
Gcn5 Promotes Acetylation, Eviction, and Methylation of Nucleosomes in Transcribed Coding Regions
Upload
independentCategory
view
1download
0
Biological Control 49 (2009) 207–213
Contents lists available at ScienceDirect
Biological Control
journal homepage: www.elsevier .com/locate /ybcon
Internal transcribed spacer-2 restriction fragment length polymorphism(ITS-2-RFLP) tool to differentiate some exotic and indigenous trichogrammatidegg parasitoids in India
G. Ashok Kumar a, Sushil K. Jalali a,*, T. Venkatesan a, Richard Stouthamer b, P. Niranjana a, Y. Lalitha a
a Insect Biotechnology Laboratory, Post Bag No. 2491, H.A. Farm Post, Bellary Road, Hebbal, Bangalore 560024, Karnataka, Indiab Department of Entomology, University of California, Riverside, CA, USA
a r t i c l e i n f o a b s t r a c t
Article history:Received 27 March 2008Accepted 6 February 2009Available online 20 February 2009
Keywords:Molecular identificationITS-2-RFLPTrichogrammatidsRestriction enzymes
1049-9644/$ - see front matter � 2009 Elsevier Inc. Adoi:10.1016/j.biocontrol.2009.02.010
* Corresponding author. Fax: +91 080 23411961.E-mail address: [email protected] (S.K. Jala
ITS-2-RFLP method was employed to distinguish 12 different trichogrammatids consisting of indigenousand exotic species such as Trichogrammatoidea armigera and Tr. bactrae, Trichogramma achaeae, T. chilonis,T. japonicum, T. embryophagum, T. pretiosum (Thelytokous Form—TF), T. brassicae, T. dendrolimi, T. evanes-cens and T. mwanzai. ITS-2 region was amplified; complete ITS-2 sequences of nine species were depos-ited in Genbank. The size of the amplified product ranged from 500 to 900 bp. Restriction enzymedigestion of ITS-2 region showed different banding profiles for these 12 species. Dichotomous keys usingthe size of the ITS-2 product and the restriction fragment length polymorphism for the enzymes (EcoRI,MseI, MvaI, and TaqI) allowed quick species identification of these trichogrammatids.
� 2009 Elsevier Inc. All rights reserved.
1. Introduction
The family Trichogrammatidae consists of about 650 species(Grissel and Schauff, 1990) and is the most widely used taxon of nat-ural enemies in various biocontrol programs worldwide. Althoughabout 210 species of Trichogramma are known to attack the eggs ofvarious crop pests (Pinto, 1998), at least 12 species are used in bio-logical control programs. With the increasing preference for sustain-able agriculture and awareness about side effects fromindiscriminate use of insecticides, biological control agents areattracting more attention. However, with so many species, ecotypesand strains available and very frequent exchange of culture material,correct identification is the first step for a successful biological con-trol program. Morphological identification of trichogrammatids re-mains difficult because of their minute size (<1 mm long), is timeconsuming and requires specialized skills (Pinto and Stouthamer,1994). Morphological identification of most species is based on sub-tle differences in male genitalia (Nagarkatti and Nagaraja, 1971,1977). Many important species share similar genital structures, forc-ing workers to rely on less dependable characters that often are in-tra-specifically variable (Pinto et al., 1989). Given the economicimportance of trichogrammatids in biological control programs,species identification must be quick, simple and widely applicable.Novel approaches that used DNA sequences of the internal tran-scribed spacer 2 (ITS-2) helped to solve this difficulty (Stouthamer
ll rights reserved.
li).
et al., 1999). Ribosomal RNA (rDNA) comprises of three genes 18S,5.8S and 28S separated by internal transcribed spacers (ITS-1 andITS-2). ITS regions evolve faster than the regions coding for the ribo-somal RNA (Hoy, 1994). The ITS region is widely used in identifica-tion studies (Hillis and Dixon, 1991). Sequence and restrictionanalysis of ITS-2 region has been used to distinguish Trichogrammaspecies collected from tomato fields in Portugal (Silva et al., 1999).
