Anatomic correlates of the face and oral cavity representations in the somatosensory cortical area...
14
Anatomic Correlates of the Face and Oral Cavity Representations in the Somatosensory Cortical Area 3b of Monkeys NEERAJ JAIN, * HUI-XIN QI, KENNETH C. CATANIA, AND JON H. KAAS Department of Psychology, Vanderbilt University, Nashville, Tennessee 37240 ABSTRACT We determined the somatotopy of the face and the oral cavity representation in cortical area 3b of New World owl monkeys and squirrel monkeys. Area 3b is apparent as a densely myelin- ated strip in brain sections cut parallel to the surface of flattened cortex. A narrow myelin-light septum that we have termed the “hand-face septum” separates the hand representation from the more lateral face and mouth representation. The face and oral cavity representation is further divided into a series of myelin-dense ovals. We show that three ovals adjacent to the hand representation correspond to the upper face, upper lip, and chin plus lower lip, whereas three or four more rostral ovals successively represent the contralateral teeth, tongue, and the ipsilateral teeth and tongue. Strips of cortex lateral and medial to the area 3b ovals, possibly corresponding to area 1 and area 3a, respectively, have similar somatotopic sequences. Although previous results suggest the existence of great variability within and across primate species, we conclude that the representations of the face and mouth are highly similar across individuals of the same species, and there are extensive overall similarities across these two species of New World monkeys. J. Comp. Neurol. 429:455– 468, 2001. © 2001 Wiley-Liss, Inc. Indexing terms: cortex; primates; Aotus; Saimiri; owl monkey; squirrel monkey Although area 3b is the most intensively studied so- matosensory cortical area in primates, the organization of the lateral portion of area 3b that represents the face and the oral cavity has been a subject of widely different in- terpretations (Pubols and Pubols, 1971; Dreyer et al., 1975; Merzenich et al., 1978; Cusick et al., 1986, 1989; Krubitzer et al., 1995; Manger et al., 1995, 1996). From these studies, it seems that the representations of these body parts vary across primate species and even between individuals of a species. Such large differences in the somatotopic representations between different individuals and species suggest that the developmental mechanisms for the organization of cortex for the face and oral struc- tures are subjected to highly individual specific regula- tions. Moreover, because the representations of other body parts such as the hand that are very uniform across indi- viduals and species (Merzenich et al., 1987; Pons et al., 1987; Jain et al., 1998), it would suggest that different regulatory mechanisms have evolved for the organization of trigeminal nerve inputs compared with those for the rest of the body parts. However, the laterally located face and mouth regions of area 3b can be extremely difficult to explore, boundaries of area 3b can be difficult to identify in this region, and comparisons across individuals can suffer from problems in identifying comparable parts of area 3b. Thus, reported variability in the organization of lateral area 3b may be more a reflection of technical difficulties and data that are difficult to interpret. To address this possibility, we reinvestigated the somatotopy of lateral area 3b in two species of monkeys and used brain sections from flattened cortex to unequivocally identify the bound- aries and extent of lateral area 3b. Histologically visible boundaries in the cortex, if present, allow clear and unambiguous delineation of areas and their subdivisions in the cortex when used in conjunc- tion with mapping data. For example, the borders of his- tologically apparent area 17 (V1) and area 18 (V2) are rarely disputed, but the boundaries and even the existence Grant sponsor: NIH; Grant number: NS 16446. *Correspondence to: Neeraj Jain, Department of Psychology, 301 Wilson Hall, Vanderbilt University, 111 21st Avenue South, Nashville, TN 37240. E-mail:[email protected] Received 4 August 2000; Revised 27 September 2000; Accepted 29 Sep- tember 2000 THE JOURNAL OF COMPARATIVE NEUROLOGY 429:455– 468 (2001) © 2001 WILEY-LISS, INC.
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of Anatomic correlates of the face and oral cavity representations in the somatosensory cortical area...

Anatomic Correlates of the Face andOral Cavity Representations in the
Somatosensory Cortical Area 3bof Monkeys
NEERAJ JAIN,* HUI-XIN QI, KENNETH C. CATANIA, AND JON H. KAAS
Department of Psychology, Vanderbilt University, Nashville, Tennessee 37240
ABSTRACTWe determined the somatotopy of the face and the oral cavity representation in cortical area
3b of New World owl monkeys and squirrel monkeys. Area 3b is apparent as a densely myelin-ated strip in brain sections cut parallel to the surface of flattened cortex. A narrow myelin-lightseptum that we have termed the “hand-face septum” separates the hand representation from themore lateral face and mouth representation. The face and oral cavity representation is furtherdivided into a series of myelin-dense ovals. We show that three ovals adjacent to the handrepresentation correspond to the upper face, upper lip, and chin plus lower lip, whereas three orfour more rostral ovals successively represent the contralateral teeth, tongue, and the ipsilateralteeth and tongue. Strips of cortex lateral and medial to the area 3b ovals, possibly correspondingto area 1 and area 3a, respectively, have similar somatotopic sequences. Although previousresults suggest the existence of great variability within and across primate species, we concludethat the representations of the face and mouth are highly similar across individuals of the samespecies, and there are extensive overall similarities across these two species of New Worldmonkeys. J. Comp. Neurol. 429:455–468, 2001. © 2001 Wiley-Liss, Inc.
Indexing terms: cortex; primates; Aotus; Saimiri; owl monkey; squirrel monkey
Although area 3b is the most intensively studied so-matosensory cortical area in primates, the organization ofthe lateral portion of area 3b that represents the face andthe oral cavity has been a subject of widely different in-terpretations (Pubols and Pubols, 1971; Dreyer et al.,1975; Merzenich et al., 1978; Cusick et al., 1986, 1989;Krubitzer et al., 1995; Manger et al., 1995, 1996). Fromthese studies, it seems that the representations of thesebody parts vary across primate species and even betweenindividuals of a species. Such large differences in thesomatotopic representations between different individualsand species suggest that the developmental mechanismsfor the organization of cortex for the face and oral struc-tures are subjected to highly individual specific regula-tions. Moreover, because the representations of other bodyparts such as the hand that are very uniform across indi-viduals and species (Merzenich et al., 1987; Pons et al.,1987; Jain et al., 1998), it would suggest that differentregulatory mechanisms have evolved for the organizationof trigeminal nerve inputs compared with those for therest of the body parts. However, the laterally located faceand mouth regions of area 3b can be extremely difficult toexplore, boundaries of area 3b can be difficult to identify in
this region, and comparisons across individuals can sufferfrom problems in identifying comparable parts of area 3b.Thus, reported variability in the organization of lateralarea 3b may be more a reflection of technical difficultiesand data that are difficult to interpret. To address thispossibility, we reinvestigated the somatotopy of lateralarea 3b in two species of monkeys and used brain sectionsfrom flattened cortex to unequivocally identify the bound-aries and extent of lateral area 3b.
Histologically visible boundaries in the cortex, ifpresent, allow clear and unambiguous delineation of areasand their subdivisions in the cortex when used in conjunc-tion with mapping data. For example, the borders of his-tologically apparent area 17 (V1) and area 18 (V2) arerarely disputed, but the boundaries and even the existence
Grant sponsor: NIH; Grant number: NS 16446.*Correspondence to: Neeraj Jain, Department of Psychology, 301 Wilson
Hall, Vanderbilt University, 111 21st Avenue South, Nashville, TN 37240.E-mail:[email protected]
Received 4 August 2000; Revised 27 September 2000; Accepted 29 Sep-tember 2000
THE JOURNAL OF COMPARATIVE NEUROLOGY 429:455–468 (2001)
© 2001 WILEY-LISS, INC.

