
Acantholaimus (chromadoridae:nematoda) from the Indian Ocean: description of seven species
18
Hydrobiologia 346: 59–76, 1997. 59 c 1997 Kluwer Academic Publishers. Printed in Belgium. Acantholaimus (chromadoridae:nematoda) from the Indian Ocean: description of seven species Agnes W. Muthumbi 1 & Magda Vincx University of Ghent, Department of Morphology, Systematics & Ecology, Marine Biology Section, K. L. Ledeganckstraat 35, B-9000 Gent, Belgium (e-mail: [email protected]; Tel: +32 (0) 9 2645216; Fax: +32 (0) 9 2645344) 1 On leave from Kenya Marine & Fisheries Research Institute, P.O. Box 81651, Mombasa, Kenya Received 15 October 1996; in revised form 7 January 1997; accepted 26 February 1997 Abstract Seven species of the Acantholaimus are described. Acantholaimus vermeuleni sp.n. is characterised by labial and cephalic sensilla that are located at the same level, two post amphidial setae on the dorso-lateral side and a poorly developed stoma without distinct teeth. Acantholaimus verscheldi sp.n. is characterised by a narrow elongate pharyngeal region, a long stoma with distinct teeth and short (4–7 m) cephalic sensilla. Acantholaimus heipi sp.n. is characterised by a narrow elongate pharyngeal region, well developed teeth in the stoma and long cephalic sensilla (11–13 m). Acantholaimus elegans Jensen 1988 has a narrow anterior pharyngeal region that increases in width posteriorly, it has setae before and after the amphids on both sides and stoma with well developed teeth. Acantholaimus gathumai sp.n. is characterised by long cephalic (10–15 m) and somatic (8–10 m) setae and lateral differentiation with fine dots (5–7 m in width). Acantholaimus geraerti sp.n. has long cephalic sensilla (15–19 m) and narrow (4–6 m) distinct lateral differentiation. Acantholaimus invaginatum sp.n. is characterised by very long cephalic setae (16–21 m), several setae at the pharyngeal region and wide lateral differentiation with fine dots, often the stoma is invaginated. Distribution of the different species of Acantholaimus encoutered in the four transects studied is also described. Introduction This work is part of the Netherlands Indian Ocean Pro- gramme of 1992 [cruise A1 (June to July 1992) and A2 (Novemberto December 1992) of the R. V. Tyro], which is aimed at assessing the monsoon effects on the Kenyan coastal ecosystems where both pelagic and benthic systems were studied. Benthic deep-sea sam- pling was carried out to assess nematode community structure. Among the deep-sea nematodes, the genus Acan- tholaimus is often one of the most abundant genus encountered. In this case, the genus is found inhab- iting sediments at depths of 500 to 2000 m. Soetaert and Heip (1995) reported that Acantholaimus showed increase in importance in terms of density with increase in water depth from the shelf break to the deep-sea sta- tions in the Mediterranean and North Atlantic. In the present work, the genus Acantholaimus was the most dominant in the family Chromadoridae and at least thirty seven morphotypes were observed but only a few species are described in this paper due to poor representation. Materials and methods Samples were taken off the Kenyan coast on four tran- sects from North to South; Kiwayu, Tana, Sabaki and Gazi, Table 1 (see also Map 1 in Muthumbi et al in press) Note that the stations are denoted by 1– for the first cruise and the equivalent are denoted by 5– for second cruise. Sampling was done using a box corer from which two sub samples were taken to a depth of 5 cm using a plastic core of diameter 2.6 cm.
Transcript of Acantholaimus (chromadoridae:nematoda) from the Indian Ocean: description of seven species

Hydrobiologia 346: 59–76, 1997. 59c 1997 Kluwer Academic Publishers. Printed in Belgium.
Acantholaimus (chromadoridae:nematoda) from the Indian Ocean:description of seven species
Agnes W. Muthumbi1 & Magda VincxUniversity of Ghent, Department of Morphology, Systematics & Ecology, Marine Biology Section,K. L. Ledeganckstraat 35, B-9000 Gent, Belgium (e-mail: [email protected]; Tel: +32 (0) 9 2645216;Fax: +32 (0) 9 2645344)1 On leave from Kenya Marine & Fisheries Research Institute, P.O. Box 81651, Mombasa, Kenya
Received 15 October 1996; in revised form 7 January 1997; accepted 26 February 1997
Abstract
Seven species of the Acantholaimus are described. Acantholaimus vermeuleni sp.n. is characterised by labial andcephalic sensilla that are located at the same level, two post amphidial setae on the dorso-lateral side and a poorlydeveloped stoma without distinct teeth. Acantholaimus verscheldi sp.n. is characterised by a narrow elongatepharyngeal region, a long stoma with distinct teeth and short (4–7 �m) cephalic sensilla. Acantholaimus heipisp.n. is characterised by a narrow elongate pharyngeal region, well developed teeth in the stoma and long cephalicsensilla (11–13 �m). Acantholaimus elegans Jensen 1988 has a narrow anterior pharyngeal region that increasesin width posteriorly, it has setae before and after the amphids on both sides and stoma with well developed teeth.Acantholaimus gathumai sp.n. is characterised by long cephalic (10–15 �m) and somatic (8–10 �m) setae andlateral differentiation with fine dots (5–7 �m in width). Acantholaimus geraerti sp.n. has long cephalic sensilla(15–19 �m) and narrow (4–6 �m) distinct lateral differentiation. Acantholaimus invaginatum sp.n. is characterisedby very long cephalic setae (16–21 �m), several setae at the pharyngeal region and wide lateral differentiation withfine dots, often the stoma is invaginated.
Distribution of the different species of Acantholaimus encoutered in the four transects studied is also described.
Introduction
This work is part of the Netherlands Indian Ocean Pro-gramme of 1992 [cruise A1 (June to July 1992) andA2 (November to December 1992) of the R. V. Tyro],which is aimed at assessing the monsoon effects onthe Kenyan coastal ecosystems where both pelagic andbenthic systems were studied. Benthic deep-sea sam-pling was carried out to assess nematode communitystructure.
Among the deep-sea nematodes, the genus Acan-tholaimus is often one of the most abundant genusencountered. In this case, the genus is found inhab-iting sediments at depths of 500 to 2000 m. Soetaertand Heip (1995) reported that Acantholaimus showedincrease in importance in terms of density with increasein water depth from the shelf break to the deep-sea sta-tions in the Mediterranean and North Atlantic.
In the present work, the genus Acantholaimus wasthe most dominant in the family Chromadoridae and atleast thirty seven morphotypes were observed but onlya few species are described in this paper due to poorrepresentation.
Materials and methods
Samples were taken off the Kenyan coast on four tran-sects from North to South; Kiwayu, Tana, Sabaki andGazi, Table 1 (see also Map 1 in Muthumbi et al inpress) Note that the stations are denoted by 1– for thefirst cruise and the equivalent are denoted by 5– forsecond cruise. Sampling was done using a box corerfrom which two sub samples were taken to a depth of5 cm using a plastic core of diameter 2.6 cm.
Article: hydr 3748 Pips nr 136018 BIO2KAP
*136018 hydr3748.tex; 24/06/1997; 18:18; v.7; p.1
60
Table 1. Location of the sampling stations
Date Station Latitude S Longitude E Depth (m)
22/6/92 105 04�24006 39�45099 511
23/6/92 106 04�20035 40�21070 1000
23/6/92 107 04�21083 41�13016 2053
28/10/92 117 03�08021 40�41080 500
29/6/92 118 03�08046 41�01077 1112
29/6/92 119 03�10067 41�14020 2007
3/7/92 132 01�56003 41�31054 1000
3/7/92 133 02�01049 41�46096 2015
4/7/92 131 02�00027 41�26062 500
6/7/92 136 02�40005 41�10017 992
3/12/92 505 04�25033 39�45021 520
4/12/92 506 04�19045 40�21080 1020
2/12/92 507 04�21031 41�13064 2088
25/11/92 517 03�09043 40�41025 508
26/11/92 518 03�07098 40�59096 963
27/11/92 519 03�09028 41�16053 2179
20/11/92 531 02�00048 41�37056 516
22/11/92 532 01�56002 41�37056 904
21/11/92 533 02�00086 41�47071 2027
7/12/92 552 04�07071 39�54067 500
Nematodes were transferred slowly to glycerine.Drawings were made with the aid of a camera lucidaon a Leitz Dialux 20 EB microscope.
Type specimens are deposited in the collection ofKoninklijk Belgisch Instituut voor Natuurwetenchap-pen (KBIN) of Brussels, slide numbers R1519-R1531and the Marine Biology section of the University ofGhent (MBRUG) slide numbers 10327-10372.