In India, twenty six species of trichogrammatids have been re-corded and among them Trichogrammatoidea armigera, Tr. bactrae,Trichogramma achaeae, T. chilonis and T. japonicum are widely dis-tributed and used against lepidopteran pests in several crops(Singh and Jalali, 1994; Jalali et al., 2003). Elsewhere in the worldT. evanescens, T. dendrolimi, T. pretiosum and T. brassicae are mostfrequently used in the field (Smith, 1996). Trichogramma galloi isnow extensively used as biocontrol agent in sugar cane fields ofBrazil (Parra and Zucchi, 2004). In the past decade, several exoticspecies have been introduced in India to target pests such as cod-ling moth (Cydia pomonella Linnaeus), diamondback moth (Plutellaxylostella Linnaeus), corn borer [Chilo partellus (Swinhoe)] andAmerican bollworm (Helicoverpa armigera Hübner) (Jalali et al.,2003). With the introduction of these non-native trichogrammatidspecies in addition to the native ones, there is a need for a quickand reliable identification technique to identify these parasitoids.While the identification of males of these species could be doneusing morphological features, many of the specimens collectedfrom the field are females that are not identifiable using morphol-ogy. In the present study we develop an identification methodbased on the ITS-2 sequences of the 12 most commonly used spe-

Table 1Trichogrammatids studied, analysis performed, identity and origin of the cultures.
Linedesignate
Studies Species Origin Collectionyear
Tra1 1,2 Trichogrammatoideaarmigera
India 1990
Trb1 1,2 Trichogrammatoidea bactrae India 1990Ta1 1,2,3 Trichogramma achaeae India 1990Tj1 1,2,3 T. japonicum India 1990Tem1 1,2,3 T. embryophagum Russia 1993Tc1 1,2,3 T. chilonis India 1990Tp1 1,2,3 T. pretiosum USA 1993Tev1 1,2,3 T. evanescens France 2004Tm1 1,2,3 T. mwanzai Kenya 2003Tbra1 1,2,3 T. pretiosum (TF) USA 1993Td1 1,2,3 T. dendrolimi Germany 1994Tbrass1 1,2,3 T. brassicae USA 2004
1, ITS-2; 2, sequencing; 3, restriction analysis.
Fig. 1. PCR amplified ITS-2 plus flanking regions of 5.8s and 28s rDNA fromdifferent trichogrammatids (Lanes: M, 50 bp DNA perfect ladder (Novagen); 1, Tr.armigera; 2, Tr. bactrae; 3, T. achaeae; 4, T. embryophagum; 5, T. japonicum; 6, T.chilonis; 7, T. dendrolimi; 8, T. evanescens; 9, T. mwanzai; 10, T. brassicae; 11, T.pretiosum (TF); 12, T. pretiosum).
208 G. Ashok Kumar et al. / Biological Control 49 (2009) 207–213
cies in India: Trichogrammatoidea armigera Manjunath, Tr. bactraeNagraja, Trichogramma achaeae Nagaraja and Nagarkatti, T. chilonisIshii, T. japonicum Ashmead, T. embryophagum Hartig, T. pretiosumRiley, T. pretiosum Riley (Thelytokous form-TF), T. brassicae Bez-denko, T. dendrolimi Matsumura, T. evanescens Westwood and T.mwanzai Schulten and Feijen.
2. Materials and methods
2.1. Parasitoid culture
We studied 12 isofemale lines identified by Dr. H. Nagarajaas Trichogrammatoidea armigera, and Tr. bactrae, Trichogramma
Fig. 2. (a) Multiple sequence alignment of ITS-2 region of group II Trichogramma specienucleotides are absent, � indicates for similar nucleotides).
achaeae, T. chilonis, T. japonicum (all indigenous), T. embryopha-gum, T. pretiosum, T. pretiosum (TF) often mistakenly referredto as T. brasiliense Ashmead (Pinto, 1997), T. brassicae, T. dendro-limi, T. evanescens and T. mwanzai (all exotic). Voucher speci-mens are kept by Dr. H. Nagaraja in PDBC (Bangalore, India).All cultures were maintained on rice moth, Corcyra cephalonica(Stainton) eggs (killed by ultraviolet irradiation) at 26 ± 1 �C,RH 60 ± 5%, LD 14:10. The origin of each species studied is pro-vided in Table 1.
s. (b) ITS-2 sequences of group III Trichogramma species (A dash indicates that the

Fig. 2 (continued)
G. Ashok Kumar et al. / Biological Control 49 (2009) 207–213 209
2.2. Isolation of DNA
DNA was extracted using chelating agent Chelex100 (5%) (Bio-rad). One or two wasp previously frozen at �80 �C were crushedwith a micro pestle in 20 ll of chelex. The homogenate was incu-bated at 56 �C for 3 h followed by 10 min at 100 �C. The superna-tant was stored at �20 �C until used.
