
Analgesic properties of S100A9 C-terminal domain: a mechanism dependent on calcium channel...
12
doi: 10.1111/j.1472-8206.2009.00686.x ORIGINAL ARTICLE Analgesic properties of S100A9 C-terminal domain: a mechanism dependent on calcium channel inhibition Camila Squarzoni Dale a,b , Christophe Altier c , Nicolas Cenac a,b , Renata Giorgi d , Maria Aparecida Juliano e , Luiz Juliano e , Gerald W. Zamponi c , Nathalie Vergnolle a,b,f * a INSERM, U563, Centre de Physiopathologie de Toulouse Purpan, Toulouse F-31300, France b Universite ´ Toulouse III Paul Sabatier, Toulouse F-31000, France c Department of Physiology and Biophysics Hotchkiss Brain Institute, University of Calgary, Calgary, AB T2N 4N1, Canada d Laboratory of Pathophysiology, Butantan Institute, Sa ˜o Paulo, SP 05505-900, Brazil (RG) e Department of Biophysics, Pharmacology Institute, Federal University of Sa ˜o Paulo, Sa ˜o Paulo, SP 04044-020, Brazil f Department of Pharmacology and Therapeutics, University of Calgary, AB T2N4N1, Canada (CSD, NC, NV) INTRODUCTION The S100A9 protein belongs to the family of S100 proteins and is a calcium binding protein of the EF-hand type [1,2]. Anti-inflammatory properties have been described for S100A9, which inhibits leukocyte traf- ficking [3], arachidonic acid metabolism [4] and is able to de-activate activated peritoneal macrophages [5]. S100A9 protein also displays potent antinociceptive properties in a mouse model of inflammatory pain [6]. S100A9, released by neutrophil, is responsible for the inhibition of pain response in a model of neutrophilic peritonitis induced by glycogen in mice [7,8]. Recent reports suggest that the C-terminal domain of the S100A9 protein (19 amino acids from the position 92–110 of murine S100A9) is responsible for most of Keywords antinociception, dorsal root ganglia neurons, inflammation, N-type calcium channels, S100A9, voltage operated calcium channels Received 10 July 2008; revised 7 November 2008; accepted 16 December 2008 *Correspondence and reprints: [email protected] ABSTRACT Calcium-binding protein S100A9 and its C-terminus peptide (mS100A9p) are anti-inflammatory and induce antinociception in rodents. We investigated the mechanisms involved in this effect, and whether they depend or not on the anti- inflammatory properties of mS100A9p. In mice, mS100A9p inhibited thermal and mechanical hyperalgesia and allodynia induced by either carrageenan or formalin, without interfering with paw edema. mS100A9p also inhibited myeloperoxidase activity (MPO), a marker of granulocyte infiltration, induced by carrageenan, but increased MPO after formalin intraplantar injection. The in vivo analgesic properties of mS100A9p were independent of opioid receptor activation. Calcium flux into dorsal root ganglia neurons induced by KCl was inhibited by mS100A9p, suggesting that this protein is able to inhibit signaling, in sensory neurons. The inhibitory effects of mS100A9p on primary afferent signaling were neither due to intracellular calcium store inhibition nor to calcium chelating properties. However, mS100A9p was able to inhibit calcium currents carried by transiently expressed N-type, but not L-type calcium channels, as demonstrated both by gene transfection techniques and electrophysiology. These data demonstrate that mS100A9p interferes with mecha- nisms involved in nociception, hyperalgesia and calcium signaling in sensory neurons, modulating primary afferent nociceptive signal by inhibiting activation of N-type voltage operated calcium channels. ª 2009 The Authors Journal compilation ª 2009 Socie ´ te ´ Franc ¸aise de Pharmacologie et de The ´ rapeutique Fundamental & Clinical Pharmacology 23 (2009) 427–438 427
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Analgesic properties of S100A9 C-terminal domain: a mechanism dependent on calcium channel...

doi: 10.1111/j.1472-8206.2009.00686.x
O R I G I N A L
A R T I C L E
Analgesic properties of S100A9 C-terminaldomain: a mechanism dependent on calciumchannel inhibition
Camila Squarzoni Dalea,b, Christophe Altierc, Nicolas Cenaca,b,Renata Giorgid, Maria Aparecida Julianoe, Luiz Julianoe,Gerald W. Zamponic, Nathalie Vergnollea,b,f*aINSERM, U563, Centre de Physiopathologie de Toulouse Purpan, Toulouse F-31300, FrancebUniversite Toulouse III Paul Sabatier, Toulouse F-31000, FrancecDepartment of Physiology and Biophysics Hotchkiss Brain Institute, University of Calgary, Calgary, AB T2N 4N1,
CanadadLaboratory of Pathophysiology, Butantan Institute, Sao Paulo, SP 05505-900, Brazil (RG)eDepartment of Biophysics, Pharmacology Institute, Federal University of Sao Paulo, Sao Paulo, SP 04044-020, BrazilfDepartment of Pharmacology and Therapeutics, University of Calgary, AB T2N4N1, Canada (CSD, NC, NV)
I N T R O D U C T I O N
The S100A9 protein belongs to the family of S100
proteins and is a calcium binding protein of the EF-hand
type [1,2]. Anti-inflammatory properties have been
described for S100A9, which inhibits leukocyte traf-
ficking [3], arachidonic acid metabolism [4] and is
able to de-activate activated peritoneal macrophages [5].
S100A9 protein also displays potent antinociceptive
properties in a mouse model of inflammatory pain [6].
S100A9, released by neutrophil, is responsible for the
inhibition of pain response in a model of neutrophilic
peritonitis induced by glycogen in mice [7,8]. Recent
reports suggest that the C-terminal domain of the
S100A9 protein (19 amino acids from the position
92–110 of murine S100A9) is responsible for most of
Keywords
antinociception,
dorsal root ganglia neurons,
inflammation,
N-type calcium channels,
S100A9,
voltage operated calcium
channels
Received 10 July 2008;
revised 7 November 2008;
accepted 16 December 2008
*Correspondence and reprints:
A B S T R A C T
Calcium-binding protein S100A9 and its C-terminus peptide (mS100A9p) are
anti-inflammatory and induce antinociception in rodents. We investigated the
mechanisms involved in this effect, and whether they depend or not on the anti-
inflammatory properties of mS100A9p. In mice, mS100A9p inhibited thermal and
mechanical hyperalgesia and allodynia induced by either carrageenan or formalin,
without interfering with paw edema. mS100A9p also inhibited myeloperoxidase
activity (MPO), a marker of granulocyte infiltration, induced by carrageenan, but
increased MPO after formalin intraplantar injection. The in vivo analgesic properties
of mS100A9p were independent of opioid receptor activation. Calcium flux into
dorsal root ganglia neurons induced by KCl was inhibited by mS100A9p, suggesting
that this protein is able to inhibit signaling, in sensory neurons. The inhibitory effects
of mS100A9p on primary afferent signaling were neither due to intracellular calcium
store inhibition nor to calcium chelating properties. However, mS100A9p was able to
inhibit calcium currents carried by transiently expressed N-type, but not L-type
calcium channels, as demonstrated both by gene transfection techniques and
electrophysiology. These data demonstrate that mS100A9p interferes with mecha-
nisms involved in nociception, hyperalgesia and calcium signaling in sensory
neurons, modulating primary afferent nociceptive signal by inhibiting activation of
N-type voltage operated calcium channels.
