
Blocking Peptides Decrease Tissue Transglutaminase Processing of Gliadin in Vitro
Upload
independentCategory
view
0download
0
EMBRYONIC STEM CELLS/INDUCED PLURIPOTENT STEM CELLS
A Small Synthetic Cripto Blocking Peptide Improves Neural
Induction, Dopaminergic Differentiation, and Functional
Integration of Mouse Embryonic Stem Cells in a Rat Model
of Parkinson’s Disease
ENZA LONARDO,aCLARE L. PARISH,
b,c,dSALVATORE PONTICELLI,
aDANIELA MARASCO,
eDIOGO RIBEIRO,
b
MENOTTI RUVO,f SANDRO DE FALCO,a ERNEST ARENAS,b GABRIELLA MINCHIOTTIa
aStem Cell Fate Laboratory, Institute of Genetics and Biophysics, Naples, Italy, bLaboratory of Molecular
Neurobiology, Department of Medical Biochemistry and Biophysics, Karolinska Institute, Stockholm, Sweden,cFlorey Neuroscience Institutes and dCentre for Neurosciences, The University of Melbourne, Parkville, Australia,eDepartment of Biological Science, University of Naples, Napoli, Italy, fInstitute of Biostructure and Bioimaging,
CNR, Naples, Italy
Key Words. Embryonic stem cells • Neural differentiation • Cell transplantation • Cellular therapy • Parkinson’s disease
ABSTRACT
Cripto is a glycosylphosphatidylinositol-anchored corecep-tor that binds Nodal and the activin type I (ALK)-4 recep-tor, and is involved in cardiac differentiation of mouse
embryonic stem cells (mESCs). Interestingly, genetic abla-tion of cripto results in increased neuralization and mid-
brain dopaminergic (DA) differentiation of mESCs, aswell as improved DA cell replacement therapy (CRT) in amodel of Parkinson’s disease (PD). In this study, we devel-
oped a Cripto specific blocking tool that would mimic thedeletion of cripto, but could be easily applied to embryonic
stem cell (ESC) lines without the need of genetic manipu-lation. We thus screened a combinatorial peptide libraryand identified a tetrameric tripeptide, Cripto blocking
peptide (BP), which prevents Cripto/ALK-4 receptor inter-action and interferes with Cripto signaling. Cripto BPtreatment favored neuroectoderm formation and pro-
moted midbrain DA neuron differentiation of mESCs invitro and in vivo. Remarkably, Cripto BP-treated ESCs,
when transplanted into the striatum of PD rats, enhancedfunctional recovery and reduced tumor formation, mim-icking the effect of genetic ablation of cripto. We therefore
suggest that specific blockers such as Cripto BP may beused to improve the differentiation of ESC-derived DA
neurons in vitro and their engraftment in vivo, bringingus closer towards an application of ESCs in CRT. STEM
CELLS 2010;28:1326–1337
Disclosure of potential conflicts of interest is found at the end of this article.
INTRODUCTION
Cripto is a glycosylphosphatidylinositol (GPI)-anchored core-ceptor that belongs to the Epidermal Growth Factor-Cripto-1/FRL-1/Cryptic (EGF-CFC) protein family and plays importantroles during embryonic development, stem cell function, andcancer progression [1, 2]. Cripto binds to ligands of trans-forming growth factor beta (TGF-b) family such as Nodal and
Nodal-related growth differentiation factor-1 or -3, and inter-acts with the activin type I (ALK-4) receptor, thereby activat-ing a complex formed with Activin type IIB serine/threoninekinase receptor [3–5]. Upon receptor activation, the intracellu-lar effectors Smad2/Smad3 become phosphorylated and to-gether with Smad4 translocate into the nucleus to regulategene expression [6]. Cripto is thus a modulator of TGF-b sig-naling that inhibits activin signaling [7, 8] and the antiproli-ferative effects of TGFb [9–11]. Consistent with that, we
Authors contributions: E.L.: conception and design, provision of study material, collection and/or assembly of data, data analysis andinterpretation, manuscript writing; C.L.P.: conception and design, provision of study material, collection and/or assembly of data, dataanalysis and interpretation, manuscript writing; S.P.: collection and/or assembly of data, data analysis and interpretation; D.M.:collection and/or assembly of data, data analysis and interpretation; D.R.: collection and/or assembly of data; M.R.: conception anddesign, collection and/or assembly of data, provision of study material, data analysis and interpretation; S.D.F.: conception and design,collection and/or assembly of data, data analysis and interpretation; E.A.: conception and design, financial support, provision of studymaterial, data analysis and interpretation, manuscript writing; G.M.: conception and design, financial support, provision of studymaterial, data analysis and interpretation, manuscript writing. E.L, C.L.P. equally contributed as first author. E.A and G.M. equallycontributed as last author.
Correspondence: Gabriella Minchiotti, Ph.D., Stem Cell Fate Laboratory, Institute of Genetics and Biophysics, ‘‘A. Buzzati-Traverso’’,CNR, Via Pietro Castellino 111, 80,131 Naples, Italy. Telephone: 39-081-6132357; Fax: 39-081-6132595; e-mail: [email protected] Ernest Arenas, MD., Ph.D., Laboratory of Molecular Neurobiology MBB, DBRM, Karolinska Institute, Scheelevag 1, 17,177Stockholm, Sweden. Telephone: 46-8-5248-7663; Fax: 46-8-34-1960; e-mail: [email protected]; Received December 22, 2009;accepted for publication May 27, 2010; first published online in STEM CELLS EXPRESS July 16, 2010. VC AlphaMed Press 1066-5099/2009/$30.00/0 doi: 10.1002/stem.458
STEM CELLS 2010;28:1326–1337 www.StemCells.com

previously showed that Cripto acts through the Nodal/ALK-4pathway to negatively regulate neural differentiation and per-mit the entry of embryonic stem cells (ESCs) into cardiac lin-eage [12]. In support of these findings, disruption of cripto inmouse embryonic stem cells (mESCs) enhanced neurogenesisand midbrain dopaminergic (DA) differentiation in vitro [13,14]. These results suggested that Cripto blocking could beused as a tool to improve the efficiency of neural differentia-tion protocols for cell replacement therapy (CRT) in neurode-generative disorders such as Parkinson’s disease (PD) [15].
PD has been identified as a priority target for CRT becausethe classical signs of the disease (tremor, rigidity, and hypoki-nesia) are caused by the progressive degeneration of a circum-scribed cell population, the substantia nigra DA neurons. Theidea of using stem cells for CRT in PD patients has beenfueled by the finding that human ventral midbrain fetal tissuegrafts, under specific conditions, improve the symptoms andreduce the need of levodopa in these patients [16, 17]. Pluripo-tent stem cells have become an attractive cell source for CRTbecause they can be easily expanded and they can give raiseany cell type in an organism. Thus, much work has focused onthe development of methods for generating midbrain DA neu-rons from ESCs [18, 19]. However, a major remaining chal-lenge in the field is the need to improve the functional outcomeand the safety of ESC grafting [20, 21]. This has lead to the de-velopment of cell sorting strategies to eliminate unwanted cellsand/or select the desired cell types [22, 23], thus reducing oreliminating uncontrolled growth of ESCs and tumor formation[24–27]. Other strategies have focused on reducing the pres-ence of pluripotent cells and proliferating neural progenitorsby gaining further control over critical developmental proc-esses such as neural induction and neuronal differentiation. Inparticular, inhibition of bone morphogenetic protein (BMP)signaling with Noggin [28, 29], or a broader inhibition ofLefty, activin and TGFb signaling, with Noggin andSB431542, have been reported to reduce the presence of pluri-potent cells and increase neural differentiation in ESC prepara-tions [28–30]. Interestingly, transplantation of Noggin-treatedhuman Embryonic Stem (hES)-derived DA cells lead toengraftment of THþ cells and behavioral improvements inhemiparkinsonian rats, but they also lead to tumor/outgrowthformation [28, 29]. Previous results from us and others haveshown that deletion of cripto in mESCs, increases DA differen-tiation [13, 14]. Although transplantation of cripto�/� mESCs-derived embryoid body (EBs) cells at high density lead to tu-mor formation [14], when grafted at low density, they abol-ished tumor/outgrowth formation by ESCs in vivo, and pro-moted functional recovery in parkinsonian rats [13]. Thisfinding suggested that strategies aiming at achieving efficientCripto blocking, other than genetic modification, could bebroadly used to improve the DA differentiation of differentpluripotent cells. We therefore endeavored to develop a com-pound capable of inhibiting Cripto signaling in ESCs, with thegoal of using such a tool to enhance DA differentiation andimprove functional recovery after transplantation in animalmodels of PD.
For this purpose, we screened a random combinatorial tet-rameric tripeptide library built with non-natural amino acidson a multimeric scaffold, which ensured enhanced recognitioncapabilities deriving from the large recognition surface andthe high flexibility of the final compounds [31]. Peptide mix-tures composing the library were utilized as competitors ofCripto/ALK-4 interaction in an enzyme-linked immunosorbentassay (ELISA)-based competition assay, following an iterativeprocess. Selected peptides were evaluated for their ability toinhibit Cripto-induced Smad2 phosphorylation in ESCs andpromote robust neuronal differentiation. These assays allowed
us to identify a tetrameric tripeptide, named Cripto blockingpeptide (Cripto BP). The action/activity of this peptide wasfurther tested on DA differentiation of mESCs in vitro. More-over, Cripto BP-treated mESCs were transplanted in parkinso-nian rats to examine their ability to promote functional recov-ery and reduce tissue overgrowth or tumor formation.
