
AAV-mediated chronic over-expression of SNAP25 in adult rat dorsal hippocampus impairs...
14
,1 ,1 ,1 ,2 ,1 ,1 *Applied Neurotherapeutics Research Group, Smurfit Institute of Genetics, Trinity College Dublin, Dublin, Ireland Applied Neurotherapeutics Research Group, UCD School of Biomolecular and Biomedical Science, UCD Conway Institute, University College Dublin, Belfield, Dublin, Ireland àFlow Cytometry Core Facilities, UCD Conway Institute, University College Dublin, Belfield, Dublin, Ireland §Discovery Neuroscience, Wyeth Research, Princeton, New Jersey, USA The ternary soluble N-ethylmaleimide-sensitive factor attach- ment protein receptor (SNARE) complex, the minimal machinery required for vesicle exocytosis, consists of the 2 Present address of William T. O’Connor is the Graduate School of Medicine, University of Limerick, Limerick, Ireland. Abbreviations used: AAV, adenoassociated virus; aCSF, artificial CSF; ADHD, attention deficit hyperactivity disorder; eGFP, enhanced green fluorescent protein; EPSP, excitatory post-synaptic potential; FACS, fluorescent-activated cell sorting; HEK293 cells, human embryonic kidney cells; hrGFP, humanized renilla green fluorescent protein; IRES, internal ribosome entry site; PBS, phosphate-buffered saline; PPF, paired-pulse facilitation; PPI, pre-pulse inhibition; rAAV, recombinant AAV; SNAP-25, synaptosomal protein of 25 kDa; SNARE, soluble N- ethylmaleimide-sensitive factor attachment protein receptor; SSC, saline sodium citrate buffer. Received August 18, 2009; revised manuscript received November 10, 2009; accepted November 22, 2009. Address correspondence and reprint requests to Alex G. McKee, Applied Neurotherapeutics Research Group, Smurfit School of Genetics, Trinity College Dublin, Dublin 2, Ireland. E-mail: [email protected] 1 These authors contributed equally to this study. Abstract Long-term memory is formed by alterations in glutamate- dependent excitatory synaptic transmission, which is in turn regulated by synaptosomal protein of 25 kDa (SNAP-25), a key component of the soluble N-ethylmaleimide-sensitive factor attachment protein receptor complex essential for exocytosis of neurotransmitter-filled synaptic vesicles. Both reduced and excessive SNAP-25 activity has been implicated in various disease states that involve cognitive dysfunctions such as attention deficit hyperactivity disorder, schizophrenia and Alzheimer’s disease. Here, we over-express SNAP-25 in the adult rat dorsal hippocampus by infusion of a recombinant adenoassociated virus vector, to evaluate the consequence of late adolescent-adult dysfunction of the soluble N-ethylma- leimide-sensitive factor attachment protein receptor protein in the absence of developmental disruption. We report a specific and significant increase in the levels of extracellular glutamate detectable by microdialysis and a reduction in paired-pulse facilitation in the hippocampus. In addition, SNAP-25 over- expression produced cognitive deficits, delaying acquisition of a spatial map in the water maze and impairing contextual fear conditioning, both tasks known to be dorsal hippocampal dependent. The high background transmission state and pre- synaptic dysfunction likely result in interference with requisite synapse selection during spatial and fear memory consolida- tion. Together these studies provide the first evidence that excess SNAP-25 activity, restricted to the adult period, is sufficient to mediate significant deficits in the memory formation process. Keywords: adenoassociated virus-mediated gene delivery, fear conditioning, paired-pulse facilitation, synaptosomal protein of 25 kDa, synaptic vesicle release, water maze. J. Neurochem. (2010) 112, 991–1004. JOURNAL OF NEUROCHEMISTRY | 2010 | 112 | 991–1004 doi: 10.1111/j.1471-4159.2009.06516.x ȑ 2009 The Authors Journal Compilation ȑ 2009 International Society for Neurochemistry, J. Neurochem. (2010) 112, 991–1004 991
Transcript of AAV-mediated chronic over-expression of SNAP25 in adult rat dorsal hippocampus impairs...

,1 ,1 ,1
,2 ,1
,1
*Applied Neurotherapeutics Research Group, Smurfit Institute of Genetics, Trinity College Dublin, Dublin, Ireland
�Applied Neurotherapeutics Research Group, UCD School of Biomolecular and Biomedical Science, UCD Conway Institute,
University College Dublin, Belfield, Dublin, Ireland
�Flow Cytometry Core Facilities, UCD Conway Institute, University College Dublin, Belfield, Dublin, Ireland
§Discovery Neuroscience, Wyeth Research, Princeton, New Jersey, USA
The ternary soluble N-ethylmaleimide-sensitive factor attach-ment protein receptor (SNARE) complex, the minimalmachinery required for vesicle exocytosis, consists of the
2Present address of William T. O’Connor is the Graduate School ofMedicine, University of Limerick, Limerick, Ireland.Abbreviations used: AAV, adenoassociated virus; aCSF, artificial CSF;
ADHD, attention deficit hyperactivity disorder; eGFP, enhanced greenfluorescent protein; EPSP, excitatory post-synaptic potential; FACS,fluorescent-activated cell sorting; HEK293 cells, human embryonickidney cells; hrGFP, humanized renilla green fluorescent protein; IRES,internal ribosome entry site; PBS, phosphate-buffered saline; PPF,paired-pulse facilitation; PPI, pre-pulse inhibition; rAAV, recombinantAAV; SNAP-25, synaptosomal protein of 25 kDa; SNARE, soluble N-ethylmaleimide-sensitive factor attachment protein receptor; SSC, salinesodium citrate buffer.
Received August 18, 2009; revised manuscript received November 10,2009; accepted November 22, 2009.Address correspondence and reprint requests to Alex G. McKee,
Applied Neurotherapeutics Research Group, Smurfit School of Genetics,Trinity College Dublin, Dublin 2, Ireland. E-mail: [email protected] authors contributed equally to this study.
Abstract
Long-term memory is formed by alterations in glutamate-
dependent excitatory synaptic transmission, which is in turn
regulated by synaptosomal protein of 25 kDa (SNAP-25), a
key component of the soluble N-ethylmaleimide-sensitive
factor attachment protein receptor complex essential for
exocytosis of neurotransmitter-filled synaptic vesicles. Both
reduced and excessive SNAP-25 activity has been implicated
in various disease states that involve cognitive dysfunctions
such as attention deficit hyperactivity disorder, schizophrenia
and Alzheimer’s disease. Here, we over-express SNAP-25 in
the adult rat dorsal hippocampus by infusion of a recombinant
adenoassociated virus vector, to evaluate the consequence of
late adolescent-adult dysfunction of the soluble N-ethylma-
leimide-sensitive factor attachment protein receptor protein in
the absence of developmental disruption. We report a specific
and significant increase in the levels of extracellular glutamate
detectable by microdialysis and a reduction in paired-pulse
facilitation in the hippocampus. In addition, SNAP-25 over-
expression produced cognitive deficits, delaying acquisition of
a spatial map in the water maze and impairing contextual fear
conditioning, both tasks known to be dorsal hippocampal
dependent. The high background transmission state and pre-
synaptic dysfunction likely result in interference with requisite
synapse selection during spatial and fear memory consolida-
tion. Together these studies provide the first evidence that
excess SNAP-25 activity, restricted to the adult period, is
sufficient to mediate significant deficits in the memory
formation process.
Keywords: adenoassociated virus-mediated gene delivery,
fear conditioning, paired-pulse facilitation, synaptosomal
protein of 25 kDa, synaptic vesicle release, water maze.
J. Neurochem. (2010) 112, 991–1004.
JOURNAL OF NEUROCHEMISTRY | 2010 | 112 | 991–1004 doi: 10.1111/j.1471-4159.2009.06516.x
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2010) 112, 991–1004 991

synaptosomal protein of 25 kDa (SNAP-25), a pre-synapticplasma membrane protein, together with synaptobrevin/vesicle-associated membrane protein and syntaxin-1A (Soll-ner et al. 1993; Wang and Tang 2006). The SNARE complexhas been repeatedly implicated in cognitive functions such asmemory formation and sensory processing (Sollner et al.1993; Hou et al. 2004, 2006; Gosso et al. 2006; O’Sullivanet al. 2007). Indeed, selective inhibition of SNAP-25 usingantisense technology has been shown to prevent neuriteoutgrowth and elongation, a morphological correlate ofmemory consolidation in the hippocampus (Osen-Sand et al.1993; O’Malley et al. 1998, 2000; Eyre et al. 2003). More-over, recent in vitro evidence in primary hippocampal cultureand in brain slices suggests that SNAP-25 is expressedpredominantly in the glutamatergic, and not GABAergic,synapses of hippocampal neurons (Verderio et al. 2004).These studies used knockdown and knock-in technologies todemonstrate that decreased and increased SNAP-25 expres-sion augments and depresses neuronal calcium responsivenessrespectively. In vivo, up- and down-regulation of SNAP-25mRNA expression has been noted to accompany hippocampallong-term potentiation and passive avoidance learning,respectively, the former a widely accepted physiologicalmodel of memory-associated synaptic plasticity (Robertset al. 1998; Hou et al. 2004, 2006; O’Sullivan et al. 2007).
