to their primary amino acid sequences, into func-tional categories on the basis of their ability to bind andpresent antigenic peptides to T cells and their associa-tion with susceptibility or resistance to disease (Ou et

222
Fig. 1 Nucleotide sequencesof Aona-DRB exon 2. In thenumbering system used, codon1 specifies the first amino acidresidue of the mature protein.The top line represents a ma-jority consensus of primate se-quences (Trtkova et al. 1993).Identity with the consensus se-quence is indicated by dashes(–)

223
Fig. 1 Continued

224
Fig. 1 Continued

225
Fig. 2 Alignment of Aona-DRB-deduced amino acid se-quences of exon 2. The topline gives the majority consen-sus of primate sequences(Trtkova et al. 1993). Identitywith the consensus sequence isindicated by dashes (–)
al. 1998). In the population of Aotus monkeys investi-gated, seven RSP could be discriminated which are po-sitively charged, di-charged, negatively charged, andneutral (Table 2). Some of these RSP are identical tohuman RSP, indicating that similar residues relevantfor antigen presentation and formation of the T-cell re-ceptor repertoire are shared between Aotus monkeysand humans.
The a-helical portion of the b1 domain has beenshown to be of importance for the restriction of T-cellresponses to a peptide antigen (Doherty et al. 1998). Acomparison of this stretch of amino acids shared byAona-DRB1*0301/0302/0308 with a group of HLA-DRB alleles revealed that this motif is 100% conservedat the amino acid level except for a single conservativeexchange of an aspartate to glutamate at position b70.

226
Fig. 3 Phylogenetic tree con-structed according to theneighbor-joining methodshowing the relationship ofexon 2 sequences between allidentified Aona-DRB allelesand selected MHC-DRB al-leles from New World mon-keys and human. Bootstrapvalues are indicated
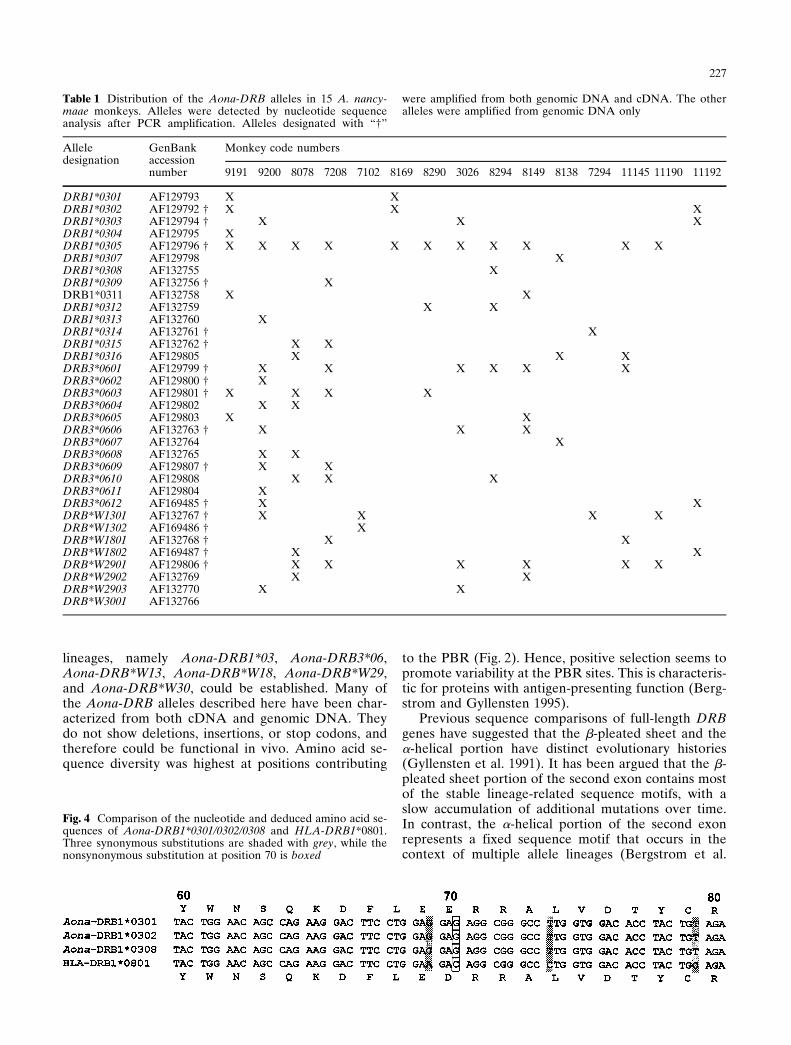
Hence, important amino acid residues contributing topocket P4 are conserved except for a conservative ex-change. A comparison at the nucleotide level (Fig. 4)demonstrates that three synonymous substitutions andone nonsynonymous substitution exist in this stretch of20 amino acids. A GenBank search revealed that onlyhumans and Aotus share this particular sequencemotif.