MATERIALS AND METHODS
Iterative Deconvolution of the TetramericTripeptide Library
The screening was performed by an ELISA assay (supportinginformation text). In the first step of the deconvolution process,the peptide pools were tested with a molar excess of 30,000-fold (calculated for each single peptide) over Cripto (8.6 �10�10 M; Fig. S1). Once identified an active pool(s), a dose-de-pendent inhibition using 200-, 500-, 1,000-, 5,000-, 12,500-and 30,000-fold excess was performed to confirm the inhibi-tory property (Fig. S2). The first active pool selected was thatlabeled as 4-23-X (X indicated random positions), where ‘‘23’’identified the amino acid L-cysteine(S-benzyl) [L-Cys(Bzl)](Fig. S2). This peptide pool was resynthesized as 30 singlepeptides and submitted to the second screening using a molarexcess of 1,000-fold (calculated for each single peptide) overCripto; the screening allowed the identification of the peptide4-23-23 as the best performing in our test (Fig. S3). To finallyinvestigate the N-terminal position, where D-Glu was initiallyarbitrarily placed, 30 single peptides labeled as B-23-23 weresynthesized and submitted to the final screening (Figs. S4, S5).The peptide indicated as 26-23-23, where ‘‘26’’ identified theamino acid L-methionine-sulfoxide [L-Met(O)], was the mole-cule showing the optimal inhibitory activity. The activity ofthis peptide was further confirmed using the high-performanceliquid chromatography (HPLC) purified molecule.
ESC Culture and Differentiation
Mouse embryonic stem (ES) cell lines, both wild-type (R1[32]) and cripto�/� [33], were used throughout the study andwere cultured as previously described [34]. For in vitro car-diac differentiation as shown in Figure 3, ESCs were aggre-gated as EBs in 15% fetal bovine serum (FBS) as previouslydescribed [12, 34]. For the dopaminergic differentiation (Fig.4A), ESCs were seeded at low density onto a stromal feederlayer for 14 days in the presence of sequential morphogensand mitogens (sonic hedgehog [Shh], fibroblast growth factor2 [FGF2], FGF-8) as well as peptides (control peptide [CP] orCripto BP). See supporting information text for details.
Western Blotting and Smad2 Phosphorylation Assay
Two-day-old cripto�/� mESC-derived EBs were serumstarved for 3 hours in Dulbecco’s modified Eagle’s medium(GIBCO, Invitrogen, Carlsbad, CA, http://www.invitrogen.com/site/us/en/home/brands/Gibco.html?CID=fl-gibco) withoutleukemia inhibitory factor (LIF) and in low serum (0.5%FBS). Following starvation, EBs were incubated for 20minutes at 37�C with recombinant mouse Cripto protein([12]; 0.5 lg/ml) either alone or in the presence of the indi-cated amount of peptides. Anti-Smad2 and anti-phospho-Smad2 (Ser465/467) Antibodies (Cell Signaling Technology,Denvers, MA, http://www.cellsignal.com/) were used, follow-ing the manufacturer’s instructions.
Lonardo, Parish, Ponticelli et al. 1327
www.StemCells.com

RNA Preparation and Quantitative ReverseTranscription Polymerase Chain Reaction
Total RNAs was extracted with TRIzol kit (Life TechnologiesInc., Carlsbad, CA, http://www.lifetechnologies.com/about-life-technologies/contact-us.html) according to manufacturer’sinstructions and quantitative polymerase chain reaction (QPCR)was performed using SYBR Green PCR master mix (Euro-Clone, Milano, IT, http://www.euroclonegroup.it/), according tomanufacturer’s instructions. Primers are described in supportinginformation Table 2.
Determination of In Vitro Dopamine andDihydroxphenylacetic Acid Levels
Dopamine release and turnover was determined in vitro usingreverse phase HPLC. DA turnover was expressed as the con-centration of dihydroxphenylacetic acid (DOPAC) to DA[DOPAC/DA]. Analysis was performed on triplicate wells onthree independent differentiation experiments.
6OHDA Lesioning and Transplantation of ESCs
All experiments were performed according to the guidelinesof the European Community and were approved by the localethical committee. Sprague-Dawley rats (250–350 g; fromARC, Australia) were housed and treated according to theguidelines of the Australian and Howard Florey Institute ani-mal ethics committee (07-030).
Thirty-seven animals received unilateral stereotaxic injec-tions of 6-Hydroxydopamine (OHD) into the substantia nigrapars compacta (SNpc) to create a complete lesion of the mid-brain DA neurons, as previously described [13]. Lesioned ani-mals were selected for transplantation based on their responseto amphetamine-induced rotational behavior as previouslydescribed [13]. Animals making greater than six rotations/mi-nute (i.e., lesions greater than 80%) were selected for grafting.Two days prior to grafting animals began cyclosporine-Aimmune suppression (10 mg/kg). Fourteen days postlesioningselected animals (based on behavioral rotations) were againanesthetized and stereotaxically injected with 1 ll of cell sus-pension at each of the four sites. Stereotaxic coordinates offirst and second injection (from Bregma): anterior 0.2 mm,lateral 3.0 mm, ventral 4.5 and 5.0 mm; third and fourthinjection, anterior 1.0 mm, lateral 2.5 mm, ventral 4.5 and 5.0mm; incisor bar 0 mm [35]). Wild-type mESCs were differen-tiated as outlined in Fig. S6a, including either CP or CriptoBP; media was also supplemented with Noggin (300 ng/ml,day 0–8, to enhance neuronal induction) as well as Wnt5a(200 ng/ml, day 8–11), to promote DA neuron differentiation.Following in vitro differentiation, ESC colonies were dis-sected away from the stromal feeder layer and dissociatedusing collagenase/dispase with agitation (700 lg/ml, Roche,for 20 minutes, 80 rpm). Cells were subsequently resuspendedat a density of 3,000 cells/ll in preparation for grafting. Fiverats received Sham injections (4 ll N2 media, 1 ll/site), 16rats received a total of 12,000 mESCs-derived cells (4 � 1 llgrafts) pretreated with the CP and a further 16 rats receivedgrafts pretreated with the Cripto BP.
Animals were tested for functional improvements at 2, 4,6, and 8 weeks post-transplantation by amphetamine-inducedrotational behavior. Eight weeks postgrafting, rats were killedby an overdose of sodium pentobarbital (100 mg/kg i.p.) andintracardially perfused as previously described [13]. Brainswere removed, postfixed for 1 hour, and then left overnight at4�C in 20% sucrose in phosphate-buffered saline (PBS). Thefollowing day, 20-lm-thick coronal sections were cut seriallythrough the striatum (12 series) and 50 lm through the SNpcand mounted directly onto chrom alum gelatinized slides.
Histological Analysis
Immunofluorescence analysis were performed as previouslydescribed [12, 13]. The following primary antibodies wereused: mouse anti-bIII-tubulin (Tuj1, 1:1,000, Promega, Madi-son, WI, http://www.promega.com); rabbit anti-tyrosinehydroxylase (TH; 1:250, PelFreez, Rogers, AR, http://www.pelfreez-bio.com/); sheep anti-TH (1:250, PelFreez,Rogers, AR, http://www.pelfreez-bio.com/); Nurr-1 (1:500,Santa Cruz, CA, USA, http://www.scbt.com/); rabbit anti-Pitx3 (1:200, a gift from P. Burbach, Rudolf Magnus Instituteof Neuroscience, Utrecht), rabbit anti-GIRK-2 (1:100, Chemi-con, Temecula, CA, http://www.millipore.com/company/cp1/redirect-ab), rabbit anti-GABA (1:500, Sigma-Aldrich, St.Louis, MO, http://www.sigmaaldrich.com/customer-service.html); rabbit anti-5HT (1:200, Biogenesis Ltd, Dorset, UK,www.biogenesis.co.uk/); rabbit anti-NET (1:20, Chemicon;rabbit anti-cytokeratin (1:4,000, DAKO, Glostrup, DK, http://www.dako.com/); rat anti-CD29 (1:200, BD Pharmingen, SanJose, CA, http://www.bdbiosciences.com/), rabbit anti-Ki67(Neomarkers Thermofisher, CA, USA, http://www.thermofisher.com/global/en/about/home.asp), mouse anti-nestin (1:200,Chemicon, Temecula, CA, http://www.millipore.com/company/cp1/redirect-ab); mouse anti-Oct4 (1:100, Santa Cruz, SantaCruz, CA, USA, http://www.scbt.com/), rabbit anti-glial fibril-lary acidic protein, (GFAP, 1:200, DAKO, Glostrup, DK,http://www.dako.com/). Appropriate fluorophore conjugatedsecondary antibodies: (Cyanin-2, Cy3, and Cy5, 1:200, JacksonImmunoResearch, Suffolk, UK, http://www.jireurope.com/home/contact.asp) were used for visualization. Labeling wasvisualized by light or fluorescent illumination using an invertedmicroscope (DMIRB, Leica Microsystems, Wetzelar, DE,http://www.leica-microsystems.com) or upright microscope(Leica DMLB2). Images were acquired on a DC 350 FX cam-era (Leica).