The abbreviations used in the text and tables are:a: body length divided by maximum body diameter,
w: body length divided by pharyngeal length, c: bodylength divided by tail length, c0: tail length divided byanal body diameter
abd: anal body diameter, amp dist: amphid distancefrom the anterior, amp wid: amphid width, bulb d: bulbdiameter
cbd: corresponding body diameter, D: water depth,gub: gubernaculum, hd: head diameter at the levelof the cephalic setae, L: total body length, L0: bodylength from the anterior to anus, M: maximum bodydiameter, spic: spicule length, V: position of vulva as apercentage of total body length, V0: position of vulvaas a percentage of body length from anterior to anus,v: vulva distance from the anterior
Formula: distance from the anterior to;head end of the pharynx M (vulva) anus
cbdAll measurements (not ratios) are in micrometers
and all curved structures are measured along the arc.
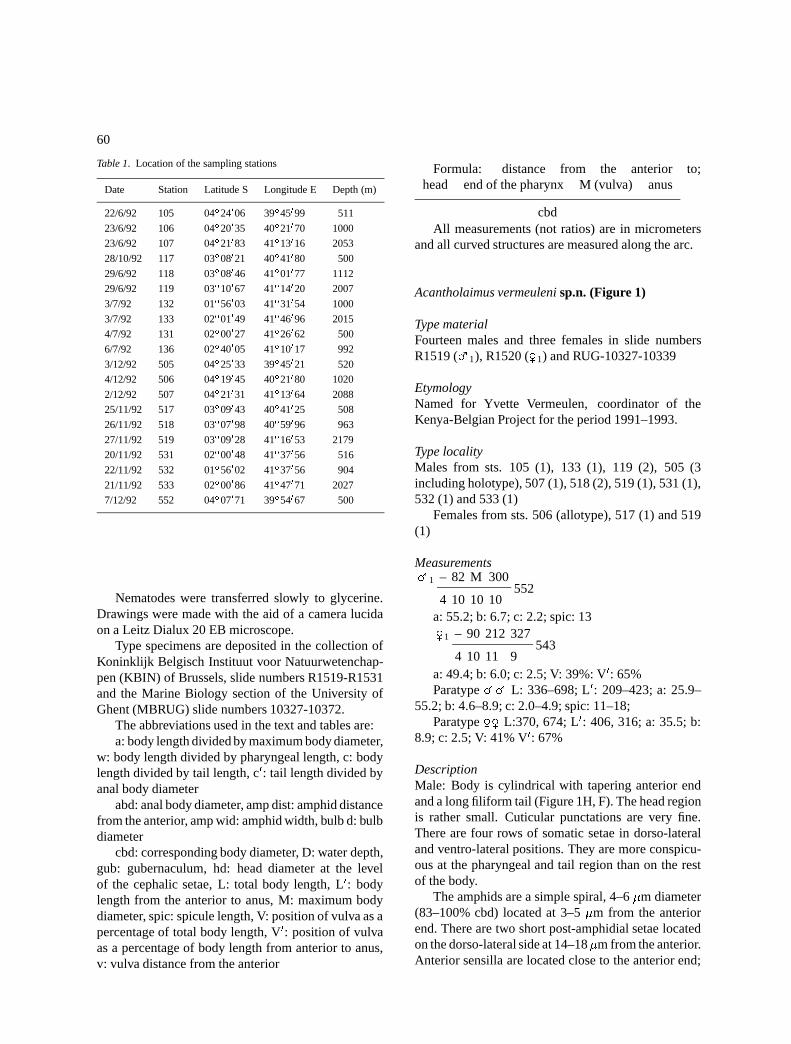
Acantholaimus vermeuleni sp.n. (Figure 1)
Type materialFourteen males and three females in slide numbersR1519 (�1), R1520 (�1) and RUG-10327-10339
EtymologyNamed for Yvette Vermeulen, coordinator of theKenya-Belgian Project for the period 1991–1993.
Type localityMales from sts. 105 (1), 133 (1), 119 (2), 505 (3including holotype), 507 (1), 518 (2), 519 (1), 531 (1),532 (1) and 533 (1)
Females from sts. 506 (allotype), 517 (1) and 519(1)
Measurements�1 – 82 M 300
5524 10 10 10
a: 55.2; b: 6.7; c: 2.2; spic: 13�1 – 90 212 327
5434 10 11 9
a: 49.4; b: 6.0; c: 2.5; V: 39%: V0: 65%Paratype �� L: 336–698; L0: 209–423; a: 25.9–
55.2; b: 4.6–8.9; c: 2.0–4.9; spic: 11–18;Paratype �� L:370, 674; L0: 406, 316; a: 35.5; b:
8.9; c: 2.5; V: 41% V0: 67%
DescriptionMale: Body is cylindrical with tapering anterior endand a long filiform tail (Figure 1H, F). The head regionis rather small. Cuticular punctations are very fine.There are four rows of somatic setae in dorso-lateraland ventro-lateral positions. They are more conspicu-ous at the pharyngeal and tail region than on the restof the body.
The amphids are a simple spiral, 4–6 �m diameter(83–100% cbd) located at 3–5 �m from the anteriorend. There are two short post-amphidial setae locatedon the dorso-lateral side at 14–18�m from the anterior.Anterior sensilla are located close to the anterior end;
hydr3748.tex; 24/06/1997; 18:18; v.7; p.2
61
Figure 1. Acantholaimus vermeuleni sp.n.
A: �1 total view
B: �1 head region
C: �1 head region
D: �2 total view minus tail
E: �2 head region
F: �2 total view
G: �1 spicule
H: �1 total view
hydr3748.tex; 24/06/1997; 18:18; v.7; p.3

62
inner labial setae are indistinct, outer labial are halfas long as the cephalic sensilla which are 3–6 �m andlocated at the same level (Figures 1B & E).
The stoma is narrow, without distinct teeth. Thepharynx is cylindrical (66–90�m long), with a slightlyswollen posterior end to form a bulb. Cardia is distinct(Figures 1F & H).
The reproductive system is monorchic with out-stretched testis; there is a short germinal zone andlarge spermatozoa with large nucleus. The vas defer-ens is short (Figures 1F & H). Spicules are 1.2–1.5 abdlong, flat on the distal end and blunt on the proximalend. There is a pair of pre-cloacal setae at 6–7 �m infront of the cloaca (Figure 1G).
The tail is conical anteriorly (35–45 �m long) andmeasures 100–349 �m. Females: They are similar tomales (Figures 1A, C and D). The reproductive sys-tem is amphidelphic with reflexed ovaries; they have ashort germinal zone, developing oocytes, and uterus ispartially filled with spermatozoa. The vulva and vaginaare simple.
Differential diagnosisAcantholaimus vermeuleni sp.n. is characterised bya narrow anterior end; short (3–6 �m long) cephalicsensilla located at the same level as the outer labialones which are half as long as the former; amphids(80–100% cbd) that are single spiral with a distinctspiral origin and two post-amphidial setae located onthe dorso-lateral position at 14–18�m from the anteriorend.
Acantholaimus vermeuleni sp.n. resembles Acan-tholaimus akvavitus Gerlach, Shrage & Riemann1979, A. gigantisetosus Vivier 1985, A. iubilus Ger-lach, Shrage & Riemann 1979, A. maks Gerlach,Shrage & Riemann 1979, A. megamphis Vivier 1985,A. microdontus Gourbault 1985, A. minutus Vitielo1970, and A. septimus Gerlach, Shrage & Riemann1979 in having two post amphidial setae on the dorso-lateral position. A. vermeuleni however differs from allthese nematodes, in that they have one or two extrapost-amphidial setae on the ventro-lateral position aswell and also in having well developed teeth in thestoma which are lacking in the former. A. microdontusdiffers from A. vermeuleni in size (L0 = 760–1105 �mand M = 33–50�m in A. microdontus compared toL0 = 200–400 �m and M = 9–19 �m in Acantholaimusvermeuleni sp.n.).
