
Percutaneous Transluminal Aortic Angioplasty: Techniques and Results

Adrenomedullin Inhibits Angiotensin II-Induced Oxidative Stress
Via Csk-Mediated Inhibition of Src Activity
Jing Liu1, Tatsuo Shimosawa2, Hiromitsu Matsui1, Fanyin Meng3, Scott C. Supowit3, Donald J.
DiPette3, Katsuyuki Ando1, Toshiro Fujita1
1Department of Endocrinology and Nephrologyዊ�2Department of Clinical Laboratory Medicine,
Faculty of Medicine, The University of Tokyo, Tokyo, Japan. 3Texas A&M University System
Health Science Center College of Medicine and Scott & White Health System, Department of
Medicine, Temple, TX.
Running title: Adrenomedullin inhibits oxidative stress
Keywords: adrenomedullin, reactive oxygen species, antioxidant, Src, lucigenin
Abstract: 168 wordsTotal: 6151 wordsFigures: 5
All correspondence to Toshiro Fujita, MD, PhD
Department of Internal Medicine, Faculty of Medicine, The University of Tokyo,7-3-1 Hongo,
Bunkyo-ku, Tokyo, 113-8655, Japan
Phone: 81-3-5800-9735
Fax: 81-3-5800-9736
E-mail: [email protected]
Page 1 of 30
Copyright Information
Articles in PresS. Am J Physiol Heart Circ Physiol (October 27, 2006). doi:10.1152/ajpheart.00486.2006
Copyright © 2006 by the American Physiological Society.

1
Abstract
We have demonstrated that adrenomedullin (AM) protects against angiotensin II (Ang II)-
induced cardiovascular damage through the attenuation of increased oxidative stress observed
in AM deficient miceዊ�However, the mechanism(s) that underlie this activity remains unclear.
To address this question we investigated the effect of AM on Ang II-stimulated reactive oxygen
species (ROS) production in cultured rat aortic vascular smooth muscle cells (VSMCs)ዊ�Ang II
markedly increased ROS production through activation of NADPH oxidase. This effect was
significantly attenuated by AM in a concentration-dependent manner. This effect was mimicked
by dibutyl-cAMP and blocked by pretreatment with H-89, a protein kinase A inhibitor, and
CGRP 8-37, an AM /CGRP receptor antagonist. This inhibitory effect of AM was also lost
following expression of a constitutively active Srcዊ�Moreover, AM intersected Ang II signaling
by inducing Csk activation, that in turn, inhibits Src activationዊ� These data, for the first time,
demonstrate that AM attenuates the Ang II-induced increase in ROS in VSMCs via activation of
Csk, thereby inhibiting Src activity.
Page 2 of 30
Copyright Information

2
Introduction
Enhanced production of reactive oxygen species (ROS) and activation of redox-dependent
signaling cascades by vasoactive agents are critical processes underlying a number of
cardiovascular and metabolic diseases (37,43). Virtually all types of vascular cells produce ROS,
such as O2- and H2O2ዊ�The major source of oxygen intermediates in the vascular wall is non-
phagocytic NADPH oxidase (6,11,20,22,34). Accumulating evidence indicates that NADPH
oxidase-derived ROS are essential participants in signaling events in vascular cells (7,28).
Angiotensin II (Ang II) is an important mediator of vascular smooth muscle cell (VSMC)
function, and a number of the effects of Ang II are mediated through the generation of ROS
(4,9,42)ዊ�In particular, Ang II-induced hypertrophy is inhibited by over expression of catalase,
which hydrolyzes hydrogen peroxide (H2O2), or by inhibition of vascular NADPH oxidases
(3,8,42).
Adrenomedullin (AM), a potent vasodilator peptide that acts primarily as a protective
autocrine/paracrine factor in the cardiovascular system, has antiproliferative and antimigrative
effects (32). We previously reported that AM gene transfer prevents the development of cuff-
induced vascular injury (31). AM deficient heterozygous knockout mice also exhibited
significant damage in multiple organ systems that was mediated by enhanced oxidative stress in
models of Ang II/salt-induced hypertension (30), aging (44), hypoxia (21) and cuff injury (15).
In addition, AM expression was stimulated by ROS in vascular endothelial cells (24). These
data are consistent with the hypothesis that AM is an endogenous anti-oxidant that plays a
protective role against increased oxidative stress. However, the underlying cellular
mechanism(s) and identification of the critical AM regulated signal transduction pathways
remained to be clarifiedዊ�
Page 3 of 30
Copyright Information

3
Several signaling molecules such as phospholipase D (PLD), protein kinase C (PKC),
phospholipase A2, and phosphatidylinositol-3-kinase (PI3K) are implicated in Ang II-evoked
NADPH oxidase activation (10,38,39,40). Whether these pathways function in parallel or in
series to stimulate this enzyme is unclearዊ�However, multiple lines of evidence strongly
indicate that Src, which influences all of these signaling molecules, is a common upstream
mediator (29)ዊ�Src kinases are a family of nonreceptor tyrosine kinases, of which the prototype,
c-Src, is the major isoform in the vasculature (23,35). Src can be switched from an inactive to
active state through control of phosphorylation at two major sites, Tyr416 and Tyr527.
Phosphorylation of Tyr416 activates Src while phosphorylation of Tyr527 inactivates this
tyrosine kinase. C-terminal Src kinase (Csk) is a key enzyme that phosphorylates Src at
Tyr527(19). In VSMCs, c-Src is rapidly activated by Ang II and plays a key role in growth
signaling (12). Furthermore, it has been shown that PKA intersects Src signaling in mammalian
cells (1). Since cAMP-PKA is the primary post-receptor signal transduction pathway of AM and
PKA upregulates Csk activity through phosphorylation, c-Src inactivation via Csk is a likely
candidate through which AM intersects Ang II-stimulated ROS signalingዊ�
Therefore, in the present study we examined whether AM directly inhibits intracellular ROS
generation stimulated by Ang II in VSMCs, and assessed the role(s) that Src and Csk play in the
regulation of the cellular signaling pathway(s) responsible for its anti-oxidant effectዊ�
Materials and Methods
Materials
Rat AM and human CGRP (8-37) were purchased from the Peptide Institute (Osaka,
Japan). Fetal bovine serum (FBS) from Trace Scientific LTD (Melbourne, Australia)ዊ�
Anti-Src and anti-phosphotyrosine mAb (4G10) antibody, a dominant-negative
Page 4 of 30
Copyright Information

