
Three new Microthoracids (Ciliophora, Nassophorea) from Austria and Venezuela

ARTICLE IN PRESS
http://www.elsevier.de/protisPublished online date 30 July 2008
1
Correspondine-mail a.warre
& 2008 Elsevdoi:10.1016/j
159, 519—533, October 2008
Protist, Vol.ORIGINAL PAPER
Analysis of the Internal Transcribed Spacer 2 (ITS2)Region of Scuticociliates and Related Taxa(Ciliophora, Oligohymenophorea) to Infer theirEvolution and Phylogeny
Miao Miaoa, Alan Warrenb,1, Weibo Songa, Shi Wangc, Huimin Shanga, and Zigui Chena
aLaboratory of Protozoology, Ocean University of China, Qingdao 266003, ChinabDepartment of Zoology, Natural History Museum, Cromwell Road, London SW7 5BD, UKcLaboratory of Molecular Genetics and Breeding of Molluscs, Ocean University of China,Qingdao 266003, China
Submitted November 25, 2007; Accepted May 31, 2008Monitoring Editor: Michael Melkonian
The ITS2 (ITS — internal transcribed spacer) region of the rDNA in 11 representative scuticociliatesand two ambiguously related genera was analyzed. In common with other eukaryotes, the putativeITS2 folding pattern consists of a closed loop with four helices supported by minimum free energy andcompensatory base changes (CBCs), although two of these helices are variable and sometimesabsent. Three topologies were obtained on the basis of traditional primary sequence analysis, ‘‘string’’strategy of secondary structure and analysis of the combined data. It was found that the secondarystructure information could help to improve alignment and utilize appropriately phylogeneticstrategies. The proposed phylogenies, though differing between sequence- and structure-basedresults, provide consistent support for high-level clades: the systematically questionable generaDexiotrichides and Cardiostomatella always cluster together in a clade basal to the scuticociliatess.s., whereas Pleuronema branches from other uronematids at a deep level, and is hence a divergenttaxon. Within the well-supported monophyletic philasterids, a sister relationship exists betweenOrchitophrya and Mesanophrys, while Uronema shows a close relationship with the group includingParanophrys and Parauronema. The positions of Metanophrys, Pseudocohnilembus and Ano-phryoides among the philasterids remain poorly resolved. Our findings firmly support the proposedevolutionary scenario inferred previously both from morphological and molecular data.& 2008 Elsevier GmbH. All rights reserved.
Key words: ITS2 secondary structure; molecular phylogeny; scuticociliates.
Introduction
The internal transcribed spacer (ITS) regions,which can excise themselves during the matura-tion of the precursor of ribosomal RNA (rRNA)
g [email protected] (A. Warren).
ier GmbH. All rights reserved..protis.2008.05.002
transcripts, are interspersed among the rRNAgenes (Maroteaux et al. 1985). The ITS regionscan be further subdivided into ITS1, which islocated between small subunit (SSU) and 5.8SrRNA genes, and ITS2, which separates the 5.8Sand large subunit (LSU) rRNA genes. Owing to

ARTICLE IN PRESS
520 M. Miao et al.
their relatively high divergence, ITS2 sequencesare widely used for phylogenetic reconstructionsat both genus and species levels (Alverez andWendel 2003; Park et al. 2003). However, ITS2 hasnot been considered appropriate for molecularphylogenetics at high taxonomic ranks, mostlydue to excessive INDELs (insertions—deletions),saturation, and/or intragenomic variation (Fabryet al. 1999; Vollmer and Palumbi 2004). It hasrecently been reported that ITS2 has a conservedcore structure that comprises four helices, thethird being the longest (Coleman 2007; Schultz etal. 2005; Wolf et al. 2005a). More than 150,000ITS2 sequences have been deposited in GenBank(NCBI), ca. 86,000 of which follow a homologymodeling of structures (via ITS2 database http://its2.bioapps.biozentrum.uni-wuerzburg.de)(Schultz et al. 2006; Selig et al. 2008). The ITS2appears to evolve at a relatively moderate rateand its value has been described for eukaryoteevolution (Coleman 2003; Schlotterer et al. 1994).The secondary structure of the ITS region hasbeen increasingly taken into account in thesequence alignments in order to compare homo-logous characteristics (Gottschling et al. 2005).Furthermore, several strategies based on struc-tural parameters have been developed to improvethe reconstruction of phylogenies (Billoud et al.2000; Subbotin et al. 2007; Wang et al. 2007).These improvements provide a well-supportedbackground for the phylogenetic application ofITS2.
Classifications of the ciliates within the subclassScuticociliatia reflect significant variations regard-ing their morphologies, life styles, and behaviors(Corliss 1979; Foissner 1996; Lynn and Small2002). Small (1967) established this taxon as anindependent lineage of hymenostomes character-ized by their mode of stomatogenesis and thepossession of the scutica or scuticovestige, whichis a group of basal bodies or kinetosomes thattypically arise posterior of, or parallel to, theparoral membrane. It is generally accepted thatthe subclass Scuticociliatia comprises threeorders: Philasterida, Pleuronematida, and Thig-motrichida (Lynn and Small 2002). Li et al. (2006)added a fourth order, Loxocephalida, although thetaxonomic rank of this group remains in question(Yi and Song, personal communication). Althoughmorphological and morphogenetic attributes areroutinely used to identify species and to deducethe relationships among them, the systematicpositions of certain taxa remain ambiguous(Morade and Small 1994; Song and Wilbert2000). In recent years, ribosomal RNA data have
provided support for the recognition of thescuticociliates as a distinct, high ranking groupwithin the class Oligohymenophorea (Li et al.2006; Ragan et al. 1996; Shang et al. 2006).Molecular data derived from random amplifiedpolymorphic DNA-fingerprinting (RAPD), andrestriction fragment length polymorphism (RFLP)have also been used to reconstruct the phyloge-netic relationships both within the Scuticociliatiaand with related taxa (Shang and Song 2002, 2005;Shang et al. 2003; Stoeck et al. 1998). However,comparisons between different studies reveal thatthere is much confusion over phylogeny andevolution of taxa within the Scuticociliatia.
In the present study, we examined the foldingpatterns of the ITS2 segment and the conserva-tion in its putative secondary structure for a rangeof scuticociliates and related species. The inclu-sion of thirteen ITS2 sequences representingeleven genera and six families allowed us toacquire a broad perspective of rRNA spacerevolution and phylogeny for the scuticociliatesand related species. The main aim of this paperwas to demonstrate the value of the ITS2 putativesecondary structure for a group of marine ciliates.To this end, two hypotheses were tested asfollows: (1) there is a conserved core structure inthe ITS2 of scuticociliates as in other eukaryotes;(2) ITS2 genealogy is congruent with SSU rRNAsequence phylogeny, moreover ITS2 putativesecondary structure can improve and enhancephylogenetic inference among scuticociliates andrelated species.
Results
Primary Sequence Comparisons
The ITS2 sequences showed considerable varia-tion among the species studied with substitutions,insertions, and deletions as presented in Table 1.The lengths of the ITS2 sequences of 13 speciesranged from 168 nt (Pleuronema coronatum) to217 nt (Cardiostomatella vermiformis) with a meanof 179 nt. The average G+C content of the ITS2sequence was 39.6%, with the highest G+Ccontent detected in C. vermiformis (47.9%) andthe lowest in Uronema elegans (31.4%).
Putative Secondary Structure Tracts inRelated Scuticociliates
Putative secondary structures of the ITS2 tran-script of the 13 species are shown in Figure 1.

ARTICLE IN PRESS
Table 1. Details of ITS regions from scuticociliates and related species.
Species Genbankaccession no.
Reference or data source(persons who sequenced thegene)
ITS2Length(nt)
ITS2 GCContent (%)
Dexiotrichides pangi AY513758 Present work (Shang) 171 36.26Cardiostomatella vermiformis EU262621 Present work (Miao) 217 47.93Anophryoides haemophila AF107779 Goggin and Murphy (2000) 174 35.63Orchitophrya stellarum AF107773 Goggin and Murphy (2000) 177 37.29Pseudocohnilembus hargisi AY513753 Present work (Shang) 178 39.89Pseudocohnilembuspersalinus
EU262622 Present work (Miao) 169 43.20
Metanophrys similis AY513757 Present work (Shang) 174 36.78Mesanophrys chesapeakensis AF107778 Goggin and Murphy (2000) 174 39.66Mesanophrys carcini AY513756 Present work (Shang) 173 39.88Paranophrys magna AY513755 Present work (Shang) 182 36.81Parauronema longum AY513759 Present work (Shang) 175 36.00Uronema elegans AY513760 Present work (Shang) 172 31.40Pleuronema coronatum AY513754 Present work (Shang) 168 47.02
The species sequenced in the senior author’s laboratory (OUC) are highlighted in boldface.