There is now substantial evidence that altered SNAP-25activity is associated with cognitive impairments seen inseveral disorders including attention deficit hyperactivitydisorder (ADHD) and schizophrenia. In humans, SNAP-25maps to 20p11.2 and a number of recent studies haveshown a possible association with ADHD (Barr et al. 2000;Brophy et al. 2002; Mill et al. 2002, 2004; Kustanovichet al. 2003). Moreover, the coloboma mouse mutant carriesa heterozygous deletion spanning four genes includingSNAP-25 and displays a hyperactivity phenotype (Hesset al. 1996). A meta-analysis of 20 separate genome-widelinkage studies for schizophrenia-associated genes reporteda significant linkage to 20p12.3-11 (an interval containingno other candidate susceptibility loci), suggesting thisinterval, and SNAP-25 in particular, to be important forthe aetiology of this complex psychiatric illness (Lewiset al. 2003). Recently, a dominant SNAP-25 mutation in theso-called blind-drunk (Bdr) mouse with a severe ataxia hasbeen characterized. This mutation, in a highly conserveddomain of SNAP-25, results in increased SNARE bindingaffinities, impairment of the exocytotic vesicle recyclingprocess, reduced glutamate release and a reduction inevoked cortical excitatory post-synaptic potentials (EPSPs;Jeans et al. 2007). This mouse also exhibits impairment insensorimotor gating and high anxiety behaviour, aspectsassociated with schizophrenia. It should also be noted thatthere is evidence from several studies utilizing immuno-histochemistry and western blot analysis of postmortembrains implicating a reduction in SNAP-25 expression in
the prefrontal cortex of schizophrenics (Mukaetova-Ladi-nska et al. 2002; Thompson et al. 2003). This may,however, reflect overall synaptic loss in the prefrontalcortex, a relatively late feature of the disease (Garey et al.1998; Glantz and Lewis 2000; Kolluri et al. 2005). Finally,human linkage studies have recently identified singlenucleotide polymorphisms in the SNAP-25 gene that arehighly correlated with measures of intelligence in the Dutchpopulation (Gosso et al. 2006). Specifically, variance in onesingle nucleotide polymorphism located in the 5¢-untrans-lated region of the SNAP-25 gene was shown to contribute3.4% of the phenotypic variation on performance IQcomponents of the Wechsler Adult Intelligence Scale III-Revised (Wechsler 1997; Gosso et al. 2006).
The contribution of developmental upset and regionallyspecific disruption to the motor and cognitive deficitsassociated with SNAP-25 activity remains unclear. Here,we have deployed recombinant adenoassociated virus(AAV)-mediated gene delivery in vivo to evaluate thefunctional consequences of increased SNAP-25 expressionrestricted in space to the dorsal hippocampus and in time tothe late adolescent–adult period of life. Specifically, we haveevaluated the consequence of dorsal hippocampal SNAP-25overactivity on short-term plasticity and transmission tone asmeasured by paired-pulse facilitation (PPF) and glutamatelevels respectively. Finally, we establish the effects of SNAP-25 over-expression on a range of cognitive-associatedbehaviours selected to test functionality in different subre-gions of the hippocampus, prefrontal cortex and motorcircuitry. Taken together, the results suggest that overactivityof SNAP-25 in the late adolescent-adult brain reduces thesignal-to-noise ratio and potential for synaptic plasticitywithin circuitry controlling spatial and fear memory consol-idation resulting in cognitive deficits akin to those observedin schizophrenia, ADHD and other disorders of higher brainfunction.
Materials and methods
PlasmidsThe SNAP-25b isoform was amplified from a cDNA library
generated from hippocampal RNA using Pfu Turbo Taq polymerase
(Stratagene, La Jolla, CA, USA) and oligonucleotides containing a
BamHI restriction site (forward) 5¢-CGGGGATCCATGGCCGAG-GACGCAGA-3¢ and a XhoI restriction site (reverse) 5¢-CCGC-TCGAGTTAACCACTTCCCAGCATCTT-3¢. The SNAP-25 cDNAwas cloned into the pAAV-IRES-hrGFP (internal ribosome entry site-
humanized renilla green fluorescent protein) plasmid (Stratagene),
flanked by the inverted terminal repeats of AAV2. pAAV-cytomega-
lovirus-enhanced GFP (eGFP); eGFP was excised from pEGFP-1
(Clontech, Mountainview, CA, USA) and blunt cloned into pAAV-
MCS (Stratagene). All cloning steps were verified by Sanger
sequencing using an ABI310 genetic analyser and BIGDYEV3.1
chemistries (Applied Biosystems, Foster City, CA, USA).
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2010) 112, 991–1004� 2009 The Authors
992 | A. G. McKee et al.

AAV Production of AAV-SNAP-25-IRES-hrGFPRecombinant AAV2/5 (rAAV2/5) viruses were generated by helper
virus free, triple transfection based on the method of Xiao et al.(1998). Human embryonic kidney (HEK293) cells (accession
number CRL-1573; ATCC, Manassas, VA, USA) were transfected
with pAAV-SNAP-25-IRES-hrGFP or control pAAV-eGFP, pRep/
Cap5 (Hildinger et al. 2001) and pHelper (Stratagene) at a ratio of
1 : 1 : 2, as previously described (O’Reilly et al. 2007). A rAAV
vector expressing SNAP-25 and hrGFP and a control vector
expressing eGFP were constructed and packaged in HEK293T.
The expression cassettes for both vectors contained a cytomegalo-
virus promoter, a b-globin intron and the human growth hormone
polyadenylation signal. The SNAP-25 expressing vector also
included an IRES linked to the hrGFP (Fig. 1a). Genomic titres
(viral particles/mL) were determined by quantitative real-time PCR
(Rohr et al. 2002). Both rAAV vectors used in this study had similar
genomic titres (�8 · 1012 and �5 · 1012 viral genomes per mL for
rAAV/SNAP-25 and rAAV/control respectively). High-titre virus
(> 1012 viral particles/mL) was used to ensure maximum transfec-
tion with a low volume (1 lL) infusion.
In vitro studiesHuman embryonic kidney 293 cells were seeded in 2 mL Dulbecco’s
modified Eagle’s medium containing 10% foetal calf serum at
1 · 106 cells per well in a six-well plate and incubated for 3 h at
37�C. Media was then aspirated and the cells were washed twice with
phosphate-buffered saline (PBS). AAV/SNAP-25 or AAV/Control
was then added to each well, at a multiplicity of infection of 2500, in
350 lL PBS. After incubation for 1 h at 37�C, 1 mL of media was
added, and the cells were incubated overnight. Cells were then
washed with PBS, fixed with 4% p-formaldehyde and counterstained
with 4¢,6-diamidino-2-phenylindole for fluorescence microscopy.
AnimalsMale Wistar rats were obtained from the Biomedical Facility in
University College Dublin (Dublin, Ireland). All animals were
postnatal day 49 on the day of surgery with subsequent testing,
behavioural and microdialysis, beginning at postnatal days 80–84.
Water and food were provided ad libitum, temperature was
maintained at 20–23�C and animals were kept on a 12 h light/dark
cycle regimen (07:00–19:00 h, light). All procedures were approved
by the Animal Research Ethics Committee of University College
Dublin and were carried out by individuals who held the appropriate
licenses issued by the Irish Department of Health.
Site-directed injection to the dorsal hippocampusAnimals were anaesthetized by intraperitoneal administration of a
solution (1 mL/kg i.p.) containing four parts ketamine hydrochloride
(Vetalar�, Pfizer Animal Health, Ringaskiddy, Cork, Ireland)
(100 mg/mL) and three parts medetomidine hydrochloride (Doma-
tor�, Pfizer Animal Health) (20 mg/mL). The depth of anaesthesia
was assessed by monitoring respiratory rate and palpebral and pedal
withdrawal reflexes. Once the animal was fully anaesthetized they
were placed in the stereotaxic frame and the skull was exposed. The
skull was trepanned (1.4 mm in diameter) bilaterally, at a position of
(a)
(b) (c)
(d) (e)
(g)(f)
Fig. 1 Construction and initial validation of the AAV/SNAP-25 delivery
tool. (a) Plasmid map of rAAV vectors. ITR, inverted terminal repeat;
hrGFP, humanized renilla green fluorescent protein; eGFP, enhanced
green fluorescent protein; CMVP cytomegalovirus promoter; BGI,
b-globin intron; IRES, internal ribosome entry site; pA, human growth
hormone polyadenylation signal. (b and c) Fluorescent microscopy
demonstrated delivery of rAAV/SNAP-25 (b) and rAAV/control (c) to
HEK293T cells in vitro. (d) Rat brain atlas at )3.8 mm with respect to
bregma showing position of injection site. (e) rAAV injection mediated
infection as indicated by the GFP expression detectable in cells of the
dentate gyrus and, in particular, the infragranular cells. High-magnifi-
cation images show GFP expression detectable in the processes and
dendrites of infected neural cells in the granule cell layer (f and g).
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2010) 112, 991–1004
Increased SNAP-25 impairs memory-associated synaptic plasticity | 993

3.8 mm posterior and 1.5 mm lateral to bregma (Paxinos and
Watson 1998). A cannula (28 gauge), loaded with either rAAV/
SNAP-25 or rAAV/Control was slowly lowered into the brain to a
depth of 3.5 mm ventral to bregma. The virus was then slowly
injected bilaterally into the brain at a rate of 0.1 lL/min over a
10 min period. After the injection, the cannula was kept in place for
a further 2 min to minimize any drawback of the virus as the
cannula was removed. After the bilateral viral injections, the
incision was closed with interrupted silk sutures and the animal was
placed in a heated cage (35�C) and monitored carefully. The
anaesthetic was reversed by administration of atipamezole hydro-
chloride (Antisedan�, Pfizer Animal Health) (2.5 mg/kg i.p.) and
carprofen (Rimadyl�, Pfizer Animal Health) was administered
(0.1 mL/kg i.p.) as a post-operative analgesic.
In preliminary experiments to optimize stereotaxic injection
procedure, we tested the effect of AAV/control viral injection on
water maze behaviour and compared it with untreated control
animals, vehicle (PBS)-injected controls and animals just exposed to
the anaesthetic. As can be seen in Fig. S1, at 3 weeks post-injection
AAV-treated animals showed a deficit in spatial learning compared
with all other controls but by 5 weeks post-injection control AAV
injected animals performed identically to untreated controls. For this
reason we carried out our behavioural, neurochemical and electro-
physiological studies 5–7 weeks post-injection.