4
mutant of Src (DN-Src), a constitutively active form of avian Src (CA-Src), and Csk-siRNA kit
came from Upstate Biotechnology (Lake Placid, NY). Lipofectin was from Invitrogen (Palo
Alto, CA), Mirus reagent was from Mirusbio (Madison, WI). Anti-Src phosphor-specific
antibody (Tyr416) was from Biosource international (Camarillo, CA), and anti-Src phosphor-
specific antibody (Tyr527) was purchased from Cell Signaling Technology (Beverly, MA)ዊ�
Anti-Csk antibody and protein A/G PLUS-Agarose were obtained from Santa Cruz
Biotechnology (Santa Cruz, CA)ዊ�Dulbecco’s modified Eagles medium (DMEM), Ang II,
rotenone, allopurinol, N-(2-[p-Bromocinnamylamino]ethyl)-5-isoquinolinesulfonamide
hydrochloride (H-89) and dibutal-cAMP, TNF-ዊ� and calcium ionophore A23187 were from
Sigma (St. Louis, MO)ዊ�
Cell Culture
Rat aortic VSMCs were prepared from the thoracic aorta of 8-week-old male Sprague-
Dawley rats using the explant method (33) and cultured in DMEM containing 10% FBS at 37˚C
in a humidified atmosphere of 95% air-5% CO2ዊ�Sub cultured cells (passages 3-12) were
deprived of serum for 48 h prior to experimental use.
Transient Transfection
Either a dominant negative mutant of Src, or a constitutive-active mutant of Src was over
expressed in VSMCs by transient transfection. All Src constructs were in the pUSEamp(+)
expression vector. Transient transfection was performed with 2 mg of total DNA per 35 mm
dish and Lipofectin according to the manufacturer’s instructions (DNA:LipofectAMINE ratio,
1:3). The transfection efficiency was confirmed by immunoblotting for total protein. Pilot
studies have documented a transfection efficiency of 15% to 25% in VSMCs using this
approach. Cells transfected with an expression vector containing the same promoter, but without
Page 5 of 30
Copyright Information

5
the subcloned transgene cassette (empty expression vector), served as a control. All Csk
siRNAs were synthesized and annealed at Dharmacon. VSMCs were plated 2 days before
transfections and grown to 60–80% confluence in 12-well plates. SMARTpool™ Csk or control
siRNA (200nM) were transfected into VSMCs with Mirus reagent (Mirusbio) according to the
manufacturer’s protocol.
Measurement of Intracellular ROS Levels
We measured ROS both in living cells and in total VSMC homogenates. ROS production in
living cells was measured by the lucigenin method (11) with minor modifications. Briefly, cells
were pretreated with or without test compounds for the indicated time periods, then stimulated
with or without Ang II (100 nmol/L) for 10 min and AM for 5 minዊ�In some experiments,
CGRP (8-37) or H-89 was added 30 min before, dibutyl-cAMP was included 1 hour before,
TNF-ዊ� 2 hours before and the calcium ionophore A23187 3 hours before AM treatmentዊ�The
cells were then trypsinized, collected by centrifugation, and the pellet washed in modified Krebs
buffer as described elsewhere (11). After washing, the cells were resuspended in Krebs buffer
with 1 mg/ml BSA, and the cell concentration was adjusted to 1x106 in 900 µl buffer. To
measure ROS production, the cell suspension was placed in a luminometer (LB 9507; Berthold,
Wildblad, Germany). Measurements were started by an injection of 100 µl lucigenin (final
concentration 5 x 10–4 M). Photon emission was counted every 1 min for up to 10 min. Modified
Krebs buffer was used as control (blank). For total cell homogenate, cells were treated the same
as in living cells and were scraped into ice-cold Hank's balanced salt solution supplemented with
MgCl 2 (0.8 mmol/L) and CaCl 2 (1.8 mmol/L)ዊ�Cells were disrupted by rapid freezing in
liquid nitrogen followed by sonicationዊ�O2- production was measured in the presence of
NADPH (100 µmol/L) for 10 min and was expressed as mean arbitrary light units per minዊ�
Page 6 of 30
Copyright Information

6
Western Blot Analysis for Phosphorylated Src
Western blot analysis was performed as described previously (17). Briefly, 30 µg of the cell
lysates were subject to 10% SDS-PAGE and proteins were transferred to nitrocellulose
membranes (Genetics, Tokyo, Japan)ዊ�The membranes were blocked for 1 hour at room
temperature with 5% skimmed milkዊ�The blots were then incubated overnight at 4˚C with anti-
Src and anti-Src phosphospecific antibodies, followed by incubation for 1 hour with a secondary
peroxide-conjugated antibody (Amersham Biosciences UK), the proteins were detected by ECL
Western blotting detection kit (Amersham Biosciences UK)ዊ�In figures showing Western blots,
the blots shown are representative of at least three independent experiments.
Immunoprecipitation and Immunoblot Analysis of Csk
Cells were lysed and centrifuged as described elsewhere (36). The supernatant was
mixed with the antibody to Csk (2 µg) and rocked at 4 °C for 2–16 hours, and then protein A/G-
Agarose was added for an additional 2 hours to overnightዊ�Immunoprecipitates were washed,
fractioned by electrophoresis, and transferred to nitrocellulose membranes (Genetics, Tokyo,
Japan)ዊ�After blocking with 5% milk, the membrane was treated with an anti-phosphotyrosine
(4G10) antibody conjugated with horseradish peroxidase (Upstate Biotechnology)ዊ�
Immunoreactive proteins were detected by enhanced chemiluminescence (Amersham Pharmacia
Biotech)ዊ�
Statistical analysis
Data were expressed as mean ± SEMዊ�Differences between groups were examined for
statistical significance using either ANOVA followed by the Dunnett’s test or unpaired t-test,
where they are appropriateዊ�P values less than 0.05 were considered statistically significantዊ�
Results
Page 7 of 30
Copyright Information