of other visual areas have been a subject of considerabledebate (see Kaas, 1997). The primary sensory areas areoften easy to visualize in brain sections, because they havehigher levels of activity of metabolic enzymes and greatermyelination. In addition, subdivisions or modules withinan area are often present because discontinuities in theperipheral receptor surface are reflected as discontinuitiesin the central representations. The barrels in the primarysomatosensory area or S1 of rats and other rodents is aparticularly striking and well-known example (e.g., Wool-sey and Van der Loos, 1970; Dawson and Killackey, 1987;Wallace, 1987; Jain et al., 1995). Other examples includerecently discovered discrete anatomically visible represen-tations of the digits and palm in primate area 3b (Jain etal., 1998), rays of the nose of the star nosed mole in areaS1 (Catania and Kaas, 1995), and blobs and stripes invisual areas V1 and V2 of primates (Wong-Riley, 1994).Other discontinuities have been described such as therepresentation of the optic disc in the layers of the lateralgeniculate nucleus of primates (Kaas et al., 1973), andsepta dividing the face, hand, and foot regions in theventroposterior nucleus of the thalamus (Kaas et al., 1984;Jones, 1985) and in the brainstem dorsal column-trigeminal nuclear complex (Belford and Killackey, 1979).Such anatomic isomorphs of the receptor surface in brain-stem and cortical representations aid in unambiguouslydelineating areal and subareal boundaries, understandingcortical organization, neuronal connections and process-ing, and plastic reorganization (e.g., Olavarria et al., 1984;Hoeflinger et al., 1995; Jain et al., 1997, 2000). Here, weshow that the representations of the face and oral struc-tures in area 3b can be consistently visualized as discretemyelin-dense ovals in brain sections of the flattened cortexin two species of New World monkeys. We have related thepattern of these ovals to the electrophysiologically deter-mined representations of these peripheral structures.
MATERIALS AND METHODS
Animals
Adult New World monkeys of either sex were used forthese experiments. We processed a total of 18 hemi-spheres from 9 owl monkeys (Aotus trivirgatus) and 8hemispheres from 4 squirrel monkeys (Saimiri sciureus).The animals were wild captured, and familial relation-ships between the animals are unknown. All the proce-dures involving animals conformed to NIH guidelines andwere approved by the Vanderbilt Animal Care and UseCommittee.
Electrophysiological mapping
Somatosensory cortex in four hemispheres from four owlmonkeys and two hemispheres from two squirrel monkeyswas mapped in the region of the face and oral cavityrepresentation. Procedures of electrophysiological map-ping and reconstruction of the mapping data are detailedelsewhere (Jain et al., 1995, 1998). The animals wereanesthetized either with a mixture of ketamine (15 mg/kg,i.m.) and xylazine (0.4 mg/kg, i.m.), or with urethane (200mg/kg as a 25% solution in 5% propylene glycol) withsupplemental dosages of ketamine as needed. The inves-tigator delimiting the receptive field was blind to the lo-cation of the electrode in the brain. We determined re-sponses to the stimulation of the cutaneous and deep
receptors, and movements of the parts of the face andmouth. Periodontal receptors were stimulated by lighttapping of the teeth. Electrolytic lesions made at selectedsites by passing cathodal current (10mA, 10 seconds) wereused to relate the electrophysiological maps to the corticalmyeloarchitecture.
Histologic procedures
At the end of the mapping procedure the animals weregiven a lethal dose of sodium pentobarbital (100 mg/kg)and perfused transcardially in a sequential manner withphosphate buffered saline (PBS, pH 7.2), buffered 2%paraformaldehyde (in PBS), and buffered 2% paraformal-dehyde with 10% sucrose (in PBS). After perfusion, cortexwas separated from the rest of the brain and flattenedbetween glass plates. The flattened cortex was post-fixedin 2% paraformaldehyde and 10% sucrose for 6–8 hoursand cryoprotected in 30% sucrose. The sections were cutparallel to the pial surface on a freezing sliding microtomeat 40 mm. The sections were stained for myelin by aslightly modified (Jain et al., 1998) Gallyas (1979) proce-dure. Cortex from the remaining hemispheres was flat-tened and stained for myelin as described above, exceptthat cortex from three hemispheres from three owl mon-keys was cut in a parasagittal plane in 40-mm-thick sec-tions. A series of sections was stained for myelin as de-scribed above, and an alternate series was stained forNissl substance to determine the staining pattern throughthe depth of cortex.
The images were acquired by using a Spot 2 digitalcamera (Diagnostic Instruments, Inc.) mounted on a Ni-kon Eclipse E800 microscope. The images were processed(Canvas 7, Deneba, Miami, FL) to adjust the overall con-trast and to create mosaics from multiple tissue sections ifrequired. No alterations were made to any localized re-gions of the images.
RESULTS
We extensively mapped the somatosensory cortex in thelateral part of the parietal and frontal lobes where faceand oral structures are represented in squirrel monkeysand owl monkeys. Our mapping results revealed orderlyprogressions of receptive fields from upper face to ipsilat-eral teeth and tongue for neurons at successively morerostral recording sites. The representations of separateparts of the face and mouth in area 3b corresponded tospecific myelin-rich ovals (Fig. 1) seen in brain sections cutparallel to the surface of flattened cortex (Figs. 2–7).
Myeloarchitectonic features of area 3b
Cortex of New World owl and squirrel monkeys is rela-tively easy to flatten, because it is largely devoid of sulci(Fig. 1). A dimple or a short shallow sulcus corresponds tothe deep central sulcus of Old World monkeys. In thesections of flattened cortex stained for myelin the fullextent of area 3b was clearly revealed. Myelin stainingwas particularly dark in layer 4 and the deepest part oflayer 3. The lower limb, trunk, and forelimb region of area3b extended from the medial wall to near the lateralsulcus and partially overlaid the central dimple (Fig. 1).The border between the hand and face representationscould be clearly visualized as a myelin light septum (Figs.1c, 2a, 3a, 4a). Laterally, a caudorostral series of six orseven myelin-rich ovals represented the face and the oral
456 N. JAIN ET AL.

cavity. In owl monkeys, the caudal part of the face repre-sentation in area 3b extended into the upper bank of thelateral sulcus. The oral cavity representation, which wasrostral to the face representation, extended all the way tothe rostral-most part of the frontal lobe and curved ontothe orbitofrontal cortex up to the orbital sulcus (Fig. 1). Insquirrel monkeys, however, all of the face and oral cavityrepresentation in area 3b was on the lateral surface of thebrain and did not extend to the orbital surface. In owlmonkeys, the linear extent of the trunk and limb repre-sentation (12.7 6 1.4 mm; n 5 4) was approximately equalto the face and mouth representation (13.7 6 1.1 mm). Insquirrel monkeys, the linear extent of the trunk and limbrepresentation (13.5 6 0.9 mm; n 5 3) was more than theface and mouth representation (11.2 6 0.4 mm).
Somatotopy in area 3b and its relationshipto the myelin-rich ovals
The face and oral cavity representations in area 3b andthe adjacent cortex were mapped in four owl monkeys andtwo squirrel monkeys. We related the mapping data to themyeloarchitectonic features by placing small electrolyticlesions at selected recording sites toward the end of themapping experiments. The overall caudal to rostral se-quence of the representation in lateral area 3b was upperface, upper lip, chin and lower lip, teeth, tongue, and teethand tongue again. The representation of each of thesebody parts could be related to a specific myelin-rich ovalvisible in the sections from flattened cortex. Three myelin-rich ovals immediately lateral to the hand representationrepresented the face. We have designated these ovals F1,F2, and F3 from caudal to rostral (Figs. 1–7). Oval F1corresponded to the representation of upper face, F2 to theupper lip, and F3 to the representation of the chin andlower lip. Although the rostral two ovals, F2 and F3, wereeasily visible in all the cases, the caudal-most oval, F1,was harder to visualize in some owl monkeys, because itcurved into the upper bank of the lateral sulcus and didnot always flatten in the same plane as the rest of area 3b.F1 was, however, clearly seen in more favorable prepara-tions (Figs. 3, 4).
Immediately rostral to the face representation, a se-quence of three or four ovals, O1, O2, O3, and O4 repre-sented the oral cavity (Figs. 1–7). The O1 oval, immedi-ately rostral to the lower lip representation, responded tothe tapping of the contralateral teeth (Figs. 6, 8). Rostralto O1, the O2 oval had neurons with receptive fields on thecontralateral and ipsilateral tongue (Figs. 6, 8). The
Fig. 1. The locations and myeloarchitectonic features of area 3b.a,b: The locations of area 3b (shaded) on outlines of the brains of owlmonkeys and squirrel monkeys. The lateral portion of area 3b thatrepresents the face and oral cavity is subdivided into several myelin-rich ovals. The ovals that represent face have been labeled F1, F2 andF3, and the ovals that represent the oral cavity have been labeled O1,O2, O3, and O4. In owl monkeys (a), ovals O3 and O4 appear as asingle elongated oval (O31O4) that extends onto the orbitofrontalcortex (lightly shaded region), and oval F1 folds into the upper bankof the lateral sulcus (LS). In squirrel monkeys (b), all of area 3b is onthe parietal surface. c: Area 3b (dark) of an owl monkey revealed in amyelin-stained brain section cut parallel to the surface of flattenedcortex. The ovals F1-F3 and O1-O4 that represent the face and theoral cavity, respectively, are labeled (c.f. a). Medial is toward the topand rostral is toward the right. Scale bar 5 1 cm in a,b; 1 mm in c.