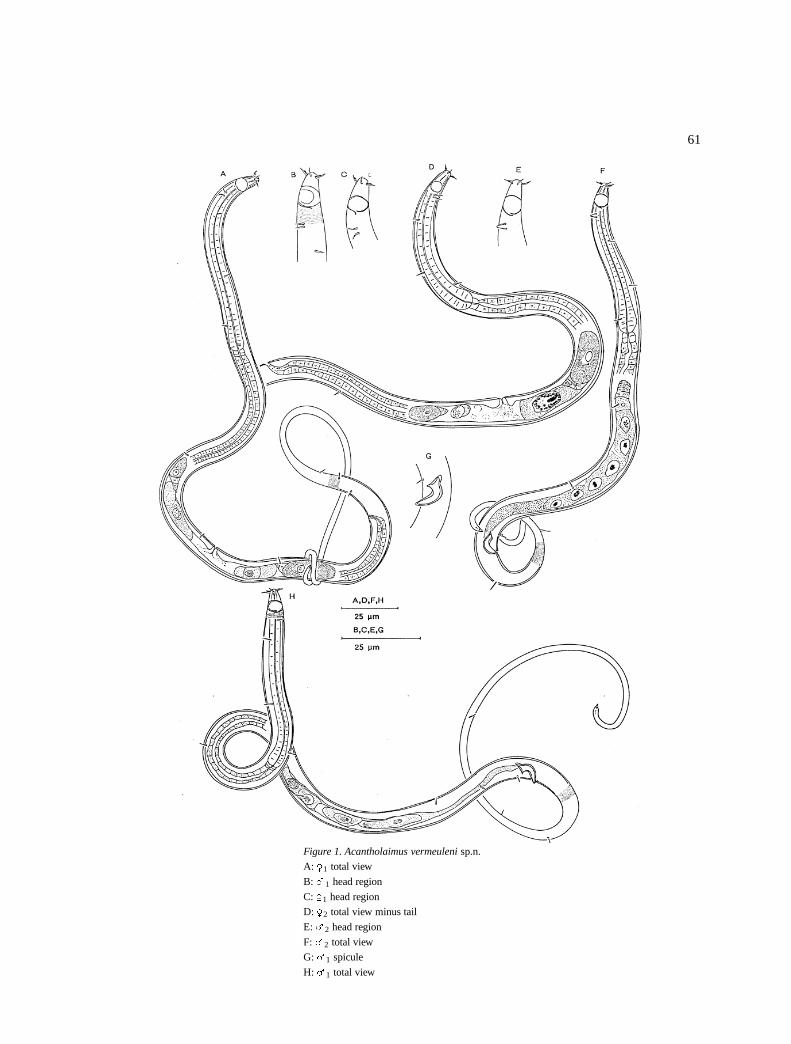
Acantholaimus verscheldi sp.n. (Figure 2)
Type materialSeven males and six females on slide numbers R1521(�1), R1522 (�1) and RUG-10332, 10334 and 10340-10346
EtymologyNamed for Dominick Verschelde of the University ofGent
Type localityMales from sts. 107 (1), 133 (1), 136 (1), 505 (2), 506(holotype) and 533 (1)
Females from sts. 133 (1), 505 (2), 506 (allotype),507 (1) and 519 (1)
Measurements� 1 – 127 M 405
5207 11 13 11
a: 40; b: 4.1; c: 4.5; spic: 11�1 – 130 260 410
5967 12 16 9
a: 37.3; b: 4.6; c: 3.2; V: 44% V0: 63%Paratype �� L: 490–621; L0: 280–467; a: 30.0–
54.1; b: 3.9–6.5; c: 2.3–4.5; spic: 11–15Paratype �� L: 471–618; L0: 290–441; a: 22.4–
36.8; b: 3.9–4.5; c: 2.3–4.6; V: 46–52%: V0: 65–67%
DescriptionMales: Body is cylindrical, narrow anterior pharyngealregion with a slight increase in width posteriorly, andthe tail is filiform (Figure 2E). Cuticular punctationsare indistinct. Somatic setae are in four longitudinalrows, that is, two dorso-lateral and two ventro-lateral.
Amphids are 4–7 �m in diameter (71–100% cbd),spiral with a single turn and they are located (posteriorof the base of the stoma) at 11–15 �m from the ante-rior end. Inner labial sensilla are inconspicuous, outerlabial are short 2–3 �m long and at the same level asthe cephalic setae. The cephalic setae are 4–7 �m longand close to the anterior. The stoma is long (9–13 �m),and it has three teeth anteriorly and highly sclerotizedwalls posteriorly (Figure 2C).
The pharynx is cylindrical with some interruptionsin the muscular part at the posterior part of the pharynxbut no well formed terminal bulb. Cardia is short butprominent (Figure 2B).
hydr3748.tex; 24/06/1997; 18:18; v.7; p.4
63
Figure 2. Acantholaimus verscheldi sp.n.
A: �1 pharyngeal region
B: �1 pharyngeal region
C: �1 head region
D: �1 head region
E: �1 total view
F: �1 total view
G: �1 reproductive system
hydr3748.tex; 24/06/1997; 18:18; v.7; p.5

64
The reproductive system is monorchic with out-stretched testis filled with large spermatozoa. The vasdeferens is short. The spicules are 1.0–1.3 abd long.There is a pair of pre-cloacal setae at 7 �m anterior ofthe cloaca (Figure 2G). The tail (115–210 �m long) isconical with a long filiform posterior end.
Females: They are similar to the males in generalshape and size (Figures 2A, D & F). The reproduc-tive system is amphidelphic with reflexed ovaries. Theanterior branch is to the right and the posterior one tothe left of the intestines. There are large spermatozoain the uterus. Vulva and vagina are simple.
Differential diagnosisAcantholaimus verscheldi sp.n. is characterised by anarrow elongate anterior pharyngeal region, short outerlabial sensilla at the same level as the cephalic setaethat are 4–7 �m long.
Acantholaimus verscheldi sp.n. resembles Acan-tholaimus elegans Jensen 1988 in body length, stomaand the position of the amphid. It also resembles Acan-tholaimus heipi sp.n. and A. septimus Gerlach, Shrage& Riemann, 1979 in the stoma. It differs from Acan-tholaimus elegans in the shape of the posterior pha-ryngeal region which is more enlarged posteriorly inA. elegans than in Acantholaimus verscheldi. Acantho-laimus verscheldi differs from Acantholaimus heipi inthe length of the anterior sensilla (labial sensilla areindistinct and the cephalic ones are 11–13 �m long inAcantholaimus heipi). It differs from A. septimus inthe position of the amphids which are within the stomaregion, the presence of two post-amphidial setae oneither side of the amphids in the A. septimus and thebody size (L0 = 575–850 and M = 21–23 �m in A. sep-timus compared to L0 = 280–467�m and M = 12–21 inAcantholaimus verscheldi sp.n.).
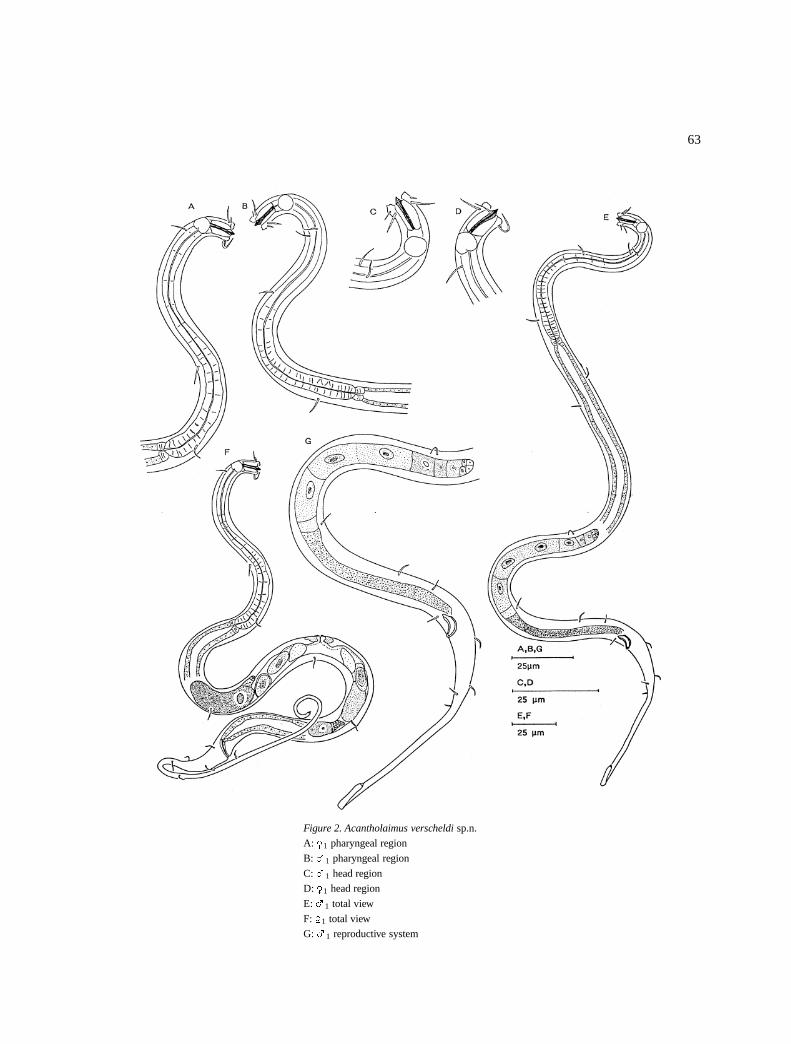
Acantholaimus heipi sp.n. (Figure 3)
Type materialFour males and two females in slide numbers R1523(�1), RUG-10314 (�1), RUG-10347-10349
EtymologyNamed for Prof. Heip of the University of Ghent
Type localityMales from sts. 133 (holotype), 517 (1) and 519 (2)
females from sts. 119 (1) and 133 (1)
Measurements� 1 – 105 M 326
6256 14 14 12
a: 44.6; b: 6.0; c: 2.0; spic: 14� 1 – 124 242 375
5597 13 17 12
a: 31.1; b: 4.5; c: 3.0; V: 43%: V0: 66%Paratype � L: 642; L0: 343; a: 35.7; b: 6.1; c: 2.1;
spic: 14–18. Two males with broken tail L: 421, 457L0: 263, 271
paratype � L: 589; L0: 312; a: 36.8; b: 5.7; c: 2.1;V: 36%: V0: 68%
DescriptionMales: Body is cylindrical and narrow on the first halfof the pharyngeal region, with a slight increase in widthtowards the end of the pharyngeal region and the tailhas a short conical anterior part and a long filiformend. Head region (6–7 �m in diameter) is continuouswith the rest of the body. Cuticular punctations areindistinct. Somatic setae (3–5 �m long) are in fourlongitudinal rows: two dorso-lateral and two ventro-lateral rows (Figure 3E).