7
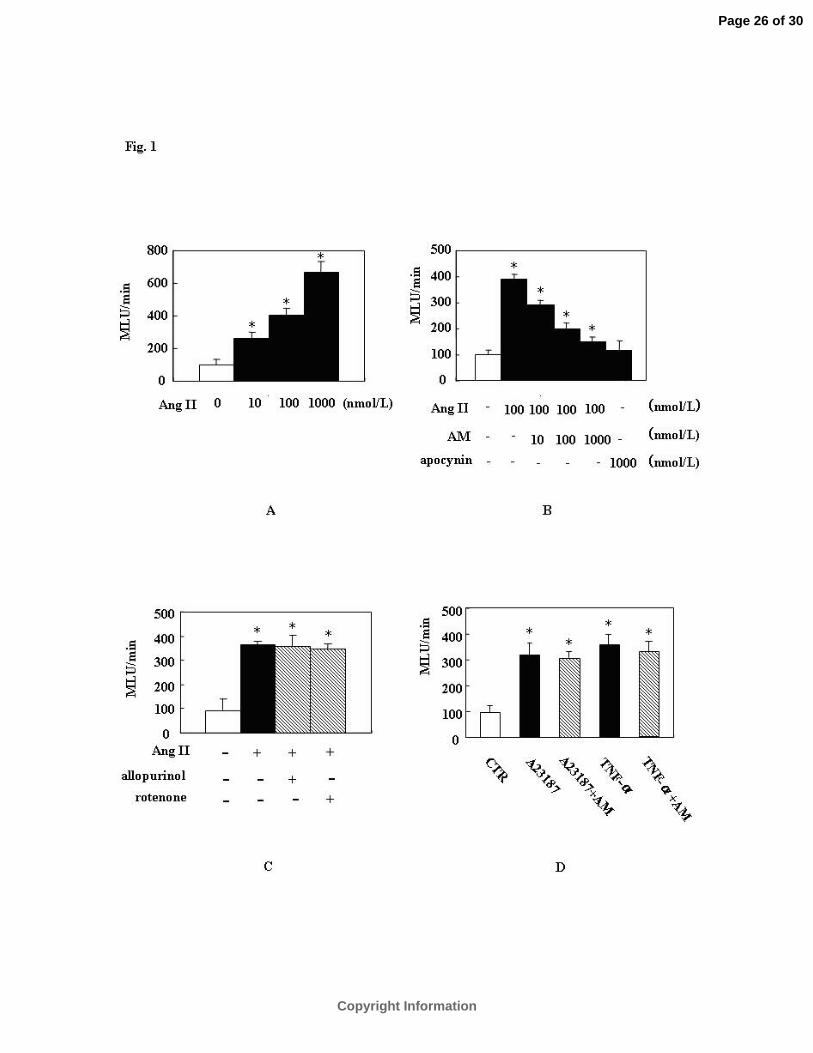
AM Attenuates Intracellular ROS Production Stimulated by Ang II.
A 10 minute treatment of VSMCs with Ang II evoked a significant increase (approximately
3- to 7-fold from the lowest to the highest concentration of Ang II) in ROS production (Fig. 1A)
which was attenuated by AM in a concentration-dependent manner (10-8-10-6 mol/L, Fig. 1B).
The highest concentration of AM (10-6 mol/L) inhibited ROS generation to almost the same
degree as the NADPH specific oxidase inhibitor, apocynin (Fig.1B), and neither inhibition of
mitochondrial electron transport by rotenone (10 µmol/L) nor inhibition of xanthine oxidase by
allopurinol (10 µmol/L) significantly altered Ang II-induced ROS production (Fig. 1C).
Likewise, AM treatment had no effect on the Ca2+ ionophore A23187 or TNF-ዊ�-induced ROS
production (Fig. 1D).
AM Inhibits Intracellular ROS Generation via an AM Receptor-Mediated and cAMP-
PKA-Dependent Pathway.
To determine whether the anti-oxidant effect of AM is mediated via the AM
receptor/cAMP-PKA-dependent pathway, we tested the effects of an AM receptor antagonist
and cAMP-related compounds both in living cells (Fig. 2A) and cell homogenates (Fig. 2B)ዊ�
Pretreatment with 3x10-6 mol/L CGRP (8-37), an AM/CGRP receptor antagonist (2), completely
abolished the AM-induced inhibition of intracellular ROS generation stimulated by Ang IIዊ�
Since the cAMP-PKA pathway is the most well documented post-receptor signal transduction
pathway of AM (5, 46), we examined the involvement of this pathway on the inhibitory
mechanism of AM on ROS generationዊ�Dibutyl-cAMP (10-3 mol/L) significantly inhibited
ROS generation by Ang II to a similar extent as AM (10-7mol/L)ዊ�In contrast, the inhibitory
effect of AM on ROS generation was completely abolished by H-89 (10-5 mol/L), a PKA
inhibitorዊ�Treatment with AM, CGRP (8-37), dibutyl-cAMP, or H-89 alone did not alter
Page 8 of 30
Copyright Information

8
intracellular ROS levels in intact cells (Fig. 2A)ዊ�Similar results were obtained with cell
homogenates (Fig. 2B).
AM Inhibits Phosphorylation of Src (Tyr416).
It has been reported that c-Src regulates Ang II-mediated NADPH oxidase-derived O2- in rat
VSMCs (29). Based on these data, experiments were performed to determine whether AM
inhibits Ang II-induced ROS production specifically through inhibition of Src (Tyr416)
phosphorylation. As shown in figure 3A, phosphorylation of Src (Tyr416) was significantly
increased in VSMCs by Ang II (10-7 mol/L) stimulation, that was, in turn, inhibited by AM in a
concentration dependent manner (10-8-10-6 mol/L). Figure 3B shows the effects of dibutyl-
cAMP (10-3 mol/L), CGRP (8-37) and H-89 (10-5 mol/L) treatment on phosphorylation. Dibutyl-
cAMP significantly (P<0.05) inhibited the phosphorylation of Src (Tyr416) by Ang II which
was comparable to that of AM (10-7 mol/L). As expected, H-89 and CGRP (8-37) completely
abolished the inhibitory effect of AM on Src (tyr416) phosphorylation. In the absence of Ang II
stimulation, treatment with AM, CGRP(8-37), dibutyl-cAMP, or H-89 alone did not
significantly change the state of Src phosphorylation.
Effects of Dominant Negative and Constituitively-Active Src on the Anti-Oxidant Activity
of AM
To confirm that the attenuation of Src activity is directly attributable to the inhibitory effect
of AM on Ang II-induced ROS generation, we undertook experiments to transfect VSMCs with
dominant negative or constituitively-active Src constructs. The transfection efficiency was
confirmed by immunoblotting for total Src protein. Figure 4A shows that both of the mutant Src
constructs were effectively over expressed in VSMCs. Then using the live transfected cells, we
assessed the effect of AM on Ang II-induced ROS production by the lucigenin
Page 9 of 30
Copyright Information