457FACE AND ORAL CAVITY REPRESENTATION IN MONKEYS
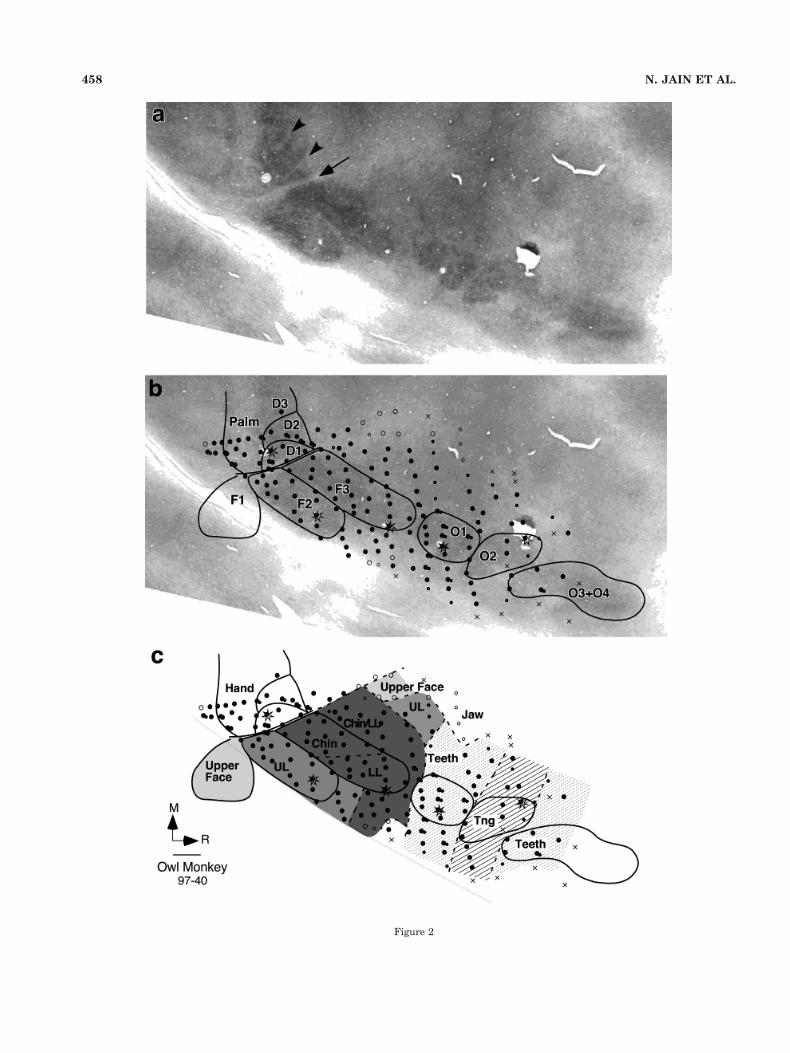
Figure 2
458 N. JAIN ET AL.

rostral-most oval was more oblong in owl monkeys. Werefer to this elongated oval of owl monkeys as the O31O4oval (Fig. 1). We were able to map only the caudal part ofthis oval, because its rostral part curved around onto theorbital cortex. In the caudal part, neurons in O31O4 ovalresponded to the tapping of the ipsilateral teeth. In squir-rel monkeys, however, this region consisted of two distinctovals, O3 and O4 (Fig. 7). The caudal O3 oval responded tothe stimulation of the ipsilateral teeth, as in owl monkeys,whereas the rostral O4 oval responded to stimulation ofthe tongue on the ipsilateral side. It is likely that theelongated oval of owl monkeys is composed of two ovalsthat did not differentiate in our preparations because ofthe extreme curvature of cortex in the region where theborder between O3 and O4 is expected. This possibility issupported by our data from owl monkey 99-12 (Figs. 5, 6,8), which was more extensively mapped in this region. Inthis monkey, we found responses to the stimulation ofipsilateral tongue in the rostral part of the O31O4 oval.
Details of the somatotopy in area 3b
The F1 oval representing upper face was mapped insquirrel monkey 97-133 (Fig. 7c) and owl monkeys 97-53(Fig. 3) and 99-12 (Figs. 5, 6). In this oval, the receptivefields spanned all of the face above the mouth. The regionscloser to the forehead were represented caudally andthose near the nose and upper lip laterorostrally, closerto oval F2.
Recordings were obtained from the F2 oval and the F3oval in squirrel monkey 97-133 (Fig. 7c) and in owl mon-keys 97-40 (Fig. 2), 97-53 (Fig. 3), and 99-12 (Figs. 5, 6). Inthe F2 oval, the lateral upper lip was represented medio-caudally and the medial upper lip was represented later-ally. In the F3 oval, the lower chin was represented closestto the hand-face border. In the rostrolateral direction inF3, the receptive fields progressed toward the hairy edgeof the lower lip. For recordings in the caudal to rostraldirection in F3, the receptive fields progressed from lat-eral to medial chin or lower lip. Most of the receptive fields
on the lips were to the stimulation of the hairy edge. Onlyfew responses were to the stimulation of the glabrous lips,and they were concentrated in the most rostral regions ofthe F2 and F3 ovals.
The O1 oval was mapped in all the monkeys. Most of theresponses were to light taps on the front four contralateralteeth. In owl monkeys, of all sites representing the teeth,including those in the regions medial and lateral to area3b, 18% responded to taps on both the upper and the lowerteeth that oppose each other, 30% were only to the upperteeth and the remaining 52% had responses only to thelower teeth (n 5 122). Although the pattern was hard todiscern, it seemed that there were more responses to up-per teeth in caudomedial O1.
Neurons in the caudal part of the O2 oval responded tothe stimulation of the contralateral tongue, and neuronsin the rostral half of O2 responded to the ipsilateraltongue (Fig. 8). The overall somatotopy suggests that thetip of the tongue is represented caudally and the root ofthe tongue is represented rostrally. In owl monkey 99-12and in squirrel monkeys 97-133 and 00-06, where the O2oval was densely mapped, neurons in a few penetrationsites near the rostral end of O2 responded to the stimula-tion of the hard palate (Figs. 5, 6, 7).
Neurons in the caudal part of the O31O4 oval of owlmonkeys and the O3 oval of squirrel monkeys respondedto stimulation of the ipsilateral teeth (Fig. 8). In addition,neurons in this region responded mainly to the stimula-tion of the teeth in the back part of the mouth. In squirrelmonkeys, where the O4 oval was clearly differentiated inmyelin preparations, we found responses to ipsilateraltongue. Neurons in the few rostral-most penetrations inO31O4 of owl monkey 99-12 also responded to the stim-ulation of the tongue, thus supporting our contention thatthe large O31O4 oval of owl monkeys is an apparentfusion of O3 and O4 of squirrel monkeys.
Nature of responses and the somatotopy ofcortex medial to area 3b
Medial to area 3b, we found another somatotopic pro-gression paralleling that in area 3b, suggesting that area3a lies adjacent to area 3b on the medial side. The soma-totopy of the face responsive cortex was a mirror image ofthat in area 3b. The chin and lower lip responses werefound immediately medial to the chin and lower lip rep-resentations in the F3 oval. More medially, neurons re-sponded to stimulation of the upper lip and upper face(Figs. 2–7). We found responses to tapping of both con-tralateral and ipsilateral teeth in the cortex medial to theO1 oval. More rostrally, there were responses to contralat-eral and ipsilateral tongue, parallel to these representa-tions in O2 of area 3b. There were also responses to thestimulation of the hard palate in squirrel monkeys (Fig. 7)and owl monkey 99-12 (Fig. 6). In owl monkeys 97-40 and98-12 (Figs. 2, 5), we found a few sites with responses tothe ipsilateral teeth that paralleled those in the caudalpart of the O31O4 oval of area 3b. In the squirrel monkey,there were many penetration sites medial to the O3 ovalwith responses to ipsilateral teeth. Much of the cortexmedial to O4 or the rostral part of O31O4 was unrespon-sive to tactile stimulation. Possibly, area 3a does not ex-tend as far rostrally as areas 3b and 1. The receptive fieldsof neurons in area 3a were larger than in area 3b, andthere were many recording sites where neurons respondedonly weakly to tactile stimulation.
