
Ang-(1–7) activates the NO/cGMP and ATP-sensitive K+ channels pathway to induce peripheral...
6
Ang-(1–7) activates the NO/cGMP and ATP-sensitive K + channels pathway to induce peripheral antinociception in rats Aline Costa a , Giovane Galdino a , Thiago Romero a , Grazielle Silva a , Steyner Cortes a , Robson Santos b , Igor Duarte a,⇑ a Department of Pharmacology, Institute of Biological Sciences, Federal University of Minas Gerais, Belo Horizonte, Brazil b Department of Physiology, Institute of Biological Sciences, Federal University of Minas Gerais, Belo Horizonte, Brazil article info Article history: Received 8 August 2013 Revised 11 November 2013 Available online 17 December 2013 Keywords: Angiotensin-(1–7) Nitric oxide K þ ATP Pain Antinociception abstract Angiotensin-(1–7) is a bioactive component of the renin–angiotensin system that is formed endoge- nously and induces nitric oxide release in several tissues. The L-arginine/NO/cyclic GMP pathway and ATP-sensitive K + channels have been proposed as the mechanism of action for the peripheral antinocicep- tion of several groups of drug and endogenous substances, including opioids, non-steroidal analgesics, acetylcholine and others. The aim of the present study was to investigate the involvement of the L-argi- nine/NO/cGMP and K þ ATP pathway on antinociception induced by angiotensin-(1–7). Paw pressure in rats was used to induce hyperalgesia via an intraplantar injection of prostaglandin E 2 (2 lg/paw). Ang-(1–7) (2, 3 and 4 lg/paw) elicited a local peripheral antinociceptive effect that was antagonized by the nonse- lective NO synthase (NOS) inhibitor L-NOarg and the selective neuronal NOS (nNOS) inhibitor L-NPA. The selective inhibition of endothelial (eNOS) and inducible (iNOS) NOS by L-NIO and L-NIL, respectively, was ineffective at blocking the effects of a local Ang-(1–7) injection. In addition, the level of nitrite in the homogenized paw tissue, as determined by a colorimetric assay, indicated that exogenous Ang-(1–7) is able to induce NO release. The soluble guanylyl cyclase inhibitor ODQ and the specific blocker of ATP-sen- sitive K + channels glibenclamide (40, 80 and 160 lg/paw) antagonized the Ang-(1–7) response. The results provide evidence that Ang-(1–7) most likely induces peripheral antinociceptive effects via the L-arginine/NO/cGMP pathway and K þ ATP pathway activation. Ó 2014 Published by Elsevier Inc. Introduction The renin–angiotensin system (RAS) plays a critical role in blood pressure control and body fluid and electrolyte homeostasis. However, the new expanded view of RAS has shown that it regu- lates the function of many more biological functions than previ- ously believed. In addition to the circulation RAS, there is a local tissue RAS in most organs and tissues studied, including the central nervous and the reproductive system, among others [1–4]. In addi- tion to angiotensin (Ang) II, other angiotensin peptides, such as Ang III [Ang-(2–8)], Ang IV [Ang-(3–8)], and Ang-(1–7), have also been shown to have important biological activities [5]. Ang-(1–7) has become an angiotensin of interest in the past few years due to the importance of the new RAS branch formed by Ang-(1–7) and its receptor Mas in many physiological systems [6]. The biological activities mediated by Ang-(1–7) have been shown to be closely related to NO release [4,7,8]. NO has been associated with numerous functions, including anti-nociception [9,10]. The molecular mechanism of this process begins with the activation of nitric oxide (NO) synthase, which produces the sec- ond messenger NO. Once generated, NO activates the guanylate cy- clase enzyme, which is directly responsible for increasing intracellular levels of cGMP [11]. ATP-sensitive K + channels K þ ATP have been implicated in several studies as the final step of the peripheral antinociceptive mecha- nism of various analgesic drugs [12–14]. When NO increases the intracellular level of cGMP, it can activate K þ ATP . Based on experi- mental evidence that the peripheral antinociceptive effects of NO donors and exogenous cGMP analogs are mediated by K þ ATP activa- tion [15,16], it has been proposed that a link exists between the L-arginine/NO/cGMP pathway and K þ ATP in the antinociceptive model. Furthermore, there is much evidence that the l-arginine=NO= cGMP=K þ ATP pathway plays an important role in the peripheral antinociception induced by cannabinoids anandamide [12,17] and N-palmitoyl-ethanolamine [18,19]. It was recently demonstrated that Ang-(1–7) exerts antinoci- ceptive activity that is likely mediated by Mas receptor activation, and there is no evidence for the participation of opioid peptides in 1089-8603/$ - see front matter Ó 2014 Published by Elsevier Inc. http://dx.doi.org/10.1016/j.niox.2013.12.007 ⇑ Corresponding author. Address: Department of Pharmacology, ICB-UFMG, Av. Antônio Carlos, 6627, Campus da Pampulha, Belo Horizonte, MG, Brasil, CEP 31270- 100, Brazil. Fax: +55 31 34092695. E-mail address: [email protected] (I. Duarte). Nitric Oxide 37 (2014) 11–16 Contents lists available at ScienceDirect Nitric Oxide journal homepage: www.elsevier.com/locate/yniox
Transcript of Ang-(1–7) activates the NO/cGMP and ATP-sensitive K+ channels pathway to induce peripheral...

