
Owl monkey MHC-DRB exon 2 reveals high similarity with several HLA-DRB lineages
17
ORIGINAL PAPER Owl monkey MHC-DRB exon 2 reveals high similarity with several HLA-DRB lineages Carlos F. Suárez & Manuel E. Patarroyo & Esperanza Trujillo & Mónica Estupiñán & Juan E. Baquero & Carlos Parra & Raúl Rodriguez Received: 25 April 2005 / Accepted: 3 May 2006 / Published online: 22 June 2006 # Springer-Verlag 2006 Abstract One hundred and ten novel MHC-DRB gene exon 2 nucleotide sequences were sequenced in 96 monkeys from three owl monkey species (67 from Aotus nancymaae, 30 from Aotus nigriceps and 13 from Aotus vociferans). Owl monkeys, like humans, have high MHC- DRB allele polymorphism, revealing a striking similarity with several human allele lineages in the peptide binding region and presenting major convergence with DRB lineages from several Catarrhini (humans, apes and Old World monkeys) rather than with others New World monkeys (Platyrrhini). The parallelism between human and Aotus MHC-DRB reveals additional similarities re- garding variability pattern, selection pressure and physico- chemical constraints in amino acid replacements. These observations concerning previous findings of similarity between the Aotus immune system molecules and their human counterparts affirm this specie’ s usefulness as an excellent animal model in biomedical research. Keywords Animal models . New World monkeys . Molecular evolution . Molecular convergence . MHC class II molecules . MHC-DRB Abbreviations PBR peptide binding region TSP trans-specific polymorphism OWM old world monkey NWM new world monkey MYA million years ago Introduction The major histocompatibility complex (MHC) is a multi- gene family belonging to the immunoglobulin super-family, encoding receptor glycoproteins expressed on cell mem- brane. MHC proteins play a central role in recognising self and non-self by presenting foreign peptides to be recog- nised by T cells, therefore constituting key pieces in defence against foreign substances. Two large groups of MHC molecules, which can be functionally and structurally distinguished. MHC class I molecules have a peptide binding region (PBR) consisting of two domains (α1 and α2) encoded by a single gene and expressed on all types of nucleated cells presenting intracellular peptides to CD8+T lymphocytes. MHC class II encoded molecules have a PBR encoded by two genes (α and β chains) expressed on antigen-presenting cells (APC), such as monocytes, B lymphocytes and macrophages to CD4+ T lymphocytes (Edwards and Hedirck 1998). MHC alleles from several non-human primate species have been studied, i.e. apes such as chimpanzees (Kenter et al. 1992; Mayer et al. 1992), Old World monkeys (OWM) such as Macaca (Zhu et al. 1991; Doxiadis et al. 2000, 2001, Otting et al. 2000) and Cercopithecus (Rosal et al. 1998), New World monkeys (NWM) such as cotton-top tamarins (Gyllensten et al. 1994; Kriener et al. 2001), Immunogenetics (2006) 58:542–558 DOI 10.1007/s00251-006-0127-0 Experiments carried out in this work complied with current Colombian Ministry of Health law and regulations governing animal care and handling. C. F. Suárez : M. E. Patarroyo (*) : E. Trujillo : M. Estupiñán : J. E. Baquero : C. Parra : R. Rodriguez Fundación Instituto de Inmunmología de Colombia (FIDIC), Carrera 50 No. 26-00, Bogotá, Colombia e-mail: [email protected] M. E. Patarroyo Universidad Nacional de Colombia, Bogotá, Colombia
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Owl monkey MHC-DRB exon 2 reveals high similarity with several HLA-DRB lineages

ORIGINAL PAPER
Owl monkey MHC-DRB exon 2 reveals high similaritywith several HLA-DRB lineages
Carlos F. Suárez & Manuel E. Patarroyo &
Esperanza Trujillo & Mónica Estupiñán &
Juan E. Baquero & Carlos Parra & Raúl Rodriguez
Received: 25 April 2005 /Accepted: 3 May 2006 / Published online: 22 June 2006# Springer-Verlag 2006
Abstract One hundred and ten novel MHC-DRB geneexon 2 nucleotide sequences were sequenced in 96monkeys from three owl monkey species (67 from Aotusnancymaae, 30 from Aotus nigriceps and 13 from Aotusvociferans). Owl monkeys, like humans, have high MHC-DRB allele polymorphism, revealing a striking similaritywith several human allele lineages in the peptide bindingregion and presenting major convergence with DRBlineages from several Catarrhini (humans, apes and OldWorld monkeys) rather than with others New Worldmonkeys (Platyrrhini). The parallelism between humanand Aotus MHC-DRB reveals additional similarities re-garding variability pattern, selection pressure and physico-chemical constraints in amino acid replacements. Theseobservations concerning previous findings of similaritybetween the Aotus immune system molecules and theirhuman counterparts affirm this specie’s usefulness as anexcellent animal model in biomedical research.
Keywords Animal models . NewWorld monkeys .
Molecular evolution .Molecular convergence .MHC class IImolecules .MHC-DRB
AbbreviationsPBR peptide binding regionTSP trans-specific polymorphismOWM old world monkeyNWM new world monkeyMYA million years ago
Introduction
The major histocompatibility complex (MHC) is a multi-gene family belonging to the immunoglobulin super-family,encoding receptor glycoproteins expressed on cell mem-brane. MHC proteins play a central role in recognising selfand non-self by presenting foreign peptides to be recog-nised by T cells, therefore constituting key pieces indefence against foreign substances.
Two large groups of MHC molecules, which can befunctionally and structurally distinguished. MHC class Imolecules have a peptide binding region (PBR) consisting oftwo domains (α1 and α2) encoded by a single gene andexpressed on all types of nucleated cells presentingintracellular peptides to CD8+T lymphocytes. MHC classII encoded molecules have a PBR encoded by two genes(α and β chains) expressed on antigen-presenting cells(APC), such as monocytes, B lymphocytes and macrophagesto CD4+ T lymphocytes (Edwards and Hedirck 1998).
MHC alleles from several non-human primate specieshave been studied, i.e. apes such as chimpanzees (Kenter etal. 1992; Mayer et al. 1992), Old World monkeys (OWM)such as Macaca (Zhu et al. 1991; Doxiadis et al. 2000,2001, Otting et al. 2000) and Cercopithecus (Rosal et al.1998), New World monkeys (NWM) such as cotton-toptamarins (Gyllensten et al. 1994; Kriener et al. 2001),
Immunogenetics (2006) 58:542–558DOI 10.1007/s00251-006-0127-0
Experiments carried out in this work complied with currentColombian Ministry of Health law and regulations governing animalcare and handling.
C. F. Suárez :M. E. Patarroyo (*) : E. Trujillo :M. Estupiñán :J. E. Baquero :C. Parra : R. RodriguezFundación Instituto de Inmunmología de Colombia (FIDIC),Carrera 50 No. 26-00,Bogotá, Colombiae-mail: [email protected]
M. E. PatarroyoUniversidad Nacional de Colombia,Bogotá, Colombia

common marmosets (Wu et al. 2000; Antunes et al. 1998),owl monkeys (Niño-Vasquez et al. 2000) and lemurs (Go etal. 2002). MHC molecules present common characteristicsin all these groups (such as polymorphism and diversifyingselection) tied to functional and structural restrictions.Some lineages are shared amongst species, but a trueorthologous relationship has only been found within simiansuborders such as Catarrhini (humans, apes and OWM) andPlatyrrhini, NWM and within Prosimians (lemurs andrelatives; see O’hUigin 1995; Kriener et al. 2000a,b,2001; Go et al. 2002; Trtkova et al. 1995). This contrastswith other MHC-II regions (DP, DQ, DO and DM)exhibiting great conservation amongst primates (thesebeing genuinely orthologous) and having similar chromo-some organisation (Satta et al. 1996).
Several New World monkey species are currently beingemployed in biomedical research, Aotus monkeys being animportant representative. They can predictably develophuman malaria, particularly Plasmodium falciparum (Collins1994; Gysin 1988; Rodriguez et al. 1990) and Plasmodiumvivax asexual/blood stage infections (Pico de Coaña et al.2003). Previous work concerning MHC-DRB has revealedthe presence of at least six different allele lineages (Niño-Vasquez et al. 2000). None of these has exhibited sequenceshaving stop codons or any kind of sequence alteration(insertions or deletions) at exon 2 level, but they haveexhibited great similarity with several HLA-DRB lineages.
Ninety-six Aotus monkeys (71 Aotus nancymaae, 15Aotus nigriceps and 10 Aotus vociferans) were sequencedin this work to establish whether they possessed allelesdifferent to those reported so far. Human and Aotus monkeyMHC-DRB characteristics were then compared. Analysiswas focused on variability patterns within the MHC-DRBβ-domain (exon 2 encoded). A phylogenetic approach wastherefore used for defining the main groups of sequences inthe Aotus species considered here and the variability soanalysed, focusing on the number and type of changesinvolved in the protein sequences. The extent and nature ofselection in these Aotus sequences by comparing them withHLA-DRB and MHC-DRB molecule sequences from otherprimates. Emphasis was placed on those residues involvedin PBR conformation for further analysis of this model’ssuitability for evaluating vaccine candidates for human use.
Materials and methods
Animals
The owl monkeys analysed in this study were caught in theAmazon area close to Leticia (Colombia) and selected atrandom to ensure the presence of a representative repertoireof different alleles. Mononuclear cells were isolated by
density gradient separation of peripheral blood obtained byvenous puncture (Garraud et al. 1994). The blood came from96 healthy monkeys from three different Aotus species: 71 A.nancymaae (29 male, 42 female), 15 A. nigriceps (six male,nine female) and 10 A. vociferans (six male, four female).
RNA extraction, polymerase chain reaction, DNA cloningand sequencing
Total cellular RNA was isolated from peripheral bloodmononuclear cells using the TRIzol one-step procedure(Invitrogen Life Technologies, CA, USA). Moloney MurineLeukemia Virus (Promega, Madison, WI, USA) was used forcDNA synthesis, according to the Manufacturer’s instructions.
cDNA samples were amplified by polymerase chainreaction (PCR) in 100-μl reaction mixtures containing5 mM MgCl2, 10 mM of each of the four dNTPs, 0.2 μMof each primer, 1× PCR buffer and 6 U of taq polymerase(Promega). A Perkin Elmer 9700 thermocycler was used forthe amplifications, using the following profile: 28 cycles at96°C, 58°C and 72°C each for 1 min, followed by 1 cycleof 72°C for 5 min. Sense (5′-GGTGCTGAGCTCCCGACTGGC-3′) and anti-sense (5′-AGGTTGTGGTGCTGCAGGGG-3′) MHC-DRB primers were used for amplify-ing exon 2, as previously described (Niño-Vasquez et al.2000). PCR products were purified with Wizard PCR prepspurification kit (Promega) and cloned into pGEM-T Easyvector (Promega). An ABI PRISM 310 genetic analyser(Perkin Elmer) was used for double-strand plasmid DNAsequencing; ABI PRISM 310 sequencing analysis 3.3 andMT Navigator 1.0.2.b3 software were used for analysingthe data. The reported alleles represented at least twoidentical clones obtained by screening 5 to 22 randomlychosen colonies from the same animal.
Sequence nomenclature
The sequences obtained were reported to the Immuno-genetics’ database at EBI http://www.ebi.ac.uk) and werenamed here according to the rules for standardised MHCnomenclature according to Klein et al. (1990).
Sequence analysis
A. nancymaae (Aona), A. nigriceps (Aoni) and A. vocifer-ans (Aovo) MHC-DRB sequences were analysed (Gen-Bank accession numbers AY563180–AY563263,DQ162624–DQ162736, respectively), together with a rep-resentative sample of simian sequences, i.e. Hominidae(human and apes), Cercopithecidae (OWM) and Platyrrhini(NWM) from the GenBank database for which completeexon 2 MHC-DRB data is available. Human sequences wereobtained from the Anthony Nolan institute (Marsh 1999,
Immunogenetics (2006) 58:542–558 543

http://www.ebi.ac.uk/imgt/hla/); only non-identical nucleotidesequences were used. A sample of DP and DQ sequences wasused for providing out-groups (one from each primatespecies). A total of 820 sequences (including Aotus MHC-DRB) were used. The simian species considered were(Ceae) Cercopithecus aethiops, (Maar) Macaca arctoides,(Mafa)Macaca fascicularis, (Mamu)Macaca mulatta, (Mane)Macaca nemestrina, (Masi) Macaca silenus, and (Paca) Papiocynocephalus anubis for OWM; (Aotr) Aotus trivirgatus,(Aona) A. nancymaae, (Aoni) A. nigriceps, (Aovo) A.vociferans, (Aoaz) Aotus azarae, (Camo) Callicebus moloch,(Caja) Callithrix jacchus, (Ceap) Cebus apella, (Saoe)Saguinus oedipus, (Sasc) Saimiri sciureus, (Pipi) Pitheciapithecia, (Capy) Callithrix pygmaea, and (Atbe) Atelesbelzebuth for NWM; and (HLA) Homo sapiens, (Gogo) Gorillagorilla and (Patr) Pan troglodytes for human and apes.
Clustal X software (Thompson et al. 1994) was used foraligning sequences; 267/89 nucleotide/amino acid positionswere analysed altogether. Phylogenetic trees were con-structed with MEGA-2.1 software (Kumar et al. 2001),using minimum-evolution (ME) and neighbour-joining (NJ)distance methods (Rzhetsky and Nei 1993). Geneticdistances were estimated by using P distance, Jukes–Cantor(Jukes 1969) and Kimura 2-parameter (Kimura 1980)methods for nucleotide sequences, and gamma and Poissoncorrected distances for amino acid sequences. Bootstrapanalysis (Hillis 1993) and internal branch test (Sitnikova1996; with 1,000 replicates for each) were used forassigning confidence levels to branch nodes.
MEGA 2.1 software was used for calculating meandifferences between and within groups (nucleotide andamino acids). Standard errors were assessed by the boot-strap method with 1,000 replicates.
MEGA 2.1 software was used for calculating synonymousand non-synonymous substitutions and associated variancerates (assessed by the bootstrap method with 1,000 repli-cates) by Nei–Gojobori’s method (Nei and Gojobori 1986).
Mismatch distribution analysis (Go et al. 2002; Figueroaet al. 2000; Di Rienzo and Wilson 1991) detectedconvergence in Aotus species MHC-DRB. Gene conversionwas estimated using the method proposed by Sawyer(1989) using GENECONV software (Sawyer 1999); thisperforms statistical procedures for aligning possible geneconversion events and provides multiple-comparison cor-rected P values for each instance.
Each position’s variation for a given set of amino-acidaligned human and Aotus MHC-DRB pockets was repre-sented by a one-letter code in its respective position withinthe sequence. Conservative and non-conservative substitu-tion could thus be determined for each position wherevariation in an amino acid symbol’s colour indicated non-conservative changes; maintaining the colour representedconservative changes based on PAM 250 substitution
matrix parameter groups (DENQH, SAT, KR, FYW, LIVM,C, G and P; see Dayhoff et al. 1978).
Results and discussion
Isolated sequences
The MHC-DRB exon 2 from 96 Aotus monkeys (71 A.nancymaae, 10 A. vociferans and 15 A. nigriceps) wasamplified by cDNA PCR. Seven hundred ninety-sevenclones were sequenced, and 67 different sequences wereobserved for A. nancymaae, 30 for A. nigriceps and 13sequences for A. vociferans. Three identical sequences werefound in previous a work on A. nancymaae (Aona-DRB3*0612, Aona-DRB*W1802 and Aona-DRB*W1302;Niño-Vasquez et al. 2000).
The Aotus MHC-DRB sequences reported in this paperwere divided into 12 lineages according to the Immuno-genetics’ database nomenclature; MHC-DRB1*03 wassubdivided into three groups of sequences (GA, GB, GC)and MHC-DRB3*06 into two groups (GA, GB) havingclearly distinctive characteristics. The most frequentlyfound alleles in the 96 owl monkeys analysed here camewithin the MHC-DRB*W13 group (23%), followed byDRB1*03 (18%), DRB*W47 (15%), DRB3*06 (12%),DRB*W29 (11%), DRB*W43 (5%), DRB*W18 (5%),DRB*W38 (3%), DRB*W42 (3%), DRB*W44 (2%),DRB*W46 (2%) and DRB*W45 (1%). The most variablegroups were DRB*W43 (mean 9.9±1.7 nt), DRB*W18(mean 6.4±1.5) and DRB*W13 (mean 5.9±1.2 nt).
Figure 1 shows the MHC-DRB exon 2 deduced proteinalignment, from groups of Aotus and some alleles from theirhuman counterparts’ lineages. The sequences were groupedinto nine blocks based on the most representative AotusDRB* lineages found (see names in the left-hand column).Each block was based on Aotus amino-acid sequencesimilarity with the closest HLA-DRB* homologue (HLA-DRB1*-0403: -0422; -0301; -0701; -0801; -1001; -1130-150101 and HLA-DRB3*-0201) and are highlighted inyellow. Amino acids shown in colour in the figure indicatethe residues forming a particular pocket (Kaufman et al.