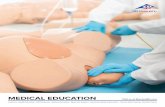
Fig. 2. Topography of the face and oral cavity representation inarea 3b and the surrounding cortex in owl monkey 97-40. a: Myelin-rich ovals in area 3b that represent different body parts. The hand-face septum separating the hand and face representation (arrow) andthe septa separating digit 1, digit 2, and digit 3 (arrowheads) aremarked. Myelin-rich ovals in the region lateral to the hand-face sep-tum represent different face and mouth parts (see below). The brainwas cut parallel to the pial surface after the cortex in the lateralsulcus was unfolded (break in the section). b: Outlines of ovals F1–F3and O1–O4 representing the face and oral cavity marked on a lowcontrast version of the image shown in a. The boundaries were drawnfrom a series of adjacent sections, and oval F1 is not clearly apparentin the section shown. The sites of electrode penetrations are markedby dots, circles, or crosses. Larger circles and dots indicate sites withbetter responses to peripheral stimulation; dots indicate responses tocutaneous stimulation or light taps, and open circles indicate re-sponses to taps. Crosses mark the locations where no neuronal re-sponses were elicited by the stimuli. Stars mark the locations of theelectrolytic lesions used to help align the myeloarchitectonic andelectrophysiological maps (compare a and b). c: Somatotopy in area 3band the cortex medial and lateral to the area 3b ovals. Solid lines showthe boundaries derived from the myeloarchitecture, dashed linesmark boundaries derived from the electrophysiological mapping data.The double hairline on the lateral side marks the location of thelateral sulcus. UL, upper lip; LL, lower lip; Tng, tongue; R, rostral; M,medial. Scale bar 5 1 mm in c (applies to a–c).
459FACE AND ORAL CAVITY REPRESENTATION IN MONKEYS
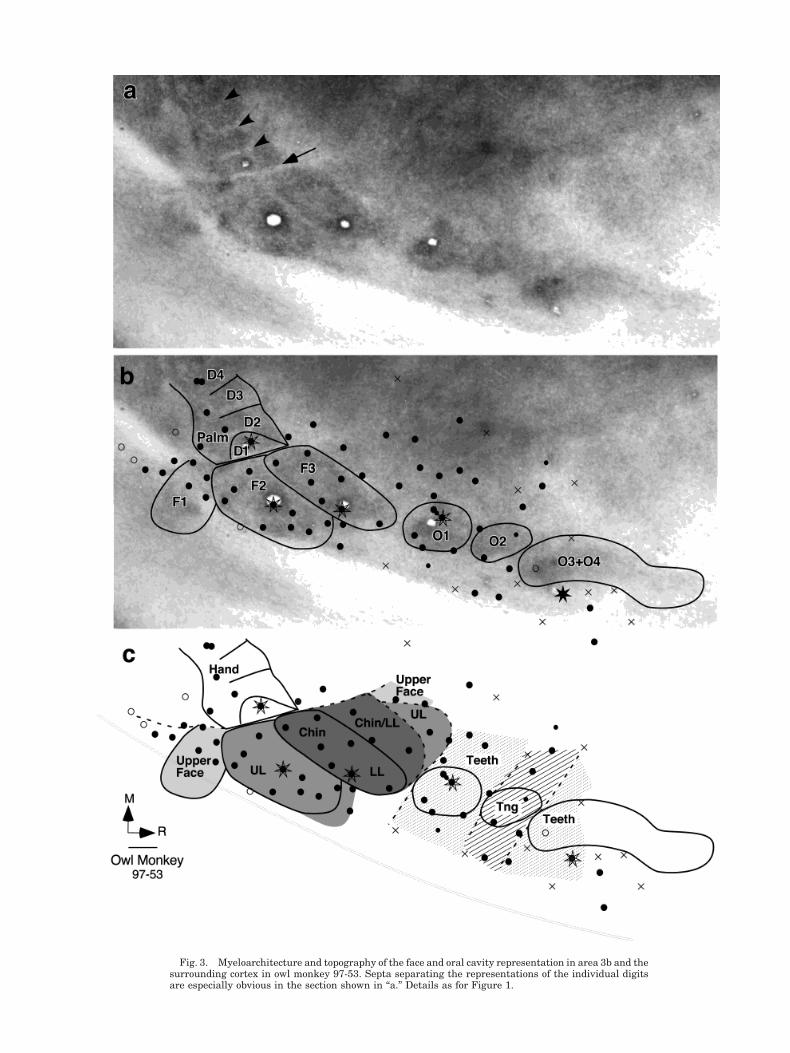
Fig. 3. Myeloarchitecture and topography of the face and oral cavity representation in area 3b and thesurrounding cortex in owl monkey 97-53. Septa separating the representations of the individual digitsare especially obvious in the section shown in “a.” Details as for Figure 1.
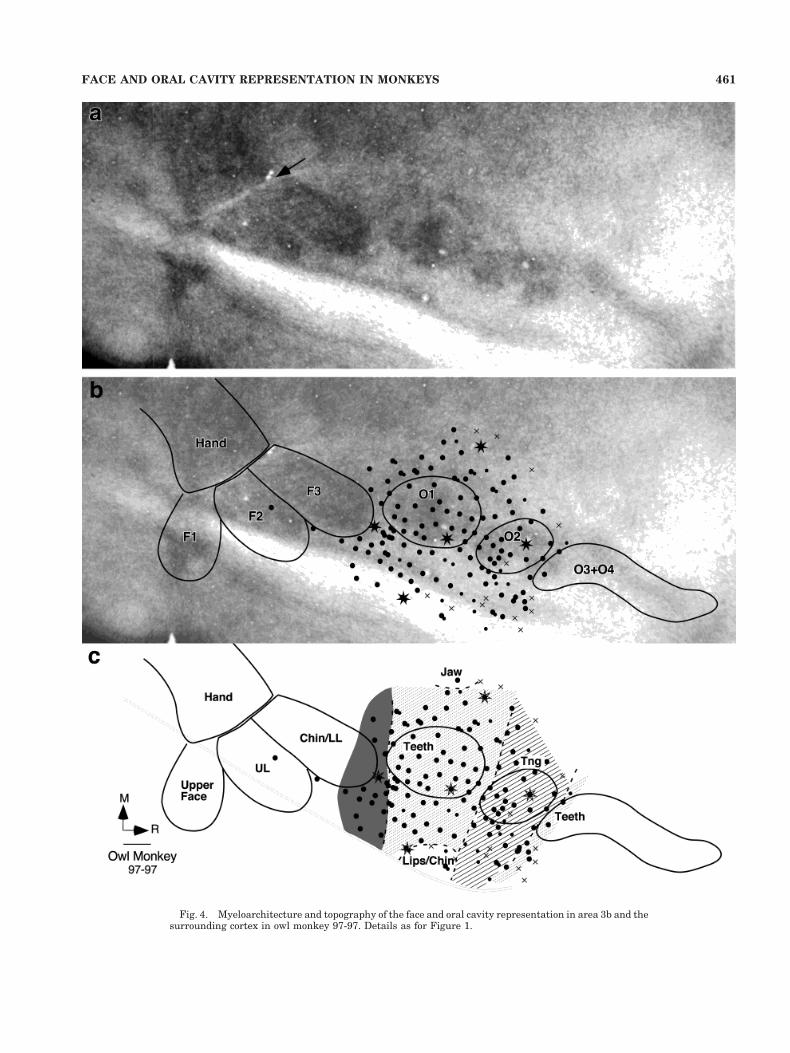
Fig. 4. Myeloarchitecture and topography of the face and oral cavity representation in area 3b and thesurrounding cortex in owl monkey 97-97. Details as for Figure 1.
461FACE AND ORAL CAVITY REPRESENTATION IN MONKEYS

In owl monkeys 97-40 and 97-97 and squirrel monkey97-133 and 00-06, cortex medial to our presumptive area3a was also explored. In this region, we found neuronsresponding to jaw movements. This cortex is likely area 4or primary motor cortex (M1).