Histological staining and immunohistochemistryFor immunohistochemical analysis, animals were killed by cervical
dislocation, brains were quickly dissected, covered in optimal
cutting temperature compound and snap frozen in CO2-cooled
n-hexane. Coronal sections (12 lm) of the dorsal hippocampus were
cut on a cryostat (HM 500 Microm, Walldorf, Germany) at )15�Cand thaw-mounted onto SuperFrost Plus slides (VWR International,
Leicestershire, UK). For histological staining, sections were stained
with toluidine blue and imaged using an Aperio Slide Scanning
system at the UCD Conway Institute (Dublin, Ireland), which was
funded under a Health Research Board of Ireland Equipment Grant.
For immunohistochemistry, sections were fixed in 70% ethanol
for 25 min, permeablised using a solution of 0.02% Triton X-100 in
PBS for 30 min, washed with PBS and incubated overnight (18 h)
with SNAP-25 primary antibody (S9684; Sigma, Dublin, Ireland).
Following two 10 min washes in PBS, sections were incubated for
3 h with an anti-rabbit IgG (H + L) secondary antibody labelled
with the fluorescent marker Cy3 (Jackson ImmunoResearch, West
Grove, PA, USA). The secondary antibody was then washed off
with PBS, the sections were mounted with Citifluor� (Leicester,
UK) and the slides were stored in darkness at 4�C. Confocal images
were taken using the 10·/0.8 W water-dipping lens (Zeiss Achro-
plan; Zeiss, Thornwood, NY, USA).
In situ hybridizationA 275 bp region of SNAP-25 cDNA (563–822) was amplified using
PCR, incorporating a SP6- and T7-RNA polymerase promoter into
the forward and reverse strand respectively. Sense and antisense
digoxigenin-UTP labelled RNA probes were then generated by
in vitro transcription using SP6- and T7-RNA polymerase respec-
tively [DIG RNA labelling kit (SP6/T7); Roche, West Sussex, UK].
Coronal sections (30 lm) of the dorsal hippocampus of both
SNAP-25-AAV and GFP-AAV were cut on a cryostat (HM 500
Microm) at )15�C and thaw-mounted onto SuperFrost Plus slides
(VWR International), one each per slide. Sections were dried at
25�C and fixed in 4% p-formaldehyde at 4�C. Following washes andpre-hybridization steps, hybridization was performed with 1 lLprobes in 100 lL hybridization buffer (Sigma, Ireland) in a
humidity chamber saturated with a 50% 2x saline sodium citrate
buffer (SSC)–50% formamide solution at 60�C for 16 h. The
sections were washed twice in 2x SSC, 1x SSC and 0.1x SSC all at
60�C. Slides then were soaked twice in buffer I (1 M Tris–HCl and
1.5 M NaCl, pH 7.5) at 25�C. Buffer I, containing 1% goat serum
and 0.1% Triton X-100, was then added to tissue sections, followed
by an incubation with anti-digoxigenin–alkaline phosphatase con-
jugate (1 : 1000, diluted with 1% normal goat serum and 0.1%
Triton X-100 in buffer I) at 25�C for 2 h. Slides were rinsed in
buffer I twice and once with buffer II (100 mM Tris–HCl, 100 mM
NaCl and 50 mM MgCl2, pH 9.5). The colour reaction was
developed in a nitro-blue tetrazolium chloride/5-bromo-4-chloro-
3¢-indolyphosphate p-toluidine salt solution (Roche) until a dark-
blue reaction product was produced in the antisense but not sense
slides. The slides were rinsed in buffer III (10 mM Tris–HCl and
1 mM EDTA, pH 8.1) to terminate the reaction, washed in distilled
water, sections coverslipped and scanned using an Aperio Slide
Scanning system.
Fluorescent-activated cell sorting and quantitative RT-PCRFollowing cervical dislocation, the dorsal hippocampus was
dissected and dissociated in trypsin (1 mg/mL in Hank’s buffered
salt solution) for 5 min at 37�C. Dissociated cells were resuspended
in Hank’s buffered salt solution and GFP-expressing cells were
identified and sorted by fluorescent-activated cell sorting (FACS;
100 000–200 000 cells per hippocampus) using a Beckman-Coulter
Altra FACS (Beckman-Coulter Inc., Fullerton, CA, USA).
Gene-specific primers for RT-PCR were designed in exonic
sequence and so that the product also spanned an intron, using the
PRIMER3 algorithm (http://www.frodo.wi.mit.edu/cgi-bin/primer3/
primer3_www.cgi). RNA was extracted using the RNeasy Mini
Kit (Qiagen, Valencia, CA, USA) and Quantitative PCR was
performed on an ABI 7300 Real-Time PCR System (Applied
Biosystems) using the Quantitech PCR kit according to manufac-
turer’s instructions (Qiagen). Serial dilutions of RNA were used to
generate standard curves of crossing cycle number versus the
logarithm of concentration for each gene of interest. A linear
regression line calculated from the standard curves allowed relative
transcript levels in each RNA sample to be determined. Values were
normalized to the relative amounts of 18s RNA present in the same
RNA preparations and a melting curve was also carried out after
each amplification step, which provides confirmation that specific
PCR products were formed in the reaction. Expression of SNAP-25
in injected hippocampus was expressed as an average increase with
control values used as the baseline.
Sodium dodecyl sulphate–polyacrylamide gel electrophoresis andimmunoblottingGreen fluorescent protein-positive cells were collected by FACS (as
above) and samples were pooled and protein lysates prepared.
Normalized proteins samples were separated on 12% polyacrylamide
minigels and electrophoretically transferred to nitrocellulose mem-
branes (Bio-Rad, Hercules, CA, USA). The nitrocellulose was
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2010) 112, 991–1004� 2009 The Authors
994 | A. G. McKee et al.

blocked in 5% non-fat milk in 10 mmol/L Tris–HCl, 150 nmol/L
NaCl and 0.05% (vol/vol) Tween 20 for 1 h at 20�C. Monoclonal
antibodies to SNAP-25 (Sigma, St Louis, MO, USA) and b-actin(Sigma, USA) were diluted in 5% milk and incubated overnight at
4�C. Themembranes were then incubated with appropriate secondary
horseradish peroxidase-linked antibodies and visualized by Super-
Signal Chemiluminescent Substrate (Pierce, Rockford, IL, USA) and
exposed to X-ray film (Kodak, NewHaven, CT, USA) for appropriate
times. Films were digitized and quantitative densitometry was
performed using NIH Image software (Version 1.61; Bethesda,
MD, USA) to determine amount of target protein (SNAP-25 or
b-actin) electrophoresed per sample.
Paired-pulse facilitationAnimals were blinded from the electrophysiological experimenters
until statistical analysis was carried out. Animals were deeply
sedated with a 1 mL intraperitoneal injection of pentobarbital
(Euthanyl; Abbott Laboratories Ireland Ltd., Ballytivnan, Sligo,
Ireland) and decapitated by guillotine. The brain was rapidly
dissected and placed into ice-cold artificial CSF (aCSF) bubbled
with 95% O2 and 5% CO2 (aCSF composition: 220 mM sucrose,
28 mM NaHCO3, 1.25 mM NaH2PO4, 2.5 mM KCl, 7 mM
MgSO4Æ7H2O, 0.5 mM CaCl2 and 7 mM D-glucose). Dorsal
hippocampal slices (400 lm) corresponding to the injection sites
were cut coronally from both hemispheres using a vibroslice
(Campden Instruments, Loughborough, Leicester, UK). Slices were
transferred to a submerged incubation chamber containing bubbled
aCSF warmed to 30 ± 0.5�C (aCSF composition: 130 mM NaCl,
24 mM NaHCO3, 1.25 mM NaH2PO4, 3.5 mM KCl, 2 mM
MgSO4Æ7H2O, 1.5 mM CaCl2 and 10 mM D-glucose) and allowed
to recover for 60 min. Thereafter, slices were kept at room
temperature (22–24�C).Following the recovery period, slices were transferred to a
recording chamber perfused with aCSF at a flow rate of 1–2 mL/min
and heated to 30 ± 0.5�C (aCSF composition: 130 mM NaCl,
24 mM NaHCO3, 1.25 mM NaH2PO4, 3.5 mM KCl, 1 mM
MgSO4Æ7H2O, 1.5 mM CaCl2 and 10 mM D-glucose). Extracellular
field EPSPs were elicited by stimulation of the Schaffer collateral
pathway of the CA1 using monopolar glass electrodes (2–4 MWresistance) at a frequency of 0.05 Hz. PPF at a stimulus interval of
50 ms was recorded. Baseline EPSPs were allowed to stabilize
before an input/output curve was generated. Responses were
recorded at stimulus strengths corresponding to 50% of the
maximum output. PPF was recorded in the CA1 for 15 min and a
digital average of 45 recordings was calculated for each hippocam-
pal slice. PPF was determined by calculating the ratio between the
amplitudes of the second and first post-synaptic potential (second/
first post-synaptic potential).