�Fig. 1 Aligning Aotus MHC-DRB deduced amino acid sequencesfrom exon 2. One hundred ten amino acid sequences were obtainedafter sequencing 797 recombinant colonies randomly screened from96 Aotus (71 Aona, 10 Aovo and 15 Aoni). Sequences previouslyreported by Niño-Vasquez et al. 2000 (21) are preceded by a dot (∙).The sequence names are provided in the left-handcolumn. Each blockwas based on Aotus amino acid sequence similarity with the closestHLA-DRB* homologue. Dashes correspond to amino-acid identity.Colouredpositions denote amino acid constituted pockets at the PBR:fuchsia P1, blue P4, ochre P6, grey P7, green P9. Secondary structuralfeatures are shown at the bottom of the figure: α1 and α2 alphahelices, β1 to β3 beta sheets, T1 to T5 turns, RC random coil
544 Immunogenetics (2006) 58:542–558

10 20 30 40 50 60 70 80 90
HLA-DRββ 1*0403 F L E Q V K H E C H F F N G T E R V R F L D R Y F Y H Q E E Y V R F D S D V G E Y R A V T E L G R P D A E Y W N S Q K D L L E Q R R A E V D T Y C R H N Y G V V E S F T V Q R RAona DRB*W4701 - - - - - - - - - - - - - - - - - - - - - - - - - - - N - - - - - - - - - - - - - F - - - - - - - - - - - - - - - - - - - - I Y Y - - - Q - - - - - - - - - - - - - - - - - - - -Aona DRB*W4702 - - - - - - - - - - - - - - - - - - - - - - - - - - - N - - - - - - - - - - - - - F - - - - - - - - - - - - - - - - - - - - M - D - - P Q - - - - - - - - - - - - - - - - - - - -Aona DRB*W4703 - - - - - - - - - - - - - - - - - - - - - - - - - - - N - - - - - - - - - - - - - F - - - - - - - - - - - - - - - - - - - - M - D - - - Q - - - - - - - - - - - - - - - - - - - -Aona DRB*W470401 - - - - - - - - - - - - - - - - - - - - - - - - - - - N - - - - - - - - - - - - - F - - - - - - - - - - - - - - - - - - - - M D - - - Q - - - - - - - - - - - G - - - - - - - -Aona DRB*W470402 S - - - - - - - - - - - - - - - - - - - - - - - - - - N - - - - - - - - - - - - - F - - - - - - - - - - - - - - - - - - - - M - D - - - Q - - - - - - - - - - - G - - - - - - - -Aona DRB*W470403 - - - - - - - - - - - - - - - - - - - - - - - - - - - N - - - - - - - - - - - - - F - - - - - - - - - - - - - - - - - - - - M D - - - Q - - - - - - - - - - - G - - - - - - - -Aona DRB*W470404 - - - - - - - - - - - - - - - - - - - - - - - - - - - N - - - - - - - - - - - - - F - - - - - - - - - - - - - - - - - - - - M - D - - - Q - - - - - - - - - - - G - - - - - - - -Aona DRB*W4705 - - - - - - - - - - - - - - - - - - - - - - - - - - - N - - - - - - - - - - - - - F - - - - - - - - - - - G H - - - - - - - M - D - - - Q - - - - - - - - - - - G - - - - - - - -Aona DRB*W4706 - - - - - - - - - - - - - - - - - - - - - - - - - - - N - - - - - - - G - - - - - F - - - - - - - - - - - - - - - - - - - - M - D - - - Q - - - - - - - - - G - - - - - - - -Aona DRB*W4707 - - - - - - - - - - - - - S - - - - - - - - - - - - - N - - - - - - - - - - - - - F - - - - - - - - - - - - - - - - - - - - M - D - - - Q - - - - - - - - - - - G - - - - - - - -Aona DRB*W4708 - - - K - - - - - - - - - - - - - - - - - - - - - - - N - - - - - - - - - - - - - F - - - - - - - - - - - - - - - - - - - - M - D - - - Q - - - - - - - - - - - G - - - - - - - -Aona DRB*W4709 - - - - - - - D - - - - - - - - - - - - - - - - - - - N - - - - - - - - - - - - - F - - - - - - - - - - - - - - - - - - - - M - D - - - Q - - - - - - - - - - - G - - - - - - - -Aona DRB*W4710 - - - - - - - - - - - - - - - - - - E - - - - - - - - N - - - - - - - - - - - - - F - - - - - - - - - - - - - - - - - - - - M - D - - - Q - - - - - - - - - - - G - - - - - - - -Aona DRB*W4711 - - - - Q V K H - - H - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - R - - - - - - - - - - - - - E - - - - - - - - - - Y - - - - A - - - - - - - -Aoni DRB*W4701 - - - - - - - - - - - - - - - - - - - Q - - - - - - - N - - - - A - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - D - - - Q - - S - - - - - - - - G - - - - E - - -Aovo DRB*W4701 - - - - - - - - - - - - - - - - - - - - - - - - - - - N - - - - - - - - - - - - - F - - - - - - - - - - - - - - - - - - - - M - D - - - Q - - - - - - - - - - - G - - - - - - - -Aovo DRB*W4702 - - - - - - - - - - - - - - - - - - - Q - - - - - - - N - G - - A - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - D - - - Q - - S - - - - - - - - G - - - - E - - -Aona DRB*W4501 - - - - - - - - - - - - - - - - - - - - - - - - - I H N - - - V - - - - - - - - - - - - - - - - - - - - - - - - - - L - - I - - D - - - Q - - - - - - - - - - - - - - - - - - - -Aona DRB*W4601 - - - - - F - P - - - - - - - - - - - - - - - - - - - N - - - - - - - - - - - - - F - - - - - - - - - Y - - - - - - L - - I - - D - - - Q - - - - - - - - - - - - - - - - - - - -Aona DRB*W4602 - - - - - F - P - - - - - - - - - - - - - - - - - - - N - - - - - - - - - - - R - F - - - - - - - - - - - - - - - - - - - - M - D - - - Q - - - - - - - - - - - G - - - - - - - -HLA DRβ 1*1122 - - - - - - - - - - - - - - - - - - - - - - - - - - - N - - - - - - - - - - - - - F - - - - - - - - - - E - - - - - - - - - F - - D - - - A - - - - - - - - - - - G - - - - - - - -
HLA-DRβ 1*0422 * * L E Q V K H E C H F F N G T E R V R F L D R Y F Y H Q E E Y V R F D S D V G E Y R A V T E L G R P D A E Y W N S Q K D L L E Q K R G R V D N Y C R H N Y G V V E S F T V Q R R• Aona DRB3*0601 R F - - L - - - - - - - - - - - - - - - Y - - - - I H - - - - V - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - F - - - - - - - -• Aona DRB3*0602 R F - - L - - - - - - - - - - - - - - - Y - - - - L - - - K - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y V - - - - - Q - - - - - - - - - - - F - - - - - - - -• Aona DRB3*0603 R F - - L - - - - - - - - - - - - - - - Y - - - - - - - - K - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - - - - - - - - - -• Aona DRB3*0604 R F - - L - - - - - - - - - - - - - - - Y - - - - - - - - K - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - G - - - - - - - -• Aona DRB3*0605 R F - - L - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - G - - - - - - - -• Aona DRB3*0606 R F - - L - - - - - - - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - G - - - - - - - -• Aona DRB3*0607 R F - - L - - - - - - - - - - - - - - - Y - - - - - - - - K - S - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y V - - - - - Q - - - - - - - - - - - F - - - - - - - -• Aona DRB3*0612 R F - - L - - - - - - - - - - - - - - - Y - - - - L - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y V - - - - - Q - - - - - - - - - - - F - - - - - - - -Aona DRB3*0613 R F - - L - - - - - - - - - - - - - - - Y - - - - I H - - - - V - - - - - - - - - - - - - - - - - - - - - - - - S - - - - Y V - - - - - - - - - - - - - - - - - - - - - - - - - -Aona DRB3*0614 R L - - L - - - - - - - - - - - - - - - Y - - - - I H - - - - V - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - - - - - - - - - -Aona DRB3*0615 R F - - L - - - - - - - - - - - - - - - Y - - - - I H - - - - V - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - - - - - - - - - -Aona DRB3*0616 R F - - L - - - - - - - - - - - - - - - Y - - - - T H - - - - V - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y V - - - - - Q - - - - - - - - - - - F - - - - - - - -Aona DRB3*0617 R F - - L - - - - - - - - - - - - - - - Y - - - - I H - - - - V - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y V - - - - - Q - - - - - - - - - - - F - - - - - - - -Aona DRB3*0618 R F - - L - - - - - - - - - - - - - - - Y - Y - - L - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y V - - - - - Q - - - - - - - - - - - F - - - - - - - -Aona DRB3*0619 R F - - L - - - - - - - - - - - - - - - Y - - - - - - - - K - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - - - - - - - - - -Aona DRB3*0620 R F - - - - - - V - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - - - - - - - - - -Aona DRB3*0621 R F - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - A - - - - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - G - - - - - - - -Aona DRB3*0622 R F - - - A - - - - - - - - - - - - - - Y - - - - - - - - K - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - G - - - - - - - -Aona DRB3*0623 R F - - L - - - - - - - - - - - - - - - Y - - - - S - - - K - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - - - - - - - - - -Aona DRB3*0624 R F - - L - - - - - - - - - - - - - - - Y - - - - S - - - K - - - - - - - - - - - - W - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - Y - - - - - - G - - - - - -Aoni DRB3*0601 R F - - L - - - - - - - - - - - - - - - Y - - - - L - - - K - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y V - - - - - Q - - - - - - - - - - - - - - - - - - - -HLA-DRβ 3*0201 R F - - L L - S - - - - - - - - - - - - - - E - H - H N - - - - A - - - - - - - - - - - - R - - - - - - - - - - - - - - - - - - - - - - Q - - - - - - - - - - - - - - - - - - - -
HLA-DRβ 1*0301 R F L E Y S T S E C H F F N G T E R V R Y L D R Y F H N Q E E N V R F D S D V G E F R A V T E L G R P D A E Y W N S Q K D L L E Q K R G R V D N Y C R H N Y G V V E S F T V Q R R• Aona DRB1*0303 - - - F Q T - - - - - - - - - - - - - - - - - - - - Y - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - - - - - Q - - - - - - - - - - - F - - - - - - - -• Aona DRB1*0304 - - - F Q T - - - - - - - - - - - - - - F - - - - - Y - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - G - - - - - - - -• Aona DRB1*0305 - - - F Q T - - - - - - - - - - - - - - - - - - - - Y - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - G - - - - - - - -• Aona DRB1*0307 - - - F Q T - - - - Y - - - - - - - - - - - - - - - Y - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - R - - - - - - - - - - - - - - - G - - - - - - - -• Aona DRB1*0309 - - - F Q T - - - - Y - - - - - - - - - F - - - - - Y - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - R - - - Q - - - - - - - - - - - G - - - - - - - -• Aona DRB1*0311 - - - F Q T - - - - - - - - - - - - - - F - - - - - Y - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - - - - - - - - - -• Aona DRB1*0312 - - - F Q T - - - - - - - - - - - - - - - - - - - - Y - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - - - - - - - - - -• Aona DRB1*0316 - - - F Q T - - - - - - - - - - - - - - - - - - - - Y - - K - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - G - - - - - - - -Aona DRB1*031701 T - - V Q T - - - - - - - - - - - - - - - - - - - - Y - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - G - - - - - - - -Aona DRB1*031702 - - - F Q T - - - - - - - - - - - - - - - - - - - - Y - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - G - - - - - - - -Aona DRB1*0318 - - - F Q T - - - - - - - - - - K - - - - - - - - - Y - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - G - - - - - - - -Aona DRB1*0319 - - - F Q T - - - - - - - - - - - - - - H - - - - - Y - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - G - - - - - - - -Aoni DRB1*0303 - - - F Q T - - - - Y - - - - - - - - - - - - - - - Y - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - R - - - - - - - - - - - - - - - G - - - - - - - -Aoni DRB1*0304 - - - F Q T - - - - - - - - - - - - - - - - - - - - Y - - K - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - G - - - - - - - -Aoni DRB1*0306 - - F Q T - - - - - - - - - - - - - - - - - - - - Y - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - G - - - - - - - -Aoni DRB1*0307 - - - F Q T - - - - - - - - - - - - - - H - - - - - Y - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - G - - - - - - - -Aovo DRB1*0301 - - - F Q T - - - - - - - - - - - - - - - - - - - - Y - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - R - - - Q - - - - - - - - - - - G - - - - - - - -Aovo DRB1*0302 - - - F Q T - - - - - - - - - - - - - - - - - - - - Y - - K - Y - - - A - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - G - - - - - - - -Aovo DRB1*0303 - - - F Q T - - - - - - - - - - - - - - - - H - - - Y - - K - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - G - - - - - - - -Aovo DRB1*0304 - - - F Q T - - - - - - - - - - - - - - - - - - - - Y - - K - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - Y V - - - - - - - - - - - - - - - - - G - - - - - - - -
HLA-DRβ 1*0701 R F L W Q G K Y K C H F F N G T E R V Q F L E R L F Y N Q E E F V R F D S D V G E Y R A V T E L G R P V A E S W N S Q K D I L E D R R G Q V D T V C R H N Y G V G E S F T V Q R RAona DRB*W3801 - - - E - A - - E - - - L - - - - - - R - - - - H I H - - - - Y A - - - - - - - - - - - - - - - - - R T - - Y - - - - - - - - - - - - A - - - - - - - - - - - - - - - - - - - - -Aona DRB*W3802 - - - E - A - - E - - - L - - - - - - R - - - - H I H - - - - Y A - - - - - - - - - - - - - - - - - R T - - Y - - - - - - - - - V - - A - - - - - - - - - - - - - - - - - - - - -Aoni DRB*W3801 - - - E - A - - E - - - L - - - - - - R - - - - H I H - - - - Y A - - - - - - - - - - - - - - - - - R T - - Y - - - - - - F - - - - - A - - - - - - - - - - - - V - - - - - - - -• Aona DRB*W3001 - - - E - V - - E - - - L - - - - - - R Y - - - - I - - R - - - - - - - - - - - - - - - - - - - - - - S - - K L - - - - - - - - - - - A S - - - Y - K - - - - - - - - - - - - - -HLA-DRβ 1*090102 - - - K - D - F E - - - - - - - - - - R Y H - G I - - - - - - V - - - - - - - - - - - - - - - - - - - - - - - - - - - F - - R - - A E - - - - - - - - - - - - - - - - - - -
HLA-DRβ 1*0801 R F L E Y S T G E C Y F F N G T E R V R F L D R Y F Y N Q E E Y V R F D S D V G E Y R A V T E L G R P S A E Y W N S Q K D F L E D R R A L V D T Y C R H N Y G V G E S F T V Q R R• Aona DRB1*0301 - - - F Q T - S - - H - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - - - - - - - - - - - R - - - - - - - - - - - - - E - - - - - - - - - - Y - - - - A - - - - - - - -• Aona DRB1*0302 - - - F Q T - S - - H - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - R - - - - - - - - - - - - - E - - - - - - - - - - Y - - - - A - - - - - - - -• Aona DRB1*0308 - - - F Q T - S - - - - - - - - - - - - Y - - - - - - - - K - - - - - - - - - - - - - - - - - - - - R - - - - - - - - - - - - - E - - - - - - - - - - Y - - - - A - - - - - - - -Aona DRB1*0320 - - - F Q T - S - - H - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - R - - - - - - - - - - - - - E - - - - - - - - - - Y - - - - A - - - - - - - -Aona DRB1*0321 - - - F Q T - S - - H - - - - - G - - - - - - - - - - - - - - Y - - - - - - - - - - - - - - - - - - R - - - - - - - - - - - - - E - - - - - - - - - - Y - - - - A - - - - - - - -Aona DRB1*0322 - - - F Q T - S - - H - - - - - G - - - - - - - - - - - - - - - - - - - C - - - - - - - - - - - - - R - - - - - - - - - - - - - E - - - - - - - - - - Y - - - - A - - - - - - - -Aona DRB1*0323 - - - F Q T - S - - H - - - - - G - - - - - - - - - - - - - - - - - - - - G - - - - - - - - - - - - R - - - - - - - - - - - - - E - - - - - - - - - - Y - - - - A - - - - - - - -Aona DRB1*0324 - - - F Q T - S - - H - - - - - G - - - - - - - - - - - - - - - - - - - - H - - - - - - - - - - - - R - - - - - - - - - - - - - E - - - - - - - - - - Y - - - - A - - - - - - - -Aona DRB1*0325 - - - F Q T - S - - H - - - - - G - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - L - - - - - - - - - - - - - E - - - - - - - - - - Y - - - - A - - - - - - - -Aona DRB1*0326 - - - F Q T - Y - - H - - - - - G - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - R - - - - - - - - - - - - - E - - - - - - - - - - Y - - - - A - - - - - - - -