The grading analysis (Fig. 3) was performed blinded bytwo investigators; to get a reliable unbiased analysis, all theEBs formed in suspension culture were individually plated in48 multiwell plates and used for morphological (Fig. 3A, 3B)and immunofluorescence analysis (Fig. 3C, 3D). The propor-tion of neuron containing colonies (labeled using the imma-ture neuronal marker bIII-tubulin, Tuj1) and TH neuron con-taining colonies were counted and expressed as a percentageof total colonies (Fig. 4). The total number of dopaminergicneurons/lm2 colony (identified using a marker for the precur-sor enzyme in DA synthesis, TH) were also counted (Fig. 4).Experiments were performed in triplicate.
Semiquantitative scoring was employed to estimate thecell composition of the tumors, as previously described [14].Tumors were stained and scored by blinded investigatorsusing the following scales: 0 ¼ no cells (0%); 1 ¼ few cells(<5%); 2 ¼ moderate cells (5–20%); 4 ¼ dominant cell type(>50%). One series of slides containing grafted striata wascounterstained using 1% Neutral red to visualize the graft/tu-mor size and the gross morphology of the striatum. Area ofgrafts and tumors was calculated using Stereo Investigatorsoftware (MicroBrightField, VT).
Statistical Analysis
Values are expressed as means SEM or SD. The statisticalsignificance of difference was determined by ANOVA fol-lowed by Tukey’s post hoc test or a Student’s t-test. Rankingof in vitro differentiation (Fig. 3C) was performed using aWilcoxon rank sum test. Statistical Package for Social Scien-ces software package, version 11.0 SPSS, Chicago, IL andSigma Stat 3.0 software were used for statistical analysis withsignificance set at p < .05.
1328 Small Molecules for Stem Cell Therapies

RESULTS
Identification of a Novel Peptide that AntagonizesCripto/ALK-4 Receptor Interaction
To identify Cripto/ALK-4 antagonists capable of disruptingdownstream signaling, we screened a tetrameric tripeptidelibrary synthesized using 29 non-natural amino acids toincrease peptide stability, plus Glycine, (supporting informa-tion Table 1). To increase the molecular surface, the librarywas assembled as previously described [31] on a peptide scaf-fold composed of a poly-lysine core [36]. To facilitate thecharacterization of the peptide mixtures, and at the same timeto expedite the screening procedure, the first residue was arbi-trarily fixed. A negatively charged amino acid, the D-glutamicacid (D-Glu, denoted 4; Fig. 1A and supporting information Ta-ble 1), was thus initially chosen as the common, N-terminalresidue, to favor peptide solubilization. The library wasarranged in 30 pools, each of them composed of 30 tetramerictripeptides, which differ for the amino acid (denoted B) on thesecond position (Fig. 1A), while the residue in the third posi-tion (denoted X) was randomly added. The total complexity ofthe library was therefore of 302 ¼ 900 different peptides. Thescreening was carried out by an ELISA-based competitionassay whereby the displacement of soluble recombinant Cripto
from coated ALK-4 by the peptide pools was evaluated. Theiterative procedure consisted of three screening rounds, whichled to the selection of the 26-23-23 tetramer formed by L-me-thionine-sulfoxide [L-Met(O)] (denoted as 26) in position 1,and S-benzylated L-cysteine [Cys(Bzl)] (denoted as 23) in posi-tions 2 and 3 of the tripeptide (Fig. 1A), (for details see sup-porting information Text and Figs. 1–4). The selected tetra-meric tripeptide (26-23-23, supporting information Fig. 5) wasthereafter named Cripto BP. This peptide blocked the interac-tion of Cripto and ALK-4 with an estimated IC50 of 400 nM,while the control tetrameric Peptide (CP), D-Ser-Ala-Cha, anda dimeric peptide (DP) variant of the 26-23-23 peptide had noeffect (Fig. 1B). We thus performed ELISA assays to evaluatewhether Cripto BP- or CP-coated to microtiter plates couldbind recombinant Cripto and/or ALK-4 receptor. Cripto butnot ALK-4 specifically recognized the immobilized Cripto BP,in a dose-dependent manner (Fig. 1C). Neither Cripto norALK-4 recognized the CP, confirming the specificity of CriptoBP for Cripto binding.
Cripto BP Antagonizes Cripto Signaling, ImpairingCardiomyogenesis, and Promoting Neurogenesisin mESCs
Given the neutralizing activity of the peptide, we first askedwhether Cripto BP was able to block Smad2-dependent Cripto
Figure 1. (A): Schematic representation of the iterative screening strategy. (1) NH2-D-Glu (D-glutamic acid, also denoted as ‘‘4’’, see supportinginformation Table 1) is the common N-terminal residue, (B) identifies the fixed residue, which denotes 1 of 30 amino acids (aa1-30, supporting in-formation Table 1) and X indicates random positions (4-B(1–30)-X). The total complexity of the first library was therefore of 302 ¼ 900 differentpeptides. In the first screening round (1), 4.23.X identifies the first active pool, where ‘‘23’’ indicates L-cysteine(S-benzyl) [L-Cys(Bzl)]. In the secondround of screening (2) the pool 4-23-X was re-synthesized in 30 single peptides (4-23-B(1–30)), and the residue ‘‘23’’ was identified in the third posi-tion (4-23-23). Finally, 30 single peptides (B(1–30)-23-23) were submitted to the third screening round (3), which identified the residue ‘‘26,’’ where‘‘26’’ indicates L-methionine-sulfoxide [L-Met(O)]. The 26-23-23 peptide is also referred to as Cripto BP. (B) Dose-dependent inhibition of Cripto/ALK-4 interaction exerted by Cripto BP, CP (i.e., tetrameric tripeptide D-Ser-Ala-Cha) and dimeric peptide (DP; i.e., dimeric tripeptide 26-23-23) inELISA-based competition assays (n ¼ 3). (C): Dose-dependent interaction of Cripto or ALK-4 to Cripto BP. The indicated peptides (BP and CP)were coated (50 lM) on the plates and increasing amounts of either Cripto or ALK-4 receptor were used in binding, ranging from 50 to 200 ng/ml.Cripto but not ALK-4 bound Cripto BP; whereas, both Cripto and ALK-4 failed to bind the CP. Mean 6 SD, *, p < .0005. Abbreviations: ALK-4,activin type I receptor; BP, blocking peptide; CP, control peptide; CRIPTO, Cripto; ELISA, enzyme-linked immunosorbent assay.
Lonardo, Parish, Ponticelli et al. 1329
www.StemCells.com

signaling in mESCs. We thus evaluated whether Cripto BP wasable to prevent Cripto-induced Smad2 phosphorylation of serumstarved 2-day-old cripto�/� EBs. Recombinant soluble Criptoprotein stimulated Smad2 phosphorylation, as assessed by West-ern blot analysis ([12] and Fig. 2); this effect was not modifiedby the presence of increasing amounts of either Cripto DP orCP (2.5 lM and 25 lM; Fig. 2). On the contrary, Cripto-induced Smad2 phosphorylation was inhibited by Cripto BPand completely blocked at 25 lM (125� molar excess; Fig. 2).We then assessed whether Cripto BP was able to antagonize theendogenous Cripto signaling and thus mimicking the effect ofgenetic ablation of cripto in a mESC differentiation assay, thatis, blocking cardiomyogenesis and inducing neuronal differen-tiation, under the well-established conditions, which promotecardiac differentiation of mESCs [12, 37]. Two day-old wild-type EBs were treated with increasing amounts of either CP orCripto BP, ranging from 5.0–25 lM, and the percentage of EBscontaining beating areas was scored from day 8 to 12 of the dif-ferentiation (n ¼ �60 EBs/group; Fig. 3B). Given that the pep-tides were dissolved in dimethyl sulfoxide (DMSO), controluntreated EBs were compared with DMSO-treated EBs. Inter-estingly, neither DMSO, the tetrameric CP (Fig. 3B), nor theDP (data not shown) affected the percentage of beating EBs.Instead, treatment of wild-type EBs with Cripto BP, induced adose-dependent inhibition of cardiomyogenesis, as shown by theprogressive reduction of rhythmically contracting EBs (at 25lM, BP 3.7% 6 3.5 vs. CP 82.9% 6 11.17, **, p < .0001;Fig. 3B).
We previously showed that cripto�/� ESCs spontaneouslydifferentiate into neurons during the time window when wild-type ESCs are primed for cardiomyogenesis by Cripto [12].Thus, if specific and effective, the Cripto BP should be ableto redirect ESC fate and to promote neural induction and neu-rogenesis in mESCs, even if they are cultivated in conditions(i.e., EBs in 15% FBS) that favor cardiac differentiation [34].