In vivo microdialysisOn postnatal day 80, rats were anaesthetized using 4% isoflurane
inhalation and maintained on 2% isoflurane in a Kopf stereotaxic
frame (David Kopf Instruments, Tujunga, CA, USA). Using similar
surgical techniques to those above for viral delivery, a microdialysis
probe of concentric design (CMA/12; Carnegie Medicin, Solna,
Sweden) with an outer diameter of 0.5 mm and a 1 mm length of
dialysing membrane was positioned in the dorsal hippocampus (AP
)3.8, ML )1.5, V )3.4; mm from bone). During surgery, the body
temperature was continuously maintained at 37�C by means of a
thermostatically regulated heating pad (CMA 150; Carnegie
Medicin). The flow rate (2 lL/min) of the perfusion medium
(sterile Ringer solution; Baxter, Thetford, Norfolk, UK – formula
per 1000 mL: sodium chloride 8.6 g, potassium chloride 300 mg
and calcium chloride 300 mg, pH �6) through the microdialysis
probes was maintained constant by a microperfusion pump (CMA
100; Carnegie Medicin) during implantation and for the duration of
the microdialysis experiments. The probe was fixed to the skull with
stainless steel screws and metacrylic cement (Svedia, Enkoping,
Sweden). At the end of the experiments, the animals were killed by
pentobarbital (Euthatal; Rhone Merieux, Harlow, UK) overdose and
the placement of the probe was verified by microscopic examination
using a Leitz Cyrostat (Leica, Ashbourne, Ireland).
The animals were given 24 h to recover post-surgery before
commencement of the microdialysis experiments. Following sur-
gery, the animals were housed individually in cages for the duration
of the experiment. For the initial 24 h after surgery, animals are
given a high calorie (5% sucrose) solution to prevent dehydration
and loss of body weight. On the day of the microdialysis
experiment, the rat was placed in a large hemispherical bowl
(diameter: 135 cm) and the microdialysis probe was connected to a
microinfusion pump (CMA 100, prior to sampling to allow for
stabilization of glutamate and GABA levels following a 24 h
recovery after probe implantation (Frantz et al. 2002). Dialysatesamples were then collected and split into two 20 lL aliquots and
analysed separately for glutamate and GABA. The results reported
are means of at least five animals in each treatment group (n = 5–8).
Neurotransmitter analysisGlutamate was determined by pre-column derivatization of a 10 lLdialysate sample with o-phtaldialdehyde/mercaptoethanol reagent
and separation by reversed-phase HPLC on a Biophase ODS 5 lMparticle column (Knauer, Berlin, Germany). The mobile phase
contained 0.1 M sodium acetate, 6.25% methanol and 1.5%
tetrahydrofurane, pH 6.95, and was perfused at a flow rate of
1 mL/min. A linear gradient system was used to clean the column
after elution of glutamate. This involved switching to 100%methanol
for 2 min before switching back to the original acetate buffer. The
excitation wavelength in the fluorescence detector (Hitachi F1000,
Tokyo, Japan) was set at 370 nm and the emission cut off filter was set
at 450 nm. The limit of detection was 0.5 pmol/sample for glutamate
(Morari et al. 1994). The GABA assay employed was based on pre-
column derivatization of a 10 lL sample with o-phtaldialdehyde/t-butylthiol reagent and separation by reversed-phase HPLC on a
Nucleosil 3 C18 column (Machery-Nagel, Duren, Germany) perfused
under isocratic conditions at the flow rate of 0.8 mL/min. The mobile
phase was 0.15 M sodium acetate, 1 mM EDTA and 50% aceto-
nitrile, pH 5.4. The BAS LC4B electrochemical detector (Bio-
analytical Systems, West Lafayette, IN, USA) was set at +0.75 V.
The limit of detection was 50 fmol/sample (Morari et al. 1994).
Open-field assessmentTo ensure that all animals were of good health prior to behavioural
analyses, all animals were weighed and their general behaviour
assessed in an open-field apparatus (62 · 62 · 15 cm) over a 5 min
period. Video tracking system (ETHOVISION 3.1; Noldus, Wagenin-
gen, the Netherlands) was used for automated recording and
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2010) 112, 991–1004
Increased SNAP-25 impairs memory-associated synaptic plasticity | 995

analyses of both distance moved and mean velocity. Other
behaviours assessed included rearing, grooming, piloerection,
defecation and posture. All observations were carried out in the
quiet room under low-level red illumination between 08:00 and
12:00 h to minimize circadian influence.
Water maze paradigmThis spatial learning task has been described in detail previously
(Murphy et al. 1996). Briefly, the water maze apparatus consisted of
a large circular pool, 1.5 m in diameter and 80 cm high. The pool
was filled with warm water (26 ± 1�C) with a platform (11 cm in
diameter) submerged 1.5 cm below the level of the water. For all
trials the platform remained in the same location, 30 cm from the
wall in the northeast quadrant. To eliminate any intra-maze cues,
both the pool floor and the platform were constructed of black poly-
vinyl plastic. The experimental room, however, contained several
extra-maze visual cues. Spatial training consisted of four training
session, spaced 24 h apart, with each session consisting of five trials
(intertrial interval, 5 min). Each trial started with the rat being
lowered into the pool facing the wall of the maze at one of three
locations. The time taken by the animal to find and climb onto the
hidden platform within the criterion of 90 s was defined as the
escape latency time. If an animal failed to find the platform within
this time it was placed on the platform for 10 s. One day following
the final training session a probe trial was conducted to examine the
animals recall for the platform location. Here, the platform was
removed and the animals swim trace monitored by a computerized
tracking system, ETHOVISION 3.1, over a 60 s period. The training
data were analysed by plotting the mean escape latency ± SEM for
both the control and treated groups. The two groups were then
compared across all sessions and within each session by Two-way
repeat measures ANOVA. Along with the p value, the F statistic is
reported with the treatment degrees of freedom (1) and the residual
degrees of freedom quoted in the square brackets. For the recall data,
the data were expressed as the mean percentage time spent in each
of the four quadrants.
Contextual fear conditioningContextual fear conditioning was carried out as described previously
(Sandi et al. 2003). Briefly, training and testing took place in a
rodent observation cage (30 · 37 · 25 cm), the side walls of which
were constructed of stainless steel, and the back walls and doors
constructed of clear Plexiglass. The floor consisted of 20 steel rods
through which a scrambled shock from a LETICA I.C shock
generator (Model LI100-26; Letica Instruments, Barcelona, Spain)
could be delivered. Each observation cage was cleaned with a 1%
acetic acid solution before and after each session. The chambers
were illuminated with a 20 W bulb. On the training day, each rat
was transported from the colony room to the laboratory (situated in
an adjacent room) and placed into the observation cage. After 3 min,
the rats received three 1-s shocks of 0.7 mA intensity. The
intershock interval was 60 s and the rats were removed from the
conditioning chambers 30 s after the final shock and returned to
their cages. Thus, a conditioning session lasted �330 s. In
experiments to determine the effects of SNAP-25 over-expression
on memory consolidation, recall of contextual fear was tested 3 and
5 days after initial conditioning. At recall testing, rats were placed
back into the same chamber as used in conditioning, in the absence
of shock, for an 8-min context test. The time spent by each rat
freezing was measured. Freezing is defined as behavioural immo-
bility except for movement needed for respiration. These data were
used to calculate percent time freezing and the mean and SEM was
calculated for each group. Difference between the groups was
assessed by Student’s t-test.
Passive avoidance conditioningThe one-trial, step-through, light–dark version of the passive
avoidance paradigm was employed as we have described previously
(Fox et al. 1995). The smaller illuminated compartment was
separated from the larger dark compartment by a shutter that
contained a small entrance. The floor of the training apparatus
consisted of a grid of stainless steel bars that would deliver a
scrambled shock (0.75 mA every 0.5 ms) for 5 s when the animal
entered the dark chamber. Recall was tested 24 h following training
by placing the animal back in the lighted chamber and noting the
time to enter the dark chamber. A criterion time of 600 s was
employed. These data were analysed by Mann–Whitney U-test withp < 0.05 considered significant.
Pre-pulse inhibition of startleThe pre-pulse inhibition (PPI) protocol employed was based on a
procedure previously described by Geyer et al. (2001). Each rat was
restrained in an appropriately sized cylindrical holder that was
placed on a movement-sensitive platform and maintained in a
soundproof chamber. The rat was allowed to habituate to a white
noise background of 70 dB for 5 min before receiving five 20 ms
startle trials of 120 dB separated by randomized intervals of 10–
20 s. Immediately thereafter, each rat received five separate
presentations with one of the pre-pulse stimuli of 72, 76, 80 or
84 dB followed 100 ms later by the 120 dB acoustic startle
stimulus. Each trial was separated by a time interval of 10–20 s.
The four pre-pulse stimuli were delivered in a randomized manner
and included periods in which there was no pre-pulse or startle
stimulus. The session terminated with five further startle trials.
Randomization of the sound signals delivered and quantification of
the startle movements were recorded from the movement-sensitive
platform. Signals were integrated by the software supplied by the
manufacturers of equipment hardware (MED-Associates Inc., St
Albans, VT, USA) with data analysed by two-way ANOVA.
Results
Infection with rAAV/SNAP-25 results in enduringover-expression of SNAP-25 in vitro and in vivoIncubation of HEK293 cells with the rAAV/control andrAAV/SNAP-25 viral vectors resulted in robust expression ofeither eGFP or hrGFP (Fig. 1b and c). Next, male lateadolescent rats (postnatal 49) were injected bilaterally intothe dorsal hippocampus with rAAV/SNAP-25 or rAAV/control. Animals were housed singly until behavioural,electrophysiological or neurochemical analyses 5–7 weekspost-injection to allow for suitable recovery from thestereotaxic injection procedure and to ensure sufficient stableviral transgene expression. Throughout the study neither viral
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2010) 112, 991–1004� 2009 The Authors
996 | A. G. McKee et al.

vector aversely affected the health of the animals as indicatedby normal weight gain (data not shown). Followingbehavioural, electrophysiological and neurochemical analy-ses, at 5–7 weeks post-injection, brains from these animalswere assessed for effectiveness of viral infection and trans-gene delivery by fluorescent microscopy, immunohistochem-istry, flow cytometry, quantitative real-time PCR or westernblotting. The rAAV vectors proved to be effective in vivodelivery vehicles as judged by the significant number of cellsin the hippocampus that expressed GFP (n = 3; Fig. 1e). Ininfected cells, GFP was expressed in the cell body and
throughout the extent of the dendritic/axonal processes andwas clearly evident in dendritic spines (Fig. 1f and g).