HLA DRB1*150101 R F L W Q P K R E C H F F N G T E R V R F L D R Y F Y N Q E E S V R F D S D V G E F R A V T E L G R P D A E Y W N S Q K D I L E Q A R A A V D T Y C R H N Y G V V E S F T V Q R RAoni DRB1*0305 - - - F Q T - S - - Y - - - - - - - - - Y - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - F - - L - - F M - E T - - A - - - - - - - - - - - - - - - - - - - -• Aona DRB1*0313 - - - F Q T - S - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - E - - - F - - L - - F M - E T - - - - - - - - K - - - - - - - - - - - - - -• Aona DRB1*0314 - - - F Q T - S - - - - - - - - - - - - Y - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - E - - - F - - L - - F M - E T - - - - - - - - K - - - - - - - - - - - - - -Aona DRB1*0327 - - - F Q T - S - - - - - - - - - - - - Y - - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - - - - - - - - - - - L M - D R - - Q - - - - - - - - - - - G - - - - - - - -HLA DRB1*160101 - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - F - - D R - - - - - - - - - - - - - G - - - - - - - -
HLA-DRβ 3*0201 R F L E L L K S E C H F F N G T E R V R F L E R H F H N Q E E Y A R F D S D V G E Y R A V R E L G R P D A E Y W N S Q K D L L E Q K R G Q V D N Y C R H N Y G V V E S F T V Q R R• Aona DRB*W1801 - - - - Q V - - - - Y - - - - - - - - Q - - - - Y - - - - - - - V - - - - - - - - - - - - T - - - - - - - K - - - G - Q - I - - L - - - - - - - - - - - - - - - G - - - - - - - -• Aona DRB*W1802 - - - - - V - - - - Y - - - - - - - - Q - - - - Y - - - - - - L V - - - - - - - - - - - - T - - - - - - - K - - - G - Q - I - - L - - - - - - - - - - - - - - - F - - - - - - - -Aona DRB*W1803 - - - - Q V - - - - Y - - - - - - - - Q - - - - Y - - - - - - F V - - - - - - - - - - - - T - - - - - - - K - - - G - Q - I - - L - - - - - - - - - - - - - - - - D - - - - - - -Aona DRB*W1804 - - - - - V - - - - Y - - - - - - - - Q - - - - Y - - - - - - L - - - - - - - - - - - - - T - - - - - - - K - - - G - Q - I - - L - - - - - - - - - - - - - - - F - - - - - - - -Aona DRB*W1805 - - - - - V - - - - Y - - - - - - - - Q - - - - Y - - - - - - L V - - - - - - - - - - - - T - - - - - - - K - - - G - Q - I - - L - - - - - - - - - - - - - - - F - - - - L - - -Aona DRB*W1806 - - - - Q V - - - - Y - - - - - - - - Q - - - - Y - - - - - - F V - - - - - - - - - - - - T - - - - - - - K - - - G - Q - I - - L - - - - - - - - - - - - - - - G - - - - - - - -Aona DRB*W1807 - - - - Q V - - - - Y - - - - - - - - Q - - - - Y - - - - - - F V - - - - - - - - - - - - T - - - - - - - K - - - G - Q - I - - L - - - - - - - - - - - - - - Q R K G - - - - - -Aoni DRB*W1801 - - - - Q V - - - - Y - - - - - - - - Q - - - - Y - - - - - - F V - - - - - - - - - - - - T - - - - - - - K - - - G - Q - I - - L - - - - - - - - - - - - - - - G - - - - - - - -• Aona DRB*W2902 - - - - Q T - - - - - - L - - - - - - - L - Q - Y - Y - - - - - - - - - - - - - - F - - - T - - - - - - - - - - - - - - - Y V - - - - - R - - - - - - - - - - - G - - - - - - - -• Aona DRB*W2903 - - - - Q T - - - - - - L - - - - - - - L - Q - Y - - - - - - L V - - - - - - - - - - - - T - - - - - - - K - - - G - Q - I - - L - - - - - - - - - - - - - - - F - - - - - - - -HLA DRB1*0422 - - - - Q V H E C - - - - - - - - - - D Y - Y H - - - V - - - - - - - - - - - T - - - - - - - - - - - - - - - - - - - - R - - - - - - V E S F T V Q R R
HLA DRB1*150101 R F L W Q P K R E C H F F N G T E R V R F L D R Y F Y N Q E E S V R F D S D V G E F R A V T E L G R P D A E Y W N S Q K D I L E Q A R A A V D T Y C R H N Y G V V E S F T V Q R R• Aona DRB*W2901 - - - E E T - S - - - - L - - - - - - - L - Q - - - - - - - - Y A - - - - - - - - - - - - - - - - - - - - - - - - - L - E R - - Y L - - - - - - - - - - - - - - F - - - - - - - -Aona DRB*W2904 - - - E E T - S - - - - L - - - - - - - L - Q - - - - S - - - Y A - - - - - - - - - - - - - - P - - - - - - - - - - L - E R - - Y L - - - - - - - - - - - - - - F - - - - - - - -Aona DRB*W2905 - - - E E T - S - - - - L - - - - - - - L - Q - - - - - - - - Y A - - - - - - - - - - - - - - - - - - - - - - - - - L - E R - - Y L - - - - - - - - - - - - - - F - - - - - - - -Aona DRB*W2906 - - - E E T - S - - - - L - - - - - - - L - Q - - - - - - - - Y A - - - - - - - - - - - - - - - - - - - - - - - - - L - E R - - Y L - - - - - - - - - - - - - - V - - - - - - - -Aona DRB*W2907 - - - E E T - S - - - - L - - - - - - - L - Q - - - - - - - - Y A - - - - - - - - - - - - - - - - - - - - - - - - - L - E R - - Y L - - - - - - - - - - - - - - - - - - A - - - -Aona DRB*W2908 - - - E E T - S - - - - L - - - - - - - L - Q - - - - - - - - Y A - - - - - - - - - - - - - - - - - - - - - - - - - L - E R - - Y L - - - - - - - - - - - - - - - - - - - - - - -Aona DRB*W2909 - - - E E T - S - - - - - T - - - - - - L - Q - - - - - - - - Y A - - - - - - - - - - - - - - - - - - - - - - - - - L - E R - - Y L - - - - - - - - - - - - - - V - G - - - - - -Aoni DRB*W2901 - - - E E T - S - - - - L - - - - - - - L - Q - - - - - - - - Y A - - - - - - - - - - - - - - - - - - - - - - - - - L - E R - - Y L - - - - - - - - - - - - - - - - - - - - - - -Aoni DRB*W2903 - - - E E T - S - - - - L - - - - - - - L - Q - - - - - - - - Y - - - - - - - - - - - - - - - - - - - - - - - - - - L - E R - - Y L - - - - - - C - - - - - - - V - - - - - - - -Aoni DRB*W290402 - - - E E T - S - - - - L - - - - - - - L - Q - - - - - - - - Y - - - - - - - - - - - - - - - - - - - - - - - - - - L - E R - - Y L - - - - - - C - - - - - - - V - - - - - - - -Aoni DRB*W2902 - - - E E T - S - - - - L - - - - - - - L - Q - - - - - - - - Y - - - V C - - - - - - - - - - - - - - - - - - - - - L - E R - - Y L - - - - - - C - - - - - - - V - - - - - - - -Aovo DRB*W2901 - - - E E T - S - - - - L - - - - - - - L - Q - - - - - - - - Y - - - - - - - - - - - - - - - - - - - - - - - - - - L - E R - - Y L - - - - - - C - - - - - - - V - - - - - - - -• Aona DRB3*0608 - - - E L V - H - - - - - - - - - - - - Y - D - - - - - - K - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - L - E R - - Y L - - - - - - - - - - - - - - F - - - - - - - -• Aona DRB3*0609 - - - E L V - H - - - - - - - - - - - - Y - D G - I H - - - - Y A - - - - - - - - - - - - - - - - - - - - - - - - - L - E R - - Y L - - - - - - - - - - - - - - V - - - - - - - -• Aona DRB3*0610 - - - E L V - H - - - - - - - - - - - - Y - D - - - - - - K - Y A - - - - - - - - - - - - - - - - - - - - - - - - - L - E R - - Y L - - - - - - - - - - - - - - F - - - - - - - -• Aona DRB1*0316 - - - F E T T S - - - - - - - - - - - - Y - D - - - - - - - - Y A - - - - - - - - - - - - - - - - - - - - - - - - - L - E R - - Y L - - - - - - - - - - - - - - F - - - - - - - -Aona DRB*W4201 - - - E E V - D - - - - L - - - - - - - F - E - - - - - - - - Y - - - - - - - - - - - - - - - - - - - - - - - - - - L - E R - - Y L - - - - - - - - K - - - - - V - - - - E - - -Aoni DRB*W4201 - - - E E V - D - - - - L - - - - - - - F - E - - - - - - - - Y - - - - - - - - - - - - - - - - - - - - - - - - - - L - E R - - Y L - - - - - - - - K - - - - - V - - - - - - - -HLA DRB1*160101 - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - F - - D R - - - - - - - - - - - - - - G - - - - - - - -
HLA-DR 1*1001 R F L E E V K F E C H F F N G T E R V R L L E R R V H N Q E E Y A R Y D S D V G E Y R A V T E L G R P D A E Y W N S Q K D L L E Q R R A A V D T Y C R H N Y G V G E S F T V Q R RAona DRB*W4101 - - - K Q - - C - - - - - - - - - - - Q - - D - H F Y - - - - - - - F - - - - - - - - - - - - - - - - - - - - F - - L - - F M - E T - - - - - - - - K - - - - - F - - - - - - - -Aoni DRB*W4301 - - - K Q - - C - - - - - - - - - - - Q - - D - H F Y - - - - - - - F - - - - - - - - - - - - - - - R T - - - - - - - - - F - - N - - - - - - - - - K - - - - - - - - - - - - - -Aoni DRB*W4302 - - - K Q - - C - - - - - - - - - - - Q - - D - H F Y - - - - - - - S - - - - - - - - - - - - - - - R T - - - - - - - - - F - - N - - - - - - - - - K - - - - - - - - - - - - - -Aoni DRB*W4303 - - - K Q - - C - - - - - - - - - - - Q - - D - H F Y - - - - - - - F - - - - - - - - - - - - - - - R T - - - - - - - - - F - - N - - - - - - - - - K - - - - - - - - L - - - - -Aoni DRB*W4305 - - - K Q - - C - - - - - - - - - - - Q - - D - H F Y - - - - V R - F - - - - - - - - - - - - - - - Q - R K - - - - - - - F - - N - - - - - - - - - K - - - - - - - - S - E - - -Aoni DRB*W4304 - - - K Q - - C - - - - - - - - - - - Q - - D - H F Y - - - - - - - F - - - E - - - - - E - - - - - R T - - - - - - - - - Y V - N - - - - E - - - R K - - - - - V - D - - - - - -Aovo DRB*W4301 - - - K Q - - C - - - - - - - - - - - Q - - D - H F Y - - - - - - - F - - - - - - - - - - - - - - - R T - - - - - - - - - F - - N - - - - - - - - - K - - - - - - - - - - - - - -HLA DRB4*0101 - - - - Q A - C - - - - L - - - - - - W N I Y I Y - - - - - - - - N - - L - - - Q - - - - - - - - - - - - - - - - - - - - - R - - - E - - - - - - Y - - - - - - - - - - - - -
HLA DRB1*150101 R F L W Q P K R E C H F F N G T E R V R F L D R Y F Y N Q E E S V R F D S D V G E F R A V T E L G R P D A E Y W N S Q K D I L E Q A R A A V D T Y C R H N Y G V V E S F T V Q R R• Aona DRB*W1302 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - F - - L - - F M - E T - - - - - - - - - - - - - - - - - - - - - - -Aona DRB*W1303 - - - E - F - L - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - E - - - F - - L - - F M - E T - - - - - - - - - - - - - - - - - - - - - - -Aona DRB*W1304 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - F - - L - - F M - E T - - - - - - - - - - - - - - - - - - A - - - -Aona DRB*W1305 - - - E - F - P - - - - - - - M - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - F - - L - - F M - E T - - - - - - - - - - - - - - - - - - - - - - -Aona DRB*W1306 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - F - - L - - F I - E T - - - - - - - - - - - - - - - - - - - - - - -Aona DRB*W1307 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - L - - L - - F M - E T - - - - - - - - - - - - - - - - - - - - - - -Aona DRB*W1308 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - E - - - F - - L - - F M - E T - - - - - - - - - - - - - - A - - - - - - - -Aona DRB*W1309 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - E - - - F - - L - - F M - E T - - - M - - - - - - - - - - A - - - - - - - -Aona DRB*W1310 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - F - - L - - F M - E T - - - - - - - - - - - - - - - - - - - - - - -Aona DRB*W1311 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - E - - - F - - L - - F M - E T - - - - - - - - K - - - - - - - - - - - - - -Aona DRB*W1312 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - F - - - - - - - - - Y - - - - - - - - R T - - - F - - L - - F M - E T - - - - - - - - K - - - - - - - - - - - - - -Aona DRB*W1313 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - P - - - - - - - E - - - F - - L - - F M - E T - - - - - - - - - - - - - - A - - - - - - - -Aoni DRB*W1301 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - F - - L - - F I - E T - - - - - - - - - - - - - - - - - - - - - - -Aoni DRB*W1302 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - - - - H F - - L - - F M - E T - - - - - - - - - - - - - - - - - - - - - - -Aoni DRB*W1303 - - - E - F - P - - - L - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - F - - L - - F M - E T - - - - - - - - - - - - - - - - - - - - - - -Aoni DRB*W1304 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - F - - L - - F M - E T - - - - - - - - - - - - - - - - - - - - - - -Aoni DRB*W1305 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - E - - - F - - L - - F M - E T - - - M - - - - - - - - - - A - - - - - - - -Aoni DRB*W1306 - - S E - F - P - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - L - - F M - E T - - - - - - - - - - - - - - - - - - - - R - -Aoni DRB*W1307 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - F - - L - - F M - E T - - - - - - - - - - - - - - G - - - - - - - -Aoni DRB*W1308 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - F - - L - - F M - E T - - - - - - - - - - D - - - - - - - - - - W -Aoni DRB*W1309 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - - - - L - - F M - E T - - - - - - - - - - - - - - - - - - - - - - -Aovo DRB*W130102 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - F - - - - - - - - - Y - - - - - - - - R T - - - F - - L - - F M - E T - - - - - - - - - - - - - - - - - - - - - - -Aovo DRB*W130101 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - F - - - - - - - - - Y - - - - - - - - R T - - - F - - L - - F M - E T - - - - - - - - K - - - - - - - - - - - - - -Aovo DRB*W1302 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - F - - - - - - - - - Y - - - - - - - - R T - - - F - - L - - F M - E T - - - - - S F - K - - - - - - - - - - - - - -Aovo DRB*W1303 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - F - - L - - F M - E T - - - - - - - - - - - - - - - - - - - - - - -Aovo DRB*W1304 - - - E - F - P - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - Y - - - - - - - - - - - - - F - - L - - F M - E T - - - - - - - - - - - - - - - - - - - A - - -HLA DRB1*160101 - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - Y - - - - - - - - - - - - - - - - - - - F - - D R - - - - - - - - - - - - - - G - - - - - - - -
HLA DRB1*1130 R F L E L L K S E C H F F N G T E R V R F L D R Y F Y N Q E E Y V R F D S D V G E F R A V T E L G R P D E E Y W N S Q K D F L E D R R A A V D T Y C R H N Y G V G E S F T V Q R RAona DRB*W4401 - - - - Q A - - - - Y - - - - - - - - - Y - - - - - - - - K - - - - - - - - - - - L - - - - - - - - R T A Q - - - - - - - - - - - H - - - - - - - - K - - - - - - - - - - - - - -Aoni DRB*W4401 - - - - Q A - - - - Y - - - - - - - - - Y - - - - - - - - K - - - - - - - - - - - L - - - - - - - - R T A Q - - - - - - - - - - - H - - - - - - - - K - K - - - - - - - - - - - -SECONDARY STRUCTURE T
5 β 1 T1 β 2 T2 α 1 α 2 RC β 3 T3 4 T4α
Immunogenetics (2006) 58:542–558 545

1984, Stern et al. 1994): fuchsia for P1, blue for P4, ochrefor P6, grey for P7 and green for P9. PBR sequence secondarystructural features are show at the bottom of the figure.
Slightly less variability was observed in Aotus MHC-DRB beta sheets cf alpha helices. The most polymorphicpositions in the Aotus MHC-DRB locus β sheet were: 9,10, 11, 13, 26, 28, 30, 31, 33 and 37. As these are highlypolymorphic residues in humans, this shows that bothgroups co-varied in these positions. Comparative analysisof alpha helices revealed that the alpha 2 helix had a greaterdegree of polymorphism and a greater number of conservedsubstitutions in each Aotus MHC-DRB lineage. The mostpolymorphic positions in the alpha chain were: 57, 67, 68,70, 73 and 74; they were also highly polymorphic inhumans. Positions 63–68 were more variable in AotusMHC-DRB than in HLA-DRB. The fragment encoding therandom coil structure between residues 86 and 94 showedsimilar polymorphism in Aotus MHC-DRB to that observedfor human lineages.