To evaluate the peptide(s) activity, 2-day-old wild-type EBswere treated with either CP, DP, or BP and both morphologi-cal and immunofluorescence analysis were performed on 13-day-old EBs (Fig. 3A). To better define the activity of thepeptides, we used anti-bIII tubulin antibodies to identify neu-rons in culture and arbitrarily defined four grades of neuronaldifferentiation (Fig. 3C) ranging from the absence of neurons(Grade 0) to full neuronal differentiation (Grade 3), that is,presence of a dense network of bIII-tubulin-positive cells.The presence of either few isolated neurons or small areas ofbIII-tubulin positive cells defined intermediate phenotypes,grade 1 and grade 2, respectively (Fig. 3C). Our data clearlyindicated that Cripto BP but not DP or CP treatment promotesrobust neuronal differentiation, characterized by a high per-centage of grade 3 (full neuronal differentiation, Fig. 3C) phe-notype (BP 45.6% 6 5.2 vs. CP 6.9% 6 0.6, Mean 6 SD *,p � .03; Fig. 3D). Accordingly, upon treatment with controlpeptides (CP or DP), the majority of the EBs scored showeda grade 0 (CP 75.9% 6 3.05 and DP 77.2% 6 7, vs. BP17.4% 6 1.15, Mean 6 SD, *, p � .03; Fig. 3D). To supportthese findings, we performed the molecular analysis andexamined the expression profile of neuronal differentiationmarkers in the same cardiac differentiation conditions, byquantitative reverse transcription polymerase chain reaction(QRT-PCR). In line with our previous findings in cripto�/�
ESCs [12], expression of the pan neuronal marker NFM wassignificantly induced in 13-day-old EBs upon treatment withCripto BP. Concomitantly, expression of the cardiac specificmarker myosin light chain-2 ventricular as well as the mesen-doderm marker alpha-fetoprotein were strongly reduced (Fig.3E). Thus, our results suggest a fate switch in ESCs frommesoderm/cardiomyocytes to neurons. Accordingly, expres-sion of the panmesodermal marker Brachyury/T was alsostrongly reduced in 2-day-old EBs upon treatment with CriptoBP (data not shown). Moreover, Cripto BP-treated wild-typeEBs also showed induction of markers typical of DA develop-ment and differentiation such as, Pitx3 [38], Shh [39], Wnt5a[40], TH (the rate limiting enzyme in DA synthesis) and do-pamine transporter (DAT; Fig. 3E), in line with previous find-ings in cripto�/� ESCs [12]. Thus, our results suggest thatCripto BP can enhance the neural differentiation of ES cells.
Increased Dopaminergic Differentiation of ESCsTreated with Cripto BP
We next examined whether Cripto BP treatment was able tofurther enhance DA differentiation of mESC, using the stro-mal cell-based protocol described by Barberi et al. [41], withminor modifications (Fig. 4A). In brief, mESCs were plated atlow density onto stromal cells (PA6) for a period of 5 days,to promote neural induction. At day 5 the media was replen-ished and supplemented with Shh and FGF-8 (for ventral mid-brain patterning). From day 8 to 11, cultures where changedinto neuronal (N2) media in the presence of Shh and FGF-8as well as basic FGF (to expand the pool of DA precursors).Finally (day 11–14), the cells were exposed to the survivalfactors brain-derived neurotrophic factor, glial cell line-derived neurotrophic factor (GDNF), and ascorbic acid. In allcultures, the CP, or Cripto BP was added to the differentiatingESCs from day 1 to day 11 (Fig. 4A).
We first examined whether immature and proliferatingcells remained in the cultures at the end of differentiation,day 14. Interestingly, Oct4þ cells (a pluripotent stem cellmarker) as well as Ki67þ cells (a nuclear cell cycle marker)were present both in CP- and BP-treated cultures. However,while extremely few/rare Oct4þ cells were identified in CP(not shown) and BP (Fig. 4B), Ki67þ cells were relatively
Figure 2. Cripto BP prevents Cripto-induced Smad2 phosphoryla-tion. Two-day-old cripto�/� mouse embryonic stem cells-derivedembryoid bodies were stimulated with recombinant Cripto protein(0.5 lg/ml) either alone or in the presence of increasing amount (2.5lM and 25 lM) of the peptides: DP (dimeric 26-23-23), CP (controltetrameric D-Ser-Ala-Cha) and BP (tertrameric 26-23-23), or leftuntreated as control. Cripto BP, but not CP or DP inhibits Cripto-induced Smad2 phosphorylation, as shown by Western blot, usinganti-phospho-Smad2 antibodies. Levels of total Smad2 were alsocompared. Smad2 phosphorylation was expressed as ratio between ar-bitrary densitometric units of P-Smad2 and Smad2. Data are mean 6SEM, n � 3. Abbreviations: BP, blocking peptide; CP, control pep-tide; DP, dimeric peptide.
1330 Small Molecules for Stem Cell Therapies

abundant both in CP (not shown) and BP cultures (Fig. 4C,BP). To further characterize the ESC cultures, we thus per-formed immunocytochemistry against nestin (neuroectoderm),cytokeratin (ectoderm/epidermis) and CD29 (mesoderm andectoderm). We found an increase in the presence of neuralrosettes and nestin staining in BP cultures (Fig. 4D, 4E) com-pared with CP cultures (Fig. 4E0). Although cytokeratinþcells were detected in both CP- and BP-differentiated cultures(Fig. 4F, BP), few CD29þ cells were present only in CP cul-tures (Fig. 4G), suggesting that Cripto BP prevented meso-derm differentiation to promote neural induction, as describedfor cripto�/� ESCs [13, 14]. The efficacy of the neural induc-tion was also evident by the abundance of both neurons(Tuj1) and glial cells (GFAP) in the BP cultures (Fig. 4H).
We subsequently assessed the effect of BP on neuronaland DA differentiation in these culture conditions. First, wefound that while treatment of cultures with CP had no effecton neural induction or TH differentiation, a clear increasewas detected with BP (Fig. 4I–4K). Following differentiationin the presence of the Cripto BP, the majority of colonieswithin the cultures were immunoreactive for bIII-tubulin
(91% compared with 66% in CP, Fig. 4I). Furthermore,Cripto BP was capable of significantly increasing the propor-tion of colonies containing TH-ir DA neurons (62%) com-pared with 34% in CP (Fig. 4J). Additionally, these TH-ir col-onies contained significantly more DA neurons (THþ cells/lm colony, 286%, Fig. 4K–4M). Interestingly, we found thatDA differentiation of Cripto BP-treated ESCs could be furtherenhanced by addition of Noggin [28, 29] and Wnt5a [40, 42](supporting information Fig. 6). Many of the TH-labeled neu-rons (in CP- and Cripto BP-treated cultures) showed a bipolarmorphology (Fig. 4N). Immunohistochemistry for Nurr1,Pitx3, and G protein-activated inwardly rectifying potassiumchannel two (GIRK2; the latter, a marker preferentiallyexpressed in SNpc DA neurons [43]), confirmed that TH-ircells within the cultures (CP or BP treated) were of a mid-brain DA phenotype (Fig. 4O–4Q) and, at least a notable pro-portion possessed a SNpc phenotype (Fig. 4Q). Interestingly,few cells within the cultures were immunoreactive for otherneuronal subpopulations markers including c-aminobutyricacid (GABA-ir) and serotonin (5HT-ir) (Fig. 4R–4S). Further-more, we analyzed the ability of the dopamine cells generated
Figure 3. Cripto BP prevents cardiomyogenesis and induces neurogenesis in wild-type ESCs. (A): In vitro cardiac differentiation scheme of wild-type ESCs. (B): Cripto BP (but not CP or DMSO) prevented cardiomyogenesis. Peptides were added to 2-day-old wild-type mouse ESCs-derivedEBs; the percentage of EBs with rhythmically contracting areas was scored on day 13 (n ¼ �60 EBs/group; data are mean 6 SD; *, p < .001, **, p< .0001). (C): Grades of neuronal differentiation of wild-type EBs, as arbitrarily defined. Grade 0: absence of neurons; Grade 1: few, isolated neuronsimmunoreactive to b-III tubulin antibodies; Grade 2: small areas of b-III tubulin-ir cells; Grade 3: dense network of b-III tubulin-ir cells. Nuclei werevisualized by 40,6-diamidino-2-phenylindole (DAPI) counterstaining. Scale bar (A–C) ¼ 100 lM, (D) ¼ 75 lM in. (D): BP (but not CP or DP) pro-moted robust neuronal differentiation in wild-type EBs. Two-day-old wild-type EBs were treated with 25 lM of the indicated peptides and immuno-fluorescence analysis was performed on day 13 (n ¼ �60 EBs/group, data are mean 6 SD; **, *, p � .03). The degree of neuronal differentiationwas scored according to panel (C). (E): Gene expression analysis of mesoderm and neuronal markers. Two-day-old wild-type EBs were treated eitherwith the indicated peptides at 25 lM (CP; BP) or vehicle (DMSO) and the expression profile of mesoderm/cardiac and neuronal markers was ana-lyzed on day 13, by quantitative reverse transcription polymerase chain reaction. mRNA was normalized to glyceraldehyde 3-phosphate dehydrogen-ase (GAPDH) and presented as fold change in gene expression relative to the control (DMSO ¼ 1). Data are mean 6 SD, n � 3; *, p < .05 **, p <.01. Abbreviations: AFP, alpha-fetoprotein; BP, blocking peptide; CP, control peptide; DAT, dopamine transporter; DMSO, dimethyl sulfoxide; DP,dimeric peptide; EB, embryoid body; ESC, embryonic stem cell; NFM, neurofilament medium polypeptide; TH, tyrosine hydroxylase.