Dorsal hippocampus was dissected and the cells dissoci-ated and sorted using FACS. When compared with anuninjected control hippocampus, both rAAV/control (eGFP)-and rAAV/SNAP-25 (hrGFP)-injected samples clearly hadgreater total fluorescence (Fig. 2a). GFP-expressing cellscomprised 8.1% (±1%, n = 6) of the total cell populationsorted by FACS. Total RNA was extracted from the GFP-positive cell-enriched fractions that resulted from the FACSanalysis and quantitative RT-PCR analysis showed signifi-
(a) (b) (c)
(d)
(f)
(g) (h) (l) (m)
(k)
(e) (i) (j)
Fig. 2 Efficient SNAP-25 expression in rAAV-infected neurons. (a)
Total fluorescence in rAAV/control-injected and non-injected control
samples. Fluorescent green cells (D) (8.1 ± 1%, n = 6) were sorted
and RNA was extracted from these cells for quantitative RT-PCR
analysis. Panel (b) indicates �100% increase in SNAP-25 mRNA
levels in GFP-expressing hippocampal cells (data are mean ± SD;
*indicates significant difference (p = 0.0001) by Student’s t-test; n = 6
animals per group). Panel (c) shows western blot data – top panel
shows SNAP-25 protein and bottom panel shows b-actin. (d–m)
Coronal sections through the rat dorsal hippocampus (�)3.3 mm with
respect to bregma). (d and i) SNAP-25 mRNA expression detected by
in situ hybridization. The level of expression is indicated by pseudo-
colour, with areas of high expression coloured red and low expression
dark blue. Areas of differential expression are indicated by arrow-
heads. (f and l) SNAP-25 protein expression was detected by immu-
nohistochemistry. (g and l) and (h and m) High power images of the
CA3-mossy fibre field and dentate gyrus apex subregions. Again,
areas with marked differential SNAP-25 expression are indicated by
arrowheads. (e and k) Toluidine blue-stained sections that illustrate
the overall morphology remained unaffected by injection of either the
control or SNAP-25 over-expressing vector. Scale bar in (d–f and
i–k) = 500 lm; Scale bar in (g, h, l and m) = 250 lm.
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2010) 112, 991–1004
Increased SNAP-25 impairs memory-associated synaptic plasticity | 997

cantly increased levels of SNAP-25 expression in rAAV/SNAP-25 injected rats compared with rAAV/control(Fig. 2b). Specifically, a 1 lL injection of high-titre rAAV/SNAP-25 into each side of the dorsal hippocampus led to anapproximate 100% increase in SNAP-25 expression withininfected cells (p = 0.0001, Student’s t-test, n = 6). Similarly,protein lysates were prepared from GFP-positive FACSsorted cells and western blot analyses confirmed that SNAP-25 was significantly up-regulated in AAV/SNAP-25 infectedcells when compared with AAV/Control cells or non-infectedcells (Fig. 2c).
In addition to the analysis outlined above, we assessedSNAP-25 expression at both mRNA and protein levels inthe dorsal hippocampus by in situ hybridization andimmunohistochemistry respectively. Comparative in situhybridization using a probe homologous to the SNAP-25mRNA sequence showed increased levels of SNAP-25mRNA in rAAV/SNAP-25 injected rats (Fig. 2d and i).Transgene expression was widespread throughout the dorsalhippocampus with highest levels of expression in the nucleiof the dentate granule neurons of the suprapyramidal bladeand the CA3 and CA1 pyramidal cells. Immunohistochem-ical analysis using an antibody targeted to SNAP-25revealed an accompanying increase in SNAP-25 proteinexpression in the dendritic fields of the primary hippocam-pal cell populations, in particular, the synaptic field of themossy fibre axons onto CA3 pyramidals (Fig. 2f–m).Finally, histological analysis revealed normal cellular mor-phology of hippocampal neurons indicating that there wereno adverse toxic effects caused by viral infection or over-expression of viral transgenes (Fig. 2e and j). The virallymediated over-expression of SNAP-25 was specific to thedorsal extent of the hippocampal structure as no differencesin SNAP-25 mRNA expression were evident by in situanalysis of the medial or ventral hippocampus at horizontallevels )5.1 mm and )7.6 mm with respect to bregmarespectively (data not shown).
rAAV/SNAP-25-mediated SNAP-25 over-expressionimpairs paired-pulse facilitation and increases basalglutamate levelsWe next assessed the effect of SNAP-25 over-expression onparameters pertinent to synapse function and plasticity in thedorsal hippocampus. In the first instance, using acute sliceelectrophysiology, we observed a significant decrease in themagnitude of PPF in the Schaffer collateral synaptic inputfield to the CA1 pyramidal neurons at 5 weeks post-injection. Figure 3a and b present representative traces fromrAAV/control and rAAV/SNAP-25 injected animals respec-tively. While the PPF obtained in the control animals was 2.1fold, that seen in the rAAV/SNAP-25 injected was only 1.75fold (Student’s t-test, p = 0.019; n = 6 animals per group)(Fig. 3c). Post-electrophysiology analysis of the acute slicesindicated robust infection rates for both viral constructs as
assessed by the number of GFP-positive cells that could beobserved (Fig. 3d).
A separate cohort of the rats injected with rAAV/SNAP-25 (n = 5) and rAAV/control (n = 8) underwent intra-hippocampal dialysis 5 weeks post-injection. A microdial-ysis probe was implanted unilaterally in the left dorsalhippocampus at precisely the same coordinates of theoriginal viral injection. Dialysate samples were collectedand analysed using reverse-phase HPLC to establish thelevels of the excitatory and inhibitory neurotransmitters,glutamate and GABA. These studies revealed that extra-cellular levels of excitatory glutamate were significantly
(a)
(b)
(d)
(e) (f)
(c)
Fig. 3 Consequence of SNAP-25 over-expression on paired-pulse
facilitation and neurotransmitter levels. (a and b) Representative tra-
ces from rAAV/eGFP control and rAAV/SNAP-25 injected animals
respectively. (c) Quantification of the paired-pulse facilitation obtained
in the rAAV/control and rAAV/SNAP-25 injected animals (data are
mean ± SEM; *indicates significant difference (p = 0.019) by Stu-
dent’s t-test; n = 6 animals per group). (d) Composite image overlay-
ing a phase contrast image and a fluorescence image showing eGFP
expression in an acute slice from a rAAV/control-injected animal.
Basal dialysate glutamate (e) and GABA (f) levels in the dorsal hip-
pocampus (data are mean ± SEM; *indicates significant difference
(p = 0.039) by Student’s t-test; n = 5–8 animals per group).
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2010) 112, 991–1004� 2009 The Authors
998 | A. G. McKee et al.
increased (Student’s t-test, p = 0.039, n = 5–8) in the dorsalhippocampus of rAAV/SNAP-25 injected rats (Fig. 3e)showing an approximate 3-fold increase in glutamate inrats over-expressing SNAP-25 (n = 5) (1.87 ± 0.97 vs.5.66 ± 1.39 lmol/L for control vs. SNAP-25 respectively),compared with control (n = 8). In contrast, no differencewas evident in the levels of GABA found in the dorsalhippocampus of the same animal cohort (9.87 ± 2.07 vs.9.74 ± 1.94 nM for control vs. SNAP-25 respectively;Fig. 3f). These findings suggest that chronic over-expres-sion of SNAP-25 resulted in a specific increase in glutamateneurotransmitter levels.