2.3. ITS-2 PCR
The PCR reaction was performed in 50 ll reaction volumesusing Biorad icycler, 2 ll DNA template, 5 ll (10�) Taq assaybuffer, 1 ll dNTP’s (each in 10 mM concentration), 1 llforward and reverse primers (10 picomoles/ll), 0.25 ll taq poly-merase (1 U). The primers used to amplify the ITS-2 region were

Fig. 2 (continued)
210 G. Ashok Kumar et al. / Biological Control 49 (2009) 207–213
50-TGTGAACTGCAGGACACATG-30 (forward) and 50-GTCTTGCCTGCTCTGAG-30 (reverse) (Stouthamer et al., 1999). The thermal cy-cling condition for PCR consisted of 30 cycles (Den: 94 �C for1 min, Ann: 55 �C for 1 min, Ext: 72 �C for 2 min, with an initialdenaturation: 95 �C for 5 min and final extension at 72 �C for10 min). PCR products were electrophoresed on 1.8% agarose gel(ACROS). Gels were stained using ethidium bromide. Molecularstandards were run along with the samples for reference.
2.4. Sequencing
The PCR products were gel eluted using Qiagen gel clean up kitand direct sequenced using an Applied Prism 310. The resulting se-quences were blasted against sequences in GenBank to confirmthat the sequence was indeed ITS-2.
Table 2Restriction sizes following digestion of PCR amplified ITS-2 plus flanking regions of 5.8s a
Species Accession No.
Group I (800–850 bp)Tr. armigera EU251072 900 bpTr. bactrae EU251071 800 bp
Group II (570–620 bp)ITS-2 size
T. achaeae EU251070 627 bpT. embryophagum DQ344044 592 bp
T. japonicum DQ471294 565 bp
Group III (500–550 bp)ITS-2 size
T. chilonis DQ220703 538 bp
T. dendrolimi DQ344045 515 bp
T. evanescens DQ381280 546 bp
T. mwanzai DQ381279 500 bp
T. brassicae DQ314611 522 bp
T. pretiosum (thelytokous form) DQ381281 525 bp
T. pretiosum DQ525178 528 bp
Note: � denotes no site present.a Only bands larger than 100 bp are listed.
2.5. Restriction enzyme analysis
Restriction digestion of the PCR product was performed in a 20 llvolume (10 ll PCR product, 2 ll (10�) reaction buffer, 1 ll reactionenzyme and 7 ll distilled water). The mixture was incubated over-night at appropriate temperatures as per manufacturer’s specifica-tions. RFLP products were analyzed by electrophoresis in 1.8%agarose gel. 50 bp DNA perfect ladder (Novagen) was included toestimate the size of restriction fragment. Four restriction enzymesused were EcoRI, MseI, MvaI and TaqI (MBI Fermentos). EcoRI wascommon enzyme selected for all the species and MseI to differentiatebetween T. chilonis, T. dendrolimi, T. evansecens, T. mwanzai, T. brass-icae and T. pretiosum. These restriction enzymes were selected usingBioedit (Hall, 1999) restriction fragment analysis, as these wereindicative for trichogrammatids used in the study.
nd 28s rDNA of 12 trichogrammatids with various restriction enzymes.