Lonardo, Parish, Ponticelli et al. 1331
www.StemCells.com

in vitro to synthesize, release and utilize dopamine. Althoughboth Cripto BP and CP-treated cultures were able to utilizeDA (as reflected by dopamine turnover; ratio of DOPAC toDA, Fig. 4T0 0 0), we noted that Cripto BP treatment of mESCs
significantly increases the KCl-evoked release of dopamine(Fig. 4T0) and DOPAC (Fig. 4T00) within the cultures, reflec-tive of enhanced DA differentiation. Thus, our results showthat Cripto BP, in addition to promoting neural induction (as
Figure 4.
1332 Small Molecules for Stem Cell Therapies
shown in EB cultures, Fig. 3), was also capable of enhancingthe number of DA neurons by 2.8-folds in a neural media(Fig. 4).
Enhanced Functional Recovery in ParkinsonianRats Receiving ESC Grafts Treated with Cripto BP
We next examined the ability of mESC-derived DA neuronsto survive, integrate and induce functional recovery in an ani-mal model of PD. Previous studies on the function of cripto
in the DA differentiation of ESCs in vivo have been per-formed using cripto�/� ESCs grown for 4 days as EBs [13,14]. Given the high yield of DA neurons in wt mESCs treatedwith Cripto BP, cells differentiated as described in supportinginformation Figure 6, in the presence of either Cripto BP orCP, were used for transplantation in 6-hydroxydopaminelesioned rats, an animal model of PD. The validity of themodel was assessed by amphetamine-induced rotationalbehavior and histological examination, which confirmed thatall animals had near complete unilateral lesions of SNpc DA
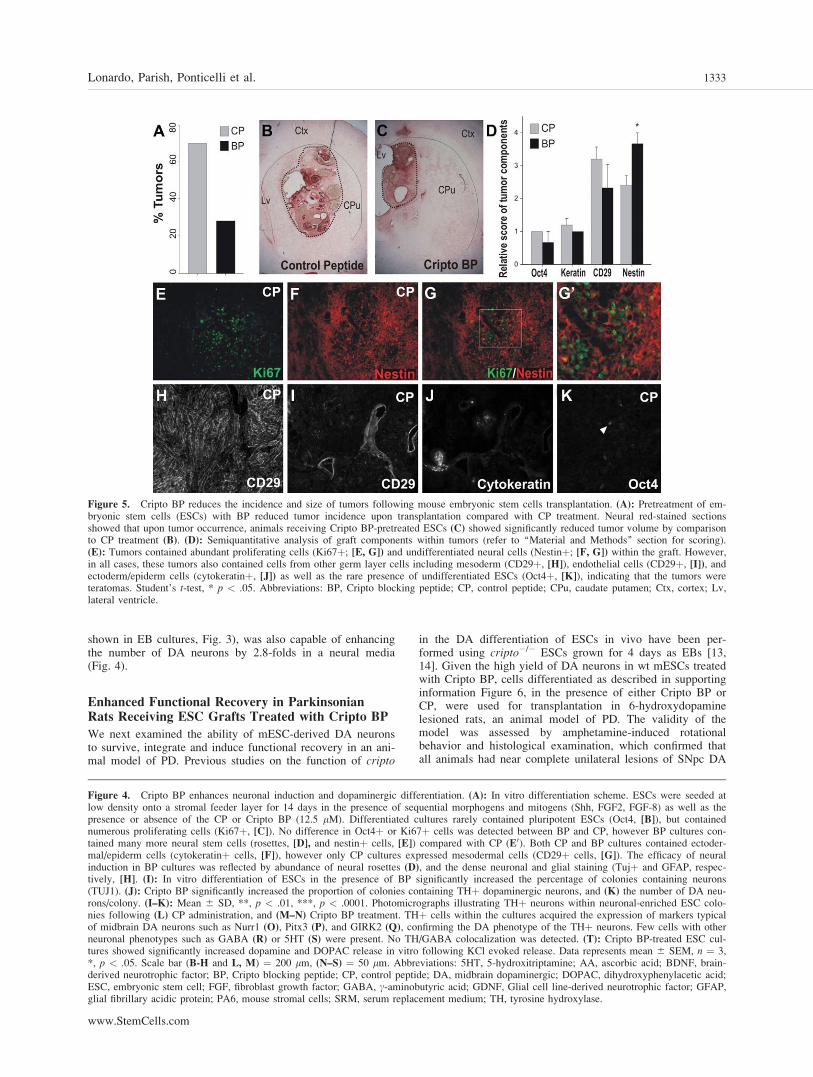
Figure 5. Cripto BP reduces the incidence and size of tumors following mouse embryonic stem cells transplantation. (A): Pretreatment of em-bryonic stem cells (ESCs) with BP reduced tumor incidence upon transplantation compared with CP treatment. Neural red-stained sectionsshowed that upon tumor occurrence, animals receiving Cripto BP-pretreated ESCs (C) showed significantly reduced tumor volume by comparisonto CP treatment (B). (D): Semiquantitative analysis of graft components within tumors (refer to ‘‘Material and Methods’’ section for scoring).(E): Tumors contained abundant proliferating cells (Ki67þ; [E, G]) and undifferentiated neural cells (Nestinþ; [F, G]) within the graft. However,in all cases, these tumors also contained cells from other germ layer cells including mesoderm (CD29þ, [H]), endothelial cells (CD29þ, [I]), andectoderm/epiderm cells (cytokeratinþ, [J]) as well as the rare presence of undifferentiated ESCs (Oct4þ, [K]), indicating that the tumors wereteratomas. Student’s t-test, * p < .05. Abbreviations: BP, Cripto blocking peptide; CP, control peptide; CPu, caudate putamen; Ctx, cortex; Lv,lateral ventricle.
Figure 4. Cripto BP enhances neuronal induction and dopaminergic differentiation. (A): In vitro differentiation scheme. ESCs were seeded atlow density onto a stromal feeder layer for 14 days in the presence of sequential morphogens and mitogens (Shh, FGF2, FGF-8) as well as thepresence or absence of the CP or Cripto BP (12.5 lM). Differentiated cultures rarely contained pluripotent ESCs (Oct4, [B]), but containednumerous proliferating cells (Ki67þ, [C]). No difference in Oct4þ or Ki67þ cells was detected between BP and CP, however BP cultures con-tained many more neural stem cells (rosettes, [D], and nestinþ cells, [E]) compared with CP (E0). Both CP and BP cultures contained ectoder-mal/epiderm cells (cytokeratinþ cells, [F]), however only CP cultures expressed mesodermal cells (CD29þ cells, [G]). The efficacy of neuralinduction in BP cultures was reflected by abundance of neural rosettes (D), and the dense neuronal and glial staining (Tujþ and GFAP, respec-tively, [H]. (I): In vitro differentiation of ESCs in the presence of BP significantly increased the percentage of colonies containing neurons(TUJ1). (J): Cripto BP significantly increased the proportion of colonies containing THþ dopaminergic neurons, and (K) the number of DA neu-rons/colony. (I–K): Mean 6 SD, **, p < .01, ***, p < .0001. Photomicrographs illustrating THþ neurons within neuronal-enriched ESC colo-nies following (L) CP administration, and (M–N) Cripto BP treatment. THþ cells within the cultures acquired the expression of markers typicalof midbrain DA neurons such as Nurr1 (O), Pitx3 (P), and GIRK2 (Q), confirming the DA phenotype of the THþ neurons. Few cells with otherneuronal phenotypes such as GABA (R) or 5HT (S) were present. No TH/GABA colocalization was detected. (T): Cripto BP-treated ESC cul-tures showed significantly increased dopamine and DOPAC release in vitro following KCl evoked release. Data represents mean 6 SEM, n ¼ 3,*, p < .05. Scale bar (B-H and L, M) ¼ 200 lm, (N–S) ¼ 50 lm. Abbreviations: 5HT, 5-hydroxitriptamine; AA, ascorbic acid; BDNF, brain-derived neurotrophic factor; BP, Cripto blocking peptide; CP, control peptide; DA, midbrain dopaminergic; DOPAC, dihydroxyphenylacetic acid;ESC, embryonic stem cell; FGF, fibroblast growth factor; GABA, c-aminobutyric acid; GDNF, Glial cell line-derived neurotrophic factor; GFAP,glial fibrillary acidic protein; PA6, mouse stromal cells; SRM, serum replacement medium; TH, tyrosine hydroxylase.
Lonardo, Parish, Ponticelli et al. 1333
www.StemCells.com
neurons (cell loss >90% and >6 rotations/minute; data notshown). When the viability of the grafts was examined, wefound that 97% of the animals (31 of 32 rats) transplantedwith 12,000 mESC-derived cells showed viable grafts at 8weeks. The majority of the rats receiving ESCs pretreatedwith CP showed the presence of tumors/overgrowths withinthe grafts (Fig. 5A, 5B). Interestingly, tumor formation wasreduced by more than half, but not abolished, in rats receivingESCs pretreated with Cripto BP (Fig. 5A, 5C). To determinethe nature of these tumors (i.e., neural overgrowths or terato-mas), we examined the grafts by immunohistochemistry andscored the cell types (Fig. 5E–5K, see ‘‘Methods’’ section).
The majority of these tumors (all CP-pretreated and 3/4 ofBP-pretreated ESCs) contained ectodermal-epidermal andmesodermal cells, indicating that they were teratomas. Thiswas confirmed by the presence of scarce Oct4þ cells in thesegrafts (Fig. 5K). Thus, while mesodermal cells were notobserved after Cripto BP treatment in vitro, these cellsappeared in vivo, indicating that the level of antagonism maynot be sufficient to prevent mesoderm formation from the fewremaining ESCs in vivo [14].
