
A synthetic amyloid lawn system for high-throughput analysis of amyloid toxicity and drug screening
7
A synthetic amyloid lawn system for high-throughput analysis of amyloid toxicity and drug screening Koyeli Girigoswami, Sook Hee Ku, Jungki Ryu, Chan Beum Park * Institute for the BioCentury and Department of Materials Science and Engineering, Korea Advanced Institute of Science and Technology, 373-1 Guseong-dong, Yuseong-gu, Daejeon 305-701, Republic of Korea article info Article history: Received 29 January 2008 Accepted 18 March 2008 Available online 5 April 2008 Keywords: Self-assembly Cell viability Surface modification Amyloid aggregation abstract Amyloid-b (Ab) is the major constituent of senile plaques in the brains of Alzheimer’s disease pa- tients. In order to develop an efficient in vitro system for studying the interaction of cells with Ab aggregates, we have prepared a synthetic amyloid lawn by immobilizing Ab peptides over a func- tionalized glass surface and subsequently incubating the template in a fresh Ab solution. On the top of different types of amyloid lawns (e.g. monomeric, oligomeric, and fibrillar), we cultivated PC12 cells, creating physical contacts between the cells and the lawns. Results indicated that cell viability was differentially affected when grown atop different Ab lawns while cells were well adhered onto the surface of these Ab lawns. The mode of cell death by Ab lawn was confirmed to be apoptotic rather than necrotic, showing that cells undergo suicide by just contact with Ab lawn. While con- ventional ‘solution-based’ methods for testing amyloid toxicity suffer from problems such as lot-to-lot variations, continued fibrillation, and heterogeneous population of aggregates, our ‘surface-based’ lawn system is suitable for high-throughput analysis of amyloid toxicity, which may enable high- throughput screening of potential drug candidates for treating amyloid diseases with the goal of reducing the cell death on the lawn. Ó 2008 Elsevier Ltd. All rights reserved. 1. Introduction Alzheimer’s disease (AD) is characterized by the deposition of insoluble amyloid aggregates as amyloid plaques in the neuropil as well as the accumulation of neuro-fibrillary tangles in the cell bodies of neurons [1]. The majority of amyloid aggregates are composed of the amyloid-b (Ab) peptides, a 39–43 amino acid residue peptide produced by cleavage from a larger amyloid pre- cursor protein, APP [2,3].Ab is present in unaffected individuals and has a normal physiological role; however, in AD patients the Ab forms ordered aggregates that deposit extracellularly as amyloid or senile plaques in the neuropil, and as vascular deposits [4,5]. Nu- merous studies indicated a central role for Ab in AD pathogenesis; however, none of these studies definitively specify the form or the site of action of Ab neurotoxicity. Strong circumstantial evidences supported the notion that prefibrillar aggregates of Ab might be important for neurotoxicity [6–9], but some studies also explicated that fibrillar Ab, rather than amorphous aggregates of Ab, were causing neuronal cell death in vitro [3,10–12]. The mode of cell death induced by Ab is also very controversial. Some groups con- firmed the necrotic mode of cell death induced by Ab [13,14], whereas several studies indicated that apoptosis was induced in the presence of this peptide [15,16]. Thus, it will be difficult to ex- plain the topography of neurodegeneration until both the form of Ab causing neurotoxicity and the mode of cell death induced by Ab is clarified. Several studies on the effect of Ab on neuronal function had been explored so far involving the addition of exogenous Ab pep- tides to neuronal preparations, with the concomitant uncertainties regarding peptide aggregation state and their access to sub-cellular compartments [17,18]. Many reports demonstrated the toxicity assay of different Ab aggregates prepared by initially dissolving fresh Ab peptides into a buffered solution [19–21], but the solution phase should become to contain Ab aggregates of different sizes/ conformations resulting in a heterogeneous mixture, and the so- lution-based method cannot avoid a continuous fibrillation of Ab during incubation. Furthermore, in vivo formation of amyloid ag- gregates had been known to be highly affected by the presence of a solid surface, and the mechanism of amyloid deposition over surface was observed to be significantly different and more physi- ologically relevant compared to that in solution phase [22,23]. Re- cently, we have successfully developed an ‘amyloid lawn’ system by * Corresponding author. Tel.: þ82 42 869 3340; fax: þ82 42 869 3310. E-mail address: [email protected] (C.B. Park). Biomaterials 29 (2008) 2813–2819 Contents lists available at ScienceDirect Biomaterials journal homepage: www.elsevier.com/locate/biomaterials 0142-9612/$ – see front matter Ó 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.biomaterials.2008.03.022 Biomaterials 29 (2008) 2813–2819
Transcript of A synthetic amyloid lawn system for high-throughput analysis of amyloid toxicity and drug screening

lable at ScienceDirect
Biomaterials 29 (2008) 2813–2819
lable at ScienceDirect
Biomaterials 29 (2008) 2813–2819
Contents lists avaiContents lists avai
Biomaterials
journal homepage: www.elsevier .com/locate/biomater ia ls
Biomaterials
journal homepage: www.elsevier .com/locate/biomater ia ls
A synthetic amyloid lawn system for high-throughput analysis of amyloidtoxicity and drug screening
Koyeli Girigoswami, Sook Hee Ku, Jungki Ryu, Chan Beum Park *
Institute for the BioCentury and Department of Materials Science and Engineering, Korea Advanced Institute of Science and Technology, 373-1 Guseong-dong,Yuseong-gu, Daejeon 305-701, Republic of Korea
a r t i c l e i n f o
Article history:Received 29 January 2008Accepted 18 March 2008Available online 5 April 2008
Keywords:Self-assemblyCell viabilitySurface modificationAmyloid aggregation
* Corresponding author. Tel.: þ82 42 869 3340; faxE-mail address: [email protected] (C.B. Park).