Nitric Oxide 37 (2014) 11–16
Contents lists available at ScienceDirect
Nitric Oxide
journal homepage: www.elsevier .com/locate /yniox
Ang-(1–7) activates the NO/cGMP and ATP-sensitive K+ channelspathway to induce peripheral antinociception in rats
1089-8603/$ - see front matter � 2014 Published by Elsevier Inc.http://dx.doi.org/10.1016/j.niox.2013.12.007
⇑ Corresponding author. Address: Department of Pharmacology, ICB-UFMG, Av.Antônio Carlos, 6627, Campus da Pampulha, Belo Horizonte, MG, Brasil, CEP 31270-100, Brazil. Fax: +55 31 34092695.
E-mail address: [email protected] (I. Duarte).
Aline Costa a, Giovane Galdino a, Thiago Romero a, Grazielle Silva a, Steyner Cortes a, Robson Santos b,Igor Duarte a,⇑a Department of Pharmacology, Institute of Biological Sciences, Federal University of Minas Gerais, Belo Horizonte, Brazilb Department of Physiology, Institute of Biological Sciences, Federal University of Minas Gerais, Belo Horizonte, Brazil
a r t i c l e i n f o
Article history:Received 8 August 2013Revised 11 November 2013Available online 17 December 2013
Keywords:Angiotensin-(1–7)Nitric oxideKþATPPainAntinociception
a b s t r a c t
Angiotensin-(1–7) is a bioactive component of the renin–angiotensin system that is formed endoge-nously and induces nitric oxide release in several tissues. The L-arginine/NO/cyclic GMP pathway andATP-sensitive K+ channels have been proposed as the mechanism of action for the peripheral antinocicep-tion of several groups of drug and endogenous substances, including opioids, non-steroidal analgesics,acetylcholine and others. The aim of the present study was to investigate the involvement of the L-argi-nine/NO/cGMP and KþATP pathway on antinociception induced by angiotensin-(1–7). Paw pressure in ratswas used to induce hyperalgesia via an intraplantar injection of prostaglandin E2 (2 lg/paw). Ang-(1–7)(2, 3 and 4 lg/paw) elicited a local peripheral antinociceptive effect that was antagonized by the nonse-lective NO synthase (NOS) inhibitor L-NOarg and the selective neuronal NOS (nNOS) inhibitor L-NPA. Theselective inhibition of endothelial (eNOS) and inducible (iNOS) NOS by L-NIO and L-NIL, respectively, wasineffective at blocking the effects of a local Ang-(1–7) injection. In addition, the level of nitrite in thehomogenized paw tissue, as determined by a colorimetric assay, indicated that exogenous Ang-(1–7) isable to induce NO release. The soluble guanylyl cyclase inhibitor ODQ and the specific blocker of ATP-sen-sitive K+ channels glibenclamide (40, 80 and 160 lg/paw) antagonized the Ang-(1–7) response. Theresults provide evidence that Ang-(1–7) most likely induces peripheral antinociceptive effects via theL-arginine/NO/cGMP pathway and KþATP pathway activation.
� 2014 Published by Elsevier Inc.
Introduction
The renin–angiotensin system (RAS) plays a critical role inblood pressure control and body fluid and electrolyte homeostasis.However, the new expanded view of RAS has shown that it regu-lates the function of many more biological functions than previ-ously believed. In addition to the circulation RAS, there is a localtissue RAS in most organs and tissues studied, including the centralnervous and the reproductive system, among others [1–4]. In addi-tion to angiotensin (Ang) II, other angiotensin peptides, such asAng III [Ang-(2–8)], Ang IV [Ang-(3–8)], and Ang-(1–7), have alsobeen shown to have important biological activities [5]. Ang-(1–7)has become an angiotensin of interest in the past few years dueto the importance of the new RAS branch formed by Ang-(1–7)and its receptor Mas in many physiological systems [6].
The biological activities mediated by Ang-(1–7) have beenshown to be closely related to NO release [4,7,8]. NO has been
associated with numerous functions, including anti-nociception[9,10]. The molecular mechanism of this process begins with theactivation of nitric oxide (NO) synthase, which produces the sec-ond messenger NO. Once generated, NO activates the guanylate cy-clase enzyme, which is directly responsible for increasingintracellular levels of cGMP [11].
ATP-sensitive K+ channels KþATP have been implicated in severalstudies as the final step of the peripheral antinociceptive mecha-nism of various analgesic drugs [12–14]. When NO increases theintracellular level of cGMP, it can activate KþATP. Based on experi-mental evidence that the peripheral antinociceptive effects of NOdonors and exogenous cGMP analogs are mediated by KþATP activa-tion [15,16], it has been proposed that a link exists between theL-arginine/NO/cGMP pathway and KþATP in the antinociceptivemodel.
Furthermore, there is much evidence that the l-arginine=NO=cGMP=KþATP pathway plays an important role in the peripheralantinociception induced by cannabinoids anandamide [12,17]and N-palmitoyl-ethanolamine [18,19].