The amphids are spiral with a single turn and thespiral origin can be seen in some specimens; they are(71–100% cbd) located at 8–11 �m from the anteriorend [(anterior most part of the amphids are at the levelof the base of the stoma (Figure 3F) or posterior to it(Figure 3B)]. Inner labial sensilla are indistinct, outerlabial short (3 �m long) and the cephalic ones are long(11–13 �m long) and close to the anterior end.
The stoma is long (8–11 �m) and it has three pro-trusible teeth at the anterior part and sclerotized wallsin the posterior part. Pharyngeal muscles surround thestoma. Pharynx (92–124�m long) is cylindrical with aslight expansion and several interruptions of the mus-cular part at the base. Cardia short (4 �m) but promi-nent (Figure 3H).
The reproductive system is long with a short germi-nal zone posterior of which are large spermatozoa filledwith a large nuclei. Vas deferens is short. Spicules are1.2–1.5 abd and typical Acantholaimid in shape (Fig-ure 3G). A pair of sub-ventral pre-cloacal setae arefound at 1–2 �m from the cloaca.
Tail is long (184–299 �m) and slightly swollen atthe end and tip is pointed. Female: They are similarto males (Figures A, C & D). The reproductive systemis amphidelphic with reflexed ovaries. Anterior branchto the right, posterior one to the left of the intestine(Figure 3I).
hydr3748.tex; 24/06/1997; 18:18; v.7; p.6
65
Figure 3. Acantholaimus heipi sp.n.
A: �1 total view
B: �2 head region
C: �1 head region
D: �1 pharyngeal region
E: �1 total view
F: �1 head region
G: �1 spicule
H: �2 pharyngeal region
I: �1 reproductive system
hydr3748.tex; 24/06/1997; 18:18; v.7; p.7

66
Differential diagnosisAcantholaimus heipi sp.n. is characterised by a narrowpharyngeal region; long (11–13 �m) cephalic setae,long stoma with three teeth, simple spiral amphidslocated close to the base of the stoma and a long filiformtail.
Acantholaimus heipi sp.n. is similar to Acantho-laimus elegans Jensen, 1988 in body length (L: 540–720�m), position of the amphids and the stoma length.However, Acantholaimus heipi differs from A. elegansin that all three crowns of anterior sensilla are distinctwith the cephalic setae being only 7–8 �m long inA. elegans and 11–13 �m in Acantholaimus heipi sp.n;the posterior part of the pharyngeal region is muchwider than the anterior part in A. elegans and it is alsoa much wider (M = 25–30 �m) species compared toAcantholaimus heipi sp.n. (M = 12–18 �m).
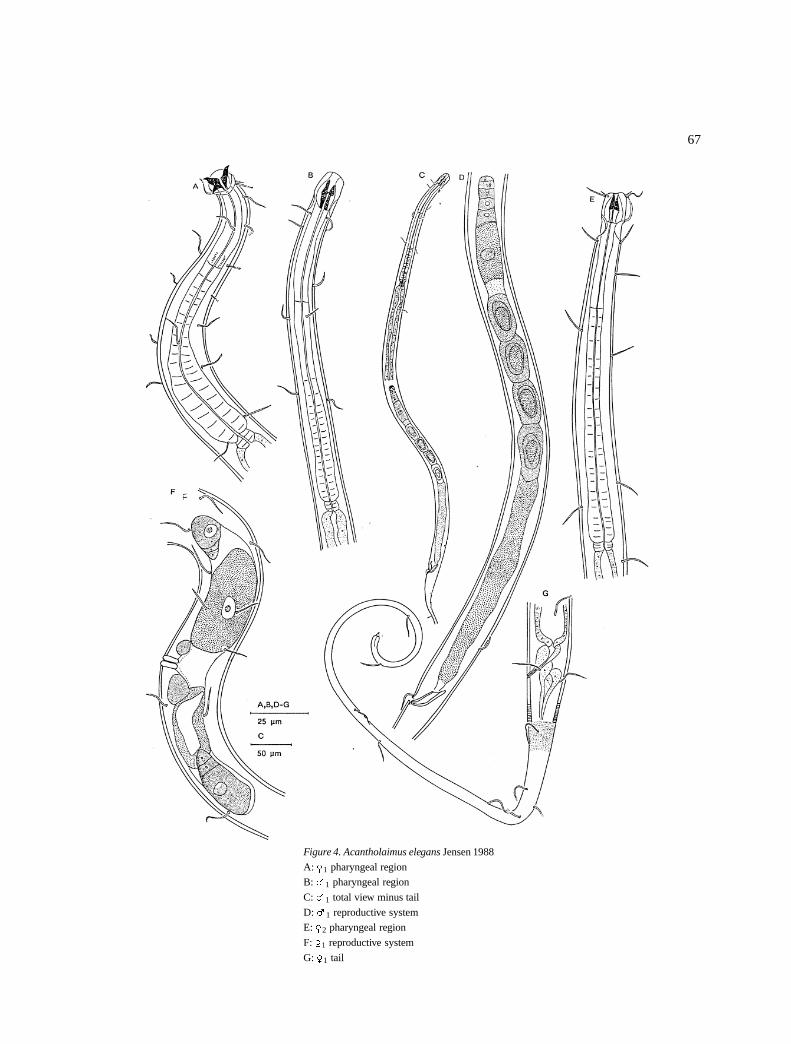
Acantholaimus elegans Jensen 1988 (Figure 4)
Material studiedThree males and two females on slide numbers R1524(�1), R1525 (�1) and RUG-10350-10352
LocalityMales from sts. 133 (1), 506 (1) and 552 (�1)
Females from sts. 117 (1) and 507 (�1)
Measurements�1 – 148 M 512(tail broken) 581
10 14 17 13spic: 15�1 – 124 298 454
73910 22 27 14
a: 27.4; b: 6.0; c: 2.6; V: 40% V0: 66%�� (tail broken in one) L: 954; L0: 388–525; a:
41.5; b: 6.4; c: 2.2; spic: 19�2 L: 739: L0: 454; a: 27.4; b: 6.0; c: 2.6; V: 44%;
V0: 68%
RedescriptionMales: Body is cylindrical, narrow in the anterior halfof the pharyngeal region, and gradually increase inwidth on the posterior half until mid body. Cuticularpunctations are very fine. The somatic setae are 8–11 �m long and in four longitudinal rows, that is twosub-dorsal and two sub-ventral positions (Figure 4C).
The amphids are single spiral, 7–11 �m (64–100%cbd) in diameter and located at (partly within the stomaregion) 7–12 �m from the anterior end. There maybefour setae before the amphidial and four posterior ofit of equal length (9–12 �m) located at sub-dorsal andsub-ventral positions similar to the somatic setae (Fig-ure 4E). Inner labial sensilla are inconspicuous andthe outer labial are 5 �m long and at the same levelas the cephalic ones which are 10–11 �m long (Fig-ure 4E). The stoma is long (12–18 �m) surroundedby well developed muscles and it has three (four?)teeth anteriorly and highly sclerotized walls posterior-ly. The pharynx is 124–159 �m long, cylindrical witha slight swollen terminal end (Figure 4B). Cardia isshort (4 �m) but conspicuous.
The reproductive system is monorchic with out-stretched testis located to the right of the intestine. Ithas a short germinal zone and large spermatozoa (fiveor six spermatozoa in the testis). The spicules are 1.2abd long. There is a pair of pre-cloacal setae locatedjust before the cloaca (Figure 4D).
The tail has a conical anterior part and filiformposterior part with several setae of the same length asthe somatic one. The most terminal setae is located at13–15 �m before the end of the tail (Figure 4G).
Females: They are similar to males in general bodyshape (Figures 4A, E & G). The reproductive systemis amphidelphic with reflexed short ovaries. The uterusis large and may contain an egg and/or spermatozoa.Vulva is simple and vagina is thick walled (Figure 4F).
DiscussionThese specimens are considered to be Acantholaimuselegans because of the shape of the stoma (long withwell developed teeth), shape of the pharyngeal region(anteriorly much narrower than the posterior part) andthe body size.
