Administration of serotonin inhibitor p-Chlorophenylalanine induces pessimistic-like judgement bias...
10
Administration of serotonin inhibitor p-Chlorophenylalanine induces pessimistic-like judgement bias in sheep Rebecca E. Doyle a,b, * , Geoff N. Hinch b , Andrew D. Fisher c , Alain Boissy d , John M. Henshall a , Caroline Lee a a CSIRO Livestock Industries, Locked Bag 1, Armidale, NSW 2350, Australia b School of Environment and Rural Science, University of New England, Armidale, NSW 2351, Australia c Faculty of Veterinary Science and Animal Welfare Science Centre, University of Melbourne, Victoria 3010, Australia d INRA, UR1213 Herbivores, F-63122 St-Gene `s-Champanelle, France Received 25 March 2010; received in revised form 23 July 2010; accepted 23 July 2010 Psychoneuroendocrinology (2011) 36, 279—288 KEYWORDS Affective state; Animal welfare; Cognitive bias; Emotion; Serotonin; Sheep Summary Judgement bias has potential as a measure of affective state in animals. The serotonergic system may be one mechanism involved with the formation of negative judgement biases. It was hypothesised that depletion of brain serotonin would induce negative judgement biases in sheep. A dose response trial established that 40 mg/kg of p-Chlorophenylalanine (pCPA) administered to sheep for 3 days did not affect feeding motivation or locomotion required for testing judgement biases. Thirty Merino ewes (10 months old) were trained to an operant task for 3 weeks. Sheep learnt to approach a bucket when it was placed in one corner of the testing facility to receive a feed reward (go response), and not approach it when in the alternate corner (no-go response) to avoid a negative reinforcer (exposure to a dog). Following training, 15 sheep were treated with pCPA (40 mg/kg daily) for an extended duration (5 days). Treated and control sheep were tested for judgement bias following 3 and 5 days of treatment, and again 5 days after cessation of treatment. Testing involved the bucket being presented in ambiguous locations between the two learnt locations, and the response of the sheep (go/no-go) measured their judgement of the bucket locations. Following 5 days of treatment, pCPA-treated sheep approached the most positive ambiguous location significantly less than control sheep, suggesting a pessimistic-like bias (treatment bucket location interaction F 1,124.6 = 49.97, p = 0.011). A trend towards a significant interaction was still evident 5 days after the cessation of pCPA treatment ( p = 0.068), however no significant interaction was seen on day 3 of testing ( p = 0.867). These results support the suggestion that judgement bias is a cognitive measure of affective state, and that the serotonergic pathway may be involved. Crown Copyright # 2010 Published by Elsevier Ltd. All rights reserved. * Corresponding author. Current address: School of Animal and Veterinary Science, Charles Sturt University, Locked Bag 588, Wagga Wagga, NSW 2678, Australia. Tel.: +61 2 6933 4721; fax: +61 2 6933 2991. E-mail address: [email protected] (R.E. Doyle). available at www.sciencedirect.com journal homepage: www.elsevier.com/locate/psyneuen 0306-4530/$ — see front matter. Crown Copyright # 2010 Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.psyneuen.2010.07.018
Transcript of Administration of serotonin inhibitor p-Chlorophenylalanine induces pessimistic-like judgement bias...

Administration of serotonin inhibitorp-Chlorophenylalanine induces pessimistic-likejudgement bias in sheep
Rebecca E. Doyle a,b,*, Geoff N. Hinch b, Andrew D. Fisher c,Alain Boissy d, John M. Henshall a, Caroline Lee a
aCSIRO Livestock Industries, Locked Bag 1, Armidale, NSW 2350, Australiab School of Environment and Rural Science, University of New England, Armidale, NSW 2351, Australiac Faculty of Veterinary Science and Animal Welfare Science Centre, University of Melbourne, Victoria 3010, Australiad INRA, UR1213 Herbivores, F-63122 St-Genes-Champanelle, France
Received 25 March 2010; received in revised form 23 July 2010; accepted 23 July 2010
Psychoneuroendocrinology (2011) 36, 279—288
KEYWORDSAffective state;Animal welfare;Cognitive bias;Emotion;Serotonin;Sheep
Summary Judgement bias has potential as a measure of affective state in animals. Theserotonergic system may be one mechanism involved with the formation of negative judgementbiases. It was hypothesised that depletion of brain serotonin would induce negative judgementbiases in sheep. A dose response trial established that 40 mg/kg of p-Chlorophenylalanine (pCPA)administered to sheep for 3 days did not affect feeding motivation or locomotion required fortesting judgement biases. Thirty Merino ewes (10 months old) were trained to an operant task for3 weeks. Sheep learnt to approach a bucket when it was placed in one corner of the testing facilityto receive a feed reward (go response), and not approach it when in the alternate corner (no-goresponse) to avoid a negative reinforcer (exposure to a dog). Following training, 15 sheep weretreated with pCPA (40 mg/kg daily) for an extended duration (5 days). Treated and control sheepwere tested for judgement bias following 3 and 5 days of treatment, and again 5 days aftercessation of treatment. Testing involved the bucket being presented in ambiguous locationsbetween the two learnt locations, and the response of the sheep (go/no-go) measured theirjudgement of the bucket locations. Following 5 days of treatment, pCPA-treated sheepapproached the most positive ambiguous location significantly less than control sheep, suggestinga pessimistic-like bias (treatment � bucket location interaction F1,124.6 = 49.97, p = 0.011). Atrend towards a significant interaction was still evident 5 days after the cessation of pCPAtreatment (p = 0.068), however no significant interaction was seen on day 3 of testing(p = 0.867). These results support the suggestion that judgement bias is a cognitive measureof affective state, and that the serotonergic pathway may be involved.Crown Copyright # 2010 Published by Elsevier Ltd. All rights reserved.
* Corresponding author. Current address: School of Animal and Veterinary Science, Charles Sturt University, Locked Bag 588, Wagga Wagga,NSW 2678, Australia. Tel.: +61 2 6933 4721; fax: +61 2 6933 2991.
E-mail address: [email protected] (R.E. Doyle).
ava i lab le at www.sc ienced i rect .com
journa l homepage: www.e l sev ie r.com/locate/psyneuen
0306-4530/$ — see front matter. Crown Copyright # 2010 Published by Elsevier Ltd. All rights reserved.
doi:10.1016/j.psyneuen.2010.07.018
280 R.E. Doyle et al.
1. Introduction
It is now increasingly accepted that animals have the abilityto experience emotions, and that the valence of their affec-tive state is an important dimension of their welfare (Paulet al., 2005; Boissy et al., 2007; Mendl et al., 2009; Veissieret al., 2009). Affective states are comprised of physiological,behavioural and cognitive components, which have evolvedto help animals avoid harm and obtain valuable resources(Rolls, 1999; Desire et al., 2002; Paul et al., 2005; Boissyet al., 2007). In addition to these, a subjective component ofaffect is also present, in humans at least, whereby theindividual is conscious of the emotion they are experiencing(Desire et al., 2002; Paul et al., 2005). While we currentlyhave strong behavioural and physiological measures of wel-fare in animals, these measures may not provide a completeassessment of an animal’s welfare (Dawkins, 2003; Gregory,2004; Paul et al., 2005). Finding additional ways to measurewelfare by focusing on the cognitive component of affectmay help improve overall welfare assessment of animals.