Discussion
So far, a few nonhuman primate species such as com-mon chimpanzees, rhesus macaques, cotton-top tamar-ins, and common marmosets have been tested thor-oughly for MHC-DRB diversity at the nucleotide se-quence level (O’hUigin et al. 1993). In the presentstudy we analyzed DRB gene nucleotide sequences ofthe A. nancymaae exon 2. The presence of six different
227
Table 1 Distribution of the Aona-DRB alleles in 15 A. nancy-maae monkeys. Alleles were detected by nucleotide sequenceanalysis after PCR amplification. Alleles designated with “†”
were amplified from both genomic DNA and cDNA. The otheralleles were amplified from genomic DNA only
Alleledesignation
GenBankaccession
Monkey code numbers
number 9191 9200 8078 7208 7102 8169 8290 3026 8294 8149 8138 7294 11145 11190 11192
DRB1*0301 AF129793 X XDRB1*0302 AF129792 † X X XDRB1*0303 AF129794 † X X XDRB1*0304 AF129795 XDRB1*0305 AF129796 † X X X X X X X X X X XDRB1*0307 AF129798 XDRB1*0308 AF132755 XDRB1*0309 AF132756 † XDRB1*0311 AF132758 X XDRB1*0312 AF132759 X XDRB1*0313 AF132760 XDRB1*0314 AF132761 † XDRB1*0315 AF132762 † X XDRB1*0316 AF129805 X X XDRB3*0601 AF129799 † X X X X X XDRB3*0602 AF129800 † XDRB3*0603 AF129801 † X X X XDRB3*0604 AF129802 X XDRB3*0605 AF129803 X XDRB3*0606 AF132763 † X X XDRB3*0607 AF132764 XDRB3*0608 AF132765 X XDRB3*0609 AF129807 † X XDRB3*0610 AF129808 X X XDRB3*0611 AF129804 XDRB3*0612 AF169485 † X XDRB*W1301 AF132767 † X X X XDRB*W1302 AF169486 † XDRB*W1801 AF132768 † X XDRB*W1802 AF169487 † X XDRB*W2901 AF129806 † X X X X X XDRB*W2902 AF132769 X XDRB*W2903 AF132770 X XDRB*W3001 AF132766
Fig. 4 Comparison of the nucleotide and deduced amino acid se-quences of Aona-DRB1*0301/0302/0308 and HLA-DRB1*0801.Three synonymous substitutions are shaded with grey, while thenonsynonymous substitution at position 70 is boxed
lineages, namely Aona-DRB1*03, Aona-DRB3*06,Aona-DRB*W13, Aona-DRB*W18, Aona-DRB*W29,and Aona-DRB*W30, could be established. Many ofthe Aona-DRB alleles described here have been char-acterized from both cDNA and genomic DNA. Theydo not show deletions, insertions, or stop codons, andtherefore could be functional in vivo. Amino acid se-quence diversity was highest at positions contributing
to the PBR (Fig. 2). Hence, positive selection seems topromote variability at the PBR sites. This is characteris-tic for proteins with antigen-presenting function (Berg-strom and Gyllensten 1995).Previous sequence comparisons of full-length DRB
genes have suggested that the b-pleated sheet and thea-helical portion have distinct evolutionary histories(Gyllensten et al. 1991). It has been argued that the b-pleated sheet portion of the second exon contains mostof the stable lineage-related sequence motifs, with aslow accumulation of additional mutations over time.In contrast, the a-helical portion of the second exonrepresents a fixed sequence motif that occurs in thecontext of multiple allele lineages (Bergstrom et al.