ª 2009 The Authors Journal compilation ª 2009 Societe Francaise de Pharmacologie et de TherapeutiqueFundamental & Clinical Pharmacology 23 (2009) 427–438 427

the anti-inflammatory and anti-nociceptive properties
of S100A9. As a matter of fact, a synthetic peptide
corresponding to this part of the C-terminal domain
of the murine calcium-binding protein S100A9
(mS100A9p) inhibits spreading and phagocytic activity
of adherent peritoneal cells [9], inhibits hyperalgesia
induced by carrageenan in rats [10] and inhibits
hyperalgesia and edema induced by a metalloprotease
[11]. Most recently, we demonstrated that mS100A9p
also inhibits the hyperalgesia induced by an agonist
peptide of protease-activated receptor 2 (PAR2), modu-
lating PAR2 activation-induced nociceptive signal on
primary afferents [12].
The C-terminus of S100A9 protein is rich in histidine
residues and it has been reported to be involved with
zinc- and arachidonic acid-binding motif of S100A9
[13,14]. This portion of the molecule shows some
similarities with the sequence of the neutrophil inhibi-
tory factor that inhibits chemotaxia and neutrophil
migration in vitro [15,16], and with the contact domain
of high molecular weight kininogen [17]. However, no
clear mechanisms have been described to explain the
anti-inflammatory or anti-nociceptive properties of
S100A9 or its C-terminal domain. In this study, we
examined the mechanisms by which the C-terminal
domain of S100A9 exerts its analgesic properties. We
determined: (i) that mS100A9p exerts analgesic proper-
ties independently of its anti-inflammatory effects; (ii)
that the analgesic properties of mS100A9p are indepen-
dent of opioid receptor activation; (iii) that mS100A9p
can act directly on sensory neurons, inhibiting calcium
signaling in primary afferents and (iv) we defined that
mS100A9p acts as a voltage operated N-type calcium
channel inhibitor.
M A T E R I A L S A N D M E T H O D S
Animals
Male C57Bl6 mice were used throughout this study.
Animals were maintained under controlled light cycle
(12/12 h) and temperature (22 ± 2 �C), with free access
to food and water. The study was conducted in accor-
dance with the guidelines of the Institutional Animal
Care and Use Committee and was approved by the Ethics
Committee on Animal Experimentation of the University
of Calgary.
Reagents
A peptide identical to the C-terminus of murine S100A9
protein (H-E-K-L-H-E-N-N-P-R-G-H-G-H-S-H-G-K-G-NH2 –
mS100A9p) was synthesized in solid phase by fluor-
enylmethyloxycarbonyl (FMOC) technique. Characteri-
zation and purification were performed by high
performance liquid chromatography, and its mass
evaluated by matrix assisted laser desorption/ionization
time-of-flight (MALDI-TOF) spectrometry. The peptide
was diluted in saline at the final concentration of 1 mg/
mL. Formalin, 5% solution, diluted in saline, was injected
by the intraplantar (i pl.) route (50 lL/paw) of mice.
Carrageenan, 2% solution, diluted in saline (carrageenan
lambda, type IV – Sigma, St Louis, MO, USA), was also
injected by the i.pl. route (50 lL/paw). Naloxone
(10 mg/kg – Sigma) was diluted in saline and injected
20 mins before carrageenan, by the subcutaneous route
[18]. Thapsigagin and calcium ionophore (A23187)
were purchased from Sigma.
Behavioral pain measures
Thermal nociception
Paw withdrawal latency to radiant heat stimulus was
measured using an Ugo Basile� Plantar test (Ugo Basile,
Comerio, Italy) essentially as preciously described [19].
The withdrawal latency was measured before and after
the i pl. injection of carrageenan (2%–50 lL), formalin
(5%–50 lL) or mS100A9p (1 lg – paw). Thermal
hyperalgesia was defined as a significant decrease in the
withdrawal latency compared with the basal measure-
ment at different times after treatment.
Mechanical hyperalgesia and allodynia
Mechanical hyperalgesia and allodynia of the hind paw
were assessed as described before [20]. Mice were placed
individually in a plastic cage. Von Frey filaments with
bending forces of 0.407 g (3.61 filament), 0.692 g (3.84
filament), and 1.202 g (4.08 filament) were pressed
perpendicularly against the plantar skin and held for 5 s.
The stimulation of the same intensity was applied three
times to each hind paw at intervals of several seconds.
The responses to these stimuli were ranked as follows: 0,
no response; 1, move away from von Frey filament; and
2, immediate flinching or licking of the hind paw.
Nociceptive score was calculated as follows:
Nociceptive score (%)
¼P
average score of each animalð Þ2� no. of animals tested
� 100
The nociceptive score was measured before and after
the i pl. injections of carrageenan (2%–50 lL), formalin
(5%–50 lL) or mS100A9p (1 lg/paw).
428 C.S. Dale et al.
ª 2009 The Authors Journal compilation ª 2009 Societe Francaise de Pharmacologie et de TherapeutiqueFundamental & Clinical Pharmacology 23 (2009) 427–438

All the nociceptive tests were performed according to
the guidelines published in a Guest Editorial in Pain on
ethical standards, for investigations of experimental pain
in animals [21].
Paw inflammation
Paw volume was measured using an electronic caliper,
before and at different times after i pl. injection of
carrageenan (2%–50 lL), formalin (5%–50 lL) and/or
mS100A9p (1 lg/paw). After 4 h of treatments, the paw
tissues of mice were assayed for myeloperoxidase activity
(MPO), as an index of tissue infiltration by granulocytes
as previously described [22–24].
Neuronal culture and calcium imaging
Dorsal root ganglia (DRG) neurons isolated from mice
were rinsed in hanks balanced salt solution (HBSS), and
incubated in HBSS containing 1% Papain (Worthing-
ton, Cedarlane, Hornby, ON, Canada) for 10 mins at
37 �C as previously described [25]. After a wash with
filtered Leibovitz’s L-15 Medium solution [glutamine
(200 mM), glucose 20%, fetal bovine serum FBS 10%],
DRG neurons were incubated in HBSS containing
collagenase (1 mg/mL-1) and dispase (4 mg/mL-1) at
37 �C for 10 mins. After titration, cells were plated in
Poly-L-ornithine-laminine (Sigma) glass-bottom petri
dishes (35 mm diameter; MatTek Corporation, Ashland,
MA, USA) and recovered with the complete culture
media [Minimum Essential Medium (MEM), 2.5% FBS,
1% penicillin/streptomycin, glutamine (200 mM), 1%
Dextrose and cytosine B-D-Arabinofuranoside (ARAC),
5-fluoro-2-desoxi-uridine (FUDR), uridine 10 lM each].