In humans, affective states influence the way informationis interpreted by altering cognitive processing (Mineka andSutton, 1992; Beevers and Carver, 2003). Negative affectivestates like anxiety and depression lead to a stronger focus on,and a stronger memory of threatening information whencompared to people in non-anxious states (Beevers andCarver, 2003). Negative affective states also increase thelikelihood of interpreting new information as being poten-tially threatening (Mineka and Sutton, 1992; Bar-Haim et al.,2007). Following the success of an innovative experiment byHarding et al. (2004), it was hypothesised that similar biasesin cognition may also be present in non-human animals, andthat cognitive biases could be used to measure a dimension oftheir welfare (Paul et al., 2005). Since then, experiments in avariety of species have shown that animals judge ambiguousinformation differently according to the conditions they havebeen exposed to prior to testing (Bateson and Matheson,2007; Burman et al., 2008; Matheson et al., 2008; Mendlet al., 2009; Doyle, 2010; Doyle et al., 2010a). Judgementbias testing in animals involves teaching an animal to performan operant task using its spatial, visual or auditory skillswhere the animal learns to distinguish between two cuesthat are reinforced differently; either positive and negative,positive and non-positive, or more positive and less positive(for review see Mendl et al., 2009). To test for biases injudgement, novel, ambiguous cues that relate to the learntones are presented to the animal, which then respondsaccording to whether it assesses the novel cue as havingpositive or negative characteristics.
Using a variety of treatments, animal studies have induceddifferent biases in judgement. Long-term negative treat-ments have resulted in more negative, or pessimistic-likejudgements (Harding et al., 2004; Bateson and Matheson,2007; Burman et al., 2008; Doyle, 2010). More positive, oroptimistic-like judgement biases were generated by anenriched environment in starlings (Matheson et al., 2008),and a short-term stressor in sheep (Doyle et al., 2010a).These studies supply evidence suggesting that judgementbias is measuring affective states in animals. A review byMendl et al. (2009) suggested that a pharmacological treat-ment may modify judgement, and thereby be used to inves-tigate mechanisms involved in the formation of judgement
biases. Following demonstrations of judgement bias in stu-dies in sheep (Doyle, 2010; Doyle et al., 2010a,b), it wasproposed that a pharmacological treatment to reduce brainserotonin levels may be an appropriate way to begin testingthe mechanisms influencing judgements.
Depletion of the neurotransmitter serotonin is thought tobe one of the main causes of clinical depression (Meltzer,1990; Sjoerdsma and Palfreyman, 1990; Stockmeier et al.,1998; Cowen, 2002; Ruhe et al., 2007; Krishnan and Nestler,2008) and also plays an important role in fear and anxiety(Graeff et al., 1996). Serotonin is synthesised in the bodyfrom the amino acid tryptophan, which is converted into 5-hydroxy-tryptophan (5-HTP). From there, 5-HTP crosses theblood-brain barrier, where it is converted into serotonin(Meltzer, 1990; Jonnakuty and Gragnoli, 2008). Depletionof brain serotonin can occur through various mechanisms,including the inhibition of 5-HTP crossing the blood—brainbarrier (Ruhe et al., 2007; Krishnan and Nestler, 2008).
One example of a serotonin antagonist is pCPA whichinhibits brain serotonin synthesis by preventing 5-HTPattach-ing to the binding sites used to transport it into the brain(Meltzer, 1990; Ruhe et al., 2007). This antagonist has beenshown to decrease brain serotonin levels and increasedepression related behaviour in a variety of species (Reveliset al., 1998), including a large number of rat studies (O’Learyet al., 2007; Kubala et al., 2008; Li et al., 2009). There are noexamples in the literature of pCPA being used to reduce brainserotonin in sheep; however, it was hypothesised that treat-ment with this drug would be an appropriate initial investi-gation. Initially, a dose response trial was conducted in sheepto determine the optimum dose of pCPA, which was thenutilised in the main experiment.
It was hypothesised that sheep treated with pCPA wouldhave a more pessimistic-like interpretation of ambiguousinformation than untreated sheep in a judgement bias test.It was further hypothesised that pCPA treatment would alsoalter behaviour in test situations to reflect a negative affec-tive state. Specifically, causing a reduction in movement andvocalisation in an open field test and decreased motivation toapproach novel objects (e.g. Desire et al., 2002; Villalbaet al., 2009).
2. Materials and methods
All experimental procedures were approved by the CSIROAnimal Ethics Committee and complied with the AustralianCode of Practice for the Care and Use of Animals for ScientificPurposes.
2.1. Animal details
Merino ewe hoggets (10—12 months of age) were housedindoors in an animal house, in group pens, on slatted floorsfor both dose response and judgement bias experiments. Allexperimental events occurred either within the animal houseor in surrounding yards. Prior to commencing the studies, thesheep were acclimated to the animal house for 2 weeks.Sheep were fed a maintenance ration once daily consisting oflucerne pellets and wheaten chaff. This was given in theafternoons following any habituation, operant task training,behavioural testing or judgement bias testing.

Administration of serotonin inhibitor p-Chlorophenylalanine 281
2.2. Drug details
The serotonin antagonist 4-chloro-DL-phenylalanine methylester hydrochloride (Sigma Aldrich, St. Louis, USA; from nowreferred to as ‘‘pCPA’’) was used because it is chemicallymore soluble than standard pCPA and can cross the bloodbrain barrier more effectively (Sigma Aldrich, 2010). Prior totreatment, pCPA was dissolved in Water for Injection BP(Baxter, Toongabbie, Australia) at a rate of 50 mg/ml. Thedose volume for each sheep was calculated according to bodyweight prior to commencing treatment. The diluted pCPA, oran equivalent volume of water for injection in the case ofcontrol sheep, was administered intraperitoneally using an18 gauge 1 inch needle (BD, New Jersey, USA). The solutionwas injected into the right paralumbar fossa, midwaybetween the iliac crest and the last rib, and about 50 mmbelow the end of the lumbar processes (Hurter et al., 1987).
2.3. Facility details
A facility (2 m � 3 m) with solid wooden walls (1.5 m high)was used as the testing facility (Fig. 1) for judgement bias,feeding motivation and novel object testing (previouslydescribed by Doyle et al., 2010a). Video cameras (Panasonic640 TVL Colour cameras, 5:50 mm lens, Osaka, Japan) wereset up at either end of the test arena tomeasure the responseof sheep to the test, and images were recorded in real timeon a digital video recorder (Pacom, Castle Hill, Australia).The sheep entered via the mesh door of the start box andexited when the test was completed by a door on the righthand side wall. Wooden panels in the front corners of thearena could be raised vertically to expose the sheep to thenegative stimulus (a dog, standing quietly) when training andtesting the sheep for judgment bias.