228
Table 2 DR RSP defined bypolymorphic residues at posi-tions 70, 71, and 74 of Aona-DR b chains
DR restrictive supermotif pattern (RSP)a
Aona-DRB YLAb ETAd ERLd DRS RKR/Qc QKR/Qd LKQalleles (n) (–) (B) (B) (c) (c) (c)
DRB3*0610 YLADRB3*0611 ETADRB1*0301 ERLDRB*W3001 DRSDRB1*0306 RKQDRB1*0307 RKRDRB1*0305 QKRDRB3*0601 QKQDRB*W2903 LKQ
a DR RSP represents motifs shared by multiple Aona-DR alleles. The one-letter amino acid code isusedb Overall charge of the DR restrictive supermotif residues is indicated by (c), positively chargedsupermotif; (–), negatively charged supermotif; (B), dicharged supermotif; (n) uncharged supermo-tifc Amino acids R and Q are considered as paired polymorphic residues which would be expected toexert similar influences on peptide binding and T-cell recognition (Ou et al. 1998)d DR RSP similar between human and A. nancymaae
1999; Gaur and Nepom 1996). Several molecular mech-anisms have been postulated to contribute to the gener-ation of genetic diversity at the MHC loci. They includepoint mutations, complex mutations, and recombina-tion events leading to combinations of the b-pleatedsheet of one allele with that of variants of the a-helix ofanother allele (Erlich and Gyllensten 1991). The Aona-DRB alleles found in this study provide evidence forthe occurrence of the first and last mechanism. Theseobservations are consistent with the idea that recombi-nation events generating new alleles generally occurover a stretch of about 120 base pairs of exon 2 encod-ing the a-helix (Bergstrom et al. 1999). The mechanismby which such exchanges occur remains to be deter-mined. A high level of recombination is also describedin the common marmoset (Antunes et al. 1998). Thedendrogram in Fig. 3 supports the notion that Aona-DRB alleles lineages follow the trans-species model ofevolution, since several Aona-DRB sequences are moresimilar to different species orthologous sequences thanto other Aona-DRB sequences (Klein 1987). MHCDRB1*03- and MHC DRB3*-like lineages can befound in other New World monkeys like Callithrix jac-chus jacchus, Saguinus oedipus, and Callicebus moloch,but has been not described in Saimiri sciureus and Ce-bus apella (Trtkova et al. 1993). Therefore, the pres-ence of exon 2 sequences with a high similarity betweenCatarrhini and Platyrrhini MHC DRB1*03 and -DRB3sequences might be explained by convergent evolution(Antunes et al. 1998; Bontrop et al. 1999; Trtkova et al.1995). In contrast to humans, the Aona-DRB3 locus ex-hibits extensive polymorphism in Aotus. Thus, diversifi-cation of MHC isotypes is not uniform in different pri-mate species (Bontrop et al. 1995, 1999). DRB1*03 andDRB3 genes occur together in individual Aotus mon-keys. The two genes may therefore reside on the samechromosome, forming a DRB1*03/DRB3 haploytpe, asit is known to occur in humans and gorillas (Klein et al.