521ITS2 Structure of Scuticociliates and Related Species
In spite of distinct sequence variation, thesetaxa shared a very similar pattern of secondarystructure with homologous sequence segmentshaving homologous locations. Therefore a gen-erally putative secondary structure model couldbe developed, the main features of which are: (1) aclosed loop with four helices (helix I, II, III and IV);(2) helix II is highly conserved containing a motif50-GYGRUUGA versus UCYCYCRY-30 at its base;(3) helix III is the longest of the four and bearsthree bulge loops; (4) helix I shows variationamong the different taxa; (5) helix IV does notalways occur (Fig. 2).
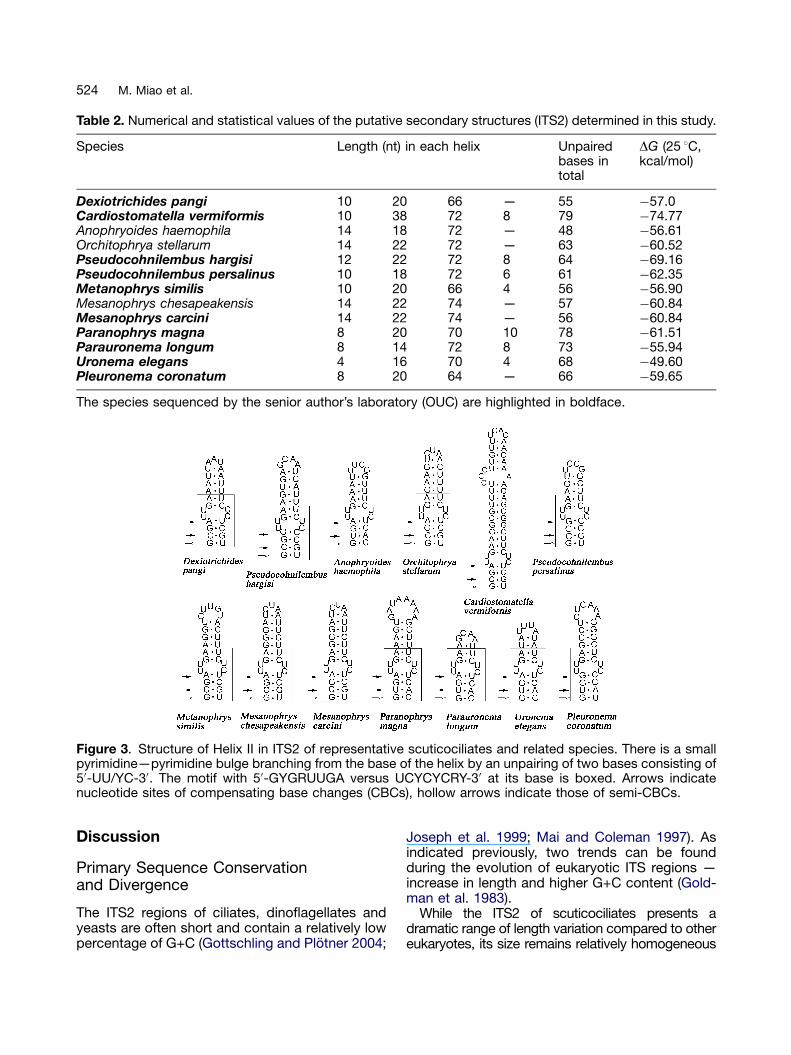
The estimated thermodynamic energy of putativesecondary structures ranges from �49.60 kcal/mol(Uronema elegans) to �74.77 kcal/mol (Cardiosto-matella vermiformis) (Table 2). The four domainsshow distinct size classes. Usually Helix I, the mostvariable region of ITS2, is 4—7 bp long but shorterin Uronema elegans (2 bp). Helix II varies from 7 bp(Parauronema longum) to 11 bp (Orchitophrya stel-larum, Pseudocohnilembus hargisi, Mesanophryschesapeakensis and M. carcini), although it isextremely elongated in C. vermiformis (19 bp). HelixIII ranges from 32 to 37 bp. Finally, Helix IV, whichis usually absent, is generally 3—5 bp long butsometimes is only 2 bp (Uronema elegans andMetanophrys similis).
The sequence domains are engaged in basepairing and are exposed as bulges or loops. Thereis a pyrimidine—pyrimidine bulge branching fromhelix II by a short base pairing consisting of 50-UU/YC-30 (50-UU/CC-30 in Dexiotrichides pangi,
50-UU/UC-30 in others) (Fig. 3). In Helix III we founda highly conserved region in all species studiedcomprising bulge B1 (highlighted gray in Fig. 4)and two small bulges (B2 and B3), positioned nextto the region presenting the motif 50-UGAAUC-GU(A)UCAGUG versus CACUGG(A)GAUUCA-30
(except Cardiostomatella vermiformis and Pleur-onema coronatum). This region contains 14 bpthat are conserved in 77% of species studied.Frequencies of bases at each position and mutualinformation on base-pair regions in Helices II andIII among ‘‘typical’’ scuticociliates (except Cardi-ostomatella vermiformis and Dexiotrichides pangi)are shown in RNA structure logos (Fig. 5)
Among the species included in the presentstudy there are several compensatory basechanges (CBCs) or hemi-CBCs within the helices:the three pairings of the bases of Helix II (Fig. 3)and the relatively conserved region of Helix III inFigure 4, marked with arrows where CBCs andhemi-CBCs occur. In fact, most of these changesconcern only one of the two paired positions.Although less stable than the Watson—Crickcomplementarities, the presence of GU apposi-tions retain the RNA helical structure.
Finally, for each taxon at least one of theMFOLD-generated structures included three orfour of the substructural features and subsequentmanual adjustments easily yielded the remainder.Therefore, sequences were resubmitted to MFOLDusing constraints for plausible base-pairing rela-tions that would generate all consensus substruc-tural features.
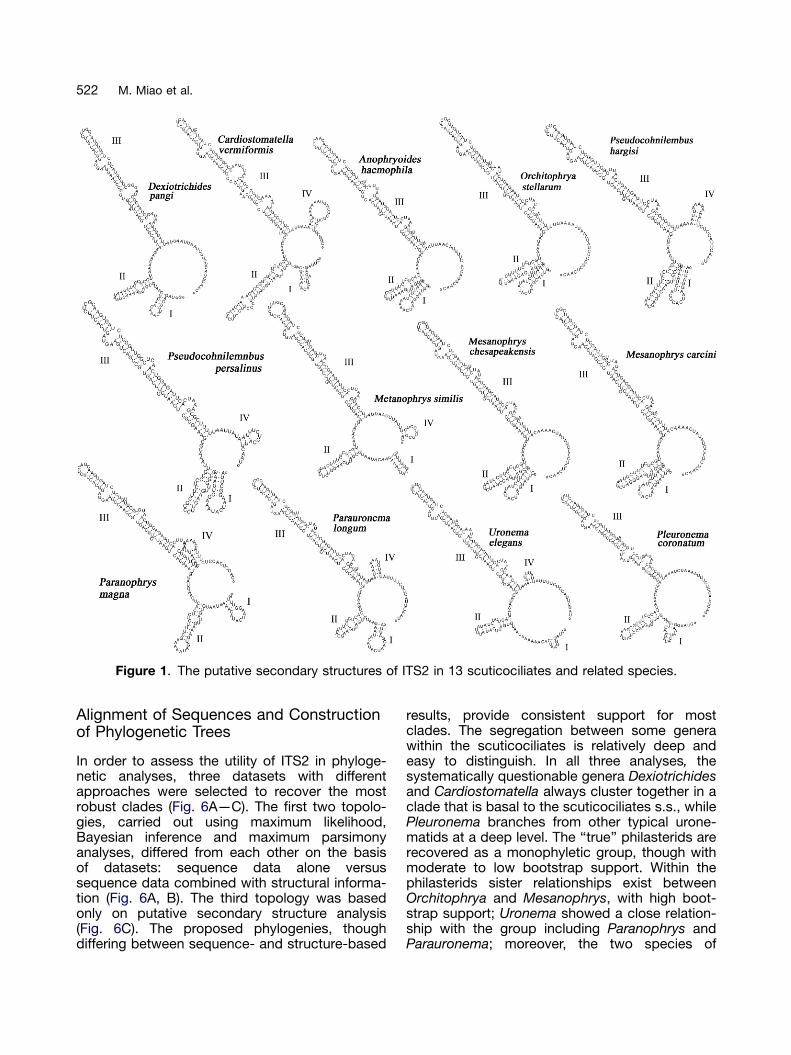
ARTICLE IN PRESS
Figure 1. The putative secondary structures of ITS2 in 13 scuticociliates and related species.