[(Figure_1)TD$FIG]
Figure 1 Testing arena used for judgement bias, feedingmotivation and novel object tests. Bucket locations for judgmentbias testing are presented for a sheep trained with the left cornerpositive. Bucket locations refer to the distance from the positivebucket: locations 0.8—1.2 m are ambiguous, and 2.0 m is thenegative location. D Signifies video camera position.
2.4. Dose response trial
Thirty sheep were habituated to the testing facility that wasused for feeding motivation and novel object tests. Initially,this involved the sheep entering the facility daily in groupsof three to receive a standard feed reward (30 g lucernepellets/sheep) in 3 identical buckets placed in the centre ofthe far wall. After 3 days, sheep then entered the facilityindividually to receive the same feed reward in a singlebucket at the same location. Every day, for a total of 8 days,each sheep entered and received the feed reward threeconsecutive times. During this period, six sheep that failedto eat in the facility were eliminated from the study group.The remaining 24 sheep were used in the dose responsetrial.
2.4.1. Drug treatmentFollowing habituation, sheep were allocated to treatmentgroups (n = 8) and weighed. Treatment groups were control(water for injection BP only), low dose (20 mg/kg pCPA), orhigh dose (40 mg/kg pCPA). The pCPA was administered forthree days: on day 1 sheep received the full dose at 0800 h(AM), and on days 2 and 3 the total dose volumewas split; halfinjected at 0800 and half at 1600 h (AM and PM). Revelis et al.(1998) concluded that administration of pCPA twice a day, at75 mg/kg of body weight, for 3 consecutive days woulddeplete brain serotonin in any species significantly. Thisand additional literature (Fletcher et al., 2001; Kubalaet al., 2008; Li et al., 2009) suggested that either long-termdoses, or localised injections of pCPA, result in depressive-like behavioural changes and decreased brain serotonin.Between these papers a trendwhereby bodyweight increaseswere inversely related to the dose of pCPA per kg required toinduce behavioural effects was also seen. As a result of this,and because a more potent compound of pCPAwas chosen forthe current study, lower dose rates of 20 mg/kg and 40 mg/kgwere used.
Refer to Table 1 for a full, chronological timeline of thedrug treatment and testing schedule.
2.4.2. Open field testThe behaviour of the sheep was tested in an open field, whichis commonly used as a test of reactivity in sheep (e.g Villalbaet al., 2009), on days 1 and 4 of the trial. The test area was3 m � 8 m, had 1 m high opaque sides. The test area wasdivided into eight 3 m � 1 m zones. The sheep individuallyentered the test for 3 min. The movement (total number ofzones crossed), the total number of vocalisations (bleats),urinations, and defecations were recorded by a motionlessperson 5 m from the outside of the test pen. It was deter-mined that the sheep had crossed a zone once its two frontfeet entered a different zone.
2.4.3. Feeding motivation testFeeding motivation was tested on days 1 and 4 of treatmentfollowing open field testing. To test for feeding motivation,sheep individually entered the testing facility and the stan-dard feed reward was presented in the bucket at the centrelocation. Sheep had 1 min to approach and eat the feed.Latency to approach the bucket and total amount of feedconsumed were measured.

Table 1 Timeline of the dose response trial.
Day Time Event
1 AM Blood samplingFull drug dose
PM Blood samplingOpen field testBlood samplingFeeding motivation testNovel object test (traffic cone)Isolation box test
2 AM Blood samplingHalf drug doseBlood samplingHalf drug dose
PM Blood samplingHalf drug dose
3 AM Blood samplingHalf drug dose
PM Blood samplingOpen field test
4 AM Blood samplingFeeding motivation testNovel object test (ball)Isolation box test
282 R.E. Doyle et al.
2.4.4. Novel object testTo test for reactivity to a novel object, sheep entered the testfacility a second time immediately after the feeding motiva-tion test on days 1 and 4. The same reward was presented atthe same location and a novel object (an orange traffic coneon day 1 and a large orange ball on day 4) was placedequidistant between the start box and the bucket. The sheephad 1 min to approach and eat the feed. Latency to exit thestart box, latency to approach the bucket, contact with thenovel object and total amount of feed consumed were allmeasured.
Video recordings of one control sheep and one low dosetreatment sheep in the feeding motivation and novel objecttests were lost from the fourth day of the experiment. Thebehavioural results of these sheep are not presented below.
2.4.5. Isolation box testOn days 1 and 4, the agitation of sheep to isolation in aconfined area was measured in the isolation box test. Sheepwere isolated in a wooden box (1.5 m � 0.75 m � 1.5 m) for30 s. Agitation was measured using an electronic meter(custom built on site), which generated a numerical valueof the sheep’s agitation based on vibrations from movementand vocalisations in the box (Blache and Ferguson, 2005;Bickell et al., 2009).
2.4.6. Blood cortisol concentrationsPrior to the experiment commencing, wool over the neck ofthe sheep was clipped to facilitate blood collection. Bloodsamples were collected from the sheep via jugular vene-puncture into 8.5 ml serum separating tubes (B.D., NewJersey, USA). Samples were collected daily before pCPAwas administered to measure baseline cortisol concentra-tions. Blood was also collected immediately before open field
testing and 20 min after release from the test to measure anyeffects the pCPA may have had on the physiological stressresponse of sheep. Following collection, samples were cen-trifuged for 10 min at 4 8C and 2060 g. Serum was stored at�20 8C for cortisol analysis.
Serum cortisol concentrations were measured using acommercial radioimmunoassay (Orion Diagnostica, Espoo,Finland), previously validated for ovine serum cortisol inthe laboratory (Paull et al., 2007). The three quality controls(QCs) contained 32.8, 67.6, and 156.4 nmol/L of cortisol withinter-assay coefficients of variation (CVs) of 14.8%, 10.7%,and 10.3% for the three QCs respectively. The intra-assay CVsfor the same QCs were 17.4%, 12.9%, and 12.1% respectively.
2.5. Judgement bias study
Thirty-six more sheep were habituated to the testing facilityas described in Section 2.4. The only exception was that thebucket was placed in the centre of the pen, rather thanagainst the back wall.
2.5.1. Judgement bias trainingOnce habituated, the sheep were trained to a spatial differ-entiation task adapted from Burman et al. (2008), and pre-viously used to test for judgement biases in sheep (Doyleet al., 2010a). The sheep were randomly assigned either theleft (n = 18) or right (n = 18) corner of the facility for positivetraining, and the feed bucket was placed in the allocatedcorner. Individually, the sheep approached and ate from thebucket on five consecutive entries per training session, with 1session per day.