It was recently demonstrated that Ang-(1–7) exerts antinoci-ceptive activity that is likely mediated by Mas receptor activation,and there is no evidence for the participation of opioid peptides in
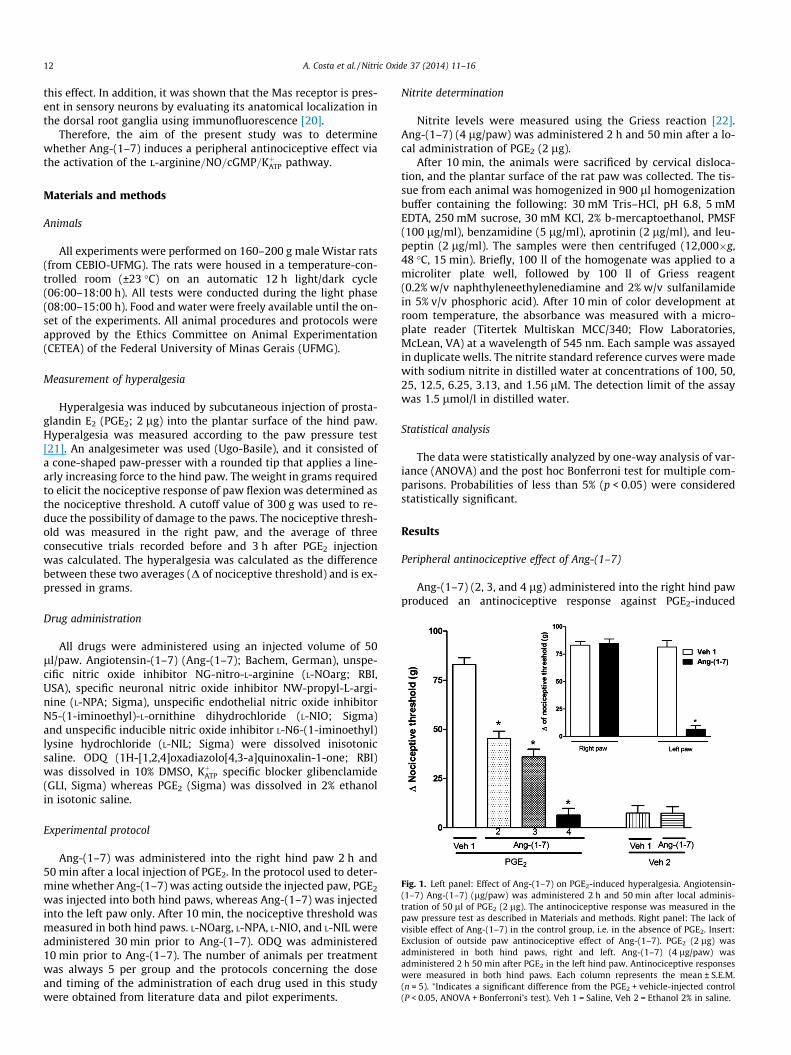
12 A. Costa et al. / Nitric Oxide 37 (2014) 11–16
this effect. In addition, it was shown that the Mas receptor is pres-ent in sensory neurons by evaluating its anatomical localization inthe dorsal root ganglia using immunofluorescence [20].
Therefore, the aim of the present study was to determinewhether Ang-(1–7) induces a peripheral antinociceptive effect viathe activation of the l-arginine=NO=cGMP=KþATP pathway.
Fig. 1. Left panel: Effect of Ang-(1–7) on PGE2-induced hyperalgesia. Angiotensin-(1–7) Ang-(1–7) (lg/paw) was administered 2 h and 50 min after local adminis-tration of 50 ll of PGE2 (2 lg). The antinociceptive response was measured in thepaw pressure test as described in Materials and methods. Right panel: The lack ofvisible effect of Ang-(1–7) in the control group, i.e. in the absence of PGE2. Insert:Exclusion of outside paw antinociceptive effect of Ang-(1–7). PGE2 (2 lg) wasadministered in both hind paws, right and left. Ang-(1–7) (4 lg/paw) wasadministered 2 h 50 min after PGE2 in the left hind paw. Antinociceptive responseswere measured in both hind paws. Each column represents the mean ± S.E.M.(n = 5). ⁄Indicates a significant difference from the PGE2 + vehicle-injected control(P < 0.05, ANOVA + Bonferroni’s test). Veh 1 = Saline, Veh 2 = Ethanol 2% in saline.
Materials and methods
Animals
All experiments were performed on 160–200 g male Wistar rats(from CEBIO-UFMG). The rats were housed in a temperature-con-trolled room (±23 �C) on an automatic 12 h light/dark cycle(06:00–18:00 h). All tests were conducted during the light phase(08:00–15:00 h). Food and water were freely available until the on-set of the experiments. All animal procedures and protocols wereapproved by the Ethics Committee on Animal Experimentation(CETEA) of the Federal University of Minas Gerais (UFMG).
Measurement of hyperalgesia
Hyperalgesia was induced by subcutaneous injection of prosta-glandin E2 (PGE2; 2 lg) into the plantar surface of the hind paw.Hyperalgesia was measured according to the paw pressure test[21]. An analgesimeter was used (Ugo-Basile), and it consisted ofa cone-shaped paw-presser with a rounded tip that applies a line-arly increasing force to the hind paw. The weight in grams requiredto elicit the nociceptive response of paw flexion was determined asthe nociceptive threshold. A cutoff value of 300 g was used to re-duce the possibility of damage to the paws. The nociceptive thresh-old was measured in the right paw, and the average of threeconsecutive trials recorded before and 3 h after PGE2 injectionwas calculated. The hyperalgesia was calculated as the differencebetween these two averages (D of nociceptive threshold) and is ex-pressed in grams.
Drug administration
All drugs were administered using an injected volume of 50ll/paw. Angiotensin-(1–7) (Ang-(1–7); Bachem, German), unspe-cific nitric oxide inhibitor NG-nitro-L-arginine (L-NOarg; RBI,USA), specific neuronal nitric oxide inhibitor NW-propyl-L-argi-nine (L-NPA; Sigma), unspecific endothelial nitric oxide inhibitorN5-(1-iminoethyl)-L-ornithine dihydrochloride (L-NIO; Sigma)and unspecific inducible nitric oxide inhibitor L-N6-(1-iminoethyl)lysine hydrochloride (L-NIL; Sigma) were dissolved inisotonicsaline. ODQ (1H-[1,2,4]oxadiazolo[4,3-a]quinoxalin-1-one; RBI)was dissolved in 10% DMSO, KþATP specific blocker glibenclamide(GLI, Sigma) whereas PGE2 (Sigma) was dissolved in 2% ethanolin isotonic saline.