0142-9612/$ – see front matter � 2008 Elsevier Ltd.doi:10.1016/j.biomaterials.2008.03.022
a b s t r a c t
Amyloid-b (Ab) is the major constituent of senile plaques in the brains of Alzheimer’s disease pa-tients. In order to develop an efficient in vitro system for studying the interaction of cells with Ab
aggregates, we have prepared a synthetic amyloid lawn by immobilizing Ab peptides over a func-tionalized glass surface and subsequently incubating the template in a fresh Ab solution. On the topof different types of amyloid lawns (e.g. monomeric, oligomeric, and fibrillar), we cultivated PC12cells, creating physical contacts between the cells and the lawns. Results indicated that cell viabilitywas differentially affected when grown atop different Ab lawns while cells were well adhered ontothe surface of these Ab lawns. The mode of cell death by Ab lawn was confirmed to be apoptoticrather than necrotic, showing that cells undergo suicide by just contact with Ab lawn. While con-ventional ‘solution-based’ methods for testing amyloid toxicity suffer from problems such as lot-to-lotvariations, continued fibrillation, and heterogeneous population of aggregates, our ‘surface-based’lawn system is suitable for high-throughput analysis of amyloid toxicity, which may enable high-throughput screening of potential drug candidates for treating amyloid diseases with the goal ofreducing the cell death on the lawn.
� 2008 Elsevier Ltd. All rights reserved.
1. Introduction
Alzheimer’s disease (AD) is characterized by the deposition ofinsoluble amyloid aggregates as amyloid plaques in the neuropil aswell as the accumulation of neuro-fibrillary tangles in the cellbodies of neurons [1]. The majority of amyloid aggregates arecomposed of the amyloid-b (Ab) peptides, a 39–43 amino acidresidue peptide produced by cleavage from a larger amyloid pre-cursor protein, APP [2,3]. Ab is present in unaffected individualsand has a normal physiological role; however, in AD patients the Ab
forms ordered aggregates that deposit extracellularly as amyloid orsenile plaques in the neuropil, and as vascular deposits [4,5]. Nu-merous studies indicated a central role for Ab in AD pathogenesis;however, none of these studies definitively specify the form or thesite of action of Ab neurotoxicity. Strong circumstantial evidencessupported the notion that prefibrillar aggregates of Ab might beimportant for neurotoxicity [6–9], but some studies also explicatedthat fibrillar Ab, rather than amorphous aggregates of Ab, were
: þ82 42 869 3310.
All rights reserved.
causing neuronal cell death in vitro [3,10–12]. The mode of celldeath induced by Ab is also very controversial. Some groups con-firmed the necrotic mode of cell death induced by Ab [13,14],whereas several studies indicated that apoptosis was induced inthe presence of this peptide [15,16]. Thus, it will be difficult to ex-plain the topography of neurodegeneration until both the form ofAb causing neurotoxicity and the mode of cell death induced by Ab
is clarified.Several studies on the effect of Ab on neuronal function had
been explored so far involving the addition of exogenous Ab pep-tides to neuronal preparations, with the concomitant uncertaintiesregarding peptide aggregation state and their access to sub-cellularcompartments [17,18]. Many reports demonstrated the toxicityassay of different Ab aggregates prepared by initially dissolvingfresh Ab peptides into a buffered solution [19–21], but the solutionphase should become to contain Ab aggregates of different sizes/conformations resulting in a heterogeneous mixture, and the so-lution-based method cannot avoid a continuous fibrillation of Ab
during incubation. Furthermore, in vivo formation of amyloid ag-gregates had been known to be highly affected by the presence ofa solid surface, and the mechanism of amyloid deposition oversurface was observed to be significantly different and more physi-ologically relevant compared to that in solution phase [22,23]. Re-cently, we have successfully developed an ‘amyloid lawn’ system by
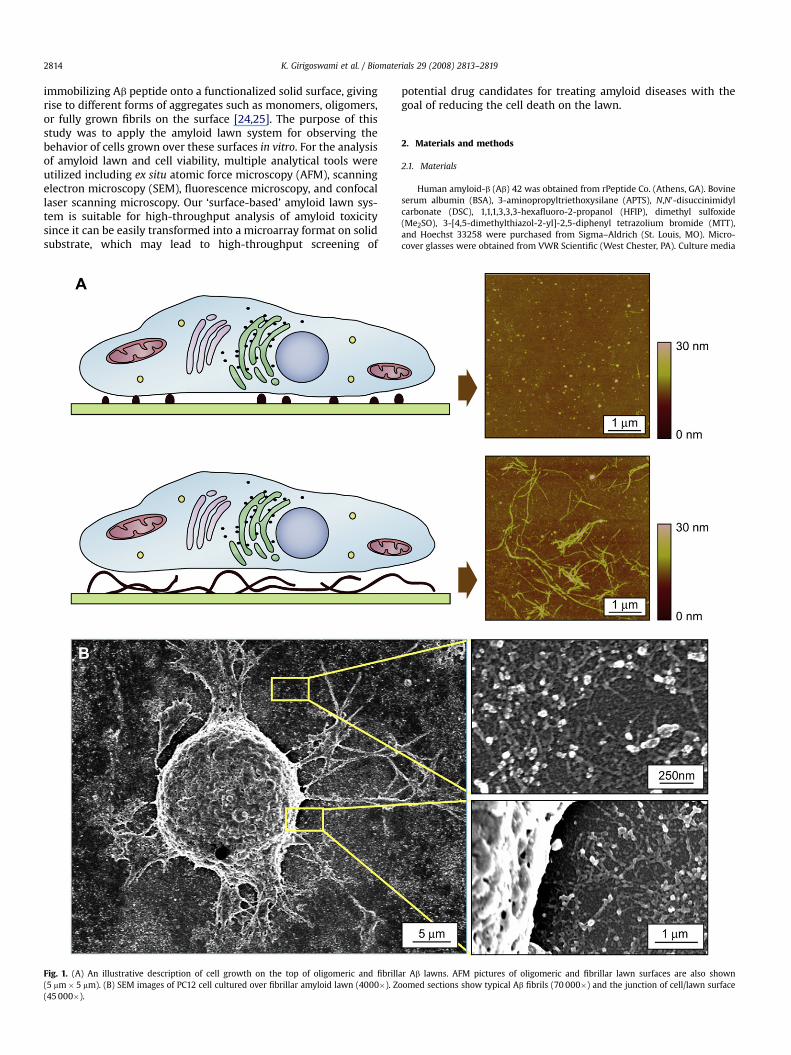
K. Girigoswami et al. / Biomaterials 29 (2008) 2813–28192814
immobilizing Ab peptide onto a functionalized solid surface, givingrise to different forms of aggregates such as monomers, oligomers,or fully grown fibrils on the surface [24,25]. The purpose of thisstudy was to apply the amyloid lawn system for observing thebehavior of cells grown over these surfaces in vitro. For the analysisof amyloid lawn and cell viability, multiple analytical tools wereutilized including ex situ atomic force microscopy (AFM), scanningelectron microscopy (SEM), fluorescence microscopy, and confocallaser scanning microscopy. Our ‘surface-based’ amyloid lawn sys-tem is suitable for high-throughput analysis of amyloid toxicitysince it can be easily transformed into a microarray format on solidsubstrate, which may lead to high-throughput screening of
Fig. 1. (A) An illustrative description of cell growth on the top of oligomeric and fibrill(5 mm� 5 mm). (B) SEM images of PC12 cell cultured over fibrillar amyloid lawn (4000�). Z(45 000�).