Cells were cultured for a minimum of 72 h. The petri
dishes were then washed twice with a Hank’s buffered
salt solution, pH 7.4 and incubated for 60 mins at
37 �C with Hank’s buffered salt solution supplemented
with 0.1% bovine serum albumin in the same solution
to which 3–5 lM of Fluo3-AM (Molecular Probes,
Eugene, OR, USA) was added. After the incubation
period, the petri dishes were washed twice again with
the assay N-2-hydroxyethylpiperazine-N¢-2-ethanesu-
fonic acid (HEPES) buffer (containing in mM: NaCl, 150;
KCl, 3; CaCl, 1.5; Hepes, 20; glucose, 10; sulfinpirazone,
0.25) of which 2 mL were left in each petridish.
Cells were observed using a wide-field fluorescence
olympus IX-70 microscope and an LC Plan FL 40X
objective. A series of 30 images in 90 s was acquired; the
first five images were used to determine the baseline.
After the fifth image, neurons were treated by an acute
administration of KCl (50 mM), calcium ionophore
A23187 (2 lM) and/or mS100A9p (0.5, 5, 50 or
100 lM). The experiment was repeated three times per
group. Fluorescence measurements reflecting elevations
of intracellular calcium were conducted at 460–490 nm
excitation and 515 nm emission in individual cells using
the acquisition program OpenLab software (Improvision
Inc a PerkinElmer Company, Waltham, MA, USA).
Transient expression of recombinant
calcium channels
Human embryonic kidney (HEK) tsA-201 cells were
cultured as described previously [26], with the exception
that cells were not moved to 28 �C after transfection.
Cells were split and plated on glass coverslips. High
voltage activated (HVA) calcium channels Cav1.2 and
Cav2.2 cDNA constructs were transfected with calcium
phosphate method at a ratio of 1 : 1 : 1 for calcium a1,
a2-d1 and b1b (2 lg each). Cells were used 24–48 h
after transfection for recording.
Electrophysiology
Electrophysiological recordings of channels expressed in
tsA-cells were conducted as described in detail previously
[26], with 20 mM barium as the charge carrier. Cells
were held at )100 mV and current voltage-relations
were acquired by stepping to various test potentials. Peak
current amplitude was normalized to cell capacitance to
obtain current densities. In all recordings, series resis-
tance was compensated by 85%. For recordings of
calcium currents in DRG neurons, current-voltage rela-
tionships were obtained by stepping to various test
potentials. Peak current was usually elicited between
)20 and 0 mV and was again normalized to cell
capacitance to obtain current densities.
The external recording solution for DRGs contained of
CaCl2 (2 mM), HEPES (10 mM), tetraethylamonium
chloride (TEACl) (160 mM) and glucose (10 mM), pH
7.4. For tsA-201 cells, it had 20 mM BaCl2 (20 mM),
MgCl2 (1 mM), HEPES (10 mM), TEACl (40 mM), glucose
(10 mM), CsCl (HVA- 65 mM) or BaCl2 (2 mM), MgCl2
(1 mM), HEPES (10 mM), TEACl (40 mM), glucose
(10 mM) and CsCl (LVA – 105 mM). Borosilicate glass
pipettes (2–4 MW) were filled with internal solution
containing CsCl (110 mM), MgCl2 (3 mM), ethylene
glycol tetraacetic acid (EGTA) (10 mM), HEPES
(10 mM), MgATP (3 mM) and GTP (0.6 mM), pH 7.2
(for DRGs) or CsMeSO4 (108 mM), MgCl2 (4 mM), EGTA
(9 mM), HEPES (9 mM), MgATP (2.6 mM) and LiGTP
(0.6 mM), pH to 7.2 (for tsA-201 cells). Peptide
(mS100A9p) was prepared daily in external solution
S100A9 analgesic properties 429
ª 2009 The Authors Journal compilation ª 2009 Societe Francaise de Pharmacologie et de TherapeutiqueFundamental & Clinical Pharmacology 23 (2009) 427–438

from 1 mM frozen stocks and was applied to cells by
microperfusion. Currents were elicited from a holding
potential of )90 mV (DRGs) or )100 mV (tsA-201 cells).
Presentation of data and statistical analysis
All data are presented as the mean ± SEM. Statistical
analysis of data was generated using GraphPAd Prism,
version 4.02 (GraphPad Software Inc., San Diego, CA,
USA). Statistical analysis between two samples was
performed using Student’s t-test. Statistical comparison
of more than two groups was performed using one way –
ANOVA with Tukey’s multiple comparisons post-test. In
all cases, a P value <0.05 was considered statistically
significant.
R E S U L T S
mS100A9p exerts antinociceptive effects
independently of anti-inflammatory properties
We investigated the anti-inflammatory and anti-nocicep-
tive effects of mS100A9p in experimental models of
inflammatory pain. Groups of mice injected by the i.pl.
route with carrageenan (2%) or formalin (5%) developed
thermal (decreased withdrawal latency) and mechanical
(increased nociceptive score) hyperalgesia (Figure 1a, b for
carrageenan and Figure 1e, f for formalin). Allodynia
(response to the innocuous stimulation of the 3.61 size
filament) was also observed after i.pl. injection of carra-
geenan (Figure 1b) or formalin (Figure 1f ). In addition,
mice injected with carrageenan or formalin showed
signs of inflammation such as edema (Figure 1c, g),
and granulocyte infiltration, as measured by MPO
activity (Figure 1d, h). Concomitant injection of 1 lg of
mS100A9p (final volume 50 lL), with carrageenan or
formalin, inhibited both thermal (Figure 1a and e) and
mechanical (Figure 1b and f ) hyperalgesia and allodynia
induced by either formalin or carrageenan. However, the
same dose of mS100A9p had no significant effect on
carrageenan- or formalin-induced edema (Figure 1c, 1g).
mS100A9p inhibited MPO activity induced by carra-
geenan injection in the mouse paw (Figure 1d), but
significantly increased MPO activity into the paw after
formalin injection (Figure 1h). We also evaluated the
effects of i pl. injections of mS100A9p alone, which did not
cause significant granulocyte infiltration compared to
vehicle-injected paws (Figure 1d) and caused a small
increase in paw volume (Figure 1c), which was not
significantly different from vehicle alone (not shown).
Overall, these data show analgesic properties for
i.pl. injection of mS100A9 peptide, which decreased
thermal and mechanical inflammatory allodynia and
hyperalgesia. The analgesic properties of mS100A9
were independent from the development of other
inflammatory parameters (edema and granulocyte
recruitment).