Some Aotus MHC-DRB sequences were identical at thenucleotide level amongst the species analysed. Only thesequence corresponding to A. nancymaae DRB*W1301was shared by the three species. Identity was only sharedby two species in another ten cases; these numbers are fartoo small to be indicative of evolutionary relationshipamongst the species being studied because of the differencein sample size. However, they do suggest a closerelationship between them. This indicates long-lastingmaintenance of allelic variants, until the ancient origin ofthis species (8 million years ago; see Setoguchi andRosenberger 1987). The Aotus DRB*W43 lineage is worthnoting, as it mainly consisted of A. nigriceps sequences.Given that this species has been less sampled compared toA. nancymaae (even with our sampling limitations), thisgroup suggested that there could be preferential represen-tation for some lineages according to a particular Aotusspecies.
Number of MHC-DRB sequences expressed in Aotus
A maximum of four sequences was observed per monkeyfor A. nancymaae, four for A. nigriceps and three for A.vociferans. The differences between different species couldhave been due to the varying number of monkeys beingsampled for each species and the random method used forchoosing the clones to be sequenced. This number agreedwith the number of MHC-DRB genes expressed in A.trivirgatus (5), determined by denaturing gradient gelelectrophoresis (Middleton et al. 2004). Duplication eventswere detected in several lineages (expression of the samelineage several times in one specimen) such as DRB1*03and DRB*W13 for all species studied, DRB*W29 for A.nancymaae and A. nigriceps, DRB*W43 for A. nigriceps
and DRB*W18, DRB*W47, DRB*W38 and DRB3*06 forA. nancymaae. This issue is still being clarified for furtherstudies.
Evolutionary analysis of Aotus and humanswithin the context of primate MHC-DRB exon 2
Figure 2 shows minimum evolution analysis with Kimura 2parameter distance, including a selection of 170 OTUs,Operational Taxonomic Units (267 nucleotide positions)from 820 primate sequences considered. A sample of allPlatyrrhini MHC-DRB sequences, an MHC-DRB sequencesample from M. mulatta and humans as Catarrhini members,and a representative sample of Aotus MHC-DRB sequencesreported in this paper were used, and some sequences ofMHC-DPB1 and MHC-DQB1 were taken as out-groups.
The tree shows that the groups made up of primateDRB sequences tended to become associated according todescribed allele lineage, but such association was seldomsupported by a statistically solid reliability value, inde-pendently of the type of sequence from which thetopology was calculated (nucleotides or amino acids).This observation is common to other studies using theMHC-DRB exon 2 (Kriener et al. 2000a; Antunes et al.1998; Wu et al. 2000; Go et al. 2002; Figueroa et al. 2000;Middleton et al. 2004), explained by the large number ofvariable positions, the enormous structural restriction cfselective pressure, molecular convergence, and the recom-bination phenomena to which this sector was submitted(Ohta 1991; Kriener et al. 2001; Hughes and Yeager 1998;Yeager and Hughes 1999; Klein et al. 1998; Kriener et al.2000a,b; O’hUigin 1995).
Seventeen groups (shown in bold in Fig. 2) of sequenceswere recovered from topologies generated just from thegenus’ sequences with the complete MHC-DRB exon 2sequence, whether by distance methods (ME, NJ) orparsimony, and independently of the type of sequence used(DNA or proteins). Some groups of Aotus sequences didnot have statistical support, even though most werewell-supported (Fig. 2). These groups were also
�Fig. 2 Minimum evolution tree for primate MHC-DRB exon 2sequences. The tree was constructed based on Kimura 2 parameterscalculated from aligning 267 nt nucleotide from primate MHC-DRBexon 2 sequences. Bootstrap values were calculated based on 1,000replicates, which only showed values higher than 60% (>70% wassignificant). Values shown in bold indicate values that were alsosignificant for internal branch test (IBT; >95%); circles denote valuesthat were only significant for IBT. We have included Catarrhini[Hominoidea (humans shown by boldgreylines) and Cercopithecoidea(OWM, Macaca mulatta) in grey] and Platyrrhini sequences (NWM)including Aotus (blackboldlines) and other NWM genera (black). DPBand DQB sequences from several primate genera were used as out-groups. The scalebar represents 0.05 substitutions per site. See“Materials and methods” section for species’ abbreviations
546 Immunogenetics (2006) 58:542–558

Aona-DRB1*0307Aoni-DRB1*0303
Aotr-DRB1*0301Aona-DRB1*0319
Aoni-DRB1*0306Aotr-DRB1*0303Aoni-DRB1*0304
Aovo-DRB1*0303Aona-DRB1*0303
Aona-DRB1*0312Aona-DRB1*0309Aovo-DRB1*0301
Aotus MHC-DRB1*03 GA
Aona-DRB1*0301Aona-DRB1*0302
Aona-DRB1*0320Aona-DRB1*0325
Aotus MHC-DRB1*03 GB
Aona-DRB1*0313Aona-DRB1*0314
Aoni-DRB1*0305Aotus MHC-DRB1*03 GC
Aona-DRB3*0601Aotr-DRB3*0602
Aona-DRB3*0613Aotr-DRB
Aona-DRB3*0605Aona-DRB3*0620
Aona-DRB3*0622Aona-DRB3*0623
Aona-DRB3*0624Aona-DRB3*0612
Aoaz-DRB3*0601Aoni-DRB3*0601
Aotus MHC-DRB3*06 GA
HLA-DRB1*030101HLA-DRB1*030201
Mamu-DRB1*0315Mamu-DRB1*0316
Catarrihini MHC-DRB1*03
Mamu-DRB*W2002HLA-DRB3*010101HLA-DRB3*0201
HLA-DRB3*030101Catarrihini MHC-DRB3
Saoe-DRB3*0508Saoe-DRB3*0510
Pipi-DRB3*0501Camo-DRB3*0501
Atbe-DRB3*0501
Platyrrhini MHC-DRB3*05
Camo-DRB3*0701Aotr-DRB*W1801
Aona-DRB*W1802Aona-DRB*W1801
Aoni-DRB*W1801Aotus MHC-DRB*W18
Aona-DRB*W2901Aoni-DRB*W2901
Aona-DRB*W2909Aoni-DRB*W2902
Aovo-DRB*W2901Aotus MHC-DRB*W29
Aona-DRB*W4201Aoni-DRB*W4201 Aotus MHC-DRB*W42Aona-DRB3*0609
Aona-DRB3*0608Aona-DRB3*0610
Aotus MHC-DRB3*06 GBCamo-DRB3*0601
Aona-DRB*W4701Aona-DRB*W4702Aona-DRB*W4706
Aona-DRB*W4709Aotus MHC-DRB*W47Aotus MHC-DRB*W45Aona-DRB*W4501Aotus MHC-DRB*W46Aona-DRB*W4601
Ceap-DRB*W1301Ceap-DRB*W1302 Ceap-DRB*W13
Aoni-DRB*W4301Aoni-DRB*W4304
Aovo-DRB*W4301Aoni-DRB*W4303
Aoni-DRB*W4305
Aotus MHC-DRB*W43
Aotus MHC-DRB*W41Aona-DRB*W4101Aona-DRB*W4401
Aoni-DRB*W4401 Aotus MHC-DRB*W44Ceap-DRB*W1303
Aona-DRB*W3001Aona-DRB*W1303
Aona-DRB*W1309Aona-DRB*W1312
Aovo-DRB*W1302Aona-DRB*W1310
Aoni-DRB*W1309Aoni-DRB*W1307
Aoni-DRB*W1303Aovo-DRB*W1304
Aoni-DRB*W1308Aona-DRB*W1306Aona-DRB*W1307
Aotus MHC-DRB*W13
HLA-DRB5*010101HLA-DRB5*0103 HLA-DRB5
Mamu-DRB*W2507Mamu-DRB*W502
Mamu-DRB19HLA-DRB1*150101
HLA-DRB1*150102 HLA-DRB1*15HLA-DRB1*160101
HLA-DRB1*160102 HLA-DRB1*16HLA-DRB1*120101
HLA-DRB1*120302 HLA-DRB1*12HLA-DRB1*140101
HLA-DRB1*1404 HLA-DRB1*14HLA-DRB1*080101
HLA-DRB1*080201 HLA-DRB1*08Mamu-DRB1*0317
Mamu-DRB1*0318 Mamu-DRB1*03HLA-DRB1*110101
HLA-DRB1*1102 HLA-DRB1*11HLA-DRB1*130101
HLA-DRB1*130201 HLA-DRB1*13Caja-DRB1*0301
Caja-DRB1*0302Capy-DRB1*0301
Saoe-DRB1*0301Camo-DRB1*0302
Camo-DRB1*0303
CallitrichidaeMHC-DRB1*03
Ceap-DRB*W1501Ceap-DRB*W1502 Camo-DRB1*03
Sasc-DRB*W1901Sasc-DRB*W1903 Sasc-DRB*W19
Caja-DRB11*0101Saoe-DRB11*0101Caja-DRB*W1611
Saoe-DRB11*0108Caja-DRB*W1601
Caja-DRB*W1605
CallitrichidaeMHC-DRB11/DRB*W16
Caja-DRB*W1201Capy-DRB*W1201
Ceap-DRB*W3201Pipi-DRB*W1201
Saoe-DRB*W1205
CallitrichidaeMHC-DRB*W12
Camo-DRB*W1401Camo-DRB*W1402
Sasc-DRB*W1401Sasc-DRB*W1404
PlatyrrihiniMHC-DRB*W14
Caja-DRB*W1606Caja-DRB*W1607 Caja-DRB*W16
Sasc-DRB*W1203Sasc-DRB*W1204 Sasc-DRB*W12
Caja-DRB*W1608Saoe-DRB*W2201
Saoe-DRB*W2209Sasc-DRB*W1201
PlatyrrihiniMHC-DRB*W22
Aona-DRB*W3801Aoaz-DRB*W3801
Aoni-DRB*W3801Atbe-DRB*W3801
Aotus MHC-DRB*W38
HLA-DRB1*0707HLA-DRB1*0705 HLA-DRB1*07
Mamu-DRB110HLA-DRB1*0406
HLA-DRB1*040101 HLA-DRB1*04HLA-DRB1*0104
HLA-DRB1*010102 HLA-DRB1*01HLA-DRB1*0902
HLA-DRB1*090102 HLA-DRB1*09Mamu-DRB51
Saoe-DRB5*0701Mamu-DRB5*0307
Mamu-DRB111HLA-DRB1*10HLA-DRB1*100101
Mamu-DRBw1Mamu-DRB41
HLA-DRB4*010101 Catarrihini MHC-DRB4HLA-DPB1*010101
Mamu-DPB1*15 MHC-DPB1Caja-DQB1*2302
HLA-DQB1*0201Mamu-DQB1*1703
MHC-DQB1
99
8499
9994
96
95
99
99
99
99
99
99
99
99
75
99
92
78
89
90
71
98
80
99
97
99
9299
99
64
60
99
98
87
99
97
96
98
99
99
73
99
96
99
83
87
90
77
82
97
65
71
75
88
92
92
76
63
86
61
91
64
70
98
85
62
98
0,05
Aotus MHC-DRB*W30
Immunogenetics (2006) 58:542–558 547

distinguished at both nucleotide and amino acid levelswhen only Aotus MHC-DRB sequences were analysed(data not shown).
No identical MHC-DRB alleles was observed betweenAotus and other primate species; cases of total identityamongst alleles from different primate species are rare(Otting et al. 2000), generally being restricted to speciesfrom the same genus and more rarely from the same family.This indicates that most alleles in primates originated inpost-speciation phenomena; even though a very few havehigh identity at genus level, this is rarely maintainedbeyond this taxonomic division.
Most NWM monkey sequences, irrespective of methodand type of sequence (NT or AA), were grouped into asingle, highly consistent group. This group includedpractically all Platyrrhini lineages (Pithecinae, Atelinae,Cebinae and Callitrichinae) but did not contain Aotus(classified as Cebidae, together with Saimiri and Cebus;Fig. 2). Aotus was seen to be a highly differentiated groupwithin the exon 2 context at nucleotide level, being mainlyrelated to Catarrhini lineages such as DRB3 (from allprimates including other NWM) and DRB1*07 (human)(Fig. 2). Some groups of Aotus MHC-DRB presented anassociation with HLA-DRB1*03 and HLA-DRB1*10 andwith HLA-DRB1*04, HLA-DRB1* 08-11-12-13 and HLA-DRB1*01 at the amino acid level, in addition to thosealready described (data not shown).
Aotus MHC-DRB variability within the context of primateMHC-DRB exon 2
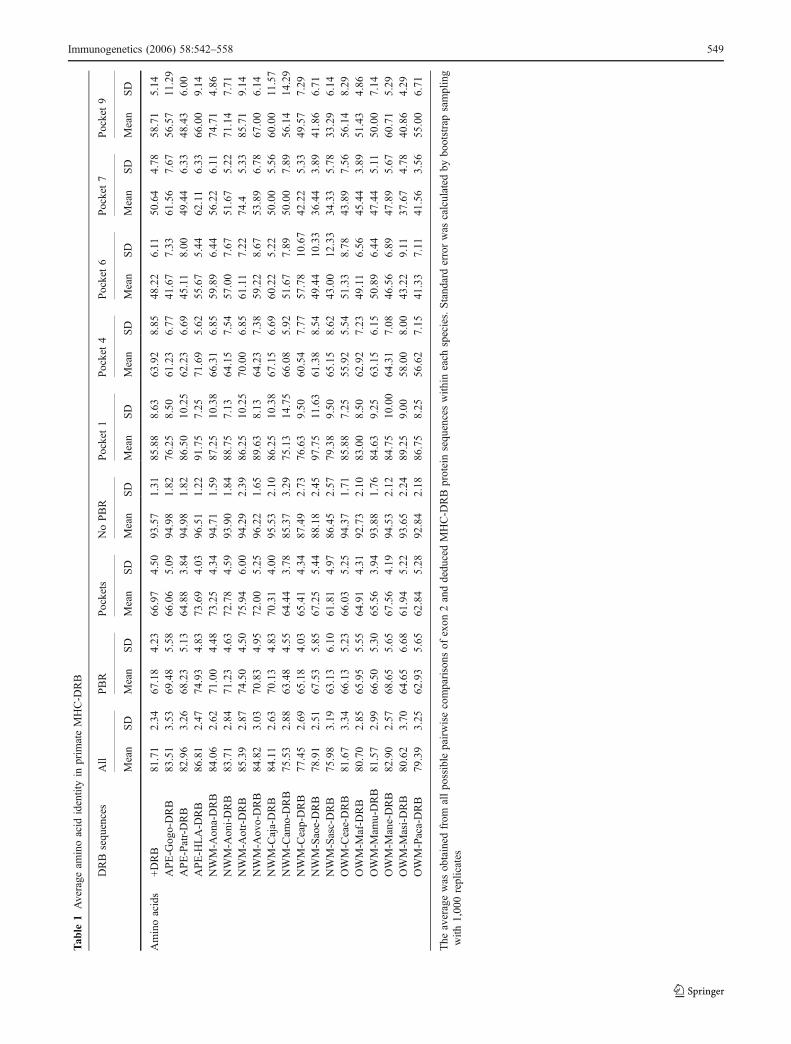
Overall, Aotus DRB had the same variability as humans andapes in the MHC-DRB β-domain at both nucleotide andamino acid levels. It was notable how this variability wasless than that shown for other NWM and OWM. These twogroups of species (the former having lower variation) couldbe distinguished at the PBR, non-PBR and pocket levels(Table 1). Like all primates considered, Aotus had greatervariability (almost double) at protein sequence than nucleo-tide level in exon 2, implying active protein variantproduction (data not shown). Variability trends were verysimilar between nucleotides and amino acids for the threespecies considered. DRB exon 2 analyses showed greatermutation accumulation in the PBR than in the region notdirectly involved in peptide binding (non-PBR in Table 1).In fact, variation at the nucleotide level in the non-PBR wasequivalent to that of amino acids, indicating evolutionarydynamics different to those of the PBR (Table 1). P1 was themost conserved pocket in primates; P4 showed mediumvariation and P6, P7 and P9 showed high variability. AotusMHC-DRB followed a similar variation pattern to thatexhibited by HLA-DRB; however (bearing in mind thedifferent number of monkeys sampled for each species), the
least variable was A. vociferans, and A. nancymaae and A.nigriceps were the most variable (with equivalent values)(Table 1).