Experimental protocol
Ang-(1–7) was administered into the right hind paw 2 h and50 min after a local injection of PGE2. In the protocol used to deter-mine whether Ang-(1–7) was acting outside the injected paw, PGE2
was injected into both hind paws, whereas Ang-(1–7) was injectedinto the left paw only. After 10 min, the nociceptive threshold wasmeasured in both hind paws. L-NOarg, L-NPA, L-NIO, and L-NIL wereadministered 30 min prior to Ang-(1–7). ODQ was administered10 min prior to Ang-(1–7). The number of animals per treatmentwas always 5 per group and the protocols concerning the doseand timing of the administration of each drug used in this studywere obtained from literature data and pilot experiments.
Nitrite determination
Nitrite levels were measured using the Griess reaction [22].Ang-(1–7) (4 lg/paw) was administered 2 h and 50 min after a lo-cal administration of PGE2 (2 lg).
After 10 min, the animals were sacrificed by cervical disloca-tion, and the plantar surface of the rat paw was collected. The tis-sue from each animal was homogenized in 900 ll homogenizationbuffer containing the following: 30 mM Tris–HCl, pH 6.8, 5 mMEDTA, 250 mM sucrose, 30 mM KCl, 2% b-mercaptoethanol, PMSF(100 lg/ml), benzamidine (5 lg/ml), aprotinin (2 lg/ml), and leu-peptin (2 lg/ml). The samples were then centrifuged (12,000�g,48 �C, 15 min). Briefly, 100 ll of the homogenate was applied to amicroliter plate well, followed by 100 ll of Griess reagent(0.2% w/v naphthyleneethylenediamine and 2% w/v sulfanilamidein 5% v/v phosphoric acid). After 10 min of color development atroom temperature, the absorbance was measured with a micro-plate reader (Titertek Multiskan MCC/340; Flow Laboratories,McLean, VA) at a wavelength of 545 nm. Each sample was assayedin duplicate wells. The nitrite standard reference curves were madewith sodium nitrite in distilled water at concentrations of 100, 50,25, 12.5, 6.25, 3.13, and 1.56 lM. The detection limit of the assaywas 1.5 lmol/l in distilled water.
Statistical analysis
The data were statistically analyzed by one-way analysis of var-iance (ANOVA) and the post hoc Bonferroni test for multiple com-parisons. Probabilities of less than 5% (p < 0.05) were consideredstatistically significant.
Results
Peripheral antinociceptive effect of Ang-(1–7)
Ang-(1–7) (2, 3, and 4 lg) administered into the right hind pawproduced an antinociceptive response against PGE2-induced

Fig. 3. Left panel: Antagonism induced by intraplantar administration of L-NPA ofthe peripheral antinociception induced by Ang-(1–7) in hyperalgesic paws (PGE2,2 lg). L-NPA (lg/paw) was administered 30 min prior to Ang-(1–7) (4 lg/paw).Right panel: The lack of effects of L-NPA 24 in the control groups, i.e. in the absenceof Ang-(1–7). Each column represents the mean ± S.E.M. (n = 4–5). # and ⁄ indicate asignificant difference compared to (PGE2 + veh 1 + veh 1) and (PGE2 + veh 1 + Ang-(1–7) 4)-injected controls, respectively (P < 0.05, ANOVA + the Bonferroni test). Veh1 = Saline, Veh 2 = Ethanol 2% in saline.
A. Costa et al. / Nitric Oxide 37 (2014) 11–16 13
hyperalgesia (2 lg/paw) in a dose-dependent manner (Fig. 1). Ang-(1–7) at the dose of 4 lg/paw when injected alone did not alter thenociceptive threshold. The protocol used to determine whetherAng-(1–7) was acting outside the injection paw demonstrated thatAng-(1–7) at a dose of 4 lg did not produce an antinociceptive ef-fect in the left paw, indicating that at this dose, Ang-(1–7) inducedan antinociceptive effect only at the injection site (Fig. 1).
Involvement of the L-Arginine/NO/cGMP Pathway in peripheralAng-(1–7)-induced antinociception
The peripheral antinociceptive effect of Ang-(1–7) (4 lg/paw)was abolished by the NOS inhibitor L-NOarg (12, 18, and 24 lg/paw) in a dose-dependent manner (Fig. 2, on left). L-NOarg didnot induce hyperalgesia or antinociception alone (Fig. 2, on right).Similarly, the selective nNOS inhibitor L-NPA (12, 18, and 24 lg/paw) completely antagonized the action of peripheral Ang-(1–7)(Fig. 3). L-NPA did not induce hyperalgesia or antinociception by it-self (Fig. 3). Endothelial and inducible selective NOS inhibitors (L-NIO and L-NIL, respectively; 24 lg/paw) were not able to blockthe Ang-(1–7)-induced antinociception (Fig. 4). The soluble gua-nylyl cyclase inhibitor ODQ (25, 50, and 100 lg/paw) blocked theantinociceptive effect of Ang-(1–7) in a dose-dependent mannerand presented no effect when injected alone in normal or hyperal-gesic paws (Fig. 5).
