
Activation and regulation of Toll-like receptors 2 and 1 in human leprosy
8
NATURE MEDICINE • VOLUME 9 • NUMBER 5 • MAY 2003 525 ARTICLES Toll-like receptors (TLRs) represent one mechanism by which the innate immune system recognizes biochemical patterns dis- played by infectious invaders. The specificity of TLRs in mediat- ing responses to defined bacterial ligands has been clearly shown: TLR2 homodimers 1,2 and TLR2-TLR1 heterodimers medi- ate the response to microbial triacylated lipoproteins 3,4 , TLR3 to double-stranded viral RNA 5 , TLR4 to lipopolysaccharide (LPS) 6–8 , TLR5 to bacterial flagellin 9 , TLR2-TLR6 to diacylated lipopep- tides 10 , TLR7 and TLR8 to imidazoquinolines 11,12 and TLR9 to bacterial CpG DNA sequences 13 . Upon activation, TLRs trigger the release of cytokines and the induction of co-stimulatory molecules that can influence the nature of the adaptive T- or B- cell response 14 . Activation of TLRs also induces antimicrobial pathways that kill intracellular organisms 15 . Leprosy provides an excellent opportunity to investigate mechanisms of innate and adaptive immunity in humans. First, leprosy primarily affects skin, so the lesions are readily accessible to study. Second, the disease presents as a clinical spectrum that correlates with the level of the immune response to the pathogen 16 . Patients with the tuberculoid form are relatively re- sistant to the pathogen; the infection is localized and the lesions are characterized by expression of the type-1 cytokines charac- teristic of cell-mediated immunity 17,18 . In contrast, patients with lepromatous leprosy are relatively susceptible to the pathogen; the infection is systemically disseminated and the lesions are characterized by the type-2 cytokines characteristic of humoral responses. To gain insight into the innate immune response in human disease, we investigated the expression and activation of TLRs in human leprosy. TLR2 and TLR2-TLR1 mediate activation by M. leprae To determine the role of TLRs in mediating cell activation by M. leprae, we used human cell lines transiently expressing TLR ho- modimers (TLR1–10) or heterodimers (TLR2-TLR1, TLR2-TLR6, TLR2-TLR10) with an endothelial leukocyte adhesion mole- cule–luciferase (ELAM.luc) reporter. These cells were activated Activation and regulation of Toll-like receptors 2 and 1 in human leprosy STEPHAN R. KRUTZIK 1,2 , MARIA TERESA OCHOA 1 , PETER A. SIELING 1 , SATOSHI UEMATSU 3 , YOLANDA W. NG 1 , ANNALIZA LEGASPI 1 , PHILIP T. LIU 1,2 , STEWART T. COLE 4 , PAUL J. GODOWSKI 5 , YUMI MAEDA 6,9 , EUZENIR N. SARNO 7 , MICHAEL V. NORGARD 8 , PATRICK J. BRENNAN 9 , SHIZUO AKIRA 3 , THOMAS H. REA 10 & ROBERT L. MODLIN 1,2 1 Division of Dermatology and 2 Department of Microbiology and Immunology, David Geffen School of Medicine at University of California Los Angeles, Los Angeles, California, USA 3 Department of Host Defense, Research Institute for Microbial Diseases, Osaka University, Japan Science and Technology Corporation, Osaka, Japan 4 Unité de Génétique Moléculaire Bactérienne, Institut Pasteur, Paris, France 5 Department of Molecular Biology, Genentech Inc., South San Francisco, California, USA 6 Department of Microbiology, Leprosy Research Center, National Institute of Infectious Diseases, Higashimurayama, Tokyo, Japan 7 Leprosy Laboratory Institute Oswaldo Cruz, Rio de Janeiro, Brasil 8 Department of Microbiology, University of Texas Southwestern Medical Center, Dallas, Texas, USA 9 Department of Microbiology, Colorado State University, Fort Collins, Colorado, USA 10 Section of Dermatology, University of Southern California School of Medicine, Los Angeles, California, USA Correspondence should be addressed to R.L.M.; e-mail: [email protected] Published online 14 April 2003; doi:10.1038/nm864 The expression and activation of Toll-like receptors (TLRs) was investigated in leprosy, a spectral disease in which clinical manifestations correlate with the type of immune response mounted to- ward Mycobacterium leprae. TLR2-TLR1 heterodimers mediated cell activation by killed M. leprae, indicating the presence of triacylated lipoproteins. A genome-wide scan of M. leprae detected 31 putative lipoproteins. Synthetic lipopeptides representing the 19-kD and 33-kD lipoproteins ac- tivated both monocytes and dendritic cells. Activation was enhanced by type-1 cytokines and in- hibited by type-2 cytokines. In addition, interferon (IFN)-γ and granulocyte-macrophage colony-stimulating factor (GM-CSF) enhanced TLR1 expression in monocytes and dendritic cells, respectively, whereas IL-4 downregulated TLR2 expression. TLR2 and TLR1 were more strongly expressed in lesions from the localized tuberculoid form (T-lep) as compared with the dissemi- nated lepromatous form (L-lep) of the disease. These data provide evidence that regulated ex- pression and activation of TLRs at the site of disease contribute to the host defense against microbial pathogens. © 2003 Nature Publishing Group http://www.nature.com/naturemedicine
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Activation and regulation of Toll-like receptors 2 and 1 in human leprosy

NATURE MEDICINE • VOLUME 9 • NUMBER 5 • MAY 2003 525
ARTICLES
Toll-like receptors (TLRs) represent one mechanism by whichthe innate immune system recognizes biochemical patterns dis-played by infectious invaders. The specificity of TLRs in mediat-ing responses to defined bacterial ligands has been clearlyshown: TLR2 homodimers1,2 and TLR2-TLR1 heterodimers medi-ate the response to microbial triacylated lipoproteins3,4, TLR3 todouble-stranded viral RNA5, TLR4 to lipopolysaccharide (LPS)6–8,TLR5 to bacterial flagellin9, TLR2-TLR6 to diacylated lipopep-tides10, TLR7 and TLR8 to imidazoquinolines11,12 and TLR9 tobacterial CpG DNA sequences13. Upon activation, TLRs triggerthe release of cytokines and the induction of co-stimulatorymolecules that can influence the nature of the adaptive T- or B-cell response14. Activation of TLRs also induces antimicrobialpathways that kill intracellular organisms15.
Leprosy provides an excellent opportunity to investigatemechanisms of innate and adaptive immunity in humans. First,leprosy primarily affects skin, so the lesions are readily accessibleto study. Second, the disease presents as a clinical spectrum that
correlates with the level of the immune response to thepathogen16. Patients with the tuberculoid form are relatively re-sistant to the pathogen; the infection is localized and the lesionsare characterized by expression of the type-1 cytokines charac-teristic of cell-mediated immunity17,18. In contrast, patients withlepromatous leprosy are relatively susceptible to the pathogen;the infection is systemically disseminated and the lesions arecharacterized by the type-2 cytokines characteristic of humoralresponses. To gain insight into the innate immune response inhuman disease, we investigated the expression and activation ofTLRs in human leprosy.