522 M. Miao et al.
Alignment of Sequences and Constructionof Phylogenetic Trees
In order to assess the utility of ITS2 in phyloge-netic analyses, three datasets with differentapproaches were selected to recover the mostrobust clades (Fig. 6A—C). The first two topolo-gies, carried out using maximum likelihood,Bayesian inference and maximum parsimonyanalyses, differed from each other on the basisof datasets: sequence data alone versussequence data combined with structural informa-tion (Fig. 6A, B). The third topology was basedonly on putative secondary structure analysis(Fig. 6C). The proposed phylogenies, thoughdiffering between sequence- and structure-based
results, provide consistent support for mostclades. The segregation between some generawithin the scuticociliates is relatively deep andeasy to distinguish. In all three analyses, thesystematically questionable genera Dexiotrichidesand Cardiostomatella always cluster together in aclade that is basal to the scuticociliates s.s., whilePleuronema branches from other typical urone-matids at a deep level. The ‘‘true’’ philasterids arerecovered as a monophyletic group, though withmoderate to low bootstrap support. Within thephilasterids sister relationships exist betweenOrchitophrya and Mesanophrys, with high boot-strap support; Uronema showed a close relation-ship with the group including Paranophrys andParauronema; moreover, the two species of

ARTICLE IN PRESS
Figure 2. The putative secondary structure modelof the ITS2 transcript in scuticociliates and relatedspecies, supported by CBCs and hemi-CBCs thatpreserve the helix pairing. The four domains, eachwith a stem—loop, are labeled I—IV, Note thecharacteristic pyrimidine—pyrimidine bulge in HelixII. In Helix III, there are three small bulges — B1, B2and B3. Helix IV occurs in some species but not alland is indicated by the dashed line. Helices II and IIIand part of the closed loop are in bold because ofthe relatively well-conserved nucleotide positions.
523ITS2 Structure of Scuticociliates and Related Species
Pseudocohnilembus and the two Mesanophrysspp., grouped together respectively.
Alignment of the ITS2 primary sequences wasconsiderably improved based on adjustment ofthe secondary structure (Fig. 6A, B). Conservedstems identified in the secondary structuraldomain provided consistent bases for correctingthe alignment of variable loop regions for thephylogenetic analyses (data not shown). The
specific instances where the topologies disagreedcould be traced to differences in bootstrapsupport (Fig. 6A, B). The genera Orchitophryaand Mesanophrys occupied a relatively basalposition within the philasterids in the trees inferredfrom ITS2 data combined with primary sequencesand putative secondary structure (Fig. 6B), ratherthan clustering with Anophryoides as in the treeinferred from ITS2 primary sequences alone (Fig. 6A).Furthermore, in the ITS2 primary sequence ana-lysis without adjustment, the well-characterizedpeniculine genus Paramecium clustered withPleuronema rather than Tetrahymena (Fig. 6A).
The topology based only on the predictedsecondary structure of the ITS2 region resolvedmost relationships among the species. The ana-lyses based on secondary structural characters,was performed using the ‘‘string’’ strategy(Fig. 6C). In this strategy, the whole secondarystructure is presented as a string in bracketnotation bonded with each other, thus it isunnecessary to deconstruct the putative second-ary structure into homologous substructural com-ponents. The neighbor-joining tree based on ITS2secondary structure alone showed that in theorder Philasterida, Metanophrys clustered in agroup with Uronema, Paranophrys and Parauro-nema whereas this affiliation is somewhat differentfrom those based on other analyses (Fig. 6A—C).
The maximum-likelihood phylogram for the SSUrRNA gene sequence data is shown in Figure 6D.The phylogeny is typical of previous SSU rRNAanalyses of the Oligohymenophorea, with highbootstrap support for all the established groups.Trees based on Bayesian inference and maximumparsimony analyses have similar topologies with thescuticociliates segregating into two monophyleticassemblages (93%ML, 1.00BI, 79%MP): a periphe-ral clade comprising Cardiostomatella and Dexio-trichides with moderate bootstrap support (55%ML,0.94BI, 66%MP), and a second clade, representingthe typical scuticociliates, with high bootstrap sup-port (97%ML, 1.00BI, 61%MP). The latter clade isfurther subdivided into two lineages: (1) the orderPleuronematida including Pleuronema, Cyclidiumand Schizocalyptra as a well-supported basal line-age (99%ML, 1.00BI, 98%MP); (2) the order Phila-sterida which is considered to be a monophyleticassemblage (96%ML, 1.00BI, 95%MP). Comparedwith the trees based on ITS2 data, Pseudocohni-lembus clusters with Metanophrys (77%ML, 0.98BI,55%MP); Parauronema is sister to Uronema andParanophrys with strong support (67%ML, 0.72BI,95%MP); Anophryoides and Mesanophrys clusterwith each other (98%ML, 1.00BI, 88%MP).
ARTICLE IN PRESS
Table 2. Numerical and statistical values of the putative secondary structures (ITS2) determined in this study.
Species Length (nt) in each helix Unpairedbases intotal
DG (25 1C,kcal/mol)
Dexiotrichides pangi 10 20 66 — 55 �57.0Cardiostomatella vermiformis 10 38 72 8 79 �74.77Anophryoides haemophila 14 18 72 — 48 �56.61Orchitophrya stellarum 14 22 72 — 63 �60.52Pseudocohnilembus hargisi 12 22 72 8 64 �69.16Pseudocohnilembus persalinus 10 18 72 6 61 �62.35Metanophrys similis 10 20 66 4 56 �56.90Mesanophrys chesapeakensis 14 22 74 — 57 �60.84Mesanophrys carcini 14 22 74 — 56 �60.84Paranophrys magna 8 20 70 10 78 �61.51Parauronema longum 8 14 72 8 73 �55.94Uronema elegans 4 16 70 4 68 �49.60Pleuronema coronatum 8 20 64 — 66 �59.65
The species sequenced by the senior author’s laboratory (OUC) are highlighted in boldface.
Figure 3. Structure of Helix II in ITS2 of representative scuticociliates and related species. There is a smallpyrimidine—pyrimidine bulge branching from the base of the helix by an unpairing of two bases consisting of50-UU/YC-30. The motif with 50-GYGRUUGA versus UCYCYCRY-30 at its base is boxed. Arrows indicatenucleotide sites of compensating base changes (CBCs), hollow arrows indicate those of semi-CBCs.
524 M. Miao et al.
Discussion
Primary Sequence Conservationand Divergence
The ITS2 regions of ciliates, dinoflagellates andyeasts are often short and contain a relatively lowpercentage of G+C (Gottschling and Plotner 2004;
Joseph et al. 1999; Mai and Coleman 1997). Asindicated previously, two trends can be foundduring the evolution of eukaryotic ITS regions —increase in length and higher G+C content (Gold-man et al. 1983).
While the ITS2 of scuticociliates presents adramatic range of length variation compared to othereukaryotes, its size remains relatively homogeneous

ARTICLE IN PRESS
Figure 4. Structure of Helix III in ITS2 of representative scuticociliates and related species. A highlyconserved region in all species studied is indicated by the shaded areas while another conserved region ineleven species (except Cardiostomatella vermiformis and Dexiotrichides pangi) is indicated by the boxes.Arrows locate nucleotide sites of compensating base changes (CBCs), hollow arrows indicate those ofsemi-CBCs.