Nature of responses and the somatotopy ofcortex lateral to area 3b
Lateral to the ovals of area 3b, neurons responded tocutaneous stimuli, and the somatotopic pattern paralleledthat found in area 3b. In a caudal to rostral sequence ofrecording sites, there were responses to the upper face,upper lip, lower lip and chin, contralateral teeth, con-tralateral tongue, ipsilateral tongue, ipsilateral teeth, andipsilateral tongue. In owl monkey 99-12 and squirrel mon-key 00-06, we also found a few sites that responded to thestimulation of the hard palate. The effective stimuli,thresholds, and sizes of receptive fields were not verydifferent from those for neurons in area 3b. This sequenceof somatotopy and nature of responses suggests that area1 borders area 3b on the lateral side as it does in theregion of the trunk and limb representations.
We did not map the opercular cortex except in thecaudal-most region in owl monkey 99-12 and, therefore,did not map the regions that would correspond to eitherthe second somatosensory area, S2, or the parietal ventralsomatosensory area, PV. However, in owl monkey 97-40and 97-97, we found three penetration sites lateral to theteeth representation where neurons responded to thestimulation of the lips. These recording sites might be themost dorsomedial part of area S2 or PV, both of whichhave face representations toward the medial side (Kru-bitzer and Kaas, 1990; Krubitzer et al., 1995).
DISCUSSION
In this report, we present a new interpretation of howthe face and the oral cavity are represented in area 3b of
monkeys. We define area 3b architectonically by usingsections of flattened cortex cut parallel to the surface. Thepattern of responses at the recording sites could be accu-rately related to a consistently apparent morphologic sub-strate. We propose that lateral area 3b is composed of aseries of 6 or 7 myelin-dense ovals, with each oval corre-sponding to the representation of a specific body part. Thisinterpretation follows our earlier demonstration that eachdigit and the palm of the hand activates a distinct myelin-dense oval in the hand region of area 3b of monkeys (Jainet al., 1998).
Defining area 3b and its myeloarchitectonicfeatures
We define area 3b by its dense myelination. Investiga-tors have long recognized myelin stains to be effective inrevealing primary sensory areas (e.g., Sanides, 1968; alsosee Krubitzer and Kaas, 1990; Jain et al., 1998). Theextent of area 3b in our myelin preparations is similar tothe area 3b described in cytochrome oxidase preparationsin owl monkeys (Tootell et al., 1985). The myelin-denseband of area 3b is subdivided by several narrow, myelin-light septa in a manner that is consistent across membersof the same species and highly similar across the twospecies of New World monkeys. A narrow septum acrossthe width of lateral area 3b separates the mediolateralportion representing the trunk, limbs, and tail from therostrally curving portion representing the face and themouth. This septum has been seen in sections from flat-tened cortex processed for myelin or cytochrome oxidase inowl monkeys (Tootell et al., 1985; Jain et al., 1998), mar-mosets (Krubitzer and Kaas, 1990), and macaque mon-keys (Jain et al., 1998). Because this myelin-light septumseems to be a consistent feature of all the monkey speciesexamined, we name this the “hand-face septum.” We havealso shown that narrow, myelin-light septa in the handregion of area 3b delineate the representation of each digitand even pads of the palm into ovals of dense myelination
Fig. 5. Topography of the face and oral cavity representation in area 3b and the surrounding cortexin owl monkey 99-12. P, hard palate. Other details as for Figure 1.
462 N. JAIN ET AL.
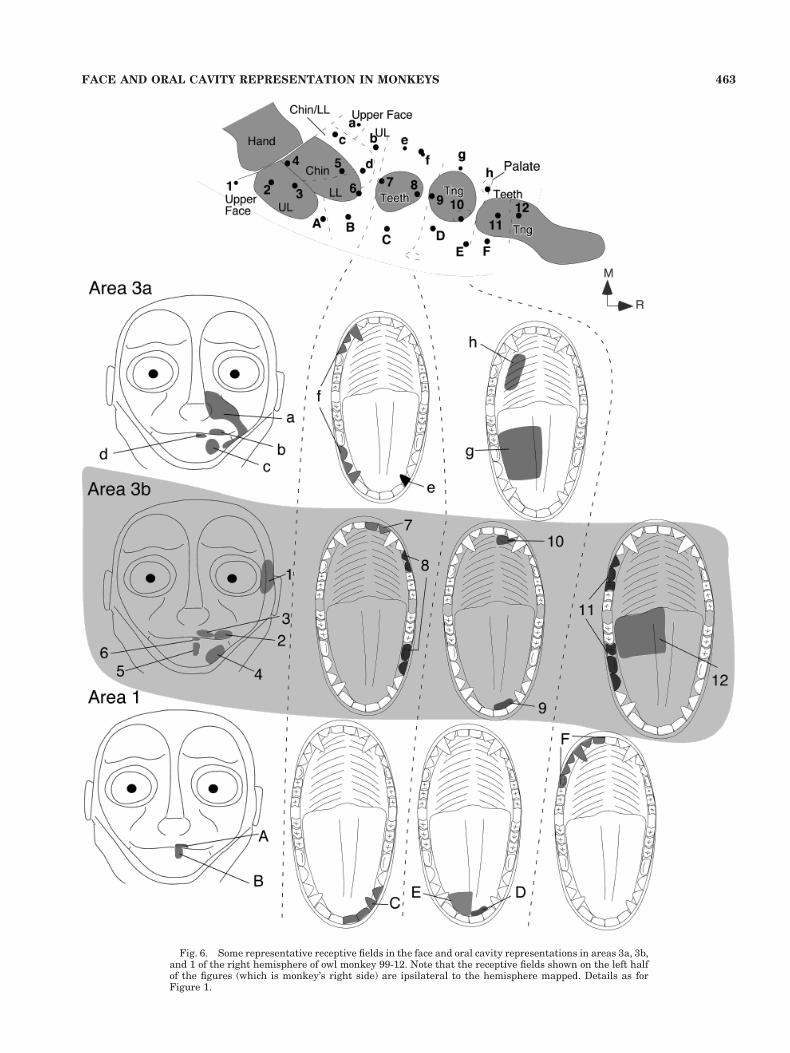
Fig. 6. Some representative receptive fields in the face and oral cavity representations in areas 3a, 3b,and 1 of the right hemisphere of owl monkey 99-12. Note that the receptive fields shown on the left halfof the figures (which is monkey’s right side) are ipsilateral to the hemisphere mapped. Details as forFigure 1.
463FACE AND ORAL CAVITY REPRESENTATION IN MONKEYS
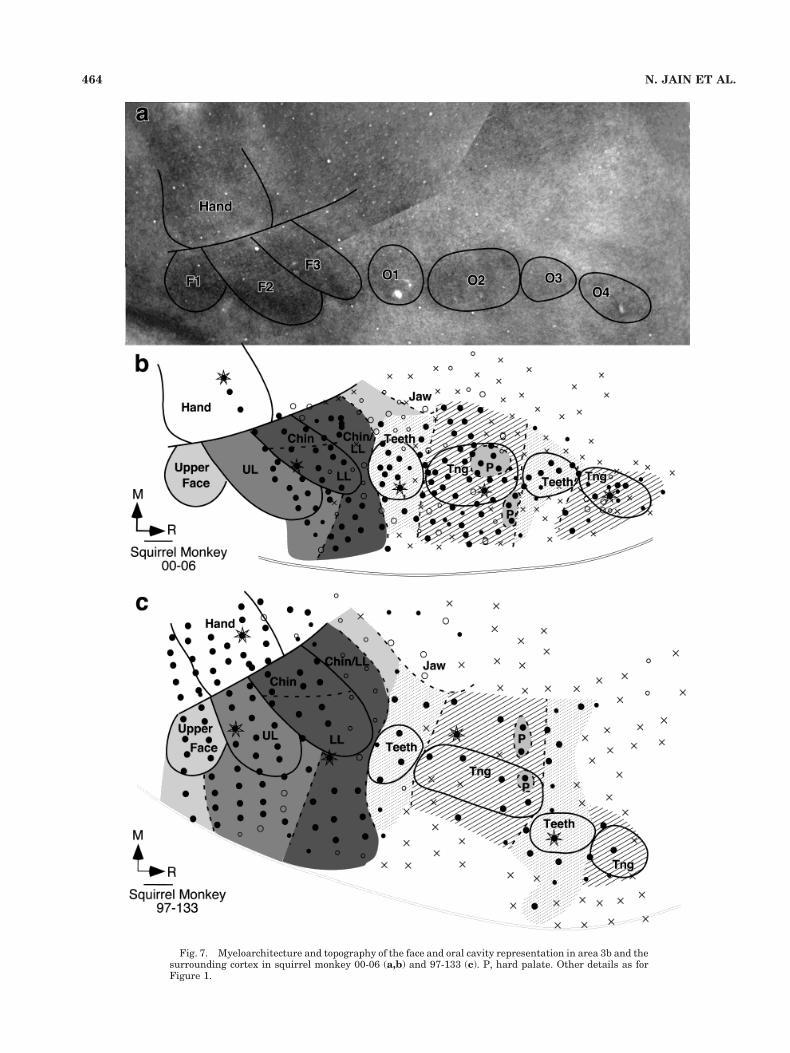
Fig. 7. Myeloarchitecture and topography of the face and oral cavity representation in area 3b and thesurrounding cortex in squirrel monkey 00-06 (a,b) and 97-133 (c). P, hard palate. Other details as forFigure 1.