TLR2 and TLR2-TLR1 mediate activation by M. lepraeTo determine the role of TLRs in mediating cell activation by M.leprae, we used human cell lines transiently expressing TLR ho-modimers (TLR1–10) or heterodimers (TLR2-TLR1, TLR2-TLR6,TLR2-TLR10) with an endothelial leukocyte adhesion mole-cule–luciferase (ELAM.luc) reporter. These cells were activated
Activation and regulation of Toll-like receptors 2 and 1in human leprosy
STEPHAN R. KRUTZIK1,2, MARIA TERESA OCHOA1, PETER A. SIELING1, SATOSHI UEMATSU3,YOLANDA W. NG1, ANNALIZA LEGASPI1, PHILIP T. LIU1,2, STEWART T. COLE4,
PAUL J. GODOWSKI5, YUMI MAEDA6,9, EUZENIR N. SARNO7, MICHAEL V. NORGARD8, PATRICK J. BRENNAN9, SHIZUO AKIRA3, THOMAS H. REA10 & ROBERT L. MODLIN1,2
1Division of Dermatology and 2Department of Microbiology and Immunology, David Geffen School of Medicineat University of California Los Angeles, Los Angeles, California, USA
3Department of Host Defense, Research Institute for Microbial Diseases, Osaka University,Japan Science and Technology Corporation, Osaka, Japan
4Unité de Génétique Moléculaire Bactérienne, Institut Pasteur, Paris, France5Department of Molecular Biology, Genentech Inc., South San Francisco, California, USA
6Department of Microbiology, Leprosy Research Center, National Institute of Infectious Diseases, Higashimurayama, Tokyo, Japan
7Leprosy Laboratory Institute Oswaldo Cruz, Rio de Janeiro, Brasil8Department of Microbiology, University of Texas Southwestern Medical Center, Dallas, Texas, USA
9Department of Microbiology, Colorado State University, Fort Collins, Colorado, USA10Section of Dermatology, University of Southern California School of Medicine, Los Angeles, California, USA
Correspondence should be addressed to R.L.M.; e-mail: [email protected]
Published online 14 April 2003; doi:10.1038/nm864
The expression and activation of Toll-like receptors (TLRs) was investigated in leprosy, a spectraldisease in which clinical manifestations correlate with the type of immune response mounted to-ward Mycobacterium leprae. TLR2-TLR1 heterodimers mediated cell activation by killed M. leprae,indicating the presence of triacylated lipoproteins. A genome-wide scan of M. leprae detected 31putative lipoproteins. Synthetic lipopeptides representing the 19-kD and 33-kD lipoproteins ac-tivated both monocytes and dendritic cells. Activation was enhanced by type-1 cytokines and in-hibited by type-2 cytokines. In addition, interferon (IFN)-γ and granulocyte-macrophagecolony-stimulating factor (GM-CSF) enhanced TLR1 expression in monocytes and dendritic cells,respectively, whereas IL-4 downregulated TLR2 expression. TLR2 and TLR1 were more stronglyexpressed in lesions from the localized tuberculoid form (T-lep) as compared with the dissemi-nated lepromatous form (L-lep) of the disease. These data provide evidence that regulated ex-pression and activation of TLRs at the site of disease contribute to the host defense againstmicrobial pathogens.
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
rem
edic
ine

526 NATURE MEDICINE • VOLUME 9 • NUMBER 5 • MAY 2003
ARTICLES
with killed M. leprae and luciferase activity was measured.Among the homodimers tested, only TLR2 was able to mediateresponsiveness toward M. leprae, activating the cells by 28-foldabove background (Fig. 1a). Co-expression of TLR1 greatly en-hanced TLR2-mediated activity, resulting in 215-fold activation(Fig. 1a). These data indicate that TLR2 homodimers or TLR2-TLR1 heterodimers can mediate cell activation by M. leprae.Studies of Tlr2–/– and Tlr1–/– mice corroborate the transfectiondata. Peritoneal macrophages from Tlr2–/– mice did not respondto M. leprae, as measured by TNF-α production, whereasmacrophages from Tlr1–/– mice had a diminished response (Fig.1b). In accordance with previous studies, macrophages fromTlr2–/– mice did not respond to Pam3Cys or macrophage-activat-ing lipopeptide (MALP)-2. Macrophages from Tlr1–/– and Tlr6–/–
mice had diminished responses to Pam3Cys and MALP-2, respec-tively (Fig. 1b).
Two groups recently showed that TLR2-TLR1 heterodimersmediate the response to triacylated lipopeptides3,4. It was thuslogical to investigate M. leprae lipoproteins and synthesize corre-sponding triacylated lipopeptides as a model for M. leprae lig-ands that stimulate TLR2 and TLR1. Unfortunately, M. lepraecannot be grown in the laboratory and it is difficult to purifyproteins from the small number of organisms that can be har-vested from armadillos. We therefore scanned the M. lepraegenome for proteins with a putative lipoprotein signal motif andidentified 31 putative lipoproteins (see Supplementary Table 1online). The M. leprae lipoproteins ML1966 and ML0603 werechosen for further study.
ML1966 (a putative M. leprae 19-kD lipoprotein) shares 47%amino acid sequence identity with the 19-kD lipoprotein ofMycobacterium tuberculosis (lpqH), a major inducer of monocytecytokine release through TLR2 (refs. 1, 3). ML1966 seems to have
arisen by means of a gene duplication event as its gene is locatedin a different site than that occupied by lpqH (Rv3763) of M. tu-berculosis. The M. leprae chromosomal locus corresponding toRv3763 contains the pseudogene ML2363, which is strictly or-thologous to lpqH, and is flanked by pseudogenes equivalent toRv3762 and Rv3764, the neighboring genes in M. tuberculosis.ML1966 mRNA has been detected in M. leprae (D. Williams andT. Gillis, personal communication). ML0603, the M. leprae 33-kDlipoprotein, was also studied because expression of this lipopro-tein by M. leprae has been detected by western blot, and becausea recombinant lipoprotein expressed in Escherichia coli triggersmonocyte cytokine release19.
Analysis by GC mass spectrometry of mycobacterial lipopro-teins provided evidence for the presence of three fatty acid sub-stitutions: two in the glycerol portion (consisting of eitherpalmitic, stearic or tuberculostearic acid) and one in the amidelinkage to the cysteine residue (palmitic acid). Although the nat-ural lipoproteins from M. leprae may contain a combination ofthree different fatty acids, tri-palmitylated lipopeptides activatea similar pattern of TLRs when compared with a purified M. tu-berculosis lipoprotein3. Therefore, hexameric peptides with a tri-palmityl modification corresponding to the M. leprae 19-kD and33-kD lipoproteins were synthesized and examined for theirability to trigger monocytes in a TLR-dependent manner.