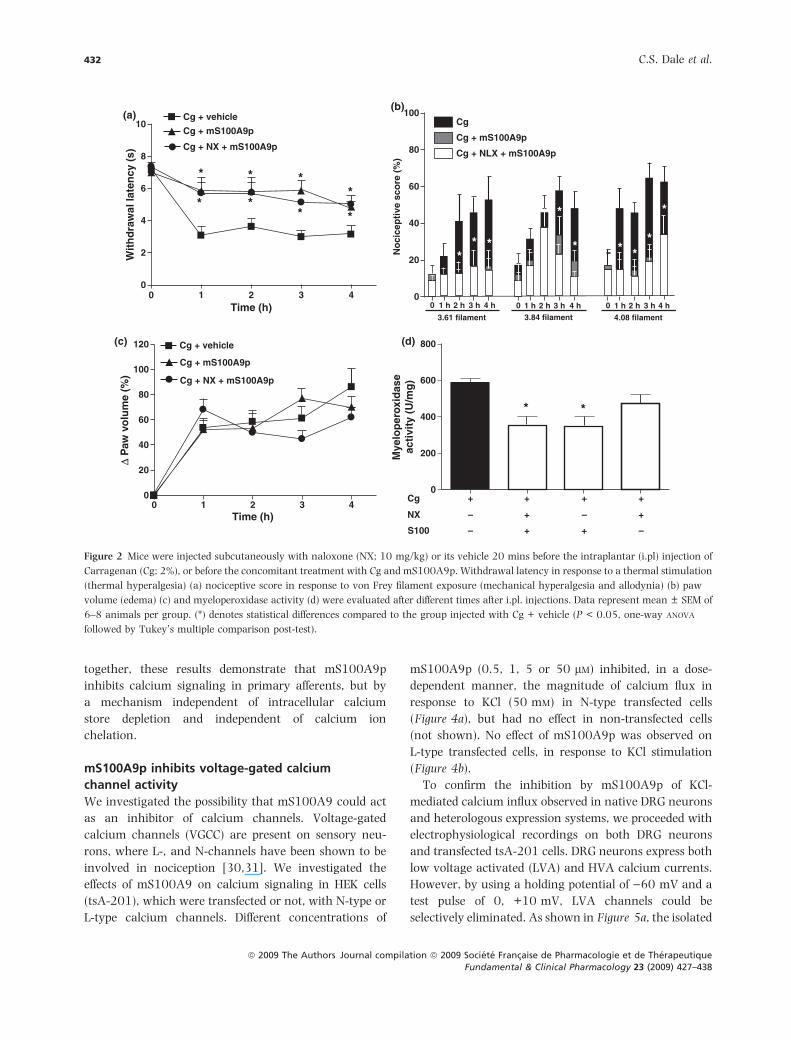
mS100A9p-induced analgesia is independent of
opioid receptor activation
Opioids are known to be important modulators of
nociception, and activation of opioid receptors on
primary afferents induces analgesia [27]. We investi-
gated whether the inhibitory effects of mS100A9p on
inflammatory hyperalgesia involved opioid receptor
activation. Naloxone treatment (10 mg/kg) antagonizes
opioid receptor activation non-selectively, inhibiting
mu-, delta- and kappa-opioid receptors [18]. The
analgesic effects of mS100A9p on carrageenan-induced
thermal (decreased withdrawal latency) or mechanical
(increased nociceptive score) hyperalgesia, and allo-
dynia were not affected by naloxone pre-treatment
(Figure 2a, b). Furthermore, naloxone treatment did not
modify carrageenan-induced edema and MPO activity
either (Figure 2c, d). These results suggest that opioid
receptor activation is not involved in the antinocicep-
tive effect of mS100A9p during carrageenan-induced
inflammation.
mS100A9p inhibits sensory neuron activity
We further investigated whether mS100A9p was able
to act directly on primary afferents, inhibiting signal-
ing in sensory neurons. Different concentrations of
mS100A9p (0.5, 5, 50 and 100 lM) did not induce
calcium mobilization in isolated DRG neurons, under
basal (unstimulated) conditions (not shown). However,
the same concentrations of mS100A9p, significantly
inhibited calcium mobilization induced by KCl (50 mM)
stimulation, in a dose-dependent manner (Figure 3a).
To investigate whether mS100A9p was able to block
calcium release from intracellular stores, DRG neurons
were treated with thapsigargin (250 nM), a specific
calcium inhibitor that promotes the discharge of
calcium from intracellular stores [28]. Pre-treatment
of DRG neurons with thapsigargin reduced KCl-
induced calcium signal, however, this treatment did
not modify the effect of mS100A9p on primary
afferents (Figure 3b). Moreover, mS100A9p had no
effect on DRG neuron signaling in response to 2 lM of
calcium ionophore, a carboxylic antibiotic that selec-
tively transfers calcium and other divalent cations
across biological membranes [29] (Figure 3c). Taken
430 C.S. Dale et al.
ª 2009 The Authors Journal compilation ª 2009 Societe Francaise de Pharmacologie et de TherapeutiqueFundamental & Clinical Pharmacology 23 (2009) 427–438

(a)
Cg
Cg + mS100A9p
mS100A9p
Vehicle
** *
* *
***
10
8
6
4
2
00 1 2 3 4
Time (h)
Wit
hd
raw
al la
ten
cy (
s)
(b) Cg + vehicleCg + mS100A9p70
60
50
40
30
20
10
00 1 h 2 h 3 h 4 h 0 1 h 2 h 3 h 4 h 0 1 h 2 h 3 h 4 h
3.61 filament
No
cice
pti
ve s
core
(%
)
3.84 filament 4.08 filament
* * *
*
*
**
*
(c) Cg + vehicle
Vehicle
100
80
60
40
20
00 1 2 3 4
Time (h)
Δ P
aw v
olu
me
(%) Cg + mS100A9p
mS100A9p
(d)
Mye
lop
erox
idas
eac
tivi
ty (
U/m
g)
600
500
400
300
200
100
0Cg Cg
+mS100A9p
mS100A9p Vehicle
**
*
(h) 400
300
200
100
0Formalin
+mS100A9p
Formalin
Mye
lop
erox
idas
eac
tivi
ty (
U/m
g)
mS100A9p Vehicle
**
(e)
Vehicle20
15
10
5
00 30 60 90 120 150 180 210 240
***
*
Formalin + mS100A9pFormalin
Time (min)
Wit
hd
raw
al la
ten
cy (
s)
(f) Formalin + mS100A9p
70
60
50
40
30
20
10
00 0 0
10 m
in
10 m
in
10 m
in
120 m
in
120 m
in
120 m
in
240 m
in
240 m
in
240 m
in
180 m
in
180 m
in
180 m
in
60 m
in
60 m
in
60 m
in
30 m
in
30 m
in
30 m
in
20 m
in
20 m
in
20 m
in
Formalin
No
cice
pti
ve s
core
(%
)
**
*
**
*
** *
*
*
(g)
VehicleFormalin + mS100A9pFormalin
0
50
100
150
200
0 30 60 90 120 120 180 210 240
Time (min)
Δ P
aw v
olu
me
(%)
Figure 1 Mice were administered intraplantar (i.pl) with carrageenan (Cg; 2%) or formalin (5%) concomitantly to vehicle, or to 1 lg of
mS100A9p, or receive i.pl. injections of vehicle alone (vehicle) or mS100A9 alone. Withdrawal latency to a thermal stimulation (thermal
hyperalgesia) (a, e) nociceptive score in response to von Frey filament exposure (mechanical hyperalgesia and allodynia), (b, f) paw volume
(edema), (c, g) and myeloperoxidase activity (d, h) were evaluated hourly after i.pl. injections. Data represent mean ± SEM of 6–8 animals per
group. (*) denotes statistical differences compared to the control groups (Cg or Formalin) (P < 0.05, one-way ANOVA followed by Tukey’s
multiple comparison post-test).