potential drug candidates for treating amyloid diseases with thegoal of reducing the cell death on the lawn.
2. Materials and methods
2.1. Materials
Human amyloid-b (Ab) 42 was obtained from rPeptide Co. (Athens, GA). Bovineserum albumin (BSA), 3-aminopropyltriethoxysilane (APTS), N,N0-disuccinimidylcarbonate (DSC), 1,1,1,3,3,3-hexafluoro-2-propanol (HFIP), dimethyl sulfoxide(Me2SO), 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl tetrazolium bromide (MTT),and Hoechst 33258 were purchased from Sigma–Aldrich (St. Louis, MO). Micro-cover glasses were obtained from VWR Scientific (West Chester, PA). Culture media
ar Ab lawns. AFM pictures of oligomeric and fibrillar lawn surfaces are also shownoomed sections show typical Ab fibrils (70 000�) and the junction of cell/lawn surface
K. Girigoswami et al. / Biomaterials 29 (2008) 2813–2819 2815
and supplements were obtained from Welgene (Korea). All other molecular biologygrade chemicals were procured from Sigma.
2.2. Preparation of fresh monomeric Ab peptide solution
Pre-existing Ab peptide aggregates were disintegrated into fresh monomers bydissolving the peptides in HFIP at 2.5 mg/ml and then sonicating the solution ina water-bath for 3 min. Ab peptide film was formed on sterile eppendorf tube wall byaliquoting the solution into the tubes and completely evaporating volatile HFIP invacuum desiccators. Then peptide films were stored at �20 �C. Immediately prior touse, Ab peptide film was dissolved in Me2SO and then diluted with phosphatebuffered saline (PBS, pH 7.4) to a desired concentration. The final concentration ofMe2SO in the peptide solution was kept at 5% (v/v).
2.3. Fabrication of synthetic amyloid lawn
Ab lawns were prepared by covalently immobilizing oligomeric or monomericAb seeds onto micro-cover glass as described earlier [24]. Briefly, cover glasses werecleaned with a piranha solution (70% H2SO4: 30% H2O2), rinsed extensively withdistilled water and dried. Hydroxyl-terminated glass surface was then reacted with3% APTS in 95% ethanol and further cured at 110 �C with 100% ethanol. For the ac-tivation of these glass surfaces with NHS, they were incubated in 20 mM DSC insodium bicarbonate buffer (50 mM, pH 8.5). The slides were then washed withdeionized water and dried with N2. A fresh Ab solution (30 mM) or 12 h pre-incubatedAb solution (30 mM) was then uniformly placed onto NHS-activated glass surface toyield monomeric and oligomeric lawns, respectively. Remaining unbound NHS-activated sites were then blocked with a 0.1% solution of BSA in 50 mM phosphatebuffer (PB) (pH 7.5). The glass slides were then extensively washed with a 50 mM PB(pH 7.5) and deionized water and were dried with N2. For fibrillar lawns, the olig-omeric surfaces were further incubated with fresh Ab (30 mM) for 24 h at 37 �C. Thetemplates were then dried with N2.
2.4. Cell culture
Undifferentiated PC12 cells were maintained in MEM with 10% heat-inactivatedFBS and 1% antibiotic solution (Invitrogen, Carlsbad, CA). Cells were subculturedevery 2 or 3 days and were maintained at 37 �C in a humidified incubator containing95% air and 5% CO2. Different Ab lawn surfaces were sterilized using 70% ethanol andplaced in a 24 well tissue culture plate after air drying. The surface morphologyremained unaltered even after sterilizing with ethanol as seen through AFM (datanot shown). Cells were added at a low density (4�104 cells/400 ml medium) andallowed to grow at 37 �C in 5% CO2 incubator, for the desired time before assay.