ITS-2 size
EcoRI�229 bp363 bp�
EcoRI MseI MvaI TaqIa
233 bp � 470 bp 141 bp305 bp 68 bp 129 bp
143 bp263 bp 434 bp 447 bp 144 bp252 bp 87 bp 68 bp 129 bp268 bp 454 bp 478 bp 305 bp278 bp 92 bp 68 bp243 bp 447 bp 432 bp 323 bp257 bp 53 bp 68 bp300 bp 412 bp 454 bp 138 bp222 bp 32 bp 68 bp 135 bp
68 bp 129 bp� � 457 bp 307 bp
68 bp� � 393 bp 309 bp
68 bp63 bp
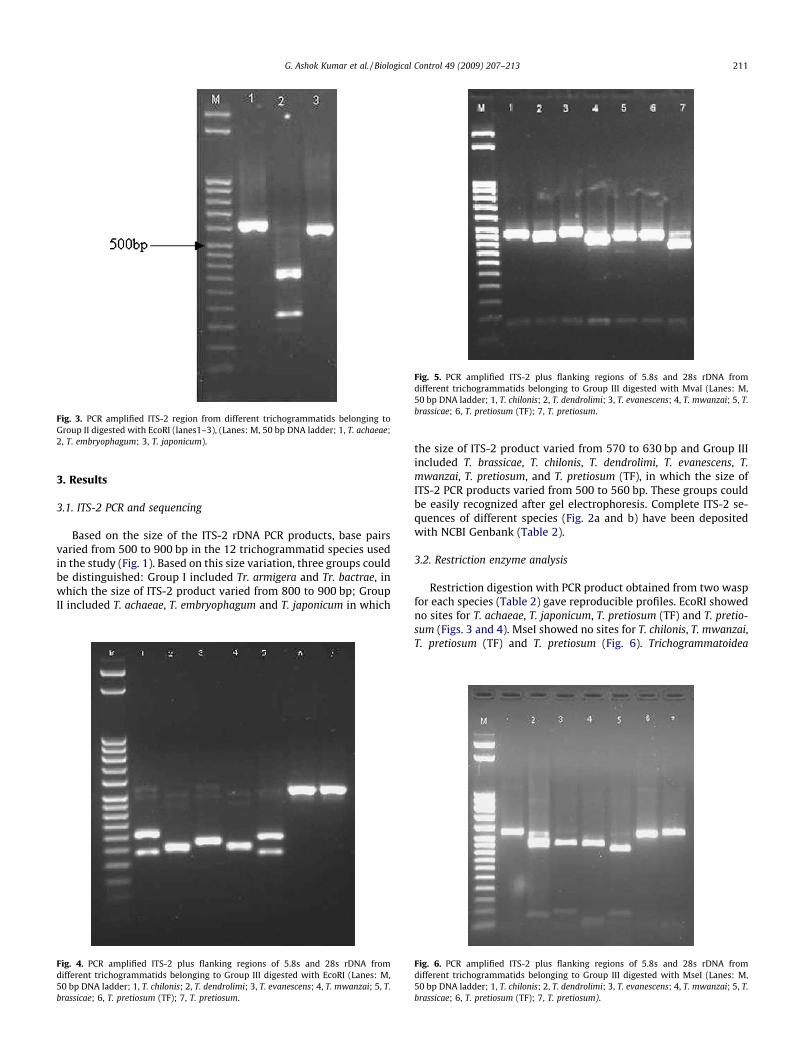
Fig. 3. PCR amplified ITS-2 region from different trichogrammatids belonging toGroup II digested with EcoRI (lanes1–3), (Lanes: M, 50 bp DNA ladder; 1, T. achaeae;2, T. embryophagum; 3, T. japonicum).
Fig. 5. PCR amplified ITS-2 plus flanking regions of 5.8s and 28s rDNA fromdifferent trichogrammatids belonging to Group III digested with MvaI (Lanes: M,50 bp DNA ladder; 1, T. chilonis; 2, T. dendrolimi; 3, T. evanescens; 4, T. mwanzai; 5, T.brassicae; 6, T. pretiosum (TF); 7, T. pretiosum.
G. Ashok Kumar et al. / Biological Control 49 (2009) 207–213 211
3. Results
3.1. ITS-2 PCR and sequencing
Based on the size of the ITS-2 rDNA PCR products, base pairsvaried from 500 to 900 bp in the 12 trichogrammatid species usedin the study (Fig. 1). Based on this size variation, three groups couldbe distinguished: Group I included Tr. armigera and Tr. bactrae, inwhich the size of ITS-2 product varied from 800 to 900 bp; GroupII included T. achaeae, T. embryophagum and T. japonicum in which
Fig. 4. PCR amplified ITS-2 plus flanking regions of 5.8s and 28s rDNA fromdifferent trichogrammatids belonging to Group III digested with EcoRI (Lanes: M,50 bp DNA ladder; 1, T. chilonis; 2, T. dendrolimi; 3, T. evanescens; 4, T. mwanzai; 5, T.brassicae; 6, T. pretiosum (TF); 7, T. pretiosum.
the size of ITS-2 product varied from 570 to 630 bp and Group IIIincluded T. brassicae, T. chilonis, T. dendrolimi, T. evanescens, T.mwanzai, T. pretiosum, and T. pretiosum (TF), in which the size ofITS-2 PCR products varied from 500 to 560 bp. These groups couldbe easily recognized after gel electrophoresis. Complete ITS-2 se-quences of different species (Fig. 2a and b) have been depositedwith NCBI Genbank (Table 2).