S100A9 analgesic properties 431
ª 2009 The Authors Journal compilation ª 2009 Societe Francaise de Pharmacologie et de TherapeutiqueFundamental & Clinical Pharmacology 23 (2009) 427–438
together, these results demonstrate that mS100A9p
inhibits calcium signaling in primary afferents, but by
a mechanism independent of intracellular calcium
store depletion and independent of calcium ion
chelation.
mS100A9p inhibits voltage-gated calcium
channel activity
We investigated the possibility that mS100A9 could act
as an inhibitor of calcium channels. Voltage-gated
calcium channels (VGCC) are present on sensory neu-
rons, where L-, and N-channels have been shown to be
involved in nociception [30,31]. We investigated the
effects of mS100A9 on calcium signaling in HEK cells
(tsA-201), which were transfected or not, with N-type or
L-type calcium channels. Different concentrations of
mS100A9p (0.5, 1, 5 or 50 lM) inhibited, in a dose-
dependent manner, the magnitude of calcium flux in
response to KCl (50 mM) in N-type transfected cells
(Figure 4a), but had no effect in non-transfected cells
(not shown). No effect of mS100A9p was observed on
L-type transfected cells, in response to KCl stimulation
(Figure 4b).
To confirm the inhibition by mS100A9p of KCl-
mediated calcium influx observed in native DRG neurons
and heterologous expression systems, we proceeded with
electrophysiological recordings on both DRG neurons
and transfected tsA-201 cells. DRG neurons express both
low voltage activated (LVA) and HVA calcium currents.
However, by using a holding potential of )60 mV and a
test pulse of 0, +10 mV, LVA channels could be
selectively eliminated. As shown in Figure 5a, the isolated
(a) Cg + vehicle
Cg + mS100A9p
Cg + NX + mS100A9p
* * * *
* * * *
10
8
6
4
2
0 0 1 2 3 4
Time (h)
Wit
hd
raw
al la
ten
cy (
s)
Cg + vehicle
Cg + mS100A9p
Cg + NX + mS100A9p
0 1 2 3 4 Time (h)
Δ P
aw v
olu
me
(%)
0
20
40
60
80
100
120 (c)
* *
Cg 0
200
400
Mye
lop
erox
idas
e ac
tivi
ty (
U/m
g) 600
800
+ + + +
+
–
– +
+ +
–
–
NX
S100
(d)
Cg100
80
60
40
20
No
cice
pti
ve s
core
(%
)
0 0 1 h 2 h 3 h 4 h 0 1 h 2 h 3 h 4 h 0 1 h 2 h 3 h 4 h
3.61 filament 3.84 filament 4.08 filament
Cg + mS100A9p
Cg + NLX + mS100A9p
* * *
*
* * * *
*
(b)
Figure 2 Mice were injected subcutaneously with naloxone (NX; 10 mg/kg) or its vehicle 20 mins before the intraplantar (i.pl) injection of
Carragenan (Cg; 2%), or before the concomitant treatment with Cg and mS100A9p. Withdrawal latency in response to a thermal stimulation
(thermal hyperalgesia) (a) nociceptive score in response to von Frey filament exposure (mechanical hyperalgesia and allodynia) (b) paw
volume (edema) (c) and myeloperoxidase activity (d) were evaluated after different times after i.pl. injections. Data represent mean ± SEM of
6–8 animals per group. (*) denotes statistical differences compared to the group injected with Cg + vehicle (P < 0.05, one-way ANOVA
followed by Tukey’s multiple comparison post-test).
432 C.S. Dale et al.
ª 2009 The Authors Journal compilation ª 2009 Societe Francaise de Pharmacologie et de TherapeutiqueFundamental & Clinical Pharmacology 23 (2009) 427–438

HVA currents were inhibited by �35% upon application
of 50 lM mS100A9 peptide, and a maximal inhibition
was observed �5 mins. after the beginning of the
response. The two major populations of HVA channels
in native DRG neurons are L-type and N-type, with
N-type channels pre-dominating [32,33]. After applica-
tion of x-conotoxin-GVIA, the selective N-type channels
blocker, effect of mS100A9p was significantly reduced
(Figure 5b, c). These results suggest that mS100A9-
(a) 1.5
1.4
1.3
1.2
1.1
1.00.5 5 50
mS100A9p (µM)
KCI (50 mM)
**
* **
**
Cal
ciu
m f
lux
inn
euro
ns
1.5
1.4
1.3
1.2
1.1
1.0
0.9
* *
* ** #
Cal
ciu
m f
lux
inn
euro
ns
KCIThapsig
mS100A9p
+ + + +
++
+ + ––
– –
(b)
2.5
2.0
1.5
1.05 50 100
mS100A9p (µM)Calcium ionophore (2 µM)
Cal
ciu
m f
lux
inn
euro
ns
(c)
Figure 3 Neurons were exposed to KCl (50 mM) concomitantly to
mS100A9p (0.5, 5, 50 or 100 lM – panel) (a) neurons treated only
with KCl were considered as control group. In panel (b), neurons
were incubated 5 mins with thapsigagrin (250 nM) and after were
exposed to KCl (50 mM) concomitantly to mS100A9p (50 lM).
Neurons treated only with KCl (control group) or KCl and
thapsigargin (Thapsg) were also evaluated. Data represent
mean ± SEM of 15–20 neurons per group. (*) denotes statistical
differences compared to KCl (P < 0.05), (***) denotes statistical
differences compared to KCl (P < 0.001), (#) denotes statistical
differences compared to KCl + thapsigargin (P < 0.05; one-way
ANOVA followed by Tukey’s multiple comparison post-test). In panel
(c), neurons were exposed to a calcium ionophore (2 lM) concom-
itantly to mS100A9p (0.5 5, 50 or 100 lM). Neurons treated only
with calcium ionophore were considered as the control group. The
calcium flux measurements were performed by fluorescence at
460–490 nm excitations and 515 nm emissions in individual cells
using a wide-field fluorescence microscope. A kinetic of 30 pictures
in 90 s was performed.
Figure 4 N-type transfected tsA-201 (a) or L-type transfected
tsA-201 (b) cells were exposed to KCl (50 mM) concomitantly to
mS100A9p (0.1, 0.5, 1, 5 or 50 lM). Data represent mean ± SEM
of 15–20 cells per group. (*) denotes statistical differences compared
to KCl (P < 0.05). The calcium flux measurements were performed
by fluorescence at 460–490 nm excitations and 515 nm emissions
in individual cells using a wide-field fluorescence microscope.
A kinetic of 30 pictures in 90 s was performed.
S100A9 analgesic properties 433
ª 2009 The Authors Journal compilation ª 2009 Societe Francaise de Pharmacologie et de TherapeutiqueFundamental & Clinical Pharmacology 23 (2009) 427–438

peptide blocks N-type but not other HVA calcium
channels (L-type, P/Q-type) in DRG neurons. Then, we
recorded from tsA-201 cells transiently expressing either
L-type or N-type channels. N-type (Cav2.2+ b1b + a2- d1)
but not L-type (Cav1.2+ b1b + a2- d1) activity was
inhibited by �40% with 25 lM peptide (Figure 5d), and
this inhibition could be partially reversed upon washout,
as seen from the raw current traces (Figure 5d), and the
representative time course of mS100A9 effect (Figure 5e).