9
chemiluminescence method. As seen in figure 4B, the dominant-negative Src transfected cells
that serve as the negative controls for this set of experiments, display basal level of ROS
generation that were unchanged following treatment with Ang II alone or Ang II and AM. In
contrast in constituitively-active Src over expressing cells, ROS generation was significantly
enhanced and was not increased further by Ang II treatment (Fig. 4C). Consistent with these
results, AM treatment failed to inhibit the elevated ROS generation (Fig. 4C).
AM and Ang II Enhanced Csk Phosphorylation Increases Src (Tyr527) Phosphorylation
and the Inhibitory effect of AM on Src activation is lost in Csk Knockdown VSMCs.
Inactivation of Src kinase involves phosphorylation of Tyr527 by Csk (19). Therefore, we
examined whether AM inhibition of Ang II-induced Src activation was associated with changes
in Csk activityዊ�Csk immunoprecipitates were probed with an anti-phosphotyrosine antibody .
Phosphorylation of Csk was significantly increased approximately 2-fold by Ang II but not by
treatment with AM alone. A much more robust increase (approximately 4-fold was observed
when the VSCMs were treated with the combinations of Ang II and either AM (10-7 mol/L) or
dibutyl-cAMP (10-3 mol/L, Fig. 5A). In contrast, treatment of the cells with Ang II did not
increase the phosphorylation of Src (Tyr527) as shown in Fig. 5C. Treatment with AM produced
an increase Src (Tyr527) phosphorylation, but it did not achieve statistical significance (data not
shown). However, the combination of Ang II and AM evoked a significant increase in Src
(Tyr527) phophorylation, again in the absence of any changes in total Src protein ( Fig. 5C). In
addition, Csk knockdown studies using siRNA also reversed the inhibitory effects of AM on
Src activation (Fig. 5C).
Page 10 of 30
Copyright Information

10
Discussion
Accumulating evidence strongly supports the hypothesis that AM possesses significant
protective properties against end organ damage, produced by a number of pathophysiological
conditions, through inhibition of oxidative stress (15,21,30,44). The mechanisms underlying this
activity vary depending on cell types and experimental conditions. In mesangial cells, AM
suppresses ROS production via the cAMP-PKA pathway (5) while in the rat ventricle increased
oxidative stress caused by ischemia/reperfusion injury can be attenuated by AM-mediated
inhibition of NADPH oxidase via the nitric oxide-cGMP signaling pathway (14). In contrast,
enhanced hypoxia-induced generation of ROS in human alveolar epithelial cells is attenuated
by a marked AM stimulated increase in the potent ROS scavenger glutathione (16).
In light of the aforementioned reports the significant findings of this study are: 1) Using rat
VSMCs in conjunction with a lucigenin chemiluminescence methodology, we have confirmed
that NADPH oxidase is the source of the marked increase in ROS following treatment with Ang
II and that the inhibitory effects of AM in this setting are mediated through the cAMP/PKA
signal transduction pathway (46). 2) The data presented herein clearly support the hypothesis
that in VSMCs, Src is the primary upstream signaling molecule that mediates the Ang II
increase in NADPH generated ROS (29). 3) We have shown, for the first time, that the
attenuation of Src activity in Ang II treated VSMCs is regulated by the phosphorylation and
subsequent activation of Csk that is dependent on AM via the cAMP/PKA signaling pathway
and may also require Ang II that could be playing an autoregulatory role by initiating a
potential negative feedback loop.
Before initiating the series of experiments on the roles that Src and Csk play in AM mediated
attenuation of Ang II evoked ROS production it was necessary to demonstrate that Ang II
treatment produced, in a concentration dependent manner, a rapid and robust enhancement in
Page 11 of 30
Copyright Information

11
VSMC ROS production that was significantly inhibited to near basal levels by AM, again in a
concentration dependent fashion. The results presented herein are consistent with those reported
by Yoshimoto et. al. in both rat aortic VSMCs (46) and endothelial cells (47). Intracellular ROS
generation stimulated by Ang II in these cell types has been shown to be mainly derived from
the activation of NADPH oxidase (11,22). This is clearly supported by our findings that
inhibitors of xanthine oxidase (allopurinol) and mitochondrial-derived ROS (rotenone) have no
significant effect on Ang II-stimulated ROS production. In contrast, treatment of the VSMCs
with apocynin, a specific NADPH oxidase inhibitor, reduced Ang II stimulated ROS generation
to levels observed in the untreated control cells which were similar to those observed following
AM treatment. However, the apocynin was somewhat more potent than AM for reasons that
remain to be clarified. Furthermore, AM was without effect on Ca2+ ionophore A23187 or
TNF-ዊ�-induced ROS production indicating that its ability to attenuate Ang II-stimulated ROS
production in cultured VSMCs is regulated by specific signaling pathway(s) that inhibit,
primarily, the activity of NADPH oxidase rather than acting as an general antioxidant.
Although the cAMP-PKA pathway is the primary signal transduction pathway stimulated by
AM, this peptide can also trigger other signaling cascades including NF-kB (27), tyrosine
kinase (13), protein phosphatase 2A (26), and PI3K/Akt (25,45) depending on the cell type or
experimental conditions. To confirm that the anti-oxidant effects of AM observed in this study
were in fact mediated primarily by the AM receptor mediated up regulation of the cAMP-PKA
signal transduction pathway, we demonstrated that the inhibitory effect of AM on Ang II
stimulated ROS generation was completely abolished by pretreatment of the VSMCs with
CGRP (8-37), an AM receptor antagonist. As expected, dibutyl-cAMP significantly inhibited
the Ang II-evoked increase in ROS generation to a similar extent as AM. Likewise, the AM
Page 12 of 30
Copyright Information

12
mediated inhibition of Ang II-evoked ROS production was abolished by H-89, a PKA inhibitor.
These data, therefore, provide additional evidence that AM inhibits Ang II stimulated NADPH
oxidase-dependent ROS production through the AM receptor mediated activation of the cAMP-
PKA pathwayዊ�
A central role for Src in the regulation of the redox status of Ang II treated VSCMs was
indicated by the observation that phosphorylation of Src (Tyr416), a key step in the activation
of this protein kinase, was markedly increased following 10 minutes of treatment with Ang II.
The link between the Ang II AT1 receptor and Src, however, is not clear. It has been reported
that in VSCMs, Ang II signaling is regulated, in part, by the transactivation of the epidermal
growth factor receptor, which serves as a scaffolding for preactivated c-Src and downstream
adaptors. Indeed, interactions between Gβγ subunits, their associated kinases, and kinase
substrates may form the signaling complex that binds c-Src (18,48). This phosphorylation of
Src was significantly attenuated in a concentration dependent manner by AM in the absence of
any change in total Src protein. Moreover this attenuation of Src phosphorylation by AM was
consistent with the concentration-dependent inhibitory effect of AM on Ang II enhanced ROS
generation. Likewise, the degree of Src Tyr416 phosphorylation, after normalization to total Src
protein, following treatment with agents that activated or blocked the AM receptor/cAMP-PKA
pathway, was in line with the ability of these agents to inhibit Ang II-evoked ROS production.
Indeed, Ang II was unable to stimulate ROS production above baseline levels in dominant
negative Src transfected VSCMs and the inhibitory effect of AM on Ang II-induced ROS
generation was lost in constitutively-active Src transfected cells, indicating that c-Src is the
target molecule through which AM inhibits ROS generation induced by Ang II in VSMCs.
As described previously, c-Src is activated by autophosphorylation of Tyr416 and inactivated
Page 13 of 30
Copyright Information