Recombination, population expansion and convergencein Aotus MHC-DRB exon 2
Mismatch distribution analysis showed that none of theAotus species analysed showed population expansion or totalrecombination at sequence or MHC-DRB secondary struc-ture level (in spite of evidence suggesting that recombinationphenomena are more frequent in alpha helices than betasheets; Go et al. 2002; see below). The frequencies shownwere clearly bimodal, disagreeing with a Poisson distribution(P<0.0001) in all cases; this was confirmed by theKolmogorov–Smirnov test (data not shown). Such distribu-tion arising from this analysis indicated the loss of allelelineages, thus producing islands of similarity detected aslocal maximums in the distribution of pairwise differencefrequencies amongst the sequences analysed. If sequencedivergence had not occurred due to a random process(bottlenecks, recombination phenomena, high rate of muta-tion provoking convergence phenomena, etc.), then histo-grams would have tended to be unimodal and would havefollowed Poisson distribution (data not shown) (Go et al.2002; Di Rienzo and Wilson 1991; Figueroa et al. 2000).This result indicated that Aotus MHC-DRB did not undergothe aforementioned processes during its evolution.
Additional analysis, following the method implementedin GENECONV software (Sawyer 1989, 1999) showed thatAotus MHC-DRB has suffered 96 significant globalrecombination events (<0.05 P values; Table 2) and 4,756significant pairwise recombination events (<0.05 P values;data not shown) involving all groups of sequences and largeexon 2 segments in each one. Gene conversion was relatedto the search for these genes’ diversity as an adaptiveresponse, increasing MHC-DRB global polymorphism asdescribed for other vertebrate species (Ohta 1991; Yeagerand Hughes 1999). It is noticeable that its very infrequentconversion events in positions 6–13, considered diagnosticfor determining allele lineage (Bergstrom et al. 1999) (e.g.the 9EYSTS13 motif), was thus characteristic of HLA-DRB1*03 (Fig. 1). This criterion was generally compliedwith in Aotus MHC-DRB lineages.
Natural selection in Aotus MHC-DRB exon 2
Two methods were used for analysing the type of selectionto which Aotus MHC-DRB has been submitted withinprimates. The first consisted of the Nei–Gojobori test fordetermining the significance of disequilibrium (if therewere any) between synonymous (dS) and non-synonymous(dN) substitutions. The second was a graphical representa-
548 Immunogenetics (2006) 58:542–558
Tab
le1
Average
aminoacid
identityin
prim
ateMHC-D
RB
DRBsequ
ences
All
PBR
Pockets
NoPBR
Pocket1
Pocket4
Pocket6
Pocket7
Pocket9
Mean
SD
Mean
SD
Mean
SD
Mean
SD
Mean
SD
Mean
SD
Mean
SD
Mean
SD
Mean
SD
Aminoacids
+DRB
81.71
2.34
67.18
4.23
66.97
4.50
93.57
1.31
85.88
8.63
63.92
8.85
48.22
6.11
50.64
4.78
58.71
5.14
APE-G
ogo-DRB
83.51
3.53
69.48
5.58
66.06
5.09
94.98
1.82
76.25
8.50
61.23
6.77
41.67
7.33
61.56
7.67
56.57
11.29
APE-Patr-DRB
82.96
3.26
68.23
5.13
64.88
3.84
94.98
1.82
86.50
10.25
62.23
6.69
45.11
8.00
49.44
6.33
48.43
6.00
APE-H
LA-D
RB
86.81
2.47
74.93
4.83
73.69
4.03
96.51
1.22
91.75
7.25
71.69
5.62
55.67
5.44
62.11
6.33
66.00
9.14
NWM-A
ona-DRB
84.06
2.62
71.00
4.48
73.25
4.34
94.71
1.59
87.25
10.38
66.31
6.85
59.89
6.44
56.22
6.11
74.71
4.86
NWM-A
oni-DRB
83.71
2.84
71.23
4.63
72.78
4.59
93.90
1.84
88.75
7.13
64.15
7.54
57.00
7.67
51.67
5.22
71.14
7.71
NWM-A
otr-DRB
85.39
2.87
74.50
4.50
75.94
6.00
94.29
2.39
86.25
10.25
70.00
6.85
61.11
7.22
74.4
5.33
85.71
9.14
NWM-A
ovo-DRB
84.82
3.03
70.83
4.95
72.00
5.25
96.22
1.65
89.63
8.13
64.23
7.38
59.22
8.67
53.89
6.78
67.00
6.14
NWM-Caja-DRB
84.11
2.63
70.13
4.83
70.31
4.00
95.53
2.10
86.25
10.38
67.15
6.69
60.22
5.22
50.00
5.56
60.00
11.57
NWM-Cam
o-DRB
75.53
2.88
63.48
4.55
64.44
3.78
85.37
3.29
75.13
14.75
66.08
5.92
51.67
7.89
50.00
7.89
56.14
14.29
NWM-Ceap-DRB
77.45
2.69
65.18
4.03
65.41
4.34
87.49
2.73
76.63
9.50
60.54
7.77
57.78
10.67
42.22
5.33
49.57
7.29
NWM-Saoe-DRB
78.91
2.51
67.53
5.85
67.25
5.44
88.18
2.45
97.75
11.63
61.38
8.54
49.44
10.33
36.44
3.89
41.86
6.71
NWM-Sasc-DRB
75.98
3.19
63.13
6.10
61.81
4.97
86.45
2.57
79.38
9.50
65.15
8.62
43.00
12.33
34.33
5.78
33.29
6.14
OWM-Ceae-DRB
81.67
3.34
66.13
5.23
66.03
5.25
94.37
1.71
85.88
7.25
55.92
5.54
51.33
8.78
43.89
7.56
56.14
8.29
OWM-M
af-D
RB
80.70
2.85
65.95
5.55
64.91
4.31
92.73
2.10
83.00
8.50
62.92
7.23
49.11
6.56
45.44
3.89
51.43
4.86
OWM-M
amu-DRB
81.57
2.99
66.50
5.30
65.56
3.94
93.88
1.76
84.63
9.25
63.15
6.15
50.89
6.44
47.44
5.11
50.00
7.14
OWM-M
ane-DRB
82.90
2.57
68.65
5.65
67.56
4.19
94.53
2.12
84.75
10.00
64.31
7.08
46.56
6.89
47.89
5.67
60.71
5.29
OWM-M
asi-DRB
80.62
3.70
64.65
6.68
61.94
5.22
93.65
2.24
89.25
9.00
58.00
8.00
43.22
9.11
37.67
4.78
40.86
4.29
OWM-Paca-DRB
79.39
3.25
62.93
5.65
62.84
5.28
92.84
2.18
86.75
8.25
56.62
7.15
41.33
7.11
41.56
3.56
55.00
6.71
The
averagewas
obtained
from
allpo
ssible
pairwisecomparisons
ofexon
2anddedu
cedMHC-D
RBproteinsequ
enceswith
ineach
species.Stand
arderrorwas
calculated
bybo
otstrapsampling
with
1,00
0replicates
Immunogenetics (2006) 58:542–558 549

tion of dN cf dS, allowing the intensity of the selectiveprocess to which different DRB exon 2 sectors have beensubmitted to be appreciated; the more the divergence fromcentral tendency (the line of neutrality) in a determinedgroup of sequences, the greater the pressure to which it hasbeen submitted. Presence in the upper left-hand sector of aspecie marker indicated positive or diversifying pressure,and presence in the lower left-hand sector indicated negative
Table 2 Gene conversion events
Sequences Begin(aln)
End(aln)
Length
Aoni_DRB1*0304–Aona_DRB3*0608 39 190 152Aovo_DRB1*0304–Aona_DRB3*0608 39 190 152Aoni_DRB1*W1308–Aona_DRB1*0309 47 181 135Aona_DRB1*0311–Aona_DRB3*0614 112 279 168Aona_DRB*W1303–Aona_DRB1*0309 47 170 124Aona_DRB*W1304–Aona_DRB1*0309 47 181 135Aona_DRB*W1306–Aona_DRB1*0309 47 181 135Aoni_DRB*W1301–Aona_DRB1*0309 47 181 135Aovo_DRB*W1304–Aona_DRB1*0309 47 181 135Aoni_DRB*W1302–Aona_DRB1*0309 47 177 131Aona_DRB*W1310–Aona_DRB1*0309 47 190 144Aoni_DRB*W1306–Aona_DRB1*0309 47 190 144Aona_DRB*W1307–Aona_DRB1*0309 47 180 134Aoni_DRB*W1303–Aona_DRB1*0309 50 181 132Aona_DRB*W1311–Aona_DRB1*0309 47 170 124Aoni_DRB*W1308–Aona_DRB1*0304 43 181 139Aona_DRB1*0303–Aona_DRB3*0616 112 279 168Aona_DRB1*0311–Aona_DRB3*0615 112 279 168Aona_DRB1*0312–Aona_DRB3*0614 112 279 168Aona_DRB*W1302–Aona_DRB1*0309 47 181 135Aovo_DRB*W1303–Aona_DRB1*0309 47 181 135Aona_DRB*W1303–Aona_DRB1*0304 43 170 128Aoni_DRB*W1309–Aona_DRB1*0309 47 190 144Aona_DRB*W1309–Aona_DRB1*0309 47 170 124Aoni_DRB*W1305–Aona_DRB1*0309 47 170 124Aovo_DRB1*0303–Aona_DRB3*0608 83 190 108Aona_DRB*W1304–Aona_DRB1*0304 43 181 139Aona_DRB*W1306–Aona_DRB1*0304 43 181 139Aoni_DRB*W1301–Aona_DRB1*0304 43 181 139Aovo_DRB*W1304–Aona_DRB1*0304 43 181 139Aoni_DRB*W1302–Aona_DRB1*0304 43 177 135Aona_DRB*W1311–Aona_DRB1*0304 43 170 128Aona_DRB*W1307–Aona_DRB1*0304 43 180 138Aoni_DRB*W1307–Aona_DRB1*0309 47 181 135Aona_DRB1*0303–Aona_DRB3*0617 112 279 168Aona_DRB1*0312–Aona_DRB3*0615 112 279 168Aona_DRB*W1310–Aona_DRB1*0304 43 190 148Aoni_DRB*W1306–Aona_DRB1*0304 43 190 148Aoni_DRB*W1303–Aona_DRB1*0304 50 181 132Aona_DRB*W1308–Aona_DRB1*0309 47 170 124Aona_DRB*W1302–Aona_DRB1*0304 43 181 139Aovo_DRB*W1303–Aona_DRB1*0304 43 181 139Aoni_DRB*W1302–Aona_DRB1*0302 43 166 124Aoni_DRB*W1302–Aona_DRB1*0326 43 166 124Aoni_DRB*W1308–Aona_DRB1*0302 43 166 124Aoni_DRB*W1308–Aona_DRB1*0326 43 166 124Aona_DRB*W4711–Aona_DRB1*0311 39 166 128Aona_DRB*W2906–Aona_DRB3*0609 95 279 185Aona_DRB*W1309–Aona_DRB1*0304 43 170 128Aoni_DRB*W1305–Aona_DRB1*0304 43 170 128Aona_DRB1*0304–Aona_DRB3*0614 112 256 145Aona_DRB1*0305–Aona_DRB3*0614 112 256 145Aona_DRB1*0318–Aona_DRB3*0614 112 256 145Aona_DRB1*0319–Aona_DRB3*0614 112 256 145Aoni_DRB1*0307–Aona_DRB3*0614 112 256 145
Table 2 (continued)
Sequences Begin(aln)
End(aln)
Length
Aotr_DRB1*0301–Aona_DRB3*0614 112 256 145Aovo_DRB1*0303–Aona_DRB3*0614 112 256 145Aona_DRB*W2902–Aona_DRB*W2905 16 190 175Aona_DRB*W2907–Aona_DRB1*0316 94 255 162Aona_DRB*W2907–Aona_DRB3*0609 95 256 162Aona_DRB1*0303–Aona_DRB3*0601 112 282 171Aoni_DRB*W1309–Aona_DRB1*0304 43 190 148Aona_DRB*W2909–Aona_DRB3*0609 95 261 167Aona_DRB1*0311–Aona_DRB3*0623 104 279 176Aovo_DRB1*0303–Aona_DRB3*0605 104 279 176Aovo_DRB1*0303–Aona_DRB3*0606 104 279 176Aona_DRB*W1303–Aona_DRB1*0302 43 166 124Aona_DRB*W1304–Aona_DRB1*0302 43 166 124Aona_DRB*W1304–Aona_DRB1*0326 43 166 124Aona_DRB*W1306–Aona_DRB1*0302 43 166 124Aona_DRB*W1306–Aona_DRB1*0326 43 166 124Aona_DRB*W1307–Aona_DRB1*0302 43 166 124Aona_DRB*W1307–Aona_DRB1*0326 43 166 124Aoni_DRB*W1301–Aona_DRB1*0302 43 166 124Aoni_DRB*W1301–Aona_DRB1*0326 43 166 124Aoni_DRB*W1302–Aona_DRB1*0320 43 166 124Aoni_DRB*W1302–Aotr_DRB*W1303 43 166 124Aoni_DRB*W1308–Aona_DRB1*0320 43 166 124Aoni_DRB*W1308–Aotr_DRB*W1303 43 166 124Aovo_DRB*W1304–Aona_DRB1*0302 43 166 124Aovo_DRB*W1304–Aona_DRB1*0326 43 166 124Aona_DRB*W4101–Aona_DRB1*0305 89 181 93Aona_DRB*W4101–Aona_DRB1*0318 89 181 93Aona_DRB*W4101–Aotr_DRB1*0301 89 181 93Aona_DRB*W4101–Aotr_DRB1*0302 89 181 93Aona_DRB1*0318–Aona_DRB3*0605 78 279 202Aona_DRB1*0319–Aona_DRB3*0605 78 279 202Aoni_DRB1*0307–Aona_DRB3*0605 78 279 202Aotr_DRB1*0301–Aona_DRB3*0605 78 279 202Aoni_DRB*W1307–Aona_DRB1*0304 43 181 139Aona_DRB*W1308–Aona_DRB1*0304 43 170 128Aona_DRB1*0308–Aona_DRB3*0622 47 166 120Aona_DRB*W2902–Aona_DRB3*0614 141 256 116Aona_DRB*W2905–Aona_DRB1*0316 94 279 186Aona_DRB*W1807–Aona_DRB*W3001 102 168 67Aotr_DRB*W1301–Aona_DRB*4701 47 173 127
Global inner significant fragments (Sim P values <0.05)were calculated with GENECONV software. The table shows thebeginning, end and length of the fragments
550 Immunogenetics (2006) 58:542–558

or conservative pressure. The same graph shows whether thegroup of sequences have (or did not have) a significantselection value according to Nei–Gojobori test (Nei andGojobori 1986).