464 N. JAIN ET AL.

in owl monkeys, squirrel monkeys, and macaque monkeys(Jain et al., 1998; see Figs. 2a, 3a). Subdivisions within therostrally extending part of area 3b representing the faceand mouth have been noted in owl monkeys (Tootell et al.,1985) and squirrel monkeys (Manger et al., 1995), butwithout an interpretation of their consistency or signifi-cance.
It is remarkable how much of area 3b is devoted to theface and oral cavity. In both squirrel and owl monkeys, theF and O ovals constitute nearly half of the length of area3b, with proportionately more of area 3b devoted to faceand mouth representation in owl monkeys than in squirrelmonkeys. In addition, the use of area 3b as a caudal andlateral boundary of frontal cortex makes it obvious thatsquirrel monkeys have much more frontal cortex than owlmonkeys.
Representation of the face and oral cavityin the F and O ovals
We have determined the major representational fea-tures and some of the internal somatotopic order of each of
the myelin dense ovals. In brief, the caudorostral series offace ovals, F1 to F3 (Fig. 1), represent in order, the upperface (F1), the upper lip (F2), and the lower lip and chin(F3). The caudorostral series of ovals, O1 to O4 (Fig. 1),represent the contralateral teeth (O1), the tongue (O2),the ipsilateral teeth (O3), and the tongue again (O4). Inowl monkeys, O3 and O4 appear to be fused, and a singlelarger O31O4 oval represents the ipsilateral teeth andtongue. Other parts of the mouth are less obviously rep-resented. Neurons responsive to the palate were recordedin O2, but the sites responsive to other parts of the innermouth were not noted. In the previous reports detailingthe representation of oral cavity in area 3b or the medialsubdivision of the ventroposterior nucleus (VPM) of thal-amus of New World monkeys, no sites have been illus-trated as responding to the stimulation of other surfaces ofthe oral cavity such as cheeks (Pubols, 1968; Pubols andPubols, 1971; Bombardiei et al., 1975; Kaas et al., 1984;Carlson et al., 1986; Cusick et al., 1986; Manger et al.,1995). We propose that these surfaces are poorly repre-sented in the cortex of owl monkeys and squirrel monkeys,
Fig. 8. Topographical details showing representations of contralateral and ipsilateral teeth andtongue in ovals O1-O4 of area 3b and the surrounding cortex. For conventions, see Figure 1.
465FACE AND ORAL CAVITY REPRESENTATION IN MONKEYS

much as the dorsal hairy surface of the hand in area 3b(see Merzenich et al., 1987; Pons et al., 1987; Jain et al.,1998). However, in the maps of the somatosensory cortexas well as the thalamus of macaque monkeys, large ex-tents of the oral cavity representation respond to the stim-ulation of the cheek pouch (Jones et al., 1986; Rausell andJones, 1991; Manger et al., 1996), likely because ma-caques use their cheek pouches to store food and othermaterials.
In the published maps of the VPM of thalamus or area3b of squirrel monkeys an ipsilateral representation of theoral cavity is either absent or relatively small (Bombardieiet al., 1975; Cusick et al., 1986; Manger et al., 1995). Incontrast, we found that approximately 50% of the totaloral cavity representation in area 3b of both owl monkeysand squirrel monkeys is occupied by ipsilateral represen-tations. This result, which is similar to the extent of theipsilateral representation in VPM and area 3b of macaquemonkeys (Jones et al., 1986; Rausell and Jones, 1991;Manger et al., 1996), likely reflects our extensive mappingof the most rostral regions of the representation.
Our data show that there is little variability in therepresentation of the face and oral cavity between individ-uals of a species, and it is highly similar between the twospecies of New World monkeys. Elements of the presentlyproposed organization of lateral area 3b can be found inprevious mapping studies of this region in owl monkeys(Merzenich et al., 1978; Cusick et al., 1989) and squirrelmonkeys (Benjamin et al., 1968; Cusick et al., 1986; Man-ger et al., 1995), but these proposals vary in detail so thatno consistent pattern emerges. We suggest that large in-terindividual and interspecific variations in the face andmouth representations in area 3b in previous reports werelargely due to difficulties in determining the boundaries ofarea 3b. Architectonic boundaries can be unclear in thisregion if the brain is cut in traditional planes. Moreover,unlike the hand region, a lack of clear progressions andreversals of progressions of receptive fields complicated bybilateral representations may have made it harder to as-sign mapping sites to specific somatosensory areas. Stud-ies of the connections of this portion of area 3b likewisecould be inconclusive if borders were not clearly identified.
It is likely that our conclusions apply more generally toother primates, including macaque monkeys for whichthere is variability in the published descriptions of howlateral area 3b is somatotopically organized (Dreyer et al.,1975; Krubitzer et al., 1995; Manger et al., 1996). Never-theless, these studies in macaque monkeys do demon-strate a general tendency for the upper face, upper lip, andlower lip to be represented in a caudorostral sequence asin the F1, F2, and F3 ovals in New World monkeys.
Significance of modular organization in theprimary somatosensory cortex
Area 3b of primates appears to be the homologue ofprimary somatosensory cortex (S1) in other mammals(Kaas, 1983). In many mammalian species, area S1 hasbeen shown to be subdivided into cell-dense, myelin-rich,and high cytochrome oxidase (CO) activity regions. Thesesubdivisions or modules are usually apparent in the re-gion of somatosensory cortex representing densely inner-vated parts of the skin. They correspond to folds, protru-sions, or discontinuities in the receptor sheet such as those
caused by digits (Welker and Seidenstein, 1959; Jain etal., 1998), whiskers on the face (Woolsey and Van derLoos, 1970; Woolsey et al., 1975), or rays on the nose(Catania and Kaas, 1995) and, thus, represent disjunc-tions in the activity patterns in inputs from the receptorsheets. The modules receive major afferents from the ven-troposterior nucleus of the thalamus, but the separatingnarrow, cell-poor, myelin and CO-light septa do not re-ceive dense inputs from the ventroposterior nucleus (Ola-varria et al., 1984). Septa seem to get less direct inputs,such as callosal inputs (Jones and Hendry, 1980; Olavar-ria et al., 1984; Krubitzer and Kaas, 1990) or inputs fromthe posterior group of thalamic nuclei (Lu and Lin, 1993).It is likely that callosal connections terminate preferen-tially in the myelin-poor regions surrounding the ovals inthe face region of area 3b, as well as in the myelin-poorhand-face septum and the septa between digits in thehand region. Gould et al. (1986) have illustrated the dis-tribution of the terminals from the callosal connections aswell as the regions that project across the hemisphere bymeans of corpus callosum by using a large number ofinjections of horseradish peroxidase tracer in the sensori-motor cortex of owl monkeys. In the region of area 3b, thecallosally connected portions form a pattern that isroughly a negative image of myelin-dark regions of area3b (see Figs. 14, 15 of Gould et al., 1986). In contrast, therostral regions of the oral cavity representation in area 3bwith predominantly ipsilateral representations appear tohave more dense callosal connections (Killackey et al.,1983; Gould et al., 1986). The callosal inputs also charac-terize parts of area 3b with bilateral representations (Kil-lackey et al., 1983; Conti et al., 1986; Manzoni et al.,1989). Other connections of the mouth representation inarea 3b remain to be determined.