These data are consistent with the results observed with
Ca2+measurements from DRG neurons and tsA-201
cells. Superimposition of normalized current-voltage
relations in the absence and presence of the peptide
reveals that the voltage-dependence of activation was not
affected (Figure 5f). Taken together, our results demon-
strate that mS100A9p inhibits native and expressed
N-type calcium currents and this block does not affect
channel activation.
D I S C U S S I O N
Although different in vivo studies have reported anti-
nociceptive properties for the calcium-binding protein
S100A9 or its C-terminus domain [6–8,11,12], the
mechanism of this effect has never been identified. As
murine S100A9 C-terminal peptide (mS100A9p) also
showed anti-inflammatory properties, it is uncertain
whether the analgesic effects of mS100A9p are due to
Figure 5 Representative time course of calcium currents from dorsal root ganglia (DRG) neurons recorded during a + 10 mV test pulse
(50 ms) from a holding potential of )60 mV before and after application of 50 lM mS100A9-peptide (a) representative time course of calcium
current using the same protocol as in (a) but in the presence of x-conotoxin-GVIA (1 lM) (b) histogram of voltage gated calcium channels
block after application of 50 lM peptide in the absence or presence of x-conotoxin-GVIA (1 lM) (I/Icontrol = 0.67 ± 0.04, n = 9 and
I/Icontrol = 0.83 ± 0.05, n = 5 respectively. *P < 0.05) (c) mS100A9-peptide inhibits N-type current in tsA-201 cells transfected with the
Cav2.2 subunit (+b1b, a2- d1 – panel d). Note that the inhibition is partially reversible. Inset: histogram summarizing block of N-type but
not L-type channels expressed transciently in tsA-201 cells (I/Icontrol = 0.59 ± 0.03 for Cav2.2 and I/Icontrol = 0.94 ± 0.02 for Cav1.2).
Numbers in parentheses are the numbers of cells analyzed. Error bars reflect SEM. Asterisks denote statistical significance (***P < 0.001).
Time course of the block of N-type (Cav2.2 + b1b + a2- d1) current amplitude by 25 lM mS100A9p in tsA-201 cells (e). Current amplitude
was elicited by a + 10 mV test pulse (150 ms) from a holding potential of )90 mV. Normalized I-V curve from tsA-201 cell transfected
with Cav2.2 N-type channel, in the absence or presence of 25 lM mS100A9p (f) no shift in the I-V relation was observed.
434 C.S. Dale et al.
ª 2009 The Authors Journal compilation ª 2009 Societe Francaise de Pharmacologie et de TherapeutiqueFundamental & Clinical Pharmacology 23 (2009) 427–438

a reduced inflammatory response and thereby to a
reduced activation of primary afferents by inflammatory
mediators, or whether mS100A9p exerts antinociceptive
properties per se. One of the first aims of this study was to
identify whether the anti-nociceptive properties of
mS100A9p depend on its anti-inflammatory properties.
We used two different inflammatory models, which both
induce all inflammatory parameters (edema, granulocyte
recruitment, vasodilation, hyperalgesia, allodynia), but
with different profiles of mediators. The major mediators
involved in carrageenan-induced inflammation are eico-
sanoids and sympathomimetic amines, whereas the
major inflammatory mediators involved in formalin-
induced inflammatory pain are serotonin and histamine,
which directly activate sensitized nociceptors [34].
Despite the distinct mechanisms involved in the inflam-
matory pain induced by carrageenan and that induced
by formalin, mS100A9p was effective in both models, to
decrease inflammatory hyperalgesia and allodynia.
Although anti-inflammatory properties have been
described for S100A9, here in both models, the inflam-
matory edema was not reduced by mS100A9p. Edema
formation plays an important part in inducing hyper-
algesia, as compression of peripheral nerves causes
nociceptor activation [35]. The fact that mS100A9p
did not interfere with edema suggests that the mecha-
nism by which mS100A9p exerts antinociceptive prop-
erties could be a direct effect on nociceptive pathways,
and not an anti-inflammatory effect. This is further
suggested by the fact that in the formalin model, where
primary afferents are activated within seconds, and pain
appears in the first minutes after the i pl. injection when
this effect is not yet dependent on inflammatory infil-
trates, mS100A9p was effective at reducing mechanical
hyperalgesia as early as 10 mins after the i pl. injection
(Figure 1f). Finally, the dichotomy between antinocicep-
tive and anti-inflammatory properties of mS100A9p was
demonstrated by the effects of mS100A9p on granulo-
cyte infiltration. In the carrageenan model, mS100A9p
was able to inhibit MPO activity, as previously reported,
but in the formalin model, mS100A9p treatment
increased MPO activity. This last result definitively
demonstrates that the analgesic properties of mS100A9p
observed in both models are independent of anti-inflam-
matory (inhibition of granulocyte infiltration) properties
of mS100A9p. It is interesting to note that mS100A9p
showed better anti-inflammatory properties in the car-
rageenan model than in the formalin model. This could
be explained by the different mediators and pathways
involved in those two models.
As one of the major pathway of antinociception
involves the activation of opioid receptors and the
release of endogenous opioid receptor agonists, we
investigated the possibility that mS100A9p could acti-
vate opioid receptor pathways. This hypothesis was
supported by the fact that immune cells, which have
been shown to be targeted by mS100A9p [36] are a
major source of endogenous opioid agonists (endor-
phins). Treatment with naloxone, a non-selective mu-,
delta- and kappa-opioid receptor antagonist failed to
inhibit the antinociceptive effects of mS100A9p in the
carrageenan model, thereby demonstrating that activa-
tion of opioid receptor pathway was not involved in this
mechanism.
We have previously demonstrated that mS100A9p
can act directly on sensory neuron activation by
inhibiting calcium signaling on DRG neurons in response
to the activation of different receptors such as PAR2, B1
bradykinin receptor or Transient potential vanilloid
receptor 1 – TRPV1 [10]. Here, the dichotomy between
antinociceptive and anti-inflammatory properties of
mS100A9p suggests also that mS100A9p could act
directly on primary afferents. We observed here that
mS100A9p inhibits calcium flux on DRG neurons
induced by KCl. This demonstrates that the inhibitory
effects of mS100A9p on primary afferent signaling might
not be specific of some receptors such as PAR2 or BK
receptors, but that mS100A9 is able to interfere with
calcium signaling in general, in DRG neurons. We
further showed that this effect was not the result of
inhibition of intracellular calcium stores, as thapsigargin
treatment had no effect on mS100A9p inhibition of KCl-
induced calcium signal. The inhibitory effect of
mS100A9p was not due either to a chelating effect on
calcium ions, as mS100A9p was not able to inhibit the
effects of a calcium ionophore.