525ITS2 Structure of Scuticociliates and Related Species
with major eukaryotic groups. The insertions anddeletions that account for a large proportion ofvariability in the ITS2 in the scuticociliates andrelated species do not seem to impede the formationof conserved structural elements. The ITS2 hasapparently evolved mostly by length variation of thenucleotide sequence, which causes changes inthe stems of the secondary structures. Perhaps thesmall size of these spacers (as compared with thosein vertebrates, for example) obviates the need forG+C-rich DNA, or the high A+Tcontent might favor astructure analogous to that favored by high G+Ccontent.
Sequence Structural Evolution of ITS2 inScuticociliatia and Implications for itsFunction
In the present investigation it was found that the13 species shared the same ITS2 putativesecondary structure model, viz. a four-fingeredhand, which is similar to that in algae, plants,mice, insects, protozoa and vertebrates (Coleman2003, 2005; Hoef-Emden 2007; Joseph et al.1999; Mai and Coleman 1997; Michot et al. 1999;Oliverio et al. 2002). Among the four helicesnucleotide sequences in scuticociliates appear

ARTICLE IN PRESS
Figure 5. ITS2 structure logo of Helix II (A) and Helix III (B) in ITS2 of typical scuticociliates (exceptCardiostomatella vermiformis and Dexiotrichides pangi). The height of a base in each column is proportionalto its frequency in multiple sequence alignment. The relative entropy method was used where the frequencyof bases in each column is compared to the background frequency of each base. A prior nucleotidedistribution is set to A:C:G:U ¼ 1:1:1:1. Inversed sequence characters indicate a less-than-backgroundfrequency. Mutual information in pairs of columns is indicated by the letter M.
526 M. Miao et al.
to have evolved most rapidly in Helix IV, and nextmost rapidly in Helix I, as in plants and green algae(Mai and Coleman 1997). Helix II is more stableand characteristically has a small bulge, similarto that found in the neogastropods (Oliverio et al.2002). Helix II in Cardiostomatella vermiformis isconspicuously longer than ‘‘typical’’ scuticocili-ates, suggesting that Cardiostomatella is anisolated genus close to the scuticociliates. HelixIII usually contains several bulges and conserva-tive regions.
There are at least two strands of evidence thatsupport the accuracy of the models of ITS2structure suggested here. Firstly, CBCs or hemi-CBCs have been found as defined previously(Gutell et al. 1994). Many of the changes in theprimary sequence are silent in terms of RNAstructure, which suggests that at least some partsof the predicted structures have functional impor-tance because they are conserved. Heredity isguaranteed by faithful DNA replication, whereasevolution depends upon errors accompanyingDNA replication. This suggests mechanisms ofindividual nucleotide replacements, deletions, andrecombination events. And secondly, the identifi-cation of homologous alignment is somewhatambiguous based on the primary sequences,which supports the existence of the helices.
Our analysis also supports the assertion that theoverall size of ITS2 is not critical for correcttranscription and, as a result, we agree that theremay be a ‘‘minimum-nucleotide formula’’ for ITS2,i.e. a minimum number of necessary nucleotides
(Lott et al. 1998). According to our present work, itis clear that the conservation of the structuraldomains (i.e. Helices II and III) found in allscuticociliates investigated to date is constrainedand probably plays a significant functional role inthe folding of the secondary structure of ITS2during rRNA primary transcript processing.Helices I and IV, which are close to the ITS2 endsand are highly variable in length and form, couldbe a part of their stop signals. However it isunknown whether other factors, such as theclosed-loop-like palm, are involved. Furthermoreit is also unknown how genetic diversity canbe generated and whether there are additionalhelices in other scuticociliates.
Two alternative models have recently beenreported for Saccharomyces cerevisiae ITS2: the‘‘hairpin model’’ and the ‘‘ring model’’ (Cote et al.2002). A functional genetic assay was carried outand both of the models were identified as beingimportant in efficient processing. Notably, thescuticociliate sequences examined thus far canadopt the ring structure but appear unable to forma structure analogous to the hairpin model. It is,however, difficult to draw any conclusions basedon these findings because taxon sampling is stillnot sufficiently representative among the scutico-ciliates. Although in Figure 1 Helix I is shownadjacent to the 50 end, it should be noted that thisfeature is variable and differs from species tospecies. Furthermore, Helix IV occurs near the30 end in some species (e.g. Cardiostomatellavermiformis, Metanophrys similis, Parauronema

ARTICLE IN PRESS
Figure 6. Phylogenetic relationships inferred from different gene sequences in thirteen species. A. Inferredfrom ITS2 primary sequences only. Numbers at the nodes represent bootstrap values (in %): 1stNo. ¼ bootstrap values derived from ML method out of 100 replications, 2nd No. ¼ Bayesian credibilityvalue using the MrBayes program, 3rd No. ¼ bootstrap values derived from MP method out of 1000replications. B. Inferred from ITS2 primary sequences and putative secondary structure. Other symbols havethe same meaning as in A. C. Inferred from ITS2 putative secondary structures. Tetrahymena americanis andParamecium tetraurelia were selected as the outgroup taxa. The thick branches denote positions that arecongruent in topologies A, B, C. D. Phylogenetic relationships inferred from SSU rRNA gene data among thescuticociliates and related species. Numbers at the nodes represent bootstrap values (in %): 1stNo. ¼ bootstrap values derived from ML method out of 100 replications, 2nd No. ¼ Bayesian credibilityvalue using the MrBayes program, 3rd No. ¼ bootstrap values derived from MP method out of 1000replications. A karyorelictid ciliate, Loxodes striatus (U24248), was selected as the outgroup species. Speciesused in this study are shown in bold. *Support values o50%/0.50 and disagreement between a method andthe reference ML tree at a given node.
527ITS2 Structure of Scuticociliates and Related Species
longum, Paranophrys magna, Pseudocohnilembushargisi, Pseudocohnilembus persalinus and Uro-nema elegans) but may be short and divergent, or
even absent, in others. In addition, a complexnetwork of interactions might take place toassemble the pre-rRNA structural features directly

ARTICLE IN PRESS
528 M. Miao et al.
involved in processing steps into a relativelycompact structure. For instance, an interactionbetween the 5.8S rRNA 30-end and LSU 50-endwhich is an essential requirement, is also commonin yeast and vertebrates (Joseph et al. 1999).Moreover, putative secondary structure modelsare not necessarily the same as the ones that formin vivo, and they are three-dimensional forms ofITS2, not two-dimensional forms. Therefore, themodel is constrained and may not accuratelyreflect the situation in life. Nevertheless, evidenceof co-variations in the ITS2 sequences wasrecovered here, which supported our structuralestimates.
Phylogenetic Comparisons withinScuticociliatia and Related Species
We can use the available data and conservedstructural elements to identify homology in ITS2evolution, but what is the phylogenetic utility of thismolecule among scuticociliates and related spe-cies? Clearly, the variable ITS2 regions are morelikely to accumulate mutations that could poten-tially record the divergence of the major (e.g. familylevel) lineages, meanwhile they are also more likelyto accrue homoplastic changes in our studies.
It is well known that alignment plays animportant role in phylogenetic research in thesense that alignment ambiguities may lead tospuriously scored synapomorphies as well aserroneously inferred indels. Since the secondarystructure of ITS2 provides recognition and dock-ing signals during the maturation of the precursorof ribosomal RNA, the structure is often moreconservative than the primary sequence (Goodet al. 1997). As a result, one cannot just usestandard multiple sequence alignment techniquessuch as Clustal W (Thompson et al. 1994), sincethese completely neglect structural information. Inour study, MARNA helped to generate multiplealignment based on pairwise sequence—structureinformation of RNAs. Undoubtedly, the processof alignment was greatly facilitated by the putativesecondary structure model predicted with com-parative analyses (Fig. 6B). The most strikingfeature of the primary sequence tree is theposition of Paramecium tetraurelia, which isnested within the scuticociliates, albeit with weakbootstrap support (Fig. 6A). This is in contrast tothe well-established position of Parameciumamong the peniculines in trees based on othermolecular (SSU rRNA) and morphological data,thus emphasizing the need for congruent results
from several different lines of evidence in order toestablish robust phylogenies.