The organization of cortex along the medialborder of the face and mouth
representation in area 3b
Area 3a forms most or all of the rostral border of theportion of area 3b that represents the trunk and limbs(Merzenich et al., 1978; Jones and Porter, 1980). Area 3aresponds to the stimulation of muscle spindle receptorsand possibly other deep receptors, as well as tactile inputsrelayed from area 3b (Lucier et al., 1975; Heath et al.,1976; Recanzone et al., 1992). Neurons in area 3a havealso been reported to respond to heating of the skin (Tom-merdahl et al., 1998). Area 3a has been only partiallyexplored, and the full extent and the somatotopic organi-zation of this area are largely unknown. In our recordingsfrom sites medial to the F and O ovals, neurons respondedvigorously to light taps on the face, teeth, and tongue. Wedid not detect a consistent change in responsiveness todeep receptors as is often observed in the transitions from3b to 3a in the hand cortex, but neurons in this regionwere often less responsive to light tactile stimuli and thereceptive fields were larger. In Nissl preparations, thisregion had a less dense, but distinct granular layer andscattered large cells in layer 5. These architectonic fea-tures are consistent with our physiological evidence thatarea 3a continues along this portion of area 3b with aparallel somatotopic organization (Phillips et al., 1971;Jones and Porter, 1980).
466 N. JAIN ET AL.

Organization of cortex along thecaudolateral border of area 3b
Along the caudal border of area 3b, a parallel represen-tation of the body surface has been described in area 1(Merzenich et al., 1978; Kaas et al., 1979; Felleman et al.,1983). As for area 3a, the existence of such a parallelrepresentation of cutaneous receptors in area 1 has beenmost firmly established for the region where trunk andlimbs are represented. In this region, an additional par-allel representation has been described in area 2. Yet,uncertainties remain about the organization of the cortexalong the ventral border of area 3b, and there are severalinterpretations of the cortical organization of this region.(1) Area 1 and 2 form two separate parallel representa-tions that curve along the complete border of area 3b(Merzenich et al., 1978; Kaas et al., 1979; Pons et al.,1985). (2) Area 1 borders area 3b along its entire caudaland ventral border, and areas S2 and PV border the faceportion of area 1, with area 2 terminating near the caudalborder of S2. This interpretation for squirrel monkeys(Cusick et al., 1986), owl monkeys (Cusick et al., 1989),and macaque monkeys (Nelson et al., 1980; Krubitzer etal., 1995) raises the question of where, if anywhere, area 2represents the oral cavity. In certain other reports, theborder between area 1 and 2 was not distinguished pastthe hand-face border and there is no clear allocation of thecortex to either area 1 or 2 in the region of the facerepresentation (Dreyer et al., 1975; Burton and Fabri,1995). (3) Areas S2 and PV directly border area 3b, andareas 1 and 2 terminate along the dorsocaudal border ofS2. This interpretation for marmosets (Krubitzer andKaas, 1990) is consistent with the evidence from nonpri-mate mammals that S2 and PV directly border S1 (Kru-bitzer et al., 1986; Krubitzer, 1995; Beck et al., 1996, alsosee Kaas, 1983). However, this interpretation leaves littlespace for the representation of the oral cavity in eitherarea 1 or 2. Our data from owl monkeys and squirrelmonkeys show a clear progression of receptive fields fromupper face to teeth and tongue lateral and parallel to themyelin-rich ovals of area 3b. This strongly suggests thatarea 1 parallels area 3b along the face and mouth repre-sentation in these two New World monkey species andfavors interpretation 2 above. Further studies on theconnections of the F and O ovals of area 3b and adjoin-ing cortex by using small amounts of neuronal tracerscould provide additional and perhaps compelling evi-dence. Area 1 should have a different pattern of connec-tions with other cortical areas and the thalamus thanPV and S2.
The region of cortex lateral to the tongue representationin area 1 where we found a nonresponsive region in squir-rel monkey may correspond to the larynx area (Jurgens,1982) because larynx was not stimulated in our experi-ments. In owl monkeys, the homologous region is likely tobe in the lateral sulcus, and it may be smaller in sizebecause owl monkeys vocalize less than squirrel monkeys.Based on previous reports it is unlikely that any of thearea 3b ovals includes taste cortex, which appears to lie inthe frontal opercular cortex in squirrel monkeys (Ben-jamin and Burton, 1968) and macaque monkeys (Scott etal., 1986).
ACKNOWLEDGMENT
We thank Dr. Christine Collins for her helpful com-ments on the manuscript.
LITERATURE CITED
Beck PD, Pospichal MW, Kaas JH. 1996. Topography, architecture, andconnections of somatosensory cortex in opossums: evidence for fivesomatosensory areas. J Comp Neurol 366:109–133.
Belford GR, Killackey HP. 1979. The development of vibrissae representa-tion in subcortical trigeminal centers of the neonatal rat. J CompNeurol 188:63–74.
Benjamin RM, Burton H. 1968. Projection of taste nerve afferents toanterior opercular-insular cortex in squirrel monkey (Saimiri sciu-reus). Brain Res 7:221–231.
Benjamin RM, Emmers R, Blomquist AJ. 1968. Projection of tongue nerveafferents to somatic sensory area I in squirrel monkey (Saimiri sciu-reus). Brain Res 7:208–220.
Bombardiei RA Jr, Johnson JI Jr, Campos GB. 1975. Species differences inmechanosensory projections from the mouth to the ventrobasal thala-mus. J Comp Neurol 163:41–63.
Burton H, Fabri M. 1995. Ipsilateral intracortical connections of physio-logically defined cutaneous representations in areas 3b and 1 of ma-caque monkeys: projections in the vicinity of the central sulcus. J CompNeurol 355:508–538.
Carlson M, Huerta MF, Cusick CG, Kaas JH. 1986. Studies on the evolu-tion of multiple somatosensory representations in primates: the orga-nization of anterior parietal cortex in the New World Callitrichid,Saguinus. J Comp Neurol 246:409–426.
Catania KC, Kaas JH. 1995. Organization of the somatosensory cortex ofthe star-nosed mole. J Comp Neurol 351:549–567.
Conti F, Fabri M, Manzoni T. 1986. Bilateral receptive fields and callosalconnectivity of the body midline representation in the first somatosen-sory area of primates. Somatosens Res 3:273–289.
Cusick CG, Wall JT, Kaas JH. 1986. Representations of the face, teeth andoral cavity in areas 3b and 1 of somatosensory cortex in squirrelmonkeys. Brain Res 370:359–364.
Cusick CG, Wall JT, Felleman DJ, Kaas JH. 1989. Somatotopic organiza-tion of the lateral sulcus of owl monkeys: area 3b, S-II, and a ventralsomatosensory area. J Comp Neurol 282:169–190.
Dawson DR, Killackey HP. 1987. The organization and mutability of theforepaw and hind paw representations in the somatosensory cortex ofneonatal rat. J Comp Neurol 256:246–256.
Dreyer DA, Loe PR, Metz CB, Whitsel BL. 1975. Representation of headand face in postcentral gyrus of the macaque. J Neurophysiol 38:714–733.
Felleman DJ, Nelson RJ, Sur M, Kaas JH. 1983. Representations of thebody surface in areas 3b and 1 of postcentral parietal cortex of Cebusmonkeys. Brain Res 268:15–26.
Gallyas F. 1979. Silver staining of myelin by means of physical develop-ment. Neurol Res 1:203–209.
Gould HJD, Cusick CG, Pons TP, Kaas JH. 1986. The relationship ofcorpus callosum connections to electrical stimulation maps of motor,supplementary motor, and the frontal eye fields in owl monkeys.J Comp Neurol 247:297–325.
Heath CJ, Hore J, Phillips CG. 1976. Inputs from low threshold muscle andcutaneous afferents of hand and forearm to areas 3a and 3b of baboon’scerebral cortex. J Physiol (Lond) 257:199–227.
Hoeflinger BF, Bennett-Clarke CA, Chiaia NL, Killackey HP, Rhoades RW.1995. Patterning of local intracortical projections within the vibrissaerepresentation of rat primary somatosensory cortex. J Comp Neurol354:551–563.
Jain N, Florence SL, Kaas JH. 1995. Limits on plasticity in somatosensorycortex of adult rats: hindlimb cortex is not reactivated after dorsalcolumn section. J Neurophysiol 73:1537–1546.