There are some differences however, in these spec-imens compared to the original description by Jensen1988 in the length of the cephalic setae (7–8 �m com-pared to 10-11 �m) and somatic setae (8–10 �m com-pared to 8–11 �m). In the description of Jensen, theholotype has extra setae within the stoma region (andthe amphids are not conspicuous) which are not foundin the allotype female and the paratype male. However,Jensen does not mention this difference. In our spec-imens, we had one male and female with extra setaesituated anterior of the amphids. But owing to the highvariability in deep-sea sediments, and the low numberof individuals per species, we shall not attempt to split
hydr3748.tex; 24/06/1997; 18:18; v.7; p.8
67
Figure 4. Acantholaimus elegans Jensen 1988
A: �1 pharyngeal region
B: �1 pharyngeal region
C: �1 total view minus tail
D: �1 reproductive system
E: �2 pharyngeal region
F: �1 reproductive system
G: �1 tail
hydr3748.tex; 24/06/1997; 18:18; v.7; p.9

68
the species on that aspect alone, besides on observa-tion of the original material the setae at 13–15�m fromthe tail terminal could be seen in all three specimens,therefore, we redescribe it as A. elegans.
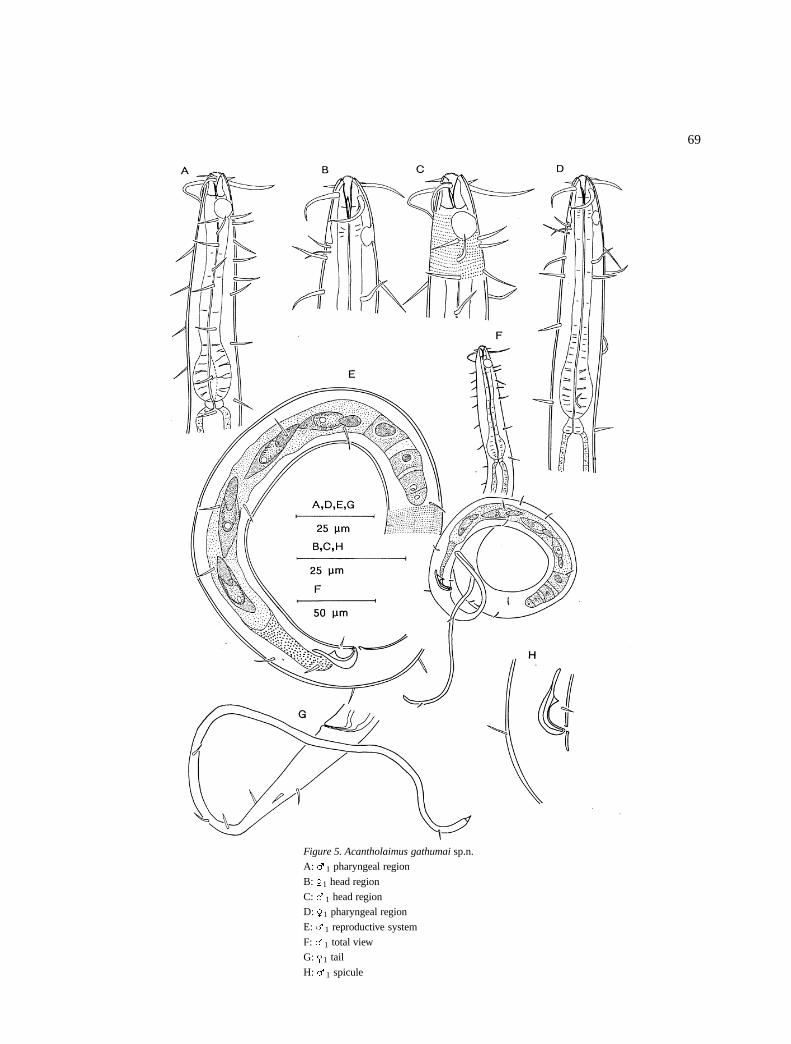
Acantholaimus gathumai sp.n. (Figure 5)
Type materialThree males and three females on slide numbers R1526�(1), R1527 (�1) and RUG-10353-10356
EtymologyName given for Prof. J. M. Gathuma of the Universityof Nairobi
Type localityMales from sts. 505 (holotype), 506 (1), 533 (1)
Females from sts. 119 (1), 505 (allotype), 507 (1)
Measurements�1 – 78 M 363
5817 16 18 14
a: 32.3; b: 7.4; c: 2.7; spic: 18�1 – 77 265 395
5907 15 17 11
a: 34.7; b: 7.7; c: 3.0; V: 50%; V0: 67%Paratype �� L: 562–608; L0: 381–395; a: 29.6–
40.5; b: 6.8–7.5; c: 2.9–3.1; spic: 23–24Paratype �� L: 556–631; L0: 395–423; a: 30.9–
33.2; b: 7.3–7.8; c: 3.0–3.7; V: 48–50%; V0: 70–71%
DescriptionMales: Body is cylindrical, anteriorly it tapers slightlyand posteriorly it ends with a filiform tail. Cuticu-lar punctations starts at the anterior most level of theamphids; the dots are arranged in regular transverserows. Laterally (5–7 �m diameter), the dots are largerthan on the dorsal and ventral sides. There are four lon-gitudinal rows of somatic setae (6–8�m long) that startimmediately posterior of the amphids at dorso-lateraland ventro-lateral positions and borders the lateral dif-ferentiation area (Figure 5F).
The amphids are a simple spiral, 5–6 �m in diam-eter or 45–56% cbd, located at 8–10 �m from theanterior end. Inner labial sensilla are inconspicuous,the outer are 3 �m long and located at the same levelas the cephalic ones which are 10–15 �m (1.5–2.0 hd)long (Figure 5C) (in most specimens the anterior sen-silla are broken off).
Stoma is long (7–8 �m), anterior part has teeth andposterior part has sclerotized walls. The pharyngealmuscles surround most of the stoma. The pharynx iscylindrical, 71–87�m long and swollen at the terminalend to form a bulb which has several interruptions.Cardia is small (Figure 5A).
The reproductive system is monorchic with out-stretched testis located to the right of the intestine; ashort germinal zone and large spermatozoa fill the restof the testis; the vas deferens is short (Figure 5E). Thespicules are curved on the proximal end and pointedon the distal end. A pair of pre-cloacal setae is presentat 4–5 �m from the cloaca (Figure 5H).
The tail is conical with a long filiform end. Females:They are similar to males in all aspects (Figures B & D).The reproductive system is amhpidelphic with reflexedovaries.
Differential diagnosisAcantholaimus gathumai sp.n. is characterised bypunctated cuticle with lateral differentiation (5–7 �min diameter) of larger dots, long (6–8�m long) somaticand cephalic (10–15 �m long) setae.
Acantholaimus gathumai sp.n. resembles Acantho-laimus arminius Gerlach, Shrage & Riemann, 1979,A. minutus Vitiello 1970, A. quintus Gerlach, Shrage& Riemann, 1979 and Acantholaimus geraerti sp.n. inthe shape of the stoma.
It differs from A. arminius in the position of theamphids which are located within the stoma region andthe body size (L0 = 1400–1600 �m) in A. arminius. Itcan be distinguished from A. minutus in that cephalicsetae are only 6 �m long compared to 10–15 �m inthe new species and somatic setae are very short inA. minutus. Acantholaimus gathumai sp.n. differs fromA. quintus in that A. quintus has at least three teeth inthe stoma and several setae at the pharyngeal region.A. gathumai can be distinguished from A. geraerti inthe lateral differentiation:Acantholaimus gathumai hasa wider (5–7 �m) lateral differentiation although it isa much thinner species (M = 15–19 �m compared toM = 29–32 �m and lateral differentiation is 4–7 �mand more conspicuous in Acantholaimus geraerti n.sp.