Interestingly, despite more nestinþ cells were detected inBP compared with CP-pretreated ES cells (Fig. 5D), we didnot detect areas with very high cell density/parenchymal
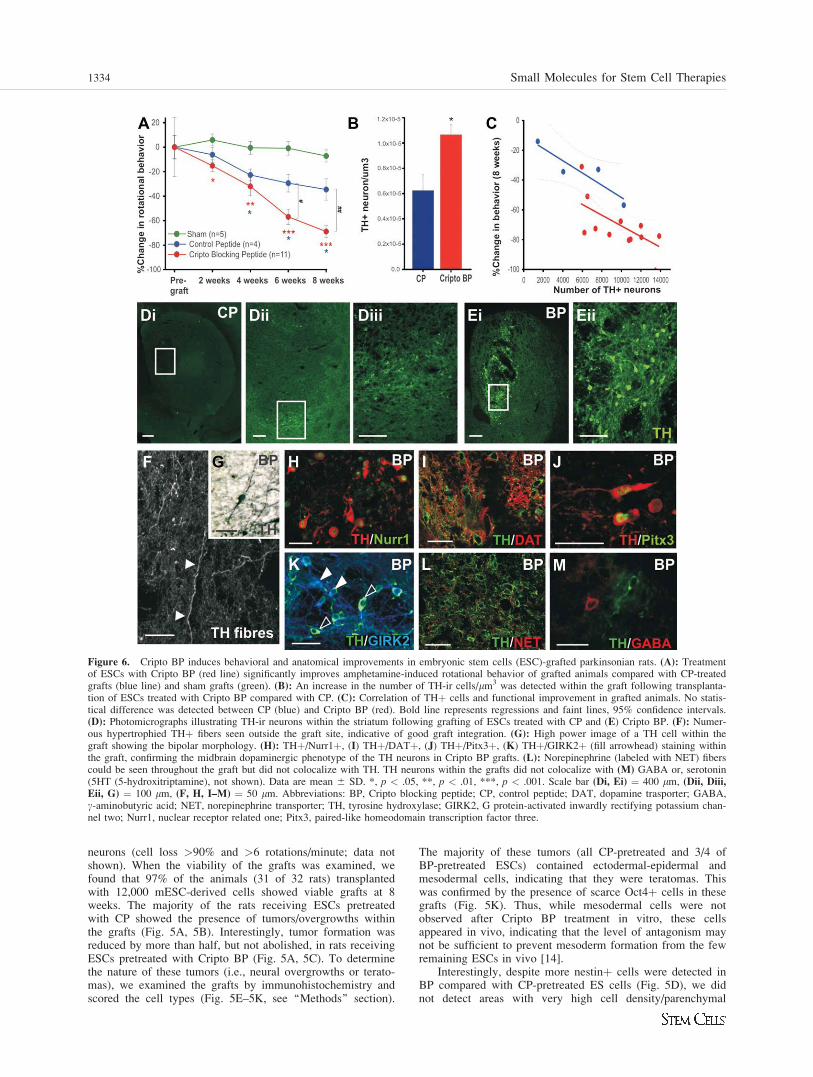
Figure 6. Cripto BP induces behavioral and anatomical improvements in embryonic stem cells (ESC)-grafted parkinsonian rats. (A): Treatmentof ESCs with Cripto BP (red line) significantly improves amphetamine-induced rotational behavior of grafted animals compared with CP-treatedgrafts (blue line) and sham grafts (green). (B): An increase in the number of TH-ir cells/lm3 was detected within the graft following transplanta-tion of ESCs treated with Cripto BP compared with CP. (C): Correlation of THþ cells and functional improvement in grafted animals. No statis-tical difference was detected between CP (blue) and Cripto BP (red). Bold line represents regressions and faint lines, 95% confidence intervals.(D): Photomicrographs illustrating TH-ir neurons within the striatum following grafting of ESCs treated with CP and (E) Cripto BP. (F): Numer-ous hypertrophied THþ fibers seen outside the graft site, indicative of good graft integration. (G): High power image of a TH cell within thegraft showing the bipolar morphology. (H): THþ/Nurr1þ, (I) THþ/DATþ, (J) THþ/Pitx3þ, (K) THþ/GIRK2þ (fill arrowhead) staining withinthe graft, confirming the midbrain dopaminergic phenotype of the TH neurons in Cripto BP grafts. (L): Norepinephrine (labeled with NET) fiberscould be seen throughout the graft but did not colocalize with TH. TH neurons within the grafts did not colocalize with (M) GABA or, serotonin(5HT (5-hydroxitriptamine), not shown). Data are mean 6 SD. *, p < .05, **, p < .01, ***, p < .001. Scale bar (Di, Ei) ¼ 400 lm, (Dii, Diii,Eii, G) ¼ 100 lm, (F, H, I–M) ¼ 50 lm. Abbreviations: BP, Cripto blocking peptide; CP, control peptide; DAT, dopamine trasporter; GABA,c-aminobutyric acid; NET, norepinephrine transporter; TH, tyrosine hydroxylase; GIRK2, G protein-activated inwardly rectifying potassium chan-nel two; Nurr1, nuclear receptor related one; Pitx3, paired-like homeodomain transcription factor three.
1334 Small Molecules for Stem Cell Therapies

distortion containing nestinþ cells and Ki67þ cells in the ab-sence of mesodermal and endodermal cells, indicating thatthere were no cases of isolated neural overgrowth (data notshown). Remarkably, in the cases where tumors wereobserved, animals receiving ESCs pretreated with Cripto BPshowed a significant reduction in tumor volume (0.94 � 1010
lm3 6 0.27 � 1010) compared with CP (2.22 � 1010 lm3 60. 44 � 1010, Mean 6 SEM, *, p < .05), as assessed in neu-ral red-stained sections (Fig. 5B–5C). Thus, our results indi-cate that Cripto BP reduces the incidence and size of mESC-derived teratomas by �60%.
We next examined the impact of ESC transplantation onamphetamine-induced circling behavior and dopamine neuronengraftment in rats that did not present any tumor (n ¼ 11Cripto BP grafts and n ¼ 4 CP grafts, 15 BP grafts and 16 CPgrafts of the total). Functionally, the motor behavior of parkin-sonian rats receiving ESCs grafts significantly improved by 8weeks compared with sham-treated animals (Fig. 6A). Graftingof 12,000 ESCs pretreated with CP showed modest levels offunctional improvement compared with shams from 4 to 8weeks. Most remarkably, animals receiving the same number ofESCs treated with the Cripto BP showed significant reductionsin amphetamine-induced rotational behavior as early as week 2postgrafting. Moreover, the reduction in motor asymmetry pro-gressively (and significantly) improved from 6 weeks postgraft-ing compared with CP grafts (Fig. 6A).
Subsequently, we examined whether the functional improve-ment observed in Cripto BP-treated animals was related to anincrease in TH-ir neuron counts within the grafts of tumor-freeanimals. Importantly, the volume of the grafts was not signifi-cantly different between treatments (CP ¼ 8.79 � 108 6 1.20 �108 and BP ¼ 9.43 � 108 6 0.88 � 108, p ¼ .687), and no THneurons were found within the striatum of animals receivingsham grafts (data not shown). When we examined the number ofTH cells, we found that animals grafted with Cripto BP-treatedESCs showed a consistent increase in THþ cells/lm3 comparedwith CP-treated ESCs (p ¼ .046, Fig. 6B–6E). However, wefound no statistically significant difference between CP and BP-pretreated ESC grafts when examining the correlation betweenbehavioral improvement and THþ neuron number (Fig. 6C),indicating that the improvement is mainly contributed by anincrease in THþ neurons. We observed that many of the THþcells within the grafts displayed a bipolar morphology (Fig. 6G)as well as an extensive network of hypertrophied neuritesthroughout the striatum (Fig. 6F). Immunohistochemistryrevealed that many of the TH-ir cells within the grafts also coex-pressed Nurr1, DAT, Pitx3, and GIRK2, verifying their midbrainDA identity (Fig. 6H–6K). Interestingly, Pitx3/TH and GIRK2/TH colabeling illustrated that several THþ neurons within thegrafts were SNpc midbrain-like DA neurons, both in CP and BPgrafts, (filled arrowheads in Fig. 6K). Although norepinephrinefibers (labeled with norepinephrine transporter, NET) could beseen within the grafts (Fig. 6L), no TH-ir cell bodies within thegraft colocalized with other neurotransmitters, including norepi-nephrine (Fig. 6L), GABA (Fig. 6J), and serotonin (not shown).
In summary, our results illustrate that a synthetic smallpeptide, Cripto BP, improves the generation of midbrain DAneurons from ESCs in vitro and in vivo, reduces the incidenceof teratomas and results in a behavioral improvement in par-kinsonian rats.