Both M. leprae lipopeptides activated primary human mono-cytes obtained from healthy donors, as measured by the release ofIL-12 p40 (Fig. 1c). A TLR2-neutralizing antibody blocked activa-tion. Similarly, the 19-kD and 33-kD lipopeptides activatedmonocyte-derived dendritic cells in a TLR2-dependent manner torelease IL-12 p40 (Fig. 1d). Together, these data provide evidencethat M. leprae and two of its constituent ligands activate cells ofthe innate immune system through TLR2 and TLR2-TLR1.
0
50
100
150
200
250
2/1 2/6 2/10
Fo
ld in
du
ctio
n
TLRs
0
50
100
150
200
250
– 1 2 3 4 5 6 7 8 9 10
Fo
ld in
du
ctio
n
TLR
a
0
200
400
600
800
1,000
1,200
0 100 1,000 10,000
TN
F-α
(p
g/m
l)
0
500
1,000
1,500
2,000
2,500
TN
F-α
(p
g/m
l)
– LPSMALP-2Pam3Cys
M. leprae (ng/ml)
b
IL-1
2 p
40 (
pg
/ml)
0
500
1,000
1,500
2,000
2,500
3,000
3,500
IL-1
2 p
40 (
pg
/ml)
Media – αTLR2 IgG1
19-kD
0
200
400
600
800
1,000
Media – αTLR2 IgG1
19-kD
c
0
500
1,000
1,500
2,000
2,500
3,000
IL-1
2 (p
g/m
l)
0
50
100
150
200
250
300
Media – αTLR2 IgG1
IL-1
2 (p
g/m
l)
33-kD
Media – αTLR2 IgG1
33-kD
d
Fig. 1 TLR2 and TLR2-TLR1 mediate the response to M. leprae and M. lep-rae 19-kD and 33-kD lipopeptides. a, HEK 293 cells transiently expressingTLR homodimers (TLR1–10) or heterodimers (TLR2/1, 2/6 or 2/10) and anELAM.luc plasmid were stimulated with killed M. leprae. Cells transfectedwith TLR4 were also transfected with MD2 (a TLR-4–associated protein). b, Left, peritoneal macrophages from wild-type (�), Tlr1–/– (�), Tlr2–/– (�)or Tlr6–/– (�) mice were activated with either medium or killed M. leprae.Right, peritoneal macrophages from wild-type (�), Tlr1–/– (�), Tlr2–/– (�), or
Tlr6–/– (�) were also activated with either media, Pam3Cys, MALP-2 or LPSto serve as controls. TNF-α production was measured by ELISA. c and d,Primary human monocytes (left) or monocyte-derived dendritic cells (right)were stimulated with the M. leprae 19-kD (c) or 33-kD (d) lipopeptides inthe presence of medium, a TLR2-blocking antibody (αTLR2) or an IgG1control antibody. IL-12 p40 production was measured by ELISA. The dataare representative of at least 2 independent experiments. Error bars repre-sent s.e.m.
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
rem
edic
ine

NATURE MEDICINE • VOLUME 9 • NUMBER 5 • MAY 2003 527
ARTICLES
Regulation of TLR2-TLR1 activation by cytokinesFor intracellular pathogens, including M. leprae, the balance oftype-1 and type-2 cytokines influences whether the clinical formof the disease is localized or disseminated, respectively. In lep-rosy, the type-1 cytokines IFN-γ, IL-12, IL-18, and GM-CSF pre-dominate in localized T-lep lesions, whereas the type-2cytokines IL-4 and IL-10 characterize disseminated L-lep le-sions17,18. We investigated whether these cytokines could influ-ence TLR2-TLR1 activation of monocytes and monocyte-deriveddendritic cells by stimulating these cells with the M. leprae 19-kDlipopeptide in the presence or absence of cytokines, and thenmeasuring cytokine release. The type-1 cytokines IFN-γ, IL-12,IL-18 and GM-CSF enhanced the ability of the M. leprae 19-kDlipopeptide to trigger monocyte release of TNF-α (Fig. 2a). Incontrast, IL-4 and IL-10 substantially inhibited the lipopeptide-induced release of TNF-α. IFN-γ also upregulated lipopeptide-induced release of IL-12 p40 from monocyte-derived dendriticcells (Fig. 2b). IL-18 had little effect, whereas IL-4 and IL-10 in-hibited IL-12 p40 release. In the absence of the 19-kD lipopep-tide, the type-1 and type-2 cytokines did not stimulate
monocyte and monocyte-derived dendritic-cell cytokine release(data not shown). These data provide evidence that the localtype-1 or type-2 cytokine environment can influence the degreeof TLR2-TLR1 activation.
Type-1 and type-2 cytokines influence TLR expressionWe next wanted to address whether type-1 and type-2 cytokine
in T-leplesions
Cytokines prominent
0.00 0.50 1.0 5.0 10 15
Change in TNF- α (ng/ml)
19-kD
+ IFN-γ+ IL-12
+ IL-18
+ GM-CSF
+ IL-4
+ IL-10
+ media
2.0
in L-leplesions
Cytokines prominent
a
0 500 1,000 1,500 2,000 2,500 3,000
+ IL-10
+ IL-4
+ IL-18
+ IFN-γ
+ media19-kD
Change in IL-12 (pg/ml)
in T-leplesions
Cytokines prominent
in L-leplesions
Cytokines prominent
b
Fig. 2 Cytokine profiles influence TLR2-TLR1 activation of monocyte andmonocyte-derived dendritic cells. a and b, Monocytes (a) or monocyte-derived dendritic cells (b) were activated with M. leprae 19-kD lipopeptidetogether with either media, IFN-γ, IL-12, IL-18, GM-CSF, IL-4 or IL-10. TNF-α (a) or IL-12 p40 (b) production was measured by ELISA. The data areshown as TNF-α or IL-12 p40 levels above media background and are rep-resentative of 3 independent experiments.
TLR2
Media IL-12 IL-18
161 127181
GM-CSF
145
126 115 81 82 88
101
IFN-γ
Monocytes
Dendriticcells
Cel
l num
ber
Cel
l num
ber
a IL-4 IL-10
34 178
39 115
Media
161
126
TLR2
Monocytes
Dendriticcells
Cel
l num
ber
Cel
l num
ber
b
TLR1
Media IL-12 IL-18 GM-CSFIFN-γ
Monocytes
Dendriticcells
<1 9 <1 <1<1
<1 <1 <1 7<1
Cel
l num
ber
Cel
l num
ber
c IL-4 IL-10Media
TLR1
Monocytes
Dendriticcells
<1 <1 7
<1 <1 <1
Cel
l num
ber
Cel
l num
ber
d
Fig. 3 Effect of cytokines on TLR2 and TLR1 expression on monocytes andmonocyte-derived dendritic cells. a, Monocytes and monocyte-derived den-dritic cells were treated with media or recombinant type-1 cytokines IFN-γ, IL-12, IL-18 or GM-CSF. Cells were labeled with a TLR2-specific monoclonalantibody or an isotype control and examined by flow cytometry.b, Monocytes and monocyte-derived dendritic cells were treated with mediaor the recombinant type-2 cytokines IL-4 or IL-10. TLR2 cell-surface expressionwas examined by flow cytometry. a and b, the data shown are representativeof at least 2 independent experiments; values at upper right of each graph rep-
resent changes in mean fluorescence intensity between TLR2 and the IgG1 iso-type control. c, Monocytes and monocyte-derived dendritic cells were treatedwith media, IFN-γ, IL-12, IL-18 or GM-CSF. Cells were labeled with a TLR1-spe-cific monoclonal antibody or an isotype control and examined by flow cytom-etry. d, Monocytes and monocyte-derived dendritic cells were treated withmedia, IL-4 or IL-10. TLR1 cell-surface expression was examined by flow cy-tometry. c and d, the data shown are representative of at least 2 independentexperiments; values at upper right of each graph represent changes in meanfluorescence intensity between TLR1 and the IgG1 isotype control.