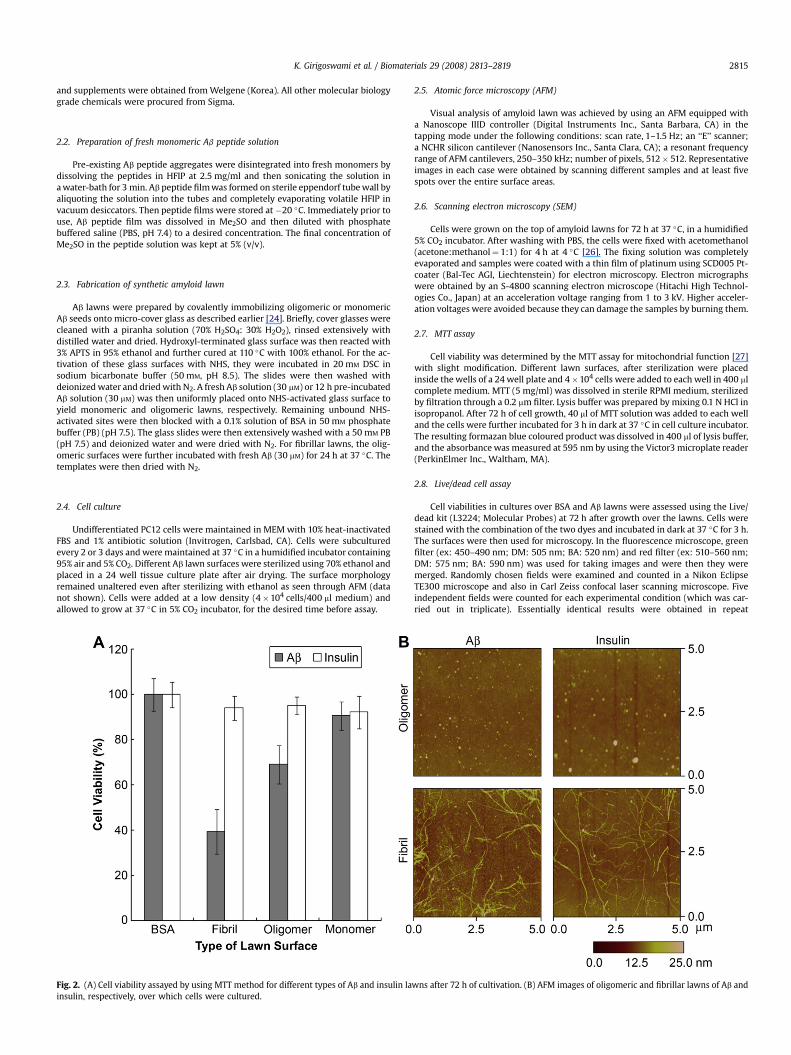
Fig. 2. (A) Cell viability assayed by using MTT method for different types of Ab and insulin lainsulin, respectively, over which cells were cultured.
2.5. Atomic force microscopy (AFM)
Visual analysis of amyloid lawn was achieved by using an AFM equipped witha Nanoscope IIID controller (Digital Instruments Inc., Santa Barbara, CA) in thetapping mode under the following conditions: scan rate, 1–1.5 Hz; an ‘‘E’’ scanner;a NCHR silicon cantilever (Nanosensors Inc., Santa Clara, CA); a resonant frequencyrange of AFM cantilevers, 250–350 kHz; number of pixels, 512� 512. Representativeimages in each case were obtained by scanning different samples and at least fivespots over the entire surface areas.
2.6. Scanning electron microscopy (SEM)
Cells were grown on the top of amyloid lawns for 72 h at 37 �C, in a humidified5% CO2 incubator. After washing with PBS, the cells were fixed with acetomethanol(acetone:methanol¼ 1:1) for 4 h at 4 �C [26]. The fixing solution was completelyevaporated and samples were coated with a thin film of platinum using SCD005 Pt-coater (Bal-Tec AGI, Liechtenstein) for electron microscopy. Electron micrographswere obtained by an S-4800 scanning electron microscope (Hitachi High Technol-ogies Co., Japan) at an acceleration voltage ranging from 1 to 3 kV. Higher acceler-ation voltages were avoided because they can damage the samples by burning them.
2.7. MTT assay
Cell viability was determined by the MTT assay for mitochondrial function [27]with slight modification. Different lawn surfaces, after sterilization were placedinside the wells of a 24 well plate and 4�104 cells were added to each well in 400 mlcomplete medium. MTT (5 mg/ml) was dissolved in sterile RPMI medium, sterilizedby filtration through a 0.2 mm filter. Lysis buffer was prepared by mixing 0.1 N HCl inisopropanol. After 72 h of cell growth, 40 ml of MTT solution was added to each welland the cells were further incubated for 3 h in dark at 37 �C in cell culture incubator.The resulting formazan blue coloured product was dissolved in 400 ml of lysis buffer,and the absorbance was measured at 595 nm by using the Victor3 microplate reader(PerkinElmer Inc., Waltham, MA).
2.8. Live/dead cell assay
Cell viabilities in cultures over BSA and Ab lawns were assessed using the Live/dead kit (L3224; Molecular Probes) at 72 h after growth over the lawns. Cells werestained with the combination of the two dyes and incubated in dark at 37 �C for 3 h.The surfaces were then used for microscopy. In the fluorescence microscope, greenfilter (ex: 450–490 nm; DM: 505 nm; BA: 520 nm) and red filter (ex: 510–560 nm;DM: 575 nm; BA: 590 nm) was used for taking images and were then they weremerged. Randomly chosen fields were examined and counted in a Nikon EclipseTE300 microscope and also in Carl Zeiss confocal laser scanning microscope. Fiveindependent fields were counted for each experimental condition (which was car-ried out in triplicate). Essentially identical results were obtained in repeat
wns after 72 h of cultivation. (B) AFM images of oligomeric and fibrillar lawns of Ab and

Fig. 3. Cells were cultivated over BSA and fibrillar Ab lawn and stained using calcein AM and ethidium homodimer dyes, which stain live and dead cells in green and red, respectively. Theimages shown are taken using (A) fluorescence microscope (200�) (B) confocal laser scanning microscope (400�). The individual image of cells stained with calcein AM and ethidiumhomodimer dyes and also the overlay of the two images is shown. (C) Bar graphs representing the percent dead cells after scoring live and dead cells under fluorescent microscope.