DISCUSSION
In this study, we report the identification (by a combinatorialscreening) of a novel small synthetic peptide (Cripto BP) that
antagonizes the onco-developmental signaling molecule,Cripto [2]. Although Cripto blocking antibodies have beenused in the past to suppress tumor cell growth [44], syntheticmolecules/peptides offer numerous advantages over biothera-peutics as they are easier and cheaper to produce, more stable,and generally free of contaminants of biological origin. Com-binatorial screenings have been successfully used in the lasttwo decades to identify small molecules/peptides for use indrug discovery and cell biology [45]. Recently, cell-basedscreenings have identified synthetic molecules that regulatestem cell differentiation/proliferation, which have been usedas probes to study the underlying biology of stem cells [46–49]. Although many basic questions regarding identificationof targets and mechanisms of action of the molecules identi-fied in cell-based screenings still remain to be answered, ithas become evident that such synthetic molecules could leadto significant advances in stem cell biology. In our study weused a reverse screening approach using a defined mecha-nism/target, the Cripto/ALK-4 receptor complex, and there-fore screened a synthetic peptide library to identify a CriptoBP. The choice of a synthetic peptide library was made aspeptides are known to be optimal probes to target the extrac-ellular domains of membrane receptors, provided their typicalhigh affinity and selectivity, minimal drug-drug interactions,low accumulation capacity, and low toxicity [31]. Moreover,tetrameric peptides are extremely flexible molecules, have ahigh-recognition surface, are highly resistant to proteases andcan thus be applied in functional assays and in a broad rangeof applications both in vitro and in vivo.
Based on these considerations, we designed an assay toidentify competitor peptides of the Cripto/ALK-4 receptorcomplex that could be used to control ESC fate. Consistentwith this idea, we found that Cripto BP prevented Cripto-de-pendent Smad2 phosphorylation, blocked cardiomyogenesisand favored neurogenesis in ESCs, mimicking the phenotypeof cripto�/� ESCs [12, 13]. Thus, these data and our in vitroresults showing that Cripto BP selectively binds Cripto andprevent its in vitro binding to ALK-4 receptor, support the ideathat, despite the function of ALK-4 as a receptor for multipleTGFb-related ligands [50], the neutralizing activities that wedetect are the result of specific targeting of the Cripto protein.
Interestingly, the BMPs antagonist Noggin was previouslyshown to induce neuroectodermal cell development andenhance production of DA neurons from hESCs [28, 29].Moreover, it has been recently reported that a combination ofNoggin and the small molecule SB431542, which inhibitsALK-4, -5, and -7 receptors [51] and blocks SMAD signaling,efficiently induce neural conversion of human pluripotentstem cells and allow their subsequent DA differentiation [30].Our protocol differs from previous studies in that we devel-oped and used a selective inhibitor (Cripto BP), instead ofbroader inhibitors such as Noggin and particularly SB431542,which blocks signaling by several TGF-b superfamily ligands.Our work indicates that a selective blocking with Cripto BPis sufficient to improve neural differentiation of mESCs invitro. This improvement was not at the expense of anincreased differentiation of Oct4þ cells, as their number didnot change by Cripto BP, but was rather due to a switch frommesodermal to neural differentiation. We thus suggest thatadditional cell sorting or BMP inhibitors could furtherimprove this protocol. In agreement with this, we found thatNoggin and Wnt5a enhanced the production of DA neuronsby Cripto BP in vitro. These findings indicate that Cripto BPcan indeed be used in combination with other factors to fur-ther improve DA differentiation.
With regard to cell transplantation in animal models ofPD, Chiba et al. [29], previously found that grafting of
Lonardo, Parish, Ponticelli et al. 1335
www.StemCells.com

hemiparkinsonian rats with hESCs-derived DA neurons pre-treated with Noggin lead to significant engraftment of DAneurons and behavioral improvement, but all animals showedcell overgrowths. These results differ from the study by Sonn-tag et al. [28], who showed teratomas in about 25% of theanimals grafted with Noggin-treated cells and behavioralimprovement that correlated with THþ cell numbers in onlytwo of the grafted animals. In our study, we found that CriptoBP treatment, led to grafts containing many more THþ cells(9,304 6 850 for BP and 5,841 6 1,939 for CP), and to aclear behavioral improvement in hemiparkinsonian rats. How-ever, our approach did not entirely eliminate tumors. CriptoBP significantly reduced the incidence of teratoma formation(by 58%) and the size of tumors, but teratomas were stillobserved in as many as 27% of the grafted rats, even in thepresence of additional factors, that is, Wnt5a and Noggin.Previous studies have shown that while transplantation of asfew as 20,00 cripto�/� ES-derived EB cells resulted in func-tional recovery and no tumor/overgrowth formation ESCs[13], 50,000 cripto�/� ES-derived EB cells did not prevent tu-mor/overgrowth formation, but lead to smaller grafts that con-tained more neurons [14]. We thus explored whether differentcell grafting densities of mESCs, differentiated as describedin Figure 4A, could reduce tumor/outgrowth formation whilemaintaining behavioral improvement, but found that cell den-sity plays a seemingly insignificant role in predifferentiatedmESCs in the presence of Cripto BP (supporting informationFig. 7). We also explored whether the efficiency of Criptoblocking with Cripto BP may have decreased in vivo, butfound that infusion of Cripto BP peptide in vivo did not fur-ther enhance the effect of Cripto BP pretreatment in vitro(data not shown). Thus, our results suggest that additional fac-tors present during DA differentiation and/or after graftingmay play an additional role in tumor/outgrowth formation.These data are thus in agreement with the results by Sonntaget al., who suggested that activation of other pathways, in acell autonomous or nonautonomous manner, may also contrib-ute to cell overgrowth in vivo [14]. Finally, it should be notedthat few previous studies have reported robust engraftment ofmES cell-derived DA neurons in animal models of PD in theabsence of tumor formation by either using transgenes [52,53], feeders [41] or knockout ES cells [13] (see [20] forreview). Our differentiation protocol, while aiming at avoid-ing genetic modification, requires the use of PA6 feeders. Inour protocol, we introduced some modifications to the proto-col by Barberi, which together with some variability intro-duced by PA6 cocultivation, may have contributed to incom-plete specification of the ESCs and tumor formation. In thefuture, it will thus be important to determine whether addi-tional instructive/differentiation factors, inhibitors of pathwaysinvolved in tumor formation and cell separation techniques,can be used to improve the functional integration of ES-
derived DA neurons in models of PD, making feeder-free andtransgene-free DA differentiation protocols safer, as well asmore reproducible and robust.
CONCLUSION
In conclusion, our strategy provides evidence that a smallsynthetic peptide obtained from an in vitro screen can notonly target specific proteins and signaling pathways in vitro,but can also enhance neural induction in mESCs, midbrainDA neuron differentiation and DA release. More importantly,Cripto BP can improve the number and the functional integra-tion of ESC-derived DA neurons, leading to behavioralimprovements in animal models of PD. This is the first timethat a reverse screening approach has been used in stem cellbiology to improve stem cell transplantation. However, thepresence of teratomas in a significant number of the graftedanimals remains as an important limiting factor. Hence, fur-ther efforts are required to elucidate the mechanisms responsi-ble for tumor formation and optimize CRT. Future studieswill examine whether hESCs respond to Cripto BP in a simi-lar manner as mESCs and whether the screening and targetingof additional signaling pathways may improve the therapeuticapplication of stem cells in CRT for PD.
ACKNOWLEDGMENTS
We are grateful to the members of the Stem Cell Fate Laboratoryand the Arenas laboratory for helpful discussion. This workwas supported by the Telethon Foundation to G.M. (grantsGGP05112, GGP08120) and S.D.F. (GGP08062), AssociazioneItaliana Ricerca sul Cancro (AIRC) to G.M. and S.D.F.; and bygrants from the European Union (Eurostemcell), the SwedishResearch Council (VR2008), Norwegian Research Council andKarolinska Institutet, to E.A. M.R. is funded by a FIRB (project,n� RBNE03PX83_005). C.L.P. was supported by a Human Fron-tiers Science Program Long-term Fellowship and a NationalHealth and Medical Research Council, Australia Career Devel-opment Award. D.R. was supported by a by a grant form the Por-tuguese Praxis XXI program of the Fundacao para a Ciencia e aTecnologia.
DISCLOSURE OF POTENTIAL CONFLICTS
OF INTEREST
The authors indicate no potential conflicts of interest.
REFERENCES
1 Strizzi L, Bianco C, Normanno N et al. Cripto-1: A multifunctionalmodulator during embryogenesis and oncogenesis. Oncogene 2005;24:5731–5741.
2 Minchiotti G. Nodal-dependant Cripto signaling in ES cells: Fromstem cells to tumor biology. Oncogene 2005;24:5668–5675.
3 Cheng SK, Olale F, Bennett JT et al. EGF-CFC proteins are essentialcoreceptors for the TGF-beta signals Vg1 and GDF1. Genes Dev2003;17:31–36.
4 Reissmann E, Jornvall H, Blokzijl A et al. The orphan receptor ALK7and the Activin receptor ALK4 mediate signaling by Nodal proteinsduring vertebrate development. Genes Dev 2001;15:2010–2022.
5 Schier AF. Chemokine signaling: Rules of attraction. Curr Biol 2003;13:R192–R194.
6 Massague J, Chen YG. Controlling TGF-beta signaling. Genes Dev2000;14:627–644.
7 Gray PC, Harrison CA, Vale W. Cripto forms a complex with activinand type II activin receptors and can block activin signaling. ProcNatl Acad Sci USA 2003;100:5193–5198.
8 Kelber JA, Shani G, Booker EC, Vale WW, Gray PC. Cripto is a non-competitive activin antagonist that forms analogous signaling com-plexes with activin and nodal. J Biol Chem 2008;283:4490–4500.