The type of selection between different Aotus MHC-DRB PBR sectors was similar to HLA-DRB. Figure 3presents the result of the above analysis. All speciesshowed a slight tendency to accumulate more non-synon-
AllGogo
Patr
HLA
Aovo
Camo
CeapSaoe
Sasc
CeaeMane
Masi
AonaAoni
Aotr
Caja
Mafa
Mamu
Paca
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35
HLA
Aoni
Aovo
Camo Ceap
SaoeSasc
MafaMasi
PacaAll
GogoPatr
AonaAotrCaja
CeaeMamuMane
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0.00 0.02 0.04 0.06 0.08 0.10 0.12 0.14 0.16 0.18
AllGogo
Patr
HLA
AonaAoni
Aotr
Aovo
Camo
Ceap
SaoeSasc
Mafa
MamuMane
Masi
Paca
Caja
Ceae
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0.04 0.06 0.08 0.10 0.12 0.14 0.16 0.18 0.20
AllGogoPatr
HLA
Aona
Aoni
Aotr
AovoCamo
Ceap
Ceae
Mane
Masi
Paca
Caja
SaoeSasc
Mafa
Mamu
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.40
0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35 0.40
All
Gogo
Patr
HLA
AonaAotr
Aovo
Camo
Ceap
Saoe
Sasc
Ceae
Mafa
Mamu
Mane
Masi
Paca
Aoni
Caja
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35
Gogo
Patr
HLAAona
Aotr
Aovo
Camo
CeapSaoe
Sasc
Ceae
Mafa
MamuMane
Paca
All
Aoni
Caja
Masi
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.05 0.10 0.15 0.20 0.25 0.30 0.35
AllGogo
Patr
HLAAona
Aoni
Aotr
Aovo
Camo
Ceap
Saoe
Sasc
Mane
Masi
PacaCajaCeae
MafaMamu
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.80
0.00 0.10 0.20 0.30 0.40 0.50 0.60 0.70 0.80
Gogo
HLA
Aoni
Aotr
Camo Ceap
Saoe
Sasc
Ceae
Mafa
Mane
Masi
Paca
AllPatr
Aona
Ao
vo
Caja
Mamu
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.00 0.10 0.20 0.30 0.40 0.50 0.60
Gogo
Aotr
Camo
Ceap
SaoeSasc
CeaeMafa
Mamu
MasiPaca
AllPatr
HLA
Aona
Aoni
Ao
vo Caja
Mane
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.00 0.10 0.20 0.30 0.40 0.50 0.60 0.70
dN
dS
whole exon 2 non-PBR PBR
Set of pocket residues
OWM NWM AotusHLA-Apes All DRB
Pocket 1 Pocket 4
Pocket 6 Pocket 7 Pocket 9
AllGogo
Patr
HLA
Aovo
Camo
CeapSaoe
Sasc
CeaeMane
Masi
AonaAoni
Aotr
Caja
Mafa
Mamu
Paca
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35
HLA
Aoni
Aovo
Camo Ceap
SaoeSasc
MafaMasi
PacaAll
GogoPatr
AonaAotrCaja
CeaeMamuMane
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0.00 0.02 0.04 0.06 0.08 0.10 0.12 0.14 0.16 0.18
AllGogo
Patr
HLA
AonaAoni
Aotr
Aovo
Camo
Ceap
SaoeSasc
Mafa
MamuMane
Masi
Paca
Caja
Ceae
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0.04 0.06 0.08 0.10 0.12 0.14 0.16 0.18 0.20
AllGogoPatr
HLA
Aona
Aoni
Aotr
AovoCamo
Ceap
Ceae
Mane
Masi
Paca
Caja
SaoeSasc
Mafa
Mamu
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.40
0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35 0.40
All
Gogo
Patr
HLA
AonaAotr
Aovo
Camo
Ceap
Saoe
Sasc
Ceae
Mafa
Mamu
Mane
Masi
Paca
Aoni
Caja
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35
Gogo
Patr
HLAAona
Aotr
Aovo
Camo
CeapSaoe
Sasc
Ceae
Mafa
MamuMane
Paca
All
Aoni
Caja
Masi
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.05 0.10 0.15 0.20 0.25 0.30 0.35
AllGogo
Patr
HLAAona
Aoni
Aotr
Aovo
Camo
Ceap
Saoe
Sasc
Mane
Masi
PacaCajaCeae
MafaMamu
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.80
0.00 0.10 0.20 0.30 0.40 0.50 0.60 0.70 0.80
Gogo
HLA
Aoni
Aotr
Camo Ceap
Saoe
Sasc
Ceae
Mafa
Mane
Masi
Paca
AllPatr
Aona
Ao
vo
Caja
Mamu
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.00 0.10 0.20 0.30 0.40 0.50 0.60
Gogo
Aotr
Camo
Ceap
SaoeSasc
CeaeMafa
Mamu
MasiPaca
AllPatr
HLA
Aona
Aoni
Ao
vo Caja
Mane
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.00 0.10 0.20 0.30 0.40 0.50 0.60 0.70
dN
dS
whole exon 2 non-PBR PBR
Set of pocket residues
OWM NWM AotusHLA-Apes All DRB
Pocket 1 Pocket 4
Pocket 6 Pocket 7 Pocket 9
AllGogo
Patr
HLA
Aovo
Camo
CeapSaoe
Sasc
CeaeMane
Masi
AonaAoni
Aotr
Caja
Mafa
Mamu
Paca
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35
HLA
Aoni
Aovo
Camo Ceap
SaoeSasc
MafaMasi
PacaAll
GogoPatr
AonaAotrCaja
CeaeMamuMane
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0.00 0.02 0.04 0.06 0.08 0.10 0.12 0.14 0.16 0.18
AllGogo
Patr
HLA
AonaAoni
Aotr
Aovo
Camo
Ceap
SaoeSasc
Mafa
MamuMane
Masi
Paca
Caja
Ceae
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0.04 0.06 0.08 0.10 0.12 0.14 0.16 0.18 0.20
AllGogoPatr
HLA
Aona
Aoni
Aotr
AovoCamo
Ceap
Ceae
Mane
Masi
Paca
Caja
SaoeSasc
Mafa
Mamu
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.40
0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35 0.40
All
Gogo
Patr
HLA
AonaAotr
Aovo
Camo
Ceap
Saoe
Sasc
Ceae
Mafa
Mamu
Mane
Masi
Paca
Aoni
Caja
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35
Gogo
Patr
HLAAona
Aotr
Aovo
Camo
CeapSaoe
Sasc
Ceae
Mafa
MamuMane
Paca
All
Aoni
Caja
Masi
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.05 0.10 0.15 0.20 0.25 0.30 0.35
AllGogo
Patr
HLAAona
Aoni
Aotr
Aovo
Camo
Ceap
Saoe
Sasc
Mane
Masi
PacaCajaCeae
MafaMamu
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.80
0.00 0.10 0.20 0.30 0.40 0.50 0.60 0.70 0.80
Gogo
HLA
Aoni
Aotr
Camo Ceap
Saoe
Sasc
Ceae
Mafa
Mane
Masi
Paca
AllPatr
Aona
Ao
vo
Caja
Mamu
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.00 0.10 0.20 0.30 0.40 0.50 0.60
Gogo
Aotr
Camo
Ceap
SaoeSasc
CeaeMafa
Mamu
MasiPaca
AllPatr
HLA
Aona
Aoni
Ao
vo Caja
Mane
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.00 0.10 0.20 0.30 0.40 0.50 0.60 0.70
dN
dS
whole exon 2 non-PBR PBR
Set of pocket residues
OWM NWM AotusHLA-Apes All DRB
Pocket 1 Pocket 4
Pocket 6 Pocket 7 Pocket 9
AB
C
D E F
G H I
Fig. 3 Primate MHC-DRB natural selection. The graphs for specificpockets show dN cf dS substitutions; the line represents the tendencytowards neutrality. The regions above and below the line of neutralitydenote positive selection pressure and negative selection pressure,respectively. Names are shown in bold and greatersymbolsizeindicates significant selection pressure as revealed by Nei–Gojobori
test. A rhombus denotes the set of all MHC-DRB studied. Afilledcircle denotes human and ape MHC-DRB. A filledsquaredenotes OWM MHC-DRB, a filledtriangle denotes NWM MHC-DRB and an emptytriangle denotes Aotus MHC-DRB sequences. Seethe “Materials and methods” section for species’ abbreviations
Immunogenetics (2006) 58:542–558 551

ymous (dN) than synonymous (dS) mutations for the wholeof exon 2 (Fig. 3a); however, this tendency was onlysignificant in the Nei–Gojobori test for some NWM species(Aoni, Caja, Sasc and Ceap, shown in bold). The closestgroup to HLA-DRB was Aotus MHC-DRB, showing thattotal variability tendency followed a similar pattern foraccumulating dN cf dS mutations.
The non-peptide binding region (non-PBR, Fig. 3b)consistently exhibited negative selective pressure andseparation between Catarrhini (OWM and humans andapes) and Platyrrhini (NWM), as the first accumulatedsynonymous cf non-synonymous mutations with signifi-cantly greater intensity, whilst NWM presented a patternclose to neutrality. Aotus MHC-DRB moved away from thistendency, and (even though not having significant negativepressure) it approached the tendency exhibited by HLA-DRB more than other NWM, once more confirming thesetwo species’ close similarity at this level.
PBR (Fig. 3c) and direct peptide binding sites (pockets)(Fig. 3d) had positive selective pressure, being significantfor most DRB exon 2 from the species analysed. AotusMHC-DRB and HLA-DRB presented similar values in bothcases, just like the whole exon.
Except for P1, primate DRB pockets (P1–P9, Figs. 3e–i)followed the trend observed in the PBR and the set ofpockets. P1 showed a tendency to have negative selectivepressure in most Catarrhini species including humans, whilstPlatyrrhini did not. Some species even exhibited significantpositive selective pressure, such as C. apella and A.nigriceps (Fig. 3e). It is notable that this highly specificlevel followed the same tendency seen when analysingvariability regarding low and high variability pockets andhigh and low variability species; a more diffuse associationwas observed between HLA-DRB and Aotus MHC-DRB.Even though HLA-DRB and Aotus MHC-DRB had a similaroverall pattern of diversity and dN cf dS, the way they accu-mulated changes at each pocket level varied more specifical-ly. Both Aotus and human species had the same tendenciesregarding selective pressures, except for P1; however, AotusMHC-DRB presented a pattern more like Catarrhini MHC-DRB, always being presented as an atypical NWM.
Some characteristics divided NWM (excluding Aotus)from OWM, humans and apes (such as greater variationand quasi-neutral non-PBR pattern and greater diversity inP1), pointing to specific specialisation between both orders.Except for this behaviour, selection tendencies in distinctprimate DRB exon 2 regions were generally similar.
Behaviour within each lineage was more conservativethan for the species’ set of DRB, each exhibiting its owncharacteristic stamp (data not shown). Such conservationcould have indicated functional specialisation within eachspecies; however, HLA-DRB and Aotus MHC-DRB line-ages shared characteristics a having common variation,
reflecting similar pressures and indicating a commonfunctional role.
Aotus showed a high degree of convergence with humanMHC-DRB at PBR
When only the PBR was considered and the same analysisperformed, it was then found that these relationships weregenerally maintained, but that more groups of Aotus MHC-DRB sequences converging with Catarrhini allele lineagesbecame clearly defined, being well above those presented inother Platyrrhini (data not shown). The relationshipbetween Aotus MHC-DRB and MHC-DRB allele lineagescharacteristic of humans and other Catarrhini became moreevident at the nucleotide and amino acid residue levels(being directly related to conforming pockets), presenting asignificant degree of identity amongst them and being veryclearly differentiated from MWM allele lineages. A tree forAotus and human sequences (Fig. 4) clearly showed theconvergences detected at this level between these twospecies’ DRB groups.
All Aotus MHC-DRB were grouped with human andother Catarrhini MHC-DRB sequences in exon 2 at thelevel of amino acids directly involved in peptide binding(35 residues). The following associations were found(Fig. 4), showing only associations with HLA-DRB:
1. HLA-DRB1* 08/11/12/13/14 converged with AotusMHC-DRB1*03 GB.
2. HLA-DRB1*1130 converged with Aotus MHC-DRB*W44.
3. HLA-DRB1*03 converged with Aotus MHC-DRB1*03GA.
4. HLA-DRB3 and HLA-DRB1*0422 converged withAotus MHC-DRB*W18 and DRB3*06 GA.
5. HLA-DRB1*04 and HLA-DRB1*1122 converged withAotus MHC-DRB*W45, W46 and W47.
6. HLA-DRB1*09 and HLA-DRB1*07 converged withAotus MHC-DRB*W30 and W38.
7. HLA-DRB4 and HLA-DRB1*10 converged with AotusMHC-DRB*W41 and W43.
8. HLA-DRB1*15 and DRB1*16 were related to AotusMHC-DRB*W29, W42, DRB6*03 GB, DRB1*03 GCand MHC-DRB*W13.