Billoud et al. (2000) focused on the rRNAmolecules of Cirripedia (barnacles) comparingprimary and secondary structure information. Inthis study ‘‘molecular morphometrics’’, i.e. themeasurable structural parameters of the mole-cules (geometrical features, bond energies, basecomposition, etc.), were used as specific char-acters to construct a phylogenetic tree. Subbotinet al. (2007) converted the original sequence datainto 28 symbol codes using secondary structureinformation. Although phylogenetic trees based onrRNA were successfully reconstructed, their meth-ods were not suitable for our study because of thedifficulty of defining substructural homology inITS2 structure alignment. Wolf et al. (2005b) wrotethe software CBCAnalyzer for detecting speciesthat are discriminated by their sexual incompat-ibility. However, due to the low number of CBCs,the block CBC Tree can be used only forreconstructing the phylogeny of a small set ofclosely related taxa (Muller et al. 2007). Someattempts have also been made to formalize thesecondary structure comparisons, although diffi-culties persist in determining a distance betweentwo related structures with variable topologies(Fontana et al. 1993; Nakaya et al. 1996). Recently,several algorithms based on the ‘‘string’’ strategyhave been proposed (Jiang et al. 2002; Zhanget al. 1999). Wang et al. (2007) developed the stringstrategy based on the highly divergent ITS1 to inferphylogeny among the Pectinidae (scallops). In ourstudy, we adopted the string strategy to revealphylogenetic relationships among scuticociliatesand related species (Fig. 6C).
The tree topologies based on the three datasetssuggest that a deep phylogenetic signal has beenretained in the ITS2 sequences of those speciesused in our study. The availability of SSU rRNAsequences allows us to compare our ITS2 results toan independent phylogeny. The putative secondarystructure of ITS2 appears to be effective and helpfulin finding the clustering patterns, although thepositions of ‘‘ambiguous’’ genera such as Dexio-trichides—Cardiostomatella complex, Pleuronemaand Pseudocohnilembus as well as the familyOrchitophryidea require further consideration.
As reported recently, the Dexiotrichides—Cardiostomatella complex exhibits a mixture ofhymenostome and scuticociliate features withrespect to: ciliary pattern; obliquely oriented Tetra-hymena-like membranelles; a deeply excavatebuccal cavity; an evenly curved paroral mem-brane; a short fragment with densely packed

ARTICLE IN PRESS
529ITS2 Structure of Scuticociliates and Related Species
dikinetids at the anterior end of SK1 (Song et al.2005). Our analysis including putative secondarystructure prediction supports the scheme pre-sented by Li et al. (2006) in which this assemblagemay represent an evolutionary divergent offshootand a new order Loxocephalida, comprisingDexiotrichides—Cardiostomatella complex, withinthe subclass Scuticociliatia is suggested. Thishypothesis awaits further reinvestigation to testthe topology of existing phylogenetic trees.
Pleuronema branches at a deep level in the SSUrRNA tree and groups with Schizocalyptra in aclade that is basal to the other scuticociliates withstrong support (Fig. 6D). By contrast, in the ITS2secondary structure trees, Pleuronema is groupedwith Cardiostomatella and Dexiotrichides in aclade that is sister to the ‘‘true’’ scuticociliatess.s. (Fig. 6C). During the divisional processes ofPleuronema, considerable parental membranellardedifferentiation occurs with changes in shape aswell as orientation (Ma et al. 2003). This is analternative kind of ‘‘scuticobuccokinetal’’ type,which is quite different from that of Dexiotrichidesor Cardiostomatella (Song et al. 2005). These datasuggest that Pleuronema should be placed withinthe ‘‘true’’ scuticociliates s.s., although a reliablealignment could not be obtained among thelineages within this complex clade becausePleuronema was the only representative of theorder Pleuronematida (Fig. 6B).
Members of the genus Pseudocohnilembus arecharacterized by their two highly specialized paroralmembranes parallel to each other and about equalin length (Song et al. 2002). Based on moleculardata, Pseudocohnilembus, Paranophrys and Uro-nema were apparently closely related (Shang et al.2006). The data presented here demonstrate thatPseudocohnilembus is a sister group to Meta-nophrys and both form an isolated position in theSSU rRNA tree (Fig. 6D); whereas Pseudocohnilem-bus clusters with the clade containing Mesanophrysand Anophryoides in the ITS2 trees (Fig. 6C).
The monophyly of the family Orchitophryidea isin doubt and phylogeny within this group is notwell resolved. Lynn and Small (2002) placedParanophrys, Mesanophrys, Metanophrys, Ano-phryoides and Orchitophrya in the family Orchito-phryidea. The molecular data reported here,however, suggest that the family Orchitophryideaappears to be paraphyletic. Paranophrys, Uro-nema and Parauronema always form a clade, thesister group of which is still ambiguous yetMetanophrys differs from the closely relatedgenus Paranophrys only in the terminal positionof the anterior end of the paroral membrane
(adjacent to the middle of M2 versus adjacent tothe anterior end of M2). The ITS2 trees revealedsimilar relationships (Fig. 6A—C), whereas in thetree based on SSU rRNA gene sequence data(Fig. 6D) Metanophrys as well as Pseudocohni-lembus branched basally within the order Philas-terida except for the two genera Miamiensis andPhilasterides. In addition, Song and Wilbert (2000)noted that the genus Anophryoides should besynonymized with Paranophrys, and that it differsfrom Mesanophrys in the structure of the buccalapparatus. The ITS2 region of Anophryoides ismore similar to the consensus sequence ofMesanophrys spp. and Orchitophrya than to thatof Paranophrys, indicating that Anophryoidesmight be phylogenetically closer to Mesanophrys.Since a few taxa are nested together, it is difficultto determine which tree has the most potential toaccurately demonstrate scuticociliate phylogeny.Clearly more data are needed in order to resolvethe early branches and the evolutionary relation-ships among the Orchitophryidea.
To summarize, ITS2 combines information mostgermane to genus- and species-level systematics,although it may also be useful at family or evenhigher levels and thus be useful for resolvingscuticociliate phylogeny. In order to produce amore stable phylogeny of the scuticociliates,however, sequence data for additional key taxaare needed. No single gene is thought to contain asufficient number of informative sites to produce anunequivocal phylogeny. The results of the presentstudy suggest that ITS2 gene sequence data mightprovide important phylogenetic evidence to sup-plement that from other genes, and at a relativelysmall cost considering its short length.
Methods
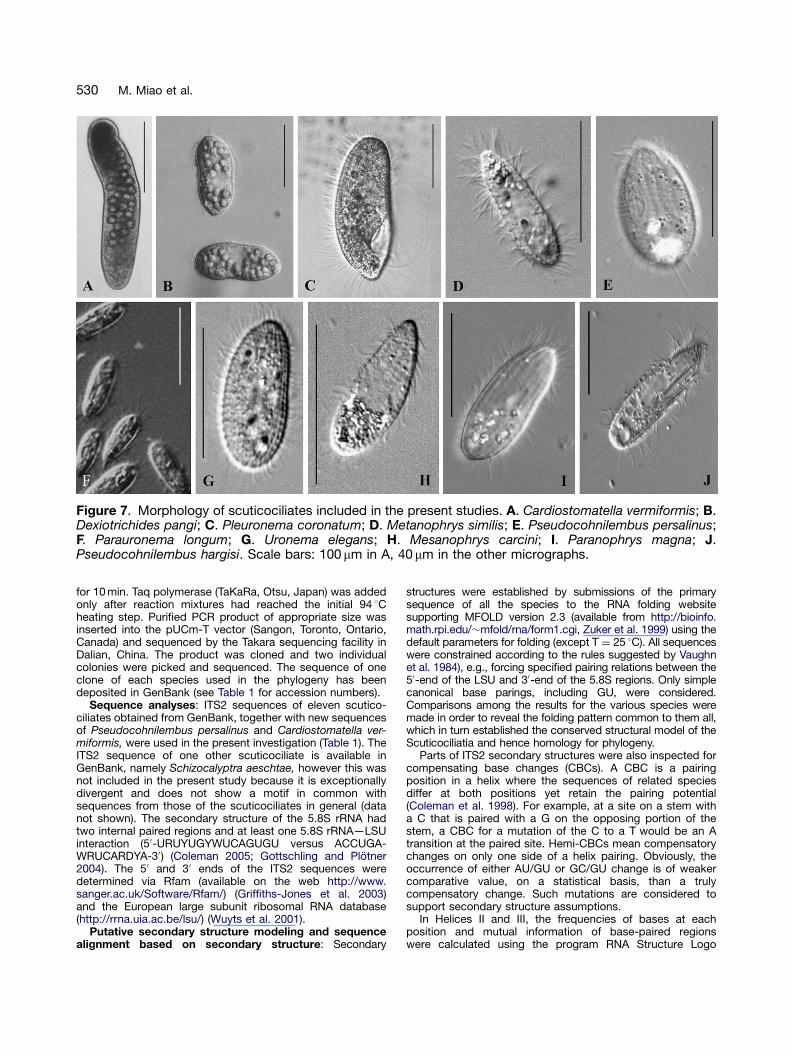
DNA extraction, amplification and sequencing: Cardiosto-matella vermiformis (Kahl 1928) and Pseudocohnilembuspersalinus Evans & Thompson, 1964 were obtained from thecell bank of the senior author’s laboratory and are availableupon request. The morphologies of these two isolates, plusthose of eight other scuticociliates used in the present studyare shown in Figure 7.