1991). Hence, it is possible that the DRB1*03 andDRB3 genes remained together in some haplotypes inboth the Catarrhini and Platyrrhini. The importance ofthe MHC DRB1*03 lineage is stressed by the observa-tion that members of this lineage have been detected inmany species and it presumably plays a distinctive rolein mounting immune responses against heat shock pro-teins of mycobacteria (Geluk et al. 1993). The emer-gence of similar motifs in several species could be anindication that the parasites or infectious threats tothese species share certain features. Antigenic threatsare time- and location-dependent and new allelesagainst specific threats will be developed in parallel tosuccessful old allelic lineages. Aona-DRB*W13, Aona-DRB*W18, Aona-DRB*W29, and Aona-DRB*W30might have been established and expanded by Aotus inresponse to its specific environmental challenges.The number of different DRB genes per individual
varied, and up to 13 different Aona-DRB genes (mon-key 9200) could be amplified (Table 1). Family data arenot available and therefore direct assignment of genesto haplotypes is not possible. It is of interest that someof the genes that occupy a single locus in anthropoidprimates are apparently multiplied in the Aotus. Multi-plication of MHC-DRB loci has been demonstrated di-rectly by cosmid cloning analysis in the orangutan(Schonbach et al. 1993), but might also be present inother species (Bontrop et al. 1990, 1995; Figueroa et al.1994; Trtkova et al. 1993).One particular motif encompassing 20 amino acids
ranging from position 60 to 80 of exon 2 is highly simi-lar to human motifs (Fig. 4). This region of the HLA-DR molecule is important for the interaction of the DRb chain with both the peptide presented and the anti-gen-specific TCR (Doherty et al. 1998). The motif foundin Aona-DRB1*0301/0302/0308 is 100% conserved be-tween Aotus and certain HLA-DRB alleles except for aconservative exchange at position b70. This particular

229
motif can be found in members of HLA-DRB1*04,HLA-DRB1*08, HLA-DRB1*11, HLA-DRB1*13,HLA-DRB1*14, HLA-DRB1*16, and HLA-DRB5*01lineages. The evolution of the MHC and TCR systemscannot be seen independent from each other, since thea-helix of DR molecules interact with the TCR (Silveret al. 1997). We have established previously the reper-toire of TCR AV, -J, and -C and TCR BV and -J genesof A. nancymaae (Favre et al. 1998; Vecino et al. 1999).All Aotus TRAV and TRBV amino acid sequencesidentified were 75 to 90% identical to their humancounterparts. One explanation for this similarity mightbe the conservation of some central motifs of MHC-DRbetween Aotus and human.The prominent hydrophobic pocket P1 accommo-
dates primary anchor residues of the peptide boundand is, apart from the glycine/valine dimorphism at po-sition b86, highly conserved among different HLA-DRmolecules (Davenport et al. 1995). This strict duality isnot exclusive for humans but can be extended to Cerco-pithecus aethiops and A. nancymaae where pairs of al-leles differing at position b86 in glycine or valine havebeen described (Bontrop et al. 1995; Rosal-Sanchez etal. 1998). When glycine is present at b86, aliphatic oraromatic residues can anchor the peptide; with valine atb86, the pocket is smaller so that tyrosine and trypto-phane cannot be accommodated. The determination ofHLA-DR peptide-binding motifs has shown that a hy-drophobic anchor residue (F, Y, W, M, I, V, or L) atposition 1 is required for binding to these molecules(Hammer et al. 1993). Position b86 of Aona-DRB is oc-cupied with amino acids glycine, valine, alanine andphenylalanine. Hence, pocket P1 of Aona-DRB mightbind a variety of amino acids with aliphatic or aromaticside chains maintaining the hydrophobic interactionsalong the side and floor of the pocket.We have compared the RSP designated by the poly-
morphic residues at positions 70, 71, and 74 of DR b-chains of humans with the corresponding residuesfound in the alleles described herein (Ou et al. 1998).We were able to distinguish seven different RSP in theAotus (Table 2), each of which are shared by differentalleles. In humans RSP can have overall negative, posi-tive, positive/negative, and neutral charges. This distri-bution of charges can also be found in the Aotus alleles,and some RSP in Aotus are very similar to human RSP,indicating that comparable ranges of T-cell epitopescould be bound and presented by human and Aona-DRB alleles. However, only detailed binding studies ofpeptides to isolated Aona-DRB alleles will confirm thisassumption. Our data support the view that A. nancy-maae is a suitable model system for testing epitope-focused peptide vaccines against P. falciparum and oth-er infectious diseases.