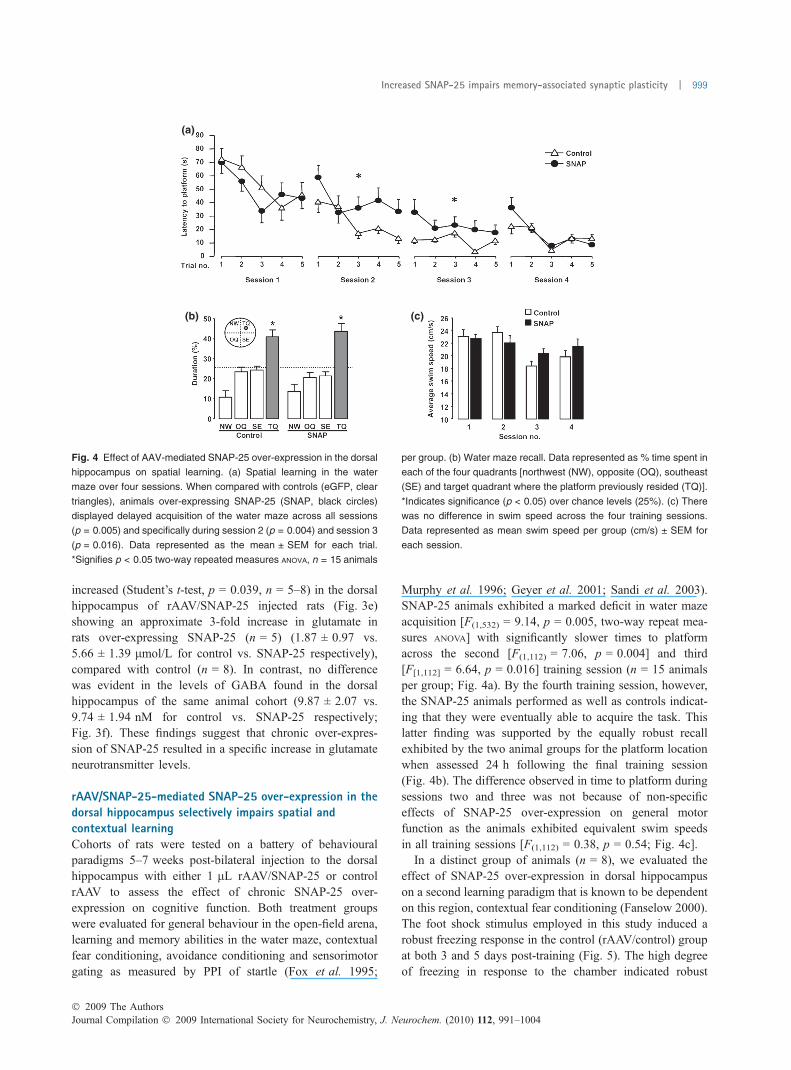
rAAV/SNAP-25-mediated SNAP-25 over-expression in thedorsal hippocampus selectively impairs spatial andcontextual learningCohorts of rats were tested on a battery of behaviouralparadigms 5–7 weeks post-bilateral injection to the dorsalhippocampus with either 1 lL rAAV/SNAP-25 or controlrAAV to assess the effect of chronic SNAP-25 over-expression on cognitive function. Both treatment groupswere evaluated for general behaviour in the open-field arena,learning and memory abilities in the water maze, contextualfear conditioning, avoidance conditioning and sensorimotorgating as measured by PPI of startle (Fox et al. 1995;
Murphy et al. 1996; Geyer et al. 2001; Sandi et al. 2003).SNAP-25 animals exhibited a marked deficit in water mazeacquisition [F(1,532) = 9.14, p = 0.005, two-way repeat mea-sures ANOVA] with significantly slower times to platformacross the second [F(1,112) = 7.06, p = 0.004] and third[F[1,112] = 6.64, p = 0.016] training session (n = 15 animalsper group; Fig. 4a). By the fourth training session, however,the SNAP-25 animals performed as well as controls indicat-ing that they were eventually able to acquire the task. Thislatter finding was supported by the equally robust recallexhibited by the two animal groups for the platform locationwhen assessed 24 h following the final training session(Fig. 4b). The difference observed in time to platform duringsessions two and three was not because of non-specificeffects of SNAP-25 over-expression on general motorfunction as the animals exhibited equivalent swim speedsin all training sessions [F(1,112) = 0.38, p = 0.54; Fig. 4c].
In a distinct group of animals (n = 8), we evaluated theeffect of SNAP-25 over-expression in dorsal hippocampuson a second learning paradigm that is known to be dependenton this region, contextual fear conditioning (Fanselow 2000).The foot shock stimulus employed in this study induced arobust freezing response in the control (rAAV/control) groupat both 3 and 5 days post-training (Fig. 5). The high degreeof freezing in response to the chamber indicated robust
(a)
(b) (c)
Fig. 4 Effect of AAV-mediated SNAP-25 over-expression in the dorsal
hippocampus on spatial learning. (a) Spatial learning in the water
maze over four sessions. When compared with controls (eGFP, clear
triangles), animals over-expressing SNAP-25 (SNAP, black circles)
displayed delayed acquisition of the water maze across all sessions
(p = 0.005) and specifically during session 2 (p = 0.004) and session 3
(p = 0.016). Data represented as the mean ± SEM for each trial.
*Signifies p < 0.05 two-way repeated measures ANOVA, n = 15 animals
per group. (b) Water maze recall. Data represented as % time spent in
each of the four quadrants [northwest (NW), opposite (OQ), southeast
(SE) and target quadrant where the platform previously resided (TQ)].
*Indicates significance (p < 0.05) over chance levels (25%). (c) There
was no difference in swim speed across the four training sessions.
Data represented as mean swim speed per group (cm/s) ± SEM for
each session.
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2010) 112, 991–1004
Increased SNAP-25 impairs memory-associated synaptic plasticity | 999

contextual memory. The percent time spent freezing uponre-exposure to the conditioning chamber was significantlyreduced in the rAAV/SNAP-25-injected group at the 3-dayrecall (Student’s t-test, p = 0.01), but was no longer differentfrom control in the second recall session on day 5 post-training.
Open-field analysis revealed no significant differences ingeneral motor function with both locomotion and rearingappearing normal in these animals (Fig. 6a). Moreover, inpassive avoidance learning, a paradigm thought to be moredependent on ventral hippocampus, an area not directlyaffected by our intervention, no difference was apparent inrecall for association between the darkened chamber and thefoot shock (median latency 600 s with interquartile range 0,0 s vs. median latency 600 s, with interquartile range 89, 0 s,rAAV/control vs. rAAV/SNAP-25 respectively; Fig. 6b).Finally, PPI of startle was evaluated as it is predominantlya prefrontal-dependent sensory processing task (Fig. 6c).Again, no difference was apparent in basal startle amplituderAAV/control (10.52 ± 14.57 au) compared with rAAV/SNAP-25 (12.88 ± 10.33 au). Although there was a ten-dency towards increased PPI in the SNAP-25 animals at thelower pre-pulse intensities, there was no overall significanteffect on sensory processing [F(1,96) = 1.44, p = 0.23].
Discussion
Long-term memory is formed by alterations in glutamate-dependent excitatory synaptic transmission, which is in turnregulated by SNAP-25, a key component of the SNAREcomplex essential for exocytosis of neurotransmitter-filledsynaptic vesicles (Lin and Scheller 2000; Lamprecht andLeDoux 2004). Moreover, SNAP-25 has been shown to playa significant role in memory-associated plasticity andcognitive function (Hou et al. 2004, 2006; Gosso et al.
2006). Specifically, the studies by Hou et al. (2004, 2006)demonstrated a memory-associated increase in SNAP-25expression and used antisense-induced knockdown ofSNAP-25 to report impairment in contextual fear memory.Together with our own findings here that over-expression ofSNAP-25 can similarly impair contextual fear memory these
(a)
(b)
(c)
Fig. 6 Effect of AAV-mediated SNAP-25 over-expression in the dorsal
hippocampus on locomotion, avoidance conditioning and sensorimotor
processing. (a) Exploratory behaviour (locomotion and rearing
behaviour) in the open field (data represented as the mean ± SEM,
n = 8). (b) Passive avoidance training and 24 h recall latencies. Data
represented as the median (thick line) latency to enter the dark com-
partment and the interquartile range illustrated by the box plot, n = 8.
(c) The percent pre-pulse inhibition at each pre-pulse intensity (data
represented as the mean ± SEM, n = 8).
Fig. 5 Effect of AAV-mediated SNAP-25 over-expression in the dorsal
hippocampus on contextual fear conditioning; rAAV/SNAP-25 injected
animals (filled columns) have a significantly reduced % time freezing
on day 3, when compared with control-injected animals (open col-
umns), although there is no significant difference by day 5. Data
represented as the mean ± SEM for each day, p = 0.01 statistical
analysis was performed by Student’s t-test, *indicates p < 0.05, n = 8.
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2010) 112, 991–1004� 2009 The Authors
1000 | A. G. McKee et al.

data suggest that SNAP-25 expression during memoryconsolidation must be tightly regulated, with any excessivedeviation from normal expression levels in either directionresulting in memory impairment. Indeed, both reduced andexcessive SNAP-25 activity has been implicated in variousdisease states that involve cognitive dysfunctions such asADHD, schizophrenia and Alzheimer’s disease (Greber et al.1999; Barr et al. 2000; Mukaetova-Ladinska et al. 2002;Mill et al. 2004; Jeans et al. 2007).
To date, no studies have characterized the consequences ofSNAP-25 overactivity for the correct functioning of the lateadolescent-adult nerve circuitry in the absence of concom-itant developmental disruption. However, one recent reportdescribes AAV-mediated over-expression of a botulinumtoxin A cleavage resistant SNAP-25 in rat spinal cord,attenuating neuromuscular paralysis (Raghunath et al. 2008).For the purpose of this study, we required the constructionand validation of viral-mediated gene delivery tools to allowthe local over-expression of SNAP-25 in vivo. In particular,high-titre rAAV design and production was necessary aslarge bolus injection volumes cannot be used in thehippocampus without risk of tissue damage and subsequentimpact on behavioural parameters. Here, rAAV viral titres of�5 · 1012 or higher were routinely achieved. The use ofthese high-titre preparations resulted in efficient infectionrates in vivo and the robust over-expression of SNAP-25 atboth a message and protein level in the absence of tissuedamage to the dorsal hippocampus.
The specificity of the role of SNAP-25 in neurotransmittervesicular release is still a topic of some debate (Verderioet al. 2004; Wang and Tang 2006; Matteoli et al. 2009). Inparticular, in the cortical regions such as hippocampus itremains unclear as to whether SNAP-25 functions exclu-sively in glutamatergic synapses or if it is also a componentof the GABA release machinery. Verderio et al. (2004)reported the lack of SNAP-25 immunoreactivity at inhibitorysynapses of mature rat hippocampal cultures and adult rathippocampus. This report has been backed up by similarfindings in adult mouse hippocampus (Frassoni et al. 2005),the rat cerebral cortex (Bragina et al. 2007) and human brain(Garbelli et al. 2008). However, contradictory findings byTafoya et al. (2006, 2008) report the presence of SNAP-25 inboth developing and adult mouse glutamatergic andGABAergic neurons. In this study, the impact of SNAP-25over-expression seemed to be restricted to glutamatergictransmission as our microdialysis studies revealed a selectiveincrease in extracellular glutamate levels in the dorsalhippocampus with no effect on the inhibitory neurotransmit-ter GABA. The possibility exists, of course, that thisdifferential effect resulted from an AAV infection preferencefor glutamatergic granule and pyramidal cells of the hippo-campus over GABAergic interneurons. However, no suchpreference has been reported previously. It is also importantto note that the observed increase in glutamate levels
achieved by SNAP-25 over-expression (from �2 lM incontrols to �6 lM in rAAV/SNAP-25-treated) did notappear to mediate excitotoxicity as indicated by the toluidineblue-stained sections illustrating no difference in overallmorphology between the control or SNAP-25 over-express-ing vector thus ensuring that effects noted on behaviour werenot because of neuronal cell death.