After 5 days of positive training, negative training wasincorporated. The negative training involved the bucketbeing placed in the alternate corner to the positive location.If the sheep approached within a 30 cm radius of the bucket,the side panel lifted to reveal the dog (sitting quietly). Assoon as the sheep retreated, the panel was lowered. Trainingcontinued for a total of 10 days.
Training sessions were split into three positive and twonegative events. This was done in a pseudo-random order toensure no more than two negative or positive events suc-ceeded each other, and each training session ended with thepresentation of the positive location. Upon each entry, thesheep had a 25-s time limit to approach the bucket beforethis was deemed a no-go response. For the sheep to beclassified as trained they had to respond correctly (go/no-go) to each bucket location for three out of four consecutivetraining sessions.
During this time, six sheep did not reach criteria and wereeliminated from the experiment. Thirty sheep were success-fully trained to the judgement bias test, 16 sheep trained tothe left side being positive and 14 sheep trained to the rightside being positive. Training was continued throughout thedrug treatment period as outlined above, apart from onjudgement bias testing days.
2.5.2. Drug treatmentThe 30 remaining sheep were randomly allocated to treat-ment groups (treatment or control, n = 15) after stratifica-tion by weight. After assessment of the results from the pilotstudy, a dose rate of 40 mg/kg/day of pCPA was chosen and

Table 2 Timeline of the judgement bias study.
Day Time Event
1 PM Half drug dose2 AM Half drug dose
PM Half drug dose3 AM Half drug dose
PM Half drug dose4 AM Half drug dose
Judgement bias testPM Half drug dose
5 AM Half drug doseFeeding motivation testNovel object testOpen field test
PM Half drug dose6 AM Half drug dose
PM Judgement bias test11 AM Post treatment
judgement bias testing
Administration of serotonin inhibitor p-Chlorophenylalanine 283
administered for 5 days at half the daily rate on 2 occasionsper day, in the same way as described in Section 2.4. The firsthalf dose was administered on day 1 (PM), and therefore thefinal dose was given on day 6 (AM). This approach was chosenso the sheep could be tested for a difference in affectivestate shortly after the final half dose was given, as opposed tothe protocol used in the dose response trial.
Refer to Table 2 for timeline of the drug treatment andtesting schedule.
2.5.3. Judgement bias testingAll sheep were tested for judgement bias following 3 and 5full days of drug treatment (on days 4 and 6), and 5 daysfollowing the cessation of drug treatment (day 11). On all 3testing days, sheep individually entered the arena five con-secutive times. The bucket was presented once in five dif-ferent locations in the following order: ambiguous locations0.8 m, 1.0 m, and 1.2 m from the positive reference location,the positive reference location and the negative referencelocation. During the tests, reference locations remainedreinforced, however ambiguous locations were not rein-forced (no dog or food if the bucket was approached). Thego/no-go response of the sheep to each bucket was mea-sured. A ‘‘go’’ was defined as a sheep approaching with theirforelimbs within a 30 cm radius of the bucket. The estimatedduration of feed deprivation prior to judgement bias testingwas between 16 and 19 h and this variation was distributedevenly across treatments.
One sheep from the pCPA-treated group did not respond toany of the judgment bias tests or the feeding motivation andnovel object tests. The data from this animal has beenexcluded from these analyses but were retained for the openfield test results.
2.5.4. Feeding motivation, novel object and open fieldtestingFollowing 4 days of treatment (day 5), sheep were tested forfeeding motivation, reactivity to a novel object (traffic cone)and behaviour in an open field test in that order. The methods
used were the same as those outlined in Section 2.4. Testingoccurred in a different order to that used in the dose responsetrial to allow for judgement bias training to be incorporatedand minimise sheep handling.
2.6. Statistical analyses
All data were assessed for normality and homoscedasticityand met these assumptions.
2.6.1. Judgement biasResults for each day were analysed separately as binary (go/no-go) data using a generalised linear mixed model with logitlink function:
logp
1� p
� �
where p is the proportion of times a bucket was approached.The linear component of the model contained a fixed term(treatment), a covariate (bucket location) and the interac-tion between these, which was the main point of interest forthe experiment. Animal was fitted as a random effect. Thefull linear component of the model was:
mþ aþ bþ a:bþ c
where a = factor (Treatment), b = variate (Bucket location)and c = random factor (Animal). ASReml (VSN InternationalLtd., Hemel Hempstead, United Kingdom) was used to fit themodel, and significance was assessed using Wald F Statistics.Because of the random factor for animal the denominatordegrees of freedom did not take integer values, and ASRemlestimated these using the method of Kenward and Roger(Gilmour et al., 2006). Bucket locations were reported astheir distance from the positive location (0, 0.8, 1.0, 1.2 and2.0 m). The judgement bias results from the three test dayswere analysed separately to account for heterogeneous vari-ance between testing days.
2.6.2. Behavioural testsAll analyses were performed in GenStat 10.2 (VSN Interna-tional Ltd., Hemel Hempstead, United Kingdom).
Where appropriate, chi square analyses were performedon all censored behaviours (those that had a cut off point;e.g. latency to approach the bucket) to examine treatmentdifferences. Following that, all behaviours were tested fortreatment differences using one-way ANOVAs, where treat-ment was the factor. Censored events in the correspondingbehavioural data were removed prior to each ANOVA. Beha-viours from the open field test and agitation scores from theisolation box test were also analysed for treatment differ-ences using one-way ANOVA with treatment as a factor.
2.6.3. Cortisol resultsAll analyses were performed in GenStat 10.2 (VSN Interna-tional Ltd., Hemel Hempstead, United Kingdom). Cortisolconcentrations from each sample point (before open fieldtesting day 1, after open field testing day 1, before open fieldtesting day 4, and after open field testing day 4) wereanalysed individually for treatment differences using anANOVA.
Baseline cortisol concentrations were analysed for treat-ment differences using a repeated measures ANOVA which

Table
3Behaviourala
ndphysiologica
lresultsfrom
theopenfield,feedingmotiva
tionan
dnove
lobject
testsofsheepin
thedose
response
trial.Themean
sofeac
htreatmentgroup
arepresented,followedbythestan
darderrorofthedifference
.