�Fig. 4 Minimum evolution tree for owl monkey–human MHC-DRBpocket positions. The tree was constructed using a P-distance matrixcalculated from aligning deduced amino acid sequences. Bootstrapvalues were calculated based on 1,000 replicates. Only values higherthan 60% are shown (>70% was significant); bold values indicatevalues that were also significant for internal branch test (IBT; >95%)and circles denote values only significant for IBT. Collapsed brancheswere denoted by triangles. Human and Aotus sequences are drawn ingrey and black, respectively. HLA DPB and DQB (grey) sequenceswere used as out-group. The scalebar represents 0.1 substitutions persite. See “Materials and methods” section for species’ abbreviations
552 Immunogenetics (2006) 58:542–558
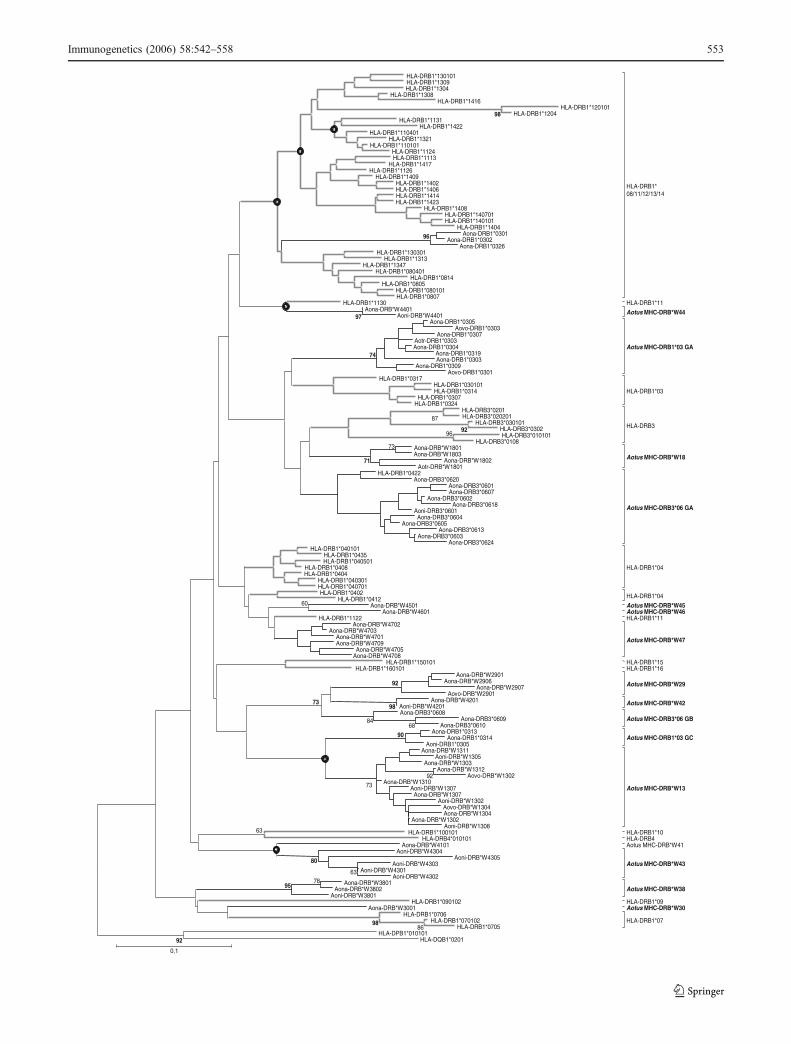
HLA-DRB1*130101HLA-DRB1*1309HLA-DRB1*1304
HLA-DRB1*1308HLA-DRB1*1416
HLA-DRB1*120101HLA-DRB1*1204
HLA-DRB1*1131HLA-DRB1*1422
HLA-DRB1*110401HLA-DRB1*1321
HLA-DRB1*110101HLA-DRB1*1124HLA-DRB1*1113
HLA-DRB1*1417HLA-DRB1*1126
HLA-DRB1*1409HLA-DRB1*1402HLA-DRB1*1406HLA-DRB1*1414HLA-DRB1*1423
HLA-DRB1*1408HLA-DRB1*140701HLA-DRB1*140101
HLA-DRB1*1404Aona-DRB1*0301
Aona-DRB1*0302Aona-DRB1*0326
HLA-DRB1*130301HLA-DRB1*1313
HLA-DRB1*1347HLA-DRB1*080401
HLA-DRB1*0814HLA-DRB1*0805
HLA-DRB1*080101HLA-DRB1*0807
HLA-DRB1*08/11/12/13/14
HLA-DRB1*11HLA-DRB1*1130Aona-DRB*W4401
Aoni-DRB*W4401 Aotus MHC-DRB*W44Aona-DRB1*0305
Aovo-DRB1*0303Aona-DRB1*0307
Aotr-DRB1*0303Aona-DRB1*0304
Aona-DRB1*0319Aona-DRB1*0303
Aona-DRB1*0309Aovo-DRB1*0301
Aotus MHC-DRB1*03 GA
HLA-DRB1*0317HLA-DRB1*030101HLA-DRB1*0314
HLA-DRB1*0307HLA-DRB1*0324
HLA-DRB1*03
HLA-DRB3*0201HLA-DRB3*020201
HLA-DRB3*030101HLA-DRB3*0302HLA-DRB3*010101
HLA-DRB3*0108
HLA-DRB3
Aona-DRB*W1801Aona-DRB*W1803
Aona-DRB*W1802Aotr-DRB*W1801
Aotus MHC-DRB*W18
HLA-DRB1*0422Aona-DRB3*0620
Aona-DRB3*0601Aona-DRB3*0607
Aona-DRB3*0602Aona-DRB3*0618
Aoni-DRB3*0601Aona-DRB3*0604
Aona-DRB3*0605Aona-DRB3*0613
Aona-DRB3*0603Aona-DRB3*0624
Aotus MHC-DRB3*06 GA
HLA-DRB1*040101HLA-DRB1*0435HLA-DRB1*040501
HLA-DRB1*0408HLA-DRB1*0404
HLA-DRB1*040301HLA-DRB1*040701
HLA-DRB1*04
HLA-DRB1*0402HLA-DRB1*0412 HLA-DRB1*04
Aotus MHC-DRB*W45Aona-DRB*W4501Aotus MHC-DRB*W46Aona-DRB*W4601HLA-DRB1*11HLA-DRB1*1122
Aona-DRB*W4702Aona-DRB*W4703
Aona-DRB*W4701Aona-DRB*W4709
Aona-DRB*W4705Aona-DRB*W4708
Aotus MHC-DRB*W47
HLA-DRB1*15HLA-DRB1*150101HLA-DRB1*16HLA-DRB1*160101
Aona-DRB*W2901Aona-DRB*W2906
Aona-DRB*W2907Aovo-DRB*W2901
Aotus MHC-DRB*W29
Aona-DRB*W4201Aoni-DRB*W4201 Aotus MHC-DRB*W42Aona-DRB3*0608
Aona-DRB3*0609Aona-DRB3*0610
Aotus MHC-DRB3*06 GB
Aona-DRB1*0313Aona-DRB1*0314
Aoni-DRB1*0305Aotus MHC-DRB1*03 GC
Aona-DRB*W1311Aoni-DRB*W1305
Aona-DRB*W1303Aona-DRB*W1312
Aovo-DRB*W1302Aona-DRB*W1310
Aoni-DRB*W1307Aona-DRB*W1307
Aoni-DRB*W1302Aovo-DRB*W1304Aona-DRB*W1304
Aona-DRB*W1302Aoni-DRB*W1308
Aotus MHC-DRB*W13
HLA-DRB1*10HLA-DRB1*100101HLA-DRB4HLA-DRB4*010101Aotus MHC-DRB*W41Aona-DRB*W4101
Aoni-DRB*W4304Aoni-DRB*W4305
Aoni-DRB*W4303Aoni-DRB*W4301
Aoni-DRB*W4302
Aotus MHC-DRB*W43
Aona-DRB*W3801Aona-DRB*W3802
Aoni-DRB*W3801Aotus MHC-DRB*W38
HLA-DRB1*09HLA-DRB1*090102Aotus MHC-DRB*W30Aona-DRB*W3001
HLA-DRB1*0706HLA-DRB1*070102
HLA-DRB1*0705HLA-DRB1*07
HLA-DPB1*010101HLA-DQB1*0201
8698
98
78
92
92
95
96
68
92
87
63
92
96
98
90
80
73
84
73
74
71
97
73
63
60
0,1
Immunogenetics (2006) 58:542–558 553
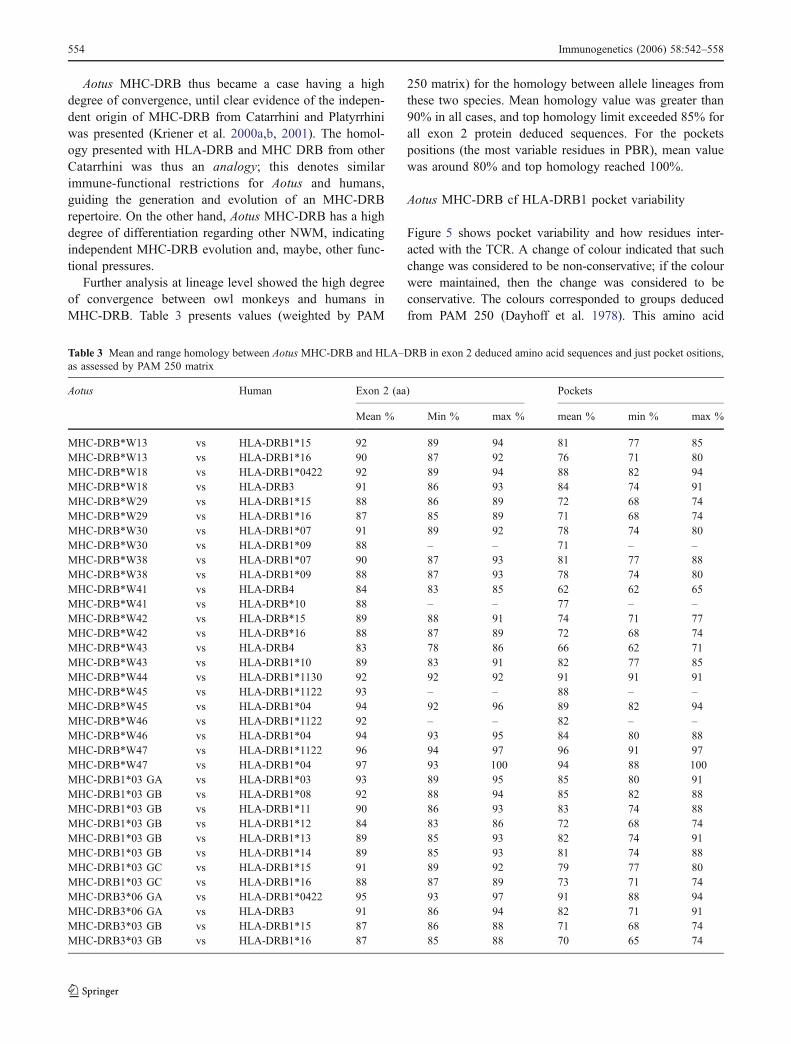
Aotus MHC-DRB thus became a case having a highdegree of convergence, until clear evidence of the indepen-dent origin of MHC-DRB from Catarrhini and Platyrrhiniwas presented (Kriener et al. 2000a,b, 2001). The homol-ogy presented with HLA-DRB and MHC DRB from otherCatarrhini was thus an analogy; this denotes similarimmune-functional restrictions for Aotus and humans,guiding the generation and evolution of an MHC-DRBrepertoire. On the other hand, Aotus MHC-DRB has a highdegree of differentiation regarding other NWM, indicatingindependent MHC-DRB evolution and, maybe, other func-tional pressures.
Further analysis at lineage level showed the high degreeof convergence between owl monkeys and humans inMHC-DRB. Table 3 presents values (weighted by PAM
250 matrix) for the homology between allele lineages fromthese two species. Mean homology value was greater than90% in all cases, and top homology limit exceeded 85% forall exon 2 protein deduced sequences. For the pocketspositions (the most variable residues in PBR), mean valuewas around 80% and top homology reached 100%.
Aotus MHC-DRB cf HLA-DRB1 pocket variability
Figure 5 shows pocket variability and how residues inter-acted with the TCR. A change of colour indicated that suchchange was considered to be non-conservative; if the colourwere maintained, then the change was considered to beconservative. The colours corresponded to groups deducedfrom PAM 250 (Dayhoff et al. 1978). This amino acid
Table 3 Mean and range homology between Aotus MHC-DRB and HLA–DRB in exon 2 deduced amino acid sequences and just pocket ositions,as assessed by PAM 250 matrix
Aotus Human Exon 2 (aa) Pockets
Mean % Min % max % mean % min % max %
MHC-DRB*W13 vs HLA-DRB1*15 92 89 94 81 77 85MHC-DRB*W13 vs HLA-DRB1*16 90 87 92 76 71 80MHC-DRB*W18 vs HLA-DRB1*0422 92 89 94 88 82 94MHC-DRB*W18 vs HLA-DRB3 91 86 93 84 74 91MHC-DRB*W29 vs HLA-DRB1*15 88 86 89 72 68 74MHC-DRB*W29 vs HLA-DRB1*16 87 85 89 71 68 74MHC-DRB*W30 vs HLA-DRB1*07 91 89 92 78 74 80MHC-DRB*W30 vs HLA-DRB1*09 88 – – 71 – –MHC-DRB*W38 vs HLA-DRB1*07 90 87 93 81 77 88MHC-DRB*W38 vs HLA-DRB1*09 88 87 93 78 74 80MHC-DRB*W41 vs HLA-DRB4 84 83 85 62 62 65MHC-DRB*W41 vs HLA-DRB*10 88 – – 77 – –MHC-DRB*W42 vs HLA-DRB*15 89 88 91 74 71 77MHC-DRB*W42 vs HLA-DRB*16 88 87 89 72 68 74MHC-DRB*W43 vs HLA-DRB4 83 78 86 66 62 71MHC-DRB*W43 vs HLA-DRB1*10 89 83 91 82 77 85MHC-DRB*W44 vs HLA-DRB1*1130 92 92 92 91 91 91MHC-DRB*W45 vs HLA-DRB1*1122 93 – – 88 – –MHC-DRB*W45 vs HLA-DRB1*04 94 92 96 89 82 94MHC-DRB*W46 vs HLA-DRB1*1122 92 – – 82 – –MHC-DRB*W46 vs HLA-DRB1*04 94 93 95 84 80 88MHC-DRB*W47 vs HLA-DRB1*1122 96 94 97 96 91 97MHC-DRB*W47 vs HLA-DRB1*04 97 93 100 94 88 100MHC-DRB1*03 GA vs HLA-DRB1*03 93 89 95 85 80 91MHC-DRB1*03 GB vs HLA-DRB1*08 92 88 94 85 82 88MHC-DRB1*03 GB vs HLA-DRB1*11 90 86 93 83 74 88MHC-DRB1*03 GB vs HLA-DRB1*12 84 83 86 72 68 74MHC-DRB1*03 GB vs HLA-DRB1*13 89 85 93 82 74 91MHC-DRB1*03 GB vs HLA-DRB1*14 89 85 93 81 74 88MHC-DRB1*03 GC vs HLA-DRB1*15 91 89 92 79 77 80MHC-DRB1*03 GC vs HLA-DRB1*16 88 87 89 73 71 74MHC-DRB3*06 GA vs HLA-DRB1*0422 95 93 97 91 88 94MHC-DRB3*06 GA vs HLA-DRB3 91 86 94 82 71 91MHC-DRB3*03 GB vs HLA-DRB1*15 87 86 88 71 68 74MHC-DRB3*03 GB vs HLA-DRB1*16 87 85 88 70 65 74
554 Immunogenetics (2006) 58:542–558
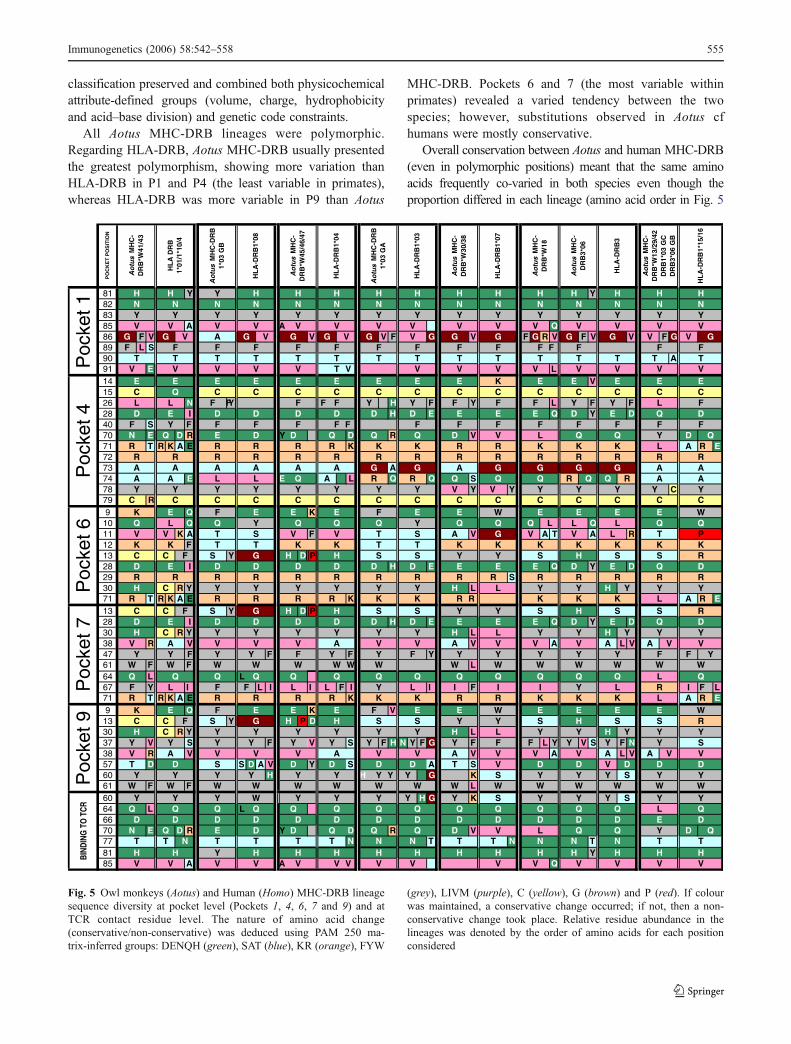
classification preserved and combined both physicochemicalattribute-defined groups (volume, charge, hydrophobicityand acid–base division) and genetic code constraints.
All Aotus MHC-DRB lineages were polymorphic.Regarding HLA-DRB, Aotus MHC-DRB usually presentedthe greatest polymorphism, showing more variation thanHLA-DRB in P1 and P4 (the least variable in primates),whereas HLA-DRB was more variable in P9 than Aotus
MHC-DRB. Pockets 6 and 7 (the most variable withinprimates) revealed a varied tendency between the twospecies; however, substitutions observed in Aotus cfhumans were mostly conservative.
Overall conservation between Aotus and human MHC-DRB(even in polymorphic positions) meant that the same aminoacids frequently co-varied in both species even though theproportion differed in each lineage (amino acid order in Fig. 5
81 Y Y828385 A A Q86 F V V V F G V F G R V F V V F G89 L S90 A91 E L14 V1526 N Y F H F Y L F F28 I H E Q Y D40 S F70 E D R Y D R V71 T R K A E K R E7273 A74 E E L Q Q S78 Y V Y C79 R9 Q K10 Q Q11 K A F V A T A R12 F13 Y D P28 I H E Q Y D29 S30 R Y L71 T R K A E K R E13 Y D P28 I H E Q Y D30 R Y L38 R V V A L V47 F F F Y61 F F L64 L L67 Y I L I I F I I F71 T R K A E K R E9 Q K V13 Y P D30 R Y L37 V S F V S F H N Y F G F L Y V S F N38 R V V A L V57 D S D A V Y S A S60 H H G K61 F F L60 H G K64 L L6670 E D R Y D R V77 N T N T81 Y85 A A Q
V
T
L
Y
Y
L Q
DY
DY
YVA VS
RS
LE W
A
L QR I
F
YVVA
RS
R
L A
RS
Q
CE W
A
QDA
F
Y
V
Y
L
A
Y
T
V G
HL
A-D
RB
1*15
/16
R
D
Q
Ao
tus
MH
C-
DR
B*W
13/2
9/42
D
RB
1*03
GC
D
RB
3*06
GB
Y
V
C
Q
R
Y
Q
H
Y
Q
Y
TH
V
QD
QT N
D
Y
HL
A D
RB
1*
01/1
*10/
4
H HN NN
HL
A-D
RB
1*04
V
F
CYA
D
C
YW
Q
C FE
CA
VG
FT
YV
G VFTT
C F
EC CF F
ER
LVK
E
FD Q
DD
A
E
AY
C CE E
EY
Q
R
VK K
RR R
YYA
A
W
DY YV AF
D
Ao
tus
MH
C-D
RB
1*
03 G
B
V
Y
H H
Ao
tus
MH
C-
D
RB
*W45
/46/
47
V
F
F
F
YR R
HD
R R
QV
C
Ao
tus
MH
C-
DR
B*W
41/4
3
HNY
VG
CY
R
D
H H
L LR R
E
Y YY Y
Ao
tus
MH
C-D
RB
1*
03 G
A
F
Y
Y
Y
T
G
Y
QT
Q
Y
T
D
Y
W
E
YW
E
TH H
Ao
tus
MH
C-
D
RB
*W18
Ao
tus
MH
C-
D
RB
*W30
/38
HL
A-D
RB
1*07
D D
G
Q
V V
HL
A-D
RB
1*08
DR
L
R
R
Y
Q
Q
GT
Ao
tus
MH
C-
D
RB
3*06
W
Y
K
HL
A-D
RB
1*03
YQ
E
F
T
Y
YS
R
S
R
Q
T
Y
WYY Y
WY Y
Y S
H YY
V DS
W
YE
SK
PO
CK
ET
PO
SIT
ION
N
FN
Y
D D D
YQ
NTQ D
T
Y
QF
FS
Y
S
L
C
D D D D
D
D
Y
A
V
H
I
A
Q Q
S
Y
Y
H
R
E
K
W
W EE
F
VD
Q
D V
R R
Q
YW
Y
R
R
A
H
H
R
E
Y
D DV V
YV V A
D
L I I
E
R
G
R
Q
T
Y
S
E
Y
E
F
Y
D D
Q
Y
R
VY
D
HR
V
KQ
N
Y
QF
C
T
W
Q
R
HF
E
W
WW
V
K
V
GE
VY
V
Y
Y
T
Y
D
Q
R
Y
HV V VV V
HV
H
L
E
VT
V
Y
W
H
S Y
HH
Q Q
Q
HN
DDDT
Q Q
F
WS
T
YW
Y
E
Y
Y
K K
Y
Q Q QW
YL
F Y
S
D
YL Y
R K K
Y
Q
YW
V
Y
YE
YW
E
V
Y YL
S
RR R
KH
KS
DES
EEC
E
YVCC
Q
Q Q
H Y Y
C G
YKY
A
KC
A L
KSS
E
A A AA
V
R
L QK
L
GGQ R
R
SR
W W
G
R R
S
R
R
R
V
R
W
D
D
QW
Y
K
F
KQ
R
EF F
FF F
FED D D
C
EF F F
RR
YL FLC
C C C C
C CC C CE E
C C C CE KE EE E EE E
Y
V
Y Y
TT
V V
VVV V V V VTT T TT T TT
GV V V
F F
VV
V
H
Y Y Y Y Y
HH H
Y
HH
FGG
N N N N
F FF FG G
N
F F FF
VA VG
BIND
ING
TO T
CR
NY Y Y
V
Poc
ket 4
Poc
ket 6
Poc
ket 7
Poc
ket 9
Poc
ket 1 N N
H Y H H
G
N
V
HL
A-D
RB
3
V
Q
W
K RS Y
V V
K
E
K
D D D E
V A
W
E
L
F
KR
Q
GR
Y
L
D
KH
Y
V VV
H
L
H
D
N
Y Y
S
V
Y
Y Y
S
A
H
Y
NH
Q Q
Q
W W
V
D
R
V
C
K
Y
LQ
Fig. 5 Owl monkeys (Aotus) and Human (Homo) MHC-DRB lineagesequence diversity at pocket level (Pockets 1, 4, 6, 7 and 9) and atTCR contact residue level. The nature of amino acid change(conservative/non-conservative) was deduced using PAM 250 ma-trix-inferred groups: DENQH (green), SAT (blue), KR (orange), FYW
(grey), LIVM (purple), C (yellow), G (brown) and P (red). If colourwas maintained, a conservative change occurred; if not, then a non-conservative change took place. Relative residue abundance in thelineages was denoted by the order of amino acids for each positionconsidered
Immunogenetics (2006) 58:542–558 555

corresponded to their frequency when appearing in allelelineage). The positions generally presented slight changesrespecting property conservation; however, some positionshad high variability, especially in P6, P7 and P9, as thesegenerally shared residues or types of property in HLA-DRBand Aotus MHC-DRB. Positions interacting with the TCR alsoshowed a high degree of conservation. It is worth noting thatpolar amino acids predominated in the positions involved(Fig. 5). The foregoing implies that similarity between humansand Aotus reflects a discretely structured DRB variabilitypattern, suggesting that both species’ MHC-DRB could besubmitted to the same functional demands within this sector(the most variable in the PBR).