The GFP visualization and FACS studies suggested that�8% of cells in the hippocampus were infected by rAAV/SNAP-25 virus and these expressed 95% additional SNAP-25 compared with normal. The over-expression of SNAP-25in the dorsal hippocampus resulted in a surprisinglysubstantial increase in basal tone of glutamate in the cellularmilieu and a defect in normal synaptic function as indicatedby the decrement in the facilitation of synaptic signallingnormally achieved by temporally close pre-synaptic activa-tions. The later observation is consistent with recent findingsin the heterozygous SNAP-25+/) mouse which displays alarger PPF response than its wildtype counterpart (Pozziet al. 2008). The reduced PPF observed in this study wasaccompanied by a specific detrimental effect on spatiallearning, a paradigm long established to be dependent on thedorsal region of the hippocampus (Morris et al. 1982; Moseret al. 1995; Bannerman et al. 2004). Moreover, this effectwas particularly pronounced on the first trial of each session.Taken with the deficit in contextual fear conditioning, thesedata offer clear evidence that over-expression of SNAP-25 inthe adult dorsal hippocampus causes dysregulation of mem-ory consolidation machinery in this brain region. Moreover,the cognitive dysfunction mediated by SNAP-25 over-expression was restricted to the dorsal hippocampal regiontargeted by the viral intervention as no deficits were observedin avoidance conditioning or PPI of startle, functionsdependent on the ventral hippocampus and prefrontal cortexrespectively (Murphy et al. 1996; Bertoglio et al. 2006).
There are several possible explanations for the nature ofthe SNAP-25 over-expression-mediated deficit on spatialmemory. In the first instance, while only �8% of cells wereinfected, a significant number of which were granule orpyramidal neurons, the exaggerated influence on bothglutamate basal tone and PPF could possibly be explainedby a knock-on dysregulation of other glutamatergic neuronsin the dorsal hippocampal circuitry perhaps resulting in agood proportion of glutamatergic neurons falsely appearingoveractive within the nerve circuitry. In most computationalmodels, it is concluded that any given memory trace willinvolve activation of �5% of the cells in the dentate granuleor CA fields (Treves and Rolls 1994; Rolls and Kesner 2006;Aimone et al. 2009). Over-expression of SNAP-25 maycreate a distracter cell group which, during the memory tracelie down, reduces the signal-to-noise ratio and therefore thefidelity of the memory trace itself.
A second possibility relates to the need for SNAP-25levels to decrease transiently during the memory consolida-
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2010) 112, 991–1004
Increased SNAP-25 impairs memory-associated synaptic plasticity | 1001

tion process following a learning event (O’Sullivan et al.2007). Using microarray analysis, quantitative RT-PCR andwestern blotting techniques, SNAP-25 expression was shownto be markedly reduced at 6–12 h following passiveavoidance training. These changes were shown to belearning-specific as they were not observed in time-matchedcontrol animals (O’Sullivan et al. 2007). Given the role ofSNARE in glutamate release and the influence of SNAP-25on neuronal calcium responsiveness, a coordinated decreasein expression of SNAP-25 may function to modulateneuronal excitability during the 12 h post-training period(O’Sullivan et al. 2007). Thus, the chronic high expressionof SNAP-25 in a significant proportion of the hippocampalglutamatergic neurons would impede such a down-regulationand again mediate a high transmission signal during a phaseof memory consolidation when quiescence may be requiredto facilitate the synapse selection process initiated at this time(Murphy and Regan 1998; Regan 2004).
Evidence suggests that aberrant SNAP-25 expression andactivity is associated with neurological disease and cognitiveimpairments (Barr et al. 2000; Brophy et al. 2002; Mill et al.2002, 2004; Kustanovich et al. 2003; Gosso et al. 2006).Here, we have shown that over-expressing SNAP-25 withinthe hippocampus leads to excessive glutamate release and asubsequent impairment in memory consolidation, providingfurther evidence that tight regulatory controls are needed forefficient neurotransmitter release and neuronal function.
In summary, the data obtained in the study provide the firstevidence that dysregulation of SNAP-25 levels in localregions of the adult brain can result in deficits in memoryformation highlighting the need for tight regulation of SNAP-25 activity for normal cognitive function. Furthermore, resultsfrom the study demonstrate that intervention using high-titreAAV vectors delivered locally to brain regions such as thehippocampus can be used effectively to explore the role ofover-expression (or knockdown) of candidate genes incognitive processes such as learning and memory.
Acknowledgements
The authors would like to thank the staff of the UCD Biomedical
Facility for technical assistance with the animal studies. The Applied
Neurotherapeutics Research Group is a Strategic Research Cluster
funded jointly by Science Foundation Ireland (03/IN3/B403C and
07/IN.1/B1778) and Wyeth Discovery. Menelas N. Pangalos is a full
time employee of Wyeth Research.
Supporting information
Additional Supporting information may be found in the online
version of this article:
Fig. S1 Optimization of the AAV injection.
As a service to our authors and readers, this journal provides
supporting information supplied by the authors. Such materials are
peer-reviewed and may be re-organized for online delivery, but are
not copy-edited or typeset. Technical support issues arising from
supporting information (other than missing files) should be
addressed to the authors.
References
Aimone J. B., Wiles J. and Gage F. H. (2009) Computational influ-ence of adult neurogenesis on memory encoding. Neuron 61,187–202.
Bannerman D. M., Rawlins J. N., McHugh S. B., Deacon R. M., YeeB. K., Bast T., Zhang W. N., Pothuizen H. H. and Feldon J. (2004)Regional dissociations within the hippocampus-memory andanxiety. Neurosci. Biobehav. Rev. 28, 273–283.
Barr C. L., Feng Y., Wigg K., Bloom S., Roberts W., Malone M.,Schachar R., Tannock R. and Kennedy J. L. (2000) Identificationof DNA variants in the SNAP-25 gene and linkage study of thesepolymorphisms and attention-deficit hyperactivity disorder. Mol.Psychiatry 5, 405–409.
Bertoglio L. J., Joca S. R. and Guimaraes F. S. (2006) Further evidencethat anxiety and memory are regionally dissociated within thehippocampus. Behav. Brain Res. 175, 183–188.
Bragina L., Candiracci C., Barbaresi P., Giovedı S., Benfenati F. and ContiF. (2007) Heterogeneity of glutamatergic and GABAergic releasemachinery in cerebral cortex. Neuroscience 146, 1829–1840.
Brophy K., Hawi Z., Kirley A., Fitzgerald M. and Gill M. (2002)Synaptosomal-associated protein 25 (SNAP-25) and attentiondeficit hyperactivity disorder (ADHD): evidence of linkage andassociation in the Irish population. Mol. Psychiatry 7, 913–917.
Eyre M. D., Richter-Levin G., Avital A. and Stewart M. G. (2003) Mor-phological changes in hippocampal dentategyrus synapses followingspatial learning in rats are transient.Eur. J. Neurosci. 17, 1973–1980.
Fanselow M. S. (2000) Contextual fear, gestalt memories, and the hip-pocampus. Behav. Brain Res. 110, 73–81.
Fox G. B., O’Connell A. W., Murphy K. J. and Regan C. M. (1995)Memory consolidation induces a transient and time-dependent in-crease in the frequency of neural cell adhesion molecule polysi-alylated cells in the adult rat hippocampus. J. Neurochem. 65,2796–2799.
Frantz K., Harte M., Ungerstedt U. and O’Connor W. T. (2002) A dualprobe characterization of dialysate amino acid levels in the medialprefrontal cortex and ventral tegmental area of the awake freelymoving rat. J. Neurosci. Methods 119, 109–119.
Frassoni C., Inverardi F., Coco S., Ortino B., Grumelli C., Pozzi D.,Verderio C. and Matteoli M. (2005) Analysis of SNAP-25 immu-noreactivity in hippocampal inhibitory neurons during develop-ment in culture and in situ. Neuroscience 131, 813–823.
Garbelli R., Inverardi F., Medici V., Amadeo A., Verderio C., MatteoliM. and Frassoni C. (2008) Heterogeneous expression of SNAP-25in rat and human brain. J. Comp. Neurol. 506, 373–386.
Garey L. J., Ong W. Y., Patel T. S., Kanani M., Davis A., MortimerA. M., Barnes T. R. E. and Hirsch S. R. (1998) Reduced dendriticspine density on cerebral cortical pyramidal neurons in schizo-phrenia. J. Neurol. Neurosurg. Psychiatry 65, 446–453.
Geyer M. A., Krebs-Thomson K., Braff D. L. and Swerdlow N. R.(2001) Pharmacological studies of prepulse inhibition models ofsensorimotor gating deficits in schizophrenia: a decade in review.Psychopharmacology (Berl.) 156, 117–154.
Glantz L. A. and Lewis D. A. (2000) Decreased dendritic spine densityon prefrontal cortical pyramidal neurons in schizophrenia. Arch.Gen. Psychiatry 57, 65–73.
Gosso M. F., de Geus E. J., van Belzen M. J., Polderman T. J., HeutinkP., Boomsma D. I. and Posthuma D. (2006) The SNAP-25 gene isassociated with cognitive ability: evidence from a family-based
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2010) 112, 991–1004� 2009 The Authors
1002 | A. G. McKee et al.

study in two independent Dutch cohorts. Mol. Psychiatry 11, 878–886.
Greber S., Lubec G., Cairns N. and Fountoulakis M. (1999) Decreasedlevels of synaptosomal associated protein 25 in the brain of patientswith Down syndrome and Alzheimer’s disease. Electrophoresis 20,928–934.