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
rem
edic
ine
528 NATURE MEDICINE • VOLUME 9 • NUMBER 5 • MAY 2003
ARTICLES
patterns could influence the level of TLR2 and TLR1 expressionon monocytes and monocyte-derived dendritic cells. Cells weretreated with recombinant cytokines and TLR expression wasmeasured by flow cytometry. The type-1 cytokines IFN-γ, IL-12,IL-18, and GM-CSF did not affect TLR2 expression (Fig. 3a). IL-10, which inhibited TLR2-TLR1 activation, did not downregu-late TLR2 expression. The type-2 cytokine IL-4, however,substantially decreased TLR2 cell surface expression on bothmonocytes and monocyte-derived dendritic cells (Fig. 3b). As
previously shown20, IL-4 downregulates TLR2 mRNA as mea-sured by quantitative PCR (data not shown).
In contrast to the downregulation of TLR2 by type-2 cy-tokines, TLR1 expression was upregulated by type-1 cytokines.Freshly isolated monocytes expressed undetectable to low levelsof TLR1, but this expression was increased through the additionof the type-1 cytokine IFN-γ (Fig. 3c). The other type-1 cytokines,IL-12, IL-18, and GM-CSF, as well as the type-2 cytokines testeddid not influence levels of TLR1 expression. In monocyte-
derived dendritic cells, the type-1 cytokine GM-CSFinduced expression of TLR1 (Fig. 3c). The type 2 cy-tokine IL-10 seemed to have a modest effect on TLR1expression in dendritic cells, perhaps from driving thedendritic cell towards a more macrophage-like cell(Fig. 3d). Taken together, our data indicate that type-1and type-2 cytokine patterns differentially regulateTLR1 and TLR2 expression, respectively.
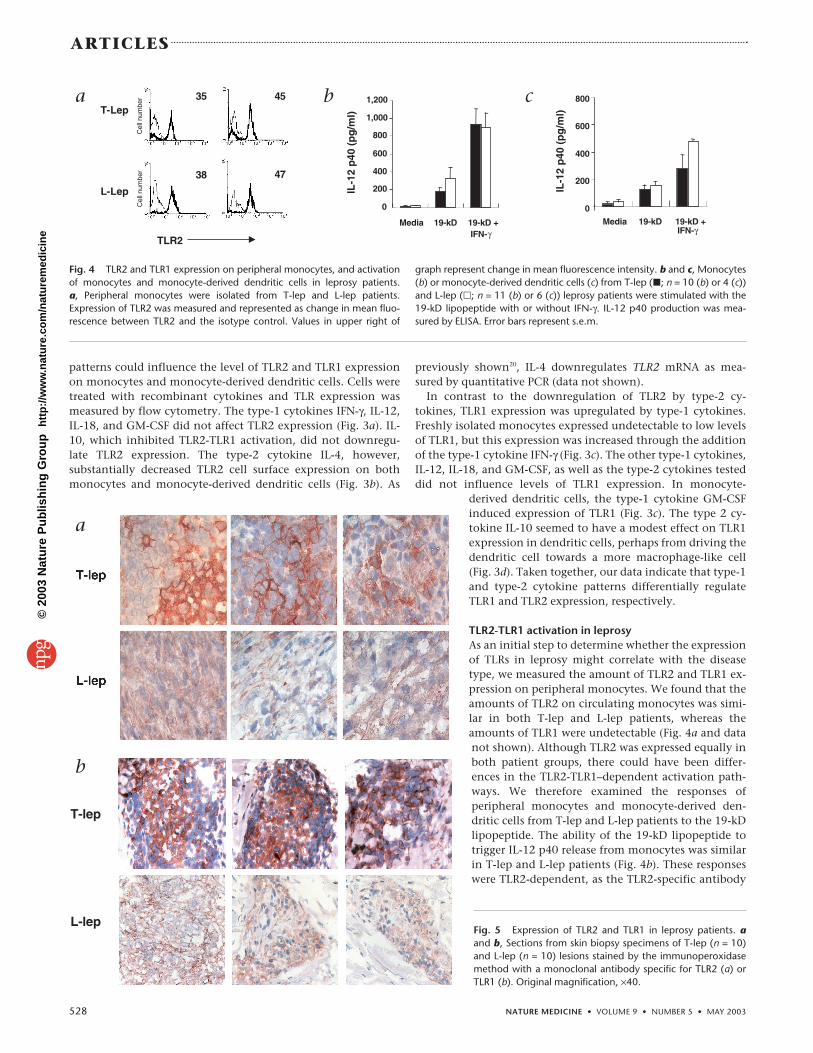
TLR2-TLR1 activation in leprosyAs an initial step to determine whether the expressionof TLRs in leprosy might correlate with the diseasetype, we measured the amount of TLR2 and TLR1 ex-pression on peripheral monocytes. We found that theamounts of TLR2 on circulating monocytes was simi-lar in both T-lep and L-lep patients, whereas theamounts of TLR1 were undetectable (Fig. 4a and datanot shown). Although TLR2 was expressed equally inboth patient groups, there could have been differ-ences in the TLR2-TLR1–dependent activation path-ways. We therefore examined the responses ofperipheral monocytes and monocyte-derived den-dritic cells from T-lep and L-lep patients to the 19-kDlipopeptide. The ability of the 19-kD lipopeptide totrigger IL-12 p40 release from monocytes was similarin T-lep and L-lep patients (Fig. 4b). These responseswere TLR2-dependent, as the TLR2-specific antibody
T-Lep
L-Lep
TLR2
35 45
38 47
Cel
l num
ber
Cel
l num
ber
a
0
200
400
600
800
1,000
1,200
Media 19-kD 19-kD +IFN-γ
IL-1
2 p
40 (
pg
/ml)
b
0
200
400
600
800
Media 19-kD 19-kD +IFN-γ
IL-1
2 p
40 (
pg
/ml)
c
Fig. 4 TLR2 and TLR1 expression on peripheral monocytes, and activationof monocytes and monocyte-derived dendritic cells in leprosy patients. a, Peripheral monocytes were isolated from T-lep and L-lep patients.Expression of TLR2 was measured and represented as change in mean fluo-rescence between TLR2 and the isotype control. Values in upper right of
graph represent change in mean fluorescence intensity. b and c, Monocytes(b) or monocyte-derived dendritic cells (c) from T-lep (�; n = 10 (b) or 4 (c))and L-lep (�; n = 11 (b) or 6 (c)) leprosy patients were stimulated with the19-kD lipopeptide with or without IFN-γ. IL-12 p40 production was mea-sured by ELISA. Error bars represent s.e.m.
a
T-lep
L-lep
b
Fig. 5 Expression of TLR2 and TLR1 in leprosy patients. aand b, Sections from skin biopsy specimens of T-lep (n = 10)and L-lep (n = 10) lesions stained by the immunoperoxidasemethod with a monoclonal antibody specific for TLR2 (a) orTLR1 (b). Original magnification, ×40.