Acknowledgements This work was supported in part by funds ofthe Swiss National Science Foundation (31-52068.97). The Institu-to de Inmunologia is financed by the Colombian Ministry ofHealth and by Colciencias.
References
Antunes SG, de Groot NG, Brok H, Doxiadis G, Menezes AA,Otting N, Bontrop RE (1998) The common marmoset: a newworld primate species with limited Mhc class II variability.Proc Natl Acad Sci USA 95 :11745–11750
Bergstrom T, Gyllensten U (1995) Evolution of Mhc class II poly-morphism: the rise and fall of class II gene function in prima-tes. Immunol Rev 143 :13–31
Bergstrom TF, Erlandsson R, Engkvist H, Josefsson A, ErlichHA, Gyllensten U (1999) Phylogenetic history of hominoidDRB loci and alleles inferred from intron sequences. Immu-nol Rev 167 :351–365
Bontrop RE, Broos LA, Pham K, Bakas RM, Otting N, Jonker M(1990) The chimpanzee major histocompatibility complexclass II DR subregion contains an unexpectedly high numberof beta-chain genes. Immunogenetics 32 :272–280
Bontrop RE, Otting N, de Groot NG, Doxiadis GG (1999) Majorhistocompatibility complex class II polymorphisms in prima-tes. Immunol Rev 167 :339–350
Bontrop RE, Otting N, Slierendregt BL, Lanchbury JS (1995)Evolution of major histocompatibility complex polymor-phisms and T-cell receptor diversity in primates. ImmunolRev 143 :33–62
Brown JH, Jardetzky T, Saper MA, Samraoui B, Bjorkman PJ,Wiley DC (1988) A hypothetical model of the foreign antigenbinding site of class II histocompatibility molecules. Nature332 :845–850
Brown JH, Jardetzky TS, Gorga JC, Stern LJ, Urban RG, Strom-inger JL, Wiley DC (1993) Three-dimensional structure of thehuman class II histocompatibility antigen HLA-DR1. Nature364 :33–39
Chicz RM, Urban RG, Gorga JC, Vignali DA, Lane WS, Strom-inger JL (1993) Specificity and promiscuity among naturallyprocessed peptides bound to HLA-DR alleles. J Exp Med178 :27–47
Chizzolini C, Sulzer AJ, Olsen-Rasmussen MA, Collins WE(1991) Epstein-Barr virus transformation of Saimiri sciureus(squirrel monkey) B cells and generation of a Plasmodiumbrasilianum-specific monoclonal antibody in P. brasilianum-infected monkeys. Infect Immun 59 :2285–2290
Ciochon RL, Chiarelli AB (1980) Paleobiogeographic perspec-tives on the origin of the Platyrrhini. In: Chichon RL, ChiarelliAB (eds) Evolutionary biology of the New World monkeysand continental drift. Plenum, New York, pp 459–493
Davenport MP, Quinn CL, Chicz RM, Green BN, Willis AC,Lane WS, Bell JI, Hill AV (1995) Naturally processed pep-tides from two disease-resistance-associated HLA-DR13 al-leles show related sequence motifs and the effects of the di-morphism at position 86 of the HLA-DR beta chain. ProcNatl Acad Sci USA 92 :6567–6571
Doherty DG, Penzotti JE, Koelle DM, Kwok WW, Lybrand TP,Masewicz S, Nepom GT (1998) Structural basis of specificityand degeneracy of T cell recognition: pluriallelic restriction ofT-cell responses to a peptide antigen involves both specificand promiscuous interactions between the T-cell receptor,peptide, and HLA-DR. J Immunol 161 :3527–3535