Hess E. J., Collins K. A. and Wilson M. C. (1996) Mouse model ofhyperkinesis implicates SNAP-25 in behavioral regulation.J. Neurosci. 16, 3104–3111.
Hildinger M., Auricchio A., Gao G., Wang L., Chirmule N. and WilsonJ. M. (2001) Hybrid vectors based on adeno-associated virus ser-otypes 2 and 5 for muscle-directed gene transfer. J. Virol. 75,6199–6203.
Hou Q., Gao X., Zhang X. et al. (2004) SNAP-25 in hippocampal CA1region is involved in memory consolidation. Eur. J. Neurosci. 20,1593–1603.
Hou Q. L., Gao X., Lu Q., Zhang X. H., Tu Y. Y., Jin M. L., Zhao G. P.,Yu L., Jing N. H. and Li B. M. (2006) SNAP-25 in hippocampalCA3 region is required for long-term memory formation. Biochem.Biophys. Res. Commun. 347, 955–962.
Jeans A. F., Oliver P. L., Johnson R. et al. (2007) A dominant mutationin Snap25 causes impaired vesicle trafficking, sensorimotor gating,and ataxia in the blind-drunk mouse. Proc. Natl Acad. Sci. USA104, 2431–2436.
Kolluri N., Sun Z., Sampson A. R. and Lewis D. A. (2005) Lamina-specific reductions in dendritic spine density in the prefrontalcortex of subjects with schizophrenia. Am. J. Psychiatry 162,1200–1202.
Kustanovich V., Merriman B., McGough J., McCracken J. T., SmalleyS. L. and Nelson S. F. (2003) Biased paternal transmission ofSNAP-25 risk alleles in attention-deficit hyperactivity disorder.Mol. Psychiatry 8, 309–315.
Lamprecht R. and LeDoux J. (2004) Structural plasticity and memory.Nat. Rev. Neurosci. 5, 45–54.
Lewis C. M., Levinson D. F., Wise L. H. et al. (2003) Genome scanmeta-analysis of schizophrenia and bipolar disorder, part II:schizophrenia. Am. J. Hum. Genet. 73, 34–48.
Lin R. C. and Scheller R. H. (2000) Mechanisms of synaptic vesicleexocytosis. Annu. Rev. Cell Dev. Biol. 16, 19–49.
Matteoli M., Pozzi D., Grumelli C., Condliffe S. B., Frassoni C.,Harkany T. and Verderio C. (2009) The synaptic split of SNAP-25:different roles in glutamatergic and GABAergic neurons? Neuro-science 158, 223–230.
Mill J., Curran S., Kent L., Gould A., Huckett L., Richards S., Taylor E.and Asherson P. (2002) Association study of a SNAP-25 micro-satellite and attention deficit hyperactivity disorder. Am. J. Med.Genet. 114, 269–271.
Mill J., Richards S., Knight J., Curran S., Taylor E. and Asherson P.(2004) Haplotype analysis of SNAP-25 suggests a role in theaetiology of ADHD. Mol. Psychiatry 9, 801–810.
Morari M., O’Connor W. T., Ungerstedt U. and Fuxe K. (1994) Dopa-mine D1 and D2 receptor antagonism differentially modulatesstimulation of striatal neurotransmitter levels by N-methyl-D-aspartic acid. Eur. J. Pharmacol. 256, 23–30.
Morris R. G., Garrud P., Rawlins J. N. and O’Keefe J. (1982) Placenavigation impaired in rats with hippocampal lesions. Nature 297,681–683.
Moser M.-B., Moser E. I., Forrest E., Andersen P. and Morris R. G. M.(1995) Spatial learning with a minislab in the dorsal hippocampus.Proc. Natl Acad. Sci. USA 92, 9697–9701.
Mukaetova-Ladinska E. B., Hurt J., Honer W. G., Harrington C. R. andWischik C. M. (2002) Loss of synaptic but not cytoskeletalproteins in the cerebellum of chronic schizophrenics. Neurosci.Lett. 317, 161–165.
Murphy K. J. and Regan C. M. (1998) Contributions of cell adhesionmolecules to altered synaptic weightings during memory consoli-dation. Neurobiol. Learn. Mem. 70, 73–81.
Murphy K. J., O’ Connell A. W. and Regan C. M. (1996) Repetitive andtransient increases in hippocampal neural cell adhesion moleculepolysialylation state following multi-trial spatial training. J. Neu-rochem. 67, 1268–1274.
O’Malley A., O’Connell C. and Regan C. M. (1998) Ultrastructuralanalysis reveals avoidance conditioning to induce a transientincrease in hippocampal dentate spine density in the6 hour post-training period of consolidation. Neuroscience 87,607–613.
O’Malley A., O’Connell C., Murphy K. J. and Regan C. M. (2000)Transient spine density increases in the mid-molecular layer ofhippocampal dentate gyrus accompany consolidation of a spatiallearning task in the rodent. Neuroscience 99, 229–232.
O’Reilly M., Palfi A., Chadderton N. et al. (2007) RNA interference-mediated suppression and replacement of human rhodopsin in vivo.Am. J. Hum. Genet. 81, 127–135.
O’Sullivan N. C., McGettigan P. A., Sheridan G. K., Pickering M.,Conboy L., O’Connor J. J., Moynagh P. N., Higgins D. G., ReganC. M. and Murphy K. J. (2007) Temporal change in geneexpression in the rat dentate gyrus following passive avoidancelearning. J. Neurochem. 101, 1085–1098.
Osen-Sand A., Catsicas M., Staple J. K., Jones K. A., Ayala G., KnowlesJ., Grenningloh G. and Catsicas S. (1993) Inhibition of axonalgrowth by SNAP-25 antisense oligonucleotides in vitro and in vivo.Nature 364, 445–448.
Paxinos G. and Watson C.. (1998) The Rat Brain in Sterotaxic Co-Ordinates, Academic Press, New York.
Pozzi D., Condliffe S., Bozzi Y., Chikhladze M., Grumelli C., Proux-Gillardeaux V., Takahashi M., Franceschetti S., Verderio C. andMatteoli M. (2008) Activity-dependent phosphorylation of Ser187is required for SNAP-25-negative modulation of neuronal volt-age-gated calcium channels. Proc. Natl Acad. Sci. USA 105, 323–328.
Raghunath A., Perez-Branguli F., Smith L. and Dolly J. O. (2008)Adeno-associated virus transfer of a gene encoding SNAP-25resistant to botulinum toxin A attenuates neuromuscular paralysisassociated with botulism. J. Neurosci. 28, 3683–3688.
Regan C. M. (2004) Cell adhesion molecules, in Memories are Made ofThese: From Messengers to Molecules (Riedel G. and Blatt B.eds). Kluwer Academic/Plenum Publishers, Norwell, MA.
Roberts L. A., Morris B. J. and O’Shaughnessy C. T. (1998)Involvement of two isoforms of SNAP-25 in the expression oflong-term potentiation in the rat hippocampus. Neuroreport 9,33–36.
Rohr U. P., Wulf M. A., Stahn S., Steidl U., Haas R. and Kronenwett R.(2002) Fast and reliable titration of recombinant adeno-associatedvirus type-2 using quantitative real-time PCR. J. Virol. Methods106, 81–88.
Rolls E. T. and Kesner R. P. (2006) A computational theory of hippo-campal function, and empirical tests of the theory. Prog. Neurobiol.79, 1–48.
Sandi C., Merino J. J., Cordero M. I., Kruyt N. D., Murphy K. J. andRegan C. M. (2003) Modulation of hippocampal NCAM polysi-alylation and spatial memory consolidation by fear conditioning.Biol. Psychiatry 54, 599–607.
Sollner T., Bennett M. K., Whiteheart S. W., Scheller R. H. and RothmanJ. E. (1993) A protein assembly–disassembly pathway in vitro thatmay correspond to sequential steps of synaptic vesicle docking,activation, and fusion. Cell 75, 409–418.
Tafoya L. C., Mameli M., Miyashita T., Guzowski J. F., Valenzuela C. F.and Wilson M. C. (2006) Expression and function of SNAP-25 as a
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2010) 112, 991–1004
Increased SNAP-25 impairs memory-associated synaptic plasticity | 1003

universal SNARE component in GABAergic neurons. J. Neurosci.26, 7826–7838.
Tafoya L. C., Shuttleworth C. W., Yanagawa Y., Obata K. and WilsonM. C. (2008) The role of the t-SNARE SNAP-25 in actionpotential-dependent calcium signaling and expression inGABAergic and glutamatergic neurons. BMC Neurosci. 9, 105.
Thompson P. M., Egbufoama S. and Vawter M. P. (2003) SNAP-25reduction in the hippocampus of patients with schizophrenia. Prog.Neuropsychopharmacol. Biol. Psychiatry 27, 411–417.
Treves A. and Rolls E. T. (1994) Computational analysis of the role ofthe hippocampus in memory. Hippocampus 4, 374–391.
Verderio C., Pozzi D., Pravettoni E. et al. (2004) SNAP-25 modulationof calcium dynamics underlies differences in GABAergic andglutamatergic responsiveness to depolarization. Neuron 41, 599–610.
Wang Y. and Tang B. L. (2006) SNAREs in neurons-beyond synapticvesicle exocytosis. Mol. Membr. Biol. 23, 377–384.
Wechsler D. (ed). (1997). WAIS-III Wechsler Adult Intelligence Scale.Psychological Corporation, San Antonio, TX.
Xiao X., Li J. and Samulski R. J. (1998) Production of high-titer re-combinant adeno-associated virus vectors in the absence of helperadenovirus. J. Virol. 72, 2224–2232.
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2010) 112, 991–1004� 2009 The Authors
1004 | A. G. McKee et al.




