Openfield
test
Feedingmotiva
tiontest
Nove
lobject
test
Treatment
Move
ment
Voca
lisation
Pre-test
cortisol
(ng/
ml)
Post-test
cortisol
(ng/
ml)
Bucket
approac
h(s)
Feed
consumed
(g)
Startbox
exit(s)
Bucket
approac
h(s)
Feed
consumed
(g)
Nove
lobject
contact
(s)
Day
1Control
33�
8.6
15�
4.6
28.8�
8.98
19.7�
6.65
2.7�
0.5
62�
7.3
7.2�
5.1
5.0�
5.7
62�
7.0
12.2�
8.8
Low
dose
35�
8.6
12�
4.6
25.9�
8.98
27.7�
6.65
3.5�
0.5
63�
7.3
4.4�
5.1
5.5�
5.7
56�
7.0
4.4�
8.8
Highdose
40�
8.6
19�
4.6
18.7�
8.98
16.2�
6.65
3.0�
0.5
55�
7.3
7.9�
5.1
11.8�
5.7
53�
7.0
28.6�
8.8
Day
4Control
43�
6.8
12�
4.3
35.4�
7.24
21.7�
4.76
8.4�
4.1
52�
8.0
6.9�
7.1
4.0�
1.1
59�
8.0
16.0�
10.3
Low
dose
33�
6.8
10�
4.3
33.5�
7.24
24.4�
4.76
3.0�
4.1
63�
8.0
18.1�
7.1
4.1�
1.1
59�
8.0
17.3�
10.3
Highdose
37�
6.8
13�
4.3
31.2�
7.24
16.4�
4.76
2.4�
4.1
59�
8.0
4.0�
7.1
5.3�
1.1
49�
8.0
12.6�
10.3
284 R.E. Doyle et al.
tested for statistically significant effects of treatment, day,and a treatment � day interaction. Baseline cortisol concen-trations taken on day 1 prior to treatment were fitted as acovariate.
3. Results
3.1. Dose response trial
3.1.1. Open field testBehavioural and serum cortisol results from the open fieldtest are summarised in Table 3. No significant treatmentdifferences were noted in any of the behaviours( p > 0.05). There were also no significant treatment differ-ences in the cortisol concentrations of the sheep pre or postthe open field test (all p > 0.05).
3.1.2. Feeding motivation and novel object testsChi square analyses were performed on censored results todetermine if a treatment difference existed between thenumber of bucket approaches and novel object approaches inthe novel object test. On day 1, 6 control sheep, 5 low and 5high dose sheep made contact with the novel object(x2 = 0.38, p = 0.83). On day 4 of the novel object test twosheep from the high treatment group did not approach thebucket (x2 = 2.09, p = 0.35). Three out of 7 control and lowdose sheep, and 5 out of 8 high dose sheep made contact withthe novel object (x2 = 0.78, p = 0.68). No chi square analyseswere performed for any data in the feeding motivation testbecause there were no censored events as all sheepapproached the bucket.
All timed behavioural results from the feeding motivationand novel object tests are summarised in Table 3. No statis-tically significant treatment differences were seen in anymeasured behaviours.
3.1.3. Isolation box testNo significant differences were seen in the isolation box teston either day of testing. On day 1, average agitation scoresfor the control, low and high dose groups were 64, 64 and 58respectively (F2,21 = 0.06, p = 0.942). On day 4 the averageagitation scores for the three groups were 112, 78 and 57respectively (F2,21 = 1.75, p = 0.198).
3.1.4. Baseline cortisol concentrationsBaseline cortisol results are presented in Table 4. No sig-nificant treatment � time interactions, nor general treat-ment differences, were identified ( p > 0.05). A significanteffect of time was present however (F4,84 = 1.00, p = 0.006).
3.2. Judgement bias study
3.2.1. Judgement biasFollowing 3 days of treatment (Fig. 2) there were no sig-nificant effects on treatment � bucket location(F1,135.4 = 0.82, p = 0.867). Treatment was also not signifi-cantly different (F1,23.1 = 0.36, p = 0.553) however bucketlocation was highly significant (F1,133.1 = 89.62, p < 0.001).
Judgement bias testing following 5 days of drug treatment(Fig. 3) resulted in a significant treatment � bucket locationinteraction (F1,131.2 = 6.69, p = 0.011). Control sheep

Table 4 Baseline serum cortisol concentrations (ng/ml) of sheep in the dose response trial. The means of each treatment at eachtime point, and the combined mean are presented. The standard error of the difference for treatment means is �6.09.
Serum cortisol concentrations (ng/ml)
Day 2 AM Day 2 PM Day 3 AM Day 3 PM Day 4 AM
Control 22.67 17.74 19.86 27.12 26.25Low dose 27.82 22.28 16.91 29.71 33.09High dose 27.71 21.30 19.31 26.84 30.78Combined mean 26.07 20.44 18.69 27.89 30.04
Administration of serotonin inhibitor p-Chlorophenylalanine 285
approached the 0.8 m and 1 m ambiguous locations morethan the pCPA-treated sheep.
Five days after the cessation of treatment judgement biaswas tested again (Fig. 4) and a treatment � bucket location[(Figure_2)TD$FIG]
Figure 2 Approach responses of sheep following 3 days of pCPAtreatment. Back-transformed approach responses presented as aproportion of 1 in the form of a bar graph. Bucket locations arerepresented as distance from the positive bucket with 0 m = pos-itive bucket location and 2 m = negative bucket location. GLMMcurves of logit-transformed data presented with standard errorsfor treated (*) and control (&) sheep.[(Figure_3)TD$FIG]
Figure 3 Approach responses of sheep following 5 days of pCPAtreatment. Back-transformed approach responses presented as aproportion of 1 in the form of a bar graph. Bucket locations arerepresented as distance from the positive bucket with 0 m = pos-itive bucket location and 2 m = negative bucket location. GLMMcurves of logit-transformed data presented with standard errorsfor treated (*) and control (&) sheep.
interaction tended towards statistical significance(F1,119.9 = 3.38, p = 0.068). Control sheep approached the0.8 m location more frequently, but approached the 1.2 mlocation less frequently than pCPA treated sheep.
3.2.2. Feeding motivation and novel object testsSome sheep did not approach the novel object or the feedbucket when the novel object was present. Chi squaredanalyses were performed on these behaviours to accountfor the censored results. Six out of 14 pCPA-treated sheepand one out of 15 controls approached the novel object,which was a significant difference (x2 = 5.18, p = 0.022).Five pCPA-treated sheep and four controls did not approachthe bucket at all during the novel object test (x2 = 0.28,p = 0.60).
All behavioural results from the feeding motivation andnovel object tests are summarised in Table 5. Latency tocontact the novel object was not analysed because only onecontrol sheep approached it.
3.2.3. Open field testNumber of vocalisations of the pCPA-treated sheep was sig-nificantly lower than that of the controls (8 vs 21 vocalisa-tions respectively; S.E.D. = 3.76, p = 0.001).
There was no effect of treatment on movement within theopen field (24 vs 25 zones crossed respectively; S.E.D. = 6.55,p = 0.864).[(Figure_4)TD$FIG]
Figure 4 Approach responses of sheep 5 days after the cessa-tion of pCPA treatment. Back-transformed approach responsespresented as a proportion of 1 in the form of a bar graph. Bucketlocations are represented as distance from the positive bucketwith 0 m = positive bucket location and 2 m = negative bucketlocation. GLMM curves of logit-transformed data presented withstandard errors for treated (*) and control (&) sheep.