13
by Csk which phosphorylates Tyr527 (1,19). Indeed we observed that the knockdown of Csk
by si RNA blocked the phosphorylation of Src527 thereby indicating a direct relation between
Csk and Src inactivation.. It was, however, surprising that treatment of the cells with Ang II
under the same conditions that enhance ROS generation, produced a significant increase in Csk
phosphorylation, one mechanism by which this tyrosine kinase is activated, while AM alone did
not. However, addition of Ang II in combination with either AM or dibutyl cAMP resulted in a
marked increase in Csk phosphorylation without a change in total Csk protein. In contrast, when
Src Tyr527 phosphorylation was assessed, neither Ang II nor AM alone was able to significantly
stimulate posphorylation at this site whereas the combination of the two did result in a
statistically significant upregulation of Try527 phosphorylation.
Although we do not have any direct evidence to support this hypothesis, it is tempting to
speculate that in order to maintain a balance between active and inactive Src, which clearly
participates in modulation of the redox status of this cell type, Ang II treatment simultaneously
activates Src and also sets the stage for the downregulation of this protein by phosphorylation
of Csk. This change in the activation status of Csk such that it cannot deactivate Src, via Tyr527
phosphorylation, except in the presence of a negative regulatory agent like AM that is able to
trigger Csk up to a level of activity that is sufficient to down regulate Src, thereby reestablishing
homeostasis of the redox state of the cell. Indeed, a report from Touyz et al (41) provides
indirect evidence for this argument. In this study they demonstrated that Ang II stimulated
VSMC growth and remodeling was mediated by the activation of c-Src and that this effect was
significantly greater in VSMCs from spontaneously hypertensive rats (SHR) compared to those
from normotensive Wistar-Kyoto (WKY) rats. Ang II mediated C-Src phosporylation (Tyr527)
was approximately 4-fold greater in SHR than WKY. However, Ang II increased Csk
phosphorylation 2-to 3-fold in WKY but not in SHR. From these data it was concluded that c-
Page 14 of 30
Copyright Information

14
Src phosphorylation and Src dependent cell growth following Ang II treatment is increased in
VSMCs from SHR, and that these processes are associated with blunted Ang II-induced
phosphorylation of Csk. Since the VSMCs used in our study came from normal rats our
tentative conclusion regarding the mechanism underlying the Csk mediated inhibition of Src
activity and ROS production are consistent with the data reported by Touyz et al (41). We are,
however, well aware that multiple mechanisms for Src and Csk regulation exist, and it still
unclear as to the exact contribution of the different regulatory mechanisms in a given
physiological setting (36).
Page 15 of 30
Copyright Information

15
References
1. Abrahamsen H, Vang T, Tasken Kዊ�Protein kinase A intersects Src signaling in membrane
microdomainsዊ�J Biol Chemዊ� 278:17170-17177, 2003ዊ�
2. Bellibas SE, Guidobono F, Bettica P, Netti C, Pecile Aዊ�Effects of pyridoxine
neurotoxicity on a distribution of calcitonin gene-related peptide binding sitesዊ�Pol J
Pharmacolዊ� 49:37-42, 1997ዊ�
3. Berk BC, Corson MAዊ�Angiotensin II signal transduction in vascular smooth
muscle: role of tyrosine kinasesዊ�Circ Resዊ� 80:607-616, 1997ዊ�
4. Chen XL, Tummala PE, Olbrych MT, Alexander RW, Medford RM. Angiotensin II
induces monocyte chemoattractant protein-1 gene expression in rat vascular smooth muscle cells.
Circ Res. 83:952-9, 1998.
5. Chini EN, Chini CC, Bolliger Cዊ�Cytoprotective effects of adrenomedullin in glomerular
cell injury: central role of cAMP signaling pathwayዊ�Kidney Intዊ� 52:917-925, 1997ዊ�
6. De Keulenaer GW, Alexander RW, Ushio-Fukai M, Ishizaka N, Griendling KKዊ�Tumor
necrosis factor alpha activates a p22phox-based NADH oxidase in vascular smooth muscleዊ�
Biochem Jዊ� 329:653-657, 1998ዊ�
7. Droge Wዊ�Free radicals in the physiological control of cell functionዊ�Physiol. Revዊ� 82:47-
95, 2002ዊ�
8. Eguchi S, Inagami Tዊ�Signal transduction of angiotensin II type 1 receptor
through receptor tyrosine kinaseዊ�Regul Peptዊ� 91:13-20, 2000ዊ�
9. Frank GD, Eguchi S, Inagami T, Motley ED. N-acetylcysteine inhibits
angiotensin II-mediated activation of extracellular signal-regulated kinase and epidermal
Page 16 of 30
Copyright Information