Jain N, Catania KC, Kaas JH. 1997. Deactivation and reactivation ofsomatosensory cortex after dorsal spinal injury. Nature 386:495–498.
Jain N, Catania KC, Kaas JH. 1998. A histologically visible representationof the fingers and palm in primate area 3b and its immutability fol-lowing long term deafferentation. Cereb Cortex 8:227–236.
Jain N, Florence SL, Qi H-X, Kaas JH. 2000. Growth of new brain stemconnections in adult monkeys with massive sensory loss. Proc NatlAcad Sci USA 97:5546–5550.
467FACE AND ORAL CAVITY REPRESENTATION IN MONKEYS

Jones EG. 1985. The thalamus. New York: Plenum Press.Jones EG, Hendry SH. 1980. Distribution of callosal fibers around the hand
representations in monkey somatic sensory cortex. Neurosci Lett 19:167–172.
Jones EG, Porter R. 1980. What is area 3a? Brain Res Brain Res Rev2:1–43.
Jones EG, Schwark HD, Callahan PA. 1986. Extent of the ipsilateralrepresentation in the ventral posterior medial nucleus of the monkeythalamus. Exp Brain Res 63:310–320.
Jurgens U. 1982. Afferents to the cortical larynx area in the monkey. BrainRes 239:377–389.
Kaas JH. 1983. What if anything is S1? The organization of the “firstsomatosensory area” of cortex. Physiol Rev 63:206–231.
Kaas JH. 1997. Theories of visual cortex organization in primates. In:Rockland KE, editors. Cerebral cortex. New York: Plenum Press. p91–125.
Kaas JH, Guillery RW, Allman JM. 1973. Discontinuities in the dorsallateral geniculate nucleus corresponding to the optic disc: a compara-tive study. J Comp Neurol 147:163–180.
Kaas JH, Nelson RJ, Sur M, Dykes RW, Merzenich MM. 1979. Multiplerepresentation of the body within “S-1” of primates. Science 204:521–523.
Kaas JH, Nelson RJ, Sur M, Dykes RW, Merzenich MM. 1984. The soma-totopic organization of the ventroposterior thalamus of the squirrelmonkey, Saimiri sciureus. J Comp Neurol 1984:111–140.
Killackey HP, Gould HJd, Cusick CG, Pons TP, Kaas JH. 1983. Therelation of corpus callosum connections to architectonic fields and bodysurface maps in sensorimotor cortex of new and old world monkeys.J Comp Neurol 219:384–419.
Krubitzer L. 1995. The organization of neocortex in mammals: are speciesdifferences really so different. Trends Neurosci 18:408–417.
Krubitzer LA, Kaas JH. 1990. The organization and connections of somato-sensory cortex in marmosets. J Neurosci 10:952–974.
Krubitzer LA, Sesma MA, Kaas JH. 1986. Microelectrode maps, myeloar-chitecture, and cortical connections of three somatotopically organizedrepresentations of the body surface in the parietal cortex of squirrels.J Comp Neurol 250:403–430.
Krubitzer L, Clarey J, Tweedale R, Elston G, Calford M. 1995. A redefini-tion of somatosensory areas in the lateral sulcus of macaque monkeys.J Neurosci 15:3821–3839.
Lu SM, Lin RC. 1993. Thalamic afferents of the rat barrel cortex: a light-and electron- microscopic study using Phaseolus vulgaris leucoagglu-tinin as an anterograde tracer. Somatosens Mot Res 10:1–16.
Lucier GE, Ruegg DC, Wiesendanger M. 1975. Responses of neurons inmotor cortex and in area 3a to controlled stretches of forelimb musclesin cebus monkeys. J Physiol (Lond) 251:833–853.
Manger PR, Woods TM, Jones EG. 1995. Representation of the face andintraoral structures in area 3b of the squirrel monkey (Saimiri sciu-reus) somatosensory cortex, with special reference to the ipsilateralrepresentation. J Comp Neurol 362:597–607.
Manger PR, Woods TM, Jones EG. 1996. Representation of face and intra-oral structures in area 3b of macaque monkey somatosensory cortex.J Comp Neurol 371:513–521.
Manzoni T, Barbaresi P, Conti F, Fabri M. 1989. The callosal connectionsof the primary somatosensory cortex and the neural bases of midlinefusion. Exp Brain Res 76:251–266.
Merzenich MM, Kaas JH, Sur M, Lin CS. 1978. Double representation ofthe body surface within cytoarchitectonic areas 3b and 1 in “SI” in theowl monkey (Aotus trivirgatus). J Comp Neurol 181:41–73.
Merzenich MM, Nelson RJ, Kaas JH, Stryker MP, Jenkins WM, Zook JM,Cynader MS, Schoppmann A. 1987. Variability in hand surface repre-sentations in areas 3b and 1 in adult owl and squirrel monkeys. J CompNeurol 258:281–296.
Nelson RJ, Sur M, Felleman DJ, Kaas JH. 1980. Representation of thebody surface in postcentral parietal cortex of Macaca fascicularis.J Comp Neurol 192:611–643.
Olavarria J, Van Sluyters RC, Killackey HP. 1984. Evidence for the com-plementary organization of callosal and thalamic connections withinrat somatosensory cortex. Brain Res 291:364–368.
Phillips CG, Powell TPS, Wiesendanger M. 1971. Projection from low-threshold muscle afferents of hand and forearm to area 3a of baboon’scortex. J Physiol (Lond) 217:419–446.
Pons TP, Garraghty PE, Cusick CG, Kaas JH. 1985. The somatotopicorganization of area 2 in macaque monkey. J Comp Neurol 241:445–466.
Pons TP, Wall JT, Garraghty PE, Cusick CG, Kaas JH. 1987. Consistentfeatures of the representation of the hand in area 3b of macaquemonkeys. Somatosen Res 4:309–331.
Pubols LM. 1968. Somatic sensory representation in the thalamic ventro-basal complex of the spider monkey (Ateles). Brain Behav Evol 1:305–323.
Pubols BH Jr, Pubols LM. 1971. Somatotopic organization of spider mon-key somatic sensory cerebral cortex. J Comp Neurol 141:63–75.
Rausell E, Jones EG. 1991. Histochemical and immunocytochemical com-partments of the thalamic VPM nucleus in monkeys and their relation-ship to the representational map. J Neurosci 11:210–225.
Recanzone GH, Merzenich MM, Jenkins WM. 1992. Frequency discrimi-nation training engaging a restricted skin surface results in emergenceof a cutaneous response zone in cortical area 3a. J Neurophysiol 67:1057–1070.
Sanides F. 1968. The architecture of the cortical taste nerve areas insquirrel monkey (Saimiri sciureus) and their relationships to insular,sensorimotor and prefrontal regions. Brain Res 8:97–124.
Scott TR, Yaxley S, Sienkiewicz ZJ, Rolls ET. 1986. Gustatory responses inthe frontal opercular cortex of the alert cynomolgus monkey. J Neuro-physiol 56:876–890.
Tommerdahl M, Delemos KA, Favorov OV, Metz CB, Vierck CJ Jr, WhitselBL. 1998. Response of anterior parietal cortex to different modes ofsame-site skin stimulation. J Neurophysiol 80:3272–3283.
Tootell RBH, Hamilton SL, Silverman MS. 1985. Topography of cyto-chrome oxidase activity in owl monkey cortex. J Neurosci 5:2786–2800.
Wallace MN. 1987. Histochemical demonstration of sensory maps in therat and mouse cerebral cortex. Brain Res 418:178–182.
Welker WI, Seidenstein S. 1959. Somatic sensory representation in thecerebral cortex of the raccoon (Procyon lotor). J Comp Neurol 111:464–501.
Wong-Riley MTT. 1994. Primate visual cortex: dynamic metabolic organi-zation and plasticity revealed by cytochrome oxidase. In: Peters A,Rockland KS, editors. Primary visual cortex in primates. New York:Plenum Press. p 141–200.
Woolsey TA, Van der Loos H. 1970. The structural organization of layer IVin the somatosensory region (SI) of mouse cerebral cortex. Brain Res17:205–242.
Woolsey TA, Welker C, Schwartz RH. 1975. Comparative anatomical stud-ies of the Sm1 face cortex with special reference to the occurrence of“barrels” in layer IV. J Comp Neurol 164:79–94.
468 N. JAIN ET AL.




