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
rem
edic
ine

NATURE MEDICINE • VOLUME 9 • NUMBER 5 • MAY 2003 529
ARTICLES
blocked the response by 80% (data not shown). The19-kD lipopeptide also induced similar levels of IL-12p40 release from monocyte-derived dendritic cellsfrom T-lep patients as compared with L-lep patients(Fig. 4c). These data indicate that in circulating blood,both T-lep and L-lep patients can mount a functionallyequivalent TLR2-TLR1 response.
Expression of TLR2 and TLR1 in leprosy lesionsThe battle between host immune response and micro-bial pathogens in leprosy is enjoined in the skin lesion,the site of disease activity. We wanted to determinewhether the expression of TLR2 and TLR1 in the le-sions of leprosy patients correlates with the local cy-tokine pattern characteristic of the different forms ofthe disease. We assessed the frequency and distributionof TLR2- and TLR1-expressing cells in biopsy speci-mens from patients with leprosy using immunoperoxi-dase labeling. Notable differences were observed at thelevel of both TLR2 and TLR1 expression. TLR2 wasstrongly expressed in T-lep lesions on large ovoid cellsand on cells with a dendritic morphology (Fig. 5a). Often T-lep lesions studied, approximately 40–50% of thecells infiltrating the granulomas were positive forTLR2. In contrast, TLR2 was only weakly expressed oncells in ten L-lep lesions. Similarly, TLR1 was muchmore strongly expressed in T-lep lesions as comparedwith L-lep lesions (Fig. 5b).
We used double immune labeling and confocal lasermicroscopy to examine the nature of the cells express-ing TLR2 and TLR1 in T-lep lesions (Fig. 6).Approximately 90–95% of TLR2-expressing cells ex-pressed both CD14 and CD68, indicating expression ofTLR2 on cells of the monocyte/macrophage lineage.An additional 5% of cells expressed both TLR2 andCD1a, indicating expression of TLR2 on dendritic cells in theselesions. Expression of TLR1 co-localized with TLR2, indicatingthat most cells expressed the TLR2-TLR1 heterodimer. Finally,TLR2-positive cells did not express the T-cell marker CD3.Together, these findings indicate that the frequency of mono-cytes and dendritic cells expressing TLR2 and TLR1 in leprosy le-sions is higher in T-lep patients, the group with localizedinfection and expression of type-1 cytokines in lesions, as com-pared with L-lep patients, the group with disseminated infectionand expression of type-2 cytokines in lesions.
DiscussionThe discovery of mammalian TLRs has provided new insightinto mechanisms of innate immunity to microbial pathogens.Activation of TLRs leads to the induction of antimicrobial path-ways central to innate defense15 as well as the upregulation ofantigen presentation molecules and secretion of cytokines thatinfluence the nature of the adaptive immune response14. Here,we undertook a comprehensive and integrated analysis of TLR
responsiveness, regulation and distribution in human disease,by analyzing human leprosy. We focused on TLR2 and TLR1 be-cause the bacteria that causes leprosy, M. leprae, was found to ac-tivate either TLR2 homodimers or TLR2-TLR1 heterodimers. Inaddition, scanning of the M. leprae genome revealed 31 lipopro-teins that could serve as pathogen-associated molecular patternsto be recognized by the innate immune system. Syntheticlipopeptides representing putative 19-kD and 33-kD lipoproteinsactivated monocytes and monocyte-derived dendritic cellsthrough TLR2. We found that expression and activation of TLR2and TLR1 were regulated by type-1 and type-2 cytokine patterns.Analysis of lesions provided in vivo evidence that the local cy-tokine environment regulates the expression of TLR2 and TLR1,thus determining the outcome of the innate immune responseagainst microbial infection.
Through our study of human leprosy, we correlated TLR2 andTLR1 expression in disease lesions with resistance and suscepti-bility to a human pathogen. Whereas TLR2 expression on pe-ripheral monocytes of patients from across the spectrum of
Fig. 6 Phenotype of cells expressing TLR2 in T-lep lesions.Two-color immunofluorescence confocal images were obtainedfor the markers CD14, CD68, CD1a, TLR1 or CD3 (green; left)and TLR2 (red; center). The 2 images were then superimposed(yellow; right). TLR2-positive cells in the T-lep lesions were posi-tive for CD14, CD68, CD1a and TLR1, but not CD3. Originalmagnification, ×63.
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
rem
edic
ine

530 NATURE MEDICINE • VOLUME 9 • NUMBER 5 • MAY 2003
ARTICLES
leprosy was equivalent, we observed differences at the sites ofdisease in the skin lesions obtained from patients. TLR2 wasstrongly expressed on monocytes and dendritic cells in lesionsfrom T-lep patients, which are self-healing and characterized byresistance to growth of M. leprae. In contrast, TLR2 was weaklyexpressed on monocytes in L-lep lesions, which are progressiveand characterized by enormous quantities of the organism.Similarly, we found much higher TLR1 expression in T-lep le-sions compared with L-lep lesions. Previous studies of leprosy le-sions indicates that the local expression of the type-1 cytokinesIFN-γ, IL-12, IL-18 and GM-CSF predominates in T-lep lesions,whereas the type-2 cytokines IL-4 and IL-10 characterize L-lep le-sions17,18,21,22. Our data corroborate this by showing that IL-4downregulates TLR2 mRNA and protein expression on mono-cytes20,23 and also downregulates TLR2 on dendritic cells. We hy-pothesize that the strong expression of the type-2 cytokine IL-4in L-lep lesions accounts for the diminished expression of TLR2.In contrast, the type-1 cytokines IFN-γ and GM-CSF upregulatelevels of TLR1 expression in vitro, suggesting that the high levelsof these cytokines in T-lep lesions accounts for the higher levelsof TLR1 expression. These findings indicate that in humans, theregulated expression of TLR2 and TLR1 contributes to the out-come between the host response and the microbial invader.These results also suggest that effects of the local cytokine envi-ronment on the expression of TLR2-TLR1 heterodimers is a pos-sible mechanism by which some individuals are hyporesponsiveto vaccination with the Borrelia burgdorferi OspA lipopeptide4.