16
growth factor receptor. Biochem Biophys Res Commun 280:1116-9, 2001.
10. Griendling KK, Sorescu D, Lassègue B, Ushio-Fukai Mዊ�Modulation of protein kinase
activity and gene expression by reactive oxygen species and their role in vascular physiology
and pathophysiologyዊ�Arterioscler Thromb Vasc Biolዊ� 20:2175-2183, 2000ዊ�
11. Griendling KK, Minieri CA, Ollerenshaw JD, Alexander RWዊ�Angiotensin II
stimulates NADH and NADPH oxidase activity in cultured vascular smooth muscle cellsዊ�Circ
Resዊ� 74:1141-1148, 1994ዊ�
12. Ishida M, Ishida T, Thomas S, Berk B. Activation of extracellular signal regulated
kinases (ERK1/2) by Ang II is dependent on c-Src in vascular smooth muscle cells. Circ Res.
82:7-12, 1998.
13. Iwasaki H, Shichiri M, Marumo F, Hirata Y. Adrenomedullin stimulates proline-rich
tyrosine kinase 2 in vascular smooth muscle cells. Endocrinology. 142:564-572, 2001
14. Kato K, Yin H, Agata J, Yoshida H, Chao L, Chao Jዊ�Adrenomedullin gene
delivery attenuates myocardial infarction and apoptosis after ischemia and reperfusion. Am J
Physiol. 285:H1506-H1514, 2003.
15. Kawai J, Ando K, Tojo A, Shimosawa T, Takahashi K, Onozato ML, Yamasaki M,
Ogita T, Nakaoka T, Fujita Tዊ�Endogenous adrenomedullin protects against vascular
response to injury in miceዊ�Circulationዊ� 109:1147-1153, 2004ዊ�
16. Kim JY, Yim JH, Cho JH, Kim JH, Ko JH, Kim SM, Park SJ, and Park JH.
Adrenomedullin Regulates Cellular Glutathione Content via Modulation of γ-Glutamate-
cysteine Ligase Catalytic Subunit Expression. Endocrinology. 147(3):1357-64, 2006
17. Kyaw M, Yoshizumi M, Tsuchiya K, Kagami S, Izawa Y, Fujita Y, Ali, N, Kanematsu
Page 17 of 30
Copyright Information

17
Y, Toida K, Ishimura K, Tamaki Tዊ�Src and Cas are essentially but differentially involved in
angiotensin II-stimulated migration of vascular smooth muscle cells via extracellular signal-
regulated kinase 1/2 and c-Jun NH2-terminal kinase activationዊ�Mol Pharmacolዊ� 65:832-841,
2004ዊ�
18. Luttrell LM, Ferguson SSG, Daaka Y, Miller WE, Maudsley S, Della Rocca GJ, Lin F-
T, Kawakatsu H, Owada K, Luttrell DK. β-Arrestin-dependent formation of β2 adrenergic
receptor-Src protein kinase complexes. Science. 283:655–661, 1999.
19. Martin GSዊ�The hunting of the Srcዊ�Nat Rev Mol Cell Biolዊ� 2:467-475, 2001ዊ�
20. Matsubara T, Ziff Mዊ�Increased superoxide anion release from human endothelial cells in
response to cytokinesዊ�J Immunolዊ� 137:3295-3298, 1986ዊ�
21. Matsui H, Shimosawa T, Itakura K, Xing GQ, Ando K, Fujita Tዊ�Adrenomedullin can
protect against pulmonary vascular remodeling induced by hypoxiaዊ�Circulationዊ� 109:2246-
2251, 2004ዊ�
22. Mohazzab KM, Kaminski PM, Wolin MSዊ�NADH oxidoreductase is a major source of
superoxide anion in bovine coronary artery endotheliumዊ�Am J Physiolዊ� 266:H2568-2572,
1994ዊ�
23. Oda Y, Renaux B, Bjorge J, Saifeddine M, Fujita DJ, Hollenberg MDዊ�c-Src is a major
cytosolic tyrosine kinase in vascular tissueዊ�Can J Physiol Pharmacolዊ� 77:606-617, 1999ዊ�
24. Ogita T, Hashimoto E, Yamasaki Mዊ�Hypoxic induction of adrenomedullin in cultured
human umbilical vein endothelial cellsዊ�J Hypertensዊ� 19:603-608, 2001ዊ�
25. Okumura H, Nagaya N, Itoh T, Okano I, Hino J, Mori K, Tsukamoto Y, Ishibashi-
Ueda H, Miwa S, Tambara K, Toyokuni S, Yutani C, Kangawa K. Adrenomedullin
Page 18 of 30
Copyright Information

18
infusion attenuates myocardial ischemia/reperfusion injury through the phosphatidylinositol 3-
kinase/Akt-dependent pathway. Circulation. 109:242-248, 2004
26. Parameswaran N, Nambi P, Hall CS, Brooks DP, Spielman WS. Adrenomedullin
decreases extracellular signal-regulated kinase activity through an increase in protein
phosphatase-2A activity in mesangial cells. Eur J Pharmacol. 388:133-138, 2000
27. Pleguezuelos O, Hagi-Pavli E, Crowther G, Kapas S. Adrenomedullin signals through
NF-kappaB in epithelial cells. FEBS Lett. 577:249-254, 2004
28. Sauer H, Wartenberg M, Heschler Jዊ�Reactive oxygen species as intracellular
messengers during cell growth and differentiationዊ�Cell Physiol Biochemዊ� 11:173-186, 2001ዊ�
29. Seshiah PN, Weber DS, Rocic P, Valppu L, Taniyama Y, Griendling KKዊ�Angiotensin
II stimulation of NAD(P)H oxidase activity: upstream mediatorsዊ�Circ Resዊ� 91:406-413, 2002ዊ�
30. Shimosawa T, Shibagaki Y, Ishibashi K, Kitamura K, Kangawa K, Kato S, Ando K,
Fujita Tዊ�Adrenomedullin, an endogenous peptide, counteracts cardiovascular damageዊ�
Circulationዊ� 105:106-111, 2002ዊ�
31. Shindo T, Kurihara H, Maemura K, Kurihara Y, Kuwaki T, Izumida T, Minamino Nዊ�
Hypotension and resistance to lipopolysaccharide-induced shock in transgenic mice
overexpressing adrenomedullin in their vasculatureዊ�Circulationዊ� 101:2309-2316, 2000ዊ�
32. Sugo S, Minamino N, Shoji H, Kangawa K, Matsuo Hዊ�Effects of vasoactive substances
and cAMP related compounds on adrenomedullin production in cultured vascular smooth
muscle cellsዊ�FEBS Lettዊ� 369:311-314, 1995ዊ�
33. Sun Ji, Marx SO, Chen HJ, Poon M, Marks AR, Rabbani LE. Role for p27Kip1 in
Vascular Smooth Muscle Cell Migration. Circulation. 103:2967-2972, 2001.
Page 19 of 30
Copyright Information