DNA was extracted following Chen and Song (2002) andused as a template for polymerase chain reactions (PCR)utilizing the primers ITS-F (50-GTA GGT GAA CCT GCG GAAGGA TCA TTA-30) and ITS-R (50-TAC TGA TAT GCT TAA GTTCAG CGG-30) that are complementary to conserved regionsand which encompass the 30 end of SSU rRNA, the whole ofthe ITS1, 5.8S rRNA and ITS2 regions, and the 50 end of LSUrRNA (Goggin and Murphy 2000). PCR conditions were asfollows: predenaturation at 94 1C for 5 min followed by thirtycycles of denaturation at 94 1C for 1 min; annealing at 63 1C for1 min; extension at 72 1C for 1 min; a final extension at 72 1C
ARTICLE IN PRESS
Figure 7. Morphology of scuticociliates included in the present studies. A. Cardiostomatella vermiformis; B.Dexiotrichides pangi; C. Pleuronema coronatum; D. Metanophrys similis; E. Pseudocohnilembus persalinus;F. Parauronema longum; G. Uronema elegans; H. Mesanophrys carcini; I. Paranophrys magna; J.Pseudocohnilembus hargisi. Scale bars: 100 mm in A, 40 mm in the other micrographs.
530 M. Miao et al.
for 10 min. Taq polymerase (TaKaRa, Otsu, Japan) was addedonly after reaction mixtures had reached the initial 94 1Cheating step. Purified PCR product of appropriate size wasinserted into the pUCm-T vector (Sangon, Toronto, Ontario,Canada) and sequenced by the Takara sequencing facility inDalian, China. The product was cloned and two individualcolonies were picked and sequenced. The sequence of oneclone of each species used in the phylogeny has beendeposited in GenBank (see Table 1 for accession numbers).
Sequence analyses: ITS2 sequences of eleven scutico-ciliates obtained from GenBank, together with new sequencesof Pseudocohnilembus persalinus and Cardiostomatella ver-miformis, were used in the present investigation (Table 1). TheITS2 sequence of one other scuticociliate is available inGenBank, namely Schizocalyptra aeschtae, however this wasnot included in the present study because it is exceptionallydivergent and does not show a motif in common withsequences from those of the scuticociliates in general (datanot shown). The secondary structure of the 5.8S rRNA hadtwo internal paired regions and at least one 5.8S rRNA—LSUinteraction (50-URUYUGYWUCAGUGU versus ACCUGA-WRUCARDYA-30) (Coleman 2005; Gottschling and Plotner2004). The 50 and 30 ends of the ITS2 sequences weredetermined via Rfam (available on the web http://www.sanger.ac.uk/Software/Rfam/) (Griffiths-Jones et al. 2003)and the European large subunit ribosomal RNA database(http://rrna.uia.ac.be/lsu/) (Wuyts et al. 2001).
Putative secondary structure modeling and sequencealignment based on secondary structure: Secondary
structures were established by submissions of the primarysequence of all the species to the RNA folding websitesupporting MFOLD version 2.3 (available from http://bioinfo.math.rpi.edu/�mfold/rna/form1.cgi, Zuker et al. 1999) using thedefault parameters for folding (except T ¼ 25 1C). All sequenceswere constrained according to the rules suggested by Vaughnet al. 1984), e.g., forcing specified pairing relations between the50-end of the LSU and 30-end of the 5.8S regions. Only simplecanonical base parings, including GU, were considered.Comparisons among the results for the various species weremade in order to reveal the folding pattern common to them all,which in turn established the conserved structural model of theScuticociliatia and hence homology for phylogeny.
Parts of ITS2 secondary structures were also inspected forcompensating base changes (CBCs). A CBC is a pairingposition in a helix where the sequences of related speciesdiffer at both positions yet retain the pairing potential(Coleman et al. 1998). For example, at a site on a stem witha C that is paired with a G on the opposing portion of thestem, a CBC for a mutation of the C to a T would be an Atransition at the paired site. Hemi-CBCs mean compensatorychanges on only one side of a helix pairing. Obviously, theoccurrence of either AU/GU or GC/GU change is of weakercomparative value, on a statistical basis, than a trulycompensatory change. Such mutations are considered tosupport secondary structure assumptions.
In Helices II and III, the frequencies of bases at eachposition and mutual information of base-paired regionswere calculated using the program RNA Structure Logo

ARTICLE IN PRESS
531ITS2 Structure of Scuticociliates and Related Species
(http://www.cbc.dtu.dk/�gorodkin/appl/slogo.html) (Gorodkinet al. 1997). Structural motifs among ‘‘typical’’ scuticociliateswere identified based on the structure logo. For each struc-tural domain, the position, number of base-pairs, unpairedbases in bulge and/or interior loops, and the GC content ofbase-paired regions were investigated and compared.
Construction of phylogenetic trees: Three differentphylogenetic analyses were carried out from different data-sets: Dataset I. Only primary sequences of ITS2 were alignedusing a computer-assisted procedure, Clustal W, ver. 1.80(Thompson et al. 1994). Dataset II. Primary sequences andsecondary structures were combined, and the alignmentcould be obtained at the MARNA web server (Siebert andBackofen 2005) (http://biwww2.informatik.uni-freiburg.de/Software/MARNA/index.html). As the default setting, the basedeletion was scored 2.0, base mismatch 1.0, arc removing2.0, arc breaking 1.5, and mismatch 1.8 with ensemble ofshaped structures. The two alignments were used to developphylogeny following multiple algorithms. The alignment wassubmitted to the best-fit model from MRMODELTEST (Nylan-der 2004) obtained by the Akaike Information Criterion (AIC) inorder to perform a search with the branch and boundalgorithm in PAUP*(V.4.0b10) (Swofford 2002). Heuristicsearches and a 100-fold bootstrap analysis were applied formaximum likelihood analysis using the PhyML V3.0 program(via http://www.phylogeny.fr/phylo_cgi/phyml.cgi). A BayesianInference (BI) tree was run in the computer program, MrBayesv3.0b4 (Huelsenbeck and Ronquist 2001), using the MarkovChain Monte Carlo (MCMC) algorithm. The chain length was5,000,000 generations with trees sampled every 100 genera-tions and an initial burn-in of 10,000. Maximum-parsimony(MP) calculation was analyzed in PAUP (V.4.0b10), andbootstrap re-sampled 1000 times. Dataset III. Only putativesecondary structures of ITS2, which were presented as stringin bracket notation, were used for structural alignment. In thebracket notion, base pairs are denoted by matching brackets,dots denote unpaired bases. The distance matrix wasproduced by the Vienna RNA distance server (http://bioweb.-pasteur.fr/seqanal/interfaces/rnadistance.html) and the fullstring algorithm was selected. The two distance matriceswere subjected to phylogenetic analysis by neighbor joiningusing MEGA 3.1. Tetrahymena americanis (GenBankaccession number AY833381) and Paramecium tetraurelia(GenBank accession number AY833390) were chosen asout-groups to the 13 species in above analyses. Differenttrees were compared to assess the stability of internalbranches in the resulting topology.