Erlich HA, Gyllensten UB (1991) Shared epitopes among HLAclass II alleles: gene conversion, common ancestry and balanc-ing selection. Immunol Today 12 :411–414
Favre N, Daubenberger C, Marfurt J, Moreno A, Patarroyo M,Pluschke G (1998) Sequence and diversity of T-cell receptoralpha V, J, and C genes of the owl monkey Aotus nancymaae.Immunogenetics 48 :253–259
Figueroa F, O’hUigin C, Tichy H, Klein J (1994 The origin of theprimate Mhc-DRB genes and allelic lineages as deduced fromthe study of prosimians. J Immunol 152 :4455–4465
Fremont DH, Hendrickson WA, Marrack P, Kappler J (1996)Structures of an MHC class II molecule with covalently boundsingle peptides. Science 272 :1001–1004

230
Fu XT, Bono CP, Woulfe SL, Swearingen C, Summers NL, Sini-gaglia F, Sette A, Schwartz BD, Karr RW (1995) Pocket 4 ofthe HLA-DR (alpha, beta 1*0401) molecule is a major deter-minant of T-cells recognition of peptide. J Exp Med181 :915–926
Garraud O, Perraut R, Gysin J, Behr C, Dubois P, BonnemainsB, Jouin H, Michel JC, Pereira da Silva L (1994) Manipulatingblood T-cells and b-cells from squirrel monkeys: some techni-cal considerations. J Immunol Methods 173 :165–173
Gaur LK, Nepom GT (1996) Ancestral major histocompatibilitycomplex DRB genes beget conserved patterns of localized po-lymorphisms. Proc Natl Acad Sci USA 93 :5380–5383
Geluk A, Elferink DG, Slierendregt BL, Meijgaarden KE van,Vries RR de, Ottenhoff TH, Bontrop RE (1993) Evolutionaryconservation of major histocompatibility complex-DR/pep-tide/T-cell interactions in primates. J Exp Med 177 :979–987
Gramzinski RA, Maris DC, Doolan D, Charoenvit Y, Obaldia N,Rossan R, Sedegah M, Wang R, Hobart P, Margalith M, Hoff-man S (1997) Malaria DNA vaccines in Aotus monkeys. Vac-cine 15 :913–915
Gyllensten UB, Sundvall M, Erlich HA (1991) Allelic diversity isgenerated by intraexon sequence exchange at the DRB1 locusof primates. Proc Natl Acad Sci USA 88 :3686–3690
Gysin J (1998) Animal models: primates. In: Sherman IW (ed)Malaria: parasite biology, pathogenesis and protection. ASMPress, Washington D.C., pp 419–439
Hammer J, Valsasnini P, Tolba K, Bolin D, Higelin J, Takacs B,Sinigaglia F (1993) Promiscuous and allele-specific anchors inHLA-DR-binding peptides. Cell 74 :197–203
Hill AV (1998) The immunogenetics of human infectious dis-eases. Annu Rev Immunol 16 :593–617
Kappes D, Strominger JL (1988) Human class II major histocom-patibility complex genes and proteins. Annu Rev Biochem57:991–1028
Kaufman JF, Auffray C, Korman AJ, Shackelford DA, Stromin-ger J (1984) The class II molecules of the human and murinemajor histocompatibility complex. Cell 36 :1–13
Klein J (1987) Origin of major histocompatibility complex poly-morphism: the trans- species hypothesis. Hum Immunol19 :155–162
Klein J, Bontrop RE, Dawkins RL, Erlich HA, Gyllensten UB,Heise ER, Jones PP, Parham P, Wakeland EK, Watkins DI(1990) Nomenclature for the major histocompatibility com-plexes of different species: a proposal. Immunogenetics31 :217–219