19
34. Sundaresan M, Yu ZX, Ferrans VJ, Irani K, Finkel Tዊ�Requirement for generation of
H2O2 for platelet-derived growth factor signal transductionዊ�Scienceዊ� 270:296-299, 1995ዊ�
35. Thomas SM, Brugge JSዊ�Cellular functions regulated by Src family kinasesዊ�Annu Rev
Cell Dev Biolዊ� 13:513-609, 1997ዊ�
36. Torgersen KM, Vang T, Abrahamsen H, Yaqub S, Hor˘ejsˇı´V, Schraven B, Rolstad B,
Mustelin T, and Taske´n K. Release from Tonic Inhibition of T Cell Activation through
Transient Displacement of C-terminal Src Kinase (Csk) from Lipid Rafts. J Biol Chem. 276:
29313–29318, 2001.
37. Touyz RMዊ�Oxidative stress and vascular damage in hypertensionዊ�Curr. Hypertens. Repዊ�
2:98-105, 2000ዊ�
38. Touyz RM, Chen X, Tabet F, Yao G, He G, Quinn MT, Pagano PJ, Schiffrin ELዊ�
Expression of a functionally active gp91phox-containing neutrophil-type NAD(P)H oxidase in
smooth muscle cells from human resistance arteries: regulation by angiotensin IIዊ�Circ Resዊ�
90:1205-1213, 2002ዊ�
39. Touyz RM, Schiffrin ELዊ�Ang II-stimulated generation of reactive oxygen species in
human vascular smooth muscle cells is mediated via PLD dependent pathwaysዊ�Hypertensionዊ�
34:976-982, 1999ዊ�
40. Touyz RM, Schiffrin ELዊ�Increased generation of superoxide by angiotensin II in smooth
muscle cells from resistance arteries of hypertensive patients role of phospholipase D-dependent,
NAD(P)H oxidase-sensitive pathwaysዊ�J Hypertensዊ� 19:1245-1254, 2001ዊ�
41. Touyz RM, Wu XH, He G, Salomon S, Schiffrin EL. Increased Angiotensin II–Mediated
Src Signaling via Epidermal Growth Factor Receptor Transactivation Is Associated With
Page 20 of 30
Copyright Information

20
Decreased C-Terminal Src Kinase Activity in Vascular Smooth Muscle Cells From
Spontaneously Hypertensive Rats. Hypertension. 39[part 2]:479-485, 2002.
42. Ushio-Fukai M, Zafari AM, Fukui T, Ishizaka N, Griendling KKዊ�p22phox is a critical
component of the superoxide-generating NADH/NADPH oxidase system and regulates
angiotensin II-induced hypertrophy in vascular smooth muscle cellsዊ�J Biol Chemዊ�
271:23317-23321, 1996ዊ�
43. Wilcox CSዊ�Reactive oxygen species: Role in blood pressure and kidney functionዊ�Curr.
Hypertens. Repዊ� 4:160-166, 2002ዊ�
44. Xing GQ, Shimosawa T, Ogihara T, Matsui H, Itakura K, Xu QY, Asano T, Ando K,
Fujita Tዊ�Angiotensin II-induced insulin resistance is enhanced in adrenomedullin deficient
miceዊ�Endocrinologyዊ� 145:3647-3651, 2004ዊ�
45. Yin H, Chao L, Chao J. Adrenomedullin protects against myocardial apoptosis after
ischemia/reperfusion through activation of Akt-GSK signaling. Hypertension. 43:109-116,
2004
46. Yoshimoto T, Fukai N, Hirata Yዊ�Anti-oxidant effect of adrenomedullin on angiotensin
II-induced ROS generation in vascular smooth muscle cellsዊ�Endocrinologyዊ� 145:3331-3337,
2004
47. Yoshimoto T, Gochou N, Fukai N, Sugiyama T, Shichiri M, and Hirata Y.
Adrenomedullin Inhibits Angiotensin II-Induced Oxidative Stress and Gene Expression in Rat
Endothelial Cells. Hypertens Res. 28: 165-172, 2005
48. Zou YZ, Komuro I, Yamazaki T, Kudoh S, Aikawa R, Zhu WD, Shiojima I, Hiroi Y,
Tobe K, Kadowaki T, Yazaki Y. Cell type-specific angiotensin II-evoked signal transduction
Page 21 of 30
Copyright Information

21
pathways: critical roles of Gβγ subunit, Src family, and Ras in cardiac fibroblasts. Circ Res.
82:337–345, 1998.
Page 22 of 30
Copyright Information

22
Figure legends
Figure 1. AM inhibits intracellular ROS generation stimulated by Ang II in VSMCsዊ�
ROS generation was detected by lucigenin chemiluminescence methodዊ�(A) Concentration
dependent response of ROS generation by Ang IIዊ�VSMCs were stimulated with various
concentrations of Ang II (10-8-10-6 mol/L ) for10 minዊ�(B) AM inhibits intracellular ROS
generation stimulated by Ang II in a concentration-dependent manner(10-8-10-6 mol/L )ዊ�Cells
pretreated with AM (10-8 -10-6 mol/L) for 5 min or apocynin (10-6 mol/L) for 2 hours were
stimulated with Ang II (10-7 mol/L ) for 10 minዊ�(C) Treatment of VSMCs with mitochondrial-
derived ROS inhibitor rotenone (10 µmol/L) and the xanthine oxidase inhibitor allopurinol (10
µmol/L) for 1 hourዊ�(D) Treatment with calcium ionophore A23187 (1µmol/L) for 3 hours or
TNF-ዊ� (10 ng/mL) for 2 hours. ROS generation is expressed as fold increase compared with
controlዊ�The values are representative of three independent experiments, each performed in
triplicate. MLU (mean light units and CTR (control). *P<0.05 vs. control; †P<0.05 vs. Ang II.
Figure 2. AM inhibits intracellular ROS generation via a receptor-mediated and
cAMP-PKA-dependent pathwayዊ�ROS production was examined by lucigenin
chemiluminescence both in living cells (A) and cell homogenates (B). VSMCs treated with or
without AM (10-7 mol/L), dibutyl-cAMP (10-3 mol/L), H-89 (10-5 mol/L) or CGRP(8-37) (3x10-5
mol/L) for 0.5-1 h were stimulated in the absence or presence of Ang II (10-7 mol/L) for10 minዊ�
In some experiments, CGRP(8-37) or H-89 was applied 30 min prior to the AM pretreatmentዊ�
Quantification of ROS levels by the lucigenin chemiluminescence method, and the calculation
and plotting of data are the same as in Fig. 1ዊ�The values are representative of three
independent experiments, each performed in triplicate. *P<0.05 vs. control; †P<0.05 vs. Ang II.
Page 23 of 30
Copyright Information