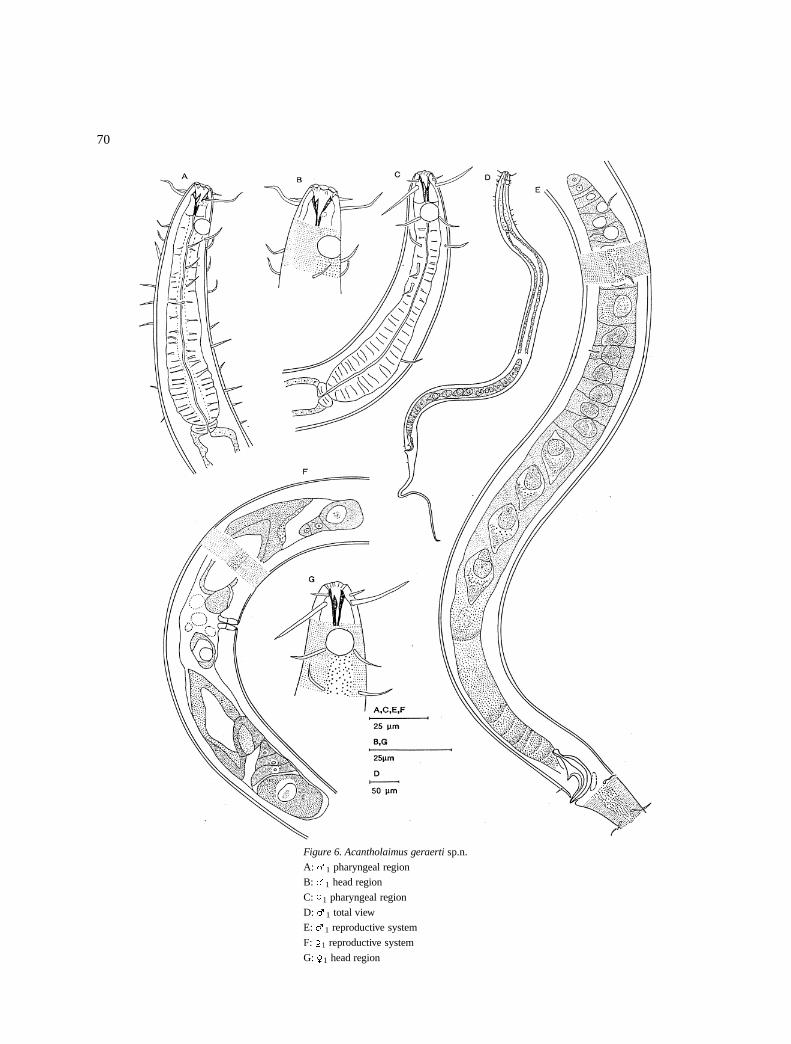
Acantholaimus geraerti sp.n. (Figure 6)
Type materialFour males and three females on slide numbers R1528�(1), R1529 (�1) and RUG-10357-10361
hydr3748.tex; 24/06/1997; 18:18; v.7; p.10
69
Figure 5. Acantholaimus gathumai sp.n.
A: �1 pharyngeal region
B: �1 head region
C: �1 head region
D: �1 pharyngeal region
E: �1 reproductive system
F: �1 total view
G: �1 tail
H: �1 spicule
hydr3748.tex; 24/06/1997; 18:18; v.7; p.11
70
Figure 6. Acantholaimus geraerti sp.n.
A: �1 pharyngeal region
B: �1 head region
C: �1 pharyngeal region
D: �1 total view
E: �1 reproductive system
F: �1 reproductive system
G: �1 head region
hydr3748.tex; 24/06/1997; 18:18; v.7; p.12

71
EtymologyName given for Prof. E. Geraert of the University ofGhent.
Type localityMales from sts. 505 (1), 518 (1), 533 (2 includingholotype)
Females from sts. 506 (1), 518 (allotype), 533 (1)
Measurements�1 – 102 M 623
85312 27 29 20
a: 29.4; b: 8.4; c:3.7; spic: 33�1 – 114 460 635
92512 26 30 19
a: 30.8; b: 8.1; c: 3.2; V: 50%; V0: 72%Paratype �� (Tails broken in two males) L: 858
L0: 551–651; a: 26.8; b: 8.5; c: 2.8: spic: 27–35Paratype �� (Tail broken in one female) L: 953; L0:
595–637; a: 29.8; b:8.5; c: 3.0; V: 49%; V0: 72–73%
DescriptionMale: The body is cylindrical, tapers slightly to theanterior end and a filiform tail end (Figure 6D). Cuti-cle is punctated with punctations starting from anteriorlevel of the amphids, the punctations are arranged intransverse rows with lateral differentiation having larg-er and more conspicuous dots than on the rest of thebody (Figure 6B); the lateral differentiation is 6 �m atthe pharyngeal region and 4–5 �m wide at mid body(Figure 6E). Somatic setae are (8–12 �m long) in fourlongitudinal rows; two in the dorso-lateral and two inthe ventro-lateral positions and bordering the lateraldifferentiation.
The amphids are spiral with a single turn locatedat 8–12 �m from the anterior; they are 6–8 �m indiameter (40–50% cbd). The inner labial sensilla areinconspicuous, the outer labial are 5�m long and at thesame level as the cephalic setae which are 15–19 �mlong (Figure 6B).
The stoma is 12–17 �m long and it has three teeth.Pharynx is cylindrical (101–114 �m), and anteriorlysurrounds the stoma; posteriorly it is expanded to formthe terminal bulb. The cardia is small but prominent(Figure 6A).
Reproductive system is monorchic with outstretch-ed testis. The spermatozoa are large, rectangular inshape and they have large nuclei. Vas deferens is short.The spicules are (1.4–1.7 cbd) curved proximally and
rounded distally. There is a pair of pre-cloacal setae at6 �m from the cloaca (Figure 6E).
Females: They are similar to males in all aspects(Figures 6C & G). The reproductive system isamphidelphic with reflexed ovaries, anterior branchto the right and the posterior one to the left of the intes-tine. There is a short germinal zone followed by growthzone which may contain only a single developed ova.The uterus is filled with large sperm cells. The vaginais thick walled and vulva is simple (Figure 6F).
Differential diagnosisAcantholaimus geraerti sp.n. is characterised by longcephalic sensilla (15–19 �m), well developed teeth inthe stoma, narrow (4–6 �m) and conspicuous lateraldifferentiation.
Acantholaimus geraerti sp.n. resembles Acantho-laimus gathumai sp.n., A. arminius Gerlach, Shrage& Riemann, 1979 and A. calathus Gerlach, Shrage &Riemann, 1979 in the stoma shape.
Acantholaimus geraerti can be distinguished fromAcantholaimus gathumai by the body size (L0 = 363–423, M = 14–19 �m in Acantholaimus gathumai andL0 = 551–651, M = 29–32 �m in Acantholaimus ger-aerti) and lateral differentiation which is narrow (4–5 �m wide) and more conspicuous in A. geraerti sp.n.compared to A. gathumai (5–6 �m wide) althoughA. geraerti is the larger of the two species. It canbe distinguished from A. arminius by the positionof the amphids which are within the stoma regionin A. arminius and the wide lateral differentiation.Acantholaimus geraerti differs from A. calathus bythe absence of longitudinal rows of dots present in thelatter.
Acantholaimus invaginatum sp.n. (Figure 7)
Type materialSix males and seven females on slide numbers R1530� 1), R1531 (�1) and RUG-10362-10370
Type localityMales from sts. 105 (1), 107 (1), 133 (1), 505 (holo-type), 507 (1) and 519 (1)
Females from sts. 133 (2), 505 (3 including allo-type), 519 (1) and 533 (1)
hydr3748.tex; 24/06/1997; 18:18; v.7; p.13
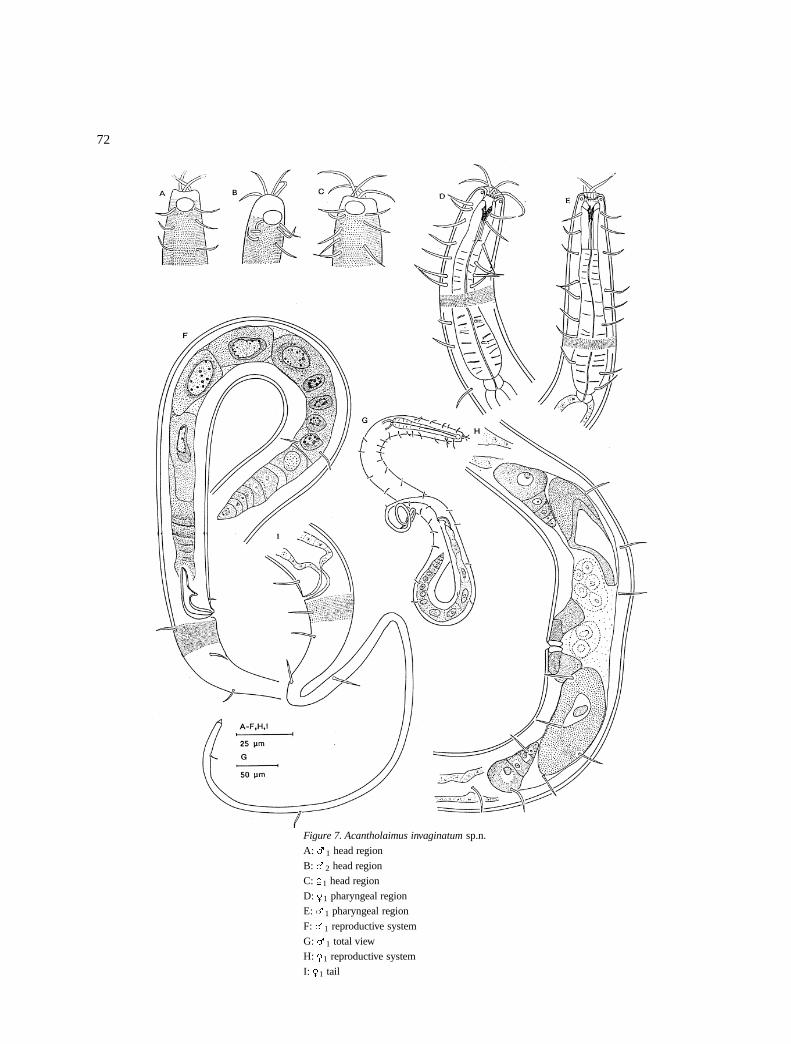
72
Figure 7. Acantholaimus invaginatum sp.n.