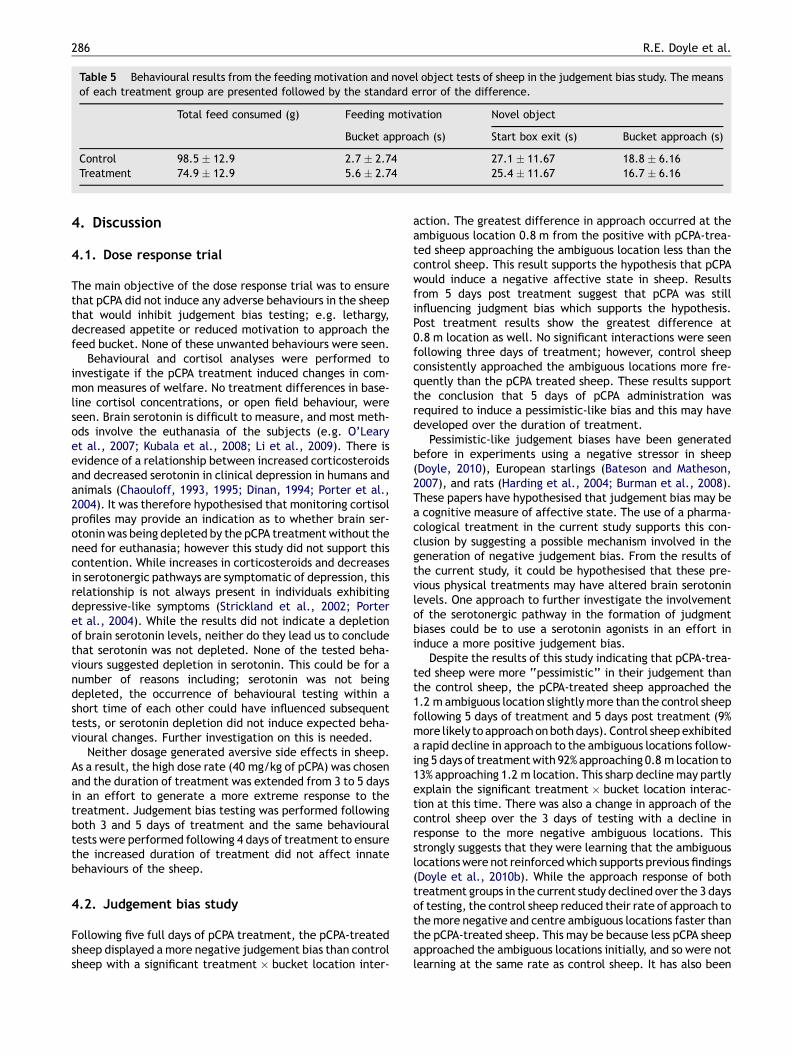
Table 5 Behavioural results from the feeding motivation and novel object tests of sheep in the judgement bias study. The meansof each treatment group are presented followed by the standard error of the difference.
Total feed consumed (g) Feeding motivation Novel object
Bucket approach (s) Start box exit (s) Bucket approach (s)
Control 98.5 � 12.9 2.7 � 2.74 27.1 � 11.67 18.8 � 6.16Treatment 74.9 � 12.9 5.6 � 2.74 25.4 � 11.67 16.7 � 6.16
286 R.E. Doyle et al.
4. Discussion
4.1. Dose response trial
The main objective of the dose response trial was to ensurethat pCPA did not induce any adverse behaviours in the sheepthat would inhibit judgement bias testing; e.g. lethargy,decreased appetite or reduced motivation to approach thefeed bucket. None of these unwanted behaviours were seen.
Behavioural and cortisol analyses were performed toinvestigate if the pCPA treatment induced changes in com-mon measures of welfare. No treatment differences in base-line cortisol concentrations, or open field behaviour, wereseen. Brain serotonin is difficult to measure, and most meth-ods involve the euthanasia of the subjects (e.g. O’Learyet al., 2007; Kubala et al., 2008; Li et al., 2009). There isevidence of a relationship between increased corticosteroidsand decreased serotonin in clinical depression in humans andanimals (Chaouloff, 1993, 1995; Dinan, 1994; Porter et al.,2004). It was therefore hypothesised that monitoring cortisolprofiles may provide an indication as to whether brain ser-otonin was being depleted by the pCPA treatment without theneed for euthanasia; however this study did not support thiscontention. While increases in corticosteroids and decreasesin serotonergic pathways are symptomatic of depression, thisrelationship is not always present in individuals exhibitingdepressive-like symptoms (Strickland et al., 2002; Porteret al., 2004). While the results did not indicate a depletionof brain serotonin levels, neither do they lead us to concludethat serotonin was not depleted. None of the tested beha-viours suggested depletion in serotonin. This could be for anumber of reasons including; serotonin was not beingdepleted, the occurrence of behavioural testing within ashort time of each other could have influenced subsequenttests, or serotonin depletion did not induce expected beha-vioural changes. Further investigation on this is needed.
Neither dosage generated aversive side effects in sheep.As a result, the high dose rate (40 mg/kg of pCPA) was chosenand the duration of treatment was extended from 3 to 5 daysin an effort to generate a more extreme response to thetreatment. Judgement bias testing was performed followingboth 3 and 5 days of treatment and the same behaviouraltests were performed following 4 days of treatment to ensurethe increased duration of treatment did not affect innatebehaviours of the sheep.
4.2. Judgement bias study
Following five full days of pCPA treatment, the pCPA-treatedsheep displayed amore negative judgement bias than controlsheep with a significant treatment � bucket location inter-
action. The greatest difference in approach occurred at theambiguous location 0.8 m from the positive with pCPA-trea-ted sheep approaching the ambiguous location less than thecontrol sheep. This result supports the hypothesis that pCPAwould induce a negative affective state in sheep. Resultsfrom 5 days post treatment suggest that pCPA was stillinfluencing judgment bias which supports the hypothesis.Post treatment results show the greatest difference at0.8 m location as well. No significant interactions were seenfollowing three days of treatment; however, control sheepconsistently approached the ambiguous locations more fre-quently than the pCPA treated sheep. These results supportthe conclusion that 5 days of pCPA administration wasrequired to induce a pessimistic-like bias and this may havedeveloped over the duration of treatment.
Pessimistic-like judgement biases have been generatedbefore in experiments using a negative stressor in sheep(Doyle, 2010), European starlings (Bateson and Matheson,2007), and rats (Harding et al., 2004; Burman et al., 2008).These papers have hypothesised that judgement bias may bea cognitive measure of affective state. The use of a pharma-cological treatment in the current study supports this con-clusion by suggesting a possible mechanism involved in thegeneration of negative judgement bias. From the results ofthe current study, it could be hypothesised that these pre-vious physical treatments may have altered brain serotoninlevels. One approach to further investigate the involvementof the serotonergic pathway in the formation of judgmentbiases could be to use a serotonin agonists in an effort ininduce a more positive judgement bias.