Calcium ions are widely recognized to play a funda-
mental role in the regulation of several biological
processes, such as modulation of neurotransmitter
release and modulation of cell membrane excitability
[30,37]. Evidence has accumulated for the involvement
of calcium ions also in nociception and anti-nociception
[30]. Particularly, the presence and functionality of L-,
N- and P/Q-type of VGCC were demonstrated in the
dorsal horn of the spinal cord [38] and to be involved
in nociceptive transmission. As in sensory neurons,
mS100A9p inhibited calcium flux induced by different
receptors [12] as well as by KCl (Figure 3); this suggests
that mS100A9p could act directly on calcium channels
present on primary afferents. Thus, we investigated
S100A9 analgesic properties 435
ª 2009 The Authors Journal compilation ª 2009 Societe Francaise de Pharmacologie et de TherapeutiqueFundamental & Clinical Pharmacology 23 (2009) 427–438

whether mS100A9p could directly interfere with the
activation of L- or N-type voltage-gated calcium chan-
nels. Our results demonstrate that mS100A9p inhibits
N-type calcium channels expressed in tsA-201 cells,
whereas no effect was observed on L-type channels.
These data were confirmed by electrophysiology in
isolated sensory neurons, in which we observed that
S100A9-C-terminal peptide could specifically block
N-type calcium channels, while mS100A9p effect was
almost abolished in the presence of N-type channel
blocker x-conotoxin-GVIA. These results indicate that
S100A9-C-terminal peptide is able to inhibit native and
expressed N-type calcium currents without affecting
channel activation. The mechanism by which S100A9
inhibits N-type calcium channel will have to be further
investigated. One possibility is that parameters of
the gating of the channel are changed by S100A9.
We know that the current-voltage relation is not
modified by mS100A9p, but if the peptide blocks
the pore of the channel, the open probability or
conductance of the channel could be affected. In
addition, recent studies have shown that reducing
agents or endogenous compounds with metal chelator
properties (particularly Zn2+ions) are able to regulate
calcium channels [39]. Similar mechanism could be
hypothesized for S100A9.
Voltage gated calcium channels as a family repre-
sent one of the most important regulators of Ca2+
concentration in neurons, as they do in many cell
types, and hence have an important role in neuronal
functions, including nociception and chronic/neuro-
pathic pain [40]. Particularly, the N-type VGCCs have
been demonstrated to be involved in short lasting
nociception, hyperexcitability, inflammation and neuro-
pathic pain [38,41–44]. Our results demonstrating the
inhibitory effects of mS100A9p on N-type VGCCs are
consistent with such a role and suggest for mS100A9p
an important role as therapeutic tool for the control
of nociception. Furthermore, our results reinforce the
concept that S100A9 protein is an important actor
of the modulation of nociceptive responses. Whether
native S100A9 plays such an endogenous role as a pain
modulator via N-type VGCC inhibition still has to be
demonstrated. However, these studies depend on the
availability of more specific tools for the study of S100A9
(knock-out mice, blocking antibodies, large supplies of
pure S100A9 native protein).
In conclusion, our study demonstrates, for the first
time the mechanisms involved in the antinociception
induced by the C-terminus of murine S100A9. Our
findings highlight mS100A9p as a new important tool
for the inhibition of N-type VGCC, and for the inhibition
of inflammatory pain.
A C K N O W L E D G E M E N T S
This work was supported by Coordenacao de Aperfeicoa-
mento de Pessoal de Nıvel Superior-CAPES, the Canadian
Institute for Health Research, and the Institut UPSA de la
douleur. CSD holds a Canadian Association of Gastroen-
terology – CAG/SHERING fellowship. NV is supported by
an AVENIR-INSERM program, by the ‘Fondation Betten-
court-Schueller’ and the ‘Fondation Schlumberger’, and
was an Alberta Heritage Foundation for Medical
Research (AHFMR) scholar, and a Canadian Institute
for Health Research Investigator. GWZ is a Canada
Research Chair and AHFMR Scientist. CA was supported
by an AHFMR Fellowship Award.
We thank Dr. Pina Colarusso from the Department of
Physiology & Biophysics of the University of Calgary for
the use of the fluorescence microscope.
R E F E R E N C E S
1 Hessian P.A., Edgeworth J., Hogg N. MRP-8 and MRP-14, two
abundant Ca(2+)-binding proteins of neutrophils and mono-
cytes. J. Leukoc. Biol. (1993) 53 197–204.
2 Roth J., Vogl T., Sunderkotter C., Sorg C. Chemotactic activity of
S100A8 and S100A9. J. Immunol. (2003) 171 5651.
3 Raftery M.J., Harrison C.A., Alewood P., Jones A., Geczy C.L.
Isolation of the murine S100 protein MRP14 (14 kDa migra-
tion-inhibitory-factor-related protein) from activated spleen
cells: characterization of post-translational modifications and
zinc binding. Biochem. J. (1996) 316 285–293.
4 Kerkhoff C., Klempt M., Kaever V., Sorg C. The two calcium-
binding proteins, S100A8 and S100A9, are involved in the
metabolism of arachidonic acid in human neutrophils. J. Biol.
Chem. (1999) 274 32672–32679.
5 Kerkhoff C., Klempt M., Sorg C. Novel insights into structure
and function of MRP8 (S100A8) and MRP14 (S100A9).
Biochim. Biophys. Acta. (1998) 1448 200–211.
6 Giorgi R., Pagano R.L., Dias M.A., Aguiar-Passeti T., Sorg C.,
Mariano M. Antinociceptive effect of the calcium-binding
protein MRP-14 and the role played by neutrophils on the
control of inflammatory pain. J. Leukoc. Biol. (1998) 64 214–
220.
7 Pagano R.L., Dias M.A., Dale C.S., Giorgi R. Neutrophils and the
calcium-binding protein MRP-14 mediate carrageenan-induced
antinociception in mice. Mediators Inflamm. (2002) 11 203–
210.
8 Pagano R.L., Mariano M., Giorgi R. Neutrophilic cell-free
exudate induces antinociception mediate by the protein
S100A9. Mediators Inflamm. (2006) 2006, 36765.
436 C.S. Dale et al.
ª 2009 The Authors Journal compilation ª 2009 Societe Francaise de Pharmacologie et de TherapeutiqueFundamental & Clinical Pharmacology 23 (2009) 427–438

9 Pagano R.L., Sampaio S.C., Juliano L., Juliano M.A., Giorgi R.
The C-terminus of murine S100A9 inhibits spreading and
phagocytic activity of adherent peritoneal cells. Inflamm. Res.
(2005) 54 204–210.
10 Dale C.S., Pagano R.L., Paccola C.C. et al. Effect of the
C-terminus of murine S100A9 protein on experimental
nociception. Peptides (2006) 27 2794–2802.
11 Dale C.S., Goncalves L.R.C., Juliano L., Juliano M.A., da Silva
A.M., Giorgi R. The C-terminus of murine S100A9 inhibits
hyperalgesia and edema induced by jararhagin. Peptides (2004)
25 81–89.
12 Dale C.S., Cenac N., Britto L.R. et al. The C-terminus of murine
S100A9 protein inhibits hyperalgesia induced by the agonist
peptide of protease-activated receptor 2 (PAR2). Br. J. Phar-
macol. (2006) 149 374–384.