A: �1 head region
B: �2 head region
C: �1 head region
D: �1 pharyngeal region
E: �1 pharyngeal region
F: �1 reproductive system
G: �1 total view
H: �1 reproductive system
I: �1 tail
hydr3748.tex; 24/06/1997; 18:18; v.7; p.14

73
Measurements�1 – 90 M 530
78212 24 29 19
a: 27; b: 8.7; c: 3.1; spic: 32�1 – 88 419 558
86115 28 34 21
a: 25.3; b: 9.8; c: 2.8; V: 49%; V0:75%Paratype �� L: 733–1019; L0: 512–735; a: 27.8–
39.2; b: 8.5–9.6; c:3.2–3.5; spic: 24–32Paratype �� L: 711–892; L0: 512–642; a: 24.5–
28.9; b: 8.5–9.7; c: 3.3–3.6; V: 49–52%; V0: 70–75%
DescriptionMales: Body is cylindrical, anterior end is blunt andposterior end has a long filiform tail (Figure 7H). Cuti-cle is punctated from the anterior level of the amphids.Punctations are in regular transverse rows. Laterally,they are larger than on the dorsal and ventral sides. Thelateral differentiation region is 10–12 �m on the pha-ryngeal region (Figure 7A) and mid-body, and 8 �m atthe anterior tail region (Figure 7F). Somatic setae [7–12 �m long at pharyngeal (Figure 7E) and 9–13 �m attail region (Figure 7F)] are in four longitudinal rows:two in dorso-lateral and two in ventro-lateral positions.
Amphids are circular, 5–8 �m (38–66% cbd) indiameter and located at 3–5 �m from the anteriorend (NB the location of the amphids from the ante-rior maybe more than what is measured here whenthe stoma is not invaginated). Cephalic setae are long(16–21 �m or 1.3–1.4 hd) (Figure 7B). Stoma (20–22 �m long) has 3 (4?) teeth and often it is invaginated(Figure 7E).
Pharynx is cylindrical (84–106�m long),anteriorlysurrounds the stoma and posteriorly it is swollen toform the terminal bulb (Figure 7E).
The reproductivesystem is monorchic with an ante-riorly outstretched testis located to the right of the intes-tine. The spermatozoa are large rectangular in shapeor elongate. Vas deferens is short. The spicules arecurved, 1.2–1.7� abd long (Figure 7F).
Tail is short conical (35–54 �m long) and longposterior filiform part.
Females: They are similar to the males in mostaspects (Figures 7A, D & I). Reproductive system isamphidelphic with reflexed ovaries, anterior to theright, posterior to the left of the intestines. Vulva issimple and vagina is 6–7 �m long and thick walled(Figure 7H).
Differential diagnosisAcantholaimus invaginatum sp.n. is characterised bycuticle with lateral differentiation of larger dots, longcephalic (16–21 �m) and somatic (7–13 �m) setae.Circular amphids (38–64% cbd) located close to theanterior end. Long stoma, with three teeth and ofteninvaginated.
Acantholaimus invaginatum sp.n. resemblesA. maks Gerlach, Shrage & Riemann, 1979 in the ante-rior region (invaginated stoma) and the arrangement ofthe somatic setae at the pharyngeal region; it resem-bles A. polydentatus Gerlach, 1951 and A. calathusGerlach, Shrage & Riemann, 1979 in the size of thecephalic setae and the arrangement of the somatic setaeand A. quintus Gerlach, Shrage & Riemann, 1979 in thearrangement of the somatic setae and the well devel-oped teeth in the stoma.
A. invaginatum sp.n. differs from A. maks in theabsence of a pair of post amphidial setae on eitherside of the amphids and the length of the somaticsetae which are rather short in A. maks. It differsfrom A. polydentatus and A. calathus in the absenceof two longitudinal rows of larger dots on the lateralsides which are present in these two species, and fromA. quintus in the length of the cephalic setae: 11 �mor 0.6� hd sensu Gerlach et al., 1979, 15 �m sen-su Gourbault & Vincx 1985 and 8–11 �m or 0.8� hdsensu Vivier 1985).
Distribution of Acantholaimus species within the fourtransects of the Indian OceanIn deep-sea studies the family Chromadoridae is one ofthe most dominant families reported (Dinet & Vivier,1979; Thistle & Sherman, 1985; Tietjen, 1989) and thegenus Acantholaimus, one of the typical deep-sea gen-era (Soetaert & Heip, 1995) is almost always present inhigh numbers in these sediments (Thistle & Sherman,1985; Tietjen, 1989).
Although some work is done on deep-sea nema-todes with identification to the genera level, only a fewstudies have dealt with assemblages at the species lev-el. However, available information points to an exis-tence of low population density and high number ofspecies in deep-sea sediments compared to shallowerhabitats. Co-existence of several species of the samegenus is common. Thistle & Sherman (1985) com-pared the number of species present per genera in theGulf of Gascony (Dinet & Vivier, 1979), VenezuelanBasin (Tietjen) and the Scotian Rise (Thistle & Sher-
hydr3748.tex; 24/06/1997; 18:18; v.7; p.15

74
man, 1985). The highest reported number of congener-ic species is 42 of Theristus in the Gulf of Gascony.
In the present study we report 38 species (morpho-types) of Acantholaimus from the four transects of theIndian Ocean studied during the two sampling cruises.A number of species occurred in more than one station,and at least 13 species were encountered only once.Soetaert & Heip (1990), argue that, for rare species,the higher the number of individuals observed, thehigher the number of species that may be encoutered.In this case, it seems also true that the more stationswere investigated the more the number of rare speciesof Acantholaimus were encountered.
Some 3–18 different species (morphotypes) ofAcantholaimus were found co-existing in one station(Tables 2 and 3). In the Gulf of Gascony, Dinet &Vivier (1979) reported six species, Thistle & Sher-man (1985) reported seven species and Tietjen (1989),reported twelve species of Acantholaimus in the PuertoRico Area and Hatteras Abyssal Plain. How is it pos-sible that such a large number of different species ofthe same genus can co-exist together? Jensen (1988a)recognised four species of Acantholaimus co-existingin one station in the Norwegian sea. He attributed thisability of co-existence to their different types of mouthopenings and buccal cavities which indicate a foodresource partitioning. We also find a similar variationin the buccal cavities of the seven species of Acantho-laimus described in this paper which were often foundtogether in some of the stations.