Effect of Ang-(1–7) on the nitrite levels in the homogenized plantarsuperficial rat paw
Fig. 6 shows that the injection of Ang-(1–7) into the rat paw in-duced an increase in the nitrite levels in PGE2-treated and non-treated paws compared with the control groups.
Involvement of the K+ channels in peripheral Ang-(1–7)-inducedantinociception
The KþATP specific blocker glibenclamide (20, 40, and 80 lg/paw)dose-dependently blocked the Ang-(1–7)-induced (4 lg/paw)
Fig. 2. Left panel: Antagonism induced by intraplantar administration of L-NOarg ofthe peripheral antinociception induced by Ang-(1–7) in hyperalgesic paws (PGE2,2 lg). L-NOarg (NOA, lg/paw) was administered 30 min prior to Ang-(1–7) (4 lg/paw). Right panel: The lack of effects of NOA 24 in the control groups, i.e. in theabsence of Ang-(1–7). Each column represents the mean ± S.E.M. (n = 4–5). # and ⁄
indicate a significant difference compared to (PGE2 + veh 1 + veh 1) and (PGE2 + veh1 + Ang-(1–7) 4)-injected controls, respectively (P < 0.05, ANOVA + the Bonferronitest). Veh 1 = Saline, Veh 2 = Ethanol 2% in saline.
Fig. 4. Left panel: Effect of intraplantar administration of L-NIO or L-NIL on theperipheral antinociception induced by Ang-(1–7) in hyperalgesic paws (PGE2, 2 lg).L-NIO or L-NIO (24 lg/paw) was administered 30 min prior to Ang-(1–7) (4 lg/paw). Each column represents the mean ± S.E.M. (n = 4–5). ⁄ Indicate a significantdifference compared to (PGE2 + veh 1 + veh 1) and (PGE2 + veh 1 + Ang-(1–7) -injected controls, respectively (P < 0.05, ANOVA + the Bonferroni test). Veh1 = Saline, Veh 2 = Ethanol 2% in saline.
peripheral antinociception (Fig. 7). Glibenclamide alone did not in-duce hyperalgesia or any overt behavioral effect (Fig. 7).
Discussion
We demonstrated in the present study that Ang-(1–7) elicits aperipheral antinociceptive effect in rat paw PGE2-induced hyperal-gesia, as observed by the mechanical paw pressure test. Further-

Fig. 5. Left panel: Antagonism induced by intraplantar administration of ODQ of theperipheral antinociception induced by Ang-(1–7) in hyperalgesic paws (PGE2, 2 lg).ODQ (lg/paw) was administered 10 min prior to Ang-(1–7) (4 lg/paw). Rightpanel: The lack of effects of ODQ 100 in the control groups, i.e. in the absence ofAng-(1–7). Each column represents the mean ± S.E.M. (n = 4–5). # and ⁄ indicate asignificant difference compared to (PGE2 + veh 1 + veh 1) and (PGE2 + veh 1 + Ang-(1–7) 4)-injected controls, respectively (P < 0.05, ANOVA + the Bonferroni test). Veh1 = 10% DMSO in saline, Veh 2 = saline, Veh 3 = 2% Ethanol in saline.
Fig. 6. Effect of Ang-(1–7) injection on nitrite concentration [NO2] in the homog-enized plantar superficial of the paw. Ang-(1–7) (4 lg/paw) was administered 2 hand 50 min after local administration of PGE2. The plantar superficial of the rat pawwas collected 3 h after local administration of PGE2. Each column represents themean ± SEM (n = 4). ⁄ Significant difference compared with PGE2 + veh 1 or veh2 + veh 1 group (P < 0.05, ANOVA 1 the Bonferroni test). Veh 1 = saline, veh2 = ethanol 2% in saline.
Fig. 7. Left panel: Antagonism by intraplantar administered glibenclamide of Ang-(1–7)-induced peripheral antinociception in hyperalgesic paws. Glibenclamide (Gli:lg/paw) was administered 5 min prior to Ang-(1–7) (4 lg/paw). Right panel: Thelack of effects of GLI 80 in the control groups, i.e. in the absence of Ang-(1–7). Eachcolumn represents the mean ± S.E.M. (n = 4). # and ⁄ indicate a significant differencecompared to PGE2 + veh 1 + veh 2 – and Ang-(1–7) + veh 2 + Ang-(1–7) 20–injectedcontrols, respectively (P < 0.05, ANOVA + the Bonferroni test). Veh 1 = 1% tween insaline, Veh 2 = saline, Veh 3 = 2% ethanol in saline.
14 A. Costa et al. / Nitric Oxide 37 (2014) 11–16
more, we found that this antinociceptive response is mediated bythe Mas receptor, as the antagonist A-779 blocked Ang-(1–7) ac-tion [20]. This method has an advantage over other hyperalgesictests because it minimizes the interference of mediators producedduring the inflammatory process [13]. To exclude central effects,PGE2 was administered to both hind paws, while Ang-(1–7) was in-jected only in the left hind paw. Under these conditions, Ang-(1–7)failed to produce antinociception in the right paw, verifying its lo-cal peripheral action. Hence, a dose of 4 lg/paw was used in sub-sequent experiments to evaluate the peripheral mechanism ofAng-(1–7).