9 Gray PC, Shani G, Aung K et al. Cripto binds transforming growthfactor beta (TGF-beta) and inhibits TGF-beta signaling. Mol Cell Biol2006;26:9268–9278.
10 Shani G, Fischer WH, Justice NJ et al. GRP78 and Cripto form acomplex at the cell surface and collaborate to inhibit transforming
1336 Small Molecules for Stem Cell Therapies

growth factor beta signaling and enhance cell growth. Mol Cell Biol2008;28:666–677.
11 Shukla A, Ho Y, Liu X et al. Cripto-1 alters keratinocyte differentia-tion via blockade of transforming growth factor-beta1 signaling: Rolein skin carcinogenesis. Mol Cancer Res 2008;6:509–516.
12 Parisi S, D’Andrea D, Lago CT et al. Nodal-dependent Cripto signal-ing promotes cardiomyogenesis and redirects the neural fate of embry-onic stem cells. J Cell Biol 2003;163:303–314.
13 Parish CL, Parisi S, Persico MG et al. Cripto as a target for improvingembryonic stem cell-based therapy in Parkinson’s disease. Stem Cells2005;23:471–476.
14 Sonntag KC, Simantov R, Bjorklund L et al. Context-dependent neu-ronal differentiation and germ layer induction of Smad4(�/�) andCripto(�/�) embryonic stem cells. Mol Cell Neurosci 2005;28:417–429.
15 Parish CL, Arenas E. Stem-cell-based strategies for the treatment ofParkinson’s disease. Neurodegener Dis 2007;4:339–347.
16 Lindvall O, Hagell P. Clinical observations after neural transplantationin Parkinson’s disease. Prog Brain Res 2000;127:299–320.
17 Winkler C, Kirik D, Bjorklund A. Cell transplantation in Parkinson’sdisease: how can we make it work? Trends Neurosci 2005;28:86–92.
18 Kriks S, Studer L. Protocols for generating ES cell-derived dopamineneurons. Adv Exp Med Biol 2009;651:101–111.
19 Koch P, Kokaia Z, Lindvall O et al. Emerging concepts in neuralstem cell research: autologous repair and cell-based disease modelling.Lancet Neurol 2009;8:819–829.
20 Li JY, Christophersen NS, Hall V et al. Critical issues of clinicalhuman embryonic stem cell therapy for brain repair. Trends NeurosciMar 2008;31:146–153.
21 Lindvall O, Kokaia Z. Prospects of stem cell therapy for replacing do-pamine neurons in Parkinson’s disease. Trends Pharmacol Sci 2009;30:260–267.
22 Jonsson ME, Ono Y, Bjorklund A et al. Identification of transplantabledopamine neuron precursors at different stages of midbrain neurogene-sis. Exp Neurol 2009;219:341–354.
23 Lindvall O, Kokaia Z. Stem cells in human neurodegenerative disor-ders—Time for clinical translation? J Clin Invest 2010;120:29–40.
24 Chung S, Shin BS, Hedlund E et al. Genetic selection of sox1GFP-expressing neural precursors removes residual tumorigenic pluripotentstem cells and attenuates tumor formation after transplantation. J Neu-rochem 2006;97:1467–1480.
25 Hedlund E, Pruszak J, Lardaro T et al. Embryonic stem cell-derivedPitx3-enhanced green fluorescent protein midbrain dopamine neuronssurvive enrichment by fluorescence-activated cell sorting and function inan animal model of Parkinson’s disease. Stem Cells 2008;26:1526–1536.
26 Villaescusa JC, Arenas E. Transplantable midbrain dopamine neurons:A moving target. Exp Neurol 2010;222:173–178.
27 Friling S, Andersson E, Thompson LH et al. Efficient production ofmesencephalic dopamine neurons by Lmx1a expression in embryonicstem cells. Proc Natl Acad Sci USA 2009;106:7613–7618.
28 Sonntag KC, Pruszak J, Yoshizaki T et al. Enhanced yield of neuroe-pithelial precursors and midbrain-like dopaminergic neurons fromhuman embryonic stem cells using the bone morphogenic protein an-tagonist noggin. Stem Cells 2007;25:411–418.
29 Chiba S, Lee YM, Zhou W et al. Noggin enhances dopamine neuronproduction from human embryonic stem cells and improves behavioraloutcome after transplantation into Parkinsonian rats. Stem Cells 2008;26:2810–2820.
30 Chambers SM, Fasano CA, Papapetrou EP et al. Highly efficient neu-ral conversion of human ES and iPS cells by dual inhibition ofSMAD signaling. Nat Biotechnol 2009;27:275–280.
31 Marino M, Ruvo M, De Falco S et al. Prevention of systemic lupuserythematosus in MRL/lpr mice by administration of an immunoglob-ulin-binding peptide. Nat Biotechnol 2000;18:735–739.
32 Nagy A, Rossant J, Nagy R et al. Derivation of completely cell cul-ture-derived mice from early-passage embryonic stem cells. Proc NatlAcad Sci USA. 1993;90:8424–8428.
33 Xu C, Liguori G, Adamson ED et al. Specific arrest of cardiogenesisin cultured embryonic stem cells lacking Cripto-1. Dev Biol 1998;196:237–247.
34 Minchiotti G, Parisi S, Persico MG. Cripto signaling in differentiatingembryonic stem cells. Methods Mol Biol 2006;329:151–169.
35 Paxinos G, Watson C. The Rat Brain in Stereotaxic Co-ordinates. 3rded. Sydney: Academic Press, 1998.
36 Tam JP. Synthetic peptide vaccine design: Synthesis and properties ofa high-density multiple antigenic peptide system. Proc Natl Acad SciUSA 1988;85:5409–5413.
37 Boheler KR, Crider DG, Tarasova Y et al. Cardiomyocytes derivedfrom embryonic stem cells. Methods Mol Med 2005;108:417–435.
38 Smidt MP, Smits SM, Burbach JP. Homeobox gene Pitx3 and its rolein the development of dopamine neurons of the substantia nigra. CellTissue Res 2004;318:35–43.
39 Ye W, Shimamura K, Rubenstein JL et al. FGF and Shh signals con-trol dopaminergic and serotonergic cell fate in the anterior neuralplate. Cell 1998;93:755–766.
40 Castelo-Branco G, Wagner J, Rodriguez FJ et al. Differentialregulation of midbrain dopaminergic neuron development by Wnt-1,Wnt-3a, And Wnt-5a. Proc Natl Acad Sci USA 2003;100:12747–12752.
41 Barberi T, Klivenyi P, Calingasan NY et al. Neural subtype specifica-tion of fertilization and nuclear transfer embryonic stem cells andapplication in parkinsonian mice. Nat Biotechnol 2003;21:1200–1207.
42 Parish CL, Castelo-Branco G, Rawal N et al. Wnt5a-treated midbrainneural stem cells improve dopamine cell replacement therapy in par-kinsonian mice. J Clin Invest 2008;118:149–160.
43 Schein JC, Hunter DD, Roffler-Tarlov S. Girk2 expression in the ven-tral midbrain, cerebellum, and olfactory bulb and its relationship tothe murine mutation weaver. Dev Biol 1998;204:432–450.
44 Adkins HB, Bianco C, Schiffer SG et al. Antibody blockade of theCripto CFC domain suppresses tumor cell growth in vivo. J ClinInvest 2003;112:575–587.
45 Li W, Ding S. Small molecules that modulate embryonic stem cellfate and somatic cell reprogramming. Trends Pharmacol Sci 2010;31:36–45.
46 Ding S, Schultz PG. A role for chemistry in stem cell biology. NatBiotechnol 2004;22:833–840.
47 Chen S, Do JT, Zhang Q et al. Self-renewal of embryonic stem cellsby a small molecule. Proc Natl Acad Sci USA 2006;103:17266–17271.
48 Huangfu D, Maehr R, Guo W et al. Induction of pluripotent stem cellsby defined factors is greatly improved by small-molecule compounds.Nat Biotechnol 2008;26:795–797.
49 Borowiak M, Maehr R, Chen S et al. Small molecules efficientlydirect endodermal differentiation of mouse and human embryonicstem cells. Cell Stem Cell 2009;4:348–358.
50 Schmierer B, Hill CS. TGFbeta-SMAD signal transduction: Molecularspecificity and functional flexibility. Nat Rev Mol Cell Biol 2007;8:970–982.
51 Inman GJ, Nicolas FJ, Callahan JF et al. SB-431542 is a potent andspecific inhibitor of transforming growth factor-beta superfamily typeI activin receptor-like kinase (ALK) receptors ALK4, ALK5, AndALK7 Mol Pharmacol 2002;62:65–74.
52 Kim JH, Auerbach JM, Rodriguez-Gomez JA et al. Dopamine neuronsderived from embryonic stem cells function in an animal model ofParkinson’s disease. Nature 2002;418:50–56.
53 Fukuda H, Takahashi J, Watanabe K et al. Fluorescence-activated cellsorting-based purification of embryonic stem cell-derived neural pre-cursors averts tumor formation after transplantation. Stem Cells 2006;24:763–771.
See www.StemCells.com for supporting information available online.
Lonardo, Parish, Ponticelli et al. 1337
Copyright © 2022 FDOKUMEN



