Despite the results of this study indicating that pCPA-trea-ted sheep were more ‘‘pessimistic’’ in their judgement thanthe control sheep, the pCPA-treated sheep approached the1.2 m ambiguous location slightlymore than the control sheepfollowing 5 days of treatment and 5 days post treatment (9%more likely to approach onbothdays). Control sheep exhibiteda rapid decline in approach to the ambiguous locations follow-ing 5 days of treatmentwith 92% approaching 0.8 m location to13% approaching 1.2 m location. This sharp decline may partlyexplain the significant treatment � bucket location interac-tion at this time. There was also a change in approach of thecontrol sheep over the 3 days of testing with a decline inresponse to the more negative ambiguous locations. Thisstrongly suggests that they were learning that the ambiguouslocationswere not reinforcedwhich supports previous findings(Doyle et al., 2010b). While the approach response of bothtreatment groups in the current study declined over the 3 daysof testing, the control sheep reduced their rate of approach tothe more negative and centre ambiguous locations faster thanthe pCPA-treated sheep. This may be because less pCPA sheepapproached the ambiguous locations initially, and so were notlearning at the same rate as control sheep. It has also been

Administration of serotonin inhibitor p-Chlorophenylalanine 287
shown in the literature that serotonin depletion decreaseslearning (Altman et al., 1989; Hritcu et al., 2007), which mayexplain the above results. However, there is also evidence ofserotonin depletion not affecting learning (Matsukawa et al.,1997; Petrasek and Stuchlik, 2009). Regardless of the influ-ence, even if pCPA inhibited learning in the current study, theresults seen on day 5 of treatment would still support thehypothesis that pCPA induced a negative judgement bias.
Following 4 days of treatment two significant treatmentdifferences in behaviour were seen. The first showed thatcontrol sheep vocalised significantly more than pCPA-treatedsheep in the open field test. Sheep vocalise strongly whenseparated from flock mates as a way of helping them recon-nect with the flock (Boissy et al., 2005). This differencesuggests that the control sheep were more motivated torejoin the flock than the treated sheep. This difference inmotivation supports the hypothesis that pCPA would reducereactivity in sheep, in a similar way to depressive-like symp-toms in humans and other animals (e.g. Kubala et al., 2008).In contrast to this result, significantly more treated sheepapproached a novel object compared to the controls. Thissuggests that treated sheep had a greater propensity forexploration than the control animals, which is an unexpectedresult. Serotonin is associated with both depressive affectivestates and anxious/fearful affective states in humans. Anxi-ety and fear tend to be associated with the avoidance ofunfamiliar stimuli, whereas depression does not induce thissame sort of avoidance. The difference in this current studycould be of use for future investigations into the identifica-tion of specific affective states beyond valence in animals. Itshould also be noted that no behavioural differences wereseen following 3 days of pCPA treatment in the dose responsetrial. While this could be a result of a difference in treatmentduration, further investigations into this are warranted.
This paper is a preliminary investigation into the mechan-isms involved in the formation of judgement biases in live-stock, and it is suggested that further research is justified.The results provide strong support to the hypothesis thatjudgement bias is measuring the influence that affectivestate valence has on cognitive processing, and thereforehas the potential to be a valuable component of the ofmeasurement of animal welfare.
Role of the funding sources
No external funding sources contributed to this research.
Conflict of interest
There are no conflicts of interest in this study.
Acknowledgements
We would like to acknowledge Ian Colditz, Delphine Aubriot,Guilleme Hervault and Jen Smith for their help conductingthe experiment and reviewing the manuscript.
References
Altman, H.J., Orgen, S.O., Berman, R.F., Normile, H.J., 1989. Theeffects of p-chloroamphetamine, a depletor of brain serotonin,
on the performance of rats in two types of positively reinforcedcomplex spatial discrimination tasks. Behav. Neural Biol. 52,131—144.
Bar-Haim, Y., Lamy, D., Pergamin, L., Bakermans-Kranenburg, M.J.,van IJzendoorn, M.H., 2007. Threat-related attentional bias inanxious and nonanxious individuals: a meta-analytic study. Psy-chol. Bull. 133, 1—24.
Bateson, M., Matheson, S.M., 2007. Performance on a categorisationtask suggests that removal of environmental enrichment induces‘pessimism’ in captive European starlings (Sturnus vulgaris).Anim. Welfare 16, 33—36.
Beevers, C.G., Carver, C.S., 2003. Attentional bias and mood persis-tence as prospective predictors of dysphoria. Cognit. Ther. Res.27, 619—637.
Bickell, S., Poindron, P., Nowak, R., Chadwick, A., Ferguson, D.,Blache, D., 2009. Genotype rather than non-genetic behaviouraltransmission determines the temperament of Merino lambs.Anim. Welfare 18, 459—466.
Blache, D., Ferguson, D., 2005. Increasing sheep meat productionefficiency and animal welfare by selection for temperament. PTFinal Report p59, Meat and Livestock Australia.
Boissy, A., Fisher, A.D., Bouix, J., Hinch, G.N., Le Neindre, P., 2005.Genetics of fear in ruminant livestock. Livest. Prod. Sci. 93,23—32.
Boissy, A., Manteuffel, G., Jensen, M.B., Moe, R.O., Sprujit, B.,Keeling, L.J., Winckler, C., Forkman, B., Dimitrov, I., Langbein,J., Bakken, M., Veissier, I., Aubert, A., 2007. Assessment ofpositive emotions in animals to improve their welfare. Physiol.Behav. 92, 375—397.
Burman, O.H.P., Parker, R., Paul, E.S., Mendl, M., 2008. A spatialjudgement task to determine background emotional state inlaboratory rats. Anim. Behav. 76, 801—809.
Chaouloff, F., 1993. Physiopharmacological interactions betweenstress hormones and central serotonergic systems. Brain Res.Rev. 18, 1—32.
Chaouloff, F., 1995. Regulation of 5-HTreceptors by corticosteroids —where do we stand. Fundam. Clin. Pharmacol. 9, 219—233.
Cowen, P.J., 2002. Cortisol, serotonin and depression: all stressedout? Br. J. Psychiatry 180, 99—100.
Dawkins, M.S., 2003. Behaviour as a tool in the assessment of animalwelfare. Zoology 106, 383—387.
Desire, L., Boissy, A., Veissier, I., 2002. Emotions in farm animals: anew approach to animal welfare in applied ethology. Behav.Process. 60, 165—180.
Dinan,T.G., 1994.Glucocorticoidsand thegenesis ofdepressive-illness— a psychobiological model. Br. J. Psychiatry 164, 365—371.
Doyle, R.E., Fisher, A.D., Hinch, G.N., Boissy, A., Lee, C., 2010a.Release from restraint generates a positive judgement bias insheep. Appl. Anim. Behav. Sci. 122, 28—34.