3.2. Restriction enzyme analysis
Restriction digestion with PCR product obtained from two waspfor each species (Table 2) gave reproducible profiles. EcoRI showedno sites for T. achaeae, T. japonicum, T. pretiosum (TF) and T. pretio-sum (Figs. 3 and 4). MseI showed no sites for T. chilonis, T. mwanzai,T. pretiosum (TF) and T. pretiosum (Fig. 6). Trichogrammatoidea
Fig. 6. PCR amplified ITS-2 plus flanking regions of 5.8s and 28s rDNA fromdifferent trichogrammatids belonging to Group III digested with MseI (Lanes: M,50 bp DNA ladder; 1, T. chilonis; 2, T. dendrolimi; 3, T. evanescens; 4, T. mwanzai; 5, T.brassicae; 6, T. pretiosum (TF); 7, T. pretiosum).

Table 3Dichotomous key to differentiate the 12 Trichogramma species based on PCR productof the ITS-2 region plus flanking regions of the 5.8S and 28S rDNA and restrictionanalysis with various restriction enzymes.
1 Size of the PCR product larger than 700 bp 2Size of the PCR product smaller than 700 bp 3
2 Size of the PCR product �850 bp Tr. armigeraSize of the PCR product �800 bp Tr. bactrae
3 Size of the PCR product larger than 560 bp 4Size of the PCR product smaller than 560 bp 6
4 PCR product restricted with EcoRI results in two bands T.embryophagum
PCR product restricted with EcoRI results in a single band 55 Size of PCR product larger than 600 bp T. achaeae
Size of the PCR product �600 bp T. japonicum6 PCR product restricted with MseI results in a single band
>500 bp7
PCR product restricted with MseI results at least one band<500 bp
8
7 PCR product restricted with EcoRI results in two bands(�240, �300 bp)
T. chilonis
PCR product restricted with EcoRI results in a single band T. pretiosum8 PCR product restricted with EcoRI results in two bands
clearly different in sizeT. brassicae
PCR product restricted with EcoRI results in two bands verysimilar in size often only visible as a single band
9
9 Size of the band 275 bp T. evanescensSize of the band 250 bp 10
10 PCR product restricted with TaqI results in largest band of>300 bp
T. mwanzai
PCR product restricted with TaqI in largest product of<150 bp
T. dendrolimi
Fig. 7. PCR amplified ITS-2 plus flanking regions of 5.8s and 28s rDNA fromdifferent trichogrammatids belonging to Group III digested with TaqI (Lanes: M,50 bp DNA ladder; 1, T. chilonis; 2, T. dendrolimi; 3, T. evanescens; 4, T. mwanzai;5, T. brassicae; 6, T. pretiosum (TF); 7, T. pretiosum).
212 G. Ashok Kumar et al. / Biological Control 49 (2009) 207–213
armigera and Tr. bactrae belonging to genus Trichogrammatoideacould be easily differentiated after amplification of ITS-2 region.Restriction digestion with EcoRI enzyme enabled differentiationof 10 species, while restriction digestion with MseI enabled differ-entiation of six species. MvaI enzyme (Fig. 5) proved useful to dif-ferentiate T. pretiosum (TF) from T. pretiosum. Cleavage activity ofTaqI (Fig. 7) enzyme aided in discrimination of T. dendrolimi andT. mwanzai. Based on restriction patterns, a dichotomous key wasconstructed for easy differentiation of these 12 trichogrammatids(Table 3).