Growing evidence indicates that some Ang-(1–7) actions areassociated with NO release. The central administration ofAng-(1–7) into the rat hypothalamus reduces norepinephrine re-lease through a nitric oxide mechanism [23]. In vitro studies have
shown NO release from catecholaminergic neurons [8]. Moreover,Ang-(1–7) stimulates NO release from endothelial cells [24,25].Further studies revealed that Ang-(1–7), acting on this G protein-coupled receptor, activated endothelial nitric oxide synthase andNO production via Akt-dependent pathways, leading to vasodila-tion in the vascular endothelium [26].
Accordingly, our experiments showed that pretreatment withthe NOS inhibitor L-NOarg abolished the Ang-(1–7)-induced antin-ociceptive response. Then, we used specific inhibitors to elucidatethe possible involvement of each specific isoform of NOS. The nNOSinhibitor L-NPA was able to block the Ang-(1–7)-induced effect, butneither the eNOS inhibitor L-NIO or the iNOS inhibitor L-NILshowed an effect, suggesting that nNOS is an important effectorin the peripheral antinociceptive effect of this peptide. A set ofexperiments revealed that nNOS is the primary isoform responsi-ble for the increased NO levels in catecholaminergic neurons fol-lowing Ang-(1–7) stimulation [8]. The Ang-(1–7)-inducedincrease in NO was mediated by Mas receptor activation. Anotherstudy showed that nNOS, but not eNOS, expression was reducedin ventricular myocytes from Mas�/� mice, suggesting that nNOSsignaling is also associated with Ang-(1–7)/Mas receptor actionin other cell types [27,28].
It has been shown that NOS activation results in an increase innitrite (NO�2 ) and nitrate (NO�3 ) ions, the degradation products ofNO [29]. The evidence that NO is metabolized to both (NO�2 ) and(NO�3 ) derives largely from experiments with intact cells, tissues,and whole animals, rather than pure aqueous systems. In chemicalstudies, it has been shown that pure aqueous solutions of NO gen-erate primarily ðNO�2 Þ [30,31]. As a result, the measurement of ni-trite levels is a good indirect measure of NO levels. In the presentstudy, we verified that the injection of Ang-(1–7) into the ratpaw is able to induce an increase in nitrite levels, and this findingconfirms the hypothesis that Ang-(1–7) activates nNOS at periph-eral sites and induces antinociception through NO release.
NO stimulates soluble guanylyl cyclase (sGC) and increasescytosolic cGMP concentrations, leading to the activation of PKG

A. Costa et al. / Nitric Oxide 37 (2014) 11–16 15
[11]. In fact, when sGC was inhibited by ODQ, Ang-(1–7) failed toinduce its peripheral antinociceptive effect, again supporting thehypothesis that NO participates through a cGMP-dependent mech-anism (present results). In accordance with this hypothesis, severalauthors have shown that the L-arginine/NO/cGMP pathway isresponsible for the effects caused by various analgesic drugs inthe periphery [9,10,32–35].
In the past, it was suggested that the activation of the NO/cGMPpathway could induce some biological effects through the openingof KþATP channels [36,37]. This hypothesis was also confirmed byantinociceptive effect when the antinociception induced by so-dium nitroprusside (an NO donor) and dibutyryl-cGMP (a mem-brane-permeable analog of cGMP) was inhibited by thesulfonylurea glibenclamide [15,16] and enhanced by diazoxide[38]. Functional KþATP channels are present in both large- andsmall-diameter sensory neurons in the dorsal root ganglia [39].The pore-forming Kir6.1 or Kir6.2 subunit determines ATP sensitiv-ity and unitary conductance, while the SUR1 or SUR2 subunit con-fers responsiveness to KþATP channel agonists and sulfonylureassuch as glibenclamide [40].
To investigate the possible participation of KþATP channels inAng-(1–7)-induced peripheral antinociception, we used the spe-cific KþATP channel blocker glibenclamide. Our results show a blockof the Ang-(1–7) peripheral antinociceptive effect by glibenclamidein a dose-dependent manner. In the same model utilized in thisstudy, experiments developed by our group have shown that theperipheral antinociceptive effects of several other drugs such asnoradrenaline [41] and xylazine [42] were antagonized by KþATP
channels. In those experiments, the participation of voltage-depen-dent K+ channels and small- and large-conductance Ca2+-activatedK+ channels in peripheral antinociception was discarded.
Increasing evidence indicates that Ang-(1–7) plays a consider-able role in modulating pain sensitivity. Until now, it had beenhypothesized that the peripheral antinociceptive effect inducedby Ang-(1–7) is dependent on Mas receptor activation and thatopioid peptides may not be involved [20]. In the present study,we observed that Ang-(1–7) induced a peripheral antinociceptiveeffect via l-arginine=NO=cGMP=KþATP pathway activation. Taken to-gether, these results contribute to our understanding of RAS phys-iological function in the sensory system, primarily with regard tonociceptive information. Additionally, they support the creationof a new class of analgesic drugs that are distinct from the NSAIDsand opiates already in use.
Acknowledgment
This research and fellowship were supported by the ConselhoNacional de Pesquisa (CNPq-Brazil).
References
[1] H.O. von Bohlen, D. Albrecht, The CNS renin–angiotensin system, Cell TissueRes. 326 (2) (2006) 599–616.
[2] V.M. Pereira, F.M. Reis, R.A. Santos, G.D. Cassali, S.H. Santos, K. Honorato-Sampaio, A.M. dos Reis, Gonadotropin stimulation increases the expression ofangiotensin-(1–7) and MAS receptor in the rat ovary, Reprod. Sci. 16 (12)(2009) 1165–1174.
[3] M. Paul, A. Poyan Mehr, R. Kreutz, Physiology of local renin–angiotensinsystems, Physiol. Rev. 86 (3) (2006) 747–803.