Phylogenetic trees based on SSU rRNA gene sequenceswere also constructed for comparative purposes. Sequencealignment was analyzed using ML, BI and MP according to thedescription above. The nucleotide sequences used in thispaper are available from the GenBank/EMBL databases underthe following accession numbers: Anophryoides haemophila(U51554), Cardiostomatella vermiformis (AY881632), Cohni-lembus verminus (Z22878), Cyclidium glaucoma (Z22879),C. plouneouri (U27816), Dexiotrichides pangi (AY212805),Entorhipidium tenue (AY541688), E. triangularis (AY541690),Epistylis chrysemydis (AF335514), Frontonia tchibisovae(DQ885987), Glaucoma chattoni (X56533), Ichthyophthiriusmultifiliis (U17354), Lembadion bullinum (AF255358), Licno-phora macfarlandi (AF527758), Loxophyllum rostratum(DQ411864), L. jini (EF123708), Mesanophrys carciniAY103189, Metanophrys similis (AY314803), Miamiensisavidus (AY550080), Paramecium woodruffi (AF255362),Paranophrys magna (AY103191), Parauronema longum(AY212807), Philasterides dicentrarchi (AY642280), Plagiopy-
liella pacifica (AY541685), Pleuronema coronatum (AY103188),Pseudocohnilembus marinus (Z22880), P. persalinus(AY551906), Pseudokeronopsis carnea (AY881633), P. flava(DQ227798), Pseudovorticella sinensis (DQ845295), Schizo-calyptra aeschtae (DQ777744), Tetrahymena australis(M98015), Thyrophylax vorax (AY541686), Uronema elegans(AY103190), U. marinum 1 (DQ867073), U. marinum 2(DQ867072), Uronemella filificum (EF486866), and Zootham-nopsis sinica (DQ190469). A karyorelictid ciliate, Loxodesstriatus (U24248), was selected as the outgroup species.
Acknowledgements
The work is supported by the ‘‘National NaturalFoundation of Science of China’’ (Project no.30670280), and the Darwin Initiative Program(Project no. 14-015) which is funded by the UKDepartment for Environment, Food and RuralAffairs. We would like to express gratitude toDr. Marc Gottschling (Freie Universitat Berlin,Germany), Prof. Annette W. Coleman (BrownUniversity, USA), Prof. Marco Oliverio (LaSapienza University, Italy), Dr. Peter Foster (Nat-ural History Museum, London) and Prof. GuanpinYang (Ocean University of China, China) for theirvaluable comments on the manuscript. We arealso grateful to Mr. Yangang Wang and HonganLong postgraduates of the Laboratory of Proto-zoology, OUC, for sampling.
References
Alverez I, Wendel JF (2003) Ribosomal ITS sequences andplant phylogenetic inference. Mol Phylogenet Evol 29: 417—434
Billoud B, Guerrucci M, Masselot M, Deutsch JS (2000)Cirripede phylogeny using a novel approach: molecularmorphometrics. Mol Biol Evol 17: 1435—1445
Chen Z, Song W (2002) Phylogenetic positions of Aspidiscasteini and Euplotes vannus within the order Euplotida(Hypotrichia: Ciliophora) inferred from complete small subunitribosomal RNA gene sequences. Acta Protozool 41: 1—9
Coleman AW (2003) ITS2 is a double-edged tool for eukaryoteevolutionary comparisons. Trends Genet 19: 370—375
Coleman AW (2005) Paramecium aurelia revisited. J EukaryotMicrobiol 52: 68—77
Coleman AW (2007) Pan-eukaryote ITS2 homologiesrevealed by RNA secondary structure. Nucleic Acids Res 35:3322—3329
Coleman AW, Preperata RM, Mehrotra B, Mai JC (1998)Derivation of the secondary structure of the ITS-1 transcript inVolvocales and its taxonomic correlations. Protist 149: 135—146
Corliss JO (1979) The Ciliated Protozoa: Characterization,Classification, and Guide to the Literature, 2nd Ed. PergamonPress, London

ARTICLE IN PRESS
532 M. Miao et al.
Cote CA, Greer CL, Peculis BA (2002) Dynamic conforma-tional model for the role of ITS2 in pre-rRNA processing inyeast. RNA 8: 786—797
Fabry S, Kohler A, Coleman AW (1999) Intraspecies analysis:comparison of ITS sequence data and gene intron sequencedata with breeding data for a worldwide collection of Goniumpectorale. J Mol Eol 48: 94—101
Fontana W, Konings DAM, Stadler PF, Schuster P (1993) Sta-tistics of RNA secondary structures. Biopolymers 33: 1389—1404
Foissner W (1996) Ontogenesis in Ciliated Protozoa withEmphasis on Stomatogenesis. In Hausmann K, Bradbury PC(eds) Ciliates: Cells as Organisms. Gustav Fischer, Stuttgart,Germany, pp 95—177
Goggin CL, Murphy NE (2000) Conservation of sequence in theinternal transcribed spacers and 5.8S ribosomal RNA amonggeographically separated isolates of parasitic scuticociliates(Ciliophora, Orchitophryidae). Dis Aquat Organ 40: 79—83
Goldman WE, Goldberg G, Bowman LH, Steinmetz D,Schlessinger D (1983) Mouse rDNA: sequences and evolu-tionary analysis of spacer and mature RNA regions. Mol CellBiol 3: 1488—1500
Good L, Intine RVA, Nazar RN (1997) The ribosomal-RNA-processing pathway in Schizosaccharomyces pombe. Eur JBiochem 247: 314—321
Gorodkin J, Heyer LJ, Brunak S, Stormo GD (1997)Displaying the information contents of structural RNA align-ments: the structure logos. Comput Appl Biosci 13: 583—586
Gottschling M, Plotner J (2004) Secondary structure modelsof the nuclear internal transcribed spacer regions and 5.8SrRNA in Calciodinelloideae (Peridiniaceae) and other dino-flagellates. Nucleic Acids Res 32: 307—315
Gottschling M, Keupp H, Plotner J, Knop R, Willems H,Kirsch M (2005) Phylogeny of calcareous dinoflagellates asinferred from ITS and ribosomal sequence data. Mol Phylo-genet Evol 36: 444—455
Griffiths-Jones S, Bateman A, Marshall M, Khanna A, EddySR (2003) Rfam: an RNA family database. Nucleic Acids Res31: 439—441
Gutell RR, Larsen N, Woese CR (1994) Lessons from anevolving rRNA 16S and 23S rRNA structures from acomparative perspective. Microbiol Rev 58: 10—26
Hoef-Emden K (2007) Revision of the genus Cryptomonas(Cryptophyceae) II: incongruences between the classicalmorphospecies concept and molecular phylogeny in smallerpyrenoid-less cells. Phycologia 46: 402—428
Huelsenbeck JP, Ronquist F (2001) MRBAYES: Bayesianinference of phylogenetic trees. Bioinformatics 17: 754—755
Jiang T, Lin G, Ma B, Zhang K (2002) A general edit distancebetween two RNA structures. J Comput Biol 9: 371—388
Joseph N, Krauskopf E, Vera MI, Michot B (1999)Ribosomal internal transcribed spacer 2 (ITS2) exhibits acommon core of secondary structure in vertebrates and yeast.Nucleic Acids Res 27: 4533—4540
Li L, Song W, Warren A, Wang Y, Ma H, Hu X, Chen Z (2006)Phylogenetic position of the marine ciliate, Cardiostomatellavermiforme (Kahl, 1928) Corliss, 1960 inferred from the
complete SSrRNA gene sequence, with establishment of anew order Loxocephalida n. ord. (Ciliophora, Oligohymeno-phorea). Europ J Protistol 42: 107—114
Lott TJ, Burns BM, Zancope-Oliveira R, Elie CM, Reiss E(1998) Sequence analysis of the internal transcribed spacer 2(ITS2) from yeast species within the genus Candida. CurrMicrobiol 36: 63—69
Lynn DH, Small E (2002) Phylum Ciliophora Doflein, 1901. InLee JJ, Bradbury PC, Leedale GF (eds) An Illustrated Guide tothe Protozoa. 2nd Ed. Allen Press Inc., Lawrence, Kansas,USA, pp 371—656
Ma H, Gong J, Song W (2003) Stomatogenesis of the marineciliate Pleuronema coronatum Kent, 1881 (Ciliophora, Scuti-cociliatida). Acta Zool Sinica 49: 829—834
Mai JC, Coleman AW (1997) The internal transcribed spacer2 exhibits a common secondary structure in green algae andflowering plants. J Mol Evol 44: 258—271