K. Girigoswami et al. / Biomaterials 29 (2008) 2813–28192816
experiments. Dead cells were also scored for both BSA and fibrillar Ab templates andthe percentage of dead cells were found as the ratio of number of dead cells to thetotal number of cells counted and also plotted in a bar graph.
2.9. Apoptosis/necrosis assay
Apoptosis or necrosis was analyzed using the Vybrant apoptosis assay kit con-taining YoPro-1/propidium iodide (PI) dyes (V13243; Molecular Probes). This assayis based on the use of the green fluorescent YoPro-1 dye that specifically stainsapoptotic cells that remain impermeant to PI (a dead cell stain). Live cells are notstained with YoPro-1. Moreover, PI is able to detect necrotic cells by binding to DNA.Cells were grown over the respective lawn surfaces for 72 h, the medium was re-moved and washed with PBS followed by staining with the dyes. Fluorescencemicroplate assay was done as described by [28,29] and the percent of apoptotic ornecrotic cells were calculated. The readings were taken using the optimum wave-lengths for YoPro-1 of 485 nm lex and 530 nm lem and for PI of 485 nm lex and642 nm lem.
2.10. Determination of apoptosis by fluorescence microscopy
Apoptosis was confirmed by morphological criteria determined by labeling thecells with the nuclear stain Hoechst 33258 and visualization by fluorescence mi-croscopy. After growing the cells over BSA and Ab lawn surfaces for 72 h, mediumwas removed and the surfaces were washed with PBS. Cells were then fixed withacetomethanol (acetone:methanol¼ 1:1) for 4 h at 4 �C [26] and stained withHoechst 33258 (5 mg/ml) and incubated in dark for 4 min at RT. Nuclei (blue orpurple) that were condensed or fragmented were scored as apoptotic. From eachlawn (in triplicate), five fields were taken randomly and more than 400 cells werescored. The percentage of apoptotic cells was plotted, taking the ratio of the numberof apoptotic cells to the total number of cells counted.
Fig. 4. Bar graph representing the fluorescent intensity corresponding to either apo-ptosis or necrosis in cells grown over BSA, monomeric, oligomeric, and fibrillar Ab
lawns, respectively.
2.11. Caspase-3 assay
Caspase-3 activity assay was performed according to the protocol given by themanufacturers (E-13183; Molecular Probes). Cells were plated in equal numbers(5�105 cells/well) in a 24 well plate over the top of BSA and fibrillar templates.After 48 h, cells were trypsinized, suspended in PBS, and centrifuged at 1500 rpmfor 5 min. The pellet was lysed using 50 ml of the supplied lysis buffer for 30 minon ice. The lysed cells and cellular debris were cleared by centrifugation at5000 rpm for 5 min in a microcentrifuge at RT. Supernatant (50 ml) was trans-ferred from each sample to individual microplate wells (96 well) and 50 ml of celllysis buffer was also taken as a no-enzyme control to determine the backgroundfluorescence of the substrate. Then, 50 ml of the prepared 2� substrate workingsolution was added to each sample and control and the plate was incubated indark at RT for 30 min. The fluorescence was measured in the Victor3 microplatereader by excitation at 355 nm and the recording of emission at 460 nm. Theassay was repeated thrice and the mean and SD were plotted for the relative foldincrease of caspase-3.
3. Results
The concept of cell culture over amyloid lawn is illustrativelydescribed in Fig. 1A. We cultured cells over Ab lawns, which con-sisted of three different forms of Ab aggregate states like – mono-meric, oligomeric, and fibrillar. The representative AFM images ofAb lawn surfaces in Fig. 1A show oligomeric or fibrillar aggregates,respectively. Cells were well adhered onto the surface of these Ab
lawns as observed under inverted optical microscope. The numberof adhered cells was similar to the number of cells adhered to a bareglass cover slip of equal size indicating similar plating efficiency ofAb lawns compared to glass (data not shown). Further, the SEMimage in Fig. 1B confirms that PC12 cells were adhered atop a typ-ical fibrillar Ab surface. As a control for the experiments, we utilizedbovine serum albumin (BSA)-immobilized surfaces, which also hada similar number of adherent cells like bare glass. The viability ofcells grown over these lawns was evaluated through MTT assay(Fig. 2A). The morphology of surfaces of the different Ab lawns wasalso visualized using ex situ AFM (Fig. 2B). Results indicated that theoligomeric and fibrillar Ab lawns were differentially cytotoxic, butthe monomeric lawn was virtually non-toxic. The cell viability onAb oligomeric lawns decreased to w69% cells, whereas only w39%
K. Girigoswami et al. / Biomaterials 29 (2008) 2813–2819 2817
of cells were viable on fibrillar lawns, which indicated that thecontact with fibrillar Ab lawns induced higher toxicity compared tooligomeric Ab lawns.