4. Discussion
The ITS-2-RFLP tool was able to differentiate the 12 Trichogram-ma species used in this study. Length variation was observed in theITS-2 region and it clearly separated the two genera, i.e., Tricho-gramma and Trichogrammatoidea from each other. Such length var-iation in the ITS region has been reported before (Orrego andAgudelo-Silva, 1993; Sappal et al., 1995). Using the length variationand the RFLP patterns we were able to construct a dichotomouskey (Table 3). Studies on ITS-2-RFLP analysis to identify Tricho-gramma species have been limited to particular geographical re-gions (Silva et al., 1999; Ciociola et al., 2001; Thompson et al.,2003) or particular crops over a wide area (Pinto et al., 2002). Thisis one of the first attempts to differentiate some indigenous andexotic species involved in various biological control programmesin India. The intra-specific variability in both total ITS-2 lengthand restriction fragments length appears to be low within thetwo genera studied, as was found earlier in the analysis of ITS-2 se-quences of some North American species (Stouthamer et al., 1999).PCR–RFLP analysis is very simple, fast and reliable. PCR–RFLP toolhas been used in detection and species identification of fungalpathogen and nematodes (Dendis et al., 2003; Roberta et al., 2003).
The restriction pattern shown for the MvaI digestion of thearrhenotokous form shows that the ITS-2 for this species is poly-morphic for the MvaI site, so within individuals of the arrhenotok-ous T. pretiosum two different ITS-2 families exist one with therestriction site and one without that site. This is visible in the gelbecause for the arrhenotokous population two bands are visible alarger band of the same size as the thelytokous form and a smallerband only found in the arrhenotokous form. Slight variation withinindividuals is commonly found in Trichogramma wasps, for in-stance Trichogramma kaykai has a similar polymorphism for anEcoRI site (Stouthamer et al., 1999). The differentiation using MvaIbetween the arrhenotokous and thelytokous form of T. pretiosumworks for these particular populations that have been importedto India. However, other arrhenotokous populations of T. pretiosumoriginating from Brazil and some from Riverside, California alsoshow the same pattern as the arrhenotokous form shown here(see for instance GenBank Accession Nos. AF082820, AY187261).
These methods are intended to replace the traditional morpho-logical methods when large number of individuals need to be iden-tified that belong to a known group of trichogrammatids. Forinstance, for checking laboratory contamination or the fate of re-leased species in the field. These DNA based methods are faster, be-cause no mounting of the specimens is required, and allowsidentification of both females and males. Application of the DNAbased methods should free up time for systematists to identifyand describe new species. From the present study it can be con-cluded that rDNA markers and restriction enzyme patterns canbe used for reliable differentiation between two genera and alsobetween various indigenous and exotic species to be field evalu-ated in India.
Acknowledgments
We thank Dr. R.J. Rabindra, Project Director, Project Directorateof Biological Control, Bangalore for his keen interest in the studyand Ms. R. Rajeshwari for valuable technical assistance.
References
Ciociola Jr., A.C., Zucchi, R.A., Stouthamer, R., 2001. Molecular key to seven Brazilianspecies of Trichogramma (Hymenoptera: Trichogrammatidae) using sequencesof the ITS-2 region and restriction analysis. Neotropical Entomology 30, 256–262.
Dendis, M., Horvath, R., Michalek, J., Ruzicka, F., Grijalva, M., Bartos, M., Bendik, J.,2003. PCR–RFLP detection and species identification of fungal pathogens in

G. Ashok Kumar et al. / Biological Control 49 (2009) 207–213 213
patients with febrile neutropenia. Clinical microbiology and infection 9, 1191–1201.
Grissel, E.E., Schauff, M.E., 1990. A handbook of the families of Nearctic Chalcidoidea(Hymenoptera). Entomological Society of Washington 1, 85.
Hall, T.A., 1999. BioEdit: a user-friendly biological sequence alignment and analysisprogram for Windows 95/98/NT. Nucleic Acids Symposium Series 41, 95–98.
Hillis, D.M., Dixon, M.T., 1991. Ribosomal DNA: molecular evolution andphylogenetic inference. Quarterly Review of Biology 66, 411–416.
Hoy, M.A., 1994. Insect Molecular Genetics: An Introduction to Principles andApplications. Academic Press, San Diego. 540pp.
Jalali, S.K., Rabindra, R.J., Rao, N.S., Dasan, C.B., 2003. Mass production oftrichogrammatids and chrysopids. Project Directorate of Biological Control,Bangalore 560024, Technical Bulletin No. 33, 16pp.