Maroteaux L, Herzog M, Soyer-Gobillard MO (1985)Molecular organization of dinoflagellate ribosomal DNA:evolutionary implications of the deduced 5.8S rRNA second-ary structure. Biosystems 18: 307—319
Michot B, Joseph N, Mazan S, Bachellerie JP (1999)Evolutionarily conserved structural features in the ITS2 ofmammalian pre-rRNAs and potential interactions with thesnoRNA U8 detected by comparative analysis of new mousesequences. Nucleic Acids Res 27: 2271—2282
Morade JF, Small EB (1994) Morphology and stomatogenesisof Mesanophrys pugettensis n. sp. (Scuticociliatida: Orchito-phryidae): a facultative parasitic ciliate of the Dungeness crab,Cancer magister (Crustacea: Decapoda). Trans Am MicroscSoc 113: 343—364
Muller T, Philippi N, Dandekar T, Schultz J, Wolf M (2007)Distinguishing species. RNA 13: 1469—1472
Nakaya A, Yonezawa A, Yamamoto K (1996) Classificationof RNA secondary structures using the techniques of clusteranalysis. J Theor Biol 183: 105—117
Nylander JAA (2004) MrModeltest Version 2. Distributed bythe author. Department of Systematic Zoology, EvolutionaryBiology Centre, Uppsala University, Uppsala, Sweden
Oliverio M, Cervelli M, Mariottini P (2002) ITS2 rRNAevolution and its congruence with the phylogeny of muricidneogastropods (Caenogastropoda, Muricoidea). Mol Phylo-genet Evol 25: 63—69
Park G, Im KI, Yong TS (2003) Phylogenetic relationship ofribosomal ITS2 and mitochondrial COI among diploid and triploidParagonimus westermani isolates. Korean J Parasitol 41: 47—55
Ragan MA, Cawthorn RJ, Despres B, Murphy CA, SinghRK, Loughlin MB, Bayer RC (1996) The lobster parasiteAnophryoides haemophila (Scuticociliatida: Orchitophryidae):nuclear 18S rDNA sequence, phylogeny and detection usingoligonucleotide primers. J Eukaryot Microbiol 43: 341—346
Schlotterer C, Hauser MT, von Haeseler A, Tautz D (1994)Comparative evolutionary analysis of rDNA ITS regions inDrosophila. Mol Biol Evol 11: 513—522
Schultz J, Maisel S, Gerlach D, Muller T, Wolf M (2005) Acommon core of secondary structure of the internal transcribedspacer 2 (ITS2) throughout the Eukaryota. RNA 11: 361—364

ARTICLE IN PRESS
533ITS2 Structure of Scuticociliates and Related Species
Schultz J, Muller T, Achtziger M, Seibel PN, Dandekar T,Wolf M (2006) The internal transcribed spacer 2 database — AWeb server for (not only) low level phylogenetic analyses.Nucleic Acids Res 34: W704—W707
Selig C, Wolf M, Muller T, Dandekar T, Schultz J (2008) TheITS2 database II: homology modelling RNA structure formolecular systematics. Nucleic Acids Res 36: D377—D380
Shang H, Song W (2002) Studies on three closely-relatedscuticociliates using RAPD fingerprinting and RFLP riboprint-ing. High Tech Lett 12: 87—90 (in Chinese)
Shang H, Song W, Warren A (2003) Phylogenetic positions oftwo ciliates, Paranophrys magna and Mesanophrys carcini(Ciliophora: Oligohymenophorea), within the subclass Scuti-cociliatia inferred from complete small subunit rRNA genesequences. Acta Protozool 42: 171—181
Shang H, Song W (2005) Separation and relationship of tenmarine scuticociliates (Protozoa, Ciliophora) using RAPDfingerprinting method. Acta Oceanol Sin 24: 78—85
Shang H, Song WB, Warren A, Li L, Chen Z (2006)Phylogenetic positions of two marine ciliates, Metanophryssimilis and Pseudocohnilembus hargisi (Protozoa, Ciliophora,Scuticociliatia), inferred from complete small subunit rRNAgene sequence. Progr Nat Sci 16: 373—378
Siebert S, Backofen R (2005) MARNA: multiple alignment andconsensus structure prediction of RNAs based on sequencestructure comparisons. Bioinformatics 21: 3352—3359
Small EB (1967) The Scuticociliatida, a new order of the classCiliatea (phylum Protozoa, subphylum Ciliophora). Trans AmMicrosc Soc 86: 345—370
Song W, Warren A, Roberts D, Shang H, Ma H, Wilbert N,Miao M, Yi Z (2005) Re-evaluation of the phylogeneticposition of the genus Dexiotrichides (Protozoa, Ciliophora,Scuticociliatida) inferred from stomatogenetic and molecularinformation for Dexiotrichides pangi. Progr Nat Sci 15:700—707
Song W, Wilbert N (2000) Redefinition and redescription ofsome marine scuticociliates from China. I, with report of a newspecies, Metanophrys sinensis nov. spec. (Ciliophora, Scuti-cociliatida). Zool Anz (Syst) 239: 45—74
Song W, Wilbert N (2002) Reinvestigations of three ‘‘well-known’’ marine scuticociliates: Uronemella filificum (Kahl,1931) nov. gen., nov. comb., Pseudocohnilembus hargisiEvans and Thompson, 1964 and Cycilidium citrullus Cohn1865, with description of the new genus Uronemella (Proto-zoa, Ciliophora, Scuticociliatida). Zool Anz 241: 317—331
Stoeck T, Przybos E, Schmidt HJ (1998) A combination ofgenetics with inter- and intra-strain crosses and RAPD-finger-
prints reveals different population structures within the Para-mecium aurelia species complex. Europ J Protistol 34: 348—355
Subbotin SA, Sturhan D, Vovlas N, Castillo P, Tambe JT,Moens M, Baldwin JG (2007) Application of the secondarystructure model of rRNA for phylogeny: D2—D3 expansionsegments of the LSU gene of plant-parasitic nematodes fromthe family Hoplolaimidae Filipjev, 1934. Mol Phylogenet Evol43: 881—890
Swofford DL (2002) PAUP*: Phylogenetic Analysis UsingParsimony (* and Other Methods). Sinauer, Sunderland, MA
Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W:improving the sensitivity of progressive multiple sequencealignment through sequence weighting, position-specific gappenalties and weight matrix choice. Nucleic Acids Res 22:4673—4680
Vaughn JC, Sperbeck SJ, Ransey WJ, Lawrence CB (1984)A universal method for the secondary structure of 5.8Sribosomal RNA molecules, their contact sites with 28Sribosomal RNAs, and their prokaryotic equivalent. NucleicAcids Res 12: 7479—7502
Vollmer SV, Palumbi SR (2004) Testing the utility of internallytranscribed spacer sequences in coral phylogenetics. MolEcol 13: 2763—2772
Wang S, Bao Z, Li N, Zhang L, Hu J (2007) Analysis of thesecondary structure of ITS1 in Pectinidae: implications forphylogenetic reconstruction and structural evolution. MarBiotechnol (NY) 9: 231—242
Wolf M, Achtziger M, Schultz J, Dandekar T, Muller T(2005a) Homology modeling revealed more than 20,000 rRNAinternal transcribed spacer 2 (ITS2) secondary structures.RNA 11: 1616—1623
Wolf M, Friedrich J, Dandekar T, Muller T (2005b)CBCAnalyzer: inferring phylogenies based on compensatorybase changes in RNA secondary structures. In Silico Biol 5: 27
Wuyts J, De Rijk P, Van de Peer Y, Winkelmans T, DeWachter R (2001) The European large subunit ribosomal RNAdatabase. Nucleic Acids Res 29: 175—177
Zhang K, Wang L, Ma B (1999) Computing Similarity betweenRNA Structures. In Crochemore M, Paterson M (eds)Proceedings of the 10th Symposium on Combinatorial PatternMatching. Lecture Notes in Computer Science, Vol. 1645.Springer, London, UK, pp 281—293
Zuker M, Mathews DH, Turner DH (1999) Algorithmsand Thermodynamics for RNA Secondary Structure Prediction:A Practical Guide. In Barciszewski J, Clark BFC (eds) RNABiochemistry and Biotechnology. NATO ASI Series. KluwerAcademic Publishers, Dordrecht, the Netherlands, pp 11—43
Copyright © 2022 FDOKUMEN




