Klein J, O’hUigin C, Kasahara M, Vincek V, Klein D, Figueroa F(1991) Frozen haplotypes in MHC evolution. In: Klein J,Klein D (eds) Molecular evolution of the major histocompati-bility complex. Springer, Berlin, pp 261–286
O’hUigin C, Bontrop R, Klein J (1993) Nonhuman primate Mhc-DRB sequences: a compilation. Immunogenetics 38 :165–183
Ou D, Mitchell LA, Tingle AJ (1998) A new categorization ofHLA DR alleles on a functional basis. Hum Immunol59 :665–676
Rosal-Sanchez M, Paz-Artal E, Moreno-Pelayo MA, Martinez-Quiles N, Martinez-Laso J, Martin-Villa JM, Arnaiz-Villena A(1998) Polymorphism of Mhc-DRB alleles in Cercopithecusaethiops (green monkey): generation and functionality. TissueAntigens 51 :541–548
Saitou N, Nei M (1987) The neighbor-joining method: a newmethod for reconstructing phylogenetic trees. Mol Biol Evol4 :406–425
Schonbach C, Vincek V, Mayer WE, Golubic M, O’hUigin C,Klein J (1993) Multiplication of Mhc-DRB5 loci in the oran-gutan: implications for the evolution of DRB haplotypes.Mamm Genome 4 :159–170
Silver J, Gulwani-Akolkar B, Leffell MS, Bias W, Kotb M, El De-hamy M, Guedez YB, Kotby A, Obata F, Kashiwagi N (1997)HLA and the T-cell receptor. In: Charron D (ed) HLA. Ge-netic diversity of HLA. Medical and Scientific InternationalPublisher, Sevres, pp 183–187
Smith KJ, Pyrdol J, Gauthier L, Wiley DC, Wucherpfennig KW(1998) Crystal structure of HLA-DR2 (DRA*0101,DRB1*1501) complexed with a peptide from human myelinbasic protein. J Exp Med 188 :1511–1520
Stern LJ, Brown JH, Jardetzky TS, Gorga JC, Urban RG, Strom-inger JL, Wiley DC (1994) Crystal structure of the humanclass II MHC protein HLA-DR1 complexed with an influenzavirus peptide. Nature 368 :215–221
Tiercy JM, Jeannet M, Mach B (1990) A new HLA-DRB1 allelewithin the DRw52 supertypic specificity (DRw13-DwHAG):sequencing and direct identification by oligonucleotide typing.Eur J Immunol 20 :237–241
Todd JA, Acha-Orbea H, Bell JI, Chao N, Fronek Z, Jacob CO,McDermott M, Sinha AA, Timmerman L, Steinman L (1988)A molecular basis for MHC class II-associated autoimmunity.Science 240 :1003–1009
Trtkova K, Kupfermann H, Grahovac B, Mayer WE, O’hUigin C,Tichy H, Bontrop R, Klein J (1993) Mhc-DRB genes of pla-tyrrhine primates. Immunogenetics 38 :210–222
Trtkova K, Mayer WE, O’hUigin C, Klein J (1995) Mhc-DRBgenes and the origin of New World monkeys. Mol PhylogenetEvol 4 :408–419
Vecino W, Daubenberger C, Rodriguez R, Moreno A, PatarroyoM, Pluschke G (1999) Sequence and diversity of T-cell recep-tor b-chain V and J genes of the owl monkey Aotus nancy-maae. Immunogenetics 49 :792–799
Wucherpfennig KW, Strominger JL (1995) Selective binding ofself peptides to disease-associated major histocompatibilitycomplex (MHC) molecules: a mechanism for MHC- linkedsusceptibility to human autoimmune diseases. J Exp Med181 :1597–1601
Copyright © 2022 FDOKUMEN


