23
Figure 3. AM inhibits Src (Tyr416) phosphorylation stimulated by Ang II via a receptor-
mediated and cAMP-PKA-dependent pathway in VSMCsዊ�(A) Concentration dependent
response of Src (Tyr416) phosphorylation by Ang IIዊ�VSMCs were pretreated with various
concentrations of AM (10-8 -10-6 mol/L) for 5 min, then stimulated with Ang II (10-7 mol/L ) for
10 minዊ�The top panel is a representative immunoblot demonstrating phosphorylation of Src
(Tyr416)ዊ�The bottom panel represents averaged data from three independent experiments
quantified by densitometry of immunoblots and expressed as fold increases in phosphorylation
compared with unstimulated cellsዊ�(B) Effect of dibutyl-cAMP, CGRP(8-37) and H-89 on
phosphorylation of Src (Tyr416)ዊ�VSMCs treated with or without AM (10-7 mol/L), dibutyl-
cAMP (10-3 mol/L), H-89 (10-5 mol/L) or CGRP(8-37) (3x10-5 mol/L) for 0.5-1 h were
stimulated with or without Ang II (10-7 mol/L) for 10 minዊ�In some experiments, CGRP(8-37)
or H-89 were applied 30 min prior to the AM pretreatmentዊ�Phosphorylation of Src (Tyr416)
was evaluated by Western blottingዊ�The top panel is a representative immunoblot of the
phosphorylation of Src (Tyr416)ዊ�The bottom panel represents averaged data from three
independent experiments quantified by densitometry of immunoblots and expressed as fold
increases compared to unstimulated cellsዊ�*P<0.05 vs. control; †P<0.05 vs. Ang II
Figure 4. Effects of AM on Ang II-induced ROS production in dominant negative or
constitutively-active Src transfected cellsዊ�(A) The transfection efficiency for these experiments
was confirmed by immunoblotting for total Src protein. ROS production was examined by
lucigenin chemiluminescence method both in dominant-negative Src (B) and constitutive-active
Src transfected living cells (C). Controls are DN-Src and CA-Src transfected cell in the absence
Page 24 of 30
Copyright Information

24
of stimulation. The values are representative of three independent experiments, each performed
in triplicate.
Figure 5. AM enhances Ang II-induced Csk and in turn Src (Tyr527) phosphorylation, the
inhibitory effect of AM on Src activation is lost in Csk knockdown VSMCs. VSMCs were
pretreated with AM (10-7 mol/L) for 5 min or dibutyl-cAMP (10-3 mol/L) for 1 hour, followed by
addition of Ang II (10-7 mol/L ) for 10 minዊ�(A) Csk was immunoprecipitated from cell lysates
with anti-Csk antibodyዊ�Immunoprecipitated proteins were subjected to SDS-PAGE and
immunoblotted with antiphosphotyrosine antibodyዊ�The top panel is a representative
immunoblot from three independent experiments assessing phosphorylated Cskዊ�The bottom
panel represents averaged data from three independent experiments quantified by densitometry
of immunoblots and expressed as fold increases in phosphorylation compared to unstimulated
cells. *P<0.05 vs control; †P<0.01 Ang II + AM and Ang II + dibutyl cAMP vs control. IP
indicates immunoprecipitation and IB, immunoblottingዊ�(B) VSMCs were transfected with
SMARTpool™ Csk or control siRNA and incubated for 48 h. The expression of Csk protein
was selectively inhibited by siRNA (top panel) without reducing β-actin control protein (bottom
panel). Each experiment was performed three times in triplicate. (C) Immunoblots and
quantification of total Src and phosphorylated Tyr416 and Tyr527 with and without Csk siRNA.
Each panel represents averaged data from three independent experiments quantified by
densitometry of immunoblots. *P<0.05 vs control.
Page 25 of 30
Copyright Information
Page 26 of 30
Copyright Information

Fig. 2A
B
Page 27 of 30
Copyright Information
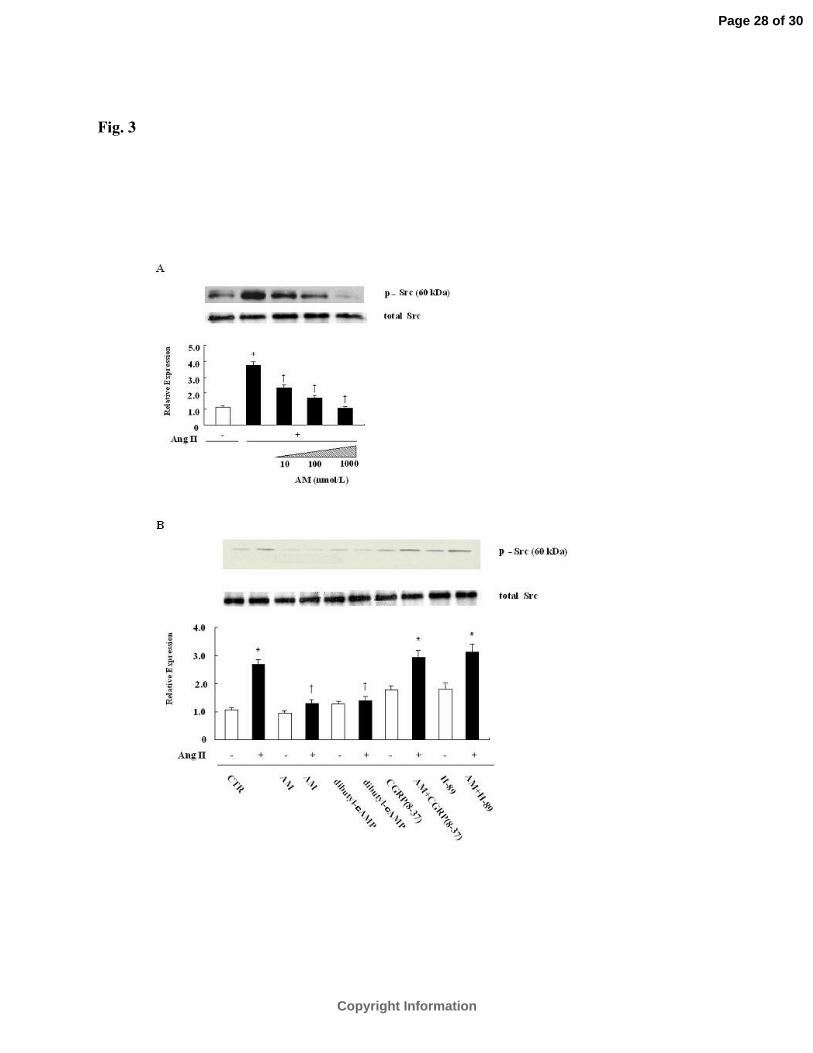
Fig. 3
Page 28 of 30
Copyright Information

Fig. 4A
B C
Page 29 of 30
Copyright Information

Fig. 5A
Fig. 5B
Fig. 5C
Page 30 of 30
Copyright Information
Copyright © 2022 FDOKUMEN




