In order to compare the cell viability on amyloid lawns com-posed of other amyloidogenic peptide, we have prepared ‘insulin’amyloid lawns in a manner similar to Ab lawns. Insulin was pre-viously reported to form amyloid fibrils on a solid surface accordingto AFM imaging, circular dichroism spectra, Fourier transform in-frared study, Thioflavin-T fluorescence, and Congo red binding [30].In the present study, we examined the viability of cells grown overeach insulin amyloid lawn surface. As shown in Fig. 2A, cell viabilitydata indicated that contact with insulin amyloids did not
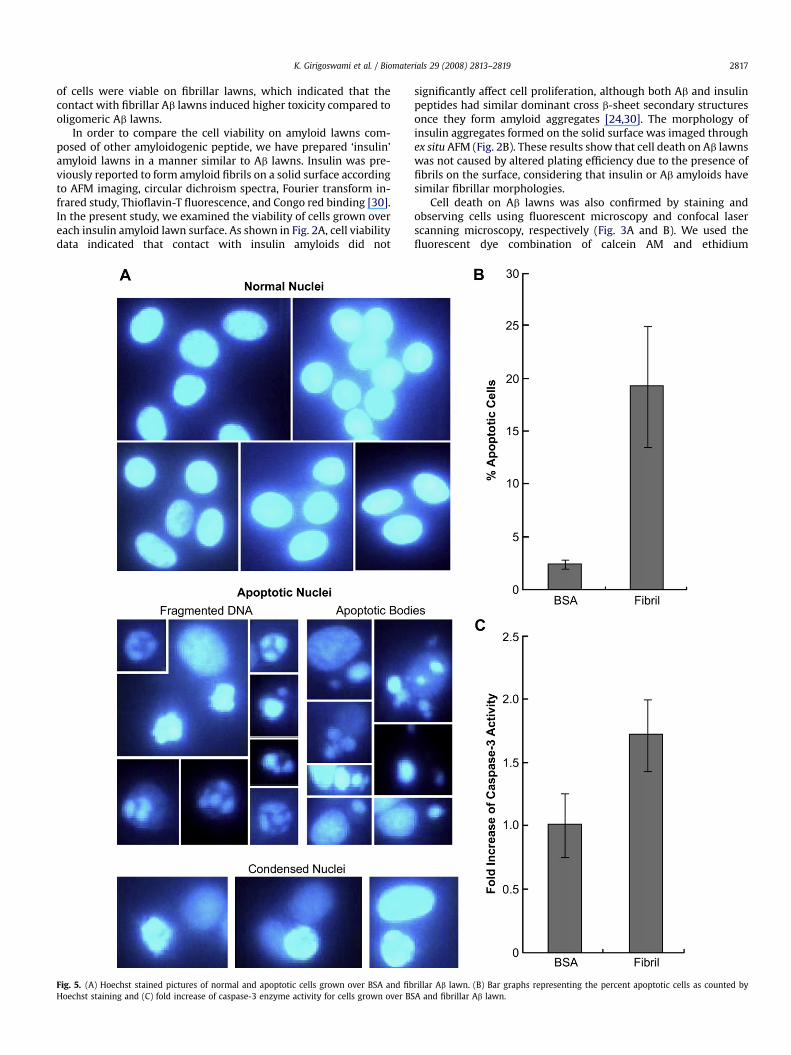
Fig. 5. (A) Hoechst stained pictures of normal and apoptotic cells grown over BSA and fibHoechst staining and (C) fold increase of caspase-3 enzyme activity for cells grown over BS
significantly affect cell proliferation, although both Ab and insulinpeptides had similar dominant cross b-sheet secondary structuresonce they form amyloid aggregates [24,30]. The morphology ofinsulin aggregates formed on the solid surface was imaged throughex situ AFM (Fig. 2B). These results show that cell death on Ab lawnswas not caused by altered plating efficiency due to the presence offibrils on the surface, considering that insulin or Ab amyloids havesimilar fibrillar morphologies.
Cell death on Ab lawns was also confirmed by staining andobserving cells using fluorescent microscopy and confocal laserscanning microscopy, respectively (Fig. 3A and B). We used thefluorescent dye combination of calcein AM and ethidium
rillar Ab lawn. (B) Bar graphs representing the percent apoptotic cells as counted byA and fibrillar Ab lawn.

K. Girigoswami et al. / Biomaterials 29 (2008) 2813–28192818
homodimer, which stains live and dead cells in green and red, re-spectively. With appropriate filters, the cells were visualized andthe percent of total dead cells were plotted as a ratio of totalnumber of dead cells to the total number of cells counted (Fig. 3C).As shown in the figures, much higher number of dead cells (red)was observed on Ab lawns, compared to BSA-immobilized surfacethat was used as a control set. This observation was consistent withthe results of MTT assay, confirming that the cell death occurreddue to contact with Ab lawns.
In order to further investigate the mode of cell death on Ab
lawns, we have carried out the apoptotic/necrotic cell death assay.The dyes used in the study stain the apoptotic cells to be green andnecrotic to be red, which can be detected using appropriate filtersin a fluorescence microplate reader. Our results demonstrated thatthere was a background necrosis for BSA surface as well as for eachtype of Ab lawns, but the extent of apoptosis was much higher inthe case of oligomeric or fibrillar Ab surfaces (Fig. 4). According tothe results, the fibrillar Ab lawns caused higher apoptosis (w1.91fold) than oligomeric lawns (w1.21 fold), compared to the BSAsurface. This results show that the mode of cell death was notnecrosis but apoptosis for both oligomeric and fibrillar Ab
assemblages.The morphology of nuclei in cells grown on the Ab lawn was
investigated by using the cell-permeable DNA-staining dye,Hoechst 33258. According to previous study [31], cells with ho-mogeneously stained nuclei were considered to be viable, whereasthe presence of chromatin condensation and/or fragmentation wasindicative of apoptosis. For this study, we have visualized the nucleiand classified the cells as either normal or apoptotic. The differentfeatures of apoptosis such as condensed nuclei, fragmented DNA,and apoptotic bodies in cells grown over amyloid lawn are shown inFig. 5A. Apoptotic cells were also scored for both BSA and fibrillarAb lawns and the ratio of number of apoptotic cells to the totalnumber of cells was plotted as percent apoptotic cells (Fig. 5B). Theresults revealed much higher number of apoptotic nuclei for cellsgrown over fibrillar Ab lawn. According to literatures, apoptoticpathways are not unified and involve either caspase dependent orindependent routes, and many caspase dependent pathways ofapoptosis converge to caspase-3 enzyme activation [32–34]. In thiswork, we measured the activity of caspase-3 in cells grown on thefibrillar Ab lawn after 48 h of cultivation. We observed a significantelevation of caspase-3 enzyme activity in the cells compared tocells grown over BSA-immobilized surface (Fig. 5C). The resultsconclude that cells died via caspase-3 dependent apoptosis throughtheir contact with fibrillar Ab aggregates on the lawn surface.