Each lineage can be analysed in detail using theinformation in Fig. 5. For example, the Aotus lineagesMHC-DRB*W45, *W46 and W*47 (taken as a group)converge with HLA-DRB1*04, showing great homology atpocket level, ranging from 80 to 100% and having 95%mean (Table 3). Figure 5 shows that variant position 86 inP1 presented G and V in both species, these residues’alternation being the tendency for other allele lineages inboth HLA-DRB and Aotus MHC-DRB. Other Aotus MHC-DRB lineages also showed F in this position.
These changes may be considered non-conservative byPAM 250, indicating major changes in physicochemical-genetic code attributes. V and G were separated by onenucleotide substitution in the second position at geneticcode level (V: GGN→G: GTN), and F was separated fromV by a change in first position (F: TTY→V: GTT, GTC).The interchange between G and F could only occur as V asan intermediary state; this change may easily have beenpresented by point mutation in positive selection stressedregions. These V and G changes shared similar surface areain folded proteins (V, 23.7 Å2 and G, 24.5 Å2) (Janin et al.1978) and the same charge. F presented a similar surfacearea as V and G (25.5 Å2) and the same polarity but sharedhigh hydrophobicity and non-polar nature with V. Themajor change between these three residues was presented interms of volume (G, 60 Å3; F, 189 Å3; V, 141.4 Å3;Goldsack and Chalifoux 1973).
This example indicates that a single point mutation mayexplain the emergence of non-conservative polymorphismin several pocket positions having variable physicochemicalproperty conservation in both HLA-DRB and Aotus MHC-DRB. For example, previously analysed lineage positions40 (F→L) and 70 (D→Y) in P4, 9 (E→K), 11 (V→F) and13 (H→P, H→D) in P6, 64 (Q→L) and 67 (L→F, I→F) inP7, 37 (Y→V, Y→S), 57 (D→Y) and 60 (Y→H) in P9 andin TCR binding position 70 (D→Y), 77 (T→N) and 91(V→E) may be explained by this process. In most cases,these non-conservative changes implied fewer sequencescompared to the predominant residue in the allele lineage.
This pattern explained most polymorphic non-conserva-tive changes in the remaining lineages shown in Fig. 5.Very distinct physicochemical properties were presented toa much lesser degree than other changes involving moresteps at the codon level. This fact, the high degree ofhomology and the co-variation exhibited could implysimilar Aotus MHC-DRB and HLA-DRB peptide presenta-tion ability, highlighting the similarity between these twospecies’ immune systems.
Concluding remarks
The hallmark of classic MHC in all vertebrates is theirenormous polymorphism as a response to the dynamics ofthe host–pathogen relationship. Such polymorphism isactively maintained by adaptive evolution in the form ofbalanced selection (Hughes and Yeager 1998). Mutationand recombination processes (cross-over, micro-recombi-nation and gene conversion) have contributed towardsmoulding MHC polymorphism (Ohta 1991; Yeager andHughes 1999). They could be trans-specific (TSP), as allelevariants increasing an individual’s suitability are maintainedbeyond the speciation process, leading to different speciesexhibiting common allele lineages (Klein et al. 1998).Natural selection could also have provoked a level ofmolecular PBR convergence amongst distantly relatedMHC alleles, indicating that the need for adapting MHCto peptide presentation may have optimum solutions at thesequence level (O’hUigin 1995). Both processes haveproduced similarity in divergent species’ PBR, clearly seenin the case of Catarrhini and Platyrrhini MHC-DRBevolution (Trtkova et al. 1995; Kriener et al. 2000a,b,2001). However, whilst similarity between genes caused byTSP denotes homology, similarity due to convergencedenotes functional analogy, as could be occurring inHLA-DRB cf Aotus MHC-DRB.
Aotus MHC-DRB was highly idiosyncratic (presentinghigh recombination rates) and preferentially presentedassociation with MHC-DRB from humans, apes andOWM rather than with NWM MHC-DRB. Aotus MHC-DRB extensively converged with HLA-DRB. This conver-gence with HLA-DRB happened at different levels; itoccurred in the amount of overall variability, types ofsubstitution found, similar natural selective pressures andcommon pocket amino acid property signatures at thesequence level. The functional implications of suchconvergence are evident. Aotus has obvious potentialamongst NWM as a promising animal model for thecomparative study of processes involving peptide presenta-tion with MHC-DRB. The overall evidence regarding bio-logical similarity between human and owl monkey immune
556 Immunogenetics (2006) 58:542–558

systems justifies its use as an excellent animal model forpharmacological and vaccine-development research.
Acknowledgements This research has been supported by theColombian President’s Office and the Colombian Ministry of SocialProtection. We would like to thank Jason Garry for patientlyreviewing the manuscript.
References
Antunes SG, de Groot NG, Brok H, Doxiadis G, Menezes AA, OttingN, Bontrop RE (1998) The common marmoset: a new worldprimate species with limited Mhc class II variability. Proc NatlAcad Sci USA 95:11745–11750
Bergstrom TF, Erlandsson R, Engkvist H, Josefsson A, Erlich HA,Gyllensten U (1999) Phylogenetic history of hominoid DRB lociand alleles inferred from intron sequences. Immunol Rev167:351–365
Collins WE (1994) The owl monkey as a model for malaria. In: BaerJF, Weller RE, Kakoma I (eds) Aotus: the owl monkey.Academic, New York, pp 245–258
Dayhoff M, Schwartz R, Orcutt B (1978) A model of evolutionarychange in proteins. In: Dayhoff M (ed) Atlas of protein sequenceand structure, vol. 5. National Biomedical Research Foundation,Washington, DC, pp 345–352
Di Rienzo A, Wilson AC (1991) Branching pattern in the evolutionarytree for human mitochondrial DNA. Proc Natl Acad Sci USA88:1597–1601
Doxiadis GG, Otting N, de Groot NG, Noort R, Bontrop RE (2000)Unprecedented polymorphism of Mhc-DRB region configurationsin rhesus macaques. J Immunol 164:3193–3199
Doxiadis GG, Otting N, de Groot NG, Bontrop RE (2001) Differentialevolutionary MHC class II strategies in humans and rhesusmacaques: relevance for biomedical studies. Immunol Rev183:76–85
Edwards S, Hedirck P (1998) Evolution and ecology of MHC mole-cules: from genomics to sexual selection. TREE 13(8)305–311
Figueroa F, Mayer WE, Sultmann H, O’hUigin C, Tichy H, Satta Y,Takezaki N, Takahata N, Klein J (2000) Mhc class II B gene evolu-tion in East African cichlid fishes. Immunogenetics 51:556–575
Garraud O, Perraut R, Gysin J, Behr C, Dubois P, Bonnemains B,Jouin H, Michel JC, Pereira da Silva L (1994) Manipulatingblood T cells and B cells from squirrel monkeys: some technicalconsiderations. J Immunol Methods 173:165–173
Go Y, Satta Y, Kawamoto Y, Rakotoarisoa G, Randrianjafy A,Koyama N, Hirai H (2002) Mhc-DRB genes evolution in lemurs.Immunogenetics 54:403–417
Goldsack DE, Chalifoux RC (1973) Contribution of the free energy ofmixing of hydrophobic side chains to the stability of the tertiarystructure of proteins. J Theor Biol 39:645–651
Gyllensten U, Bergstrom T, Josefsson A, Sundvall M, Savage A,Blumer ES, Giraldo LH, Soto LH, Watkins DI (1994) Thecotton-top tamarin revisited: Mhc class I polymorphism of wildtamarins, and polymorphism and allele diversity of the class IIDQA1, DQB1, and DRB loci. Immunogenetics 40:167–176
Gysin J (1988) Animal models: primates. In: Sherman IW (ed)Malaria: parasite biology, pathogenesis and protection. ASM,Washington DC, pp 419–439
Hillis DM (1993) An empirical test of bootstrapping as a method forassessing confidence in phylogenetic analysis. Syst Biol 42:182–192
Hughes AL, Yeager M (1998) Natural selection at major histocompat-ibility complex loci of vertebrates. Annu Rev Genet 32:415–35
Janin J, Wodak S, Levitt M, Maigret B (1978) The conformation ofamino acid side chains in proteins. J Mol Biol 125:357–386
Jukes TH (1969) Evolution of protein molecules. In: Munro HN(ed) Mammalian protein metabolism. Academic, New York, pp21–132
Kaufman JF, Auffray C, Korman AJ, Shackelford DA, Strominger J(1984) The class II molecules of the human and murine majorhistocompatibility complex. Cell 36:1–13
Kenter M, Otting N, Anholts J, Jonker M, Schipper R, Bontrop RE(1992) Mhc-DRB diversity of the chimpanzee (Pan troglodytes).Immunogenetics 37:1–11
Kimura M (1980) A simple method for estimating evolutionary ratesof base substitutions through comparative studies of nucleotidesequences. J Mol Evol 16:111–120
Klein J, Bontrop RE, Dawkins RL, Erlich HA, Gyllensten UB, HeiseER, Jones PP, Parham P, Wakeland EK, Watkins DI (1990)Nomenclature for the major histocompatibility complexes ofdifferent species: a proposal. Immunogenetics 31:217–219
Klein J, Sato A, Nagl S, O’hUigín C (1998) Molecular Trans-SpeciesPolymorphism. Annu Rev Ecol Syst 29:1–21
Kriener K, O’hUigin C, Klein J (2000a) Alu elements supportindependent origin of prosimian, platyrrhine, and catarrhineMhc-DRB genes. Genome Res 5:634–643
Kriener K, O’hUigin C, Tichy H, Klein J (2000b) Convergentevolution of major histocompatibility complex molecules inhumans and New World monkeys. Immunogenetics 51:169–178
Kriener K, O’hUigin C, Klein J (2001) Independent origin offunctional MHC class II genes in humans and New Worldmonkeys. Hum Immunol 62:1–14
Kumar S, Tamura K, Jakobsen IB, Nei M (2001) MEGA2: molecularevolutionary genetics analysis software. Bioinformatics 17:1244–1245
Mayer WE, O’hUigin C, Zaleska-Rutczynska Z, Klein J (1992) Trans-species origin of Mhc-DRB polymorphism in the chimpanzee.Immunogenetics 37:12–23
Middleton S, Anzenberger G, Knapp L (2004) Identification of NewWorld monkey MHC-DRB alleles using PCR, DGGE and directsequencing. Immunogenetics 55:785–790
Nei M, Gojobori T (1986) Simple methods for estimating the numbersof synonymous and nonsynonymous nucleotide substitutions.Mol Biol Evol 3:418–426
Niño-Vasquez JJ, Vogel D, Rodriguez R, Moreno A, Patarroyo ME,Pluschke G, Daubenberger CA (2000) Sequence and diversity ofDRB genes of Aotusnancymaae, a primate model for humanmalaria parasites. Immunogenetics 51:219–230
Ohta T (1991) Role of diversifying selection and gene conversion inevolution of Major Histocompatibility Complex loci. Proc NatlAcad Sci USA 15:6716–6720
O’hUigin C (1995) Quantifying the degree of convergence in primateMhc-DRB genes. Immunol Rev 143:123–140
Otting N, de Groot NG, Noort MC, Doxiadis GG, Bontrop RE (2000)Allele diversity of Mhc-DRB alleles in rhesus macaques. TissueAntigens 56:58–68
Pico de Coaña Y, Rodriguez J, Guerrero E, Barrero C, Rodriguez R,Mendoza M, Patarroyo MA (2003) A highly infective Plasmo-dium vivax strain adapted to Aotus monkeys: quantitativehaematological and molecular determinations useful for P. vivaxmalaria vaccine development. Vaccine 21:3930–3937
Rodriguez R, Moreno A, Guzman F, Calvo M, Patarroyo ME (1990)Studies in owl monkeys leading to the development of asynthetic vaccine against the asexual blood stages of Plasmodiumfalciparum. Am J Trop Med Hyg 43:339–354
Rosal-SanchezM, Paz-Artal E, Moreno-PelayoMA,Martinez-Quiles N,Martinez-Laso J, Martin-Villa JM, Arnaiz-Villena A (1998)Polymorphism of Mhc-DRB alleles in Cercopithecus aethiops
Immunogenetics (2006) 58:542–558 557

(green monkey): generation and functionality. Tissue Antigens51:541–548
RzhetskyA,NeiM (1993) Theoretical foundation of theminimum-evolutionmethod of phylogenetic inference. Mol Biol Evol 10:1073–1095
Satta Y, Mayer WE, Klein J (1996) Evolutionary relationship of HLA-DRB genes inferred from intron sequences. J Mol Evol 42:648–657
Sawyer S (1989) Statistical tests for detecting gene conversion. MolBiol Evol 6:526–538
Sawyer SA (1999) GENECONV: A computer package for thestatistical detection of gene conversion. Distributed by the author.http://www.math.wustl.edu/~sawyer/geneconv/ Department ofMathematics, Washington University in St. Louis
Setoguchi T, Rosenberger AL (1987) A fossil owl monkey from LaVenta, Colombia. Nature 326(6114):692–694
Sitnikova T (1996) Bootstrap method of interior branch test forphylogenetic trees. Mol Biol Evol 13:605–611
Stern LJ, Brown JH, Jardetzky TS, Gorga JC, Urban RG, StromingerJL, Wiley DC (1994) Crystal structure of the human class IIMHC protein HLA-DR1 complexed with an influenza viruspeptide. Nature 368:215–221
Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W:improving the sensitivity of progressive multiple sequencealignment through sequence weighting, position-specific gappenalties and weight matrix choice. Nucleic Acids Res22:4673–4680
Trtkova K, Mayer WE, O’Huigin C, Klein J (1995) Mhc-DRB genesand the origin of New World monkeys. Mol Phylogenet Evol4:408–419
Wu MS, Tani K, Sugiyama H, Hibino H, Izawa K, Tanabe T,Nakazaki Y, Ishii H, Ohashi J, Hohjoh H, Iseki T, Tojo A,Nakamura Y, Tanioka Y, Tokunaga K, Asano S (2000) MHC(major histocompatibility complex)-DRB genes and polymor-phisms in common marmoset. J Mol Evol 51:214–222
Yeager M, Hughes AL (1999) Evolution of the mammalian MHC:natural selection, recombination, and convergent evolution.Immunol Rev 167:45–58
Zhu ZF, Vincek V, Figueroa F, Schonbach C, Klein J (1991) Mhc-DRB genes of the pigtail macaque (Macaca nemestrina):implications for the evolution of human DRB genes. Mol BiolEvol 8:563–578
558 Immunogenetics (2006) 58:542–558




