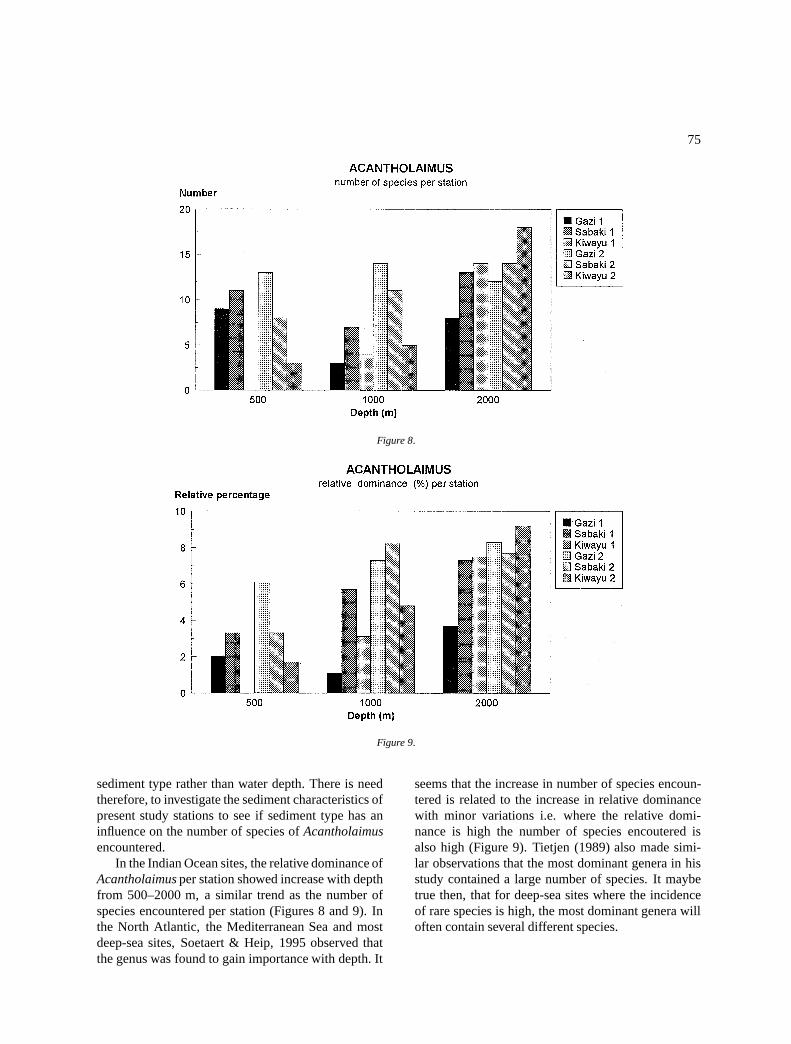
What factors are likely to influence the number ofspecies encountered per station? The number of indi-viduals observed per station (Soetaert & Heip, 1990)may explain the high numbers of species in some sta-tions like 105 and 505. In spite of the difference inthe number of individuals observed per station in ourstudy, there is a tendency of the number of speciesencountered to increase with depth from 500–2000 m(Figure 8). Tietjen (1976) studied nematode speciesdistribution with depth in North Carolina and foundthat species diversity (diversity is largely a functionof species richness) decreased with water depth. Inhis study area, water depth was related to sedimenttype where shallower stations had sandy sediments anddeeper stations had clayey-silt sediments. He arguedthat sandy sediments provide higher microhabitats andthereby a higher diversity. Tietjen (1984) also foundspecies richness and dominance to be related to grainsize distribution and organic input in three stationsin the Venezuelan Basin. It is possible then, that theincrease in species number with depth is in response to
Table 2. Acantholaimus species identified from the IndianOcean from the first cruise A1 (Jun-Jul/ 1992) (N. observed=total number of individuals observed, A. observed= Acantho-laimus individuals observed, z= individuals not identified)
Station 105 106 107 117 118 119 133 136
Depth 500 1000 2000 500 1000 2000 2000 1000
N.observed 884 281 269 479 159 358 535 163
A.observed 18 3 10 17 9 26 41 5
A. vermeuleni 2 4 1 5 3 2
A. verscheldi 1 2 2 3 1
A. heipi 1 1 3 4
A. elegans 3 1 3 2
A. gathumai 2 1 1 4 3
A. geraerti
A.invaginat 2 1 5
A. sp. 1 1 2 1
2 4
3
4 4 1 1 1
5 1 1 1
6 1
7 1 1
8 1 1 1 2 2
9
10 1
11 1 3
12 1 1
13
14
15 2 1 2 1 3
16 1 1
17
18 1 1
19
20 2 1
21
22 1
23
24 1 3
25 1
26
27 1
28
29 4
30 1
z 1 2
No. of species 9 3 8 11 7 13 14 4
hydr3748.tex; 24/06/1997; 18:18; v.7; p.16
75
Figure 8.
Figure 9.
sediment type rather than water depth. There is needtherefore, to investigate the sediment characteristics ofpresent study stations to see if sediment type has aninfluence on the number of species of Acantholaimusencountered.
In the Indian Ocean sites, the relative dominance ofAcantholaimus per station showed increase with depthfrom 500–2000 m, a similar trend as the number ofspecies encountered per station (Figures 8 and 9). Inthe North Atlantic, the Mediterranean Sea and mostdeep-sea sites, Soetaert & Heip, 1995 observed thatthe genus was found to gain importance with depth. It
seems that the increase in number of species encoun-tered is related to the increase in relative dominancewith minor variations i.e. where the relative domi-nance is high the number of species encoutered isalso high (Figure 9). Tietjen (1989) also made simi-lar observations that the most dominant genera in hisstudy contained a large number of species. It maybetrue then, that for deep-sea sites where the incidenceof rare species is high, the most dominant genera willoften contain several different species.
hydr3748.tex; 24/06/1997; 18:18; v.7; p.17

76
Table 3. Acantholaimus species identified from the Indian Ocean fromthe second cruise A2 (Nov.-Dec./ 1992) (Abbreviations are the sameas in table 2).
Station 505 506 507 517 518 519 531 532 533 552
Depth 500 1000 2000 500 1000 2000 500 1000 2000 500
No. observed 739 343 302 371 294 323 353 188 337 277
A. observed 45 25 25 12 24 25 6 9 31 19
A. vermeuleni 7 2 2 1 4 3 1 3 3 1
A. verscheldi 5 2 4 2 1 1 1 4
A. heipi 2 2 1
A. elegans 5 2 1 1 3 2 2
A. gathumai 5 3 2 1 3 2 2 3 1
A. geraerti 1 1 2 3 1
A. invaginat 8 2 4 1 2 1
A. sp. 1 3 3
A. sp. 2 2 2
A. sp. 3 2 2
4 1 1
5 3 2 1
6 1 2 1 2 1
7 3 1 1 3 2 3
8 1 1 1
9 1
10 1
11
12 1 1 1 1
13 1 1
14 2
15 3 3 2 2 1 3
16 1 2
17 1
18 2
19 1
20 1 1
21 1 1 1 2
22
23 2
24 2
25 1 1
26 2 1 1 1
27
28 2 1
29
30
z 1 1 1 3 1 1 1
No of species 13 14 12 8 11 14 3 5 18 11
Acknowledgments
The authors would like to thank the following organisa-tions who made it possible for this work to be complet-
ed: Algemeen Bestuur voor de Ontwikkelingssamen-werking (ABOS, Belgium), Netherlands Indian OceanProgramme 1990–1995, Kenya Marine & FisheriesResearch Institute in Mombasa (KMFRI) and ResearchProject 2 0086 96 of the National Science Foundationin Belgium. Sincere gratitudes to Rita Van Driessche,Guy and Mirriam for their technical help. Thanks alsoto Christ Heino for helping in translation of some Ger-man manuscripts. Our sincere thanks also to Dr JohnTietjen for reading the manuscript.
References
Dinet, A. & M. H. Vivier, 1979. Le meiobenthos Abyssal du Gulfede Gascogne. II Les peuplements de nematodes et leur diversitespecifique. Cah. Biol. mar. 20: 109–123.
Gerlach, S. A., M. Shrage & F. Riemann, 1979. Die GattungAcantholaimus (Nematoda, Chromadoridae), und Beobachtun-gen uber einen mutmaßlichen Transportmechanimus fur Sper-mien bei A. calathus sp.n. In Veroff, Inst. Meeresforschung Bre-merhaven 18: 35–67.
Gourbault, N. & M. Vincx, 1985. Nematodes abyssaux du cam-pagne Walda du NØ‘Jean Charcot’. VI. Le genre Acantholaimus:especes nouvelles et etude de l‘appareil reproducteur a spermato-zoides geants. Bull. Mus. natn. Hist. nat. Paris 4e ser 7: 611–632.
Jensen, P., 1988. Four new nematode species abundant in the deep-sea benthos of the Norwegian Sea. Sarsia 73: 149–155.
Jensen, P., 1988a. Nematode assemblages in the deep-sea benthosof the Norwegian Sea. Deep-Sea Res. 35: 1173–1184.
Muthumbi, A. W., K. Soetaert & M. Vincx, 1997. Deep-sea nema-todes from the Indian Ocean: new and known species of thefamily Comesomatidae. Hydrobiologia 346: 25–57.
Soetaert, K. & C. Heip, 1990. Sample-size dependence of diversityindices and the determination of sufficient sample size in a high-diversity deep-sea enviroment. Mar. Ecol. Prog. Ser. 59: 305–307.
Soetaert, K. & C. Heip, 1995. Nematode assemblages of deep-seaand shelf break sites of the North Atlantic and the MediterraneanSea. Mar. Ecol. Prog. Ser. 125: 171–183.
Thistle, D. & K. M. Sherman, 1985. The nematode fauna of a deep-sea site exposed to strong near-bottom currents. Deep-Sea Res.32: 1077–1088.
Tietjen, J. H., 1976. Distribution and species diversity of deep-seanematodes off North Carolina. Deep-Sea Res. 23: 755–768.
Tietjen, J. H., 1984. Distribution and species diversity of deep-seanematodes in the Venezuela Basin. Deep-Sea Res. 31: 119–132.
Tietjen, J. H., 1989. Ecology of deep-sea nematodes from the PuertoRico Trench area and Hatteras Abyssal Plain. Deep-Sea Res. 36:1579–1594.
Vivier, M. H., 1985. Especes du genre Acantholaimus (Nematoda,Chromadoridae, Spilipherinae). Peuplements profonds du golfede Gascogne L. Laubier & Cl. Monniot (eds). Ifremer: 331–349.
hydr3748.tex; 24/06/1997; 18:18; v.7; p.18




