Doyle, R.E., 2010. Judgement bias: a cognitive measure of affectivestate in sheep. Unpublished PhD Thesis. University of New Eng-land.
Doyle, R.E., Vidal, S., Fisher, A.D., Hinch, G.N., Boissy, A., Lee, C.,2010b. The effect of repeated testing on judgement biases ofsheep. Behav. Process. 83, 349—352.
Fletcher, P.J., Selhi, Z.F., Azampanah, A., Sills, T.L., 2001. Reducedserotonin activitiy disrupts prepulse inhibition of the acousticstartle reflex: effects of 5,7-dihydroxytryptamine and p-chloro-phenylalanine. Neuropsychopharmacology 24, 399—409.
Gilmour, A.R., Gogel, B.J., Cullis, B.R., Thompsom, R., 2006. ASRemlUser Guide Release 2.0. VSN International, Hemel Hempstead.
Graeff, F.G., Guimaraes, F.S., De Andrade, T.G.C.S., Deakin, J.F.W.,1996. Role of 5-HT in stress, anxiety and depression. Pharmacol.Biochem. Behav. 54, 129—141.
Gregory, N.G., 2004. Physiology and Behaiour of Animal Suffering.Blackwell Science, Oxford.
Harding, E.J., Paul, E.S., Mendl, M., 2004. Cognitive bias and affec-tive state. Nature 427, 312.

288 R.E. Doyle et al.
Hritcu, L., Clicinschi, M., Nabeshima, T., 2007. Brain serotonindepletion impairs short-term memory, but not long-term memoryin rats. Physiol. Behav. 91, 567—652.
Hurter, L.R., Mullerstraat, G., Pietersburg, W., 1987. Preliminaryinvestigation into the efficacy of intraperitoneal vaccination ofsheep and goat kids against heartwater. Onderstepoort J. Vet.Res. 54, 507—508.
Jonnakuty, C., Gragnoli, C., 2008. What do we know about serotonin?J. Cell Biol. 217, 301—306.
Krishnan, V., Nestler, E.J., 2008. The molecular neurobiology ofdepression. Nature 455, 894—902.
Kubala, K.H., McGinnis, M.Y., Anderson, G.M., Lumia, A.R., 2008. Theeffect of an anabolic androgenic steroid and low serotonin on socialand non-social behaviors in male rats. Brain Res. 1232, 21—29.
Li, H.P., An, F.L., An, S.C., 2009. Orbitofrontal cortex action of 5-hydroxytryptamine and its receptor in an acute forced swimmingstress-induced depression model. Neural Regen. Res. 4, 530—535.
Matheson, S.M., Asher, L., Bateson, M., 2008. Larger, enriched cagesare associated with ‘optimistic’ response biases in captive Euro-pean starlings (Sturnus vulgaris). Appl. Anim. Behav. Sci. 109,374—383.
Matsukawa, M., Ogawa, M., Nakadate, K., Maeshima, T., Ichitani, Y.,Kawai, N., Okado, N., 1997. Serotonin and acetylcholine arecrucial to maintain hippocampal synapses and memory acquisi-tion in rats. Neurosci. Lett. 230, 13—16.
Meltzer, H., 1990. Role of serotonin in depression. Ann. N.Y. Acad.Sci. 600, 486—500.
Mendl, M., Burman, O.H.P., Richard, M.A., Paul, E.S., 2009. Cognitivebias as an indicator of animal emotion and welfare: emergingevidence and underlying mechanisms. Appl. Anim. Behav. Sci.118, 161—181.
Mineka, S., Sutton, S.K., 1992. Cognitive biases and the emotionaldisorders. Psychol. Sci. 3, 65—69.
O’Leary, O.F., Bechtholt, A.J., Crowley, J.J., Hill, T.E., Page, M.E.,Lucki, I., 2007. Depletion of serotonin and catecholamines blockthe acute behavioural response to different classes of antide-pressany drugs in the mouse tail suspension test. Psychopharma-cology 192, 357—371.
Paul, E.S., Harding, E.J., Mendl, M., 2005. Measuring emotionalprocesses in animals: the utility of a cognitive approach. Neu-rosci. Biobehav. Rev. 29, 469—491.
Paull, D.R., Lee, C., Colditz, I.G., Atkinson, S.J., Fisher, A.D., 2007.The effect of a topical anaesthetic formulation, systemic flunixinand carprofen, singly or in combination, on cortisol and beha-vioural responses of Merino lambs to mulesing. Aust. Vet. J. 85,98—106.
Petrasek, T., Stuchlik, A., 2009. Serotonin-depleted rats are capableof learning in active place avoidance, a spatial task requiringcognitive coordination. Physiol. Res. 58, 299—303.
Porter, R.J., Gallagher, P., Watson, S., Young, A.H., 2004. Cortico-steroid-serotonin interactions in depression: a review of thehuman evidence. Psychopharmacology 173, 1—17.
Revelis, J., Thompson, A.M., Hill Britton, B., Thompson, G.C., 1998.Effects of para-chlorophenylalanine (pCPA) on the bush babyauditory brainstem response. Hearing Res. 116, 119—130.
Rolls, E.T., 1999. The Brain and Emotion. Oxford University Press,Oxford.
Ruhe, H.G., Mason, N.S., Schene, A.H., 2007. Mood is indirectlyrelated to serotonin, norepinephrine and dopamine levels inhumans: a meta-analysis of monoamine depletion studies. Mol.Psychiatry 12, 331—359.
Sigma Aldrich, 2010. Specification sheet. In: http://www.sigmaal-drich.com/catalog/ProductDetail.do?lang=en&N4=C3635.SIGMA&N5=SEARCH_CONCAT_PNO.BRAND_KEY&F=SPEC (Ed. by SigmaAldrich). St Louis, USA.
Sjoerdsma, A., Palfreyman, M.G., 1990. History of serotonin andserotonin disorderes. Ann. N.Y. Acad. Sci. 600, 1—8.
Stockmeier, C.A., Shaprio, L.A., Dilley, G.E., Kolli, T.N., Friedman,L., Rajkowska, G., 1998. Increas in serotonin-1A autoreceptors inthe midbrain of suicuide victims with major depression — post-mortem evidence for decreased serotonin activity. J. Neurosci.18, 7394—7401.
Strickland, P.L., Deakin, J.F.W., Percival, C., Dixon, J., Gater, R.A.,Goldberg, D.P., 2002. Bio-social origins of depression in thecommunity. Interactions betweeen social adversity, cortisoland serotonin neurotransmission. Br. J. Psychiatry 180, 168—173.
Veissier, I., Boissy, A., Desire, L., Greiveldinger, L., 2009. Animals’emotions: studies in sheep using appraisal theories. Anim. Wel-fare 18, 347—354.
Villalba, J.J., Manteca, X., Provenza, F.D., 2009. Relationship be-tween reluctance to eat novel foods and open-field behavior insheep. Physiol. Behav. 96, 276—281.