4. Discussion
Over the last two decades, accumulating evidences had impli-cated abnormal protein aggregation as a mechanistic feature ofmany amyloid diseases, including Alzheimer’s disease [5,35]. Al-though significant progress has been made in understanding themechanism of amyloid fibril formation [36], there still remainsuncertainty with regard to Ab-mediated neuronal cell death [37].Studies on the neurotoxic properties of Ab aggregates had proven itdifficult to interpret because of significant lot-to-lot variabilityfound in their preparations [38]. Furthermore, the use of variousculture systems and functional endpoints when assessing the cel-lular toxicity associated with Ab aggregate made the research quiteinconclusive and uncertain. Thus, it is critical that future re-searchers employ a relatively uniform and stationary form of Ab
aggregates for the study of Ab aggregates-associated toxicity. Inaddition, investigators reported that Ab fibril formation was ac-celerated significantly in the presence of solid surfaces, and themechanism of fibril formation on the surfaces was more physio-logically relevant because amyloid depositions are associated with
surfaces in vivo [22,23]. In order to overcome these problems, wehave introduced the amyloid lawn system for the first time, toobserve cell behaviors over different types of Ab lawns, specifically,monomeric, oligomeric, and fibrillar. We then employed cells inorder to examine viability, apoptosis, necrosis, and caspase-3 ac-tivities. The results of the present study suggested an induction ofapoptotic cell death via caspase-3 dependent pathway by merecontact with Ab lawn, rather than their cellular uptake.
Many different peptides or proteins had been reported to formamyloids having dominant cross b-sheet structure as a commonfeature, independently of their primary sequences [39,40]. In thisstudy, other than Ab amyloid, we have evaluated the toxicity ofinsulin amyloid as a comparison. The results highlighted that, al-though insulin forms similar amyloid fibrils on a solid surface likeAlzheimer’s Ab, its aggregates are not toxic to cells. This indicatedthat the contact of amino acid side chains of these two amyloidswith the cells might elicit differential death signals, and the toxicityof Ab amyloid may not be caused by the secondary structure (i.e.cross b-sheet) itself. Further studies are needed to investigate thespecific interactions of the side chains of Ab aggregates withconstituents of cell membrane.
5. Conclusion
We have established an in vitro system for evaluating the cel-lular toxicity of amyloid by introducing a new concept of amyloidlawn. According to our results, cell viability was differentially af-fected when grown on the top of the synthetic amyloid lawn,depending on the precursor peptide (i.e. Ab and insulin) and theaggregate size (i.e. monomeric, oligomeric, and fibrillar). The re-sults suggest that amyloid aggregates can cause cell death throughcontact, not by their uptake inside cell, since the amyloid assem-blages in the system were anchored to the solid surface and theircytotoxicity could be yielded only by contiguity. By addressingcurrent problems of ‘solution-based’ methods such as variations inpreparation and continued fibrillation during incubation, our ‘sur-face-based’ lawn system can be applied for the high-throughputanalysis of amyloid toxicity and drug screening by further adoptingmicroarray format on solid substrate.
Acknowledgement
This research was supported by grants from the KAIST Institutefor the BioCentury, the BioGreen 21 Program (20070301034038),and the Korea Research Foundation (KRF-2006-D00078). The au-thors thank the National NanoFab Center (NNFC) in Korea fortechnical supports in the characterization.
References
[1] Selkoe DJ. The molecular pathology of Alzheimer’s disease. Neuron 1991;6:487–98.
[2] Tanzi RE, Gusella JF, Watkins PC, Bruns GA, St. George-Hyslop P, VanKeuren ML, et al. Amyloid beta protein gene: cDNA, mRNA distribution, andgenetic linkage near the Alzheimer locus. Science 1987;235:880–4.
[3] Kang J, Lemaire H-G, Unterbeck A, Salbaum JM, Masters SL, Grzesckik KH, et al.The precursor of Alzheimer’s disease amyloid A4 protein resembles a cell-surface receptor. Nature 1987;325:733–6.
[4] Knowles RB, Wyart C, Buldyrev SV, Cruz L, Urbanc B, Hasselmo ME, et al.Plaque-induced neurite abnormalities: implications for disruption of neuralnetworks in Alzheimer’s disease. Proc Natl Acad Sci USA 1999;96:5274–9.
[5] Selkoe DJ. Folding proteins in fatal ways. Nature 2003;426:900–4.[6] Lambert MP, Barlow AK, Chromy BA, Edwards C, Freed R, Liosatos M, et al.
Diffusible, nonfibrillar ligands derived from Abeta1-42 are potent centralnervous system neurotoxins. Proc Natl Acad Sci USA 1998;95:6448–53.
[7] Walsh DM, Hartley DM, Kusumoto Y, Fezoui Y, Condron MM, Lomakin A, et al.Amyloid b-protein fibrillogenesis. Structure and biological activity of proto-fibrillar intermediates. J Biol Chem 1999;274:25945–52.
[8] Dahlgren KN, Manelli AM, Stine Jr WB, Baker LK, Krafft GA, LaDu MJ. Oligo-meric and fibrillar species of amyloid-beta peptides differentially affectneuronal viability. J Biol Chem 2002;277:32046–53.

K. Girigoswami et al. / Biomaterials 29 (2008) 2813–2819 2819
[9] Hamley IW. Peptide fibrillization. Angew Chem Int Ed Engl 2007;46:8128–47.[10] Pike CJ, Burdick D, Walencewicz AJ, Glabe CG, Cotman CW. Neurodegeneration
induced by beta-amyloid peptides in vitro: the role of peptide assembly state.J Neurosci 1993;13:1676–87.
[11] Lorenzo A, Yankner BA. Beta-amyloid neurotoxicity requires fibril formationand is inhibited by congo red. Proc Natl Acad Sci USA 1994;91:12243–7.