13 Loomans H.J., Hahn B.L., Li Q.Q., Phadnis S.H., Sohnle P.G.
Histidine-based zinc-binding sequences and the anti-
microbial activity of calprotectin. J. Infect. Dis. (1998)
177 812–814.
14 Sopalla C., Leukert N., Sorg C., Kerkhoff C. Evidence for the
involvement of the unique C-tail of S100A9 in the binding of
arachidonic acid to the heterocomplex S100A8/A9. Biol. Chem.
(2002) 383 1895–1905.
15 Watt K.W., Brightman I.L., Goetzl E.J. Isolation of two
polypeptides comprising the neutrophil-immobilizing factor of
human leucocytes. Immunology (1983) 48 79–86.
16 Freemont P., Hogg N., Edgeworth J. Sequence identity. Nature
(1989) 339 516.
17 Hessian P.A., Wilkinson L., Hogg N. The S100 family protein
MRP-14 (S100A9) has homology with the contact domain of
high molecular weight kininogen. FEBS Lett. (1995) 371 271–
275.
18 Juni A., Klein G., Kest B. Morphine hyperalgesia in mice is
unrelated to opioid activity, analgesia, or tolerance: evidence for
multiple diverse hyperalgesic systems. Brain Res. (2006) 1070
35–44.
19 Hargreaves K., Dubner R., Brown F., Flores C., Joris J. A new
and sensitive method for measuring thermal nociception in
cutaneous hyperalgesia. Pain (1988) 32 77–88.
20 Takasaki I., Andoh T., Nojima H., Shiraki K., Kuraishi Y.
Gabapentin antinociception in mice with acute herpetic pain
induced by herpes simplex virus infection. J. Pharmacol. Exp.
Ther. (2001) 296 270–275.
21 Zimmermann M. Ethical guidelines for investigations of exper-
imental pain in conscious animals. Pain (1983) 6 109–110.
22 Bradley P.P., Priebat D.A., Christensen R.D., Rothstein G.
Measurement of cutaneous inflammation: estimation of neu-
trophil content with an enzyme marker. J. Invest. Dermatol.
(1982) 78 206–209.
23 Vergnolle N., Hollenberg M.D., Sharkey K.A., Wallace J.L.
Characterization of the inflammatory response to proteinase-
activated receptor-2 (PAR2)-activating peptides in the rat paw.
Br. J. Pharmacol. (1999) 127 1083–1090.
24 Asfaha S., Cenac N., Houle S. et al. Protease-activated receptor-
4: a novel mechanism of inflammatory pain modulation. Br. J.
Pharmacol. (2007) 150 176–185.
25 Cenac N., Andrews C., Holzhausen M. et al. Proteases are
released in Irritable Bowel Syndrome: a role in visceral pain.
J. Clin. Invest. (2007) 117, 636–647.
26 Beedle A.M., McRory J.E., Poirot O. et al. Agonist-independent
modulation of N-type calcium channels by ORL1 receptors. Nat.
Neurosci. (2004) 7 118–125.
27 Vanderah T.W. Pathophysiology of pain. Med. Clin. North Am.
(2007) 91 1–12.
28 Thastrup O., Cullen P.J., Drobak B.K., Hanley M.R., Dawson
A.P. Thapsigargin, a tumor promoter, discharges intracellular
Ca2+ stores by specific inhibition of the endoplasmic reticulum
Ca2(+)-ATPase. Proc. Natl Acad. Sci. USA (1990) 87 2466–
2470.
29 Luckasen J.R., White J.G., Kersey J.H. Mitogenic properties of a
calcium ionophore, A23187. Proc. Natl. Acad. Sci. USA (1974)
71 5088–5090.
30 Prado W.A. Involvement of calcium in pain and antinocicep-
tion. Braz. J. Med. Biol. Res. (2001) 34 449–461.
31 Lee Y., Lee C.H., Oh U. Painful channels in sensory neurons.
Mol. Cells. (2005) 20 315–324.
32 Sutton K.G., Martin D.J., Pinnock R.D., Lee K., Scott R.H.
Gabapentin inhibits high-threshold calcium channel currents in
cultured rat dorsal root ganglion neurons. Br. J. Pharmacol.
(2002) 135 257–265.
33 Altier C., Khosravani H., Evans R.M. et al. ORL1 receptor-
mediated internalization of N-type calcium channels. Nat.
Neurosci. (2006) 9 31–40.
34 Parada C.A., Tambeli C.H., Cunha F.Q., Ferreira S.H. The major
role of peripheral release of histamine and 5-hydroxytryptamine
in formalin-induced nociception. Neuroscience (2001) 102
937–944.
35 Dray A., Bevan S. Inflammation and hyperalgesia: highlighting
the team effort. Trends Pharmacol. Sci. (1993) 14 287–290.
36 Striz I., Trebichavsky I. Calprotectin – a pleiotropic molecule in
acute and chronic inflammation. Physiol. Res. (2004) 53 245–
253.
37 Zamponi G.W., Snutch T.P. Modulation of voltage-dependent
calcium channels by G proteins. Curr. Opin. Neurobiol. (1998)
8 351–356.
38 Vanegas H., Schaible H. Effects of antagonists to high-threshold
calcium channels upon spinal mechanisms of pain, hyperalge-
sia and allodynia. Pain (2000) 85 9–18.
39 Nelson M.T., Joksovic P.M., Su P. et al. Molecular mechanisms
of subtype-specific inhibition of neuronal T-type calcium
channels by ascorbate. J. Neurosci. (2007) 27 12577–
12583.
40 Gribkoff V.K. The role of voltage-gated calcium channels in
pain and nociception. Semin. Cell Dev. Biol. (2006) 17 555–
564.
41 Maggi C.A., Tramontana M., Cecconi R., Santicioli P. Neuro-
chemical evidence for the involvement of N-type calcium chan-
nels in transmitter secretion from peripheral endings of sensory
nerves in guinea pigs. Neurosci. Lett. (1990) 114 203–206.
42 Santicioli P., Del Bianco E., Tramontana M., Geppetti P., Maggi
C.A. Release of calcitonin gene-related peptide like-immuno-
reactivity induced by electrical field stimulation from rat spinal
S100A9 analgesic properties 437
ª 2009 The Authors Journal compilation ª 2009 Societe Francaise de Pharmacologie et de TherapeutiqueFundamental & Clinical Pharmacology 23 (2009) 427–438

afferents is mediated by conotoxin-sensitive calcium channels.
Neurosci. Lett. (1992) 136 161–164.
43 Smith M.T., Cabot P.J., Ross F.B., Robertson A.D., Lewis R.J. The
novel N-type calcium channel blocker, AM336, produces potent
dose-dependent antinociception after intrathecal dosing in rats
and inhibits substance P release in rat spinal cord slices. Pain
(2002) 96 119–127.
44 Altier C., Dale C.S., Kisilevsky A.E. et al. Differential role of N-
type calcium channel splice isoforms in pain. J. Neurosci.
(2007) 27 6363–6373.
438 C.S. Dale et al.
ª 2009 The Authors Journal compilation ª 2009 Societe Francaise de Pharmacologie et de TherapeutiqueFundamental & Clinical Pharmacology 23 (2009) 427–438




