A notable finding of the present study was that type-1 andtype-2 cytokine patterns differentially affected TLR2 and TLR1activation by an M. leprae lipopeptide. The type-1 cytokines IFN-γ, GM-CSF, IL-12 and IL-18 enhanced TLR2 and TLR1 activation,whereas the type-2 cytokines IL-4 and IL-10 inhibited activation.It is noteworthy that IL-10 inhibited TLR2 and TLR1 activationbut did not downregulate TLR2 or TLR1 expression as comparedwith IL-4, which inhibited both TLR2 expression and TLR2-TLR1heterodimer activation. The type-1 cytokines IFN-γ, IL-12 and IL-18 upregulated TLR2-TLR1 heterodimer responses without af-fecting TLR2 expression. The type-1 cytokines IFN-γ andGM-CSF, however, increased TLR1 expression in monocytes andmonocyte-derived dendritic cells, respectively. Thus, the localcytokine pattern triggered during microbial infection regulatesthe innate immune system through two different and indepen-dent mechanisms, one directly affecting TLR activation and theother modulating TLR expression. It is well known that the in-nate immune response serves an instructional role in shapingthe adaptive immune response. Our data indicate that the adap-tive immune response, by releasing cytokines from T cells thatregulate TLR activation, can also influence the magnitude of theinnate immune response.
The ability of the innate immune system to recognize andrapidly respond to microbial pathogens is crucial to the host de-fense against infection. The Toll family of receptors seems to beintegral in this role, given that it has been conserved throughouthundreds of millions of years of evolution, and is found in bothinsects and mammals. In Drosophila, mutations in Toll or down-stream signaling molecules substantially increase susceptibilityto fungal and bacterial infections. Mice with spontaneous or tar-geted mutations in TLRs are more susceptible to bacterial infec-tion, further implicating TLRs as critical receptors in mammalianhost defense24–29. Recent studies also indicate that TLRs are re-quired for optimal induction of innate immunity in mouse mod-els of microbial infection30,31. Although genetic analysis of TLR2
in L-lep patients detected a polymorphism in the cytoplasmicsignaling domain of the TLR2 protein32, we found that there wasno difference in the ability of the TLR2-TLR1 ligand to trigger IL-12 p40 release from peripheral blood monocytes obtained fromL-lep as compared with T-lep patients. Further experiments arewarranted to determine whether such polymorphisms affect TLRactivation pathways.
We found that M. leprae activates cells through TLR2 andTLR2-TLR1. Of the potential ligands that activate TLR2-TLR1,microbial lipoproteins have long been known to be potent im-munostimulatory molecules. Experiments have shown thatTLR2 mediates responsiveness to lipoproteins and lipopep-tides1,33. Additional studies have shown that triacylatedlipoproteins and lipopeptides activate a TLR2-TLR1 het-erodimer3,4. Using a genome-wide scan, 31 putative lipoproteingenes were identified in the annotated genome sequence of M.leprae34, together with 3 genes encoding enzyme systems re-quired for processing the lipoprotein precursor and attachingthe lipid group. All of these genes have orthologs in M. tubercu-losis35, which also contains about 60 additional genes forlipoproteins that have either been lost by the leprosy bacillusor retained in inactive form as pseudogenes. Among the lattergroup is ML1966, a gene orthologous to lpqH (Rv3763), whichencodes the well-characterized 19-kD lipoprotein antigen ofM. tuberculosis36,37. A synthetic M. leprae 19-kD lipopeptide in-duced IL-12 p40 release from human monocytes and mono-cyte-derived dendritic cells through TLR2. Similarly, a 33-kDlipoprotein known to be expressed by M. leprae19 was studiedand found to activate monocytes and monocyte-derived den-dritic cells through TLR2. Because our data suggest a promi-nent role for TLR2 and TLR1 in mediating the response toM. leprae, we cannot rule out other TLR ligands such as lipoara-binomannan and peptidoglycan activating through TLR2 (refs.38, 39). In addition, although the response through TLR4 wasweak using the available killed M. leprae, previous studies sug-gest that live mycobacteria can activate TLR4 (ref. 38). Furtherinvestigation to identify the TLR4 ligand(s) in live M. tuberculo-sis should clarify this issue.
Our data suggest that the outcome of the immune responsein leprosy is determined in part by the influence of the localcytokine environment on the regulated expression and activa-tion of TLR2 and TLR1. Although TLR activation is a centralpart of innate immunity, TLR activation can also lead to im-munopathology and tissue damage, as in the apoptosis and in-flammation-mediated tissue injury seen in septic shock. Evenin the T-lep form of leprosy, where the level of TLR activationis strong and the pathogen is contained, inflammation-medi-ated nerve injury is a prominent feature. It should be possibleto design therapeutic agents that regulate TLR expression andactivation. Although such interventions could be used as anadjuvant for immune responses for vaccines or to treat activeinfection, the potential consequence of tissue injury requiresscrutiny.
MethodsTLR ligands. Killed M. leprae was obtained from P. Brennan (Colorado StateUniversity, Fort Collins, Colorado). M. leprae hexameric peptides with a tri-palmityl modification corresponding to 19-kD and 33-kD lipoproteins fromM. leprae (Pam3CSGGTQ and Pam3CLPAWK, respectively) and peptideslacking the tri-palmityl modification were synthesized (Norgard andBachem, Torrance, California). Salmonella minnesota LPS (Sigma, St. Louis,Missouri) was purified as described40. MALP-2 and Pam3Cys-Ser-(Lys)4
(CSK4) were used as described2,41.
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
rem
edic
ine

NATURE MEDICINE • VOLUME 9 • NUMBER 5 • MAY 2003 531
ARTICLES
Patients and clinical specimens. Patients with leprosy were classified ac-cording to the criteria of Ridley and Jopling16. Scalpel or punch skinbiopsy specimens (6 mm diameter) were obtained after informed consent(Institutional Review Board, University of Southern California School ofMedicine) from 10 T-lep and 10 L-lep leprosy patients at the time of diag-nosis. Specimens were embedded in OCT medium (Ames, Elkhart,Indiana), snap-frozen in liquid nitrogen and stored at –70 °C until sec-tioning.
Antibodies and cytokines. We used antibodies against TLR2 (2392;Genentech, San Francisco, California), TLR1 (eBioscience, San Diego,California), CD14 (Zymed, San Francisco, California), CD1a (OKT6 IgG1;ATCC, Manassas, Virginia), CD3 (OKT3 IgG1; ATCC), CD68 (Y1/82A; BDPharmingen, San Diego, California) and IgG controls (Sigma). The follow-ing cytokines were used: IFN-γ (Endogen, Woburn, Massachusetts), IL-4(Peprotech, Rocky Hill, New Jersey), IL-10 (BD Pharmingen), IL-12 (BDPharmingen), IL-18 (R&D Systems, Minneapolis, Minnesota) and GM-CSF(Immunex, Seattle, Washington).