[4] R.D. Machado, R.A. Santos, S.P. Andrade, Mechanisms of angiotensin-(1–7)-induced inhibition of angiogenesis, Am. J. Physiol. Regul. Integr. Comp. Physiol.280 (4) (2001) R994–R1000.
[5] F. Fyhrquist, O. Saijonmaa, Renin–angiotensin system revisited, J. Intern. Med.264 (3) (2008) 224–236.
[6] R.A. Santos, A.J. Ferreira, A.C. Simões e Silva, Recent advances in theangiotensin-converting enzyme 2-angiotensin-(1-7)-Mas axis, Exp. Physiol.93 (5) (2008) 519–527.
[7] M.C. Zimmerman, Angiotensin II and angiotensin-1-7 redox signaling in thecentral nervous system, Curr. Opin. Pharmacol. 11 (2) (2011) 138–143.
[8] R.F. Yang, J. Yin, Y. Li, M.C. Zimmerman, H.D. Schultz, Angiotensin-(1–7)increases neuronal potassium current via a nitric oxide-dependentmechanism, Am. J. Physiol. Cell Physiol. 300 (1) (2011) C58–C64.
[9] I.D.G. Duarte, B.B. Lorenzetti, S.H. Ferreira, Peripheral analgesia and activationof nitric oxide-cyclic GMP pathway, Eur. J. Pharmacol. 186 (1990) 289–293.
[10] I.D.G. Duarte, B.B. Lorenzetti, I.R. Santos, S.H. Ferreira, Analgesia by directantagonism of noceptor sensitization involves the arginine-nitric oxide-cGMPpathway, Eur. J. Pharmacol. 217 (1992) 225–227.
[11] S. Moncada, R.M.J. Palmer, E.A. Higgs, Nitric oxide: physiology,pathophysiology and pharmacology, Pharmacol. Rev. 43 (1991) 109–142.
[12] G.M. Reis, D. Pacheco, A.C. Perez, A. Klein, I.D.G. Duarte, Opioid receptor andNO/cGMP pathway as a mechanism of peripheral antinociceptive action of thecannabinoid receptor agonist anandamide, Life Sci. 85 (2009) 351–356.
[13] D.P. Alves, I.D. Duarte, Involvement of ATP-sensitive K(+) channels in theperipheral antinociceptive effect induced by dipyrone, Eur. J. Pharmacol. 444(1–2) (2002) 47–52.
[14] D.P. Alves, M. Tatsuo, R. Leite, I.D. Duarte, Diclofenac-induced peripheralantinociception, Life Sci. 74 (20) (2004) 2577–2591.
[15] A.C. Soares, R. Leite, M.A. Tatsuo, I.D. Duarte, Activation of ATP-sensitive K(+)channels: mechanism of peripheral antinociceptive action of the nitric oxidedonor, sodium nitroprusside, Eur. J. Pharmacol. 400 (2000) 67–71.
[16] A.C. Soares, I.D.G. Duarte, Dibutyryl-cyclic GMP induces peripheralantinociception via activation of ATP-sensitive K+ channels in the rat PGE2-induced hyperalgesic paw, Br. J. Pharmacol. 134 (2001) 127–131.
[17] G.M. Reis, M.A. Ramos, A.C. Perez, A. Klein, I.D.G. Duarte, Endogenouscannabinoid receptor agonist anandamide induces peripheralantinociception by activation of ATP-sensitive K+, Life Sci. 88 (2011) 653–657.
[18] T.R.L. Romero, G.S. Galdino, G.C. Silva, L.C. Resende, A.C. Perez, S.F. Cortes, I.D.G.Duarte, Involvement of the L-arginine/nitricoxide/cyclic guanosinemonophosphatepathway in peripheral antinociceptioninduced by n-palmitoyl-ethanolamine in rats, J. Neurosci. Res. 90 (2012) 1474–1479.
[19] T.R.L. Romero, I.D.G. Duarte, N-palmitoyl-ethanolamine (PEA) inducesperipheral antinociceptive effect by ATP-sensitive K+ channels activation, J.Pharmacol. Sci. 118 (2012) 156–160.
[20] A.C. Costa, L.K. Becker, E.R. Moraes, T.R.L. Romero, L. Guzzo, R.A. Santos, I.D.G.Duarte, Angiotensin-(1–7) induces peripheral antinociception through Masreceptor activation in an opioid-Independent pathway, Pharmacology 89(2012) 137–144.
[21] L.D. Randall, J.J. Selitto, A method for measurement of analgesic activity oninflamed tissues, Arch. Int. Pharmacol. 113 (1957) 233–249.
[22] L.C. Green, D.A. Wagner, J. Glogowski, P.L. Skipper, J.S. Wishnok, S.R.Tannenbaum, Analysis of nitrate, nitrite, and [15 N] nitrate in biologicalsamples, Anal. Biochem. 126 (1982) 131–138.
[23] M.M. Gironacci, M. Vatta, M. Rodriguez-Fermepín, B.E. Fernández, C. Peña,Angiotensin-(1–7) reduces norepinephrine release through a nitric oxidemechanism in rat hypothalamus, Hypertension 35 (2000) 1248–1252.
[24] N. Toda, K. Ayajiki, T. Okamura, Interaction of endothelial nitric oxide andangiotensin in the circulation, Pharmacol. Rev. 59 (2007) 54–87.
[25] H. Heitsch, S. Brovkovych, T. Malinski, G. Wiemer, Angiotensin-(1–7)-stimulated nitric oxide and superoxide release from endothelial cells,Hypertension 37 (2001) 72–76.