Nagarkatti, S., Nagaraja, H., 1971. Redescription of some known species ofTrichogramma showing the importance of male genitalia as a diagnosticcharacter. Bulletin of Entomological Research 61, 13–31.
Nagarkatti, S., Nagaraja, H., 1977. Biosystematics of Trichogramma andTrichogrammatoidea species. Annual Review of Entomology 22, 157–176.
Orrego, C., Agudelo-Silva, F., 1993. Genetic variation in the parasitoid wasp Trichogramma(Hymenoptera: Trichogrammatidae) revealed by DNA amplification of a section of thenuclear ribosomal repeat. Florida Entomologist 76, 519–524.
Parra, J.R.P., Zucchi, R.A., 2004. Trichogramma in Brazil: feasibility of use aftertwenty years of research. Neotropical Entomology 33, 261–271.
Pinto, J.D., 1997. Trichogrammatoidea brasiliensis (Ashmead)—New combination fora species historically placed in Trichogramma. Proceedings of the BiologicalSociety of Washington 99, 593–596.
Pinto, J.D., 1998. Systematics of the North American species of Trichogramma(Hymenoptera: Trichogrammatidae). Memoirs of Entomological Society ofWashington 22, 1–287.
Pinto, J.D., Stouthamer, R., 1994. Systematics of the Trichogrammatidae withemphasis on Trichogramma. In: Wajnberg, E., Hassan, S.A. (Eds.), BiologicalControl with Egg Parasitoids. CAB International, Oxon, UK, pp. 1–36.
Pinto, J.D., Koopmanschap, A.B., Platner, G.R., Stouthamer, R., 2002. The NorthAmerican Trichogramma (Hymenoptera: Trichogrammatidae) parasitizingcertain Tortricidae (Lepidoptera) on apple and pear with ITS-2 DNAcharacterization and redescription of a new species. Biological Control 23,134–142.
Pinto, J.D., Velten, R.K., Platner, G.R., Oatman, E.R., 1989. Phenotypic plasticity andtaxonomic characters in Trichogramma. Annals of the Entomological Society ofAmerica 85, 413–422.
Roberta, L.C., Omar, S.C., Cristine, L.F.G.M., Carlos, G.T., Marcia, C.F.S., Renata, B.,Rafel, M., Walter, S.L., Henrique, L.L., 2003. Molecular differentiation ofAngiostrongylus castaricensis, A. cantonensis and A. vasorum by polymerasechain reaction–restriction fragment length polymorphism. Mem Inst OswaldoCruz Rio de Janeiro 98, 1039–1043.
Sappal, N.P., Jeng, R.S., Hubbes, M., Liu, F., 1995. Restriction fragment lengthpolymorphisms in polymerase chain reaction amplified ribosomal DNAs ofthree Trichogramma (Hymenoptera: Trichogrammatidae) species. Genome 38,419–425.
Silva, I.M.M.S., Honda, J., van Kan, F., Hu, S.J., Neto, L., Pintureau, B., Stouthamer, R.,1999. Molecular differentiation of five Trichogramma species occurring inPortugal. Biological Control 16, 177–184.
Singh, S.P., Jalali, S.K., 1994. Trichogrammatids. Project Directorate of BiologicalControl, H.A. Farm Post, Bangalore 560024, Technical Bulletin No. 7, 95pp.
Smith, S.M., 1996. Biological control with Trichogramma: Advances, successes, andpotential of their use. Annual Review of Entomology 41, 375–406.
Stouthamer, R., Hu, J., van Kan, F.J.P.M., Platner, G.R., Pinto, J.D., 1999. The utilityof internal transcribed spacer 2 DNA sequences of the nuclear ribosomalgene for distinguishing sibling species of Trichogramma. BioControl 43, 421–440.
Thompson, L.J., Rundle, B.J., Carew, M.E., Hoffmann, A.A., 2003. Identification andcharacterization of Trichogramma species from south-eastern Australia usingthe internal transcribed spacer 2 (ITS-2) region of the ribosomal gene complex.Entomologia Experimentalis et Applicata 106, 235–240.
Copyright © 2022 FDOKUMEN




