Monocytes, dendritic cells and ELISA. Monocytes were isolated by adher-ence in 1% FCS as described1. For dendritic cells, adherent cells were cul-tured in RPMI with 1,000 U/ml IL-4 and 750 U/ml GM-CSF and matured for7 d. Monocytes and dendritic cells were stimulated with medium, killed M.leprae, or M. leprae 19-kD or 33-kD lipopeptides in the presence or absenceof IFN-γ (100 U/ml), IL-12 (100 ng/ml), IL-18 (100 ng/ml), GMCSF (800U/ml), IL-4 (1,000 U/ml) or IL-10 (100 U/ml). Antibody blocking was per-formed as described1. IL-12 p40 or TNF-α production was measured byELISA after 24 h (BD Pharmingen).
Bacterial genomics. During the annotation of the M. leprae genome se-quence34, lipoproteins were identified by the presence of an appropriatelypositioned PROSITE motif (PS00013 prokaryotic membrane lipoproteinlipid attachment site42) following a signal peptide, allowing no mismatches.Lipoprotein gene and protein sequences were obtained from the Lepromadatabase (http://genolist.pasteur.fr/Leproma/).
Transfection and reporter assays. Human embryonic kidney 293 cellswere transiently transfected as described3. Twenty-four hours after transfec-tion, cells were stimulated with M. leprae for 8 h. Cells were lysed and lu-ciferase activity was measured as described3.
Mouse peritoneal macrophages and ELISA. Tlr1–/–, Tlr2–/–, and Tlr6–/– micewere generated as described3,8,10. Isolation of peritoneal macrophages wasdone as described15. Macrophages (105) were stimulated for 24 h. TNF-α(Genzyme Techne, Minneapolis, Minnesota) and IL-6 (R&D Systems) pro-duction were measured by ELISA.
Cell surface labeling. Surface expression of TLR1, TLR2, CD14 and CD1awas determined using marker-specific antibodies. TLR1 and TLR2 signalswere amplified with a biotin-conjugated goat antibody against mouse IgG(Southern Biotechnology Associates, Birmingham, Alabama) and a strepta-vidin-phycoerythrin conjugate (Caltag, Burlingame, California). Cells werefixed with 1% paraformaldehyde, acquired on a Becton Dickinson FacsScan(Mountain View, California) and analyzed using WinMDI 2.8 (J. Trotter,Scripps Research Institute, San Diego, California).
Real-time quantitative PCR. Monocytes were stimulated for 12 h withmedium or IL-4 (0.01–1,000 U/ml). RNA was isolated using Trizol Reagent(Invitrogen, Carlsbad, California). cDNA was synthesized using the SUPER-SCRIPT Choice System Kit (Invitrogen). The following h36B4 primers andprobes were designed using Primer Express (Applied Biosystems,Branchburg, New Jersey): 5′-CCACGCTGCTGAACATGCT-3′ (forward), 5′-TCGAACACCTGCTGGATGAC-3′ (reverse), 5′-AACATGTCCCCCTTCTC-CTTTGGGCT-3′ (probe); TLR2 primers and probe sequences5′-GGCCAGCAAATTACCTGTGTG-3′ (forward), 5′-AGGCGGACATCCT-GAACCT-3′ (reverse), 5′-TCCATCCCATGTGCGTGGCC-3′ (probe).Reactions were carried out in TaqMan Universal PCR Master Mix (AppliedBiosystems) under recommended conditions, run on the ABI PRISM 7700and analyzed with Sequence Detector Software (Applied Biosystems). Therelative quantities of TLR2 per sample were calculated using the ∆∆ CT for-
mula as described43. The data were normalized by fold change to mediacontrol. The assay was repeated 3 times and values were averaged.
Immunoperoxidase labeling, immunofluorescence labeling and mi-croscopy. Immunoperoxidase labeling of cryostat sections was done as de-scribed44. TLR1- and TLR2-positive cells in dermal granulomas werequantitated by calculating the percentage of positive cells based on thetotal number of cells within the granuloma, as described44. Double im-munofluorescence was performed by serially incubating sections with anti-bodies against CD markers and TLR1, followed by incubation withisotype-specific fluorochrome (Caltag, Burlingame, California). Sectionswere washed with PBS containing 2% goat serum, incubated with TLR2-specific antibody for 1 h, then incubated with a tetramethyl rhodamineisothiocyanate (TRITC)–conjugated antibody against mouse IgG1(Southern Biotechnology). Sections were washed and mounted inVectashield medium (Vector laboratories, Burlingame, California). Controlswere performed as described45,46. Double immunofluorescence was exam-ined with a Leica-TCS-SP inverted confocal laser scanning microscope asdescribed46.
Note: Supplementary information is available on the Nature Medicine website.
AcknowledgmentsWe thank D. Williams and T. Gillis (National Hansen’s Disease Programs) foruse of their unpublished data on expression of ML1966 mRNA, W. Chung fortechnical assistance and S. Smale for critical comments on the manuscript andscientific discussion. This work was supported in part by grants from theNational Institutes of Health (AI07126, AI22553 and AI47866) and the WorldHealth Organization (IMMLEP).
Competing interests statementThe authors declare that they have no competing financial interests.
RECEIVED 22 NOVEMBER 2002; ACCEPTED 21 MARCH 2003
1. Brightbill, H.D. et al. Host defense mechanisms triggered by microbial lipopro-teins through Toll-like receptors. Science 285, 732–736 (1999).
2. Aliprantis, A.O. et al. Cell activation and apoptosis by bacterial lipoproteinsthrough Toll-like receptor-2. Science 285, 736–739 (1999).
3. Takeuchi, O. et al. Role of TLR1 in mediating immune response to microbiallipoproteins. J. Immunol. 169, 10–14 (2002).
4. Alexopoulou, L. et al. Hyporesponsiveness to vaccination with Borrelia burgdorferiOspA in humans and in TLR1- and TLR2-deficient mice. Nat. Med. 8, 878–884(2002).
5. Alexopoulou, L., Holt, A.C., Medzhitov, R. & Flavell, R.A. Recognition of double-stranded RNA and activation of NF-κB by Toll-like receptor 3. Nature 413,732–738 (2001).
6. Yang, R.B. et al. Toll-like receptor-2 mediates lipopolysaccharide-induced cellularsignalling. Nature 395, 284–288 (1998).
7. Poltorak, A. et al. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice:mutations in Tlr4 gene. Science 282, 2085–2088 (1998).
8. Takeuchi, O. et al. Differential roles of TLR2 and TLR4 in recognition of gram-neg-ative and gram-positive bacterial cell wall components. Immunity 11, 443–451(1999).
9. Hayashi, F. et al. The innate immune response to bacterial flagellin is mediated byToll- like receptor 5. Nature 410, 1099–1103 (2001).
10. Takeuchi, O. et al. Discrimination of bacterial lipoproteins by Toll-like receptor 6.Int. Immunol. 13, 933–940 (2001).