[12] St. John PA. Differential binding and activation of caspase-3 in cultured hippo-campal neuronsbyassembly forms of A beta 1-42. J Neurosci Res2007;85:1205–14.
[13] Behl C, Davis JB, Klier FG, Schubert D. Amyloid beta peptide induces necrosisrather than apoptosis. Brain Res 1994;645:253–64.
[14] Suzuki A. Amyloid b-protein induces necrotic cell death mediated by ICEcascade in PC12 cells. Exp Cell Res 1997;234:507–11.
[15] Loo DT, Copani A, Pike CJ, Whittemore ER, Walencewicz AJ, Cotman CW.Apoptosis is induced by b-amyloid in cultured central nervous systemneurons. Proc Natl Acad Sci USA 1993;90:7951–5.
[16] LeBlanc AC. The role of apoptotic pathways in Alzheimer’s disease neuro-degeneration and cell death. Curr Alzheimer Res 2005;2:389–402.
[17] Lee M, You HJ, Cho SH, Woo CH, Yoo MH, Joe EH, et al. Implication of the smallGTPase Rac1 in the generation of reactive oxygen species in response to beta-amyloid in C6 astroglioma cells. Biochem J 2002;36:937–43.
[18] Deshpande A, Mina E, Glabe C, Busciglio J. Different conformations of amyloidbeta induce neurotoxicity by distinct mechanisms in human cortical neurons.J Neurosci 2006;26:6011–8.
[19] Bush AI, Pettingell WH, Multhaup G, d Paradis M, Vonsattel JP, Gusella JF, et al.Rapid induction of Alzheimer A beta amyloid formation by zinc. Science 1994;265:1464–7.
[20] Esler WP, Stimson ER, Jennings JM, Ghilardi JR, Mantyh PW, Maggio JE. Zinc-induced aggregation of human and rat b-amyloid peptides in vitro. J Neuro-chem 1996;66:723–32.
[21] Atwood CS, Moir RD, Huang X, Scarpa RC, Bacarra NME, Romano DM, et al.Dramatic aggregation of Alzheimer Ab by Cu(II) is induced by conditionsrepresenting physiological acidosis. J Biol Chem 1998;273:12817–26.
[22] Zhu M, Souillac PO, Ionescu-Zanetti C, Carter SA, Fink AL. Surface-catalyzedamyloid fibril formation. J Biol Chem 2002;277:50914–22.
[23] Pellarin R, Guarnera E, Caflisch A. Pathways and intermediates of amyloidfibril formation. J Mol Biol 2007;374:917–24.
[24] Ha C, Park CB. Ex situ atomic force microscopy analysis of b-amyloid self-as-sembly and deposition on a synthetic template. Langmuir 2006;22:6977–85.
[25] Ha C, Ryu J, Park CB. Metal ions differentially influence the aggregation anddeposition of Alzheimer’s b-amyloid on a solid template. Biochemistry 2007;46:6118–25.
[26] Pathak R, Dey SK, Sarma A, Khuda-Bukhsh AR. Genotoxic effects in M5 cellsand Chinese hamster V79 cells after exposure to 7Li-beam (LET¼ 60 keV/microm) and correlation of their survival dynamics to nuclear damages andcell death. Mutat Res 2007;628:56–66.
[27] Carmichael J, DeGraff WG, Gazdar AF, Minna JD, Mitchell JB. Evaluation ofa tetrazolium-based semiautomated colorimetric assay: assessment ofchemosensitivity testing. Cancer Res 1987;47:936–42.
[28] Wronski R, Golob N, Grygar E, Windisch M. Two color, fluorescence-based microplate assay for apoptosis detection. Biotechniques 2002;32:666–8.
[29] Plantin-Carrenard E, Bringuier A, Derappe C, Pichon J, Guillot R, Bernard M,et al. A fluorescence microplate assay using yopro-1 to measure apoptosis:application to HL60 cells subjected to oxidative stress. Cell Biol Toxicol 2003;19:121–33.
[30] Ha C, Park CB. Template-directed self-assembly and growth of insulin amyloidfibrils. Biotechnol Bioeng 2005;90:848–55.
[31] Lizard G, Fournel S, Genestier L, Dhedin N, Chaput C, Flacher M, et al. Kineticsof plasma membrane and mitochondrial alterations in cells undergoingapoptosis. Cytometry 1995;21:275–83.
[32] Susin SA, Daugas E, Ravagnan L, Samejima K, Zamzami N, Loeffler M, et al.Two distinct pathways leading to nuclear apoptosis. J Exp Med 2000;192:571–80.
[33] Thornberry NA, Lazebnik Y. Caspases: enemies within. Science 1998;281:1312–6.
[34] Hengartner MO. The biochemistry of apoptosis. Nature 2000;407:770–6.[35] Selkoe DJ. Alzheimer’s disease: genes, proteins, and therapy. Physiol Rev 2001;
81:741–66.[36] Harper JD, Lansbury PT. Models of amyloid seeding in Alzheimer’s disease
and scrapie: mechanistic truths and physiological consequences of thetime dependent solubility of amyloid proteins. Annu Rev Biochem 1997;66:385–407.
[37] Caughey B, Lansbury PT. Protofibrils, pores, fibrils, and neurodegeneration:separating the responsible protein aggregates from the innocent bystanders.Annu Rev Neurosci 2003;26:267–98.
[38] May PC, Gitter BD, Waters DC, Simmons LK, Becker GW, Small JS, et al.b-Amyloid peptide in vitro toxicity: lot-to-lot variability. Neurobiol Aging1992;13:605–7.
[39] Fandrich M, Dobson CM. The behavior of polyamino acids reveals an in-verse side chain effect in amyloid structure formation. EMBO J 2002;21:5682–90.
[40] Dobson CM. Protein folding and misfolding. Nature 2003;426:884–90.




