[26] W.O. Sampaio, R.A. Souza dos Santos, R. Faria-Silva, L.T. da Mata Machado, E.L.Schiffrin, R.M. Touyz, Angiotensin-(1–7) through receptor Mas mediatesendothelial nitric oxide synthase activation via Akt-dependent pathways,Hypertension 49 (2007) 185–192.
[27] A.C. da Costa-Goncalves, R. Leite, R.A. Fraga-Silva, S.V. Pinheiro, A.B. Reis, F.M.Reis, R.M. Touyz, R.C. Webb, N. Alenina, M. Bader, R.A. Santos, Evidence thatthe vasodilator angiotensin-(1–7)-Mas axis plays an important role in erectilefunction, Am. J. Physiol. Heart Circ. Physiol. 293 (2007) H2588–H2596.
[28] E.R. Gomes, A.A. Lara, P.W. Almeida, D. Guimaraes, R.R. Resende, M.J.Campagnole-Santos, M. Bader, R.A. Santos, S. Guatimosim, Angiotensin-(1–7)prevents cardiomyocyte pathological remodeling through a nitric oxide/guanosine 30 ,50-cyclic monophosphate-dependent pathway, Hypertension 55(2010) 153–160.
[29] V.P. Reutov, E.G. Sorokina, NO-synthase and nitrite-reductase components ofnitric oxide cycle, Biochemistry 63 (1998) 874–884.
[30] S.E. Schwartz, Trace Atmospheric Constituents: Properties, Transformation andFates, first ed., John Wiley & Sons, New York, 1983.
[31] D.A. Wink, J.F. Darbyshire, R.W. Nims, J.E. Saavedra, P.C. Ford, Reactions of thebioregulatory agent nitric oxide in oxygenated aqueous media: determinationof the kinetics for oxidation and nitrosation by intermediates generated in theNO/O2 reaction, Chem. Res. Toxicol. 6 (1993) 23–27.
[32] T.R.L. Romero, I.D.G. Duarte, a2 Adrenoceptor agonist xylazine inducesperipheral antinociceptive effect by activation of the L-arginine/nitric oxide/cyclic GMP pathway in rat, Eur. J. Pharmacol. 613 (2009) 64–67.
[33] M. Déciga-Campos, F.J. López-Muñoz, Participation of the L-arginine-nitricoxide-cyclic GMP-ATP-sensitive K+ channel cascade in the antinociceptiveeffect of rofecoxib, Eur. J. Pharmacol. 484 (2004) 193–199.
[34] E.K. Perimal, M.N. Akhtar, A.S. Mohamad, M.H. Khalid, O.H. Ming, S. Khalid,L.M. Tatt, M.N. Kamaldin, Z.A. Zakaria, D.A. Israf, N. Lajis, M.R. Sulaiman,Zerumbone-induced antinociception: involvement of the L-arginine-nitricoxide-cGMP-PKC-K+ ATP channel pathways, Basic Clin. Pharmacol. Toxicol.108 (2011) 155–162.
[35] R. Negrete, A. Hervera, S. Leánez, J.M. Martín-Campos, O. Pol, Theantinociceptive effects of JWH-015 in chronic inflammatory pain are

16 A. Costa et al. / Nitric Oxide 37 (2014) 11–16
produced by nitric oxide-cGMP-PKG-KATP pathway activation mediated byopioids, PLoS One 6 (2011) e26688.
[36] M. Kubo, Y. Nakaya, S. Matsuoka, K. Saito, Y. Kuroda, Atrial natriureticfactor and isosorbide dinitrate modulate the gating of ATP-sensitive K+
channels in cultured vascular smooth muscle cells, Circ. Res. 74 (1994)471–476.
[37] M.E. Murphy, J.E. Brayden, Nitric oxide hyperpolarizes rabbit mesentericarteries via ATP-sensitive K+ channels, J. Physiol. 486 (1995) 47–58.
[38] D.P. Alves, A.C. Soares, J.N. Francischi, M.S.A. Castro, A.C. Perez, I.D.G. Duarte,Additive antinociceptive-effect of combination of diazoxide, an activator ofATP-sensitive K+ channels, and sodium nitroprusside and dibutyryl-cGMP, Eur.J. Pharmacol. 489 (2004) 59–65.
[39] T. Kawano, V. Zoga, J.B. McCallum, H.E. Wu, G. Gemes, M.Y. Liang, S. Abram,W.M. Kwok, Q.H. Hogan, C.D. Sarantopoulos, ATP-Sensitive potassium currentsin rat primary afferent neurons: biophysical, pharmacological properties, andalterations by painful nerve injury, Neuroscience 162 (2) (2009) 431–443.
[40] A.P. Babenko, L. Aguilar-Bryan, J. Bryan, A view of sur-KIR6.X, KATP channels,Annu. Rev. Physiol. 60 (1998) 667–687.
[41] T.R.L. Romero, L. Guzzo, A.C. Perez, I.D.G. Duarte, Noradrenaline activates theNO/cGMP/ATP-sensitive K+ channels pathway to induce peripheralantinociception in rats, Nitric Oxide 1 (2012) 157–161.
[42] T.R. Romero, I.D. Duarte, Involvement of ATP-sensitive K(+) channels in theperipheral antinociceptive effect induced by the alpha(2)-adrenoceptoragonist xylazine, J. Pharmacol. Sci. 111 (4) (2009) 323–327.




