11. Hemmi, H. et al. Small anti-viral compounds activate immune cells via the TLR7MyD88- dependent signaling pathway. Nat. Immunol. 3, 196–200 (2002).
12. Jurk, M. et al. Human TLR7 or TLR8 independently confer responsiveness to theantiviral compound R-848. Nat. Immunol. 3, 499 (2002).
13. Hemmi, H. et al. A Toll-like receptor that recognizes bacterial DNA. Nature 408,740–745 (2000).
14. Medzhitov, R., Preston-Hurlburt, P. & Janeway, C.A.J. A human homologue ofthe Drosophila Toll protein signals activation of adaptive immunity. Nature 388,394–397 (1997).
15. Thoma-Uszynski, S. et al. Induction of direct antimicrobial activity through mam-malian toll-like receptors. Science 291, 1544–1547 (2001).
16. Ridley, D.S. & Jopling, W.H. Classification of leprosy according to immunity. Afive-group system. Int. J. Lepr. 34, 255–273 (1966).
17. Yamamura, M. et al. Defining protective responses to pathogens: cytokine pro-files in leprosy lesions. Science 254, 277–279 (1991).
18. Salgame, P. et al. Differing lymphokine profiles of functional subsets of human
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
rem
edic
ine

532 NATURE MEDICINE • VOLUME 9 • NUMBER 5 • MAY 2003
ARTICLES
CD4 and CD8 T cell clones. Science 254, 279–282 (1991).19. Maeda, Y. et al. Novel 33-kilodalton lipoprotein from Mycobacterium leprae.
Infect. Immun. 70, 4106–4111 (2002).20. Staege, H., Schaffner, A. & Schneemann, M. Human toll-like receptors 2 and 4
are targets for deactivation of mononuclear phagocytes by interleukin-4.Immunol. Lett. 71, 1–3 (2000).
21. Sieling, P.A. et al. IL-12 regulates T helper type 1 cytokine responses in human in-fectious disease. J. Immunol. 153, 3639–3647 (1994).
22. Garcia, V.E. et al. IL-18 promotes type 1 cytokine production from NK cells and Tcells in human intracellular infection. J. Immunol. 162, 6114–6121 (1999).
23. Flo, T.H. et al. Differential expression of Toll-like receptor 2 in human cells. J.Leukoc. Biol. 69, 474–481 (2001).
24. O’Brien, A.D. et al. Genetic control of susceptibility to Salmonella typhimurium inmice: role of the LPS gene. J. Immunol. 124, 20–24 (1980).
25. Anderson, G.W.J. & Osterman, J.V. Host defenses in experimental rickettsialpox:genetics of natural resistance to infection. Infect. Immun. 28, 132–136 (1980).
26. Cross, A. et al. The importance of a lipopolysaccharide-initiated, cytokine-medi-ated host defense mechanism in mice against extraintestinally invasiveEscherichia coli. J. Clin. Invest. 96, 676–686 (1995).
27. Weinstein, D.L., Lissner, C.R., Swanson, R.N. & O’Brien, A.D. Macrophage defectand inflammatory cell recruitment dysfunction in Salmonella susceptibleC3H/HeJ mice. Cell. Immunol. 102, 68–77 (1986).
28. Medina, E. & North, R.J. Resistance ranking of some common inbred mousestrains to Mycobacterium tuberculosis and relationship to major histocompatibilitycomplex haplotype and Nramp1 genotype. Immunology 93, 270–274 (1998).
29. Takeuchi, O., Hoshino, K. & Akira, S. Cutting edge: TLR2-deficient and MyD88-deficient mice are highly susceptible to Staphylococcus aureus infection. J.Immunol. 165, 5392–5396 (2000).
30. Wang, X. et al. Toll-like receptor 4 mediates innate immune responses toHaemophilus influenzae infection in mouse lung. J. Immunol. 168, 810–815(2002).
31. Wooten, R.M. et al. Toll-like receptor 2 is required for innate, but not acquired,host defense to Borrelia burgdorferi. J. Immunol. 168, 348–355 (2002).
32. Kang, T.J. & Chae, G.T. Detection of Toll-like receptor 2 (TLR2) mutation in thelepromatous leprosy patients. FEMS Immunol. Med. Microbiol. 31, 53–58 (2001).
33. Aliprantis, A.O., Yang, R.B., Weiss, D.S., Godowski, P. & Zychlinsky, A. The apop-totic signaling pathway activated by Toll-like receptor-2. EMBO J. 19, 3325–3336(2000).
34. Cole, S.T. et al. Massive gene decay in the leprosy bacillus. Nature 409,1007–1011 (2001).
35. Cole, S.T. et al. Deciphering the biology of Mycobacterium tuberculosis from thecomplete genome sequence. Nature 393, 537–544 (1998).
36. Post, F.A. et al. Mycobacterium tuberculosis 19-kilodalton lipoprotein inhibitsMycobacterium smegmatis-induced cytokine production by human macrophagesin vitro. Infect. Immun. 69, 1433–1439 (2001).
37. Yeremeev, V.V. et al. Deletion of the 19kDa antigen does not alter the protectiveefficacy of BCG. Tubercle Lung Dis. 80, 243–247 (2000).
38. Means, T.K. et al. Human Toll-like receptors mediate cellular activation byMycobacterium tuberculosis. J. Immunol. 163, 3920–3927 (1999).
39. Means, T.K. et al. The CD14 ligands lipoarabinomannan and lipopolysaccharidediffer in their requirement for Toll-like receptors. J. Immunol. 163, 6748–6755(1999).
40. Hirschfeld, M., Ma, Y., Weis, J.H., Vogel, S.N. & Weis, J.J. Cutting edge: repurifi-cation of lipopolysaccharide eliminates signaling through both human andmurine toll-like receptor 2. J. Immunol. 165, 618–622 (2000).
41. Takeuchi, O. et al. Cutting edge: preferentially the R-stereoisomer of the my-coplasmal lipopeptide macrophage-activating lipopeptide-2 activates immunecells through a toll-like receptor 2- and MyD88-dependent signaling pathway. J.Immunol. 164, 554–557 (2000).
42. Bairoch, A., Bucher, P. & Hofmann, K. The PROSITE database, its status in 1997.Nucleic Acids Res. 25, 217–221 (1997).
43. Monney, L. et al. Th1-specific cell surface protein Tim-3 regulates macrophageactivation and severity of an autoimmune disease. Nature 415, 536–541 (2002).
44. Modlin, R.L. et al. In situ identification of cells in human leprosy granulomas withmonoclonal antibodies to interleukin 2 and its receptor. J. Immunol. 132,3085–3090 (1984).
45. Stenger, S. et al. An antimicrobial activity of cytolytic T cells mediated by gran-ulysin. Science 282, 121–125 (1998).
46. Ochoa, M.T. et al. T-cell release of granulysin contributes to host defense in lep-rosy. Nat. Med. 7, 174–179 (2001).
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
rem
edic
ine




















